honeybee kenyon cells are regulated by a tonic gaba receptor conductance
TRANSCRIPT

doi:10.1152/jn.00180.2014 112:2026-2035, 2014. First published 16 July 2014;J NeurophysiolMary J. Palmer and Jenni Harveyreceptor conductanceHoneybee Kenyon cells are regulated by a tonic GABA
You might find this additional info useful...
55 articles, 22 of which can be accessed free at:This article cites /content/112/8/2026.full.html#ref-list-1
including high resolution figures, can be found at:Updated information and services /content/112/8/2026.full.html
can be found at:Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of November 3, 2014.
American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at http://www.the-aps.org/.(monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2014 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on Novem
ber 3, 2014D
ownloaded from
on Novem
ber 3, 2014D
ownloaded from

Honeybee Kenyon cells are regulated by a tonic GABA receptor conductance
Mary J. Palmer and Jenni HarveyDivision of Neuroscience, Medical Research Institute, University of Dundee, Dundee, United Kingdom
Submitted 6 March 2014; accepted in final form 13 July 2014
Palmer MJ, Harvey J. Honeybee Kenyon cells are regulated by atonic GABA receptor conductance. J Neurophysiol 112: 2026–2035,2014. First published July 16, 2014; doi:10.1152/jn.00180.2014.—The higher cognitive functions of insects are dependent on theirmushroom bodies (MBs), which are particularly large in social insectssuch as honeybees. MB Kenyon cells (KCs) receive multisensoryinput and are involved in associative learning and memory. In additionto receiving sensory input via excitatory nicotinic synapses, KCsreceive inhibitory GABAergic input from MB feedback neurons.Cultured honeybee KCs exhibit ionotropic GABA receptor currents,but the properties of GABA-mediated inhibition in intact MBs arecurrently unknown. Here, using whole cell recordings from KCs inacutely isolated honeybee brain, we show that KCs exhibit a toniccurrent that is inhibited by picrotoxin but not by bicuculline. Bathapplication of GABA (5 �M) and taurine (1 mM) activate a toniccurrent in KCs, but L-glutamate (0.1–0.5 mM) has no effect. The toniccurrent is strongly potentiated by the allosteric GABAA receptormodulator pentobarbital and is reduced by inhibition of Ca2� chan-nels with Cd2� or nifedipine. Noise analysis of the GABA-evokedcurrent gives a single-channel conductance value for the underlyingreceptors of 27 � 3 pS, similar to that of resistant to dieldrin (RDL)receptors. The amount of injected current required to evoke actionpotential firing in KCs is significantly lower in the presence ofpicrotoxin. KCs recorded in an intact honeybee head preparationsimilarly exhibit a tonic GABA receptor conductance that reducesneuronal excitability, a property that is likely to contribute to thesparse coding of sensory information in insect MBs.
honeybee; Kenyon cell; GABA receptor; tonic current
HIGHER COGNITIVE PROCESSING in insects utilizes the mushroombodies (MBs), paired neuronal structures that receive multisen-sory information and are involved in associative learning andmemory (Heisenberg 1998; Menzel 2012). In honeybees, eachMB contains �170,000 Kenyon cells (KCs; Witthöft 1967),with the KC somata densely packed in the center of the medianand lateral MB calyces. The calyceal rims contain the largedendritic arborizations of the KCs, which receive excitatorysynaptic input from sensory projection neurons. KC axonsproject via the MB pedunculus to the �/�-lobes, from wherethey output to multisensory extrinsic neurons (Mobbs 1982). Inaddition, KCs receive GABAergic input from a small numberof MB feedback neurons, each of which extensively innervatesa particular subregion of both the �-lobe and calyceal rim(Mobbs 1982; Bicker et al. 1985; Gronenberg 1987; Rybak andMenzel 1993; Grünewald 1999a; Ganeshina and Menzel2001). GABAergic input to the MBs is known to contribute tothe sparse coding of sensory information in KCs (Perez-Oriveet al. 2002; Papadopoulou et al. 2011; Froese et al. 2013; Leiet al. 2013; Lin et al. 2014). Sparse coding is an efficient and
powerful strategy for processing sensory information in whichonly a small proportion of the neuronal population responds toa given stimulus, with each neuron narrowly tuned in itsresponse selectivity. As sparse coding is also used in vertebratecortexes, it may represent a ubiquitous strategy for higher levelsensory processing (Olshausen and Field 2004).
The vertebrate central nervous system is known to utilizedifferent forms of ionotropic GABA receptor-mediated inhibi-tion for different functions. In addition to phasic inhibition,characterized by rapid transient postsynaptic responses, tonicGABA receptor currents regulate synaptic integration, neuro-nal gain, and network excitability over longer timescales (Se-myanov et al. 2004; Farrant and Nusser 2005). Phasic inhibi-tion and tonic inhibition are mediated by GABAA receptorscontaining different subunits that are localized to synaptic andextrasynaptic sites, respectively (Glykys and Mody 2007;Belelli et al. 2009). However, it is unknown whether insectneurons also use both phasic inhibition and tonic inhibition.The honeybee genome contains orthologs of the DrosophilaGABA receptor subunit resistant to dieldrin (RDL) and theputative GABA receptor subunits GABA and glycine-likereceptor of Drosophila (GRD) and ligand-gated chloride chan-nel homologue 3 (LCCH3; Buckingham et al. 2005; Jones andSattelle 2006). All three subunits are expressed in honeybeebrain, with RDL and LCCH3 being detected in cultured anten-nal lobe neurons (Dupuis et al. 2010). GABA-evoked currentsin these neurons are largely inhibited by picrotoxin (PTX) butare insensitive to bicuculline, a pharmacological profile that isconsistent with the predominant expression of homomericRDL receptors (Buckingham et al. 2005; Dupuis et al. 2010).Similarly, cultured KCs from both honeybees and Drosophilaexhibit GABA receptor currents that are inhibited by PTX butnot bicuculline (Lee et al. 2003; Su and O’Dowd 2003;Grünewald and Wersing 2008). KCs in acutely isolated cock-roach brain exhibit PTX-sensitive IPSPs (Demmer and Klop-penburg 2009), but the properties of ionotropic GABA recep-tors in intact MBs and the types of inhibition that regulate KCfunction are currently not well understood.
To examine the endogenous GABAergic input to honeybeeKCs, we have utilized a recently developed technique to makewhole cell patch-clamp recordings from KCs in acutely iso-lated honeybee brain (Palmer et al. 2013) and extended this toan intact head preparation. We find that honeybee KCs exhibita PTX-sensitive tonic GABA receptor conductance that mod-ulates neuronal excitability, thus providing the first evidencethat tonic inhibition serves a similar function in insect andvertebrate neurons.
METHODS
Whole cell recordings were made from KCs in acutely isolatedhoneybee brain as previously described (Palmer et al. 2013). In brief,
Address for reprint requests and other correspondence: M. J. Palmer,Division of Neuroscience, Medical Research Institute, Ninewells MedicalSchool, Mailbox 6, Univ. of Dundee, Dundee, DD1 9SY, UK (e-mail:[email protected]).
J Neurophysiol 112: 2026–2035, 2014.First published July 16, 2014; doi:10.1152/jn.00180.2014.
2026 0022-3077/14 Copyright © 2014 the American Physiological Society www.jn.org
on Novem
ber 3, 2014D
ownloaded from

adult worker honeybees (Apis mellifera mellifera) were anesthetizedon ice and the intact brain was isolated while submerged in extracel-lular solution (as below). Surrounding tissue and membranes wereremoved by a combination of manual dissection and treatment for 10min with papain (0.3 mg/ml), L-cysteine (1 mg/ml), collagenase (64�g/ml), and dispase (0.7 mg/ml; Husch et al. 2009). The removal ofcovering membranes was necessary to obtain successful whole cellrecordings from KCs. The brain was normally hemisected toreduce animal use, transferred to the recording chamber, securedwith a mesh weight, and continuously perfused with extracellularsolution comprising the following (in mM): 140 NaCl, 5.0 KCl, 1.0MgCl2, 2.5 CaCl2, 4.0 NaHCO3, 1.2 NaH2PO4, 6.0 HEPES, and 14glucose, adjusted to pH 7.4 with NaOH, 326 mosM (Oleskevich1999).
Whole cell KC recordings were also made in an intact honeybeehead preparation. Following cooling and decapitation, the piece ofhead capsule overlying the MB calyces was removed, leaving the restof the head intact. The ocelli and covering membranes were removedby manual dissection and enzyme treatment as above, and the head wastransferred to a custom-built recording chamber. The exposed MB caly-ces were perfused with extracellular solution and visualized usingepi-illumination. All honeybee dissections and neuronal recordingswere performed at room temperature (18–22°C).
