hindy_mangione_keys-vegetative effects_finalv1.2 (1) pdf
TRANSCRIPT

Vegetative effects on grassland bird biodiversity in Southwest Michigan
MICHAEL HINDY, Cornerstone University, 1001 E Beltline NE, Grand Rapids, MI 49525,
USA
TRAVIS MANGIONE, Cornerstone University, 1001 E Beltline NE, Grand Rapids, MI 49525,
USA
ROBERT S. KEYS, Cornerstone University, 1001 E Beltline, NE, Grand Rapids, MI 49525,
USA
ABSTRACT In response to the decline of grassland birds and their habitat, the Pierce Cedar
Creek Institute has been working to restore portions of their property to prairie habitat. Obligate
grassland bird species, including the Henslow's sparrow (Ammodramus henslowii) and
grasshopper sparrow (Ammodramus savannarum), have avoided these grasslands despite a
decade of the prairie restoration. In the summer of 2016 we investigated the vegetation of
grasslands and restored prairies throughout Southwest Michigan to determine if vegetation
characteristics within these restored prairies are driving site selection. Our bird and vegetation
surveys found significant differences in vegetative density, litter depth, vegetative height and
cool/warm season mixes between restored prairies and other grasslands in regards to obligate
grassland birds. We found obligate grassland birds preferred a mean vertical density of 46.6 cm,
3.56 cm litter depth, 70.3 cm vegetation height, and 61.32% cool-season plant dominance. These
parameters significantly differ than what is found in restored prairies, which were nearly void of
obligate grassland birds. This information helps to inform the Institute’s management plans to
increase grassland bird biodiversity in their prairies.
KEY WORDS: biodiversity, cool-season plants, grasshopper sparrow, grassland birds,
Henslow's sparrow, Pierce Cedar Creek Institute, prairie management, prairie restoration,
Southwest Michigan, vegetation characteristics

Hindy, Mangione and Keys 2
INTRODUCTION
The decrease in grassland bird species over the last 30 years has put grasslands in the
conservation spotlight (Johnson and Igl 2001; Ribic and Sample 2001; Bakker et al. 2002;
Norment 2002; Herkert et al. 2003; Brennan and Kuvlesky Jr. 2005). Eighty percent of
grasslands in the United States have been converted to other uses such as development and
agricultural use (Rahmig et al. 2009). The State of Michigan has also seen 99% of its original
grasslands converted to other uses over the last one hundred years (Chapman and Brewer 2008;
Parker 2012). The Pierce Cedar Creek Institute (Institute) (42°32'07.0"N 85°18'06.7"W) has
been mitigating this loss by restoring 30.35 ha of tallgrass prairie and increasing the contiguous
prairie footprint to make their grasslands attractive for grassland birds. Remaining native
grasslands are in danger of natural plant succession (Norment 2002), thus increasing the patch
size of the Institute’s grasslands is a priority in slowing succession (Fletcher and Koford 2002;
Norment 2002).
The two target bird species of the Institute's restored prairie are the state endangered
Henslow’s sparrow (Ammodramus henslowii) and the state "species of special concern"
grasshopper sparrow (Ammodramus savannarum) (BirdLife International 2012; Howell 2015).
Surveys prior to the Institute's prairie expansion recorded only three sightings of Henslow’s
sparrow over the past six years (Oosterhouse et al. 2012; Cushman et al. 2013; eBird 2014).
Research completed last year at the Institute indicated no Henslow’s sparrows in the new
contiguous prairie environments (Lambert et al. 2015). In contrast, two other regional grassland
sites (Soya Conservation Reserve Program grassland and Gerald R Ford International Airport)
show healthy breeding population combinations of Henslow’s, grasshopper, savannah

Hindy, Mangione and Keys 3
(Passerculus sandwichensis), and/or vesper sparrows (Pooecetes gramineus), along with
bobolink (Dolichonyx oryzivorus) and eastern meadowlarks (Sturnella magna) (Lambert et al.
2015).
The disparity between these sites and the desire to make modifications at the Institute to
attract these species has prompted questions regarding occupation factors present in other sites.
Lambert et al. (2015) found cool season domestic fescue grasses (Festuca sp.) had the highest
correlation with grassland bird diversity over goldenrods (Solidago sp.) and big bluestem
(Andropogon gerardii) dominated grasslands (Lambert et al. 2015). Over time, big bluestem can
come to dominate in abundance and create a monoculture in restored prairies (Kindscher and
Tieszen 1998). This dominance of big bluestem could deter grassland birds from inhabiting
restored prairies (Lambert et al. 2015).
This led us to hypothesize the driving factor contributing to the absence of obligate grassland
birds at the Institute was the vegetative characteristics of the prairies. Grassland vegetation has
characteristics that affect grassland bird diversity, such as vertical density, litter depth, vegetative
height, and ground cover (Fletcher and Koford 2002; Rahmig et al. 2009). For instance, studies
have noted Henslow's sparrows prefer a high vertical density (Navarette-Tindall 2010), tall
vegetation (Herkert 1994), and a deep litter layer (Zimmerman 1988). Conversely, other studies
found that grasshopper sparrows prefer sparse, low vegetation (Jobin and Falardeau 2010; Ruth
2015). Aspects such as this make it difficult to manage for multiple species within the same
grassland. While the habitat components for Henslow's sparrows is similar in nature to the
grasslands at the Institute, Henslow’s sparrows have rarely been found in the tallgrass prairies on
the site. This structural component of grasslands will be a part of our investigation into the low
grassland bird biodiversity on the Institute property.

