human cytomegalovirus infection induces a rapid … cytomegalovirus infection induces a rapid and...
TRANSCRIPT

of May 14, 2018.This information is current as
Cells T+Expression of NK Cell Receptors on CD8
Rapid and Sustained Change in the Human Cytomegalovirus Infection Induces a
van LierBaas, Eddy Roosnek, Ineke J. M. ten Berge and René A. W. Amber van Stijn, Ajda T. Rowshani, Sila L. Yong, Frank
http://www.jimmunol.org/content/180/7/4550doi: 10.4049/jimmunol.180.7.4550
2008; 180:4550-4560; ;J Immunol
Referenceshttp://www.jimmunol.org/content/180/7/4550.full#ref-list-1
, 21 of which you can access for free at: cites 36 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2008 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Human Cytomegalovirus Infection Induces a Rapid andSustained Change in the Expression of NK Cell Receptors onCD8� T Cells1
Amber van Stijn,2,3*† Ajda T. Rowshani,2† Sila L. Yong,2* Frank Baas,2‡ Eddy Roosnek,2§
Ineke J. M. ten Berge,2† and Rene A. W. van Lier2*
The CD8� T cell compartment of human CMV-seropositive individuals characteristically contains a high proportion of cells that expressNK cell receptors (NKRs) which may contribute to the surveillance of virus-infected cells. To test whether this enhanced expression isa direct and immediate result of CMV infection, we used DNA microarrays to analyze putative changes in the RNA expression level of39 NKRs in CMV-specific CD8� T cells of renal transplant recipients experiencing primary CMV infection. Already in the acute phaseof infection 29 NKRs were induced, of which 19 remained high 1 year after cessation of viral replication. Activating and inhibitory NKRswere induced to a similar extent. Detailed longitudinal flow cytometric analyses confirmed NKR changes at the protein level. Strikingly,a strong induction of CD94 on CD3� T cells was observed with surface expression of activating CD94dim NKG2C dimers appearingbefore inhibitory CD94bright NKG2A ones. After the acute phase of infection, the balance between inhibitory and activating receptorsdid not change. Thus, CMV infection induces a rapid and lasting change in the expression of NKRs on human CD8� T cells. TheJournal of Immunology, 2008, 180: 4550–4560.
C ytomegalovirus is a persistent � herpes virus that in theWestern world infects 60–90% of the population. In im-munocompetent individuals, primary CMV infection is
infrequently diagnosed because of the lack of prototypical symp-toms. Immunocompromised patients, however, may experience se-vere complaints due to pulmonary and gastrointestinal pathology(1). In the suppression of initial CMV replication and the mainte-nance of viral latency, both NK cells and T cells have been im-plicated (2). CMV appears to be unique among the persisting vi-ruses in that it stably augments the number of circulating restingcytolytic T cells (3). Within the CD4� pool, these cells are char-acterized by the absence of CD28 and the presence of cytolyticgranules containing perforin and granzyme B (4). For a fraction ofthese cells, specificity for CMV peptides presented by HLA classII molecules can indeed be demonstrated (5). Also, the CD8� Tcell compartment of CMV carriers contains an expanded fractionof perforin- and granzyme-expressing cells (6). These cells havebeen characterized by various phenotypic markers (for a review,see Ref. 7) and CMV peptide-specific CD8� T cells are often
contained within this distinctive subset (3). Frequently, CMV-spe-cific CD8� T cells in the latency stage lack the lymph node-hom-ing receptor CCR7 and the costimulatory receptors CD27 andCD28, but express CD57 and CD45RA. Longitudinal studies inkidney transplant recipients have indicated that the acquisition ofthis phenotype by Ag-specific CD8� T cells is a stepwise processthat continues for an extended period after primary CMV infection(8). Because these cells typically lack expression of the classicalcostimulatory receptors, it has been suggested that other surfacemolecules might contribute to their activation upon encounteringvirus-infected cells (9–11).
NK cell receptors (NKRs)4 have first been described as surfacereceptors on NK cells that bind to specific HLA class I molecules(12–15). These receptors may belong to either the Ig- or C-typelectin superfamilies and transmit, upon binding their ligands, in-hibitory or activating signals to the cell’s interior. The quality ofthe signal depends on the composition of the intracellular parts ofthe individual receptors or their associated molecules. Addition-ally, NK cells express natural cytotoxic receptors (NCR), likeNKp44 and NKp46, for which no ligands have been conclusivelydefined but that appear to function as strong activators of NK cellfunction. Apart from NK cells, expression of NKRs is also foundon the resting cytotoxic effector cells of CMV carriers (16). Inagreement with this, transsectional cohort studies have shown thatCMV infection leaves an imprint in the NKR repertoire on T cells(17). Whether NKRs on T cells regulate recognition and subse-quent elimination of CMV-infected cells is not clear. Still, certainNKRs are able to down-modulate cytotoxicity induced via TCR/CD3 or to inhibit cytolysis after cross-linking other NKRs (18–21). Triggering of the C-type lectin receptor NKG2D on Ag-spe-cific CD8� cells does not trigger cytokine production, calcium
*Department of Experimental Immunology and †Department of Internal Medicine,Division of Nephrology and ‡Department of Neurogenetics, Academic Medical Cen-ter, Amsterdam, The Netherlands; and §Division of Hematology, Geneva UniversityHospital, Geneva, Switzerland
Received for publication August 21, 2007. Accepted for publication January 30, 2008.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Grant C03.2034 from the Dutch Kidney Foundation (toA.v.S.). R.A.W.v.L. was supported by VICI Grant 918.46.606 from The NetherlandsOrganization of Scientific Research.2 A.v.S. designed research, analyzed data, and wrote the article. A.T.R. designedresearch. S.L.Y. performed research and analyzed data. F.B. contributed vital newreagents and analytical tools. E.R. contributed vital new reagents and analytical tools.I.J.t.B. designed research and revised the manuscript critically for important intellec-tual content. R.A.W.v.L. designed research and revised the manuscript critically forimportant intellectual content and wrote the article.3 Address correspondence and reprint requests to Dr. Amber van Stijn, Department ofExperimental Immunology, Academic Medical Center, Meibergdreef 9, 1105AZ Am-sterdam, The Netherlands. E-mail address: [email protected]
4 Abbreviations used in this paper: NKR, NK cell receptor; NCR, natural cytotoxicreceptor; p.i., postinfection; KIR, killer Ig-like receptor; aNKR, activating NKR;iNKR, inhibitory/inhibiting NKR.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

mobilization, nor cytotoxicity as it does in NK cells, but, instead,it augments cytotoxic and proliferative responses of T cells (9).Other NKRs may lower activation thresholds (22) and therebyfunction as costimulatory receptors for T cells. Ibegbu et al. (23)described the presence of the inhibitory C-type lectin KLRG1 incombination with CD57 on terminally differentiated memoryCD8� cells. These cells secrete cytokines but have a poor repli-cative proliferation, suggesting a modifying role of KLRG1 inthese CD8� cells.