Whole cell voltage-clamp and current-clamp recordings were ob-tained from visually identified KCs. Patch pipettes (6–8 M�) werepulled from borosilicate glass and filled with either K-gluconate orCsCl-based intracellular solution (K-gluc. or CsCl int. soln.) compris-ing the following (in mM): 110 K-gluconate, 25 HEPES, 10 KCl, 5MgCl2, 3 Mg-ATP, 0.5 Na-GTP, and 0.5 EGTA, pH 7.2, 284 mosM;and 115 CsCl, 25 HEPES, 10 TEA-Cl, 3 Mg-ATP, 0.5 Na-GTP, and0.5 EGTA, pH 7.2, 270 mosM. Membrane current (Im) and voltage(Vm) were recorded via an EPC-10 patch-clamp amplifier controlledby Patchmaster software (HEKA). Holding potentials (Vh) and mea-sured Vm were corrected after the experiment for the liquid junctionpotential (13 mV with K-gluc. int. soln.; 5 mV with CsCl int. soln.).Passive membrane properties and series resistance (Rs) were measuredfrom the capacitative current response to �10-mV, 10-ms voltagesteps from Vh. Recordings were not used if Im or Vm changes wereaccompanied by changes in Rs. Recorded KCs were only used forexperiments if the capacitative current decayed with a double-expo-nential time course and the input resistance (Ri) was �3 G�. Thistype of KC recording exhibits rapidly adapting action potential (AP)firing and spontaneous membrane currents and is presumed to haveintact neurites (unpublished observations).
Drugs were bath applied via the extracellular solution (flow rate �2ml/min). Transient GABA and L-glutamate-mediated responses wereevoked via pressure application (10–20 psi using a Picospritzer II) ofagonist from a glass micropipette positioned 25–50 �m from therecorded KC soma. Evoked currents were quantified by measurementof charge or peak amplitude.
Im variance (�2) was measured by averaging the squared deviationof points from the current mean, using 10-s current traces that werewell fit by a straight line. For noise analysis to estimate single-channelconductance, mean current amplitude (I) was obtained by subtractingthe PTX-insensitive current component of Im. A plot of �2 against Iwas fit with: �2 � iI � I2/N � b, to yield estimates for i (single-channel current), N (number of receptors), and b (background vari-ance). Single-channel conductance (�) was obtained from � � i/V,with V being the driving force for Cl�.
Offline analysis was performed using IgorPro software (WaveMetrics).Pooled data are expressed as means � SE; n numbers refer to thenumber of tested KCs, each of which was from a different honeybee.Statistical significance was assessed using paired or unpaired Stu-dent’s t-tests as appropriate, with P � 0.05 considered significant.
RESULTS
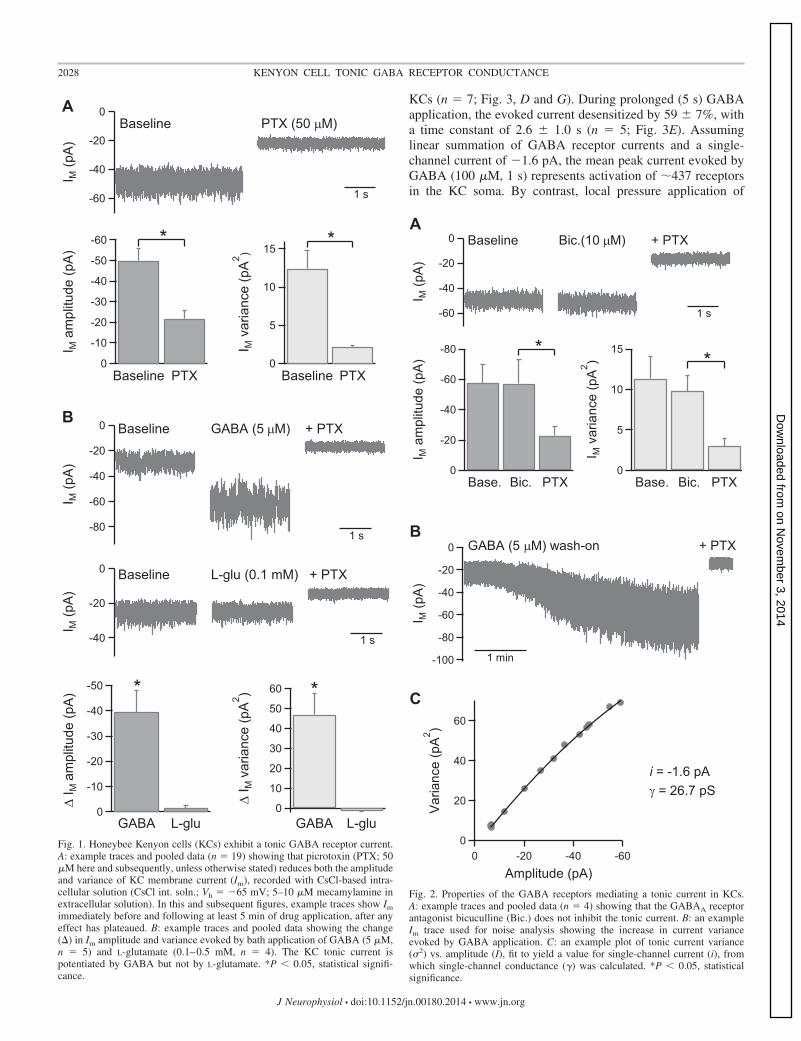
With the use of whole cell voltage-clamp recordings (K-gluc. int. soln., Vh � �73 mV) from visually identified KCs inacutely isolated honeybee brain, it was observed that bathapplication of PTX (50 �M here and subsequently) caused apositive shift in the membrane current (Im), consistent withinhibition of a tonic current. To further examine the propertiesof this current, recordings were made with a high intracellularCl� concentration (CsCl int. soln., Vh � �65 mV) withbath-applied mecamylamine (5–10 �M) to inhibit nicotinicACh receptor currents. Under these conditions, PTX reducedIm amplitude by 28 � 4 pA (n � 19; P � 0.01; Fig. 1A) andnoticeably decreased the recorded current noise. This wasquantified by measuring Im variance, which PTX reduced by10.3 � 2.3 pA2 (n � 19; P � 0.01; Fig. 1A). These effects wereassociated with a 2.5-fold increase in KC input resistance from1.07 � 0.14 to 2.65 � 0.57 G� (n � 15; P � 0.01). HoneybeeKCs therefore exhibit a PTX-sensitive tonic current that con-tributes substantially to the membrane conductance. It was notpossible to distinguish individual synaptic events within thetonic current noise.
PTX antagonizes ionotropic GABA receptors and gluta-mate-activated Cl� channels (GluCl receptors), both of whichare expressed in honeybee KCs (Grünewald and Wersing 2008;El Hassani et al. 2012). To examine the roles of these recep-tors, the effect of bath application of GABA and L-glutamateon KC Im was investigated. GABA (5 �M) increased Imamplitude from �28 � 5 to �68 � 12 pA (n � 5; P � 0.01),which was subsequently reduced to �19 � 6 pA by addition ofPTX (n � 5; P � 0.01; Fig. 1B). GABA also increased Imvariance from 7.2 � 1.8 to 54.3 � 12.0 pA2 (n � 5; P � 0.05),which was subsequently reduced to 1.9 � 0.2 pA2 by PTX(n � 5; P � 0.05). By contrast, L-glutamate (0.1–0.5 mM) hadno significant effect on either Im amplitude or variance (n � 4;Fig. 1B). The tonic current in KCs is therefore likely to bemediated via GABA receptors rather than GluCl receptors.
Most native insect GABA receptors and homomeric RDLreceptors are insensitive to the GABAA receptor antagonistbicuculline (Buckingham et al. 2005). Consistent with this,bicuculline (10 �M) did not inhibit the tonic GABA receptorcurrent in KCs (n � 4; Fig. 2A). To obtain an estimate of thesingle-channel conductance of the receptors mediating thetonic current, noise analysis of current traces during GABA (5�M) wash-on was performed (Fig. 2, B and C; see METHODS fordetails of analysis). Fits of the current variance/amplitude plotsobtained from five KCs gave a single-channel current (i) valueof �1.6 � 0.2 pA, from which single-channel conductance (�)was calculated to be 27 � 3 pS (n � 5).
