human monoclonal antibodies reactive with - cancer research
TRANSCRIPT

(CANCER RESEARCH 49, 1665-1670, April 1, 1989)
Human Monoclonal Antibodies Reactive with Human MyelomonocyticLeukemia Cells1
Marshall R. Posner,2 David J. Santos, Hillary S. Elboim, Marea B. Tumber, and A. Raymond Frackelton, Jr.
From the Division of Hematology/Oncology, Department of Medicine, Roger Williams General Hospital, The Roger Williams Cancer Center, and the Brown UniversityMedical School, Providence, Rhode Island 02908
ABSTRACT
Peripheral blood mononuclear cells from a patient with chronic mye-logenous leukemia (CML), in remission, were depleted of CDS-positiveT-cells and cultured with Epstein-Barr virus. Four of 20 cultures (20%)secreted human IgG antibodies selectively reactive with the cell surfacesof certain human leukemia cell lines. Three polyclonal, Epstein-Barrvirus-transformed, B-cell lines were expanded and fused with the human-mouse myeloma analogue HMMA2.11TG/O. Antibody from secretingclones HL 1.2 (IgG,), HL 2.1 (IgG3), and HL 3.1 (IgG,) have beencharacterized. All three react with HL-60 (promyelocytic), RWLeu4(CML promyelocytic), and U937 (monocytic), but not with KG-1 (mye-loblastic) or K562 (CML erythroid). There is no reactivity with T-celllines, Burkitt's cell lines, pre-B-leukemia cell lines, or an undifferentiated
CML cell line, BV173. Leukemic cells from two of seven patients withacute myelogenous leukemia and one of five with acute lymphocyticleukemia react with all three antibodies. Normal lymphocytes, mono-cytes, polymorphonuclear cells, red blood cells, bone marrow cells, andplatelets do not react. Samples from patients with other diverse hema-topoietic malignancies showed no reactivity. Immunoprecipitations suggest that the reactive antigen(s) is a lactoperoxidase iodinatable series ofcell surface proteins with molecular weights of 42,000-54,000 and anoniodinatable protein with a molecular weight of 82,000. Based on thesedata these human monoclonal antibodies appear to react with myelomon-ocytic leukemic cells and may detect a leukemia-specific antigen or ahighly restricted differentiation antigen.
INTRODUCTION
Autoantibody responses to autologous tumors have beennoted in a variety of human malignancies. Of particular interest,serological studies of patients with diverse acute and chronicleukemias have demonstrated the presence of human antibodieson fresh leukemic cells. Reactivity of these antibodies wasfrequently found to extend to allogeneic leukemias but notnormal cells (1-4). These studies suggest that tumor-specificantigens are present on the cell surfaces of human leukemicblasts and that the patient can recognize these antigens asforeign or abnormal. Such tumor-specific or tumor-related antigens may represent virally encoded antigens such as thoseseen with human T-cell leukemia virus I-associated lymphomasand leukemias, altered growth factor receptors, oncofetal antigens, or normal cellular antigens presented in an abnormalcontext.
The role of these antibodies and the antigens with which theyreact may be important in understanding the pathogenesis ofthese diseases and in developing therapeutic strategies. However, with the exception of human T-cell leukemia virus Iantigens and antibodies, evaluation of these responses has notproceeded (5, 6). In part this may be due to the polyclonalnature of the human humoral response and the difficulty in
Received 6/7/88; revised 11/29/88; accepted 12/29/88.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
'This work was supported by a grant (RO1-CA38687) from the National
Cancer Institute.2To whom requests for reprints should be addressed, at Division of Oncology/
Hematology, Roger Williams General Hospital, Brown University MedicalSchool, 825 Chalkstone Avenue, Providence, Rl 02908.
performing reproducible studies with these polyclonal sera andheterogenous leukemias. It is apparent that to study theseantibodies it would be important to obtain them in large quantities and pure form. Human monoclonal antibody technologycan provide these reagents but until recently has not beenavailable (7).
In the present study we describe the production and initialcharacterization of three human monoclonal antibodies obtained from a single patient with CML.3 These antibodies reactwith leukemic cells of myelomonocytic origin, recognizing an-tigenic determinants on the cell surfaces of several humanmyelomonocytic cell lines and leukemic blasts from patientswith certain acute leukemias, but not with normal hemato-poietic cells of diverse origin.
MATERIALS AND METHODS
Cell Culture. Cell lines and established hybridomas were grown in a-minimum essential medium lacking nucleosides, supplemented with: 1HIMsodium pyruvate, 2 HIML-glutamine, 1% (v/v) nonessential aminoacids, 10% (v/v) fetal bovine serum (high cloning efficiency and growthpromotion; GIBCO, Grand Island, NY), 0.22% (w/v) sodium bicarbonate, and 50 ^g/ml gentamicin. All other cell culture was performedwith the same media containing 20% fetal bovine serum. Other additives were included as indicated. Cultures growing in flasks were flushed,sealed with a 5% COz/air (v/v) mixture and maintained at 37°C.