Hindy, Mangione and Keys 4
The vegetation at the Institute consists mainly of warm-season grasses and forbs (Lambert
et al. 2015). While required for a prairie ecosystem, it may indirectly affect the use by grassland
birds in this region (Lambert et al. 2015). Warm-season plants grow in mid-summer and are
potentially not suitable for the early migration of obligate grassland birds. Cool-season
herbaceous plants are beneficial because they grow earlier in the season and provide ample cover
and food for the grassland birds as they return from the wintering grounds (Navarette-Tindall
2010). Lambert et al. (2015) found the sites with the greatest grassland bird diversity consisted
of a combination of warm-season and cool-season plants, with more of the latter than the former.
This prompted us to consider if this was an important vegetative characteristic at the point of
migratory return. We matched sites attracting target species with randomly selected non-use sites
to determine if vegetative characteristics in the early season drive site selection. Regardless of
the vegetative characteristics, the success of a grassland is related to its ability to act as a source
site and not a sink for the target grassland species (McCoy et al. 2001).
Our primary objective is to build on Lambert et al. (2015) to ascertain what vegetative
characteristics in restored prairie grasslands act as attractors or detriments to the grassland bird
diversity of eight obligate grassland species.
Based on this objective we have hypothesized:
H1: the initial condition of vegetative factors at migratory arrival grasslands will determine
choice for breeding use amongst obligate grassland bird species.
H2: forb/grass height, litter depth, and vertical density will be determining factors in grassland
choice by target species.
H3: higher diversity of grass and forb species will result in higher diversity of obligate grassland
birds.

Hindy, Mangione and Keys 5
H4: obligate grassland birds will occur in grasslands that have greater coverage of cool-season
grasses and forbs than warm-season.
Developing a better understanding of the grassland characteristics in restored prairie
grasslands and those sought by grassland birds will help further the management techniques at
the Institute and other restored prairies in increasing grassland bird biodiversity.
METHOD
Study Sites
Grassland sites at 20 locations were surveyed in Barry, Kent and Kalamazoo Counties,
Michigan, USA 23 May- 19 July 2016. Grassland sites were subdivided into two categories
based on management practices and vegetative characteristics: restored prairie (RP) and mixed-
management grasslands (MM). Mixed-management grasslands are grasslands which have not
been restored and are generally fallow hay fields or maintained as grassland for other purposes.
They may use various methods of management including mowing, seeding, burning, or no
management whatsoever. Grassland sites ranged in size from 1 ha to 62 ha (Appendix A). Each
category of grassland included matched sites of similar size (Table 1).
Table 1. Number of grassland sites surveyed based on category and size range, 23 May – 19 July
2016, Barry, Kent and Kalamazoo Counties, Michigan, USA.
Grassland Category <10 ha 10-30 ha >30 ha
Restored Tallgrass Prairie 6 4 1
Mixed Management 3 4 2
Study Species
Our study focuses on eight species which are dependent on grasslands for breeding and
reproduction (obligate grassland species): Henslow’s sparrow, grasshopper sparrow, savannah
sparrow, vesper sparrow, dickcissel (Spiza americana), bobolink, eastern meadowlark and

Hindy, Mangione and Keys 6
upland sandpiper (Bartramia longicauda). We also collected data on species not fully dependent
on grasslands for nesting and reproduction (semi-obligate species): field sparrow (Spizella
pusilla), song sparrow (Melospiza melodia), and common yellowthroat (Geothlypis trichas).
Bird Survey
We conducted point count surveys of all obligate and semi-obligate grassland birds at
each site during the breeding season, 23 May – 20 June 2016 using a 10-minute, 50 m radius,
fixed point count method (Ralph et al. 1993). Surveys were conducted between sunrise and
10:00 AM; when breeding birds are most active. We recorded all target species seen or heard,
along with sex of individual (if identifiable). 100 x 100 m grids were layered over a digital
orthographic projection of each site using ArcGIS 10.4 (ESRI 2015), with each grassland cell
considered a potential point count sampling unit. We stratified sites into sections of 6-10
potential sampling units based on the sizes of the site and then randomized for point count visits
to cover the entire range of variability within each site. Sampling units were not repeated over
the survey period to ensure the most complete sampling of each site (Fletcher and Koford 2002).
Point count observations were inputted into ArcGIS 10.4 and analyzed with territory
buffers of each target species to negate pseudo-replication among sighted birds. Territory buffers
of birds overlapping at least 50% between point count sampling units were considered one
individual bird.
Habitat Measurements
Pre-arrival. Initial vegetation height, vertical density, and litter depth were made in early
May 2016 at point of initial migratory return at 13 point count locations known to have had
target species in the 2015 survey. These were paired with 13 point count sites with no target
species in the 2015 survey (Lambert et al. 2015).

