hox homeotic selector genes: key regulators of embryogenesis
TRANSCRIPT

Article ID: WMC002573 ISSN 2046-1690
Hox Homeotic Selector Genes: Key Regulators ofEmbryogenesisCorresponding Author:Dr. Antony J Durston,Professor, Sylvius Laboratory, Institute of Biology, Wassenaarseweg 72, 2333 BE - Netherlands
Submitting Author:Dr. Antony J Durston,Professor, Sylvius Laboratory, Institute of Biology, Wassenaarseweg 72, 2333 BE - Netherlands
Article ID: WMC002573
Article Type: Review articles
Submitted on:08-Dec-2011, 08:56:03 AM GMT Published on: 08-Dec-2011, 05:44:11 PM GMT
Article URL: http://www.webmedcentral.com/article_view/2573
Subject Categories:EMBRYOLOGY
Keywords:Hox, Collinearity, Evolution, Xenopus, Drosophila
How to cite the article:Durston A J. Hox Homeotic Selector Genes: Key Regulators of Embryogenesis .WebmedCentral EMBRYOLOGY 2011;2(12):WMC002573
Copyright: This is an open-access article distributed under the terms of the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the originalauthor and source are credited.
Source(s) of Funding:
None
Competing Interests:
None
WebmedCentral > Review articles Page 1 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Hox Homeotic Selector Genes: Key Regulators ofEmbryogenesisAuthor(s): Durston A J
Abstract
The Hox genes are major regulators of the animalbodyplan. They determine the identities of sequentialzones along the main body axis. The Hox genes showcollinearity– spatial and temporal ordering of theirexpression corresponding to their genomic order- aspectacular phenomenon that has excited lifescientists since its discovery in 1978. Threemechanisms have been proposed to explain thisspatially sequential axial pattern of Hox geneexpression in animal embryonic development:1) That it is secondary and regulated by other spatiallyordered genes.2) That it is regulated by interactions among Hoxgenes, or3) It is regulated by the progressive opening ofchromatin in the Hox clusters, from 3’ to 5’.A review of the evidence across different species anddevelopmental stages points to the universalinvolvement of trans-acting factors and cell–cellinteractions. The evidence focuses attention oninteractions between Hox genes and on the vertebratesomitogenesis clock. These novel conclusions opennew perspectives for the field.
Introduction
1. The Body Axes and Developmental ControlGenesTypical vertebrates- like dogs and cats and fish -usually have their head-tail body axis parallel to theground. The head is at the front end and the tail at theback. All limbs (legs or fins) are used for locomotion.In this configuration, we know the head- tail axis as theanteroposterior (main) axis. The upper side of theanimal is called its dorsal side and the lower side itsventral side. In humans, the anteroposterior axis isheld upright. Only the hind limbs are used for walking.Your front is your ventral side and your back yourdorsal side. We use the terminology for a typicalvertebrate in the sections that follow.During embryonic development, a developing animalis built by a hierarchy of genes. These include effectorgenes, encoding building blocks of the embryo- likemuscle actin and keratin. They also include
developmental control genes, which control theexpression or action of other genes. These can begenes encoding proteins controlling the genesis,secretion or transduction of intercellular signals orgenes encoding proteins controlling transcription ortranslation or protein action. Such developmentalcontrol genes regulate each other and may beorganised in very large hierarchies. Hox genes aredevelopmental control genes.2. Discovery and cloning of the Hox genes, theirrole and regulation in Drosophila. Hox genes were first discovered as homeotic genes inthe fruitfly Drosophila. They are sometimes referred toas: homeotic selector genes or: HOMC genes. Theyare characterised by the fact that a gain or loss offunction mutation in a typical Hox gene can result inconversion of one large or small part of the main bodyaxis to another. These are clearly developmentalcontrol genes acting high up in the hierarchy. In thecase of the Hox genes, the conversions take placebetween different parts of the anteroposterior axis.One famous example is: Bithorax, discovered byNobel prize winner Ed Lewis, which makes a fourwinged fly in its loss of function format. Drosophilanormally has only two wings, on the anterior thorax.The posterior thorax has vestigial ‘halteres’ . Bithoraxis a gene for posterior thorax which converts this toanterior thorax by loss of function (Lewis, 1978, 1995).In another equally famous example, discovered byWalter Gehring, Antennapedia, a gene for anteriorthorax converts part of the fly’s head to anterior thoraxand therefore antennae to legs by misregulated gain offunction (Carrasco et al., 1984) Vertebrate Hox genessimilarly have drastic phenotypes but loss of functionphenotypes are more difficult to visualise becauseeach vertebrate Hox function is mediated by multipleHox genes and these must all be knocked out. SeeFig.1.These genes typically determine the identity ofindividual Drosophila body segments or groups ofadjacent segments. In the early 80’s strategies weredeveloped for cloning developmental control genes.The first genes cloned were the hox genes Bithorax-by Hogness and his colleagues (Bender et al., 1983) (and Antennapedia- by the Gehring group (Carrasco etal., 1984). This was possible because thesetranscription factor genes contain a large highlyconserved region- the homeobox- which encodes a 60
WebmedCentral > Review articles Page 2 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
amino acid DNA binding domain and can be picked upby homology screening. It has, in fact emerged thatHox genes encode a subfamily of transcription factorsand that the homeobox and another conserved region,the haxapeptide, are important in determining theirspecificity.3. Hox clustering and colinearity: The key property. A key property of Hox genes is that they are oftenclustered in complexes. Hox complexes are amongthe most remarkable regions of the genome. A Hoxcomplex usually consists of up to 9-13 closely relatedHox genes arranged in tandem. These genes specifypatterning along body axes in all bilateria (Gehring etal., 2009, Duboule, 2007). Invertebrates have a singleHox complex, or dispersed Hox genes, but tetrapodvertebrates typically possess four similar Hoxcomplexes (HoxA–D), located on differentchromosomes (Duboule, 2007). (Fig. 2) The Hoxcomplexes also contain 5 micro RNA (miRNA) genesintercalated at homologous positions (Pearson et al.,2005; Yekta et al., 2004, 2008; Woltering and Durston,2007; Ronshaugen et al., 2005).The 3’ to 5’ order of Hox genes along a chromosomecorresponds to the order in which they act along bodyaxes; this collinear property links clustering to function,emphasizing that Hox complexes are functional unitsor meta genes No one Hox gene can pattern an axisbut a whole Hox cluster can. (Mainguy et al., 2007,Duboule 2007). Hox collinearity is crucial inembryogenesis and includes 3 important andinterrelated properties: functional colinearity describesthe spatial order in which Hox genes act along a bodyaxis; spatial colinearity refers to the spatial order inwhich the Hox genes are expressed, and temporalcolinearity is the time sequence in which they areexpressed (Text Box 1). . The organization of Hoxcomplexes is highly conserved, and Hox and mirgenes not only have remained clustered throughbilaterian evolution but are also in close proximity toeach other despite their very complex and dynamicexpression patterns. Individual Hox genes are alsovery highly conserved in Evolution.CollinearityCollinearity describes the sequential expression of agenomic cluster of Hox genes along an embryonic axisand associated properties.There are three important forms of collinearity: Spatialcollinearity is the sequential 3’ to 5’ expression of Hoxgenes along a body axis. This occurs from anterior toposterior along the main body axis and also in otheraxes, for example from proximal to distal in developinglimbs. Spatial colinearity can be associated with timedependence. The most 3’ gene is expressed first and