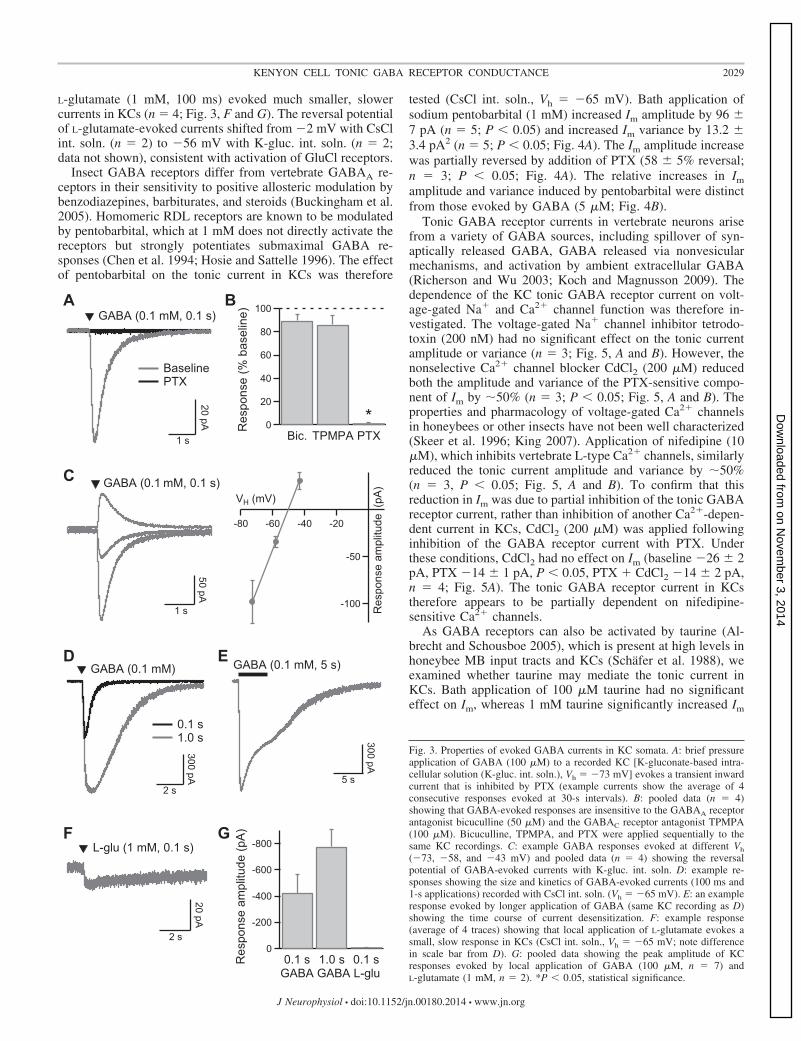
Receptor properties were further investigated by local pres-sure application of GABA to the soma of recorded KCs. Withthe use of K-gluc. int. soln. (Vh � �73 mV), GABA applica-tion (100 �M, 100 ms) evoked rapidly activating and deacti-vating inward currents (Fig. 3A). GABA-evoked currents wereinsensitive to bicuculline (50 �M; n � 4) and the GABACRantagonist TPMPA (100 �M; n � 4) but fully inhibited byPTX (n � 4; P � 0.05; Fig. 3, A and B). They reversed polarityat approximately �50 mV (n � 4; Fig. 3C), close to theexpected Cl� equilibrium potential of �51 mV. With the useof CsCl int. soln. (Vh � �65 mV), local application of GABA(100 �M, 100 ms/1 s) evoked very large inward currents in
2027KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from
KCs (n � 7; Fig. 3, D and G). During prolonged (5 s) GABAapplication, the evoked current desensitized by 59 � 7%, witha time constant of 2.6 � 1.0 s (n � 5; Fig. 3E). Assuminglinear summation of GABA receptor currents and a single-channel current of �1.6 pA, the mean peak current evoked byGABA (100 �M, 1 s) represents activation of �437 receptorsin the KC soma. By contrast, local pressure application of
-80
-60
-40
-20
0
I M (p
A)
1 s
Baseline GABA (5 μM) + PTX
-40
-20
0
I M (p
A)
1 s
Baseline L-glu (0.1 mM) + PTX
-60
-40
-20
0
I M (p
A)
1 s
Baseline PTX (50 μM)
-60
-50
-40
-30
-20
-10
0
I M a
mpl
itude
(pA
)
Baseline PTX
*15
10
5
0
I M v
aria
nce
(pA
2 )
Baseline PTX
*
-50
-40
-30
-20
-10
0 Δ I M
am
plitu
de (p
A)
GABA L-glu
* 60
50
40
30
20
10
0 Δ I M
var
ianc
e (p
A2 )
GABA L-glu
*
A
B
Fig. 1. Honeybee Kenyon cells (KCs) exhibit a tonic GABA receptor current.A: example traces and pooled data (n � 19) showing that picrotoxin (PTX; 50�M here and subsequently, unless otherwise stated) reduces both the amplitudeand variance of KC membrane current (Im), recorded with CsCl-based intra-cellular solution (CsCl int. soln.; Vh � �65 mV; 5–10 �M mecamylamine inextracellular solution). In this and subsequent figures, example traces show Im
immediately before and following at least 5 min of drug application, after anyeffect has plateaued. B: example traces and pooled data showing the change() in Im amplitude and variance evoked by bath application of GABA (5 �M,n � 5) and L-glutamate (0.1–0.5 mM, n � 4). The KC tonic current ispotentiated by GABA but not by L-glutamate. *P � 0.05, statistical signifi-cance.
A
B
-60
-40
-20
0
I M (p
A)
1 s
Baseline Bic.(10 μM) + PTX
-80
-60
-40
-20
0
I M a
mpl
itude
(pA
)
Base. Bic. PTX
* 15
10
5
0
I M v
aria
nce
(pA
2 )
Base. Bic. PTX
*
C60
40
20
0
Var
ianc
e (p
A2 )
-60-40-200Amplitude (pA)
i = -1.6 pAγ = 26.7 pS
-100
-80
-60
-40
-20
0
I M (p
A)
1 min
GABA (5 μM) wash-on + PTX
Fig. 2. Properties of the GABA receptors mediating a tonic current in KCs.A: example traces and pooled data (n � 4) showing that the GABAA receptorantagonist bicuculline (Bic.) does not inhibit the tonic current. B: an exampleIm trace used for noise analysis showing the increase in current varianceevoked by GABA application. C: an example plot of tonic current variance(�2) vs. amplitude (I), fit to yield a value for single-channel current (i), fromwhich single-channel conductance (�) was calculated. *P � 0.05, statisticalsignificance.
2028 KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from
L-glutamate (1 mM, 100 ms) evoked much smaller, slowercurrents in KCs (n � 4; Fig. 3, F and G). The reversal potentialof L-glutamate-evoked currents shifted from �2 mV with CsClint. soln. (n � 2) to �56 mV with K-gluc. int. soln. (n � 2;data not shown), consistent with activation of GluCl receptors.
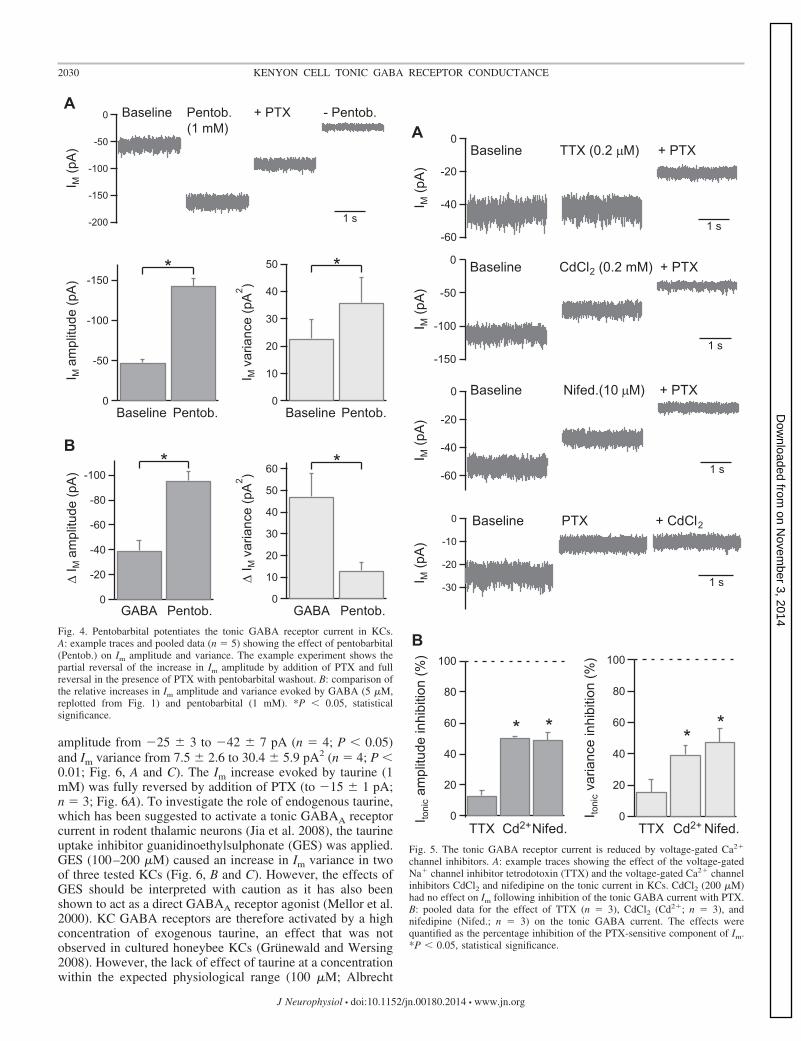
Insect GABA receptors differ from vertebrate GABAA re-ceptors in their sensitivity to positive allosteric modulation bybenzodiazepines, barbiturates, and steroids (Buckingham et al.2005). Homomeric RDL receptors are known to be modulatedby pentobarbital, which at 1 mM does not directly activate thereceptors but strongly potentiates submaximal GABA re-sponses (Chen et al. 1994; Hosie and Sattelle 1996). The effectof pentobarbital on the tonic current in KCs was therefore
tested (CsCl int. soln., Vh � �65 mV). Bath application ofsodium pentobarbital (1 mM) increased Im amplitude by 96 �7 pA (n � 5; P � 0.05) and increased Im variance by 13.2 �3.4 pA2 (n � 5; P � 0.05; Fig. 4A). The Im amplitude increasewas partially reversed by addition of PTX (58 � 5% reversal;n � 3; P � 0.05; Fig. 4A). The relative increases in Im
amplitude and variance induced by pentobarbital were distinctfrom those evoked by GABA (5 �M; Fig. 4B).