Hindy, Mangione and Keys 7
Breeding Season. We measured vegetation type, percent cover, vertical density, vegetation
height, and litter depth at 35 point count sites containing obligate target species from our 2016
surveys. Each of these was paired with a random point count site containing no obligate target
species to test for vegetative effects between occupied sites and sites lacking obligate grassland
birds. Vegetation type and cover was measured using 0.25m2 Daubenmire frame (Rahmig et al.
2009) at four plots within each site: 1 at the center of the point-count area and 3 located at 0o,
120o, and 240o 30m from the center point (Fletcher and Koford 2002). Litter depth and
vegetation cover was measured in centimeters at each plot site, taking the average of the litter
depths in each corner of the frame (Rahmig et al. 2009). Vegetation height was defined as height
of tallest green vegetation measured in centimeters at each corner of the Daubenmire frame
(Fisher and Davis 2010). Plants within the Daubenmire frame were identified to the species
whenever possible, and defined by growing season. Cool-season vegetation was defined as
primarily growing during spring and fall seasons. Warm-season vegetation was defined as
primarily growing during the summer. Vertical density was determined by measuring visual
obstruction of grasses and forbs looking through a sighting pole 1 m from ground level and 4 m
from a Robel pole in each of the four cardinal directions (Fletcher and Koford 2002; Smith
2008).
Statistical Analysis
We modeled statistical analysis after similar studies on grassland birds conducted by
Fletcher and Koford (2002), Rahmig et al. (2009) and Thomas et al. (2010). Species richness
was defined as number of obligate grassland breeding species observed (Fletcher and Koford
2002). Due to a small sample size of vegetation characteristics examined during migratory
return, a non-parametric Wilcoxon Signed-Ranks test was used to determine differences between

Hindy, Mangione and Keys 8
sites with obligate grassland birds and sites without obligate grassland birds. Independent
sample T-tests were used to determine the differences between litter depth, vertical density,
vegetative height, and ground cover during breeding season. A multiple linear regression was
used to determine the most parsimonious model between Shannon diversity indices of vegetation
and obligate grassland birds (H'), with obligate grassland birds serving as the response variable.
Vegetative models between observed and random points were compared on a seasonal plant-type
basis using Chi-square to determine if specific seasonal plant types drive the selection of habitat
by species.
RESULTS
Survey results
Point count surveys showed obligate grassland birds had highest occurrences in mixed
management grasslands greater than 30 ha. Concurrently, semi-obligate grassland birds occurred
in high numbers in restored prairies at all grassland sizes. Mixed-management grasslands had
158.67% more individual obligate birds than restored prairie grasslands (Table 2 and 3). Obligate
grassland birds occurred in fields as small as 9 ha and as large as 62 ha.
Table 2. Number of obligate and semi-obligate grassland birds counted in 10-minute point counts
in grasslands based on grassland management practice and size differential. 23 May – 20 June
2016, Barry, Kent and Kalamazoo Counties, Michigan, USA. <10 ha 10-30 ha >30 ha
P3 O4 SO5 P O SO P O SO
RP1 31 1 94 48 0 139 59 2 150
MM2 25 8 19 80 101 125 107 367 37
1Restored Prairie 2Mixed Managed Grassland 3Point Count Sites 4Obligate Grassland Birds 5Semi-obligate Grassland Birds

Hindy, Mangione and Keys 9
Table 3. Number of each species recorded in point counts, number of grasslands they were found
in, and number of those birds occurring in restored prairies (RP) and mixed-management
grasslands (MM); 23 May – 20 June 2016, Barry, Kent, and Kalamazoo Counties, Michigan,
USA.
Species # Observed # Grasslands # of birds in RP # of birds in MM
HESP1 135 7 2 133
GRSP2 13 2 0 13
SAVS3 120 3 0 120
VESP4 1 1 1 0
DICK5 6 1 0 6
BOBO6 158 6 0 158
EAME7 44 5 0 44
UPSA8 2 1 0 2
FISP9 141 16 92 49
SOSP10 209 20 121 88
COYE11 271 18 193 78
1Henslow's sparrow 2grasshopper sparrow 3savannah sparrow 4vesper sparrow 5dickcissel 6bobolink 7Eastern meadowlark
8upland sandpiper 9field sparrow 10song sparrow 11common yellowthroat
Migratory vegetation differences
A Wilcoxon Signed-Ranks Test of 2016 migratory arrival vegetative data between 13
sites with obligate grassland birds and 13 sites without obligate grassland birds (based on 2015
point count survey) showed no significant difference in litter depth (Z = -1.745, p = 0.081),
vegetation height (Z = 0.075, p = 0.916), and vertical density (Z = -0.524 , p = 0.600).
Mid-Breeding Season Vegetation differences
Independent Samples T-tests showed significant differences in litter depth (cm),
vegetation height (cm) and vertical density (cm) between sites with and without obligate
grassland birds. Litter depth was 67.4% greater in sites without obligate grassland birds (M =
7.17 cm, SD = 2.904) than with obligate grassland birds (M = 3.56 cm, SD =1.941); T(68) =