Repeated gassings with the CU2 were performed as needed to maintainproper pH. Cultures in microtiter plates or multiwell plates wereincubated in a 5% CO2 atmosphere at 37°Cin a humidified incubator.
Cell Lines. The HMMA2.11TG/O cell line, a nonsecreting humanmouse myeloma analogue, developed in this laboratory, was used forfusions (HB 9583, ATCC, Rockville, MD) (7). The B95-8 marmosetcell line was used as a source of EBV for cell transformation. TheOKT8 hybridoma was obtained from the ATCC (CRL 8014, ATCC,Rockville, MD). The origins of the leukemic cell lines used in thesestudies are described in several reviews (8-10). These cell lines werekindly supplied by a number of investigators.4
Volunteer and Patient Cells. After informed consent, PBM and bonemarrow mononuclear cells were obtained by venipuncture or bonemarrow aspiration in preservative-free heparin and separated fromcontaminating cells by density gradient separation as previously described (7). If not used immediately, cells were stored by cryopreserva-tion in liquid nitrogen after resuspension in media containing 10%dimethyl sulfoxide.
Cell Fusion. Fusions were performed according to our standardprocedure (7). In brief, the human-mouse myeloma analogueHMMA 2.11TG/O was fused in a minimum ratio of 2:1 with the otherparental cells. Fused cells were distributed in 96-well microtiter plates
3The abbreviations used are: CML, chronic myelogenous leukemia; EBV,Epstein-Barr virus; HAT, hypoxanthine, aminoptermin, thymidine; HT, hypo-xanthine, thymidine; HuMoab, human monoclonal antibody; SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis; ALL, acute lymphocyticleukemia; PBM, peripheral blood mononuclear cells; PSG, Puck's saline G; I'M.WO, Puck's saline G without calcium and magnesium; PBS, phosphate bufferedsaline; PSG2.5, Puck's saline G with 2.5% fetal bovine serum; RIPA, radio
immunoprecipitation assay.4Cell lines were kindly supplied by Dr. H. Lazarus (Centecor Corporation,
Malvern, PA), Dr. R. Todd (University of Michigan, Ann Arbor, MI), Dr. J. Ritz(Dana Farber Cancer Institute, Boston, MA), Dr. J. Pesando (BiomembraneInstitute, Seattle, WA), and Dr. M. Weimann (Roger Williams General Hospital,Providence, RI).
1665
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HUMAN MONOCLONAL ANTIBODIES AND LEUKEMIA
in HAT-containing media. The number of cells seeded into each wellin any experiment was based on the maximum number of potentialhybrids given a hypothetical 1/1 fusion efficiency. The final volume ineach well was 200 /il. In all experiments 10 MMouabain in 50 n\ ofmedia with HT was added to yield a 2 /i\i ouabain concentration ineach well 24 h after fusion to prevent the growth of normal or transformed cells. A concentration of 2 ?M ouabain was maintained in thewells for 1 week, after which routine feeding was performed. Fusionswere fed at 4-7-day intervals by removal of 100-150 ^1 of media andreplacement with an equal volume of media containing HT. Afterselection for expansion, cells were transferred to 24-well multiwellplates in media containing HT and were maintained in HT until theywere passaged once in flasks. Hybridomas were cloned by resuspensionin media containing HT and distributed in microtiter plates such thatan average of 1 cell/100 ¿¿I/wellwas obtained. Cloned cells were fed atweekly intervals with media lacking HT.
Complement Depletion of Selected T-Cells and EBV Transformation.PBM were selectively depleted of CDS-positive T-cells by antibody-mediated complement lysis. Cells were suspended at a density of 10 x106/ml in media and 100 ^1 of culture supernatant from an OKT8
culture were added. The cells were incubated for 30 min at roomtemperature, rabbit complement (Pel Freeze, Brown Deer, WI) wasadded at a final concentration of 1:5, and the cells incubated for l h at37"C. The cells were then washed three times with Puck's Saline G
without calcium or magnesium.For EBV transformation, PBM were placed in 24-well plates such
that there were 0.2-0.3 x 10* cells per well, in 1.0 ml of mediacontaining a 1:10 dilution of stock B95-8 supernatant according to theprelysis cell number. EBV-transformed cultures were maintained byfeeding weekly with the addition of small amounts of media until cellgrowth was observed and then every 3-4 days. Growing cultures weretreated as above.
Indirect Immunofluorescence of Cell Surface Antigens. Indirect im-munofluorescence for the detection of cell surface antigens was performed as previously described with some modifications (11). In brief,1.0 x IO6 live cells per sample were washed with PBS. To block Fc
binding, cells were incubated with 100 /.Aof nonspecific mouse ascitesfor 15 min at 4"( ' and then washed once. 100 //I of culture fluid
containing monoclonal antibody were added to the cell pellet andincubated at 4°Cfor 30 min. The cells were then washed twice with
PBS, followed by the addition of 100 p\ of fluorescein-conjugatedF(ab')2 goat anti-human IgG or polyspecific immunoglobulins (Tago,
Inc., Burlingame, CA) diluted in PSG 2.5%. The sample was incubatedfor 30 min at 4°Cand then washed once in PBS. Following this, the
pellet of live cells was resuspended in 250 n\ PBS and then 250 M'of1% (v/v) formaldehyde in PBS was added to fix the sample. The fixedcells were resuspended and stored up to 5 days until analysis at 4°C.
