herpesvirus active replication in multiple sclerosis: a genetic control?
TRANSCRIPT

Journal of the Neurological Sciences 311 (2011) 98–102
Contents lists available at SciVerse ScienceDirect
Journal of the Neurological Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / jns
Herpesvirus active replication in multiple sclerosisA genetic control?
Marta Garcia-Montojo a, Alfonso Martinez b, Virginia De Las Heras a, Maria Inmaculada Dominguez-Mozo a,Maria del Carmen Cenit b, Maria López-Cavanillas b, Angel Garcia-Martinez a, Ana Maria Arias-Leal a,Emilio Gomez de la Concha b, Elena Urcelay b, Rafael Arroyo a, Roberto Alvarez-Lafuente a,⁎a Servicio de Neurología, Hospital Clínico San Carlos, Madrid 28040, Spainb Servicio de Inmunología, Hospital Clínico San Carlos, Madrid 28040, Spain
⁎ Corresponding author at: Neurology Service, HospitaMartín Lagos s/n. 28040 Madrid, Spain. Tel.: +34 913303457.
E-mail address: [email protected] (R. Alvar
0022-510X/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.jns.2011.09.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 29 April 2011Received in revised form 1 September 2011Accepted 5 September 2011Available online 1 October 2011
Keywords:HHV-6MHC2TACD46Multiple sclerosisPCR
Although the etiology of multiple sclerosis (MS) is unknown, it is generally believed that genetic, immuno-logic, and environmental factors are involved. The objectives of this study were: 1. to analyze if a genetic con-trol could explain why HHV-6 would be able to actively replicate in a subset of MS patients but not incontrols; 2. to study if MS patients with HHV-6 active replication are clinically different from those withoutHHV-6 active replication. A total of 195 MS patients and 195 controls were analyzed for two SNPs at theMHC2TA locus and two SNPs at the CD46 locus. Furthermore, the MS cohort was analyzed by PCR for thedetection of HHV-6 genomes in five serum samples collected every six months along two-year follow-up.We found that 59/195 (30.2%) MS patients had at least one HHV-6 positive serum sample. No statistical sig-nificant difference was found for the two genes when the comparison was made between MS patients andcontrols; however, a statistical significance was found for the two polymorphisms of MHC2TA when we com-pared MS patients with active replication and controls (p=0.0000004 for rs4774C and p=0.011 forrs3087456G). Furthermore, increased significant differences were found for MHC2TA and CD46 when wecompared interferon beta responders and non-responders within MS patients. In conclusion, we describe agene–environment interaction in MS patients between HHV-6 and MHC2TA and CD46 that should be furtherstudied to clarify if that interaction could be a genetic control. The results show that MS patients withoutHHV-6 active replication are better responders to interferon beta treatment than those with HHV-6 activereplication.
l Clínico San Carlos, C./Profesor3303000x7440; fax: +34 91
ez-Lafuente).
l rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Multiple sclerosis (MS) is the most common inflammatory diseaseof the central nervous system. The pathogenesis of MS is not wellunderstood; however, there are strong evidences for an autoimmunecomponent. Although the etiology of MS is unknown, it is generallybelieved that genetic, immunologic, and environmental factors areinvolved. Viruses have been long associated with MS, and amongthem, human herpesviruses like human herpesvirus 6 (HHV-6) havebeen deeply studied in the last years. The HHV-6 is a lymphotropicvirus closely related to human cytomegalovirus (CMV) and humanherpesvirus-human 7 (HHV-7). It has a worldwide distribution anda high prevalence in the general adult population [1]. Many recent
publications that have been published on MS and viruses analyzethe active replication rate of HHV-6 in MS patients versus controlsthrough the analysis of the presence of HHV-6 genomes in serumsamples [2–24]. As we can see in Table 1, while the mean DNAprevalence of HHV-6 in MS patients is more than seventeen percent, controls only reach one point eight; then, the active replicationof HHV-6 in MS patients increased tenfold. If we take together allthese results, we would obtain a high statistically significant differ-ence (p=0.000001, odds ratio=11.7). Then, is there a genetic con-trol that could explain that a high prevalent virus in the worldwidepopulation is able to actively replicate in a subset of MS patients butnot in controls? Are these MS patients with HHV-6 active replicationclinically different from those without HHV-6 active replication? Tostudy these questions we analyzed in a MS cohort: two polymor-phisms of MHC2TA, a gene that has been previously associated withMS, [25] and also with HHV-6 [20], two polymorphisms of CD46,the gene that codes for the cellular receptor of HHV-6, [26] and theHHV-6 replicative status through the analysis of HHV-6 genomesin serum samples by quantitative real-time PCR along two-yearfollow-up.