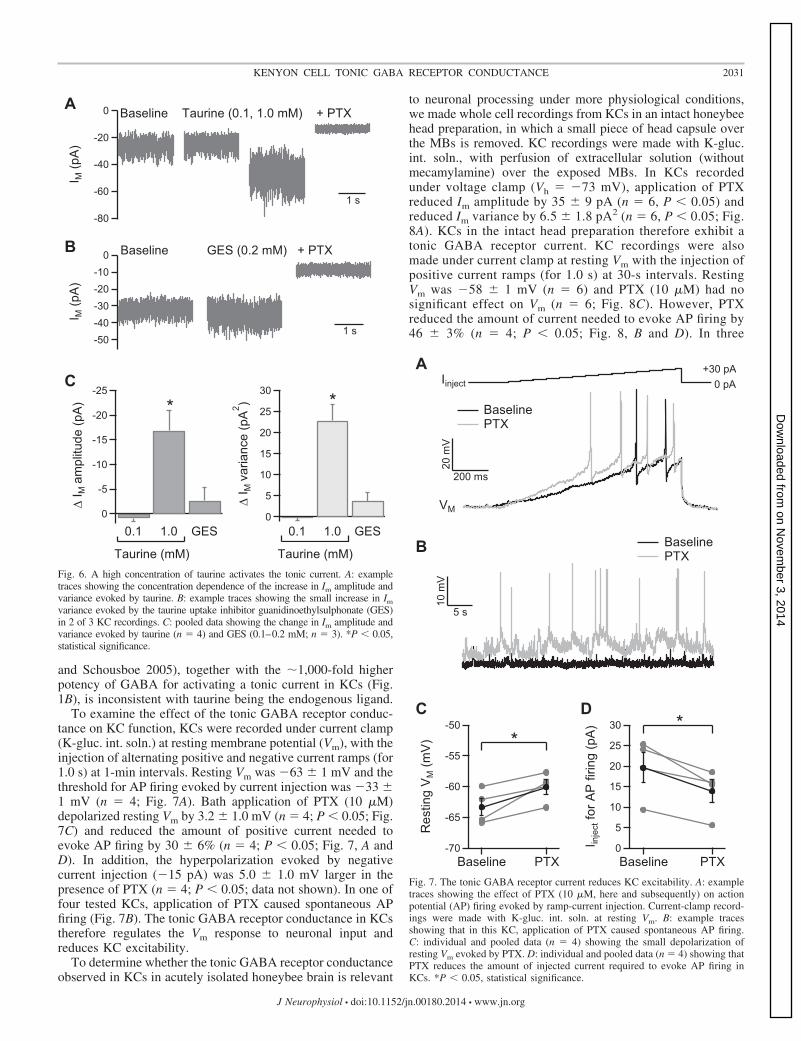
Tonic GABA receptor currents in vertebrate neurons arisefrom a variety of GABA sources, including spillover of syn-aptically released GABA, GABA released via nonvesicularmechanisms, and activation by ambient extracellular GABA(Richerson and Wu 2003; Koch and Magnusson 2009). Thedependence of the KC tonic GABA receptor current on volt-age-gated Na� and Ca2� channel function was therefore in-vestigated. The voltage-gated Na� channel inhibitor tetrodo-toxin (200 nM) had no significant effect on the tonic currentamplitude or variance (n � 3; Fig. 5, A and B). However, thenonselective Ca2� channel blocker CdCl2 (200 �M) reducedboth the amplitude and variance of the PTX-sensitive compo-nent of Im by �50% (n � 3; P � 0.05; Fig. 5, A and B). Theproperties and pharmacology of voltage-gated Ca2� channelsin honeybees or other insects have not been well characterized(Skeer et al. 1996; King 2007). Application of nifedipine (10�M), which inhibits vertebrate L-type Ca2� channels, similarlyreduced the tonic current amplitude and variance by �50%(n � 3, P � 0.05; Fig. 5, A and B). To confirm that thisreduction in Im was due to partial inhibition of the tonic GABAreceptor current, rather than inhibition of another Ca2�-depen-dent current in KCs, CdCl2 (200 �M) was applied followinginhibition of the GABA receptor current with PTX. Underthese conditions, CdCl2 had no effect on Im (baseline �26 � 2pA, PTX �14 � 1 pA, P � 0.05, PTX � CdCl2 �14 � 2 pA,n � 4; Fig. 5A). The tonic GABA receptor current in KCstherefore appears to be partially dependent on nifedipine-sensitive Ca2� channels.
As GABA receptors can also be activated by taurine (Al-brecht and Schousboe 2005), which is present at high levels inhoneybee MB input tracts and KCs (Schäfer et al. 1988), weexamined whether taurine may mediate the tonic current inKCs. Bath application of 100 �M taurine had no significanteffect on Im, whereas 1 mM taurine significantly increased Im
A B
C
D
F G
2 s
20 pA
L-glu (1 mM, 0.1 s)
1 s50 pA
GABA (0.1 mM, 0.1 s)
-800
-600
-400
-200
0
Res
pons
e am
plitu
de (p
A)
0.1 s 1.0 s 0.1 sGABA GABA L-glu
E
2 s
300 pA
GABA (0.1 mM)
0.1 s 1.0 s
100
80
60
40
20
0Res
pons
e (%
bas
elin
e)
Bic. TPMPA PTX*
1 s
20 pA
GABA (0.1 mM, 0.1 s)
Baseline PTX
-100
-50
Res
pons
e am
plitu
de (
pA)
-80 -60 -40 -20
VH (mV)
5 s
300 pA
GABA (0.1 mM, 5 s)
Fig. 3. Properties of evoked GABA currents in KC somata. A: brief pressureapplication of GABA (100 �M) to a recorded KC [K-gluconate-based intra-cellular solution (K-gluc. int. soln.), Vh � �73 mV] evokes a transient inwardcurrent that is inhibited by PTX (example currents show the average of 4consecutive responses evoked at 30-s intervals). B: pooled data (n � 4)showing that GABA-evoked responses are insensitive to the GABAA receptorantagonist bicuculline (50 �M) and the GABAC receptor antagonist TPMPA(100 �M). Bicuculline, TPMPA, and PTX were applied sequentially to thesame KC recordings. C: example GABA responses evoked at different Vh
(�73, �58, and �43 mV) and pooled data (n � 4) showing the reversalpotential of GABA-evoked currents with K-gluc. int. soln. D: example re-sponses showing the size and kinetics of GABA-evoked currents (100 ms and1-s applications) recorded with CsCl int. soln. (Vh � �65 mV). E: an exampleresponse evoked by longer application of GABA (same KC recording as D)showing the time course of current desensitization. F: example response(average of 4 traces) showing that local application of L-glutamate evokes asmall, slow response in KCs (CsCl int. soln., Vh � �65 mV; note differencein scale bar from D). G: pooled data showing the peak amplitude of KCresponses evoked by local application of GABA (100 �M, n � 7) andL-glutamate (1 mM, n � 2). *P � 0.05, statistical significance.