Hindy, Mangione and Keys 10
6.119, p < 0.0001. Similarly, vegetation height was 18% greater in sites without obligate
grassland birds (M = 84.22 cm, SD = 14.793) than with them (M = 70.33 cm, SD = 17.801);
T(68) = 3.551, p = 0.001. Vertical density of grasslands was also 39.8% greater at sites without
obligate grassland birds (M = 69.74 cm, SD = 21.908) than it was at sites where they were
present (M = 46.61 cm, SD = 19.690); T(68) = 4.645, p < 0.0001.
An Independent Samples T-Test showed no significant differences in ground cover at
sites with obligate grassland birds (M = 3.08, SD = 0.632) and sites without obligate grassland
birds (M = 2.96, SD = 0.841); T(68) = -0.676, p = 0.502.
Vegetation preferences
Using an adjusted Braun-Blanquet cover-abundance scale (Ralph et al. 1993), we defined
dominance as the addition of this scale for species according to growing season. We found cool-
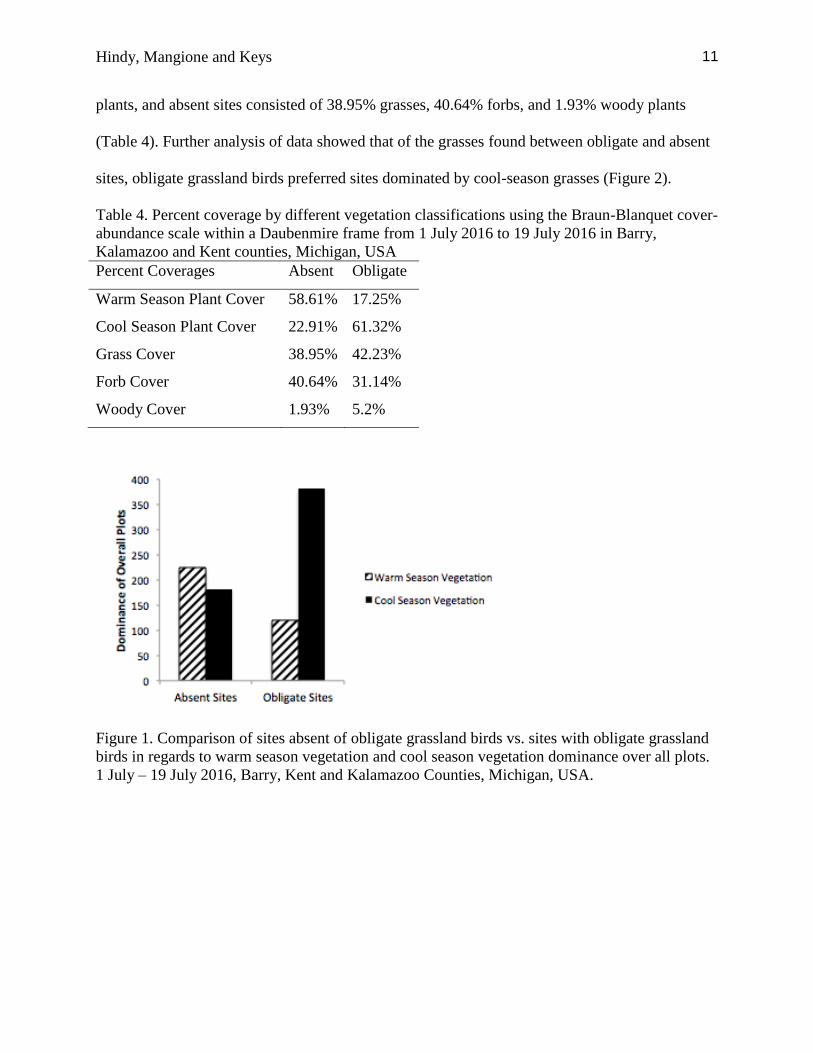
season vegetation occurred more frequently in sites with obligate grassland birds (n = 382), as
opposed to warm-season vegetation (n = 120). This was inversely projected by sites absent of
obligate grassland birds which had a higher frequency of warm-season plants (n = 224), as
opposed to cool-season plants (n =182) (Figure 1).
Percent coverage followed a similar pattern as occurrence of plant type in regards to
growing season and bird diversity. Sites with obligate grassland birds exhibited a higher percent
coverage of cool-season plants (61.32%) as opposed to warm-season plants (17.25%). The
opposite held true for sites absent of obligate grassland birds, with warm-season plants
dominating vegetation (58.61%) and cool-season plants consisting of a smaller percentage of
land cover (22.91%) (Table 4).
The percentage of grasses, forbs, and woody plants did not differ greatly between absent
and obligate sites. Obligate sites consisted of 42.23% grasses, 31.14% forbs, and 5.2% woody
Hindy, Mangione and Keys 11
plants, and absent sites consisted of 38.95% grasses, 40.64% forbs, and 1.93% woody plants
(Table 4). Further analysis of data showed that of the grasses found between obligate and absent
sites, obligate grassland birds preferred sites dominated by cool-season grasses (Figure 2).
Table 4. Percent coverage by different vegetation classifications using the Braun-Blanquet cover-
abundance scale within a Daubenmire frame from 1 July 2016 to 19 July 2016 in Barry,
Kalamazoo and Kent counties, Michigan, USA
Percent Coverages Absent Obligate
Warm Season Plant Cover 58.61% 17.25%
Cool Season Plant Cover 22.91% 61.32%
Grass Cover 38.95% 42.23%
Forb Cover 40.64% 31.14%
Woody Cover 1.93% 5.2%
Figure 1. Comparison of sites absent of obligate grassland birds vs. sites with obligate grassland
birds in regards to warm season vegetation and cool season vegetation dominance over all plots.
1 July – 19 July 2016, Barry, Kent and Kalamazoo Counties, Michigan, USA.