Analysis was performed on an Epics C cell sorter (Coulter, Hialeah,FL). Cells fixed with formaldehyde after labeling with antibodies havebeen shown to retain, without significant alteration, prefixation findingsupon flow cytofluorimetric analysis. Fixation permits storage of thesample for delayed evaluation and neutralizes human pathogens thatmight be present in the sample. Our own experiments and those ofothers have shown no effect of fixation on analysis under these conditions (12, 13). Negative controls for these experiments consisted ofnormal human sera diluted 1:1000 (approximately 10-20 jig/ml of IgG)and the monoclonal IgG immunoglobulins and antibodies 7C7 (IgGjimmunoglobulin) and I I 11)12 (IgGi antitetanus human monoclonalantibody) (7). All negative controls gave the same results in multipleassays.
Radioimmunoprecipitation of Antigens. U937 cells were surface labeled by the lactoperoxidase method with '"I (1.0 mCi/107 cells) and
lysed with RIPA buffer (14). After centrifugation to remove nuclei anddebris, the lysates were incubated with 25 n\ of protein A-Sepharose-4B CL (Pharmacia, Uppsala, Sweden) that had been preincubated with100 //I of goat anti-human immunoglobulins (Tago, Burlingame, CA),and 500 /il of HuMoab supernatant. After washing away unboundproteins, bound antigens were eluted by boiling in Laemmli buffer andsubjected to SDS-PAGE under reducing conditions.
RWLeu4 and U937 cells were metabolically labeled with [35S]methi-
onine by culture in methionine-free media supplemented with 20% cellculture medium described above and 0.3 mCi of [35S]methionine/5 xIO5cells for 14 h. The cells were centrifuged and lysed with RIPA andthe lysates immunoprecipitated with sepharose conjugated to goat anti-human IgG (Caltag, San Francisco, CA) that had been preincubatedwith HuMoabs. After unbound lysate protein had been removed by fivewashes in RIPA buffer, the bound antigen was eluted by boiling inLaemmli buffer and subjected to SDS-PAGE under reducing conditions. Unreduced eluates were diluted in RIPA buffer to reduce theconcentration of SDS and then reprecipitated with the identicalHuMoabs. These secondary immunoprecipitates were washed, eluted,reduced, and analyzed by SDS-PAGE as above.
Determination of Antibody Isotype and Secretion. Supernatants fromtest wells or bulk cultures were tested for IgG isotype using a modification of a microelisa assay as described previously (7). In brief, testwells (Immulon 2; Dynatech, Alexandria, VA) were coated with 50 M'of mouse monoclonal antibody to isotypes IgGi, IgGz, IgGs, or IgG4(Caltag Laboratories) and incubated for at least 2 h. Plates were thenblocked with PSG with 2.5% fetal bovine serum (v/v) (PSG 2.5%) fora minimum of 2 h, washed twice with PBS with 0.5% tween 20 (v/v)(PBS-tween) and twice with PBS. Then 100 ¿ilof test supernatant wereadded. The wells were incubated for 2 h, washed as above, and 75 p\ ofperoxidase-conjugated goat anti-human immunoglobulins (IgM, IgG,and IgA) or specific peroxidase-conjugated goat anti-human IgG, IgM,or IgA (Tago, Inc., Burlingame, CA) diluted 1:3000 in PSG2.5% wereadded and incubated for 1 h. The wells were washed three times withPBS-tween and three times with PBS after which 100 n\ of O-phen-ylenediamine in citrate buffer were added. Plates were read at 5-45min by observing a color change and scoring from negative to 4~.
Quantitation of secretion was performed by a modification of theassay described above. Wells were coated with 75 n\ of a 30-^g/mlsolution of goat anti-human immunoglobulins. Supernatants from routinely growing cultures were collected every 3-4 days and pooled.Dilutions in PSG2.5% were tested as above using peroxidase-conjugated goat anti-human IgG antibodies. A standard curve was generatedby serial dilution of a purified preparation of ¡sotypespecific humanmonoclonal antibody of known antibody content, and the antibodycontent of the experimental Supernatants determined simultaneously.Optical density was quantitatively determined on a microelisa readerand used to quantify the reaction.
