host factors in the resistance of newborn mice to k1 escherichia coli infection
TRANSCRIPT

Microbial Pathogenesis 1988; 4: 93-102
Host factors in the resistance of newborn mice toK1 Escherichia coli infection
Gerd Pluschke and Sinikka Pelkonen
Basel Institute for Immunology, Grenzacherstrasse 487, 4058 Basel, Switzerland
(Received October 19, 1987 ; accepted November 28, 1987)
Pluschke G . (Basel Institute for Immunology, Grenzacherstrasse 487, 4058 Basel, Switzerland)and S. Pelkonen. Host factors in the resistance of newborn mice to K1 Escherichia coli in-fection . Microbial Pathogenesis 1988; 4: 93-102 .
It is not clear which factors are responsible for the deficient resistance of human neonates toK1 E. coli sepsis and meningitis. To evaluate the relative importance of different defensemechanisms against bacterial invasion, we have analyzed the sensitivity of newborn mice withknown immune deficiencies to infection after oral challenge with virulent K1 E. coli. T and Blymphocyte and complement (C5) defects had no significant effect on natural resistance . Incontrast, both endotoxin-hyporesponsive mouse strains tested were highly sensitive . Thissusceptibility to infection was strongly age dependent .
Infant endotoxin-hyporesponsive mice were killed by i .p. injection of less than ten virulentK1 E. coli cells. In contrast, endotoxin-responsive animals and F1 hybrids derived from crossesbetween endotoxin-responsive and hyporesponsive mice survived an injection with up to 10 4bacteria . Mutants of a virulent 01 8:K1 E. coli strain defective in the synthesis of the capsularpolysaccharide or the O-antigen of lipopolysaccharide were avirulent as were 01 :1<1 bacteria,which are under-represented among E. coli isolates from neonatal meningitis . Endotoxin-hyporesponsive mice were protected from lethal bacterial challenge by monoclonal IgG specificfor the O-antigen of the challenge strain or by human recombinant interleukin 1 .
A fulminant bacterial multiplication in the bloodstream of endotoxin-hyporesponsive micewas observed after i .v. injection of 100 virulent K1 E. coli cells. Persistent bacteremia with 105to 10' bacteria per ml of blood resulted in death of the animals one to two days after challenge .In the bloodstream of endotoxin-responsive mice the bacteria proliferated to a comparableextent within the first 6 h after challenge. Thereafter they were rapidly cleared from thecirculation and the animals recovered from the infection .
Key words : Neonatal sepsis and meningitis ; natural resistance ; K1 E. coli; endotoxin .
Introduction
The incidence of bacterial sepsis and meningitis during the first few weeks of life isfar greater than during the rest of the individual's life span . Escherichia coli and groupB streptococci are the most common causes of neonatal invasive bacterial infections .'Most of the E. coli strains isolated from neonatal meningitis possess the K1 capsularantigen.` K1 E. coli have been compared with non-K1 strains and with K_ mutantsin animal disease models.` The results demonstrate that the K1 capsule significantlycontributes to the virulence. Like other sialic acid containing cell surface structures,'"'
Author to whom correspondence should be addressed : Dr Gerd Pluschke, Basel Institute for Immunology,Grenzacherstrasse 487, 4058 Basel, Switzerland .
0882-4010/88/020093+10 $03 .00/0
© 1988 Academic Press Limited

94
G. Pluschke and S . Pelkonen
it is able to impede activation of the alternative complement pathway .' This propertycontributes to the ability of K1 E. coil to resist antibody-independent serum bactericidalactivity' and opsonophagocytosis .' 4-16 Not all K1 strains, however, have the samevirulence properties . 0-group related differences were found when 95 different isolatesof the common 0-antigen groups 01, 07 and 018 were tested for the ability to causebacteremia after colonizing the gut of newborn rats . 17 In contrast to 07 and 018strains, 01 :1<1 E. coil were avirulent . Antibody-independent activation of the classicalcomplement pathway by the 01 antigen seems to be responsible for the serumsensitivity and avirulence of 01 :K1 bacteria for neonates .'" Human epidemiologicaldata, consistent with the animal infection studies, reveal that 07 :K1 and 018:K1bacteria are common and 01 :K1 bacteria are relatively rare among isolates from casesof neonatal meningitis,6,17,19,2° although all three groups of strains are common amongfecal isolates ." Furthermore, 01 :K1 strains are common among isolates from urinarytract infections,' 1,21 where virulence factors other than those in neonatal sepsis andmeningitis seem to play a crucial role . 20
Only neonates (0-4-weeks-old) are highly susceptible to invasion by K1 E. coli., 22,23
In most cases of sepsis neonatorum the infection of the bloodstream has no obviousfocus; spread to the meninges occurs in many of the affected infants .' K1 E. coli aredetected in rectal swabs of 20% to 40% of individuals of all ages, including healthynewborns . 2,24 Although many newborns carry potentially pathogenic K1 strains, onlya few (up to one per 2000 live births) develop E. coli meningitis . Several risk factors,such as obstetrical complications, premature delivery, galactosemia and necrotizingenterocolitis have been recognized, but no obvious risk factor can be associated witha substantial proportion of infected newborns .' A21
In this report the relative importance of different defense mechanisms for naturalresistance to bacterial invasion was evaluated by analysing K1 E. coil infections witha neonatal mouse infection model .