more 5’ genes are expressed sequentially later. This isdefined as temporal collinearity and, in earlyvertebrate development, spatial collinearity isgenerated from pre-existing temporal collinearity bytime space translation. The gastrula’s organiserinteracts with Hox expressing non organisermesoderm to translate a temporal sequence of Hoxcodes to a spatially collinear pattern. We also define athird property, functional collinearity- — which is thecapacity of Hox genes to coll inearly defineregion-specific structures along an axis.Hox collinearity and the organisation of the Hoxcomplexes are phenomena that have long fascinateddevelopmental, molecular and evolutionary biologists.These phenomena represent an important example ofgenomic regulation. Understanding the structure andfunction of Hox genes is crucially important, becausethey are implicated in a growing number of diseases,including important cancers (Grier et al., 2005). Seealso below.Research and thinking on Hox collinearity hasconcentrated on three aspects. First, there is thequestion of how collinearity evolved, which is clearlyone of the keys to understanding this phenomenon.Second, there are three mechanistic ideas. The first isthat Hox spatial colinearity is secondary and due to anupstream hierarchy of spatially ordered genes. Hoxcollinearity is thus not utilised. The second is thatcollinearity is based on transcriptional regulation, andspecifically that it is limited by the progressive 3’ to 5’opening of Hox cluster chromatin and/or mediated byglobal control regions. The third model is thatcollinearity depends on interactions among the Hoxgenes themselves. These interactions include‘posterior prevalence’, - a negative interaction amongHox proteins that clearly relates to functionalcollinearity in Drosophila (and possibly also to spatialand temporal collinearity; see Text Box 1). In this article, we review the basis of Hox evolutionand of the three longstanding mechanistic hypothesesto explain Hox gene collinearity.. But we also proposea new explanation. Based on evidence fromAmphibian and other vertebrate embryos, we reasonthat synchronised temporally collinear expression ofthe Hox complexes in early vertebrate embryosinvolves trans-acting factors and intercellularinteractions. We review data implicating activating aswell as repressive interactions among the Hox genesthemselves, and timed signals from the somitogensisclock. This model provides a mechanistic link betweenthe different aspects of collinearity . A review ofpotential collinearity mechanisms is now opportunebecause new data that have never been reviewed inthe literature are now available and because the
WebmedCentral > Review articles Page 3 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
existing, entrenched models are limiting in the sensethat they direct research in the same direction- that ofchromatin opening and transcriptional control- and thatthey do not explain all of the facts (below). This hasspurred us to interpret the data in a different light. Thefield gains a new perspective from this new synthesisof the data4. The evolution of Hox genes in different taxa,including vertebrates. Hox genes are available in all metazoans that havebeen studied. In all bilateria where there is information,they are concerned with patterning the main bodyaxis. Invertebrates have one Hox gene complex:vertebrates have 4 or 8. The 4 Hox gene complexestypically present in most tetrapod vertebrates arosethrough 2 rounds of genome duplication duringevolution. Xenopus laevis and teleost fishes have 8Hox complexes because of 3 genome duplications.Even the individual Hox genes are strongly conservedin evolution throughout the animal kingdom (Carrascoet al., 1984; Gehring et al. 2009; Duboule 2007,DeRobertis, 2008) and are recognisable by havingdistinct conserved sequences. The Hox genescorresponding to the same position in each of thedifferent vertebrate Hox complexes are conserved.They have very similar homeoboxes and hexapeptidesand are called a paralogue group. Hox genes ininvertebrates may be clustered and show collinearityor they may be scattered in the genome to variousextents. Different extents of fragmentation, fromatomised to fully clustered have been identified. Theclustered format is thought to be ancestral. An evolutionary explanation of collinearityIt has been proposed that colinearity evolved byrepeated tandem duplication of an ancestral ur-Hoxgene and sequential evolutionary modifications of theduplicates, leading to generation of an organised genearray from an evolutionary ground state . This idea canconceivably explain how a genomic sequence couldrelate to a spatial or temporal sequence of geneexpression. Please note that, if this is the explanationof collinearity, it is the explanation and obviates theneed for an explicit collinearity mechanism involvinginteractions between or clustering of the Hox genes.The upstream mechanism for Hox expression will bewhatever it evolved to be in order to regulate thespatially collinear localised expression of the individualHox genes- as with the segmentation gene hierarchyin Drosophila. The spatially collinear axial expressionpattern of the Drosophila Hox genes is thus secondaryand determined by the spatially ordered expressionpatterns of the gap genes. Nonetheless, we think thatexplicit collinearity mechanisms evolved- see main text. Evolution of Hox collinearity is particularly important
because it can potentially offer an explanation of howcollinear properties connect to Hox complex structure.The only other potential explanation for this comesfrom the chromatin opening model (below). It shouldbe noted that whereas clustered Hox genes inorganisms having Hox clusters show the normalspatially collinear sequence of Hox gene expression,so do Hox genes in fragmented clusters, from the splitcluster seen in Drosophila to atomised Hox genes inorganisms having no clustering- like Oikopleura(Duboule 2007, Seo et.al. 2004). These show ‘transcollinearity’. It is thus clear that the spatial ordering ofHox gene expression does not rely soley onclustering. Presumably, Hox spatial collinearityevolved in an ancestral organism with clustered Hoxgenes and and persisted after cluster disintegrationduring evolution. This already demonstrates that Hoxcollinearity properties can persist in the absence Hoxclustering and therefore of progressive chromatinopening. It has been proposed that a Hox complex,whose function is to pattern an axis, acts as a metagene or functional unit, where no one Hox gene canexecute the whole function but the whole complexdoes (Mainguy et al., 2007, Duboule, 2007). It hasalso been proposed that spatial collinearity has been aselective pressure that drives Hox clustering ratherthan vice versa. (Duboule 2007) . It has been proposed that Hox colinearity evolved byrepeated tandem duplication of an ancestral ur-Hoxgene and stepwise sequential evolutionarymodifications of the duplicates, leading to generationof an organised gene array from an evolutionaryground state (Lewis 1978 1995; Gehring et al.,2009)(Text Box 2, Fig.3A). Lewis proposed that themodifications arose by unequal recombinationbetween adjacent Hox genes. This idea canconceivably explain how a genomic sequence couldgenerate ordered properties like spatial or temporalsequences of gene expression. Please note that, if thisis the explanation of collinearity, it obviates any needfor a dedicated collinearity mechanism. The upstreammechanism for Hox expression will be whatever itevolved to be in order to regulate the correctlylocalised expression of the individual Hox genes. Thisis the case with the gap-segmentation gene hierarchyin Drosophila, (see below). Nonetheless, we think thatdedicated collinearity mechanisms evolved. Lewisshowed that 5’ posterior drosophila Hox genes areepistatic to the Hox gene Antennapedia. If they areectopically expressed in the normal Antennapediadomain, the most posterior Hox gene expresseddominates. If the most posterior Hox gene is deleted,the phenotype obtained is that of the most posteriorHox gene still expressed. And so on. This interaction
WebmedCentral > Review articles Page 4 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
was called posterior prevalence (below) and wasthought by Lewis to reflect the fact that Antennapediarepresents an ancestral ground state, while posteriorHox genes are derived from the ground state bytandem duplication and stepwise sequentialmodification (as above). It has been reported relativelyrecently by Gehring et al., (2009) that the anteriorDrosophila Hox genes have also evolved from theAntennapedia ancestral ground state. 5. The Mechanism Of Hox CollinearityThere are various ideas about this (Fig. 3). 1) In the section above, we have described the ideathat collinear Hox complexes arose by tandemduplication and sequential modification of an ancestralur-Hox gene. In this case, no special mechanism isrequired to generate spatial collinearity. The upstreammechanism for Hox expression will be whatever itevolved to be in order to regulate the correctlylocalised expression of the individual Hox genes. Thisis the case with the gap-segmentation gene hierarchyin Drosophila (Nuesslein- Volhard, 1995), where thespatial ordering of the Hox genes is secondary. Thespatially expressed gap genes are the primarydeterminats of the spatially ordered Hox geneexpression pattern (Kehle et al., 1998, Mito et al.,2006) . Later on, other genes, including the Hox genesthemselves, the cofactor teashirt, polycomb groupgenes and segmentation genes play a role ( Gebeleinand Mann, 2007, Rusch and Kaufman, 2000 Mito etal., 2006) (Fig 3A, 3B) . 2) The idea has developed in the mouse that temporalcollinearity is due to progressive opening of Hoxcomplex chromatin, from 3’ to 5’ (Fig 3C). (. This ideahas become rather popular. There is some evidencefor this (Soshnikova and Duboule 2009, Cambeyronand Bickmore, 2004, Van der Hoeven et al, 1996,Kmita et al., 2000)but the idea has limited application. It can not apply inanimals with dispersed Hox genes that behavecolinearly. It is not even the whole story in vertebrates,presumably including the mouse. Synchronisedtemporal colinearity between the different Hoxcomplexes during gastrulation (Wacker et al., 2004,Durston et al., 2010, 2011) indicates the importance oftrans acting factors and intercellular signals.3) There is evidence that interactions between Hoxgenes are important. These can obviously not accountfor the relation between Hox complex structure andcollinear properties but they are part of the story.Working in D. melanogaster, E. B. Lewis showed thatloss–of-function mutations in posterior Hox genesdrive the segmental phenotype towards that of themore anterior thoracic segment T2, which isdetermined by the Hox gene Antennapedia (Lewis,