2029KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from
amplitude from �25 � 3 to �42 � 7 pA (n � 4; P � 0.05)and Im variance from 7.5 � 2.6 to 30.4 � 5.9 pA2 (n � 4; P �0.01; Fig. 6, A and C). The Im increase evoked by taurine (1mM) was fully reversed by addition of PTX (to �15 � 1 pA;n � 3; Fig. 6A). To investigate the role of endogenous taurine,which has been suggested to activate a tonic GABAA receptorcurrent in rodent thalamic neurons (Jia et al. 2008), the taurineuptake inhibitor guanidinoethylsulphonate (GES) was applied.GES (100–200 �M) caused an increase in Im variance in twoof three tested KCs (Fig. 6, B and C). However, the effects ofGES should be interpreted with caution as it has also beenshown to act as a direct GABAA receptor agonist (Mellor et al.2000). KC GABA receptors are therefore activated by a highconcentration of exogenous taurine, an effect that was notobserved in cultured honeybee KCs (Grünewald and Wersing2008). However, the lack of effect of taurine at a concentrationwithin the expected physiological range (100 �M; Albrecht
A
B
-200
-150
-100
-50
0
I M (p
A)
1 s
Baseline Pentob. + PTX - Pentob.
-150
-100
-50
0
I M a
mpl
itude
(pA
)
Baseline Pentob.
* 50
40
30
20
10
0
I M v
aria
nce
(pA
2 )
Baseline Pentob.
*
-100
-80
-60
-40
-20
0
Δ I M
am
plitu
de (p
A)
GABA Pentob.
* 60
50
40
30
20
10
0
Δ I M
var
ianc
e (p
A2 )
GABA Pentob.
*
(1 mM)
Fig. 4. Pentobarbital potentiates the tonic GABA receptor current in KCs.A: example traces and pooled data (n � 5) showing the effect of pentobarbital(Pentob.) on Im amplitude and variance. The example experiment shows thepartial reversal of the increase in Im amplitude by addition of PTX and fullreversal in the presence of PTX with pentobarbital washout. B: comparison ofthe relative increases in Im amplitude and variance evoked by GABA (5 �M,replotted from Fig. 1) and pentobarbital (1 mM). *P � 0.05, statisticalsignificance.
A
B
-60
-40
-20
0
I M (p
A)
1 s
Baseline TTX (0.2 μM) + PTX
-60
-40
-20
0
I M (p
A)
1 s
Baseline Nifed.(10 μM) + PTX
-150
-100
-50
0
I M (p
A)
1 s
Baseline CdCl2 (0.2 mM) + PTX
-30
-20
-10
0
I M (p
A)
1 s
Baseline PTX + CdCl2
100
80
60
40
20
0I toni
c var
ianc
e in
hibi
tion
(%)
TTX Cd2+ Nifed.
**
100
80
60
40
20
0I toni
c am
plitu
de in
hibi
tion
(%)
TTX Cd2+Nifed.
* *
Fig. 5. The tonic GABA receptor current is reduced by voltage-gated Ca2�
channel inhibitors. A: example traces showing the effect of the voltage-gatedNa� channel inhibitor tetrodotoxin (TTX) and the voltage-gated Ca2� channelinhibitors CdCl2 and nifedipine on the tonic current in KCs. CdCl2 (200 �M)had no effect on Im following inhibition of the tonic GABA current with PTX.B: pooled data for the effect of TTX (n � 3), CdCl2 (Cd2�; n � 3), andnifedipine (Nifed.; n � 3) on the tonic GABA current. The effects werequantified as the percentage inhibition of the PTX-sensitive component of Im.*P � 0.05, statistical significance.
2030 KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from
and Schousboe 2005), together with the �1,000-fold higherpotency of GABA for activating a tonic current in KCs (Fig.1B), is inconsistent with taurine being the endogenous ligand.
To examine the effect of the tonic GABA receptor conduc-tance on KC function, KCs were recorded under current clamp(K-gluc. int. soln.) at resting membrane potential (Vm), with theinjection of alternating positive and negative current ramps (for1.0 s) at 1-min intervals. Resting Vm was �63 � 1 mV and thethreshold for AP firing evoked by current injection was �33 �1 mV (n � 4; Fig. 7A). Bath application of PTX (10 �M)depolarized resting Vm by 3.2 � 1.0 mV (n � 4; P � 0.05; Fig.7C) and reduced the amount of positive current needed toevoke AP firing by 30 � 6% (n � 4; P � 0.05; Fig. 7, A andD). In addition, the hyperpolarization evoked by negativecurrent injection (�15 pA) was 5.0 � 1.0 mV larger in thepresence of PTX (n � 4; P � 0.05; data not shown). In one offour tested KCs, application of PTX caused spontaneous APfiring (Fig. 7B). The tonic GABA receptor conductance in KCstherefore regulates the Vm response to neuronal input andreduces KC excitability.
To determine whether the tonic GABA receptor conductanceobserved in KCs in acutely isolated honeybee brain is relevant
to neuronal processing under more physiological conditions,we made whole cell recordings from KCs in an intact honeybeehead preparation, in which a small piece of head capsule overthe MBs is removed. KC recordings were made with K-gluc.int. soln., with perfusion of extracellular solution (withoutmecamylamine) over the exposed MBs. In KCs recordedunder voltage clamp (Vh � �73 mV), application of PTXreduced Im amplitude by 35 � 9 pA (n � 6, P � 0.05) andreduced Im variance by 6.5 � 1.8 pA2 (n � 6, P � 0.05; Fig.8A). KCs in the intact head preparation therefore exhibit atonic GABA receptor current. KC recordings were alsomade under current clamp at resting Vm with the injection ofpositive current ramps (for 1.0 s) at 30-s intervals. RestingVm was �58 � 1 mV (n � 6) and PTX (10 �M) had nosignificant effect on Vm (n � 6; Fig. 8C). However, PTXreduced the amount of current needed to evoke AP firing by46 � 3% (n � 4; P � 0.05; Fig. 8, B and D). In three
A
C
-80
-60
-40
-20
0
I M (p
A)
1 s
Baseline Taurine (0.1, 1.0 mM) + PTX
-50-40-30-20-10
0
I M (p
A)
1 s
Baseline GES (0.2 mM) + PTX B
-25
-20
-15
-10
-5
0 Δ I M
am
plitu
de (p
A)
0.1 1.0 GES
*
Taurine (mM)
30
25
20
15
10
5
0
Δ I M
varia
nce
(pA
2 )
0.1 1.0 GES
*
Taurine (mM)Fig. 6. A high concentration of taurine activates the tonic current. A: exampletraces showing the concentration dependence of the increase in Im amplitude andvariance evoked by taurine. B: example traces showing the small increase in Im
variance evoked by the taurine uptake inhibitor guanidinoethylsulphonate (GES)in 2 of 3 KC recordings. C: pooled data showing the change in Im amplitude andvariance evoked by taurine (n � 4) and GES (0.1–0.2 mM; n � 3). *P � 0.05,statistical significance.
A
B
C D
200 ms
20 m
V
Baseline PTX
Iinject
VM
+30 pA 0 pA
5 s
10 m
V
Baseline PTX
-70
-65
-60
-55
-50
Res
ting
VM
(mV
)
Baseline PTX
*30
25
20
15
10
5
0I inje
ct fo
r AP
firin
g (p
A)
Baseline PTX
*
Fig. 7. The tonic GABA receptor current reduces KC excitability. A: exampletraces showing the effect of PTX (10 �M, here and subsequently) on actionpotential (AP) firing evoked by ramp-current injection. Current-clamp record-ings were made with K-gluc. int. soln. at resting Vm. B: example tracesshowing that in this KC, application of PTX caused spontaneous AP firing.C: individual and pooled data (n � 4) showing the small depolarization ofresting Vm evoked by PTX. D: individual and pooled data (n � 4) showing thatPTX reduces the amount of injected current required to evoke AP firing inKCs. *P � 0.05, statistical significance.