Results
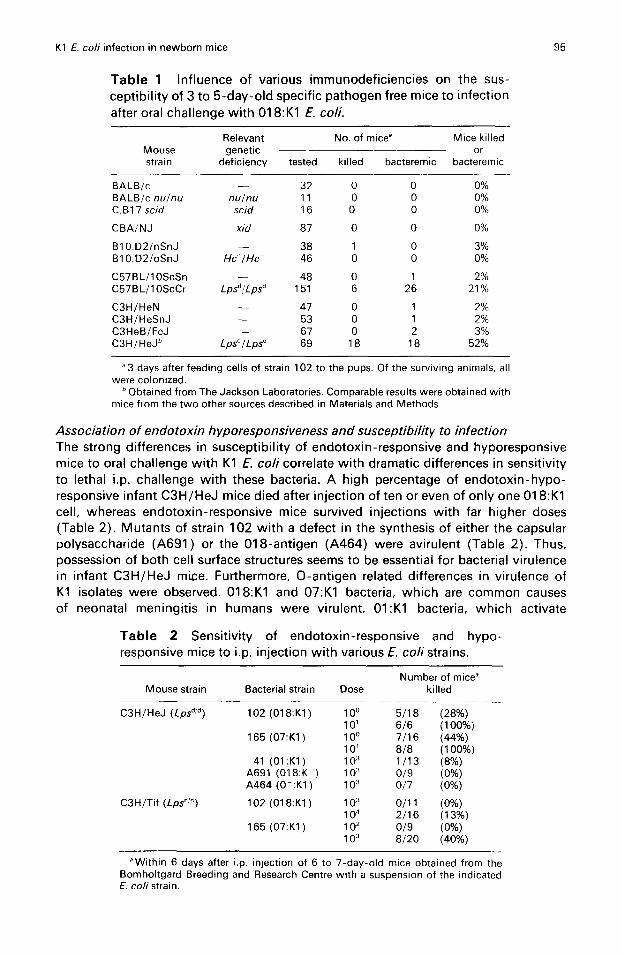
Susceptibility of newborn mice to infection after oral challenge with K1 E . coliThe ability of K1 E. coli bacteria to cause bacteremia and death in newborn mice wastested . A droplet of a suspension of the 018:K1 strain 102 was fed to 3- to 5-day-oldspecific-pathogen-free mice with a plastic pipette tip . Strain 102 is a representativemember of the 018 :K1/OMP6 clonal group, which is common among isolates fromneonatal meningitis . 20 The pups were returned to the mother and 3 days latercolonization, bacteremia and death were scored . All animals tested were colonized . Incontrast to newborn rats, 9 ' 17 newborn mice of most strains tested were relativelyresistant to invasion by the K1 E. coli bacteria (Table 1) . Several strains of mice withknown immunodeficiencies were included in this study . Most of these mutant micewere also resistant . These include (Table 1) : T cell deficient BALB/c nu/nu mice, C.B17scid mice, in which the differentiation of T and B lymphocytes is severely impaired, 25
CBA/NJ mice which carry a X-linked recessive allele designated xid which confers avariety of B-lymphocyte functional abnormalities, and B10 .D2/oSnJ mice with agenetic deficiency of the 5th component of complement . In contrast, both mousestrains tested that express the LpS' phenotype (C3H/HeJ and C57BL/1 OScCr) whichconfers endotoxin-hyporesponsiveness, were susceptible to infection (Table 1) .Closely related endotoxin-responsive C57BL/10ScSn mice and three strains of theC3H lineage with a normal (Lps"/") genotype including C3HeB/FeJ mice, which carrya Salmonella susceptibility gene distinct from Lps,26 were resistant .
K1 E. coli infection in newborn mice
95
Table 1 Influence of various immunodeficiencies on the sus-ceptibility of 3 to 5-day-old specific pathogen free mice to infectionafter oral challenge with 01 8 :K1 E. coli.