Table 1Detection of HHV-6 in serum/plasma from MS cases and controls by PCR.
Year Authors MS cases Controls
+/n % +/n %
1994 Wilborn et al. [2] 0/21 0 0/16 01997 Martin et al. [3] 0/20 0 0/20 01997 Soldan et al. [4] 15/50 30.0 0/47 01998 Ablashi et al. [5] 1/3 33.3 –/– –
1998 Fillet et al. [6] 2/32 6.3 1/34 2.91999 Goldberg et al. [7] 1/24 4.2 0/30 01999 Mirandola et al. [8] 0/32 0 0/12 02000 Akhyani et al. [9] 8/34 23.5 0/19 02001 Tomsone et al. [10] 14/38 36.8 0/43 02002 Berti et al. [11] 39/167 23.4 0/70 02002 Gutiérrez et al. [12] 0/41 0 0/98 02002 Tejada-Simon et al. [13] 22/33 66.7 7/21 33.32003 Al-Shammari et al. [14] 0/24 0 1/33 3.02003 Chapenko et al. [15] 8/26 30.8 0/150 02003 Tejada-Simon et al. [16] 10/12 83.3 6/11 54.52003 Villoslada et al. [17] 0/151 0 0/50 02004 Alvarez-Lafuente et al. [18] 17/105 16.2 0/49 02005 Fögdell-Hahn et al. [19] 4/42 9.5 0/123 02007 Martinez et al. [20] 22/99 22.2 –/– –
2008 Kuusisto et al. [21] 0/34 0 –/– –
2009 Ahram et al. [22] 9/36 25 –/– –
2009 Franciotta et al. [23] 0/54 0 0/25 02010 Alvarez-Lafuente et al. [24] 41/149 27.5 –/– –
213/1227 17.4 15/851 1.8
99M. Garcia-Montojo et al. / Journal of the Neurological Sciences 311 (2011) 98–102
2. Materials and methods
2.1. Subjects
A total of 195 patients with clinically definite relapsing–remittingMS (RRMS) were included in the study (63 males, age ranging be-tween 21 and 59 years, and 132 females, age ranging between 20and 56 years). All patients were characterized as having RRMS formore than 2 years. These patients had been treated, at least, duringtwo years, with interferon beta: IFN-beta-1a (Avonex, n=28) 30 μgintramuscularly once weekly, IFN-beta-1b (Betaferon, n=101)8 MIU subcutaneously every other day, or IFN-beta-1a (Rebif,n=66) 22 μg subcutaneously three times weekly, for more thantwo years. A control group of 195 healthy Spanish individuals wasincluded for comparative purposes in the genetic analyses. RRMSpatients and controls were paired by age and sex. This study wasapproved by the local Ethic Committee and all the participants gaveinformed consent before the enrolment.
2.2. Clinical data
EDSS of the patients was determined at each visit. At the end ofthe study we analyzed the following variables: 1) proportion ofpatients with and without relapses (the definition of relapse was: aperiod in which a person with MS experiences an acute worseningof function that lasts for at least 24 h followed by an improvementthat lasts for at least one month, confirmed by neurological examina-tion), 2) number of relapses, 3) months elapsed between the first andthe second relapses, 4) proportion of patients experiencing progres-sion in the EDSS score (the definition of progression was differentdepending on the pre-treatment EDSS score: i) increase ≥1.5 pointsif pre-treatment EDSS=0; ii) increase ≥1 point if pre-treatmentEDSS≥1 and ≤5; iii) increase ≥0.5 points if pre-treatmentEDSS≥5.5; to be regarded as progression, the increase in EDSSscore had to persist in a visit performed at least three months later),5) MS patients without progression in the first two years after diag-nosis, 6)response to interferon-beta treatment (that was defined asabsence of relapses and progression after two years of treatment),7) sex, 8) starting age, 9) starting symptoms (optical neuritis, brainstem, spinal cord, and other).
2.3. Samples
At the time of visit 10 ml of peripheral blood were drawn by veinpuncture into sterile tubes with EDTA and directly used for DNA extrac-tion, and 2 ml of serumwere isolated in serum separator tubes by cen-trifugation and then were separate into aliquots and stored at −80 °C.Among RRMS patients, two samples, blood and serum, were collectedat the recruitment, and a second pair of sampleswere obtained six (ran-ged 5–7), twelve (ranged 11–13), eighteen (ranged 16–19), and twentyfourmonths (ranged 22–26) after the recruitment; all the sampleswerecollected when the MS patients were in remission. For the controlgroup, we only collected one blood sample for each patient. For thegenetic study, only basal samples were analyzed.