Hindy, Mangione and Keys 12
Figure 2. Percentage composition of warm-season and cool-season grasses at sites absent of
obligate grassland birds vs. sites with obligate grassland bird populations overall all survey
points. 1 July - 19 July 2016, Barry, Kent and Kalamazoo Counties, Michigan, USA.
Vegetation diversity and grassland bird diversity
Linear regression modeling showed only a weak correlation between grassland vegetation
diversity to bird diversity (R2 = 0.289) (Figure 3).
Figure 3. Linear regression of vegetation diversity to bird diversity of each grassland with
obligate grassland birds. 1 July- 19 July 2016, Barry, Kent and Kalamazoo Counties, Michigan,
USA.
-1
0
1
2
3
2 2.2 2.4 2.6 2.8 3 3.2 3.4
Bir
d D
ive
rsit
y
Vegetation Diversity

Hindy, Mangione and Keys 13
DISCUSSION
We had hypothesized that initial vegetative conditions at migratory return would be
different at sites with obligate grassland birds than sites not previously having them. Our results
indicated no statistical difference between the initial conditions of vegetation for site selection
during migratory return. Vegetation conditions during migratory return may not be a factor in
site selection for establishing territories. However, sample size may play a factor in these results
(N=13 for each variable).
We also hypothesized forb/grass height, litter depth, and vertical density would be
determining factors in grassland choice by target species. This hypothesis was supported by our
results. Obligate grassland birds preferred sites with a shorter mean vegetative height (70.33 cm),
a shallower mean litter depth (3.55 cm), and lower mean vertical density (46.61 cm), than sites
without grassland birds. Our results paralleled previous studies by Lambert et al. (2015), Winter
et al. (2005) and Skinner (1975) relating to habitat selection by obligate grassland bird species.
This could be a result of adaptations to new habitat, given the loss of prairie habitats in this
region over 100 years ago. Obligate grassland bird preferences may have shifted as increasing
grassland habitats were converted to agricultural and urban uses in Michigan and the United
States and less of their original habitat remained (Rahmig et al. 2009, Parker 2012). This could
also be a result of locational differences in surveys conducted.
We also hypothesized higher diversity of grass and forb species would result in higher
diversity of grassland birds. This hypothesis was not supported by the data. Many of our restored
prairie grasslands, other than those taken over by big bluestem, had high vegetative diversity.
The plant species richness though was not as important as the structure of the vegetation, which
was why they were void of obligate grassland birds (Bakker et al. 2002). Some of the grasslands