Elegant studies in mice have revealed that NKRs are able tomodify the functional competence of virus-specific T cells in vivo.CD94-NKG2A, an inhibitory receptor of the C-type lectin class,was shown to induce anergy in a polyomavirus-specific CD8� Tcell population, contributing to the development of tumors (24).Thus, NKRs on T cells may be indispensable for an adequate bal-ance of activation and control of T cells during viral infection.
The development of CMV-specific T cell responses can longi-tudinally be followed in recipients of kidney allografts that en-counter CMV for the first time as a result of virus reactivation fromthe transplanted organ (25). We used DNA microarrays combinedwith flow cytometry to obtain a comprehensive view on the reg-ulation of NKR expression on CMV-induced CD8� T cells. Spe-cifically, we asked whether 1) NKR expression is a direct andimmediate consequence of CMV infection and 2) whether the bal-ance between activating NKRs (aNKRs) and inhibitory NKRs(iNKRs) changes between the acute and latent stage.
Materials and MethodsSubjects
Renal transplant recipients were treated with basiliximab prophylacticallyand with basic immunosuppressive drug therapy consisting of pred-nisolone, mycophenolate mofetil, and cyclosporin. Thirteen of 15 patients
experienced a primary CMV infection. Antiviral therapy consisted of gan-ciclovir. Cells from three patients were used in microarray analyses. Lon-gitudinal samples from 10others were used for detailed flow cytometry.Two CMV-seronegative renal transplant patients who did not develop pri-mary CMV infection were used to verify whether the phenotypical changeswere indeed inflicted by CMV infection rather than by the transplantationitself. Patient characteristics are listed in Table I. All patients gave writteninformed consent and the study was approved by the local medical ethicalcommittee.
Tetrameric complexes
Allophycocyanin-conjugated HLA-A2 tetramer loaded with the CMVpp65-derived NLVPTMVATV peptide and allophycocyanin-conjugatedHLA-B7 tetramer loaded with the CMV pp65-derived TPRVTGGGAMpeptide were obtained from Sanquin.
Cells
PBMC were isolated from heparinized blood using standard density gra-dient centrifugation and subsequently cryopreserved in liquid nitrogen untilthe day of analysis.
To isolate naive CD8� cells, cells were isolated from buffy coatsfrom healthy donors by a two-step procedure. After Ficoll, CD8� Tcells were isolated by CD8� microbeads (Miltenyi Biotec) and storedovernight at 4°C in 10% (v/v) serum-containing medium. CD8� T cellswere then labeled with CD27-FITC (7C9, homemade), CD45RA-RD1(Coulter), and CD8-allophycocyanin (BD Pharmingen) and FACSsorted using a FACS Aria (BD Biosciences) in naive CD8� T cells(CD8�CD45RAhighCD27high). To isolate CMV-specific CD8� effectorcells at the peak of the CMV response, PBMC were stained with HLA-DR-FITC (BD Biosciences, CD38-PE (BD Biosciences), and CD8-allo-phycocyanin (BD Pharmingen). HLA-DRhighCD38highCD8� were sortedusing a FACS Aria (BD Biosciences).
To obtain CMV-specific CD8� cells in the latency phase, PBMC ob-tained from 40 to 60 wk after transplantation (long term, 1 year postinfec-tion (p.i.)) were stained with allophycocyanin-conjugated tetramers andsubsequently allophycocyanin microbeads (Miltenyi Biotec) were used toisolate the cells. Upon reanalysis, the purified populations contained be-tween 95 and 97% tetramer-binding cells.
Table I. Patient characteristics
PatientAge
(Years)Days to FirstPositive PCR
No. of Dayswith CMV� PCR HLA Typing of Recipient (R) and Donor (D) Assays
1 69 31 219 R: A2/3 B7/39(16) DR15(2)/12(5) MicroarrayD: A2/23(9) B62(15)/49(21) DR11(5)/13(6)
2 35 45 99 R: A2/3 B7/35 DR1/15(2) MicroarrayD: A2/11 B7/8 DR4/15(2)
3 27 62 144 R: A1/3 B7/35 DR15(2)/4 MicroarrayD: A1/3 B7/35 DR4
4 35 47 68 R: A11/3 B51(5)/39(16) DR1/15(2) FACSD: A11/28 B51(5)/27 DR1/9
5 73 25 122 R: A1/2 B7/60(40) DR13(6) FACSD: A1 B63(15)/8 DR17(3)/13(6)
6 42 46 118 R: A26(10)/32(19) B44(12)/8 DR3/12(5) FACSD: A1/31(19) B39(16)/8 DR17(3)/8
7 35 25 79 R: A3/11 B7/51(5) DR15(2)/14(6) FACSD: A11/2 B51(5) DR12(5)/14(6)
8 47 28 83 R:A1/3 B7/8 DR15(2)/3 FACSD: A3/11 B7/45(12) DR15(2)/4
9 30 29 80 R: A1/2 B8/8 DR17(3) FACSD: A2 B8/41 DR3/7
10 33 NA 0 R: A3/24(9) B51(5)/55(22) DR4/13(6) FACSD: A2/24(9) B51(5)/55(22) DR13(6)/14(6)
11 49 NA 0 R: A2/3 B60(40)/13 DR13(6)/7 FACSD: A1/3 B8/13 DR3/7
12 32 96 14 R: A28 B60(40)/44(12) DR1/13(6) FACSD: A2/28 B44(12)/57(22) DR1/7
13 26 33 37 R: A3/31(19) B51(5) DR3/14(6) FACSD: A2/3 B27/61(40) DR16(2)/17(3)
14 32 61 45 R: A11/31(19) B51(5)/60(40) DR1/4 FACSD: A2/3 B13/51(5) DR1/7
15 32 49 74 R: A3/11 B7/48 DR15(2) 15(2) FACSD: A3/11 B7/18 DR15(2)/3
NA, Not applicable.
4551The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

RNA isolation, amplification, labeling, and hybridization
RNA was isolated using the nucleospin RNA isolation kit (Machery-Na-gel) according to the manufacturer’s instructions. mRNA was amplifiedusing the MessageAmp II Kit (Ambion). Labeling, hybridization, and dataextraction were performed at ServiceXS (Leiden, The Netherlands).Briefly, 100 ng of total RNA was mixed with 1 �l of T7 oligo(dT) primerin a total volume of 12 �l. Primer and template were denatured by incu-bating at 70°C for 10 min and annealed by putting the reaction tubes on ice.The first-strand reaction was performed by adding 8 �l of Reverse Tran-scription Master Mix (containing 10� First Strand buffer, RNase inhibitor,dNTP mix, and reverse transcriptase) and incubating at 42°C for 2 h. Sec-ond- strand cDNA synthesis was done by adding 63 �l of Nuclease-FreeWater, 10 �l of 10� second-strand buffer, 4 �l of dNTP mix, 2 �l of DNApolymerase, and 1 �l of RNase H and incubating at 16°C for 2 h. cDNApurification was done according to the manufacturer’s protocol (Ambion).In vitro transcription was initiated by addition of 2 �l of aaUTP Solution(50 mM), 12 �l of ATP, CTP, and GTP mix (25 mM), 3 �l of UTPSolution (50 mM), 4 �l of T7 10� reaction buffer, and 4 �l of T7 enzymemix and incubated at 37°C for 9 h. Machery-Nagel RNA Clean up mini-spin columns were used for purification of the cRNA. Dye coupling reactionwas performed using 5 �g of amino allyl aRNA in 3.33 �l, 5 �l of DMSO, and1.66 �l of NaCO3 buffer added to the monoreactive dye, prepared accordingto the manufacturer’s protocol (Amersham Biosciences). After an incubationat room temperature for 60 min, 4.5 �l of 4 M hydroxylamine was added andincubated at room temperature for 15 min. Dye-labeled aRNA was purifiedwith Machery-Nagel RNA Clean up mini-spin columns and the samples werechecked on concentration and dye incorporation on the Nanodrop ND-1000.Hybridization was performed with 600 ng of each labeled target along withfragmentation and hybridization buffer at 60°C for 17 h onto Human WholeGenome (WHG) 44K Oligo Microarrays from Agilent Technologies per themanufacturer’s protocol.