1978, 1995). Struhl used esc- Drosophila embryos,which show constitutive activation of gene expression,in combination with Hox loss of function mutations toelucidate the functional hierarchy of Drosophila Hoxgenes (Struhl, 1983). All Drosophila segments weretransformed to the phenotype of the most posteriorfunctional Hox gene expressed. Posterior prevalencein Drosophila has been thought to underly functionalcollinearity only, not spatial collinearity. Experimentallyderived ubiquitous expression of Hox genes underpromoters that are known to be transcriptionallyirrepressible leads to transformations only in regionsanterior to the functional domain of the gene. Forexample, the thoracic Antennapedia, whenubiquitously expressed, suppresses Hox genes of thehead, resulting in posterior transformation of headsegments towards a thoracic identity while notaffecting the abdomen — here, the effect of Antp isphenotypically suppressed by bithorax-complex genessuch as Ubx (Gonzalez-Reyes et al., 1990 Gibson andGehring, 1988 ). However, posterior prevalence occurs not only postranslationally (Plaza et al., 2008)but also at the levels of transcription (Beachy et al.,1988, Hafen et al., 1984, Appel and Sakonju, 1993,Struhl and White, 1985) and posttranscriptionalregulation of mRNA abundance (Yekta et al., 2004,2008, Woltering and Durston, 2007, Ronshaugen etal.,2005 ) (Text box 3). It can thus also potentiallyregulate the mRNA expression of Hox genes. Namely,spatial and temporal collinearity. Hox interactions alsooccur during vertebrate gastrulation. These includeposterior prevalence (Hooiveld et al, 1999, Wolteringand Durston, 2007) but also 3’ to 5’ activation of Hoxgene expression (McNulty et al, 2006, Hooiveld et al.,1999) See Fig. 3E, Fig. 5The Level Of ActionAll effects above on activation or repression of Hoxgenes during gastrulation result in more or less HoxmRNA.but not all act on transcription. Recentevidence shows that Hox complex mRNA availability isstrongly regulated posttranscriptionally, involving suchphenomena as polycistronic transcripts, sense/antisense transcript interactions and alternativesplicing. At least one early vertebrate Hox interaction;downregulation of more 3’ Hox mRNA’s by Hoxb4 ismicro RNA mediated (posttranscriptional). We notethat the important parameter for colinearity is the sumtotal of the (activating and repressing) inputs on eachHox gene (there may be many). We think it verysignificant that posterior prevalence (pp) acts at 3different levels. If a Hox gene is activatedtranscriptionally, its mRNA can still be destabilised bypp miR action. If the Hox protein is made, it can still beinactivated by pp protein-protein interactions. We think
WebmedCentral > Review articles Page 5 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
that pp is the most important Hox-Hox colinearityinteraction and that it needs to be dominant, to ensurethe 3’ to 5’ directionality of colinearity.
6. Hox function in vertebrates.
Hox genes have several different roles in developmentVertebrates are unique in being the only Type ofmetazoan animals in which the ancestral Hox clusterhas been duplicated due to genome duplications. Inmost tetrapod vertebrates, there are four Hox clusters,on different chromosomes, presumably due to 2genome duplications. Teleost fish have 8 clusters, dueto 3 genome duplications.6.1 Hox genes in the developing CNS andhindbrainThere is much evidence that Hox genes are importantin early anteroposterior patterning of the vertebratecentral nervous system. There is an approximatelyspatially collinear sequence of Hox expression in theearly neural plate and neural tube. Anterior boundariesfor expression of different Hox genes distinguishbetween different parts of the developing CNS- forexample, some boundaries distinguish between thedifferent segments in the hindbrain. Much work hasbeen done to characterise the regulatory genenetworks that regulate Hox expression and Hoxfunction in the developing CNS, particularly those thatpattern the developing hindbrain: a segmentedstructure. These networks do not appear to containany mechanism that mediates collinearity, which ispresumably set up earlier in the mesoderm andtransferred to the developing CNS. (see below). Thehindbrain regulators seem to maintain this earlypattern or alter it. They have an ad hoc nature, as dothe upstream regulators in Drosophila. They do notnecessarily maintain spatial collinearity. For example,the primary Hoxb1 expression domain is at a noncollinear position. This work has been reviewedextensively in recent review articles (Wright, 1993,K rum lau f , 1994 , Tumpe l e t a l . , 2009 ,Schneider-Manoury et al., 1998) and will not bediscussed further here. The role of Hox genes inpatterning the developing vertebrate CNS is limited tothe hindbrain and spinal cord. The fore- and mid- brainare patterned by other regulators, including the Otxand Emx gene families (Cecchi et al., 2000). Thepatterning of the anterior CNS by these gene familiesis actually conserved in Drosophila, but the anteriorCNS region where they act here is very small,compared with the vertebrate forebrain.6.2 Hox genes in axial mesoderm Besides specifying A-P levels early on, in the
developing central nervous system, Hox genes specifyA-P levels in mesoderm. We are talking here about theaxial and paraxial mesoderm. Hox genes areexpressed in and specify A-P levels in, the presomiticand somitic mesoderm and the lateral plate mesoderm.Hox genes are not expressed in and do not specifyA-P levels in the early notochord, which is derivedfrom the Hox-negative organiser mesoderm in thegastrula. Hox patterning of axial mesoderm is coveredby excellent recent reviews (Carapuco et al.,2005,Burke et al., 1995). We will not discuss it further here,except for one aspect (below).The expression of Hox genes in the presomitic andsomitic mesoderm is interesting because it correlateswith the process of somitogenesis, the primaryprocess of segmentation in the early vertebrateembryo, which occurs in this mesoderm. Vertebratesomitogenesis (segmentation of axial mesoderm)works via a mechanism where an oscillating system ofgene expression generates a spatial pattern bytime–space translation, just as in genesis of thevertebrate axial Hox pattern (see below and text box1). The temporal oscillation in gene expression(somitogenesis clock) generates spatially periodicsegments in the axial mesoderm: the somites(Palmeirim et al., 1997). This is closely linked tocollinear Hox expression. Hox spatial expressionboundaries coincide with somite/segment boundariesand several vertebrate somitogenesis genes areknown to regulate Hox expression (Peres et al., 2006;Dubrulle et al., 2001, Dubrulle and Pourquie, 2004,Zakany et al., 2001). 6.3 Hox genes in gastrulationHox genes are expressed earlier in development thanin the developing central nervous system and axialmesoderm. This is interesting because the Hox genesset up the primary axial pattern during these earlystages. The Hox genes are already expressed duringgastrulation. For example, in the non-organisermesoderm (NOM) of the Xenopus laevis gastrula,where Hox genes are first expressed in the embryoand are expressed with temporal colinearity (Fig.4a).This mesoderm manifests a sharply timed temporallycollinear sequence of Hox gene expression that istranslated in time and space by interactions with theSpemann organiser (SO) to to generate a spatiallycollinear pattern of Hox gene expression along themain body axis of the organism (Wacker et al., 2004a;Durston et al., 2010, 2011) The mechanism for this isshown in Figure 4b. .In short, the temporal sequenceof Hox gene expression in the mesoderm issequentially frozen, from anterior to posterior and istransferred to the developing neural plate, whichoverlies the internalising mesoderm, in the gastrula.