2.4. DNA extraction
Total DNA was isolated by DNA spin column technique of QIAampDNA BloodMini Kit (QIAGEN, Hilden, Germany), from 0.2 ml of blood,and QIAamp Ultrasens Virus Kit (QIAGEN), from 1 ml of serum,according to the manufacturer's instructions.
2.5. MHC2TA genotyping
Two polymorphisms in the MHC2TA gene (rs4774 and rs3087456)were analyzed by TaqMan Assay-on-Demand (C___381733_10) andAssay-by-Design (C__15793789_10), respectively, fromApplied Biosys-tems (Foster City, CA, USA), following manufacturer's suggestions. Bothgenetic markers conformed to Hardy–Weinberg equilibrium propor-tions in the control population (pN0.05).
2.6. CD46 genotyping
Two polymorphisms in the CD46 gene were analyzed: A/G locatedat the 5′ flanking sequence near gene (rs2796267), and A/T located atthe intron 4 (rs2724385). TaqMan Assay-on-Demand assays wereused under conditions recommended by the manufacturer (AppliedBiosystems, Foster City, CA, USA) in a HT7900 fast real time PCR plat-form. Genetic markers conformed to Hardy–Weinberg equilibriumproportions in the control population (pN0.05).
2.7. Quantitative real-time PCR for HHV-6
PCR assays were performed in a Rotor-Gene 3000 (CorbettResearch, Sydney, Australia). Primers and probes for detection ofHHV-6 and PCR protocol are previously published [27].
2.8. Statistical analysis
Qualitative variables were presented by its frequency distribution.The association among qualitative variables was analyzed by the Fisherexact test. Quantitative variables were studied through its medianvalues. Kruskall–Wallis analysis was used to test differences in continu-ous variables. We considered statistically significant differences whenpb0.05. For the genetic study, allele and genotype frequencies in pa-tients and controlswere compared by the χ2 test; p values were consid-ered significant at a level of b0.05. Odds ratio (OR) and p values werecalculated using a standard computer package (Epi Info v. 6.02, CDC,Atlanta, USA).
3. Results
3.1. HHV-6 prevalence
When we analyzed the prevalences of HHV-6 in the five serumsamples collected in each MS patient along the two-year follow-up
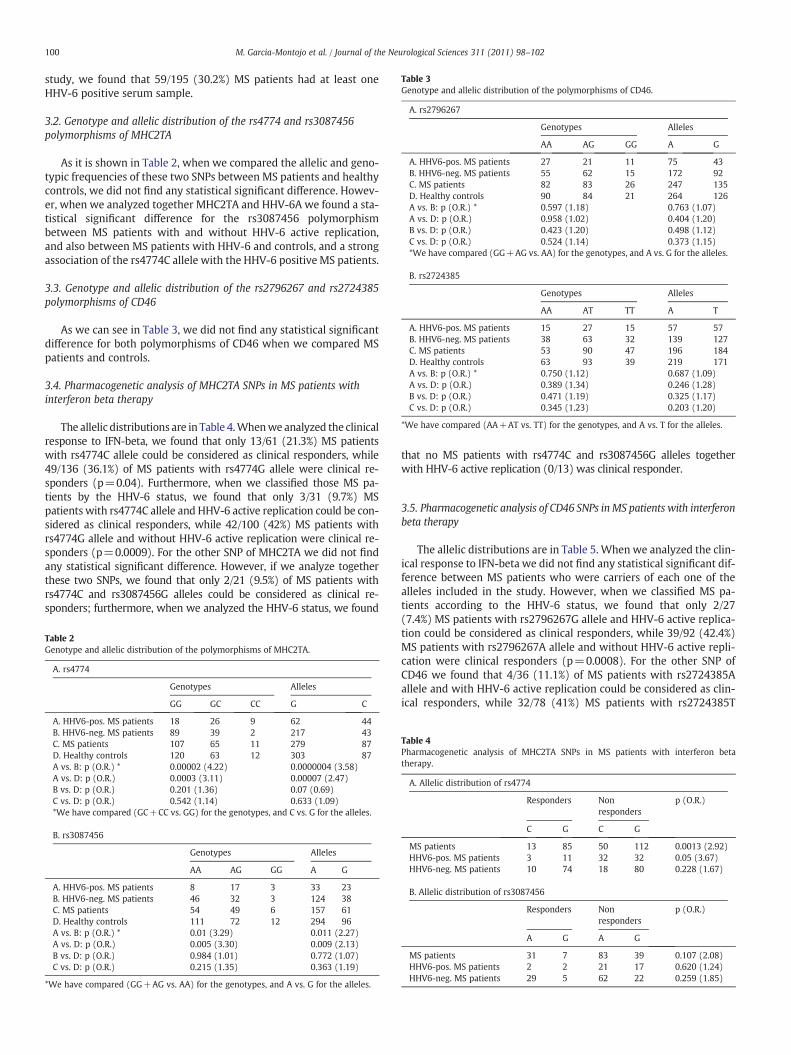
Table 3Genotype and allelic distribution of the polymorphisms of CD46.