3 days after feeding cells of strain 102 to the pups . Of the surviving animals, allwere colonized .
Obtained from The Jackson Laboratories . Comparable results were obtained withmice from the two other sources described in Materials and Methods .
Association of endotoxin hyporesponsiveness and susceptibility to infectionThe strong differences in susceptibility of endotoxin-responsive and hyporesponsivemice to oral challenge with K1 E. coli correlate with dramatic differences in sensitivityto lethal i.p. challenge with these bacteria . A high percentage of endotoxin-hypo-responsive infant C3H/HeJ mice died after injection of ten or even of only one 01 8 :K1cell, whereas endotoxin-responsive mice survived injections with far higher doses(Table 2) . Mutants of strain 102 with a defect in the synthesis of either the capsularpolysaccharide (A691) or the 018-antigen (A464) were avirulent (Table 2) . Thus,possession of both cell surface structures seems to be essential for bacterial virulencein infant C3H/HeJ mice . Furthermore, 0-antigen related differences in virulence ofK1 isolates were observed . 018:K1 and 07 :K1 bacteria, which are common causesof neonatal meningitis in humans were virulent . 01 :K1 bacteria, which activate
Table 2 Sensitivity of endotoxin-responsive and hypo-responsive mice to i .p. injection with various E. coli strains .
'Within 6 days after i .p . injection of 6 to 7-day-old mice obtained from theBomholtgard Breeding and Research Centre with a suspension of the indicatedE. coli strain .
Mouse strain Bacterial strain DoseNumber of mice'
killed
C3H/HeJ (Lps d,'d ) 102 (018:K1) 100 5/18 (28%)101 6/6 (100%)
165 (07:K1) 100 7/16 (44%)101 8/8 (100%)
41 (01 :K1) 103 1/13 (8%)A691 (018 :K ) 103 0/9 (0%)A464 (0 -:K1) 10, 0/7 (0%)
C3H/Tif (Lps ") 102 (018:K1) 103 0/11 (0%)104 2/16 (13%)
165 (07:K1) 102 0/9 (0%)103 8/20 (40%)
Mousestrain
Relevantgenetic
deficiency
No. of mice' Mice killedor
bacteremictested killed bacteremic
BALB/c - 32 0 0 0%BALB/c nu/nu nu/nu 1 1 0 0 0%C.B17 scid scid 16 0 0 0%
CBA/NJ xid 87 0 0 0%
B10.D2/nSnJ 38 1 0 3%B10.D2/oSnJ Hc"/Hc 46 0 0 0%
C57BL/10ScSn 48 0 1 2%C57BL/10ScCr Lpsd/Lpsd 151 6 26 21%
C3H/HeN 47 0 1 2%C3H/HeSnJ 53 0 1 2%C3HeB/FeJ 67 0 2 3%C3H/HeJ 0 Lpsd/Lps d 69 18 18 52%

96
Table 3 Correlation between endotoxin-hypo-responsiveness and resistance to i .p. injectionwith 018:K1 E. coli .
' C3H/HeJ = HeJ ; C3H/OuJ = OuJ ; C57BL/1 OSnJ =SnJ; C57BL/1 OScCr = ScCr . The C3H/HeJ mice used inthis experiment were obtained from the Bomholtgard Breed-ing and Research Centre .'As determined 6 days after i .p . injection of 10-day-old
mice with cells of strain 102 .
complement' $ and rarely cause invasive diseases in human neonates were also avirulentin newborn C3H/HeJ mice .
Further evidence for the association of the Lpsdid genotype with susceptibility to K1E. coli infection was obtained by experiments conducted with F1 hybrids fromendotoxin-responsive and hyporesponsive strains of the C3H and the C57BL/10lineage. The Lps gene defects in C3H/HeJ and C57BL/lOScCr mice seem to be theresult of mutational events that occurred independently ." F1 hybrids derived fromcrosses between these two mutant strains remain hyporesponsive to endotoxin, 27
suggesting that both strains carry defective alleles at the same Lps locus. F1 hybridsderived from crosses between endotoxin-responsive and hyporesponsive mice yieldeither intermediate or high-responder offspring, depending on the activity assessedand/or the strains used ." Regardless of the genetic background, all F1 hybridsderived from crosses between Lpsd l d mice and endotoxin-responsive C57BL/1 OSnJ orC3H/OuJ mice were as resistant to K1 E. coli infection as animals homozygous forthe Lps" allele (Table 3) . Accordingly, if the susceptibility gene of C3H/HeJ (Lpsd id )
and C57BL/lOScCr (Lpsd l d ) mice differs, F1 hybrids derived from crosses betweenthese two strains should be resistant . These F1 hybrids were, however, as sensitive asthe two parental strains (Table 3) . These results suggest that the Lpsd mutation conferssusceptibility to invasion by K1 E. coli.