2031KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from
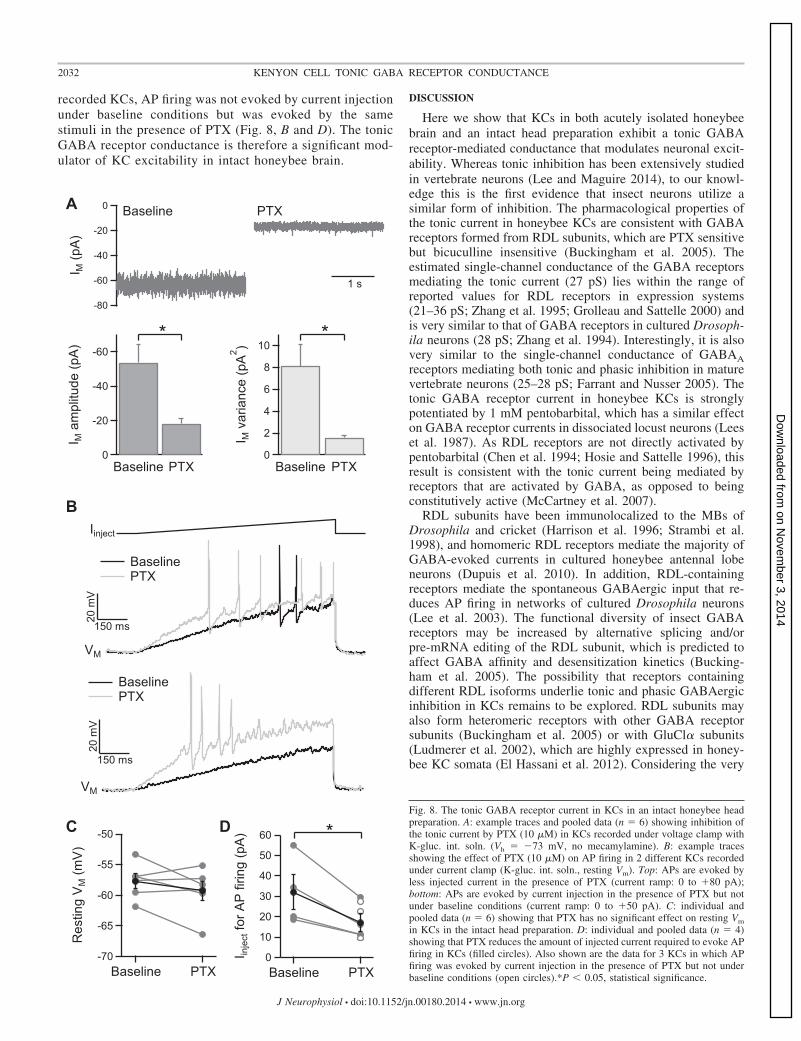
recorded KCs, AP firing was not evoked by current injectionunder baseline conditions but was evoked by the samestimuli in the presence of PTX (Fig. 8, B and D). The tonicGABA receptor conductance is therefore a significant mod-ulator of KC excitability in intact honeybee brain.
DISCUSSION
Here we show that KCs in both acutely isolated honeybeebrain and an intact head preparation exhibit a tonic GABAreceptor-mediated conductance that modulates neuronal excit-ability. Whereas tonic inhibition has been extensively studiedin vertebrate neurons (Lee and Maguire 2014), to our knowl-edge this is the first evidence that insect neurons utilize asimilar form of inhibition. The pharmacological properties ofthe tonic current in honeybee KCs are consistent with GABAreceptors formed from RDL subunits, which are PTX sensitivebut bicuculline insensitive (Buckingham et al. 2005). Theestimated single-channel conductance of the GABA receptorsmediating the tonic current (27 pS) lies within the range ofreported values for RDL receptors in expression systems(21–36 pS; Zhang et al. 1995; Grolleau and Sattelle 2000) andis very similar to that of GABA receptors in cultured Drosoph-ila neurons (28 pS; Zhang et al. 1994). Interestingly, it is alsovery similar to the single-channel conductance of GABAAreceptors mediating both tonic and phasic inhibition in maturevertebrate neurons (25–28 pS; Farrant and Nusser 2005). Thetonic GABA receptor current in honeybee KCs is stronglypotentiated by 1 mM pentobarbital, which has a similar effecton GABA receptor currents in dissociated locust neurons (Leeset al. 1987). As RDL receptors are not directly activated bypentobarbital (Chen et al. 1994; Hosie and Sattelle 1996), thisresult is consistent with the tonic current being mediated byreceptors that are activated by GABA, as opposed to beingconstitutively active (McCartney et al. 2007).
RDL subunits have been immunolocalized to the MBs ofDrosophila and cricket (Harrison et al. 1996; Strambi et al.1998), and homomeric RDL receptors mediate the majority ofGABA-evoked currents in cultured honeybee antennal lobeneurons (Dupuis et al. 2010). In addition, RDL-containingreceptors mediate the spontaneous GABAergic input that re-duces AP firing in networks of cultured Drosophila neurons(Lee et al. 2003). The functional diversity of insect GABAreceptors may be increased by alternative splicing and/orpre-mRNA editing of the RDL subunit, which is predicted toaffect GABA affinity and desensitization kinetics (Bucking-ham et al. 2005). The possibility that receptors containingdifferent RDL isoforms underlie tonic and phasic GABAergicinhibition in KCs remains to be explored. RDL subunits mayalso form heteromeric receptors with other GABA receptorsubunits (Buckingham et al. 2005) or with GluCl� subunits(Ludmerer et al. 2002), which are highly expressed in honey-bee KC somata (El Hassani et al. 2012). Considering the very
A
B
-60
-40
-20
0
I M a
mpl
itude
(pA
)
Baseline PTX
*10
8
6
4
2
0
I M v
aria
nce
(pA
2 )
Baseline PTX
*-80
-60
-40
-20
0
I M (p
A)
1 s
Baseline PTX
-70
-65
-60
-55
-50
Res
ting
VM
(mV
)
Baseline PTX
150 ms
20 m
V
Baseline PTX
VM
Iinject
150 ms
20 m
V
VM
Baseline PTX
C D 60
50
40
30
20
10
0I inje
ct fo
r AP
firin
g (p
A)
Baseline PTX
*Fig. 8. The tonic GABA receptor current in KCs in an intact honeybee headpreparation. A: example traces and pooled data (n � 6) showing inhibition ofthe tonic current by PTX (10 �M) in KCs recorded under voltage clamp withK-gluc. int. soln. (Vh � �73 mV, no mecamylamine). B: example tracesshowing the effect of PTX (10 �M) on AP firing in 2 different KCs recordedunder current clamp (K-gluc. int. soln., resting Vm). Top: APs are evoked byless injected current in the presence of PTX (current ramp: 0 to �80 pA);bottom: APs are evoked by current injection in the presence of PTX but notunder baseline conditions (current ramp: 0 to �50 pA). C: individual andpooled data (n � 6) showing that PTX has no significant effect on resting Vm
in KCs in the intact head preparation. D: individual and pooled data (n � 4)showing that PTX reduces the amount of injected current required to evoke APfiring in KCs (filled circles). Also shown are the data for 3 KCs in which APfiring was evoked by current injection in the presence of PTX but not underbaseline conditions (open circles).*P � 0.05, statistical significance.
2032 KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from

small size of L-glutamate-evoked currents relative to GABA-evoked currents in the present study, the identity and functionof receptors containing GluCl� in honeybee KCs require fur-ther investigation. It is also likely that GABAB receptorscontribute to MB inhibition, given the observed functionaleffects of modulating GABAB receptor activation in insectKCs (Demmer and Kloppenburg 2009; Froese et al. 2013;Nakamura and Yoshino 2013).
GABA-evoked currents in honeybee KCs desensitize rela-tively slowly, exhibiting a decay time constant of 2.6 s whenstimulated with a high concentration of GABA. A slow rate ofdesensitization is required for tonic receptor activation and is acommon property of GABA receptors mediating tonic currentsin vertebrate neurons (Glykys and Mody 2007; Belelli et al.2009). The magnitude of the transient current evoked by localapplication of GABA suggests that KCs have the capacity forsubstantial GABAergic input and that the tonic current repre-sents the activation of a small proportion of KC GABAreceptors. The size of the tonic current may be modulated notonly by changing the number of activated receptors but also bychanging their responsiveness to GABA. For example, GABA-evoked currents in cultured honeybee neurons are positivelyregulated by intracellular Ca2�, possibly via kinase activation(Grünewald and Wersing 2008; Dupuis et al. 2010). It istherefore possible that the reduction in the KC tonic currentevoked by CdCl2 and nifedipine results from decreased post-synaptic Ca2� influx rather than a presynaptic effect on GABArelease. Insect GABA receptor subunits have multiple phos-phorylation sites in the cytoplasmic loop between transmem-brane domains 3 and 4 (Jones and Sattelle 2006), which mayenable activity-dependent regulation of the tonic current viachanges in receptor properties or membrane expression levels.