A. rs2796267
Genotypes Alleles
AA AG GG A G
A. HHV6-pos. MS patients 27 21 11 75 43B. HHV6-neg. MS patients 55 62 15 172 92C. MS patients 82 83 26 247 135D. Healthy controls 90 84 21 264 126A vs. B: p (O.R.) * 0.597 (1.18) 0.763 (1.07)A vs. D: p (O.R.) 0.958 (1.02) 0.404 (1.20)B vs. D: p (O.R.) 0.423 (1.20) 0.498 (1.12)C vs. D: p (O.R.) 0.524 (1.14) 0.373 (1.15)*We have compared (GG+AG vs. AA) for the genotypes, and A vs. G for the alleles.
B. rs2724385
Genotypes Alleles
AA AT TT A T
A. HHV6-pos. MS patients 15 27 15 57 57B. HHV6-neg. MS patients 38 63 32 139 127C. MS patients 53 90 47 196 184D. Healthy controls 63 93 39 219 171A vs. B: p (O.R.) * 0.750 (1.12) 0.687 (1.09)A vs. D: p (O.R.) 0.389 (1.34) 0.246 (1.28)B vs. D: p (O.R.) 0.471 (1.19) 0.325 (1.17)C vs. D: p (O.R.) 0.345 (1.23) 0.203 (1.20)
*We have compared (AA+AT vs. TT) for the genotypes, and A vs. T for the alleles.
100 M. Garcia-Montojo et al. / Journal of the Neurological Sciences 311 (2011) 98–102
study, we found that 59/195 (30.2%) MS patients had at least oneHHV-6 positive serum sample.
3.2. Genotype and allelic distribution of the rs4774 and rs3087456polymorphisms of MHC2TA
As it is shown in Table 2, when we compared the allelic and geno-typic frequencies of these two SNPs between MS patients and healthycontrols, we did not find any statistical significant difference. Howev-er, when we analyzed together MHC2TA and HHV-6A we found a sta-tistical significant difference for the rs3087456 polymorphismbetween MS patients with and without HHV-6 active replication,and also between MS patients with HHV-6 and controls, and a strongassociation of the rs4774C allele with the HHV-6 positive MS patients.
3.3. Genotype and allelic distribution of the rs2796267 and rs2724385polymorphisms of CD46
As we can see in Table 3, we did not find any statistical significantdifference for both polymorphisms of CD46 when we compared MSpatients and controls.
3.4. Pharmacogenetic analysis of MHC2TA SNPs in MS patients withinterferon beta therapy
The allelic distributions are in Table 4.Whenwe analyzed the clinicalresponse to IFN-beta, we found that only 13/61 (21.3%) MS patientswith rs4774C allele could be considered as clinical responders, while49/136 (36.1%) of MS patients with rs4774G allele were clinical re-sponders (p=0.04). Furthermore, when we classified those MS pa-tients by the HHV-6 status, we found that only 3/31 (9.7%) MSpatients with rs4774C allele and HHV-6 active replication could be con-sidered as clinical responders, while 42/100 (42%) MS patients withrs4774G allele and without HHV-6 active replication were clinical re-sponders (p=0.0009). For the other SNP of MHC2TA we did not findany statistical significant difference. However, if we analyze togetherthese two SNPs, we found that only 2/21 (9.5%) of MS patients withrs4774C and rs3087456G alleles could be considered as clinical re-sponders; furthermore, when we analyzed the HHV-6 status, we found
Table 2Genotype and allelic distribution of the polymorphisms of MHC2TA.