Microarray imaging and data analysis
The microarray slides were washed following the user manual instructions andscanned using the Agilent dual-laser DNA microarray scanner. Default set-tings of Agilent Feature Extraction preprocessing protocols were used to ob-tain normalized expression values from the raw scans. Exact protocol andparameter settings are described in the Agilent Feature Extraction SoftwareUser Manual 7.5 (http://chem.agilent.com/scripts/LiteraturePDF.asp?iWHID�37629). The default Agilent normalization procedure, calledLinear & Lowess, was applied. Rosetta Resolver (Rosetta Biosoftware,Seattle, WA) was used for analysis of the data.
Flow cytometric analyses
PBMC were washed in PBS containing 0.01% (w/v) NaN3 and 0.5% (w/v)BSA (PBA). PBMC (500,000) were incubated with appropriate concen-trations of tetrameric complexes in a small volume for 30 min at 4°C,protected from light. Then fluorescent-labeled conjugated mAbs (concen-trations according to the manufacturer’s instructions) were added and in-cubated for 30 min at 4°C, protected from light. For analysis of expressionof surface markers, the following mAbs were used in different combina-tions: CD56-allophycocyanin, NKG2D-allophycocyanin (BD Pharmin-gen), KIR2DS1/L1-PE (CD158a and h, clone EB6), KIR2DL2/L3/S2-PE(CD158b and j, clone GL183), NKp44-PE, NKp46-PE, NKG2A-PE (allfrom Beckman Coulter), NKG2C-PE (R&D Systems), CD27-FITC (home-made clone 3A12 for FACS analysis and homemade clone 7C9 for FACSsorting), CD27-PE (clone L128), CD3-PerCP-Cy5.1, CD94-PE, CD94-al-lophycocyanin, NKB1 (KIR3DL1)-FITC, CD38-PE, CD45RA-FITC, anti-HLA-DR-FITC (all BD Biosciences). Cells were washed in PBA and an-alyzed using a FACSCalibur flow cytometer (BD Biosciences) or FACSCanto flow cytometer (BD Biosciences) and CellQuest software (BD Bio-sciences) or FlowJo software (Tree Star).
Quantitative PCR for KIR2DL2/DL3/DS2
From patients 4 and 8 (see Table I), CD8�CD158� cells were sorted at twotime points: at the peak of the viral load (T1) and 2 mo later (T2: patient4) or 6 mo later (T2: patient 8) when the virus was not detectable. RNA wasisolated and quantitative PCR was performed on cDNA with the Light-Cycler System (Roche Diagnostics) in microcapillary tubes with a Quan-tiTect SYBR green PCR kit solution (Qiagen). After a 15-min denaturationstep at 95°C, 50 PCR cycles of 15 s at 94°C, 30 s at 58°C, 45 s at 72°C,and 5 s at 79°C were performed. To confirm the purity and specificity ofthe reaction, a melting curve analysis was performed at the end of the PCRby slowly increasing (0.1°C/s) the temperature of the reaction from 65 to95°C. A standard curve was established with a dilution of purified PCRproduct from 10 to 107 molecules. GAPDH was used as an internal
FIGURE 1. Confirmation of the microarray results by FACS analyses ofselected CD8� T cell proteins that are known to be regulated during a primaryCMV infection. Graphs show the RNA fold changes found by microarrayanalysis (as compared with naive CD8� cells) for the genes indicated. Theprotein confirmation of the selected genes is performed by flow cytometry.Density plots are gated on CD8� T cells. Depicted on the x-axis is the pp65CMV-specific tetramer staining, on the y-axis is the protein as indicated. peak,Peak of CMV infection, 1 year; �1 year after CMV infection. Quadrantsdepicted as percentages of total CD8� T cells. Shown is one representativepatient (patient 15).
4552 NKRs ON CD8� T CELLS DURING PRIMARY CMV
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

reference. The killer Ig-like receptor (KIR) primers used were either the sameas those published by Uhrberg et al. (26) or one nucleotide modificationsthereof: KIR2DS2, 5�-CTGCACAGAGAGGGGAAGTA-3�; reverse, 5�-ACGCTCTCTCCTGCCAA-3�; KIR2DL2, 5�-CATGATGGGGTCTCCA
AA-3�; reverse, 5�-GCCCTGCAGAGAACCTACA-3�; KIR2DL3, 5�-CCACTGAACCAAGCTCCG-3�; reverse, 5�-GCAGGAGACAACTTTGGATCA-3�; and GAPDH, 5�-GGACCTGACCTGCCGTCTAG-3�; reverse,5�-CCACCACCCTGTTGCTGTAG-3�.
Table II. Changes in NKR during primary CMVa
Alternative Name Naiveb Peakc 1 Yeard mAbse
aNKRMHC class I specific
KIRsKIR2DS2 CD158j 1 3,6 1,4 GL183f
KIR2DS4 CD158l 1,2 6,4 1,9KIR3DL5A CD158f 1,1 2,3 1,1
Leukocyte Ig-likeLILRA1 LIR6/CD85l 0,9 10,6 13,4LILRA3 ILT6/LIR4/CD85e 0,9 2 2,9LILRA4 ILT7 0,9 1,3 1,3LILRA5 CD85f 0,9 3,7 2,3
Non-MHC class I specificNCRs
NCR1 NKp46 0,9 47,4 39,1 NKp46NCR2 NKp44 1,1 0,9 0,9 NKp44NCR3 NKp30 1,1 1,4 0,9
C-type lectin receptorsKLRC2 NKG2C 1 9,8 7,8 NKG2CKLRK1 NKG2D 1 3,6 2,3 NKG2DKLRC3 NKG2E 1 2,8 1,2KLRC4 NKG2F 1 2,2 1,9KLRF1 0,9 5 12,8CLEC2B AICL 1 3,8 2,2
OtherCD16 FcyIIIa 0,9 8,5 14CD244 2b4 1,1 13,5 10,6NKG7 1 8,5 14,9
iNKRMHC class I specific
KIRsKIR2DL1 CD158a 0,8 6 1,6 EB6g
KIR2DL2 CD158b1 1 5 1,5 GL183f
KIR2DL4 CD158d 1 2,9 1KIR3DL1 CD158e1/NKB1 1 2,9 1,1 NKB1KIR3DL2 CD158k 1,1 7,1 1,4KIR3DL3 CD158z 1 1,1 1,3
Leukocyte Ig-like receptorsLILR1 0,9 2,3 2,4LILRB1 ILT2/LIR1/CD85j 1 28,3 61,4LILRB2 ILT4/LIR2/CD85d 1,6 13,9 17,9LILRB3 ILT5/LIR3/CD85a 1 2,1 2,2LILRB4 ILT3/LIR5/CD85k 1 0,9 1,1LILRB5 LIR8/CB85c 0,9 1,4 1
Non-MHC class I specificNCRs
NoneC-type lectin receptors
KLRA1 1 2,2 2,4KLRB1 CD161 1 9,3 17,1KLRC1 NKG2A/CD159A 1,1 9 8 NKG2AKLRG1 MAFAL 0,9 11,5 14,5
OtherLAIR1 1 1,1 2,6LAIR2 1,2 11,8 34,4
Either aNKR or iNKRLeukocyte Ig-like receptors
LILRP2 ILT10/CD85m 1 1,5 2,1C-type lectin receptors
KLRD1 CD94 1 36,5 38,9 CD94
a Bold numbers represent fold changes of significantly changed genes according to the criteria described in Materials and Methods.b Naive: CD8�CD45�CD27�.c Peak: CD8�CD38�HLA-DR�.d One year: CMV-specific tetramer pp65� cells.e mAbs used in FACS analyses.f Clone GL183 detects KIR2DS2/2DL2/2DL3.g Clone EB6 detects KIR2DS1/L1.