Hindy, Mangione and Keys 14
with high bird diversity (Soya East), did not have high vegetative diversity, and were composed
primarily of the dominant hayfield-type grasses.
Finally, we hypothesized that grassland birds would occur in grasslands having greater
coverage of cool-season grasses and forbs than warm-season. This hypothesis was supported by
our data. These results differ with Jaster et al. (2013) who found Henslow's sparrows did not
prefer cool-season over warm-season, but are consistent with McCoy et al. (2001) who found
Henslow's sparrows preferred grasslands dominated by cool season vegetation over warm season
vegetation. Our results suggest grasslands dominated by cool-season vegetation result in greater
abundance and higher diversity of obligate grassland bird species.
Our results also indicate obligate grassland birds are found in grasslands varying in size,
from 9 ha to 62 ha. This finding suggests that obligate grassland birds may not be dependent
upon grassland size for site selection. Earlier research indicated that while obligate grassland
birds can be found in smaller patch sizes, they exhibited higher densities in large grasslands
(Herkert 1998; Swanson 1998; Winter and Faaborg 1999; Dechant et al. 2001). Larger
grasslands in our study showed a higher diversity, as well as a greater populations, of obligate
grassland birds as would be expected with island biogeographic theory (MacArthur and Wilson
1967).
Restored prairie plantings in Michigan have not attracted obligate grassland birds despite
the good intentions of organizations such as the Institute. We did not expect to find even the
largest restored prairies virtually empty of obligate grasslands birds. If obligate grassland birds
are one of the main drivers for prairie restorations, then current restoration methods need to be
adapted to replicate the vegetative structure of sites with obligate grassland bird populations or

Hindy, Mangione and Keys 15
adjust management practices to better exemplify other Midwest prairie sites where obligate
grassland birds are present.
While obligate grassland birds can inhabit small grasslands (9-13 ha), they were more
abundant in extensive grasslands (Herkert 1998; Swanson 1998; Winter and Faaborg 1999;
Dechant et al. 2001). Therefore, removing additional fencerows to connect all of the prairies at
the Institute could be beneficial. In addition, reducing the impact of warm-season grasses,
specifically avoiding a big bluestem monoculture, by planting the prairies with fewer grasses and
more forbs would make restored prairies more similar to historic Michigan prairies (Grman et al.
2013).
Prescribed fire has been the management practice of choice for many Michigan
organizations managing restored prairies. Fire is a necessary component of the prairie ecosystem,
but it must be done little by little over many years (Schramm 1990). It is effective in reducing the
litter depth, but only in the first few years (Zimmerman 1988). Some obligate birds however,
avoid sites burned in the last 3 years (Cully and Michaels 2000). Since the Institute has not had
obligate birds, prescribed fire might not be enough for the prairies to be attractive to grassland
birds.
Another management practice that has been fairly common is mowing. Mowing is
usually done in the late summer after the birds have nested and fledged (NRCS 1999). This can
be efficient in controlling growth of unwanted plants and trees. Studies have suggested that
mowing is not a good substitute for fire, as it may cause negative results on plant productivity
(Schramm 1990). While it is not the most natural method of management, it has been the chosen
method of the Gerald R Ford International Airport, which has the highest grassland bird diversity
in Southwest Michigan.

Hindy, Mangione and Keys 16
The last popular management practice is rotational grazing. Grazing is allowing certain
ungulates to graze in the prairies. This creates a varied pattern of plant height in the grasslands
and can be a good control for unwanted plant growth. Certain ungulates, such as elk and bison,
only eat grasses and leave the forbs (Undersander et al. 2000), which makes them the best
choices for grazing over goats, horses, cows, and sheep. Grassland birds have been shown to
benefit from rotational grazing (Undersander et al. 2000).
All of these management techniques have been used throughout the Midwest. Each
management practice has various benefits and each has been shown to increase bird biodiversity.
Finding what best works for Southwest Michigan will likely require multiple management
practices. As of now, prescribed fire alone has not given the desired results, thus trying different
management techniques to inhibit warm-season plant dominance may have better results in
increasing not only obligate grassland bird populations, but in creating healthier prairie
ecosystems. Therefore, our research recommends that PCCI increases grassland size, reduce
warm-season grass dominance, and combine mowing with prescribed burning to provide more
suitable habitat for obligate grassland birds to thrive.
ACKNOWLEDGEMENTS
The authors would like to acknowledge the financial support of the Willard G. and Jesse M.
Pierce Foundation and project support of the Pierce Cedar Creek Institute without which this
project would not have occurred. We would also like to thank C Scholten, K Strydhorst, and D
Proppe from Calvin College for their collaborative support on this project. Finally, we thank the
Michigan Audubon Society, Gerald R. Ford International Airport, Kalamazoo Nature Center,
and Barry County landowners G Cullers, B Olmstead, W Alward, J Soya, and A Jones for their
support and land access throughout this project.