RESULTS
Production of Human Monoclonal Antibodies. Peripheralblood mononuclear cells were obtained from a patient withCML in hematological remission following one cycle of intensive chemotherapy. The PBM were depleted of CDS-positiveT-cells and then distributed into 20 wells of a 24-well cultureplate at 250,000 cells/well with EBV. Within 3 weeks all 20 ofthe cultures showed evidence of transformation. Supernatantsfrom the EBV-transformed cultures were initially screened forreactivity with the human leukemic cell lines RWLeu4, HL60,and K562 by cell surface indirect immunofluorescence. Negative controls included monoclonal IgM and IgG immunoglobulins, sera, and antitetanus human monoclonal antibodies ofIgG and IgM classes. Positive controls were not available. Fourpresumed polyclonal EBV-transformed B-cell cultures reactedwith the acute promyelocytic cell line HL60 and the myeloidCML cell line RWLeu4, but were negative with the CMLerythroleukemia cell line K562. The remaining 16 Supernatantswere negative with all three cell lines.
All four of the polyclonal EBV-transformed B-cell lines secreting reactive antibodies were expanded and three were fusedwith the human-mouse myeloma analogue HMMA2.11TG/O.The results of these fusions are shown in Table 1. The fusionefficiency was high and, in two fusions, the majority of thehybridomas secreted antibody reactive with the two myeloidleukemic cell lines. These results are in contrast to our previous
1666
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HUMAN MONOCLONAL ANTIBODIES AND LEUKEMIA
Table 1 Results of the fusion ofHMMA2.11TG/O with antileukemic antibodysecreting polyclonal B-cell lines
Fusion Density" Wells Growth AB AB Cloned
F37F38F4012,00010,0005,2003843841921009921,92*3211001.652212
" Fused EBV transformants/well.* 192 wells each from fusions F37 and F38 were tested by indirect immunoflu-
orescence.
Table 2 Reactivity of human monoclonal antibodies with cell lines ofhematopoietic origin
Antibody
CelllineHL60RWLeu4BV173K562KG1U937CEMMOLT4HPB-ALLRAMOSJD39NALM1NALM6HL1.13V2+0004+0000000HL2.13+2+0004+0000000HL3.13+2+0004+0000000
" Cell surface reactivity by indirect ¡mmunofluorescence above background. 0•¿�•0-10%, 1+ = 11-25%, 2+ = 26-50%, 3+ = 51-75%, 4+ = 76-100%.
studies with tetanus toxoid in which fusion efficiency with EBVtransformants is slightly lower and the percentage of antibody-secreting wells substantially lower. Antibody-secreting hybri-domas from each fusion were randomly selected, cloned andrecloned. Recloned hybridomas were then used to generateculture supernatants containing human monoclonal antibodieswhich were used for subsequent studies. Antibody secretion byrecloned hybridomas has been stable for periods of up to 6months without recloning. Cultures were terminated at thattime without loss of secretion for reasons of economy. Thethree human monoclonal antibodies have been designatedHL1.1, HL2.1, and HL3.1. Isotype analysis indicates that theyare of the IgGi, IgG3, and IgGi isotypes, respectively. Routinelygrowing cultures were found to yield 11, 4, and 3.5 ¿¿g/mlofmonoclonal antibody respectively in pooled supernatants.
Reactivity with Human Hematopoietic Cells. The three humanmonoclonal antibodies demonstrated identical patterns of cellsurface reactivity with established human hematopoietic celllines of diverse origin. As shown in Table 2, they react with apromyelocytic cell line (HL60), a promyelocytic CML cell line(RWLeu4), and a monoblastoid cell line (U937), but not withother myeloid or undifferentiated cell lines (KG1, BV173), anerythroleukemia CML cell line (K562), T-cell lines (CEM,MOLT4, HPB-ALL), Burkitt's lymphoma cell lines (JD39,RAMOS), or pre-B-leukemia cell lines (NALM1, NALM6).Thus, the reactivity of these antibodies appears to be restrictedto cell lines of myelomonocytic origin. Examples of the cyto-fluorographic reactivity of each of the antibodies with one ofthe cell lines, HL60, RWLeu4, or U937, are shown in Fig. 1.
The antibodies were further tested for reactivity with normalhematopoietic cells and leukemic cells from patients with diverse forms of chronic and acute leukemia. As shown in Table3, no reactivity was observed with normal peripheral blood cellsor bone marrow cells. Leukemic cells from two of seven patientswith acute myelogenous or myelomonocytic leukemia and onepatient with ALL were reactive with all three antibodies whileleukemic cells from patients with other forms of leukemia wereunreactive. Interestingly, the antibodies did not react with the
A
B
C
Fig. 1. Representative cytofluorographic determinations of indirect immuno-fluorescent cell surface reactivity of the human monoclonal antibodies on the celllines U937,1II 60, and RWLeu4. One antibody is shown with each cell line sinceall react in the same fashion. Each figure has a negative control antibody to theleft of the arrow, overlapping (darkly shaded area) of the negative and positiveantibody, and positive antibody to the right of the arrow. The curves are expressedas log green fluorescence (X-axis) by cell number (Y-axis). /4, U937 and HL1.1(85% positive); B, HL60 and HL2.1 (50% positive); and C, RWLeu4 and HL3.1(28% positive). The negative control antibody in A-C is F11DE2, an IgGiantitetanus human monoclonal antibody, and is adjusted to 7% positivity in theassays.
chronic phase PBM of the patient from whom the hybridomaswere derived, perhaps because of modulation of antigenic determinants by preexisting serum antibody or the later stage ofdifferentiation of the circulating cells. Blast phase cells werenot available from this patient. Analysis of the ALL patient's
cells with monoclonal antibodies demonstrated reactivity witha typical pre-B-cell leukemia pattern of CD 10, CD 19, andCD20 positivity. In addition, however, a fraction of the cellsexpressed MY7, a myeloid marker, indicating some lineageinfidelity (15).