4553The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Statistics
Trend plotting was performed in a Rosetta Resolver with a fold changecutoff of �2 and a one-way ANOVA test with a p value cutoff of �0.05with the Benjamini and Hochberg false-discovery rate correction to selectsequences with differences across the different cell populations. The paired,nonparametric Wilcoxon-signed rank test was used for analysis of genesthat differ between different cell populations. The nonparametric Mann-Whitney U test was used for analysis of differences between aNKRs andiNKRs. Linear regression analysis was performed to show the relationshipbetween NKG2A and CD94bright expression. Significance was indicated byp � 0.05.
ResultsCMV infection induces a rapid and persistent induction of NKRexpression
To analyze regulation of NKR gene expression by CMV in hu-mans, we took advantage of the fact that dynamic change in virus-specific T cell populations can carefully be tracked in CMV-sero-negative renal transplant recipients that experience a primaryCMV infection due to the transplantation of a CMV-carrying do-nor kidney. As described earlier (8), circulating CMV-tetramer-binding CD8� cells emerge �2–4 wk after the first positive CMVPCR, but their number is too low to obtain enough cells for mi-croarray analyses. To be able to include enough cells eligible foranalysis, we isolated all virus-activated CD8� T cells by sortingCD8�HLA-DR�CD38� T cells. We previously showed that theseactivated lymphocytes appear at a high frequency early in the anti-CMV response and that at this point all CMV-tetramer� cells arecontained within this population (8). To obtain virus-specific cellsin the latency stage, CMV-specific pp65 tetramer� cells were iso-
lated from the same patients between 40 and 60 wk after the pri-mary response. At this time point, no CMV could be detected inthe blood by sensitive PCR. Because we used the two-color mi-croarray Agilent WHG chips to perform RNA expression profiling,the expression level of each gene in a specific population is givenas a ratio relative to the expression level of naive CD8� cellsobtained from healthy donors (subset CD8� cells/naive CD8�
cells).Microarray analysis revealed a number of changes in gene ex-
pression levels of several molecules (Fig. 1). The changes inmRNA expression levels of Ki-67, CD28, CCR7, CD27, CD127,perforin, granzyme B, and IFN-� were confirmed at the proteinlevel by FACS analyses; Ki-67, a proliferation marker, was onlyelevated during the peak of viral replication. In contrast, CD28,CD27, CD127, and CCR7 expression was low on virus-specific Tcells. Finally, the cytolytic proteins granzyme B and perforin andthe antiviral cytokine IFN-� were high in virus-specific cells.
On the Agilent’s WHG array, 39 NKR-related sequences werepresent. Changes in their relative expression levels are representedin Table II and Fig. 2. Three distinct expression profiles could bedistinguished. First, the expression of only eight NKRs such asLILRA4 or KIR3DL3 did not show significant changes (Table II).Second, a limited number of receptors, including KIR2DS2,KIR2DS4, and KIR3DL2, were up-regulated during the peak ofthe CMV infection but returned to baseline levels in the latencystage (Fig. 2A). However, the majority of NKR genes were in-duced early in the response and remained high in the follow-up.Two NKRs (LAIR1 and LILRP2) were only induced in the latentstate (Table II).
Remarkably, KIRs, that bind epitopes on HLA-C and HLA-Bclass I alleles were almost exclusively present in the second pro-file, e.g., induced during the acute phase of the viral infection, butabsent in CMV pp65-specific tetramer-positive CD8� cells in therenal transplant patients 1 year p.i. The other classes of receptorsdid not show predominance for a particular expression profile.
FIGURE 2. Two expression profiles of significantly regulated NKRsupon primary CMV infection. Trend plots are made with the Rosetta Re-solver software; bold lines connect significantly changed NKRs. Back-ground lines represent all gene expression data of the microarray. A, Tran-sient up-regulation during acute phase of infection. B, Up-regulation duringthe acute phase and �40 wk p.i.
FIGURE 3. Acquisition and maintenance of iNKRs and aNKRs duringprimary CMV infection detected by microarray. A, Cumulative fold changewas calculated by summing all of the fold changes of genes that change �2at the peak and/or 1 year p.i. observed from aNKRs (n � 15) or iNKRs(n � 13); all NKRs (n � 31, including unknown NKRs). �, p � 0.001;Wilcoxon-signed rank test. Multiple observations and genes that did notchange � (n � 8) were ignored. B, Mean fold changes and SD werecalculated. #, Mann-Whitney U test.
4554 NKRs ON CD8� T CELLS DURING PRIMARY CMV
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 4. Induction of NKRs on CD3� cells during primary CMV infection detected by FACS analysis. Dot plots are gated on CD3� T cells in whichthe CD27 marker is plotted against the staining for EB6 (KIR2DS1/KIR2DL1), GL183 (KIR2DL2, KIR2DL3, KIR2DS2), NKB1 (KIR3DL1), NKG2D,NKp44, NKp46, or CD94. Quadrant percentages depicted as percentages from CD3� cells. Shown is one representative patient (patient 4). PCR viral loadis given as copies per ml. Time is defined as weeks from the peak of the PCR viral load. All stainings are CD27-FITC, NKR-PE, CD3-PerCP, andCD56-allophycocyanin, except in the NKB1 staining, which is FITC labeled and CD27-PE is used.