WebmedCentral > Review articles Page 6 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
(Text Box 1, Fig. 4a,b). A striking feature of the Xenopus gastrula mesoderm’stemporally collinear Hox expression sequence is thatexpression of Hox genes from different Hoxcomplexes occurs in the same perfectly temporallycollinear sequence (Fig. 4A). The temporal collinearityof the different Hox complexes is thereforesynchronised (Wacker et al., 2004a; Durston et al.,2010, 2011). The different Hox paralogues (ie thedifferent copies of each different Hox gene type,produced by the vertebrate genome duplications) inthe different complexes are on different chromosomes,ruling out that Hox colinearity simply reflectscis-localised progressive opening of Hox complexchromatin for transcription. Trans acting signals areclearly needed to synchronise the different Hoxcomplexes and, since we are dealing with a cell massrather than a single cell, intercellular signals are alsorequired. We note that these trans-acting factors andintercellular signals must be very sharply timed toenable synchronisation of the different Hox complexesand are probably timed to trigger expression ofdifferent Hox genes at different times. This conclusionwas not a complete surprise. It is known that transacting factors must mediate collinearity in organismswith dispersed Hox genes. This is, however, the firstevidence that vertebrate temporal collinearity is alsomediated by trans acting factors. The involvement of trans acting factors andintercellular signals has been investigated and threesectors of the regulatory gene hierarchy have becomeinteresting.1) There is evidence that the Hox genes themselvesare involved, via Hox-Hox interactions includingposterior prevalence and via interactions involvingmicro RNA’s. These interactions drive initiation of Hoxcomplex expression as well as progression oftemporally collinear expression through the Hoxcomplexes (Hooiveld et al., 1999, Woltering andDurston, 2008, McNulty et al., 2005) (Fig. 5A). Thereis much evidence that Hox genes in vertebrates andDrosophila show activating as well as repressiveinteractions, including posterior prevalence McNulty etal., 2006; Hooiveld et al., 1999; Woltering and Durston,2008; Le Pabic et al., 2010; Lobe. 1995, Maconochieet al., 1997; Gould et al., 1997; Bergson andMcGinnis,1990 ; Miller et al., 2001, Wellik andCapecchi, 2003) and that they drive conventionalintercellular signalling pathways(eg. Graba et al. 1995,Bruhl 2004, Manak et al1994, Michaut et al., 2011,Morsi el Kadi et al., 2002, Pearson et al., 2005) aswell as acting as signalling molecules themselves(Bloch- Gallego et al., 1993, Chatelin et al., 1996).2) There is evidence that the signalling factor Wnt 8
acts as a signal to initiate synchronous expression ofthe different Hox complexes. (In der Rieden et al.,2010)3) There is evidence that the somitogenesis clock isinvolved (Fig. 5B). Vertebrate somitogenesis(segmentation of axial mesoderm) works via amechanism where an oscillating system of geneexpression generates a spatial pattern by time–spacetranslation, just as in genesis of the vertebrate axialHox pattern (see above and text box 1). This dynamicprocess is known to start during gastrulation in chickenand Xenopus ( Peres et al, 2006; Jouve et al., 2002) and it drives activation of Hox gene expression.Xdelta2 is a Xenopus oscillating somitogenesis gene(Jen et al., 1997, 1999). It is already expressed duringgastrulation and then generates presomitic stripes soits expression is already oscillatory. It regulatesexpression of Hox genes during gastrulation (Peres etal., 2006). This gene could help to drive synchronisedtemporally collinear expression of the Hox complexes.It could do so either by regulating only initiation ofexpression of Hox complexes (via labial Hox genes) orby driving initiation and 3’ to 5’ progression,(repeatedly inducing expression of different Hoxgenes). We note that XDelta2 drives expression of atleast 3 different Hox paralog groups including labial). Ifdelta drives progression as well as initiation, arepeated periodic pulsatile signal is required. The ideathat the somitogenesis clock drives Hox temporalcollinearity is very attractive because both of thesetimers are known to operate already in the gastrulaand because of the evidence linking Hox patterningand segmentation (above). Such a signalling pathwaymight act separately from the Hox genes or be downstream of them. XDelta 2 is indeed downstreamof Hox genes as well as upstream. There is a positivefeedback loop (McNulty et al., 2006, Peres et al.,2006). XDelta 2 may thus mediate Hox inducedsignalling. These findings indicate that the axialsegmentation mechanism may help to drive Hoxexpression in vertebrates, just as in Drosophila.The X. laevis example was chosen because the dataare most complete for this system; however, theconclusions are strongly supported by many findingsin other vertebrates (zebrafish, chicken and mouse)(Gaunt and Strachan, 1996, Alexandre et al. 1996,Deschamps et al., 1999). This example illustrates thatHox colinearity cannot depend solely on the collinearopening of chromatin. Because the Hox complexesare synchronised, trans-acting factors and intercellularsignals must be involved — trans-acting factors wouldbe necessary for coordinating the sequential 3’ to 5’activation of Hox genes in and between Hox clusters,and intercellular signals would enable the coordinated
WebmedCentral > Review articles Page 7 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
activation of Hox gene expression between cells in atissue. An alternative explanation is that only the most3’ Hox genes (Hox1) transactivate, and the remainingtiming is provided by synchronised opening of the Hoxcomplexes. The different structures of the 4 primaryvertebrate Hox complexes (with different Hoxparalogues missing from each) would, however, makeit difficult for progressive opening of different Hoxcomplexes to stay synchronous. Since the gastrulamesoderm is a cell mass, not a single cell,trans-activation needs to be accompanied byintercellular signalling.6.4 Hox genes in later development and in. limbs,hairs, haematopoiesis, the pancreas, etc.Vertebrate Hox genes have many other functions thanspecifying levels in the main body axis, in the centralnervous system and in axial mesoderm. They regulatethe axial patterning of limbs (Zakany and Duboule,2007). They mediate patterning and differentiation inhairs (Awgulevich, 2003), the gut (Kawazoe et al.,2002), the pancreas (Gray et al., 2011), the blood(Magli et al.,. See articles for details. These otherfunctions will not be discussed further here. Many ofthese Hox functions have been elucidated by gain andloss of function expts. In general, loss of functionmutation in a single vertebrate Hox gene delivers adeceptively mild phenotype. This has bedevilled theanalysis of Hox function using mouse genetics. It isbecause each vertebrate Hox gene is a member of aparalogue group of at up to 4 or 8 Hox genes whichhave parallel and shared functions. Where measureshave been takn to knock out a whole paralogue group,a suitably dramatic phenotype is obtained. See Fig. 1. 6.5 Modified use of Hox genes in elongatedvertebrates: snakes and Caecilians.The elongated, snake-like skeleton, as it hasconvergently evolved in numerous reptilian andamphibian clades, is from a developmental biologist’spoint of view amongst the most fascinating anatomicalpeculiarities in the animal kingdom. This kind of bodyplan is characterized by a greatly increased number ofvertebrae, a reduction of skeletal regionalization alongthe primary body axis and loss of the limbs. Recentstudies conducted in both mouse and snakes now hintat how changes in gene regulatory circuitries of theHox genes and the somitogenesis clock could underliethese striking departures from standard tetrapodmorphology. These studies show that particular snakeHox genes have changed their specificities bymutations in the homeobox. This leads to their failingto specify the expected axial boundaries and enablesparticular body regions, especially the thorax, tobecome drastically extended (Woltering et al., 2009, DiPoi et al., 2010).