Hindy, Mangione and Keys 17
LITERATURE CITED
Bakker KK, Naugle DE, Higgins KF. 2002. Incorporating Landscape Attributes into Models for
Migratory Grassland Bird Conservation. Conserv. Biol. 16:1638–1646.
BirdLife International. 2012. Ammodramus henslowii. IUCN Red List Threat. Species 20143.
Brennan LA, Kuvlesky Jr. WP. 2005. North American grassland birds: an unfolding
conservation crisis? J. Wildl. Manag. 69:1–13.
Chapman KA, Brewer R. 2008. Prairie and savanna in southern lower Michigan: history,
classification, ecology. Mich. Bot. 47:1–48.
Cully JF, Michaels HL. 2000. Henslow’s sparrow habitat associations on Kansas tallgrass
prairie. Wilson Bull. 112:115–123.
Cushman D, Johnson T, Keys R. 2013. Using electronic field recordings to increase the
detectability of bird species at point count sites. Pierce Cedar Creek Institute.
Dechant J, Sondreal M, Johnson D, Igl L, Goldade C. 2001. Effects of Management Practices on
GrasslandBirds: Bobolink. [accessed 2016 Sep 2].
http://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1123&context=usgsnpwrc
eBird Basic Dataset. Version: EBD_relDecember-2014. 2014. Ithaca, NY: Cornell Lab of
Ornithology.
ESRI. 2015. ArcGIS Desktop: Release 10.4. Redlands, CA: Environmental Systems Research
Institute.

Hindy, Mangione and Keys 18
Fisher RJ, Davis SK. 2010. From wiens to robel: a review of grassland-bird habitat selection. J.
Wildl. Manag. 74:265–273.
Fletcher RJ, Koford RR. 2002. Habitat and landscape associations of breeding birds in native and
restored grasslands. J. Wildl. Manag. 66:1011–1022.
Grman E, Bassett T, Brudvig LA. 2013. EDITOR’S CHOICE: Confronting contingency in
restoration: management and site history determine outcomes of assembling prairies, but
site characteristics and landscape context have little effect. J. Appl. Ecol. 50:1234–1243.
Herkert JR. 1994. The effects of habitat fragmentation on Midwestern grassland bird
communities. Ecol. Appl. 4:461–471.
Herkert JR. 1998. Effects of management practices on grassland birds: Henslow’s sparrow.
Jamestown, ND: U.S. Geological Survey. [accessed 2016 Sep 2].
http://pubs.er.usgs.gov/publication/70159918
Herkert JR, Reinking DL, Wiedenfeld DA, Winter M, Zimmerman JL, Jensen WE, Finck EJ,
Koford RR, Wolfe DH, Sherrod SK, et al. 2003. Effects of prairie fragmentation on the
nest success of breeding birds in the midcontinental United States. Conserv. Biol.
17:587–594.
Howell J. 2015. Current Prairie Management Practices at Pierce Cedar Creek Institute, Barry
County, Michigan.

Hindy, Mangione and Keys 19
Jaster L, Jensen WE, Forbes AR. 2013. Abundance, territory sizes, and pairing success of male
Henslow’s sparrows in restored warm- and cool-season grasslands. J. Field Ornithol.
84:234–241.
Jobin B, Falardeau G. 2010. Habitat Associations of Grasshopper Sparrows in Southern Québec.
Northeast. Nat. 17:135–146.
Johnson DH, Igl LD. 2001. Area requirements of grassland birds: A regional perspective. The
Auk 118:24–34.
Kindscher K, Tieszen L. 1998. Floristic and Soil Organic Matter Changes after Five and Thirty-
Five Years of Native Tallgrass Prairie Restoration. [accessed 2016 Sep 2].
http://www.wildflower.org/lrp_client_docs/docs_edgworkshops/Webinar%202:%20Site
%20Preparation/Kindscher%201998%20-%20Floristic%20and%20Soil.pdf
Lambert W, Lozon D, Keys R. 2015. The influence of habitat and landscape associations on
breeding birds in managed grasslands of Southwest Michigan. Pierce Cedar Creek
Institute.
MacArthur RH, Wilson EO. 1967. The theory of island biogeography. Princeton: Princeton
University Press.
McCoy TD, Ryan MR, Burger, Jr. LW, Kurzejeski EW. 2001. Grassland bird conservation: CP1
vs. CP2 plantings in conservation reserve program fields in Missouri. Am. Midl. Nat.
145:1–17.

Hindy, Mangione and Keys 20
Navarette-Tindall N. 2010. Native cool-seaon grasses in Missouri. [accessed 2016 Sep 2].
http://www.moprairie.org/wp-content/uploads/2013/04/NativeCool-
SeasonGrassesinMissouri_MPF_Vol31No02_Page20-25.pdf
Norment C. 2002. On grassland bird conservation in the northeast. The Auk 119:271–279.
NRCS. 1999. Grassland Birds. [accessed 2016 Sep 1].
http://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs143_009930.pdf
Oosterhouse J, Vander Geld A, Keys R. 2012. Analysis of vegetative structure in the choice of
nesting habitat by midwestern songbirds. Pierce Cedar Creek Institute.
Parker M. 2012. Landowner Incentive Program: Wildlife Habitat Managemnet Program.
[accessed 2016 Aug 24].
http://www.mynaturecenter.org/Resources/Documents/Prairie%20Restoration%20PLan.p
df
Rahmig CJ, Jensen WE, With KA. 2009. Grassland bird responses to land management in the
largest remaining tallgrass prairie. Conserv. Biol. 23:420–432.
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF. 1993. Handbook of field methods for
monitoring landbirds. Lincoln, NE: USDA Forest Service, University of Nebraska -
Lincoln.
Ribic CA, Sample DW. 2001. Associations of grassland birds with landscape factors in southern
Wisconsin. Am. Midl. Nat. 146:105–121.