4555The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
The balance between inhibitory and activating receptors doesnot change in the course of infection
To visualize the CMV-induced changes on NKR gene expression inCD8� T cells, we calculated a cumulative fold change and the meanfold change of all receptors. When all NKR genes were grouped to-gether, a strong increase of NKR expression was observed both in theacute and latency stages (Fig. 3A). Of the 39 NKR genes on the chip,19 are activating, 18 are inhibiting, while 2 have yet an unknownfunction. CD94 forms heterodimers with either inhibiting NKG2A oractivating NKG2C. Separating the NKRs in aNKRs and iNKRs re-vealed a similar expression pattern for both types of receptors (Fig.3B). Since a small change level in RNA can have a major impact atthe protein level, we also counted the number of aNKRs and iNKRsthat changed during infection. This showed that there was no selectiveinduction of either aNKRs or iNKRs at a particular stage; at the peakof CMV, 16 aNKRs, 12 iNKRs and CD94 were elevated, whereas 11aNKRs, 8 iNKRs, and 2 “unknown function” NKRs (LILRP2 andCD94) were up-regulated after �1 year of infection. Thus, the arraydata did not provide any indication that the balance between activatingand inhibitory receptor genes differs between virus-specific cells in theacute vs the latent stage. It should be mentioned that in the acute phaseCMV-specific CD8� T cells containing activated T cells were stud-ied, while in the latent phase pp65-tetramer� CMV-specific cells werestudied. This could potentially influence the results, although it is not
expected to radically change the results since tetramer� T cells alsohave a CD38�HLA-DR� phenotype in the acute phase.
Individual NKRs show different kinetics in cell surfaceexpression
To obtain a more thorough insight into the detailed kinetics ofNKR expression, four-color flow cytometric analyses were per-formed. Due to the limited number of fluorescence channels, wechose to analyze the NKR phenotype in combination with CD3,CD56, and CD27 markers. CD27 was included to monitor thematuration of virus-specific cells by the gradual loss of CD27. Thecombination of CD3 and CD56 proved to be a good combinationto distinguish between virus-specific T cells, including CD4� andCD8� T cells, and NK cells.
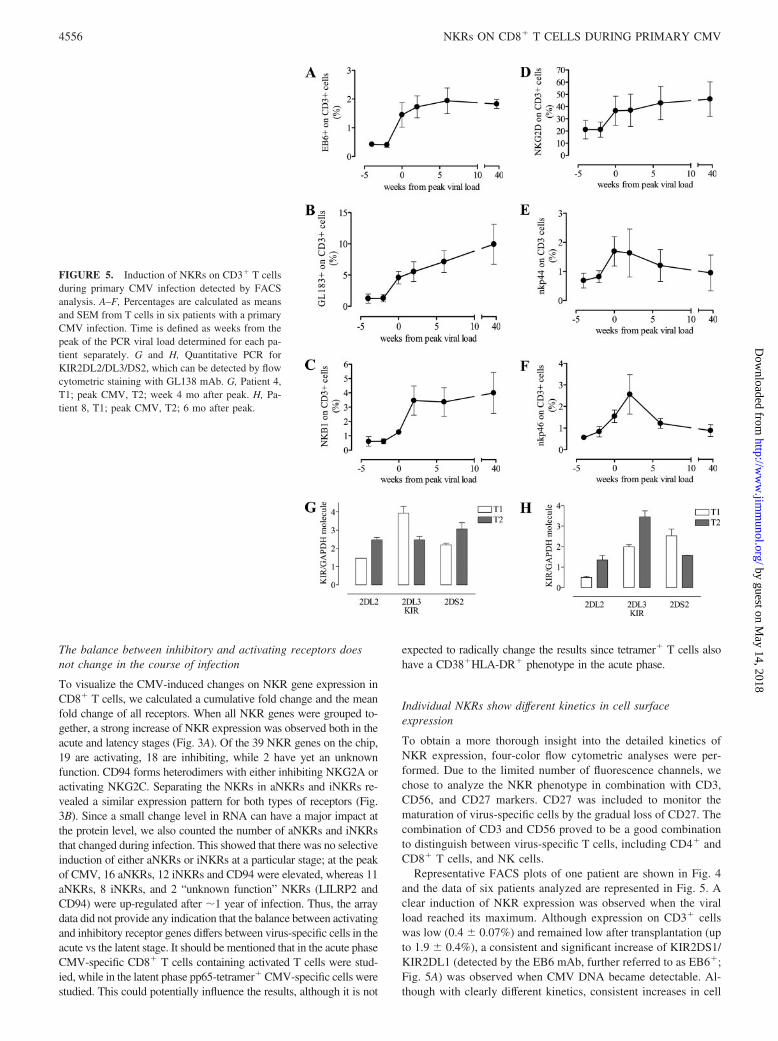
Representative FACS plots of one patient are shown in Fig. 4and the data of six patients analyzed are represented in Fig. 5. Aclear induction of NKR expression was observed when the viralload reached its maximum. Although expression on CD3� cellswas low (0.4 � 0.07%) and remained low after transplantation (upto 1.9 � 0.4%), a consistent and significant increase of KIR2DS1/KIR2DL1 (detected by the EB6 mAb, further referred to as EB6�;Fig. 5A) was observed when CMV DNA became detectable. Al-though with clearly different kinetics, consistent increases in cell
FIGURE 5. Induction of NKRs on CD3� T cellsduring primary CMV infection detected by FACSanalysis. A–F, Percentages are calculated as meansand SEM from T cells in six patients with a primaryCMV infection. Time is defined as weeks from thepeak of the PCR viral load determined for each pa-tient separately. G and H, Quantitative PCR forKIR2DL2/DL3/DS2, which can be detected by flowcytometric staining with GL138 mAb. G, Patient 4,T1; peak CMV, T2; week 4 mo after peak. H, Pa-tient 8, T1; peak CMV, T2; 6 mo after peak.
4556 NKRs ON CD8� T CELLS DURING PRIMARY CMV
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

surface expression were also found for KIR2DS2/2DL2/2DL3 (de-tected by the GL183 mAb, further referred to GL183�; Fig. 5B),KIR3DL1 (NKB1; Fig. 5C), NKG2D (Fig. 5D), the NCRs NKp44
and NKp46 (Fig. 5, E and F), and CD94 (Fig. 6A). In two CMV-seronegative individuals that received a CMV-negative donor kid-ney, no changes in NKR expression were observed (data not
FIGURE 6. CD94dim, CD94bright, NKG2A,and NKG2C on T cells during primary CMV.Total CD94 (A), CD94dim (B), and CD94bright
(C) expression on CD3� T cells. D, Linear re-gression analysis was performed betweenCD94bright and NKG2A expression on CD3�
cells (R2 � 0.98). NKG2A (E) and NKG2C (F)expression on CD8� cells. Shown is one repre-sentative patient of three patients. NKG2A (G)and NKG2C (H) on CMV-specific CD8� pp65-tetramer� cells. Shown is one representative pa-tient (patient 4). Dot plots are gated on vitalcells. Numbers indicate the percentages withinthe corresponding quadrants from the gated pop-ulation. Time is defined as weeks from the peakof the PCR viral load determined for each pa-tient separately. PreTx, Pretransplantation; n.a.,not applicable.
4557The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

shown), showing that the alterations in NKR expression were in-deed a consequence of the CMV infection and not related to thetransplantation procedure. In the follow-up periods of �1 year, wedid not observe a specific association of NKR with CD27 expres-sion, indicating that induction of these molecules is independentfrom other differentiation steps of human CMV-specific CD8� Tcells (Fig. 4).