Factors affecting the susceptibility of C3H/HeJ mice to infectionResistance of C3H/HeJ mice to infection increased with age (Table 4) . Adult animalssurvived injections with high doses of 01 8 :K1 E. coil. This age-dependent increase inresistance is not dependent on contact of the animals with enteric bacteria, sincegerm-free adult C3H/HeJ mice also survived high doses of i .p . injected bacteria (Table4) . In newborn rats we have observed slight differences in the virulence of different018:K1 E. coli isolates ." These differences were more pronounced in adult C3H/HeJ
G. Pluschke and S . Pelkonen
Mice' Lps genotype LD5Ob
(HeJxScCr)F, d/d <10(ScCr x HeJ) F, d/d <10C3H/HeJ d/d <10C57BL/10ScCr d/d <10HeJxOuJ)F, d/n 5x10'(HeJxSnJ)F, d/n 9x10'(ScCr x OuJ) F, d/n 2 x 10'(OuJ x HeJ)F, n/d 5 x 10'(OuJ x ScCr) F, n/d 7 x 10°(SnJ x HeJ)F, n/d 7 x 10`(SnJ x ScCr) F, n/d 7 x 10'(SnJ x OuJ) F, n/n 7 x 10 4C3H/OuJ n/n 6 x 10 4C57BL/1 OSnJ n/n 9 x 10°

K1 E. coli infection in newborn mice
Table 4 Age dependence of the sensitivity to i .p. injectionwith 018:K1 E. coli
'Within 5 days after i .p. injection with cells of strain 102 .
Table 5 Protection of C3H/HeJ miceagainst infection with 01 8 :K1 E. coli by injec-tion with human recombinant interleukin 1or with monoclonal IgG2b against the 018antigen
'10-day-old C3H/HeJ mice were challenged i .p . with100 K1 E. coli cells . Simultaneously they were injectedwith 20 yl of saline (s .c .) saline containing monoclonalantibodies (s .c .) or saline containing 500 U of IL-1 (i .p .) .
'Within 6 days after challenge .
mice (data not shown) . Some bacterial strains had a lower LD 50 for i .p. injection thanthe 01 8:K1 strain 102 used for most of the experiments described in this report . Thesedifferences will be investigated further .
Monoclonal IgG specific for the 018 antigen protected newborn C3H/HeJ micefrom being killed by i .p. injection with 018 :K1 bacteria (Table 5) . The protective effectof the anti-018 IgG was specific, since no protection was observed when the animalswere challenged with 07 :K1 bacteria . Protection could also be achieved by injectionof the mice with human recombinant interleukin 1 (Table 5) .
Influence of the Lps gene on bacterial multiplication in the bloodstreamOne-hundred 018 :K1 cells were injected i .v. into ten-day-old mice . Independently ofthe Lps phenotype a fulminant multiplication of the bacteria was observed for the first6 h after challenge (Fig . 1) . In endotoxin-hyporesponsive C3H/HeJ mice the bacterial
97
Mouse strain AgeDose ofbacteria
Number ofanimals killed'
C3H/HeN (Lps"i") 4d 10 3 0/13 (0%)10 4 7/7 (100%)
10d 104 0/12 (0%)10 5 0/13 (0%)
C3H/HeJ (Lps' ) 10d 10' 14/15 (93%)19d 10 1 0/11 (0%)
10 2 21/35 (60%)adult 10" 0/15 (0%)
10 5 4/8 (50%)adult germ-free 10 3 0/5 (0%)
10 4 0/5 (0%)
Challengedstrain Injected agent
Number ofmice killed'
102 (018:K1) saline 57/58 (99%)heat inactivated rIL1 13/14 (93%)
active rlLl 0/23 (0%)
5 pg anti-018 IgG 0/7 (0%)1 pg anti-018 IgG 0/6 (0%)
0.5 pg anti-018 IgG 1 /8 (13%)0.1 pg anti-018 IgG 5/8 (63%)
165 (07 :K1) saline 10/11 (91%)5 pg anti-018 IgG 11 /11 (100%)

98
G. Pluschke and S . Pelkonen
10
Time (hours)
Fig . 1 . Multiplication of i .v . injected bacteria in the bloodstream of infant mice . Ten-day-old C3H/HeJ(49) or C3H/OuJ (0) mice were injected i .v . with 100 01 8 :K1 E. coli cells . At different times after injectionanimals were killed and the blood concentration of viable bacteria was determined . The blood of at leastfive animals was tested per time point and the arithmetic mean counts ± standard error are given .
concentration reached a level of about 105-106 cells per ml of blood . This bacteremiapersisted and the animals died between the first and second day after challenge . Incontrast, about 6 h after challenge the concentration of viable bacteria in the blood ofendotoxin -responsive C3H/OuJ mice started to decrease dramatically (Fig . 1) . Twenty-four hours after injection no viable bacteria were left and all of these animals survivedthe challenge .