Conclusion
Hox genes are upstream regulators in thedevelopmental hierarchy that are of great importancefor the bodyplan. They specify and differentiatebetween different zones along the main body axis.These genes show collinearity- clustering associatedwith acquisition of ordered properties within the genecluster- a spectacular phenomenon that has attractedmuch interest. A Hox cluster is actually a metagene. It,but not an individual Hox gene, can fulf i l adevelopmental function- patterning the body axis. InDrosophila, and probably in all other invertebrates- thefull potential of the Hox genes is not realised. Theexpression of each individual Hox gene is regulated byother spatially regulated genes and so Hox collinearityis not used to pattern the main body axis. Invertebrates, temporal collinearity has been developedand this is used to pattern the main body axis anddevelop spatial collinearity, by time-space translation.It is presently generally assumed that the mechanismof temporal collinearity is progressive 3’ to 5’ openingfor transcription of Hox complexes. This may beimportant. However, we develop a differentmechanistic hypothesis: that collinearity is partlymediated by Hox gene interactions. This idea wasalready indicated by earlier investigations of posteriorprevalence. We review new evidence that trans-actingfactors and intercellular signals mediate vertebrateHox collinearity; that these include interactions amongHox genes, including posterior prevalence, as well assomitogenesis signals. We propose that these Hoxinteractions have a role in generating Hox temporaland spatial collinearity as well as functional collinearity.We note also that an evolutionary explanation forcollinearity actually probably obviates any requirementfor a dedicated collinearity mechanism. Ourconclusions open new perspectives for research intothe mechanisms underlying collinearity. Testing thismodel will require a much more extensive investigationand description of early vertebrate Hox temporalcollinearity.
References
1. Alexandre D, Clarke JD, Otoxby E, Yan YL,Jowett T, Holder N. (1996)Ectopic expression ofHoxa-1 in the zebrafish alters the fate of themandibular arch neural crest and phenocopies ar e t i n o i c a c i d - i n d u c e d p h e n o t y p e .Development.;122(3):735-746.2. Appel B, Sakonju S, (1993) Cell-type-specific
WebmedCentral > Review articles Page 8 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
mechanisms of transcriptional repression by thehomeotic gene products UBX and ABD-A inDrosophila embryos. EMBO J. 12(3): 1099–1109. 3. Awgulewitsch A. Hox in hair growth anddevelopment. Naturwissenschaften. 2003May;90(5):193-211. Epub 2003 Apr 26.4. Beachy PA, Krasnow MA, Gavis ER, HognessDS.(1988) An Ultrabithorax protein binds sequencesnear its own and the Antennapedia P1 promoters. Cell.1988;55(6):1069-1081.5. Bender W, Spierer P, Hogness DS Chromosomalwalking and jumping to isolate DNA from the Ace androsy loci and the bithorax complex in Drosophilamelanogaster. J Mol Biol. 1983 Jul 25;168(1):17-33.Bergson C, and McGinnis W, (1990) Anautoregulatory enhancer element of the Drosophilahomeot ic gene Deformed EMBO Journal9,.13.4287-4297, 6. Bloch- Gallego E, Le Roux I, Joliot AH, Volovitch M,Henderson CE, Prochiantz A. (1993) Antennapediahomeobox peptide enhances growth and branching ofembryonic chicken motoneurons in vitro. J Cell Biol.120(2):485-492.7 . Bruh l , T . e t a l . (2004) Homeobox A9transcriptionally regulates the EphB4 receptor tomodulate endothelial cell migration and tube formation.Circ. Res. 94, 743–751 8. Burke AC, Nelson CE, Morgan BA, Tabin C. Hoxgenes and the evolution of vertebrate axialmorphology. Development. 1995 Feb;121(2):333-46.9. Cambeyron S, Bickmore WA, (2004) Chromatindecondensation and nuclear reorganization of theHoxB locus upon induction of transcription. Genes Dev.18(10): 1119–1130.10. Carapuco M, Novoa A, Bobola N, Mallo M (2005)Hox genes specify vertebral types in the presomiticmesoderm. Genes Dev.;19(18):2116-2121 11. Carrasco AE, McGinnis W, Gehring WJ, DeRobertis EM, (1984) Cloning of an X. laevis geneexpressed during early embryogenesis coding for apeptide region homologous to Drosophila homeoticgenes. Cell.;37(2):409-414.12. Cecchi C, Mallamaci A, Boncinelli E. Otx and Emxhomeobox genes in brain development. Int J Dev Biol.2000;44(6):663-8.13. Chatelin J, Volovitch M, Joliot AH, Perez F,Prochiantz A. (1996) Transcription factor hoxa-5 istaken up by cells in culture and conveyed to theirnuclei. Mech Dev. 55(2):111-117.14. De Robertis EM, (2008) Evo-devo: variations onancestral themes. Cell. 132(2):185-195.15. Deschamps J, Van den Akker E, Forlani S, DeGraaff W, Oosterveen T, Roelen B, Roelofsma J.(1999) Initiation, establishment and maintenance of
Hox gene expression patterns in the mouse. Int J DevBiol. 43(7):635-650.16. Di-Poï N, Montoya-Burgos JI, Miller H, Pourquié O,Milinkovitch MC, Duboule D.Changes in Hox genes' structure and function duringthe evolution of the squamate body plan. Nature. 2010Mar 4;464(7285):99-103.17. Duboule D, (1994).Temporal colinearity and thephylotypic progression: a basis for the stability of avertebrate Bauplan and the evolution of morphologiesthrough heterochrony. Dev Suppl. 135-4218. Duboule D,(2007) The rise and fall of Hox geneclusters. Development 134(14): 2549-2460.Dubrulle J.,. McGrew MJ, Pourquié O, (2001).Cell 106,219 19. Dubrulle J, Pourquie O. (2004) Couplings e g m e n t a t i o n t o a x i s f o r m a t i o n .Development.;131(23):5783-579320. Durston A., Jansen HJ Wacker SA,.(2010) Review:Time-Space Translation Regulates Trunk AxialPatterning In The Early Vertebrate Embryo. Genomics95, 250-255.21. Durston A, Jansen H, and Wacker S, (2011)Time-Space Translation: A Developmental Principle.The Scientific World In Press.22. Gaunt SJ, Strachan L. (1996) Temporal colinearityin expression of anterior Hox genes in developingchick embryos. Dev Dyn.;207(3):270-28023. Gebelein B, Mann RS Compartmental modulationof abdominal Hox expression by engrailed andsloppy-paired patterns the fly ectoderm. Dev Biol.2007 Aug 15;308(2):593-605. Epub 2007 May 24.24. Gehring WJ, Kloter U, and Suga H, (2009).Evolution of the Hox Gene Complex from anEvolutionary Ground State. Current Topics inDevelopmental Biology 88: 35-6125. Gibson G, Gehring W. (1988) Head and thoracictransformations caused by ectopic expression ofAntennapedia during Drosophila development.Development , 102, 657–675. 26. Gomez C, Ozbudak EM, Wunderlich J, BaumannD, Lewis J, Pourquié O. Control of segment number inve r teb ra te embryos . Na tu re . 2008 Ju l17;454(7202):335-9. Epub 2008 Jun 18.27. Gonzalez Reyes G, Urquia N, Gehring WJ, StruhlG Morata G, (1990), Are cross-regulatory interactionsbetween homoeotic genes functionally significant?,Nature 344 78–80.28. Gould A, Morrison A, Sproat G, White RA,Krumlauf R. (1997) Positive cross-regulation andenhancer sharing: two mechanisms for specifyingoverlapping Hox expression patterns. Genes Dev.11(7):900-13.29. Graba Y. et al. (1995) DWnt-4, a novel Drosophila