We investigated whether the presence of KIR ligands corre-sponded to the elevation of specific KIRs during the primary CMVinfection. No evidence for this assumption was found, becausepatients 5, 8, and 9 nor their transplanted kidneys expressed HLA-Bw4, the ligand for NKB1, but still NKB1 was strongly induced inthese individuals (data not shown).
The increase in GL183� was very prominent, as at the end ofthe follow-up up to 13% of the circulating CD3� pool expressedthis marker. Because the Ab GL183 recognizes both activating(KIR2DS2) and inhibitory members (KIR2DL2 and DL3) of theKIR family, we analyzed, by quantitative PCR, whether CMV in-duced the selective outgrowth of a monoclonal population express-ing only one KIR species. CD8�GL183� cells from two patients(patients 4 and 8) at two time points were obtained and in thesecells all three transcripts could be detected with values that are inthe normal range of KIR-purified cells (Fig. 5G). This shows thatthe strongly expanded GL183� T cell population does not repre-sent the outgrowth of a monoclonal KIR� population.
CD94 dimerizes with different kinetics with NKG2C and NKG2A
The increase in CD94 expression was quite prominent on CD3�
T cells (Figs. 4 and Fig. 6A), and we therefore analyzed in moredetail whether the molecule would operate in transmitting ac-tivating and/or inhibiting signals. CD94 can only exert its sig-naling function when heterodimerized with either NKG2A (in-hibiting dimer) or NKG2C (activating dimer). The intensity ofCD94 expression is, as reported by Arlettaz et al. (27), high inconjunction with NKG2A but low when bound to NKG2C. Dur-ing the first weeks of the CMV response, predominantlyCD94dim cells appeared in the circulation (Fig. 6B) and some-what later CD94bright cells became detectable (Fig. 6C). As ex-pected (27), we found a correlation between the CD94bright andNKG2A expression in our samples (Fig. 6D). Because the ma-jority of CD94-positive T cells are CTLs, we continued theanalysis on CD8� T cells. Already before CMV infection,NKG2C is present on a small subset of CD8� cells, whereasNKG2A is almost absent on CD8� cells. Confirming the dif-ferential rise in the dimerization pattern of CD94 with NKG2Cand NKG2A, we found that a NKG2C increase on CD8� cellscould be detected shortly after the start of the infection (2 wkbefore CMV peak; Fig. 6F), whereas the increase of NKG2Astarted later in the response (Fig. 6E). To analyze the expressionof these dimers on established virus-specific cells, pp65 tetram-ers were used to analyze cells of a HLA-A2� individual. Theinhibitory CD94:NKG2A dimer was found on the majority ofthe CMV-specific tetramer� cells irrespective of the stage ofinfection (Fig. 6G). In marked contrast, the activating NKG2Cdimer was induced on the CD3� cells but conspicuously absentfrom the tetramer-binding cells (Fig. 6H). The absence ofCD94/NKG2C on cells with proven virus-specificity is remi-niscent of earlier findings showing that tetramer-binding cellsalso lack EB6 and GL183 (CD158a,h and CD158b, j expres-sion) (16).
DiscussionIn this study, we show the impact of a primary CMV response onNKR virus-induced CD8� T lymphocytes. Using a microarray ap-
proach, we found that mRNA expression levels of different mem-bers of all NKR families, including KIRs, leukocyte Ig-like recep-tors, NCRs, and C-type lectin receptors in virus-induced CD8� Tcells, increased during the infection. Many of these genes re-mained increased up to 50 wk after the peak of the initial viralburst when the virus load had long dropped below detection bysensitive PCR.
Changes in KIR2DL1/S1 (EB6), KIR2DL2/S2 (GL183), andKIR3DL1 (NKB1) mRNA were observed in the acutely acti-vated CD8�HLA-DR�CD38� cells, but expression levels inthe pp65 tetramer CD8� cells obtained during latency werecomparable to those of naive CD8� T cells. This observation isin agreement with previous observations (16, 28) where expres-sion of these KIRs was found on the CD8�CD45RA�CD27�
effector-type cells typical for CMV- infected individualspresent during CMV infection but absent on the pp65 tetramer-binding cells. The reason for this difference is unclear.CD8�CD45RA�CD27� only expand to high numbers afterCMV infection (6) and it therefore seems likely that manyCMV-reactive cells are contained within this subset. Elegantstudies by Picker et al. (29) have shown that the T cell responseto CMV is very robust and is directed against many peptides ofthe virus. Studies until now have mainly focused on pp65-spe-cific T cells and therefore it is possible that CMV-specificCD8� T cells, reactive toward epitopes generated from otherviral proteins, will have different phenotypes with regard to, forexample, KIR.
NKR expression on CD8� T cells is not associated with otherlatent viruses such as EBV and HIV (28), suggesting that expres-sion of these molecules has a particular function in the anti-CMVresponse. Since CMV-specific T cells generally reach later differ-entiation stages than T cells specific for other persisting viruses(3), one might assume that the expression of NKRs is an integralpart of the late differentiation phenotype. However, our array dataappear to argue against this notion. Up-regulation of a vast numberof NKRs already takes place during the early phase of the responsewhen cells have an activated early differentiation phenotype (16).Rather, our data suggest that the expression of NKRs as a conse-quence of CMV infection is an early and adaptive response of theCD8� effector cell pool to the virus. Whether specific viral prod-ucts and/or the virus-induced modulation of HLA expression havecreated an evolutionary necessity for this broad expression of theNKR repertoire will have to be resolved.
The early up-regulation of the NCR NKp44 was found by flowcytometry but was not revealed by the microarrays. This is perhapsnot surprising since the up-regulation of NKp44 in CD3� cells istransient and low, reaching a maximum at the peak of the viral loadof only mean 1.7%. Two weeks after the peak of the viral load, thefrequency of NKp44-expressing cells is already decreasing in mostpatients. It is possible that the patients selected for the RNA iso-lation had too low levels to be detectable by microarray analysis.In any case, it suggests that NKp44, as in NK cells (30, 31), ispreferentially expressed on the mitogenically activated cells thatcan only be found early in infection (8).
CD94 was found to be expressed on both CD3� cells (pro-tein) and CD8� CMV-specific cells (RNA). CD94 forms het-erodimers with NKG2x proteins (where x can be A, B, C, E, orH). The NKG2A/B/CD94 is an inhibitory receptor, whereas theNKG2C/CD94 and NKG2E/H/CD94 are activating receptors(32). Arlettaz et al. (27) reported that CD94bright expression isassociated with NKG2A and CD94dim is associated withNKG2C, although also other NKG2 proteins may dimerize withCD94. Our observation that CD94dim was primarily enhanced
4558 NKRs ON CD8� T CELLS DURING PRIMARY CMV
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

during the acute phase of the infection, whereas CD94bright ac-counted for the latent phase was confirmed by the quick rise inNKG2C followed by robust expansion of NKG2A which con-tinued to expand for at least 40 wk after the peak of the viralload. A strict correlation between microarray data and surfaceexpression of NKG2C was not apparent in our analyses.