Discussion
The results in this report indicate that inflammatory responses induced by LPS arehighly important for resistance to neonatal infection by K1 E. coli. Mice with the Lpsdid
genotype which are highly resistant to the toxicity of endotoxin and refractory to otherbiological effects of purified LPS were highly sensitive to K1 E. coli infections . F1hybrids derived from crosses between endotoxin-responsive and hyporesponsive micewere as resistant as mice homozygous for the Lps" allele. Similar to these findings, ithas been shown in a mouse model of ascending urinary tract infection that impairedclearance of uropathogenic E. coilfrom the kidney of C3H/HeJ compared to C3H/HeNmice correlates with endotoxin-hyporesponsiveness . 29 Such a difference was foundonly with LPS-containing bacteria . 30 In this model system an inverse relationshipexists between the recruitment of neutrophils to the urinary tract and persistance of E.coil cells in the kidney ." Furthermore, the Lpsdld genotype also seems to be linked tosusceptibility to Salmonella typhimurium ." This facultative intracellular pathogen is

K1 E. coli infection in newborn mice
99
able to proliferate in apparently uncontrolled fashion in the liver and spleen ofendotoxin-hyporesponsive mice . Here susceptibility has been attributed to the inabilityof Lpsd i d macrophages to be activated by LPS . 32
Host defenses rapidly activated on contact with LPS thus seem to be crucial elementsof host resistance to Gram-negative bacteria in general . It seems likely that differenteffector mechanisms are responsible for restriction of intracellular growth of S.typhimurium in the reticuloendothelial system, extracellular multiplication of K1 E. coliin the bloodstream of infant animals or the localized persistence of uropathogenic E.coli in the kidney. With the different infection models now available it should bepossible to sort out which of the many effects of LPS play a role in natural resistanceto Gram-negative bacterial pathogens . The availability of pure effector substancessuch as interleukins may help to elucidate the relevant defense mechanisms . Forexample, it is shown in this report that interleukin 1 can substitute at least partially forthe lack of response to LPS . The Lpsd mouse model for K1 E. coli infections sharesfeatures such as serotype specificity and age dependence of susceptibility of the hostwith the infection of human newborns . Compared to the infant rat model, it has theadvantage that endotoxin, which is a common contaminant even of sterile laboratorymaterial, is not likely to influence the results of protection experiments .
It should be noted that human newborns that have developed a K1 E. coli meningitisseem to be able to raise an acute phase response ." The LPS responsiveness oflymphoreticular cells from newborns prone or not prone to K1 E. coil infection has,however, to our knowledge, never been compared experimentally .As expected with respect to the fulminating course of the infection, B- and T-cell
defects of the newborn mice did not play a role in the natural resistance to virulent K1bacteria . On the other hand, monoclonal antibodies to the 0-antigen of the challengeK1 E. coli strain are highly protective . 34 In contrast to IgG against the K1 capsule, 35
antibodies against 0-antigens of E. coil are commonly found in the sera of healthyadults36 and newborns 37 and it thus seems likely that many of the human newbornscolonized by potentially invasive K1 E. coli are protected against infection by thetransplacental transfer of O-antigen specific maternal IgG . Because neonatal bacterialinvasive infections still produce significant morbidity and mortality in spite of antibiotictreatment, adjunctive immunotherapeutic treatment with pools of antibodies to themost common causative pathogens may become important in the management ofthese infectionS .38
Materials and methods
Bacteria/ strains . The E. coli strains with sequential numbers 41 (01 :K1), 102 (018:K1), 105(018:K1) and 165 (07 :K1) are selected representatives taken from our collection of K1 E. colistrains and have been characterized in detail . 17,111,20,39-41 Strain A691 is a spontaneous mutantof strain 102 and produces no detectable K1 capsule .7,41 Strain A464 is a transposon-inducedmutant of strain 102 and produces no lipopolysaccharide molecules with long 0-antigenchains . 41
Mice. Mice were obtained from Harlan Olac Ltd . (Blackthorn, England; C57BL/10ScSn)from the Jackson Laboratory (Bar Harbor, ME ; C57BL/10SnJ, C3HeB/FeJ, C3H/HeSnJ,C3H/HeJ, C3H/OuJ, B10 .D2/oSnJ, B10 .D2/nSnJ, CBA/NJ), from Charles River Wiga (Sulz-feld, Germany; C3H/HeN) from the Bomhaltgard Breeding and Research Centre Ltd. (Ry,Denmark; C57BL/10ScCr, C3H/HeJ, C3H/Tif) and from the Institut fur biomedizinischeForschung (Fullinsdorf, Switzerland ; BALB/c, BALB/c nu/nu, C3H/HeJ) . C.B17 scid micewere bred at the Basel Institute for Immunology . C3H/HeJ mice obtained from three differentsources yielded comparable results. Unless stated otherwise, experiments with C3H/HeJ mice

100
G. Pluschke and S . Pelkonen
were done with animals from the Institut fur biomedizinische Forschung . Animals were kept inisolators throughout the experiments .