WebmedCentral > Review articles Page 9 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Wnt gene acts downstream of homeotic complexgenes in the visceral mesoderm. Development 121,209–218.30. Gray S, Pandha HS, Michael A, Middleton G,Morgan R.HOX genes in pancreatic development and cancer.JOP. 2011 May 6;12(3):216-9.31. Grier DG, Thompson A, Kwasniewska A,McGonig le GJ, Hal l iday HL, (2005) Thepathophysiology of HOX genes and their role in cancer.J. Pathol205(2): 154–71.32. Hafen E, Levine M, Gehring WJ.(1984) Regulation of Antennapedia transcript distribution byt h e b i t h o r a x c o m p l e x i n D r o s o p h i l a .Nature.;307(5948):287-289.33. Hooiveld M, Morgan R, In Der Rieden P, Houtzager E, Pannese M,, Damen K, Boncinelli E, Durston A, (1999) Novel colinear interactions betweenvertebrate Hox genes. Int. J. Dev. Biol. 43:665-67434. In Der Rieden PMJ, Lloret Vilaspasa F, DurstonAJ, (2010). Xwnt8 directly initiates expression of labialHox genes. Dev. Dynamics 29: 226-239 35. Jen WC, Wettstein D, Turner D, Chitnis A, KintnerC. (1997) The Notch ligand, X-Delta-2, mediatessegmentation of the paraxial mesoderm in Xenopusembryos. Development. 1997;124(6):1169-1178.36. Jen WC, Gawantka V, Pollet N, Niehrs C, KintnerC. (1999) Periodic repression of Notch pathway genesgoverns the segmentation of Xenopus embryos.Genes Dev.;13(11):1486-149937. Jouve C, Iimura T, Pourquie O.(2002) Onset of thesegmentation clock in the chick embryo: evidence foroscillations in the somite precursors in the primitivestreak. Development.;129(5):1107-111138. Kawazoe Y, Sekimoto T, Araki M, Takagi K, ArakiK, Yamamura K.Region-specific gastrointestinal Hox code duringmurine embryonal gut development. Dev Growth Differ.2002 Feb;44(1):77-84.39. Kehle J, Beuchle D, Treuheit S, Christen B,Kennison JA, Bienz M, Müller J.dMi-2, a hunchback-interacting protein that functionsin polycomb repression. Science. 1998 Dec4;282(5395):1897-900.40. Kmitam M., et al., (2000) Mechanisms of Hox genecolinearity: transposition of the anterior Hoxb1 geneinto the posterior HoxD complex. Genes Dev.14(2):198-211..41. Krumlauf R, (1994) Hox genes in vertebratedevelopment Cell 78, 2, 191-20142. Le Pabic P, Scemama JL, Stellwag EJ. (2010)Role of Hox PG2 genes in Nile tilapia pharyngeal archspecification: implications for gnathostome pharyngeal
arch evolution. 12(1):45-60.43. Lewis EB, (1978) A Gene Complex ControllingSegmentation in Drosophila Nature 276: 565-568 44. Lewis EB, Nobel lecture, December 8, 1995. Thebithorax complex: The first fifty years. In: H. Lifshitz,Editor, Genes, Development and Cancer. The Life andWork of Edward B. Lewis, Kluwer AcademicPublishers, Norwell, MA 45. Lobe CG, (1995) Activation of Hox geneexpression by Hoxa-5. DNA Cell Biol.;14(10):817-23.46. Maconochie MK, et al, (1997) Cross-regulation inthe mouse HoxB complex: the expression of Hoxb2 inrhombomere 4 is regulated by Hoxb1. Genes Dev. 11,1885–1895.47. Mainguy G, Koster J, Woltering J, Jansen H,Durston A, (2007).Extensive polycistronism andantisense transcription in the Mammalian Hox clusters.PLoS ONE 2(4):e356.48. Magli MC, Largman C, Lawrence HJ. Effects ofHOX homeobox genes in bloodcell differentiation. JCell Physiol. 1997 Nov;173(2):168-77.49. Manak JR, Mathies LD, Scott MP (1994)Regulation of a decapentaplegic midgut enhancer byhomeotic proteins. Development 120, 3605–3612 50. McNulty C, Peres J, Van Den Akker W, Bardine N, Durston A. (2005) A Knockdown of the completeHox paralogous group 1 leads to dramatic hindbraina n d n e u r a l c r e s t d e f e c t s .Development.;132(12):2861-287151. Michaut L, Jansen H, Bardine N, Durston A,Gehring WJ, (2011) Analysing the function of a hoxgene: an evolutionary approach. Evolution andDevelopment under review52. Miller DF, Rogers BT, Kalkenbrenner A, HamiltonB, Holtzman SL, Kaufman T.(2001) Cross-regulationof Hox genes in the Drosophila melanogaster embryo.Mech Dev.102(1-2):3-16.53. Mito T, Okamoto H, Shinahara W, Shinmyo Y,Miyawaki K, Ohuchi H, Noji S.Krüppel acts as a gap gene regulating expression ofhunchback and even-skipped in the intermediate germcricket Gryllus bimaculatus. Dev Biol. 2006 Jun15;294(2):471-81. Epub 2006 Apr 17.54. Morsi El Kadi A, In Der Rieden P, Durston A,Morgan R, (2002). The small GTPase Rap-1 is animmediate downstream target for Hoxb4transcriptional regulation. Mech. Dev.113, 131-139 55. Nuesslein- Volhard (1995) Nobel lecture 56. Palmeirim I, Henrique D, Ish Horowicz D, PourquieO, (1997) Avian hairy gene expression identifies amolecular clock linked to vertebrate segmentation andsomitogenesis. Cell.;91(5):639-48.57. Pearson JC, Lemons D, McGinnis W, (2005) Modulating Hox Gene Functions During Animal Body
WebmedCentral > Review articles Page 10 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Patterning. Nature Reviews Genetics 6, 893, 58. Peres J, McNulty C, Durston A. (2006) Interactionbetween X-Delta-2 and Hox genes regulatessegmentation and patterning of the anteroposterioraxis. Mech Dev.;123(4):321-333.59. Plaza S, Prince F, Adachi Y, Punzo C, Cribbs DL,Gehring WJ. (2008) Cross-regulatory protein-proteininteractions between Hox and Pax transcription factors.Proc Natl Acad Sci U S A.;105(36):13439-13444. 60. Ronshaugen M, Biemar F, Piel J, Levine M, LaiEC,, (2005).The Drosophila microRNA iab-4 causes adominant homeotic transformation of halteres to wings.Genes Dev.;19(24):2947-2952.61. Rusch DB, Kaufman TC.Regulat ion ofproboscipedia in Drosophila by homeotic selectorgenes.Genetics. 2000 Sep;156(1):183-94.62. Schneider-Maunoury S, Gilardi-Hebenstreit P,Charnay P. How to build a vertebrate hindbrain.Lessons from genetics. C R Acad Sci III. 1998Oct;321(10):819-34.63. Seo HC, Edvardsen RB, Maeland A D, Bjordal M,Jensen MF, Hansen A, Flaat M, Weissenbach J,Lehrach H, Wincker P, (2004). Hox clusterdisintegration with persistent anteroposterior order ofexpression in Oikopleura dioica. Nature 431: 67 -71.64. Soshnikova N, Duboule D, (2009) EpigeneticTemporal Control of Mouse Hox Genes in VivoScience 324, 1320-1323. 65. Struhl, (1983) Role of the esc+ gene product inensuring the selective expression of segment-specifichomeotic genes in Drosophila. J. Embryol. Exp.Morphol. 76:, 297–331 66. Struhl G, White RA. Regulation of the Ultrabithoraxgene of Drosophila by other bithorax complex genes.Cell. 1985;43(2 Pt 1):507-519.67. Tümpel S, Wiedemann LM, Krumlauf R.Hox genesand segmentation of the vertebrate hindbrain. CurrTop Dev Biol. 2009;88:103-3768. Van Der Hoeven F, Zakany J, Duboule D, (1996)Transpositions in the HoxD complex reveal a hierarchyof regulatory controls. Cell.;85(7):1025-1035.69. Wacker SA, Jansen HJ, McNulty CL, Houtzager E,Durston AJ. (2004a). Timed interactions between theHox expressing non-organiser mesoderm and theSpemann organiser generate positional informationdur ing vertebrate gastrulat ion. Dev Biol .;268(1):207-21970. Wellik DM, Capecchi MR, (2003) Hox10 andHox11 genes are required to globally pattern themammalian skeleton, Science 301 363–367.71. Woltering JM, and Durston A, (2008) MiR10represses HoxB1a and HoxB3a in Zebrafish. PLoSONE.;3(1):e1396.72. Woltering Joost M., Freek J. Vonk, Hendrik Müller3,