Indeed, it has been reported (27, 33) that if NKG2A is ex-pressed, no surface NKG2C expression could be observed evenwith abundant transcription of NKG2C. This discrepancy might bedue to different affinities of CD94 for binding partners and a pref-erence to dimerize with NKG2A. Also in our analyses, whereasNKG2A was expressed at the cell surface of tetramer� cells, noNKG2C expression could be detected in apparent contrast with thehigh NKG2C mRNA content detected in the microarray analysis.
The coordinated changes of aNKRs and iNKRs on recentlyprimed and late differentiated CD8� T cells at different timepoints during the viral infection suggest that NKRs contributeto the fine tuning of antiviral effector functions, keeping thedelicate balance between efficient destruction of virus-infectedcells and loss of organ function due to immunopathology. Thisidea is in line with several observations on the function of KIRsand leukocyte Ig-like receptors showing that they can, for ex-ample, down-regulate killing efficiency (34), inhibit cytokineproduction (35), and weaken proliferative potential (28) ofCD8� T cell clones. Henel et al. (36) recently demonstrated thatwith low Ag concentrations, KIR expression on memory CD4�
T cells uncoupled granule release from gene activation, reason-ing that a general inflammation is inhibited to circumventpathological events. In view of this, it can be reasoned that inimmune reactions against persistent viruses, like CMV, a neg-ative regulation of general inflammation is essential to preventdamage by continued activation of T lymphocytes.
Cohort studies have indicated that the NKR profile on NK andT cells is imprinted by CMV (17). We here have demonstrated astrong induction of NKRs on T cells as a direct result of CMVinfection. These NKRs might provide essential control mecha-nisms for the activation of T cells by regulating costimulatorysignals, dampening T cell activation when viral loads diminishes,and controlling harmful activation to the host during latent viralinfections.
AcknowledgmentsWe thank Dr. Perry Moerland for assistance with the bioinformatics andstatistics of the microarray analysis.
DisclosuresThe authors have no financial conflict of interest.
References1. Rowshani, A. T., F. J. Bemelman, E. M. van Leeuwen, R. A. van Lier, and
I. ten Berge. 2005. Clinical and immunologic aspects of cytomegalovirus infec-tion in solid organ transplant recipients. Transplantation 79: 381–386.
2. Polic, B., H. Hengel, A. Krmpotic, J. Trgovcich, I. Pavic, P. Luccaronin,S. Jonjic, and U. H. Koszinowski. 1998. Hierarchical and redundant lymphocytesubset control precludes cytomegalovirus replication during latent infection.J. Exp. Med. 188: 1047–1054.
3. Appay, V., P. R. Dunbar, M. Callan, P. Klenerman, G. M. Gillespie, L. Papagno,G. S. Ogg, A. King, F. Lechner, C. A. Spina, et al. 2002. Memory CD8� T cellsvary in differentiation phenotype in different persistent virus infections. Nat. Med.8: 379–385.
4. Appay, V., J. J. Zaunders, L. Papagno, J. Sutton, A. Jaramillo, A. Waters,P. Easterbrook, P. Grey, D. Smith, A. J. McMichael, et al. 2002. Characterizationof CD4� CTLs ex vivo. J. Immunol. 168: 5954–5958.
5. van Leeuwen, E. M., E. B. Remmerswaal, M. T. Vossen, A. T. Rowshani,P. M. Wertheim-van Dillen, R. A. van Lier, and I. ten Berge. 2004. Emergenceof a CD4�CD28�granzyme B�, cytomegalovirus-specific T cell subset afterrecovery of primary cytomegalovirus infection. J. Immunol. 173: 1834–1841.
6. Kuijpers, T. W., M. T. Vossen, M. R. Gent, J. C. Davin, M. T. Roos,P. M. Wertheim-van Dillen, J. F. Weel, P. A. Baars, and R. A. van Lier. 2003.
Frequencies of circulating cytolytic, CD45RA�CD27�, CD8� T lymphocytesdepend on infection with CMV. J. Immunol. 170: 4342–4348.
7. van Lier, R. A., I. ten Berge, and L. E. Gamadia. 2003. Human CD8� T-celldifferentiation in response to viruses. Nat. Rev. Immunol. 3: 931–939.
8. Gamadia, L. E., E. B. Remmerswaal, J. F. Weel, F. Bemelman, R. A. van Lier,and I. ten Berge. 2003. Primary immune responses to human CMV: a critical rolefor IFN-�-producing CD4� T cells in protection against CMV disease. Blood101: 2686–2692.
9. Groh, V., R. Rhinehart, J. Randolph-Habecker, M. S. Topp, S. R. Riddell, andT. Spies. 2001. Costimulation of CD8�� T cells by NKG2D via engagement byMIC induced on virus-infected cells. Nat. Immunol. 2: 255–260.
10. Mandelboim, O., D. M. Davis, H. T. Reyburn, M. Vales-Gomez, E. G. Sheu,L. Pazmany, and J. L. Strominger. 1996. Enhancement of class II-restricted T cellresponses by costimulatory NK receptors for class I MHC proteins. Science 274:2097–2100.
11. Namekawa, T., M. R. Snyder, J. H. Yen, B. E. Goehring, P. J. Leibson,C. M. Weyand, and J. J. Goronzy. 2000. Killer cell activating receptors functionas costimulatory molecules on CD4�CD28null T cells clonally expanded in rheu-matoid arthritis. J. Immunol. 165: 1138–1145.
12. Moretta, A., C. Bottino, D. Pende, G. Tripodi, G. Tambussi, O. Viale, A. Orengo,M. Barbaresi, A. Merli, E. Ciccone, et al. 1990. Identification of four subsets ofhuman CD3�CD16� natural killer (NK) cells by the expression of clonally dis-tributed functional surface molecules: correlation between subset assignment ofNK clones and ability to mediate specific alloantigen recognition. J. Exp. Med.172: 1589–1598.
13. Moretta, A., M. Vitale, C. Bottino, A. M. Orengo, L. Morelli, R. Augugliaro,M. Barbaresi, E. Ciccone, and L. Moretta. 1993. P58 molecules as putative re-ceptors for major histocompatibility complex (MHC) class I molecules in humannatural killer (NK) cells: anti-p58 antibodies reconstitute lysis of MHC classI-protected cells in NK clones displaying different specificities. J. Exp. Med. 178:597–604.
14. Vitale, M., S. Sivori, D. Pende, L. Moretta, and A. Moretta. 1995. Coexpressionof two functionally independent p58 inhibitory receptors in human natural killercell clones results in the inability to kill all normal allogeneic target cells. Proc.Natl. Acad. Sci. USA 92: 3536–3540.
15. Bottino, C., M. Vitale, L. Olcese, S. Sivori, L. Morelli, R. Augugliaro,E. Ciccone, L. Moretta, and A. Moretta. 1994. The human natural killer cellreceptor for major histocompatibility complex class I molecules: surface modu-lation of p58 molecules and their linkage to CD3 � chain, Fc�RI � chain, and thep56lck kinase. Eur. J. Immunol. 24: 2527–2534.
16. Gamadia, L. E., R. J. Rentenaar, P. A. Baars, E. B. Remmerswaal, S. Surachno,J. F. Weel, M. Toebes, T. N. Schumacher, I. ten Berge, and R. A. van Lier. 2001.Differentiation of cytomegalovirus-specific CD8� T cells in healthy and immu-nosuppressed virus carriers. Blood 98: 754–761.