A. rs4774
Genotypes Alleles
GG GC CC G C
A. HHV6-pos. MS patients 18 26 9 62 44B. HHV6-neg. MS patients 89 39 2 217 43C. MS patients 107 65 11 279 87D. Healthy controls 120 63 12 303 87A vs. B: p (O.R.) * 0.00002 (4.22) 0.0000004 (3.58)A vs. D: p (O.R.) 0.0003 (3.11) 0.00007 (2.47)B vs. D: p (O.R.) 0.201 (1.36) 0.07 (0.69)C vs. D: p (O.R.) 0.542 (1.14) 0.633 (1.09)*We have compared (GC+CC vs. GG) for the genotypes, and C vs. G for the alleles.
B. rs3087456
Genotypes Alleles
AA AG GG A G
A. HHV6-pos. MS patients 8 17 3 33 23B. HHV6-neg. MS patients 46 32 3 124 38C. MS patients 54 49 6 157 61D. Healthy controls 111 72 12 294 96A vs. B: p (O.R.) * 0.01 (3.29) 0.011 (2.27)A vs. D: p (O.R.) 0.005 (3.30) 0.009 (2.13)B vs. D: p (O.R.) 0.984 (1.01) 0.772 (1.07)C vs. D: p (O.R.) 0.215 (1.35) 0.363 (1.19)
*We have compared (GG+AG vs. AA) for the genotypes, and A vs. G for the alleles.
that no MS patients with rs4774C and rs3087456G alleles togetherwith HHV-6 active replication (0/13) was clinical responder.
3.5. Pharmacogenetic analysis of CD46 SNPs inMS patients with interferonbeta therapy
The allelic distributions are in Table 5. When we analyzed the clin-ical response to IFN-beta we did not find any statistical significant dif-ference between MS patients who were carriers of each one of thealleles included in the study. However, when we classified MS pa-tients according to the HHV-6 status, we found that only 2/27(7.4%) MS patients with rs2796267G allele and HHV-6 active replica-tion could be considered as clinical responders, while 39/92 (42.4%)MS patients with rs2796267A allele and without HHV-6 active repli-cation were clinical responders (p=0.0008). For the other SNP ofCD46 we found that 4/36 (11.1%) of MS patients with rs2724385Aallele and with HHV-6 active replication could be considered as clin-ical responders, while 32/78 (41%) MS patients with rs2724385T
Table 4Pharmacogenetic analysis of MHC2TA SNPs in MS patients with interferon betatherapy.
A. Allelic distribution of rs4774
Responders Nonresponders
p (O.R.)
C G C G
MS patients 13 85 50 112 0.0013 (2.92)HHV6-pos. MS patients 3 11 32 32 0.05 (3.67)HHV6-neg. MS patients 10 74 18 80 0.228 (1.67)
B. Allelic distribution of rs3087456
Responders Nonresponders
p (O.R.)
A G A G
MS patients 31 7 83 39 0.107 (2.08)HHV6-pos. MS patients 2 2 21 17 0.620 (1.24)HHV6-neg. MS patients 29 5 62 22 0.259 (1.85)

Table 5Pharmacogenetic analysis of CD46 SNPs in MS patients with interferon beta therapy.
A. Allelic distribution of rs2796267
Responders Nonresponders
p (O.R.)
A G A G
MS patients 69 29 109 61 0.294 (1.33)HHV6-pos. MS patients 11 3 43 27 0.222 (2.30)HHV6-neg. MS patients 58 26 66 34 0.660 (1.15)
B. Allelic distribution of rs2724385
Responders Nonresponders
p (O.R.)
A T A T
MS patients 45 53 87 83 0.407 (1.23)HHV6-pos. MS patients 5 9 37 31 0.202 (2.15)HHV6-neg. MS patients 40 44 50 52 0.849 (1.06)
101M. Garcia-Montojo et al. / Journal of the Neurological Sciences 311 (2011) 98–102
allele and without HHV-6 active replication were clinical responders(p=0.001).