Nabila Bardine, Merijn A.G. de Bakker, Antony J.Durston & Michael K. Richardson (2009) Molecularregionalisation of the snake and caecilian body plan. Dev Bio. 332(1):82-9. 73. Wright CV. (1993) Hox genes and the hindbrain.Curr Biol. 1993 Sep 1;3(9):618-21.74 . Yek ta S , Sh ih H , Bar te l DP (2004)MicroRNA-directed cleavage of HOXB8mRNA. Science 304(5670): 594–596.75. Yekta S, Tabin CJ, Bartel DP (2008) MicroRNAsin the Hox network: an apparent link to posteriorprevalence Nat Rev Genet. 9(10): 789–796.76. Zakany J, Kmita M, Alarcon P, De La Pompa L,Duboule D (2001) Localized and transient transcriptionof Hox genes suggests a link between patterning andthe segmentation clock. Cell.;106(2):207-217.77. Zakany J, Duboule D. The role of Hox genesduring vertebrate limb development. Curr Opin GenetDev. 2007 Aug;17(4):359-66. Epub 2007 Jul 20.
WebmedCentral > Review articles Page 11 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Illustrations
Illustration 1
Image 1
WebmedCentral > Review articles Page 12 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Fig.1 Hox gene phenotypesThe function of Hox genes is indicated by gain and loss of function phenotypes. The figure shows this inDrosophila and vertebrates. A. A wild type Drosophila fly This has two wings on the anterior thorax andtwo halteres (red arrow) on the posterior thorax. B. A four winged fly, caused by a loss of function mutationin ultrabithorax, a gene for posterior thorax (Lewis, 1995). The halteres are transformed to wings. CAntennapedia mutation: anterior thoracic legs replace antennae on the head, due to a misregulated gain offunction mutation for the gene Antennapedia (a gene for anterior thorax), leading to its expression in thehead segments (Gehring, 1987). D. In vertebrates, mouse genetics has been bedevilled by the fact that thereare 4 Hox clusters, with parallel functions. This once led to the erroneous idea that vertebrate Hox loss offunction mutations have mild phenotypes. In fact, if you knock out all of the paralogues of a particular Hoxparalogue group (pg), or ectopically express a Hox gene this can give a dramatic phenotype. Left: wild typeXenopus hindbrain. This has 8 segments (rhombomeres) 2-8 each express a different combination of Hoxgenes and so have different identities, indicated by the different colours. 1 (white) expresses no Hox genes.Its identity is determined by the gene Gbx2. Middle: hindbrain in Xenopus where Hox pg1 has beenknocked down using morpholinos. The hindbrain is drastically anteriorised to the identity of r1. It is alsoshorter (redrawn from McNulty et al., 2005). Right: Skeletons of two mice. Above: wild type. Below, amouse ectopically expressing HoxC10. The HoxC10 mouse is drastically different. For example, it lacksribs (Mallo et al., 2007). The thoracic vertebrae are posteriorised to abdominal identity. This is becauseHox pg10 controls the transition from tthorax to abdomen, in the vertebral column.
WebmedCentral > Review articles Page 13 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Fig. 2 Hox Spatial and Functional Collinearity The four human and one Drosophila Hox complexes are homologues. The colour coding in Panels A and B shows thecorrespondence between the genomic order of Hox genes in the Hox complexes (A) and their spatial sequence ofexpression and action zones along the main body axis in Drosophila and human (B).
Illustration 2
Image 2
WebmedCentral > Review articles Page 14 of 21
WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Illustration 3
Image 3
WebmedCentral > Review articles Page 15 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
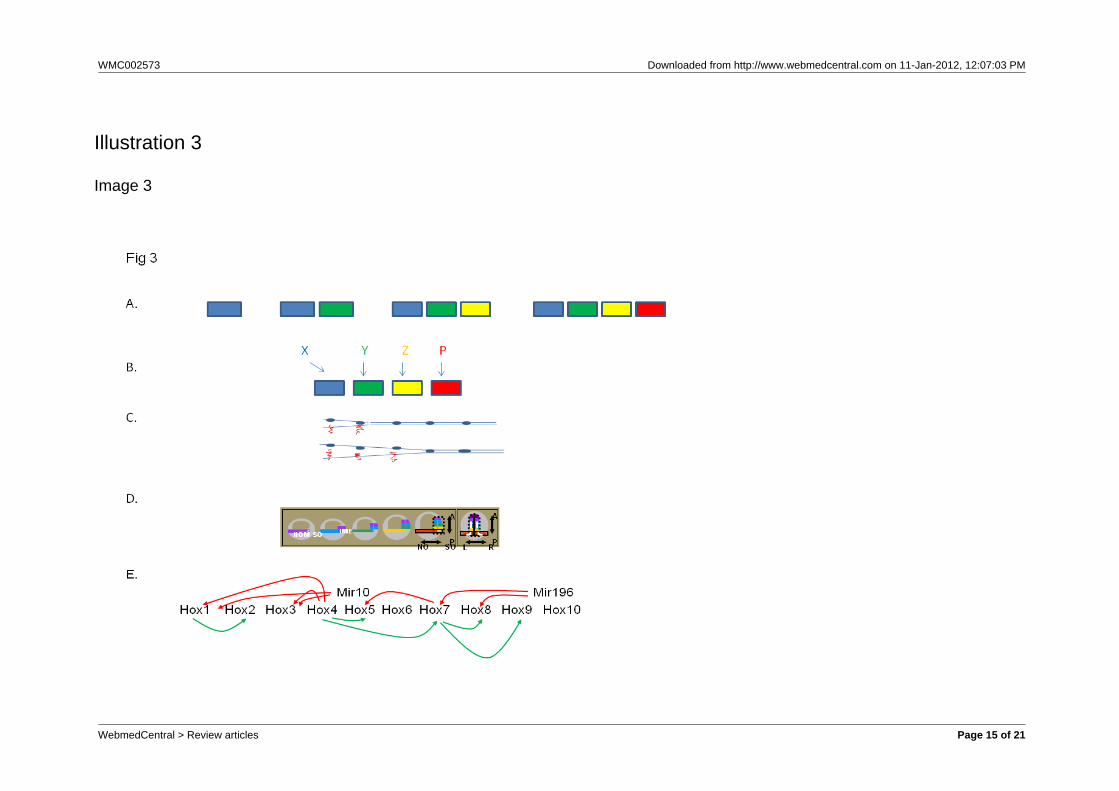
Fig. 3 Some facts and ideas about Hox colinearityA. Tandem duplication and sequential modification. Clustered Hox genes are thought to have evolved bytandem duplication of an ancestral Ur-Hox gene. The duplicates are then thought to have been progressivelymodified, so they become more and more different from each other. The figure shows tandem duplicationand progressive modification towards the right. The ur- Hox gene (left, blue) duplicates and the right handdaughter is modified (green). The green Hox gene duplicates again and its right hand daughter is modified(yellow). The yellow Hox gene duplicates again and its right hand daughter is modified (red). This type ofmechanism can give collinear properties.B. The associated upstream mechanism needed to generate spatial collinearity. If such a Hox cluster is togenerate spatial colinearity without an explicit colinearity mechanism, an individual input is needed to turnon each Hox gene to ensure it is expressed at exactly the right axial position. The inputs concerned aregoing to need an axial pattern themselves. This kind of mechanism is used in Drosophila, where the gapgenes provide the inputs. Gap genes specify the primary axial positions where the Hox genes are expressedand segmentation genes, the Hox genes themselves, polycomb group genes and cofactors like teashirtrefine this information, restricting Hox expression by specific segment boundaries. In this situation, the Hoxgenes thus do not provide the primary axial patterning information. They are secondary. It is likely that thiskind of mechanism is general in invertebrates, which probably have no temporal colinearity or colinearitymechanism and have had to evolve an ad hoc mechanism to generate spatial collinearity. Something likethis may also occur in the vertebrate hindbrain, where the gastrula’s colinearity mechanism is presumablythe primary patterning mechanism and hindbrain genes confirm or alter the patterning information.C. Progressive chromatin opening: the basic idea. This is an idea proposed by Duboule and colleagues toaccount for vertebrate temporal collinearity. The Hox complex chromatin opens from 3’ to 5’. This openingprogressively permits Hox gene transcription, from 3’ to 5’.D. Time- space translation. Vertebrates show early Hox collinearity. There is a temporally collinearsequence of Hox gene expression in the gastrula. This is used to generate a spatially collinear axialsequence of Hox gene expression. For details, see Fig. 4.E. Hox interactions.What regulates vertebrate temporal collinearity? Not just chromatin opening, asproposed by Duboule. The different vertebrate Hox clusters are expressed with synchronous temporalcollinearity. What may be involved are interactions between different Hox genes. The figure shows someinteractions between Hox genes in the vertebrate gastrula.