Sensitivity of mice to oral infection ." Three- to five-day-old mice were fed 10 µl of bacterialsuspensions (2 x 108 E. coil cells per ml, grown in L broth) with an Eppendorf plastic pipettetip. The pups were returned to their mothers immediately after feeding. Colonization, bacteremiaand death were scored 3 days after inoculation . To detect colonization, the anal area wassampled. Blood (100 ul per animal) was taken by cardiac puncture of mice anaesthetizedwith carbon dioxide vapours . Samples were cultured on minimal agar plates containingmeningococcus group B antiserum which strongly reacts with the E. coil K1 capsule." Thepurity of bacterial cultures and the identity of K1 bacteria within the samples was establishedby observing halo formation on the plates .
Sensitivity to i.p. injected bacteria . 20 pl of bacterial suspensions were injected i .p. using anEppendorf Multipette with 0.5 ml Combitips and 30 GZ Yale hypodermic needles . In protectionexperiments mice received at the same time 20 pl of saline or saline containing differentconcentrations of the monoclonal antibody BA1 .2 (s .c .) . In other experiments 5000 of humanrecombinant interleukin-1a (rIL-1) were injected together with the bacterial suspension . rIL-1,lot 14993-74/102, 3 x 106 U/mI was a gift of Drs A . Stern and P . T. Lomedico (Hoffmann-LaRoche, Inc., Nutley, NJ) . For control experiments, IL-1 samples were heat-inactivated at 100°Cfor 60 min . Isolation of the rat-mouse hybridoma BA1 .2 and affinity-purification of the ratIgG2b monoclonal antibodies specific for the 018-antigen produced by this cell line by usingsepharose-beads coupled with rhamnose has been described elsewhere .43 LD50 values werecalculated with the bioassay as described 44 after injecting groups of mice with serial 10-folddilutions of bacterial suspensions (at least 10 animals per dilution step) and determination ofthe proportion of animals killed within 6 days after challenge .
Multiplication of i.v . injected bacteria . 50 ul of saline containing 100 cells of strains 105(018:K1) were injected into ten-day-old mice via a tail vein using 30 Gz hypodermic needles .At different times after challenge animals were killed and blood samples were taken by cardiacpuncture. The blood concentration of viable bacteria was determined by distributing serialdilutions of blood on L-plates .
We are greatly indebted to Drs D . Gray, M . Flajnik, and J . Pelkonen for critical reading of themanuscript, to G . Bordmann for technical assistance in the production of monoclonal antibodiesand to Drs A . Stern and P . T. Lomedico from the Hoffmann-La Roche Research Centre, Nutley,NJ, for providing us with recombinant IL-1 . We thank C . Platner and J . Hossmann for typingthe manuscript. The Basel Institute for Immunology was founded and is supported by F .Hoffmann-La Roche & Co . Ltd., CH-4005 Basel, Switzerland .