4. Discussion
As we have shown in this study, a gene–environment interactionoccurs in MS patients. In a cohort of 195 MS patients, 59 (30.2%)had detectable HHV-6 in their serum samples at least once alongthe two-years follow-up study; this percentage of HHV-6 positivityin MS patients is significantly higher than the results obtains for con-trols, as we can see in Table 1. And this condition, the presence or theabsence of HHV-6 seems to be related to the presence or the absenceof different polymorphisms in the two genes that we have included inthis study. While no statistical significant differences could be foundfor these two genes when the comparison was made between MS pa-tients and controls, higher statistical significations were found for thetwo polymorphisms of MHC2TA when we compared MS patientswith active replication and controls. Furthermore, more statistical sig-nifications were found for MHC2TA and CD46 when we comparedthose MS patients that could be considered or not as clinical re-sponders to IFN-beta. The aim of this study was to answer two ques-tions: is there a genetic control that could explain that a highprevalent virus in the worldwide population were able to activelyreplicate in a subset of MS patients but not in controls? Are theseMS patients with HHV-6 active replication clinically different fromthose without HHV-6 active replication? Regarding the second ques-tion, our results show that those MS patients without HHV-6 activereplication are better responders to IFN-beta treatment than thoseMS patients with HHV-6 active replication, as we have shown inother previous study [28]. Then, we returned to the first question: isthere a genetic control? These two genes, MHC2TA and CD46, are re-lated to herpesviruses. MHC2TA gene is chromosome 16; this genecodes for a transcription factor (named CIITA) involved in MHCclass II expression [29]. Although CIITA does not bind directly toDNA, it regulates the transcription by recruiting additional transcrip-tion factors and cofactors, particularly chromatin modifiers [30]. Thisgene had a triple interest in our study: first, the strongest (and tosome researchers, the only) bona fide genetic factor associated withthe disease is DRB1*1501, an allele of the HLA class II molecule DR.The expression of these molecules is regulated by MHC2TA. Second,it has been recently reported that a promoter polymorphism(−168A/G) in this MHC2TA gene is associated with MS and rheuma-toid arthritis. Third, it has been described that cytomegalovirus (CMV,another b-herpesvirus) eludes immune detection at least in part byinhibiting HLA class II expression by means of down-regulatinginterferon-g-mediated expression of MHC2TA [31]. Then, we could
speculate with a similar interaction for HHV-6, that should be deeperstudied; furthermore, both polymorphisms described in this studymust be analyzed in order to establish their effect on MHC2TAexpression, and therefore, their possible effect on HHV-6 active repli-cation. Regarding CD46, it was first identified as the membrane cofac-tor protein (MCP) and a member of the regulators of complementgene cluster (RCA). CD46 is a cell surface, type 1 transmembrane gly-coprotein of 57–67 kD ubiquitously expressed on the surface of allnucleated human cells [32]; it belongs to the family of complementactivation regulators and binds complement factors to prevent selfcell destruction, and it has been also identified as a receptor for sever-al human pathogens: Measles virus [33], Streptococcus pyogenes [34],Neisseria gonorrhoeae [35], Human herpesvirus 6 (HHV-6), and vari-ous group B adenovirus [36]. Finally, several CD46 isoforms (mem-brane and soluble) can be expressed on T cells as a result ofalternative splicing [37]. Then, differences in CD46 gene could yielddifferent isoforms that can join HHV-6 conditioning its entry in thecell. Again, further studies are required for the polymorphisms de-scribed in this study in order to analyze their effect in CD46 expres-sion and their possible effect on HHV-6 active replication.
In conclusion, here we describe a gene–environment interactionin MS patients between HHV-6 and MHC2TA and CD46 that shouldbe deeper study to clarify if that interaction could be a genetic con-trol; that control could condition the replication status of HHV-6and also, the clinical evolution of the disease in a subset of MSpatients.
Conflict of interest
No conflicts of interest.
Acknowledgments
Roberto Alvarez-Lafuente is recipient of a research contract of theInstituto de Salud Carlos III-Fondo Europeo de Desarrollo Regional(Feder) (CP07/00273). Garcia-Martinez is recipient of a techniciancontract from “REEM: Red Española de Esclerosis Múltiple” (RETICS-REEM RD07/0060; www.reem.es). This work was financially sup-ported by grants from the Instituto de Salud Carlos III-Fondo Europeode Desarrollo Regional (Feder) (FIS No 09/02074), “Fundación MutuaMadrileña”, “Fundación Alfonso Martín Escudero”, and “FundaciónLAIR”. In addition, we are indebted to María Concepción Ramírezand María Jesús Díez who collected the specimens.
References
[1] Clark DA. Human herpesvirus 6 and human herpesvirus 7: emerging pathogens intransplant patients. Int J Hematol 2002;76(Suppl. 2):246–52.
[2] Wilborn F, Schmidt CA, Brinkmann V, et al. A potential role for human herpesvirustype 6 in nervous system disease. J Neuroimmunol 1994;49:213–4.