17. Guma, M., A. Angulo, C. Vilches, N. Gomez-Lozano, N. Malats, andM. Lopez-Botet. 2004. Imprint of human cytomegalovirus infection on the NKcell receptor repertoire. Blood 104: 3664–3671.
18. Ferrini, S., A. Cambiaggi, R. Meazza, S. Sforzini, S. Marciano, M. C. Mingari,and L. Moretta. 1994. T cell clones expressing the natural killer cell-related p58receptor molecule display heterogeneity in phenotypic properties and p58 func-tion. Eur. J. Immunol. 24: 2294–2298.
19. Mingari, M. C., C. Vitale, A. Cambiaggi, F. Schiavetti, G. Melioli, S. Ferrini, andA. Poggi. 1995. Cytolytic T lymphocytes displaying natural killer (NK)-like ac-tivity: expression of NK-related functional receptors for HLA class I molecules(p58 and CD94) and inhibitory effect on the TCR-mediated target cell lysis orlymphokine production. Int. Immunol. 7: 697–703.
20. Mingari, M. C., F. Schiavetti, M. Ponte, C. Vitale, E. Maggi, S. Romagnani,J. Demarest, G. Pantaleo, A. S. Fauci, and L. Moretta. 1996. Human CD8� Tlymphocyte subsets that express HLA class I-specific inhibitory receptors repre-sent oligoclonally or monoclonally expanded cell populations. Proc. Natl. Acad.Sci. USA 93: 12433–12438.
21. Mingari, M. C., M. Ponte, C. Cantoni, C. Vitale, F. Schiavetti, S. Bertone,R. Bellomo, A. T. Cappai, and R. Biassoni. 1997. HLA-class I-specific inhibitoryreceptors in human cytolytic T lymphocytes: molecular characterization, distri-bution in lymphoid tissues and co-expression by individual T cells. Int. Immunol.9: 485–491.
22. Jamieson, A. M., A. Diefenbach, C. W. McMahon, N. Xiong, J. R. Carlyle, andD. H. Raulet. 2002. The role of the NKG2D immunoreceptor in immune cellactivation and natural killing. Immunity 17: 19–29.
23. Ibegbu, C. C., Y. X. Xu, W. Harris, D. Maggio, J. D. Miller, and A. P. Kourtis.2005. Expression of killer cell lectin-like receptor G1 on antigen-specific humanCD8� T lymphocytes during active, latent, and resolved infection and its relationwith CD57. J. Immunol. 174: 6088–6094.
24. Moser, J. M., J. Gibbs, P. E. Jensen, and A. E. Lukacher. 2002. CD94-NKG2Areceptors regulate antiviral CD8� T cell responses. Nat. Immunol. 3: 189–195.
25. Snydman, D. R., B. G. Werner, B. Heinze-Lacey, V. P. Berardi, N. L. Tilney,R. L. Kirkman, E. L. Milford, S. I. Cho, H. L. Bush, Jr., A. S. Levey, et al. 1987.Use of cytomegalovirus immune globulin to prevent cytomegalovirus disease inrenal-transplant recipients. N. Engl. J. Med. 317: 1049–1054.
26. Uhrberg, M., N. M. Valiante, B. P. Shum, H. G. Shilling, K. Lienert-Weidenbach,B. Corliss, D. Tyan, L. L. Lanier, and P. Parham. 1997. Human diversity in killercell inhibitory receptor genes. Immunity 7: 753–763.
27. Arlettaz, L., J. Villard, C. de Rham, S. Degermann, B. Chapuis, B. Huard, andE. Roosnek. 2004. Activating CD94:NKG2C and inhibitory CD94:NKG2A re-ceptors are expressed by distinct subsets of committed CD8� TCR �� lympho-cytes. Eur. J. Immunol. 34: 3456–3464.
4559The Journal of Immunology
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

28. Anfossi, N., J. M. Doisne, M. A. Peyrat, S. Ugolini, O. Bonnaud, D. Bossy,V. Pitard, P. Merville, J. F. Moreau, J. F. Delfraissy, et al. 2004. Coordinatedexpression of Ig-like inhibitory MHC class I receptors and acquisition of cyto-toxic function in human CD8� T cells. J. Immunol. 173: 7223–7229.
29. Waldrop, S. L., C. J. Pitcher, D. M. Peterson, V. C. Maino, and L. J. Picker. 1997.Determination of antigen-specific memory/effector CD4� T cell frequencies byflow cytometry: evidence for a novel, antigen-specific homeostatic mechanism inHIV-associated immunodeficiency. J. Clin. Invest 99: 1739–1750.
30. Cantoni, C., C. Bottino, M. Vitale, A. Pessino, R. Augugliaro, A. Malaspina,S. Parolini, L. Moretta, A. Moretta, and R. Biassoni. 1999. NKp44, a triggeringreceptor involved in tumor cell lysis by activated human natural killer cells, is anovel member of the immunoglobulin superfamily. J. Exp. Med. 189: 787–796.
31. Vitale, M., C. Bottino, S. Sivori, L. Sanseverino, R. Castriconi, E. Marcenaro,R. Augugliaro, L. Moretta, and A. Moretta. 1998. NKp44, a novel triggeringsurface molecule specifically expressed by activated natural killer cells, is in-volved in non-major histocompatibility complex-restricted tumor cell lysis.J. Exp. Med. 187: 2065–2072.
32. Kaiser, B. K., F. Barahmand-Pour, W. Paulsene, S. Medley, D. E. Geraghty, andR. K. Strong. 2005. Interactions between NKG2x immunoreceptors and HLA-E
ligands display overlapping affinities and thermodynamics. J. Immunol. 174:2878–2884.
33. Noppen, C., C. Schaefer, P. Zajac, A. Schutz, T. Kocher, J. Kloth, M. Heberer,M. Colonna, G. De Libero, and G. C. Spagnoli. 1998. C-type lectin-like receptorsin peptide-specific HLA class I-restricted cytotoxic T lymphocytes: differentialexpression and modulation of effector functions in clones sharing identical TCRstructure and epitope specificity. Eur. J. Immunol. 28: 1134–1142.
34. Saverino, D., M. Fabbi, F. Ghiotto, A. Merlo, S. Bruno, D. Zarcone, C. Tenca,M. Tiso, G. Santoro, G. Anastasi, et al. 2000. The CD85/LIR-1/ILT2 inhibitoryreceptor is expressed by all human T lymphocytes and down-regulates their func-tions. J. Immunol. 165: 3742–3775.
35. D’Andrea, A., C. Chang, J. H. Phillips, and L. L. Lanier. 1996. Regulation of Tcell lymphokine production by killer cell inhibitory receptor recognition of selfHLA class I alleles. J. Exp. Med. 184: 789–794.
36. Henel, G., K. Singh, D. Cui, S. Pryshchep, W. W. Lee, C. M. Weyand, andJ. J. Goronzy. 2006. Uncoupling of T-cell effector functions by inhibitory killerimmunoglobulin-like receptors. Blood 107: 4449–4457.
4560 NKRs ON CD8� T CELLS DURING PRIMARY CMV
by guest on May 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from