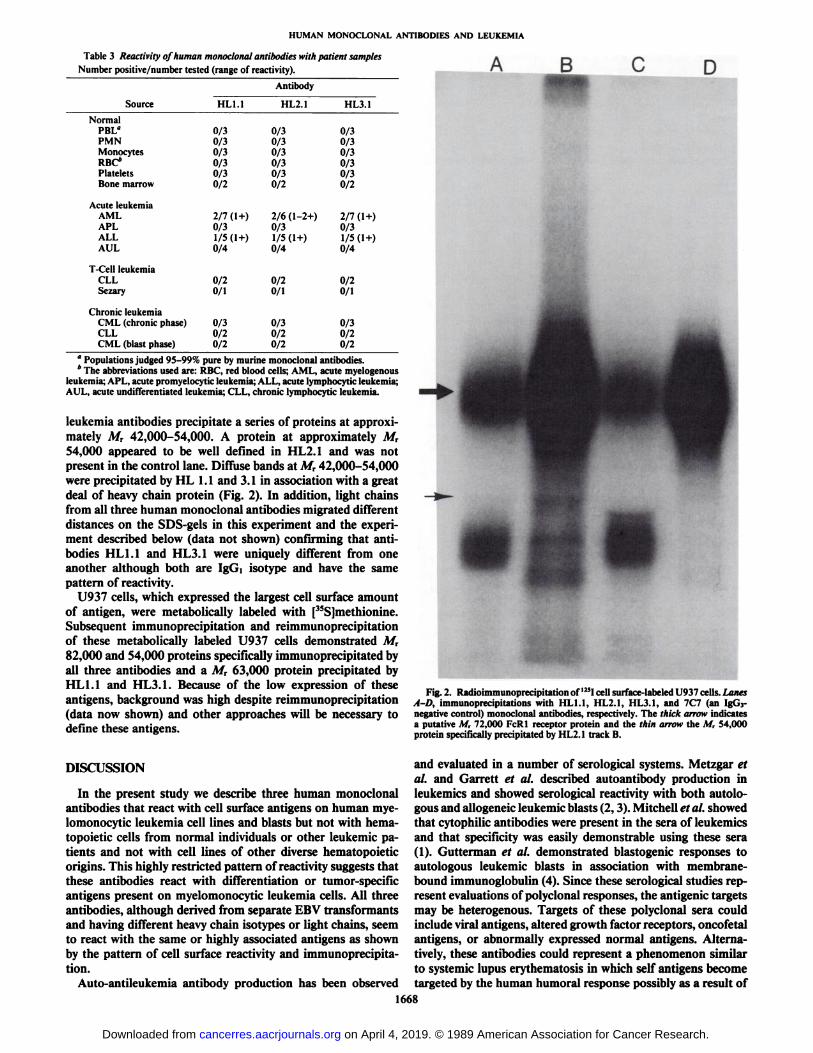
Radioimmunoprecipitation of Antigen. In order to assess thecell surface expression of these proteins, U937 cells were surfacelabeled with I25I, immunoprecipitated, and the immunoprecip-itates resolved by SDS-PAGE. In addition to the known M,72,000 Fc receptor, precipitated by the negative control IgGhuman monoclonal antitetanus antibody (16), the human anti-
1667
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
HUMAN MONOCLONAL ANTIBODIES AND LEUKEMIA
Table 3 Reactivity of human monoclonal antibodies with patient samplesNumber positive/number tested (range of reactivity). B D
SourceNormalPBL"PMNMonocytesRBC*PlateletsBone
marrowAcute
leukemiaAMLAPLALLAULT-Cell
leukemiaCLLSezaryChronic
leukemiaCML(chronicphase)CLLCML
(blast phase)HL1.10/30/30/30/30/30/22/7
(1+)0/31/5(1+)0/40/20/10/30/20/2AntibodyHL2.10/30/30/30/30/30/22/6(1-2+)0/31/5(1+)0/40/20/10/30/20/2HL3.10/30/30/30/30/30/22/7(1+)0/31/5(1+)0/40/20/10/30/20/2
" Populations judged 95-99% pure by murine monoclonal antibodies.* The abbreviations used are: RBC, red blood cells; AML, acute myelogenous
leukemia; APL, acute promyelocytic leukemia; ALL, acute lymphocytic leukemia;AUL, acute undifferentiated leukemia; CLL, chronic lymphocytic leukemia.
leukemia antibodies precipitate a series of proteins at approximately A/r 42,000-54,000. A protein at approximately A/r54,000 appeared to be well defined in HL2.1 and was notpresent in the control lane. Diffuse bands at A/r 42,000-54,000were precipitated by HL 1.1 and 3.1 in association with a greatdeal of heavy chain protein (Fig. 2). In addition, light chainsfrom all three human monoclonal antibodies migrated differentdistances on the SDS-gels in this experiment and the experiment described below (data not shown) confirming that antibodies HL1.1 and HL3.1 were uniquely different from oneanother although both are Igd isotype and have the samepattern of reactivity.