[3] Martin C, Enbom M, Söderström M, Fredrikson S, Dahl H, Lycke J, Bergström T,Linde A. Absence of seven human herpesviruses, including HHV-6, by polymerasechain reaction in CSF and blood from patients with multiple sclerosis and opticneuritis. Acta Neurol Scand 1997;95:280–3.
[4] Soldan SS, Berti R, Salem N, et al. Association of human herpes virus 6 (HHV-6)with multiple sclerosis: increased IgM response to HHV-6 early antigen anddetection of serum HHV-6 DNA. Nat Med 1997;3:1394–7.
[5] Ablashi DV, Lapps W, Kaplan M, et al. Human herpesvirus-6 (HHV-6) infection inmultiple sclerosis: a preliminary report. Mult Scler 1998;4:490–6.
[6] Fillet AM, Lozeron P, Agut H, et al. HHV-6 and multiple sclerosis. Nat Med 1998;4:537.
[7] Goldberg SH, Albright AV, Lisak RP, et al. Polymerase chain reaction analysis ofhuman herpesvirus-6 sequences in the sera and cerebrospinal fluid of patientswith multiple sclerosis. J Neurovirol 1999;5:134–9.
[8] Mirandola P, Stefan A, Brambilla E, Campadelli-Fiume G, Grimaldi LM. Absence ofhumanherpesvirus 6 and7 from spinalfluid and serumofmultiple sclerosis patients.Neurology 1999;53:1367–8.
[9] Akhyani N, Berti R, Brennan MB, Soldan SS, Eaton JM, McFarland HF, Jacobson S.Tissue distribution and variant characterization of human herpesvirus (HHV)-6:increased prevalence of HHV-6A in patients with multiple sclerosis. J Infect Dis2000;182:1321–5.

102 M. Garcia-Montojo et al. / Journal of the Neurological Sciences 311 (2011) 98–102
[10] Tomsone V, Logina I, Millers A, et al. Association of human herpesvirus 6 andhuman herpesvirus 7 with demyelinating diseases of the nervous system. J Neuro-virol 2001;7:564–9.
[11] Berti R, Brennan MB, Soldan SS, et al. Increased detection of serum HHV-6 DNA se-quences during multiple sclerosis (MS) exacerbations and correlation withparameters of MS disease progression. J Neurovirol 2002;8:250–6.
[12] Gutiérrez J, Vergara MJ, Guerrero M, Fernández O, Piédrola G, Morales P, MarotoMC. Multiple sclerosis and human herpesvirus 6. Infection 2002;30:145–9.
[13] Tejada-Simon MV, Zang YC, Hong J, et al. Detection of viral DNA and immuneresponses to the human herpesvirus 6 101-kilodalton virion protein in patientswith multiple sclerosis and in controls. J Virol 2002;76:6147–54.
[14] Al-Shammari S, Nelson RF, Voevodin A. HHV-6 DNAaemia in patients with multi-ple sclerosis in Kuwait. Acta Neurol Scand 2003;107:122–4.
[15] Chapenko S, Millers A, Nora Z, et al. Correlation between HHV-6 reactivation andmultiple sclerosis disease activity. J Med Virol 2003;69:111–7.
[16] Tejada-Simon MV, Zang YC, Hong J, et al. Cross-reactivity with myelin basic pro-tein and human herpesvirus-6 in multiple sclerosis. Ann Neurol 2003;53:189–97.
[17] Villoslada P, Juste C, Tintore M, Llorenç V, Codina G, Pozo-Rosich P, Montalban X.The immune response against herpesvirus is more prominent in the early stagesof MS. Neurology 2003;60:1944–8.
[18] Alvarez-Lafuente R, De las Heras V, Bartolomé M, Picazo JJ, Arroyo R. Relapsing–remitting multiple sclerosis and HHV-6 active infection. Arch Neurol 2004;61:1523–7.
[19] Fogdell-Hahn A, Soldan SS, Shue S, et al. Co-purification of soluble membranecofactor protein (CD46) and human herpesvirus 6 variant A genome in serumfrom multiple sclerosis patients. Virus Res 2005;110:57–63.
[20] Martínez A, Álvarez-Lafuente R, Mas A, et al. Environment–gene interaction in mul-tiple sclerosis: human herpesvirus 6 and MHC2TA. Hum Immunol 2007;68:685–9.
[21] Kuusisto H, Hyöty H, Kares S, Kinnunen E, Elovaara I. Human herpes virus 6 andmultiple sclerosis: a Finnish twin study. Mult Scler 2008;14:54–8.