Honeybee KCs exhibit spontaneous GABAergic input in theform of a tonic membrane current, rather than phasic inhibitorypostsynaptic currents (IPSCs). The lack of detectable IPSCs isunlikely to be due to the electrical properties of the KCrecordings as spontaneous nicotinic EPSCs are commonlyobserved (unpublished observations); however, it is possiblethat small amplitude IPSCs are indistinguishable within thenoise associated with the tonic current. Similarly, GABAergicIPSCs are not readily observed in recordings from DrosophilaKCs (Gu and O’Dowd 2006), whereas cockroach KCs exhibita high frequency of spontaneous IPSPs, which were suggestedto suppress spontaneous KC activity and sharpen odor re-sponses (Demmer and Kloppenburg 2009). In vertebrate neu-rons, phasic inhibition and tonic inhibition differ in theiractivation mechanisms. Tonic inhibition can be mediated byambient extracellular GABA or the release of nonvesicularGABA acting on extrasynaptic GABA receptors, providingdistinct functional effects from inhibition mediated by synapticGABA release (Farrant and Nusser 2005; Belelli et al. 2009).The tonic GABA receptor current in honeybee KCs is likely toarise, at least in part, from nonsynaptic GABA, as a componentof the tonic current is independent of voltage-gated Ca2�
channel activation. The relationship between tonic and phasicinhibition in KCs and their respective roles in the regulation ofKC function require further experimental investigation.
Within honeybee MBs, the dendritic fields of individual KCsarborize within one calyceal subregion (lip, collar, or basalring) and their axons project to a corresponding sublayer of the�-lobe (ventral, median and dorsal, respectively; Mobbs 1982;
Strausfeld 2002). These MB subregions receive and processsensory input from a specific modality (olfactory, visual, andmixed input, respectively). The MB feedback neurons consistof a group of �50 GABAergic cells located in the lateralprotocerebral lobe with extensive, highly branched neuritesinnervating specific areas of the MB �/�-lobes and peduncu-lus and the whole of one subregion of both the medial andlateral calyces (Gronenberg 1987; Rybak and Menzel 1993;Grünewald 1999a). Within the MB calyx, the majority ofGABAergic synapses are formed with KC dendrites, but re-ciprocal synapses with projection neuron boutons are alsofound (Ganeshina and Menzel 2001), potentially enablingmodulation of inhibitory input by local microcircuits. Foursubtypes of MB feedback neuron have been described in thehoneybee on the basis of their morphology, at least three ofwhich innervate corresponding �-lobe and calyceal subregions(Grünewald 1999a; Strausfeld 2002). Despite their very spe-cific MB innervation patterns, most honeybee feedback neu-rons respond to multisensory stimulation with an increased rateof AP firing (Gronenberg 1987; Grünewald 1999b). In locustand Drosophila, a single GABAergic feedback neuron hasbeen identified that similarly extensively innervates both theoutput and input regions of the MB, and responds to odorstimulation with graded depolarizations rather than APs(Leitch and Laurent 1996; Liu and Davis 2009; Papadopoulouet al. 2011). Furthermore, the responses of MB feedbackneurons exhibit plasticity related to associative learning(Grünewald 1999b; Liu and Davis 2009).
GABAergic input to the MBs contributes to a markedchange in the odor response profiles of neurons in the insectolfactory pathway, from dense representations in antennal lobeprojection neurons to sparse coding in KCs (Perez-Orive et al.2002; Turner et al. 2008). Depolarization of the locust MBfeedback neuron has been shown to reduce KC excitability andodor responses and consequently reduce odor responses in MBoutput neurons (Papadopoulou et al. 2011). In Drosophila,downregulation of RDL receptors or of GABA synthesis in theMB feedback neuron increases the proportion of KCs respond-ing to an odor and reduces the odor selectivity of individualneurons (Lei et al. 2013). Furthermore, the inhibitory feedbackloop in Drosophila MBs was recently shown to decorrelate KCresponses to similar odors, enabling discrimination betweenthese odors in an olfactory learning task (Lin et al. 2014). Inhoneybees, in vivo Ca2� imaging of KC dendritic responseshas shown that GABAergic inhibition reduces KC odor re-sponses in a concentration-dependent manner (Froese et al.2013). MB feedback inhibition therefore appears to act as again control mechanism for KC activation, maintaining sparsecoding over a wide range of input intensities (Papadopoulou etal. 2011; Lei et al. 2013). The presence of a tonic GABAreceptor current in honeybee KCs in the absence of sensorystimulation suggests that GABAergic input regulates KC ex-citability under basal conditions, in addition to being recruitedby KC activation as a form of classical feedback inhibition.Tonic inhibition is therefore likely to contribute to sparsecoding in KCs, in combination with other factors such as theirintrinsic membrane properties (Perez-Orive et al. 2002; Dem-mer and Kloppenburg 2009).
In conclusion, honeybee KCs exhibit a tonic GABA receptorconductance that modulates excitability and may contribute tosparse coding of sensory information in the MBs. The tonic
2033KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from

GABA receptor current in KCs therefore shares functionalsimilarities with tonic GABA currents in rodent brain regionssuch as the hippocampus, cerebellum, and neocortex, wherethey act to modulate neuronal gain and network dynamics (Leeand Maguire 2014). The relationship between the tonic GABAreceptor current and phasic inhibitory currents in KCs remainsto be determined, in addition to the specific subunit composi-tion, localization, and functional properties of the GABAreceptors mediating these forms of inhibition. By extending thestudy of endogenous inhibitory mechanisms to other honeybeebrain regions and to other model organisms such as Drosoph-ila, the wider role of tonic inhibition in sensory processing andhigher cognitive function in insects may be elucidated.
ACKNOWLEDGMENTS
We thank Dr. Chris Moffat for bee husbandry.
GRANTS
This work has been funded jointly by the Biotechnology and BiologicalSciences Research Council; the Department for Environment, Food and RuralAffairs; the Natural Environment Research Council, the Scottish Government;and the Wellcome Trust under the Insect Pollinators Initiative (UK) Grant No.BB/1000313/1.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: M.J.P. and J.H. conception and design of research;M.J.P. performed experiments; M.J.P. analyzed data; M.J.P. interpreted resultsof experiments; M.J.P. prepared figures; M.J.P. drafted manuscript; M.J.P. andJ.H. edited and revised manuscript; M.J.P. and J.H. approved final version ofmanuscript.
REFERENCES
Albrecht J, Schousboe A. Taurine interaction with neurotransmitter receptorsin the CNS: an update. Neurochem Res 30: 1615–1621, 2005.
Belelli D, Harrison NL, Maguire J, Macdonald RL, Walker MC, CopeDW. Extrasynaptic GABA(A) receptors: form, pharmacology, function. JNeurosci 29: 12757–12763, 2009.
Bicker G, Schäfer S, Kingan TG. Mushroom body feedback interneurons inthe honeybee show GABA-like immunoreactivity. Brain Res 360: 394–397,1985.
Buckingham SD, Biggin PC, Sattelle BM, Brown LA, Sattelle DB. InsectGABA receptors: Splicing, editing, and targeting by antiparasitics andinsecticides. Mol Pharmacol 68: 942–951, 2005.
Chen RT, Belelli D, Lambert JJ, Peters JA, Reyes A, Lan NC. Cloning andfunctional expression of a Drosophila �-aminobutyric-acid receptor. ProcNatl Acad Sci USA 91: 6069–6073, 1994.
Demmer H, Kloppenburg P. Intrinsic membrane properties and inhibitorysynaptic input of Kenyon cells as mechanisms for sparse coding? J Neuro-physiol 102: 1538–1550, 2009.
Dupuis JP, Bazelot M, Barbara GS, Paute S, Gauthier M, Raymond-Delpech V. Homomeric RDL and heteromeric RDL/LCCH3 GABA recep-tors in the honeybee antennal lobes: two candidates for inhibitory transmis-sion in olfactory processing. J Neurophysiol 103: 458–468, 2010.
El Hassani AK, Schuster S, Dyck Y, Demares F, Leboulle G, ArmengaudC. Identification, localization and function of glutamate-gated chloridechannel receptors in the honeybee brain. Eur J Neurosci 36: 2409–2420,2012.
Farrant M, Nusser Z. Variations on an inhibitory theme: phasic and tonicactivation of GABA(A) receptors. Nat Rev Neurosci 6: 215–229, 2005.
Froese A, Szyszka P, Menzel R. Effect of GABAergic inhibition on odorantconcentration coding in mushroom body intrinsic neurons of the honeybee.J Comp Physiol A 1–13, 2013.
Ganeshina O, Menzel R. GABA-immunoreactive neurons in the mushroombodies of the honeybee: an electron microscopic study. J Comp Neurol 437:335–349, 2001.
Glykys J, Mody I. Activation of GABA(A) receptors: views from outside thesynaptic cleft. Neuron 56: 763–770, 2007.
Grolleau F, Sattelle DB. Single channel analysis of the blocking actions ofBIDN and fipronil on a Drosophila melanogaster GABA receptor (RDL)stably expressed in a Drosophila cell line. Br J Pharmacol 130: 1833–1842,2000.
Gronenberg W. Anatomical and physiological-properties of feedback neuronsof the mushroom bodies in the bee brain. Exp Biol 46: 115–125, 1987.