U937 cells, which expressed the largest cell surface amountof antigen, were metabolically labeled with [35S]methionine.
Subsequent immunoprecipitation and reimmunoprecipitationof these metabolically labeled U937 cells demonstrated A/r82,000 and 54,000 proteins specifically immunoprecipitated byall three antibodies and a Afr 63,000 protein precipitated byHL1.1 and HL3.1. Because of the low expression of theseantigens, background was high despite reimmunoprecipitation(data now shown) and other approaches will be necessary todefine these antigens.
DISCUSSION
In the present study we describe three human monoclonalantibodies that react with cell surface antigens on human mye-lomonocytic leukemia cell lines and blasts but not with hema-topoietic cells from normal individuals or other leukemic patients and not with cell lines of other diverse hematopoieticorigins. This highly restricted pattern of reactivity suggests thatthese antibodies react with differentiation or tumor-specificantigens present on myelomonocytic leukemia cells. All threeantibodies, although derived from separate EBV transformantsand having different heavy chain isotypes or light chains, seemto react with the same or highly associated antigens as shownby the pattern of cell surface reactivity and immunoprecipitation.
Auto-antileukemia antibody production has been observed
Fig. 2. Radioimmunoprecipitation of 125Icell surface-labeled U937 cells. LanesA-D, immunoprecipitations with HL1.1, HL2.1, HL3.1, and 7C7 (an IgG3-negative control) monoclonal antibodies, respectively. The thick arrow indicatesa putative M, 72,000 FcRl receptor protein and the thin arrow the M, 54,000protein specifically precipitated by HL2.1 track B.
and evaluated in a number of serological systems. Metzgar etal. and Garrett et al. described autoantibody production inleukemics and showed serological reactivity with both autolo-gous and allogeneic leukemic blasts (2,3). Mitchell et al. showedthat cytophilic antibodies were present in the sera of leukemicsand that specificity was easily demonstrable using these sera(1). Gutterman et al. demonstrated blastogenic responses toautologous leukemic blasts in association with membrane-bound immunoglobulin (4). Since these serological studies represent evaluations of polyclonal responses, the antigenic targetsmay be heterogenous. Targets of these polyclonal sera couldinclude viral antigens, altered growth factor receptors, oncofetalantigens, or abnormally expressed normal antigens. Alternatively, these antibodies could represent a phenomenon similarto systemic lupus erythematosis in which self antigens becometargeted by the human humoral response possibly as a result of
1668
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HUMAN MONOCLONAL ANTIBODIES AND LEUKEMIA
immune dysregulation induced by the disease or the therapy(17). Study of these antigens and antibodies might be importantin understanding the pathogenesis of these diseases, the role ofthe host immune response in disease progression, and in thedevelopment of potentially therapeutic reagents. The development of murine monoclonal antibody technology shifted theemphasis away from studies of these autochthonous systems.Unfortunately, murine monoclonal antibody technology doesnot address the repertoire of the human auto-antitumor response. This is inherent in the murine system because themurine host preferentially reacts with differentiation and human specific antigens (7). Thus human monoclonal antibodieswill be useful in capturing these antibodies so that we maystudy the repertoire of these responses and the impact of theseantibodies and antigens on the progression of these diseases.
Solid tumor-reactive human monoclonal antibodies from patients with cancer have been reported by several investigators(18-23). Frequently, these human monoclonal antibodies, particularly IgM antibodies, have been found to react with a broadrange of normal and abnormal cells. Occasionally, a normalintracellular component, such as cytokeratin, which is not normally antigenic, is found to be the target of antibody reactivityand confers an element of tumor tissue specificity (18, 22).Although IgG human monoclonal antibodies have been rare,the antisolid tumor human monoclonal IgG antibodies havehad similar patterns of reactivity (20, 23). Our own studies haveshown that IgM human monoclonal antibodies with diffuse cellsurface reactivity to neoplastic and normal hematopoietic cellsare relatively easy to obtain from both diseased and healthyindividuals5'6 while tumor-specific antibodies may be both dif
ficult to obtain and difficult to prove as being tumor specific.Relatively specific IgG human monoclonal antibodies reac
tive with cell surface antigens on human leukemia cell lines andleukemic blasts have been reported by Andreasen and Olsson.They were able to prepare two separate IgG HuMoabs withsimilar reactivities from a patient with acute myelogenous leukemia and a patient with CML (24). The pattern of reactivitymay have been a function of their screening method whicheliminated reactivity with a variety of other hematopoietic celllines. The antibody described by these coworkers reacted withan intracellular antigen in normal bone marrow cells and anabnormal A/r 18,000 antigen expressed on the cell surfaces ofleukemic cells.