References
1 . Siegel JD, McCracken GH . Sepsis neonatorum . N Engl J Med 1981 ; 304 : 642-47 .2. Sarff LD, McCracken GH, Schiffer MS, Glode MP, Robbins JB, Orskov I, Orskov F . Epidemiology of
Escherichia coil K1 in healthy and diseased newborns . Lancet 1975 ; is 1099-104 .3. Schiffer MS, Oliveira E, Glode MP, McCracken GH, Sarff LD, Robbins JB . A review : relation between
invasiveness and the K1 capsular polysaccharide of Escherichia coli . Pediatr Res 1976 ; 10 : 82-87 .4. Robbins JB, McCracken GH, Gotschlich EC, Orskov F, Orskov I, Hanson LA . Escherichia coli K1
capsular polysaccharide associated with neonatal meningitis . N Engl J Med 1974; 290 : 1216-20 .5. Handrick W, Steinri ck H, Spencker F-B, Braun W, Vogtmann C . Infektionen durch E. coli in der
Neonatalperiode. Dtsch Gesundheits-wes 1982 ; 52 : 2212-19 .6. Mulder CJJ, van Alphen L, Zanen HC . Neonatal meningitis caused by Escherichia coli in the
Netherlands. J Infect Dis 1984 ; 150: 935-40 .7. Pluschke G, Mayden J, Achtman M, Levine RP . Role of the capsule and the 0 antigen in resistance of
018:K1 Escherichia coli to complement mediated killing. Infect Immun 1983 ; 42: 907-13 .8. Smith HW, Huggins MB . The association of the 018, K1 and H7 antigens and the Co/V plasmid of a
strain of Escherichia coli with its virulence and immunogenecity. J Gen Microbiol 1980; 121 : 387-400.
9. Glode MP, Sutton A, Moxon ER, Robbins JB . Pathogenesis of neonatal Escherichia coli meningitis :induction of bacteremia and meningitis in infant rats fed E. coli K1 . Infect Immun 1977 ; 16 : 75-80 .
10. Edwards MS, Kasper DL, Jennings HJ, Baker CJ, Nicholson-Weller A . Capsular sialic acid prevents

K1 E. coli infection in newborn mice
101
activation of the alternative complement pathway by type III, group B streptococci . J Immunol 1982 ;128:1278-83 .
11 . Jarvis GA, Vedros NA. Sialic acid of group B Neisseria meningitidis regulates alternative complementpathway activation. Infect Immun 1987 ; 55 : 174-80 .
12 . Okada N, Yasuda T, Okada H. Restriction of alternative complement pathway activation by sialosyl-glycolipids . Nature 1982; 299: 261-63 .
13. Pangburn MK, Muller-Eberhard HJ . Complement C3 convertase : cell surface restriction of /f1 H controland generation of restriction on neuraminidase-treated cells. Proc Nat] Acad Sci USA 1978; 75 : 2416--20 .
14. Stevens P, Young LS, Adamu S . Opsonization of various capsular (K) E. coli by the alternativecomplement pathway . Immunology 1983 ; 50 : 497-502 .
15 . Stevens P, Huang S, Welch WD, Young LS . Restricted complement activation by Escherichia coli withthe K1 capsular serotype : a possible role in pathogenicity . J Immunol 1978 ; 121 : 2174-80 .
16. Bortolussi R, Ferried P, Bjorksten B, Quie PG . Capsular K1 polysaccharide of Escherichia coli: relation-ship to virulence in newborn rats and resistance to phagocytosis . Infect Immun 1979; 25 : 293-98 .
17. Pluschke G, Mercer A, Kusecek B, Pohl A, Achtman M . Induction of bacteremia in newborn rats byEscherichia coli K1 is correlated with only certain 0 (lipopolysaccharide) antigen types . Infect Immun1983;39:599-608 .
18. Pluschke G, Achtman M . Degree of antibody-independent activation of the classical complementpathway by K1 Escherichia coli differs with 0 antigen type and correlates with virulence of meningitisin newborns. Infect Immun 1984 ; 43 : 684-92 .
19 . Korhonen TK, Valtonen MV, Parkkinen J . Vaisanen-Rhen V, Finne J, Orskov F, Orskov I, Svenson SB,Makela PH . Serotypes, hemolysin production and receptor recognition of Escherichia coli strainsassociated with neonatal sepsis and meningitis . Infect Immun 1985 ; 48 : 486-91 .
20 . Achtman M, Pluschke G . Clonal analysis of descent and virulence among selected Escherichia coli.Ann Rev Microbiol 1986; 40 :185-210 .
21 . Vaisanen-Rhen V, Elo J, Vaisanen E, Siitonen A, Orskov I, Orskov F, Svenson SB, Makela PH, KorhonenT. P-fimbriated clones among uropathogenic Escherichia coli strains . Infect Immun 1984 ; 43 : 149-55 .
22 . Makela PH . Functional epidemiology of bacterial meningitis . Infection [Suppl .] 1984 ; 12 : S29-34 .23. Glode MP, Sutton A, Robbins JB, McCracken GH, Gotschlich EC, Kaijser B, Hanson LA . Neonatal
meningitis due to Escherichia coli. J Infect Dis [Suppl .] 1977 ; 136: S93-97 .24. Orskov F, Sorensen KB . Escherichia coli serogroups in breast-fed and bottle-fed infants . Acta Pathol
Microbiol Scand Sect B 1975 ; 83 : 25-30 .25. Bosma GC, Custer RP, Bosma MJ . A severe combined immunodeficiency mutation in the mouse .