Grünewald B. Morphology of feedback neurons in the mushroom body of thehoneybee, Apis mellifera. J Comp Neurol 404: 114–126, 1999a.
Grünewald B. Physiological properties and response modulations of mush-room body feedback neurons during olfactory learning in the honeybee,Apis mellifera. J Comp Physiol A 185: 565–576, 1999b.
Grünewald B, Wersing A. An ionotropic GABA receptor in cultured mush-room body Kenyon cells of the honeybee and its modulation by intracellularcalcium. J Comp Physiol A 194: 329–340, 2008.
Gu HY, O’Dowd DK. Cholinergic synaptic transmission in adult DrosophilaKenyon cells in situ. J Neurosci 26: 265–272, 2006.
Harrison JB, Chen HH, Sattelle E, Barker PJ, Huskisson NS, Rauh JJ, BaiD, Sattelle DB. Immunocytochemical mapping of a C-terminus anti-peptideantibody to the GABA receptor subunit, RDL in the nervous system ofDrosophila melanogaster. Cell Tissue Res 284: 269–278, 1996.
Heisenberg M. What do the mushroom bodies do for the insect brain? Anintroduction. Learn Mem 5: 1–10, 1998.
Hosie AM, Sattelle DB. Allosteric modulation of an expressed homo-oligo-meric GABA-gated chloride channel of Drosophila melanogaster. Br JPharmacol 117: 1229–1237, 1996.
Husch A, Paehler M, Fusca D, Paeger L, Kloppenburg P. Distinct electro-physiological properties in subtypes of nonspiking olfactory local interneu-rons correlate with their cell type-specific Ca2� current profiles. J Neuro-physiol 102: 2834–2845, 2009.
Jia F, Yue M, Chandra D, Keramidas A, Goldstein PA, Homanics GE,Harrison NL. Taurine is a potent activator of extrasynaptic GABA(A)receptors in the thalamus. J Neurosci 28: 106–115, 2008.
Jones A, Sattelle D. The cys-loop ligand-gated ion channel superfamily of thehoneybee, Apis mellifera. Invert Neurosci 6: 123–132, 2006.
King GF. Modulation of insect Ca-v channels by peptidic spider toxins.Toxicon 49: 513–530, 2007.
Koch U, Magnusson AK. Unconventional GABA release: mechanisms andfunction. Curr Opin Neurobiol 19: 305–310, 2009.
Lee D, Su HL, O’Dowd DK. GABA receptors containing Rdl subunitsmediate fast inhibitory synaptic transmission in Drosophila neurons. JNeurosci 23: 4625–4634, 2003.
Lee V, Maguire J. The impact of tonic GABAA receptor-mediated inhibitionon neuronal excitability varies across brain region and cell type. FrontNeural Circuits 8: 3, 2014.
Lees G, Beadle DJ, Neumann R, Benson JA. Responses to GABA byisolated insect neuronal somata: pharmacology and modulation by a benzo-diazepine and a barbiturate. Brain Res 401: 267–278, 1987.
Lei Z, Chen K, Li H, Liu H, Guo A. The GABA system regulates the sparsecoding of odors in the mushroom bodies of Drosophila. Biochem BiophysRes Commun 436: 35–40, 2013.
Leitch B, Laurent G. GABAergic synapses in the antennal lobe and mush-room body of the locust olfactory system. J Comp Neurol 372: 487–514,1996.
Lin AC, Bygrave AM, de Calignon A, Lee T, Miesenbock G. Sparse,decorrelated odor coding in the mushroom body enhances learned odordiscrimination. Nat Neurosci 17: -559–U116, 2014.
Liu X, Davis RL. The GABAergic anterior paired lateral neuron suppressesand is suppressed by olfactory learning. Nat Neurosci 12: 53–59, 2009.
Ludmerer SW, Warren VA, Williams BS, Zheng Y, Hunt DC, Ayer MB,Wallace MA, Chaudhary AG, Egan MA, Meinke PT, Dean DC, GarciaML, Cully DF, Smith MM. Ivermectin and nodulisporic acid receptors inDrosophila melanogaster contain both �-aminobutyric acid-gated Rdl andglutamate-gated GluCl� chloride channel subunits. Biochemistry 41: 6548–6560, 2002.
McCartney MR, Deeb TZ, Henderson TN, Hales TG. Tonically activeGABAA receptors in hippocampal pyramidal neurons exhibit constitutiveGABA-independent gating. Mol Pharmacol 71: 539–548, 2007.
Mellor JR, Gunthorpe MJ, Randall AD. The taurine uptake inhibitorguanidinoethyl sulphonate is an agonist at gamma-aminobutyric acid(A)
2034 KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from

receptors in cultured murine cerebellar granule cells. Neurosci Lett 286:25–28, 2000.
Menzel R. The honeybee as a model for understanding the basis of cognition.Nat Rev Neurosci 13: 758–768, 2012.
Mobbs PG. The brain of the honeybee Apis-mellifera. 1. The connections andspatial-organization of the mushroom bodies. Philos T Roy Soc B 298:309–354, 1982.
Nakamura A, Yoshino M. A novel GABAergic action mediated byfunctional coupling between GABAB-like receptor and two differenthigh-conductance K� channels in cricket Kenyon cells. J Neurophysiol109: 1735–1745, 2013.
Oleskevich S. Cholinergic synaptic transmission in insect mushroom bodies invitro. J Neurophysiol 82: 1091–1096, 1999.
Olshausen BA, Field DJ. Sparse coding of sensory inputs. Curr OpinNeurobiol 14: 481–487, 2004.
Palmer MJ, Moffat C, Saranzewa N, Harvey J, Wright GA, Connolly CN.Cholinergic pesticides cause mushroom body neuronal inactivation in hon-eybees. Nat Comm 4: 1634, 2013.
Papadopoulou M, Cassenaer S, Nowotny T, Laurent G. Normalization forsparse encoding of odors by a wide-field interneuron. Science 332: 721–725,2011.
Perez-Orive J, Mazor O, Turner GC, Cassenaer S, Wilson RI, Laurent G.Oscillations and sparsening of odor representations in the mushroom body.Science 297: 359–365, 2002.
Richerson GB, Wu Y. Dynamic equilibrium of neurotransmitter transporters:not just for reuptake anymore. J Neurophysiol 90: 1363–1374, 2003.
Rybak J, Menzel R. Anatomy of the mushroom bodies in the honey-beebrain–the neuronal connections of the alpha-lobe. J Comp Neurol 334:444–465, 1993.
Schäfer S, Bicker G, Ottersen OP, Stormmathisen J. Taurine-like immu-noreactivity in the brain of the honeybee. J Comp Neurol 268: 60–70, 1988.
Semyanov A, Walker MC, Kullmann DM, Silver RA. Tonically activeGABA(A) receptors: modulating gain and maintaining the tone. TrendsNeurosci 27: 262–269, 2004.
Skeer JM, Norman RI, Sattelle DB. Invertebrate voltage-dependent calciumchannel subtypes. Biol Rev 71: 137–154, 1996.
Strambi C, Cayre M, Sattelle DB, Augier R, Charpin P, Strambi A.Immunocytochemical mapping of an RDL-like GABA receptor subunit andof GABA in brain structures related to learning and memory in the cricketAcheta domesticus. Learn Mem 5: 78–89, 1998.
Strausfeld NJ. Organization of the honey bee mushroom body: representation ofthe calyx within the vertical and gamma lobes. J Comp Neurol 450: 4–33, 2002.
Su HL, O’Dowd DK. Fast synaptic currents in Drosophila mushroom bodyKenyon cells are mediated by alpha-bungarotoxin-sensitive nicotinic ace-tylcholine receptors and picrotoxin-sensitive GABA receptors. J Neurosci23: 9246–9253, 2003.
Turner GC, Bazhenov M, Laurent G. Olfactory representations by Dro-sophila mushroom body neurons. J Neurophysiol 99: 734–746, 2008.
Witthöft W. Absolute anzahl und verteilung der zellen im hirn der honigbiene.Z Morphol Tiere 61: 160–184, 1967.
Zhang HG, ffrench-Constant RH, Jackson MB. A unique amino-acid of theDrosophila GABA receptor with influence on drug-sensitivity by 2 mecha-nisms. J Physiol 479: 65–75, 1994.
Zhang HG, Lee HJ, Rocheleau T, ffrench-Constant RH, Jackson MB.Subunit composition determines picrotoxin and bicuculline sensitivity ofDrosophila gamma-aminobutyric-acid receptors. Mol Pharmacol 48: 835–840, 1995.
2035KENYON CELL TONIC GABA RECEPTOR CONDUCTANCE
J Neurophysiol • doi:10.1152/jn.00180.2014 • www.jn.org
on Novem
ber 3, 2014D
ownloaded from