WebmedCentral > Review articles Page 16 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Illustration 4
Image 4
WebmedCentral > Review articles Page 17 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Fig. 4 Temporal Collinearity AndTime space translation.a) Temporal Collinearity In the Xenopus GastrulaThe figure shows Hox expression patterns at sequential stages during gastrulation in Xenopus. The embryosare seen from underneath, where a ring (the blastopore) shows the position where mesoderm tissueinvaginates during gastrulation. This ring gets smaller as gastrulation proceeds and the upper tissues in theembryo spread out and cover the lower part of the embryo (epiboly).The expression of several differentHox genes, seen as blue colour by in situ hybridisation, is in each case initially in the gastrula mesoderm inthe zone above (outside) the ring. Hox expression is thus seen as a blue ring, and since it is initially only inpart of the mesoderm (non organiser nesoderm), the ring is initially broken. This ring of Hox expressiongets smaller as the blatopore ring gets smaller and mesoderm invaginates into the embryo.The figure showsexpression of a sequence of Hox genes with different paralogue numbers, from 1 to 9. It will be seen thatthe Hox gene with the lowest paralogue number starts expression first and later numbers start sequentiallylater. It will also be seen that the Hox genes in this time sequence include members of all of the 4 primaryvertebrate paralogue groups (a,b,c,d).
b) Time-space translationTimed interactions between the Hox expressing non-organiser mesoderm and the Spemann organiser
WebmedCentral > Review articles Page 18 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
generate positional information during vertebrate gastrulation. The drawings show simplified 2-dimensionalrepresentations of Xenopus gastrulae. The first 5 drawings show parasagittal (ventral to dorsal) twodimensional representations of gastrula profiles, starting at the beginning of gastrulation and then atsequential stages till the end. The last (6th.) drawing shows the end of gastrulation, from the dorsal side(profile at the level of the dorsal axial mesoderm). Hox expressing tissue (NOM (NO and I) and, late ingastrulation neurectoderm (N)) is represented by different colours, each of which represents a different hoxcode. Initially, the coloured bar represents the broken ring of NOM in the wall of the embryo. The laterinternal coloured blocks at the dorsal side of the embryo represent the involuted NOM mesoderm. Thecoloured blocks next to them in the wall of the embryo represent the overlying neurectoderm, which alsocomes to express hox genes. Hox expression is copied from the gastrula mesoderm to the neurectoderm.The SO is shown only in the last drawing, as the heavy median black line. By this stage, it has become thenotochord and a head mesodermal portion. The first 5 drawings represent paraxial profiles, where theorganiser is not available. The black dotted line in the last drawing depicts the sphere of influence of theSO. N: neurectoderm, NO: non-organiser mesoderm; S,: Spemann organiser; A: Anterior; P: Posterior; L:Left; R: Right. N nonorganiser; S Spemann organiser. The white arrows reflect directions of cell movementflow. To dorsal, anterior and internal(drawings 1 and 6). -There is a collinear time sequence of hoxexpression in non involuted non-organiser mesoderm (NOM) in the gastrula (depicted by the spectralsequence of colours). -During gastrulation involution movements continuously bring cells from the NOMinto the inside of the embryo See stack of blocks of different colours, reflecting a history of the collinearhox mesodermal time sequence, in the internal involuted mesoderm. -Stable (ectodermal) Hox expression isinduced by a combination of signals from the SO and the Hox expressing NOM. See corresponding blocksof sequential spectral colours in the gastrula's mesoderm and outer layer, reflecting a vertical transfer of theHox codes from involuted mesoderm to overlying neurectoderm. A “Hox stripe” aspart of the anterior–posterior Hox pattern is thus formed at the dorsal side.
WebmedCentral > Review articles Page 19 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
Fig. 5 Regulators of vertebrate Hox temporal colinearity: Hox-Hox interactions and Somitogenesis oscillationsA. some cross interactions between Hox genes and Mirs in the vertebrate Hox complexes during vertebrategastrulation. Red: repression. Green: activationB. The somitogenesis clock and Hox temporal collinearity. We show an oscillating concentration of XDelta2.Sequential peaks of XDelta2 activate expression of different Hox genes. [XDelta2*]; The threshold concentration ofXDelta2 at which Hox expression is activated.
Illustration 5
Image 5
WebmedCentral > Review articles Page 20 of 21

WMC002573 Downloaded from http://www.webmedcentral.com on 11-Jan-2012, 12:07:03 PM
DisclaimerThis article has been downloaded from WebmedCentral. With our unique author driven post publication peerreview, contents posted on this web portal do not undergo any prepublication peer or editorial review. It iscompletely the responsibility of the authors to ensure not only scientific and ethical standards of the manuscriptbut also its grammatical accuracy. Authors must ensure that they obtain all the necessary permissions beforesubmitting any information that requires obtaining a consent or approval from a third party. Authors should alsoensure not to submit any information which they do not have the copyright of or of which they have transferredthe copyrights to a third party.
Contents on WebmedCentral are purely for biomedical researchers and scientists. They are not meant to cater tothe needs of an individual patient. The web portal or any content(s) therein is neither designed to support, norreplace, the relationship that exists between a patient/site visitor and his/her physician. Your use of theWebmedCentral site and its contents is entirely at your own risk. We do not take any responsibility for any harmthat you may suffer or inflict on a third person by following the contents of this website.
WebmedCentral > Review articles Page 21 of 21