Nature 1983; 301 : 527-30 .26. O'Brien AD . Influence of host genes on resistance of inbred mice to lethal infection with Salmonella
typhimurium . Curr Top Microbiol Immunol 1986; 124 : 37-48 .27. Coutinho A, Meo T . Genetic basis for unresponsiveness to lipopolysaccharide in C57BL/1 OCr mice .
Immunogenetics 1978 ; 7 : 17-23 .28. Scibienski RJ . Defects in murine responsiveness to bacterial lipopolysaccharide . In : Gershwin ME,
Merchant B, eds, Immunologic defects in laboratory animals, 2 . New York : Plenum, 1981 ; 241-258 .29. Svanborg Eden C, Briles D, Hagberg L, McGhee J, Michalek S . Genetic factors in host resistance to
urinary tract infection . Infection 1984 ; 12 : 118-123 .30 . Hagberg L, Hull R, Hull S, McGhee JR, Michalek SM, Svanborg Eden C . Difference in susceptibility
to gram-negative urinary tract infection between C3H/HeJ and C3H/HeN mice . Infect Immun 1984 ;46:839-44 .
31 . Shahin RD, Engberg I, Hagberg L, Svanborg Eden C . Neutrophil recruitment and bacterial clearancecorrelated with LPS responsiveness in local gram-negative infection . J Immunol 1987 ; 138: 3475-80 .
32. O'Brien AD, Metcalf ES, Rosenstreich DL . Defect in macrophage effector function confers Salmonellatyphimurium susceptibility on C3H/HeJ mice. Cell Immunol 1982; 67 : 325-33 .
33. Pluschke G, Jenni L, van Alphen L, Lefkovits I . Complete two-dimensional gel electrophoresis patternof de novo synthesized acute phase reactants . Clin exp Immunol 1986 ; 66 : 331-39 .
34. Pluschke G, Achtman M . Antibodies to 0-antigen of Iipopolysaccharide are protective against neonatalinfection with Escherichia coli K1 . Infect Immun 1985; 49 : 365-70 .
35. Cross AS, Zollinger W, Mandrell R, Gemski P, Sadoff J . Evaluation of immunotherapeutic approachesfor the potential treatment of infections caused by K1-positive Escherichia coli. J Infect Dis 1983 ; 147 :68-76 .
36. Oiling S, Hanson LA, Holmgren J, Jodel U, Lincoln K, Lindenberg U . The bactericidal effect of normalhuman serum on E. coli strains from normals and from patients with urinary tract infections . Infection1973; 1 : 24-31 .
37. Stephens S . Development of a sensitive solid-phase radioimmunoassay technique for quantification ofclass-specific Escherichia coli antibodies. J Immunol Meth 1984; 71 : 203-10 .
38. Gotoff SP . Immunoprophylaxis and immunotherapy of neonatal group B streptococcal infections .Infection 1984 ; 12 : 151-56 .
39. Achtman M, Mercer A, Kusecek B, Pohl A, Heuzenroeder M, Aaronson W, Sutton A, Silver RP . Sixwidespread bacterial clones among Escherichia coli K1 isolates . Infect Immun 1983; 39 : 315-35 .
40. Ochman H, Selander RK . Evidence for clonal population structure in Escherichia coli. Proc Natl AcadSci USA 1984 ; 81 : 198-201 .

102
G. Pluschke and S . Pelkonen
41 . Kusecek B, Wloch H, Mercer A, Vaisanen V, Pluschke G, Korhonen T, Achtman M . Lipopolysaccharide,capsule and fimbriae as virulence factors among 01, 07, 016, 018 or 075 and K1, K5 or K100 Escherichiacoil. Infect Immun 1984; 43 : 368-79 .
42. Kasper DL, Winkelhake JL, Zollinger WD, Brandt BL, Artenstein MS . Immunochemical similaritybetween polysaccharide antigens of Escherichia coil 07 : K1 (L) : N M and group B Neisseria meningitidis .J Immunol 1973 ; 110: 262-68 .
43. Pluschke G, Bordmann G . Isolation of rat immunoglobulin class switch variants of rat-mouse hybridomasby enzyme-linked immunosorbent assay and sequential sublining . Eur J Immunol 1987 ; 17 : 413-416 .
44. Sachs L. Angewandte Statistik . Springer-Verlag, Berlin, Heidelberg, New York, Tokyo . 1984 ; 179-81 .