The antibodies described in the present study have a significantly different pattern of reactivity with cell lines and lessbroad reactivity with AML patients than those described byAndreasen and Olsson. In addition, all three antibodies immu-noprecipitate proteins of M, 82,000 and 54,000 from metabol-icaly labeled leukemic cells and a series of iodinatable cellsurface-associated proteins in the range of M, 42,000-55,000.The data from these experiments are consistent with the M,80,000 protein being an intracellular antigen while the M,42,000-54,000 proteins are surface antigens. Alternatively,these antigens may be unrelated and share epitopes or may bedisulfide-linked chains of a heterooligomeric protein. The differences also suggest that HL1.1 and HL3.1 are detectingidentical antigens, while HL2.1 has a different but highly related specificity.
5 M. R. Posner, H. J. Barrach, H. S. Elboim, K. Nievens, D. J. Santos, C. O.
Chichester, and E. V. Lally. The generation of hybridomas secreting humanmonoclonal antibodies reactive with type II collagen. Hybridoma, in press.
6 M. R. Posner, H. S. Elboim, and M. Tumbor. Human monoclonal antibodies
from a patient with acute lymphocytic leukemia that react with cell surfaceantigens on normal and leukemic cell populations. Manuscript in preparation.
Unlike the system described by Andreasen and Olsson, in thepresently described system for HuMoab production, preselection of EBV transformants on the basis of specificity, for fusion,is possible. Fusions were performed with three of the fourantibody-producing transformants, but all four had the samereactivity pattern in the initial screening against HL60, K562,and RWLeu4. If reactivity patterns had varied, preselection forfusion would have been performed. This is an important potential advantage of the EBV transformation/fusion method withthe high fusion efficiency cell line HMMA2.11TG/O. Of interest, fusion efficiency was 25-50% higher than that seen in ourtetanus toxoid or collagen experimental systems,5 and equivalent to that seen in other antileukemia antibody experiments.6
Furthermore, compared to the tetanus or collagen systems, thefrequency of hybridomas producing the desired antibody is 10-20 times greater in the antileukemia experiments. Taken together, these data support the notion that a large fraction ofcirculating B-cells in this leukemic patient were able to produceantibodies to these antigens and indicate that these antigens arehighly immunogenic. One may speculate an important biological role for either the antigens or the humoral response.
In the present study, we have demonstrated that auto-antileu-kemia human monoclonal antibodies were readily obtainablefrom a patient with CML. These antibodies have highly restricted patterns of reactivity with established human leukemiacell lines and leukemic blasts, and lack significant reactivitywith normal hematopoietic cells, suggesting that they react withtumor-specific antigen(s) on myelomonocytic leukemia cells, orhighly restricted differentiation antigen(s). The biological importance and potential therapeutic use of these antibodies andantigens will be the subject of future studies.
REFERENCES
1. Mitchell, M. S., Mokyr, M. B., Aspnes, G. T., and Mclntosh, S. Cytophilicantibodies in man. Ann. Intern. Med., 79: 333-339, 1973.
2. Metzgar, R. S., Mohanakumar, T., and Miller, D. S. Membrane-boundimmunoglobulins on human leukemic cells. J. Clin. Invest., 56: 331-338,1975.
3. Carre«, T. J., Takahashi, T., Clarkson, B. D., and Old, L. J. Detection ofantibody to autologous human leukemia cells by immune adherence assays.Proc. Nati. Acad. Sci. USA, 74:4587-4590, 1977.
4. ( ¡interinan.J. U., Rossen, R. D., Butler, W. T., McCredie, K. B., Bodey, G.P., Freireich, E. J„and Hersh, E. M. Immunoglobulin on tumor cells andtumor-induced lymphocyte blastogenesis in human acute leukemia. N. Engl.J. Med., 288: 169-175, 1973.
5. Posner, L. E., Rogert-Guroff, M., Kalyanaraman, V. S., Poiesz, B. J.,Ruscelli, F. W., Fossieck, B., Bunn, Jr., P. A., Minna, J. D., and Gallo, R.C. Nalural antibodies to Ihe human T cell lymphoma virus in patienls wilhculaneous T cell lymphomas. J. Exp. Med., 154: 333-346, 1981.
6. Mann, D. L., DeSantis, P., Mark, G., Pfeifer, A., Newman, M., Gibbs, N.,Popovic, M., Sarngadharan, M. G., Gallo, R. C., Clark, J., and Blallner, W.HTLV-1-Associated B-cell CLL: Indirecl role for relrovirus in leukemo-genesis. Science (Wash. DC), 236: 1103-1106, 1987.
7. Posner, M. R., Elboim, H., and Sanios, C. The conslruction and use of ahuman-mouse myeloma analogue suitable for ihe routine production ofhybridomas secreting human monoclonal anlibodies. Hybridoma, 6: 611-625, 1987.