Hindy, Mangione and Keys 21
Ruth J. 2015. Status Assessment and Conservation Plan for the Grasshopper Sparrow
(Ammodramus savannarum). [accessed 2016 Sep 2]. https://www.fws.gov/mountain-
prairie/species/birds/grasshoppersparrow/GRSP%20Status%20Assessment%20and%20C
onservation%20Plan%20FINAL.pdf
Schramm P. 1990. Prairie Restoration: A 25 year perspective on establishment and management.
[accessed 2016 Sep 1].
http://images.library.wisc.edu/EcoNatRes/EFacs/NAPC/NAPC12/reference/econatres.na
pc12.pschramm2.pdf
Skinner RM, editor. 1975. Grassland Use Patterns and Prairie Bird Populations in Missouri. In:
Prairie: A Multiple View. p. 171–180.
Smith MA. 2008. Robel pole technique and data interpretation. University of Wyoming
Cooperative Extension Service
Swanson DA. 1998. Effects of management practices of grassland birds: Savannah Sparrow.
Jamestown, ND: U.S. Geological Survey. [accessed 2016 Sep 2].
http://pubs.er.usgs.gov/publication/70159923
Thomas L, Buckland ST, Rexstad EA, Laake JL, Strindberg S, Hedley SL, Bishop JRB, Marques
TA, Burnham KP. 2010. Distance software: design and analysis of distance sampling
surveys for estimating population size. J. Appl. Ecol. 47:5–14.
Undersander D, Temple S, Sample D, Bartlet J, Paine L. 2000. Grassland Birds: Fostering
habitats using rotational grazing. [accessed 2016 Sep 1].
http://learningstore.uwex.edu/assets/pdfs/A3715.pdf

Hindy, Mangione and Keys 22
Winter M, Faaborg J. 1999. Patterns of Area Sensitivity in Grassland-Nesting Birds. Conserv.
Biol. 13:1424–1436.
Winter M, Johnson DH, Shaffer JA. 2005. Variability in vegetation effects on density and
nesting of grassland birds. Boal, editor. J. Wildl. Manag. 69:185–197.
Zimmerman J. 1988. Breeding season habitat selection by the Henslow’s sparrow (Ammodramus
henslowii) in Kansas. [accessed 2016 Sep 2].
https://sora.unm.edu/sites/default/files/journals/wilson/v100n01/p0017-p0024.pdf

Hindy, Mangione and Keys 23
APPENDIX A
Grassland site locations (with GPS) surveyed from 23 May to 19 July, 2016, in Barry,
Kalamazoo and Kent Counties, Michigan USA.
Grassland Name Type Size (ha) County GPS
Cullers RP 1 Barry 42°33'33.7"N, 85°17'58.6"W Harris East RP 3 Kalamazoo 42°19'43.8"N, 85°40'07.6"W Harris North RP 4 Kalamazoo 42°19'54.0"N, 85°40'20.3"W Harris West RP 7 Kalamazoo 42°19'45.8"N, 85°40'18.9"W Olmstead East MM 7 Barry 42°33'1.62"N, 85°13'50.43"W
Olmstead North RP 7 Barry 42°32'58.69"N, 85°14'4.37"W
Alward RP 9 Barry 42°33'13.1"N, 85°17'43.7"W Jones MM 9 Barry 42°32'55.65"N, 85°18'22.98"W
GRR North MM 9 Kent 42°53'49.9"N, 85°31'13.7"W PCCI East RP 10 Barry 42°32'0.38"N, 85°18'4.89"W
PCCI North RP 11 Barry 42°32'30.91"N, 85°18'9.36"W
PCCI West RP 12 Barry 42°32'5.23"N, 85°18'18.12"W
Olmstead South MM 13 Barry 42°32'46.96"N, 85°13'58.67"W
Otis Farm RP 15 Barry 42°36'05.1"N, 85°23'20.4"W Olmstead Southeast MM 16 Barry 42°32'53.22"N, 85°13'43.85"W
GRR Mid MM 24 Kent 42°52'32.8"N, 85°31'14.8"W Soya West MM 27 Barry 42°32'5.13"N, 85°16'57.19"W
Soya East MM 46 Barry 42°32'1.34"N, 85°16'29.15"W
KNC Prairie RP 59 Barry 42°21'09.1"N, 85°35'11.3"W GRR South MM 61 Kent 42°51'33.50"N, 85°31'14.51"W