[22] AhramM, El-Omar A, Baho Y, Lubad MA. Association between human herpesvirus6 and occurrence of multiple sclerosis among Jordanian patients. Acta NeurolScand 2009;120:430–5.
[23] Franciotta D, Bestetti A, Sala S, Perucca P, Jarius S, Price RW, Di Stefano AL, CinqueP. Broad screening for human herpesviridae DNA inmultiple sclerosis cerebrospinalfluid and serum. Acta Neurol Belg 2009;109:277–82.
[24] Alvarez-Lafuente R,Martinez A, Garcia-MontojoM,Mas A, De Las Heras V, Dominguez-Mozo MI, Maria Del Carmen C, López-Cavanillas M, Bartolome M, Gomez de la ConchaE, Urcelay E, Arroyo R. MHC2TA rs4774C and HHV-6A active replication in multiplesclerosis patients. Eur J Neurol 2010;17:129–35.
[25] Swanberg M, Lidman O, Padyukov L, Eriksson P, Akesson E, Jagodic M, Lobell A,Khademi M, Börjesson O, Lindgren CM, Lundman P, Brookes AJ, Kere J, Luthman
H, Alfredsson L, Hillert J, Klareskog L, Hamsten A, Piehl F, Olsson T. MHC2TA isassociated with differential MHC molecule expression and susceptibility to rheu-matoid arthritis, multiple sclerosis and myocardial infarction. Nat Genet 2005;37:486–94.
[26] Santoro F, Kennedy PE, Locatelli G, Malnati MS, Berger EA, Lusso P. CD46 is acellular receptor for human herpesvirus 6. Cell 1999;99:817–27.
[27] Hymas W, Stevenson J, Taggart EW, Hillyard D. Use of lyophilized standards forthe calibration of a newly developed real time PCR assay for human herpes typesix (HHV6) variants A and B. J Virol Methods 2005;128:143–50.
[28] Garcia-Montojo M, De Las Heras V, Dominguez-Mozo M, Bartolome M, Garcia-Martinez MA, Arroyo R, Alvarez-Lafuente R. On behalf of the HHV-6 and MultipleSclerosis Study Group. Human herpesvirus 6 and effectiveness of interferon beta1b in multiple sclerosis patients. Eur J Neurol Apr 25 2011, doi:10.1111/j.1468-1331.2011.03410.x Epub ahead of print.
[29] LeibundGut-Landmann S, Waldburger JM, Krawczyk M, Otten LA, Suter T, FontanaA, Acha-Orbea H, Reith W. Mini-review: specificity and expression of CIITA, themaster regulator of MHC class II genes. Eur J Immunol 2004;34:1513–25.
[30] Zika E, Ting JP. Epigenetic control of MHC-II: interplay between CIITA and histone-modifying enzymes. Curr Opin Immunol 2005;17:58–64.
[31] Le Roy E, Muhlethaler-Mottet A, Davrinche C, Mach B, Davignon JL. Escape ofhuman cytomegalovirus from HLA-DR-restricted CD4(+) T-cell response is medi-ated by repression of gamma interferon-induced class II transactivator expres-sion. J Virol 1999;73:6582–9.
[32] Liszewski MK, Post TW, Atkinson JP. Membrane cofactor protein (MCP or CD46):newest member of the regulators of complement activation gene cluster. AnnuRev Immunol 1991;9:431–55.
[33] Dörig RE, Marcil A, Chopra A, Richardson CD. The human CD46 molecule is areceptor for measles virus (Edmonston strain). Cell 1999;75:295–305.
[34] Okada N, Liszewski MK, Atkinson JP, Caparon M. Membrane cofactor protein(CD46) is a keratinocyte receptor for the M protein of the group A streptococcus.Proc Natl Acad Sci U S A 1995;92:2489–93.
[35] Källström H, Liszewski MK, Atkinson JP, Jonsson AB. Membrane cofactor protein(MCP or CD46) is a cellular pilus receptor for pathogenic Neisseria. Mol Microbiol1997;25:639–47.
[36] Gaggar A, ShayakhmetovDM, Lieber A. CD46 is a cellular receptor for group B adeno-viruses. Nat Med 2003;9:1408–12.
[37] Pollard AJ, Flanagan BF, Newton DJ, Johnson PM. A novel isoform of human mem-brane cofactor protein (CD46) mRNA generated by intron retention. Gene1998;212:39–47.