8. Minowada, J., Koshiba, H., Sagawa, K., et al. Marker profiles of humanleukemia and lymphoma cell lines. J. Cancer Res. Clin. Oncol., 101:91-100,1981.
9. Wiemann, M. C., Hollmann, A., Posner, M., Arlin, Z., Friedland, M., andCalabresi, P. Eslablishment of a new Philadelphia chromosome posilivehuman chronic myeloid leukemia cell line. Clin. Res., 33:461 A, 1985.
10. Posner, M. R., Antoniou, D., Griffin, J., Schlossman, S. F., and Lazarus, H.An enzyme-linked immunosorbent assay (elisa) for ihe detection of monoclonal antibodies to cell surface antigens on viable cells. J. Immunol. Methods, 4«:23-31, 1982.
11. Posner, M. R., Schlossman, S. F., and Lazarus, H. Novel approach to theconstruclion of human "myeloma analogues" for ihe produclion of humanmonoclonal antibodies. Hybridoma, 2: 369-381, 1983.
12. Posner, M. R., Reinherz, E. L., Breard, J., Nadler, L. M., Rosenlhal, D. S.,and Schlossman, S. F. Lymphoma subpopulalions of peripheral blood andspleen in untreated Hodgkin's disease. Cancer (Phila.), 48:1170-1176,1981.
13. Lifson, J. D., Sasaki, D. T., and Engelman, E. G. Utilily of formaldehyde
1669
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

HUMAN MONOCLONAL ANTIBODIES AND LEUKEMIA
fixation for flow cytometry and inactivation of the AIDS associated retrovi- Peters, L. C, Hoover, H. C, and Hanna, M. G. Generation of tumor cell-rus. J. Immunol. Methods, 86: 143-149, 1986. reactive human monoclonal antibodies using peripheral blood lymphocytes
14. Gielkins, A. L. J., Van Zaane, D., Bloemers, H. P. J., and Bloemendal, H. from actively immunized colorectal carcinoma patients. Cancer Res., 45:Synthesis of Rauscher murine leukemia virus-specific polypeptides in vitro. 3951-3961, 1985.Proc. Nati Acad. Sci USA, 73' 356-360 1976 20. Kan-Mitchell, J., Imam, A., Kempf, R. A., Taylor, C. R., and Mitchell, M.
15. Foon, K. A., and Todd, III, R. F. Immunologie classification of leukemia s- Human monoclonal antibodies directed against melanoma tumor associ-and lymphoma. Blood, 68:1-31, 1986. ated antigens. Cancer Res., 46: 2490-2496, 1986.
16. Liesveld, J. L., Abboud, C. N., Looney, R. J., Ryan, D. H., and Brennan, J. 21" Smith, L. "•'Y'". A." »"ber, M., and Teng, N. N. H. Generation of humanK. Expression of IgG Fc receptors in myeloid leukemic cel, lines. J. Immu- ---•.£*--* y-ggg^
n M. K ju? r M T'n .-r , t.t. , , 22. Ho, M-K, Geist, C, Murray, J., Kalo, K., DeLellis, R., Wolfe, H., Carney,17. Mmota, S., and Wmfield, J. B. Identification of three major target molecules w Rabm „¿�and Dur^ >p Distribution ^ immunochemical character-
of IgM antilymphocyte autoantibodies m systemic lupus erythematosus. J. ¡zationof a keratjn.iike antigen ¡nepithelial tumors using mouse and humanImmunol., 139: 3644-3651, 1987. monoclonal antibodies. Cancer Res., 48:4969-4975, 1988.
18. Skaletsky, E., Oh, E., Rulot, C., Baird, S. M., Burnett, K. G., Masuho, Y., 23. Kjeldsen, T. B., Rasmussen, B. R., Rose, C., and Zeuthen, J. Human-humanAstanta, R. W., Haghighi, P., Collins, H., Starr, R., Carlo, D. J., Royston, hybridomas and human monoclonal antibodies obtained by fusion of lymphI., and Soboi, R. E. A human monoclonal antibody to cytokeratin interme- „¿�odelymphocytes from breast cancer patients. Cancer Res., 48: 3208-3214,diate filament antigens derived from a tumor draining lymph node. Hybrid- 1988.oma, 7: 367-376, 1988. 24. Andreasen, R. B., and Olsson, L. Antibody-producing human-human hybri-
19. Haspel, M. V., McCabe, R. P., Pomato, N., Janesch, N. J., Knowlton, J. V., domas. J. Immunol. Methods, 137:1083-1090, 1986.
1670
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1989;49:1665-1670. Cancer Res Marshall R. Posner, David J. Santos, Hillary S. Elboim, et al. Myelomonocytic Leukemia CellsHuman Monoclonal Antibodies Reactive with Human
Updated version
http://cancerres.aacrjournals.org/content/49/7/1665
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/49/7/1665To request permission to re-use all or part of this article, use this link
on April 4, 2019. © 1989 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from