ginsenoside rg5 improves cognitive dysfunction and beta-amyloid deposition in stz-induced memory...
TRANSCRIPT

International Immunopharmacology 19 (2014) 317–326
Contents lists available at ScienceDirect
International Immunopharmacology
j ourna l homepage: www.e lsev ie r .com/ locate / in t imp
Short communication
Ginsenoside Rg5 improves cognitive dysfunction and beta-amyloiddeposition in STZ-induced memory impaired rats via attenuatingneuroinflammatory responses
Shenghui Chu a,1, Junfei Gu b,e,1, Liang Feng b,⁎, Jiping Liu d, Minghua Zhang b, Xiaobin Jia b,Min Liu c,⁎⁎⁎, Danian Yao a,⁎⁎a College of Agriculture, Anhui Agricultural University, Anhui, Hefei, 230036, PR Chinab Key Laboratory of New Drug Delivery Systems of Chinese Meteria Medica, Jiangsu Provincial Academy of Chinese Medicine, Jiangsu, Nanjing, 210028, PR Chinac Department of Pharmacy, Wenzhou Medical University, Wenzhou, Zhejiang, 325035, PR Chinad Department of Pharmacology, Shaanxi University of Chinese Medicine, Xianyang 712046, PR Chinae Department of Pharmacy, Nanjing University of Chinese Medicine, Jiangsu, Nanjing, 210023, P. R. China
⁎ Correspondence to: L. Feng, 100# Shizi street, HongTel.: +86 25 856378091; fax: +86 25 85637809.⁎⁎ Correspondence to: D. Yao, No. 130# ChangjiangTel.: +86 551 5786213; fax: +86 551 5786869.⁎⁎⁎ Correspondence to: M. Liu, 82#, XueyuanzhonTel./fax: +86 577 86699350.
E-mail addresses:[email protected] (L. Feng), [email protected] (D. Yao).
1 Shenghui Chu and Junfei Gu contributed equally to th
1567-5769/$ – see front matter © 2014 Elsevier B.V. All rihttp://dx.doi.org/10.1016/j.intimp.2014.01.018
a b s t r a c t
a r t i c l e i n f oArticle history:Received 18 December 2013Received in revised form 7 January 2014Accepted 15 January 2014Available online 4 February 2014
Keywords:Ginsenoside Rg5β-AmyloidAlzheimer's diseaseInflammation
Neuroinflammatory responses play a crucial role in the pathogenesis of Alzheimer's disease (AD). GinsenosideRg5 (Rg5), an abundant natural compound in Panax ginseng, has been found to be beneficial in treating AD. Inthe present study, we demonstrated that Rg5 improved cognitive dysfunction and attenuatedneuroinflammatory responses in streptozotocin (STZ)-induced memory impaired rats. Cognitive deficits wereameliorated with Rg5 (5, 10 and 20 mg/kg) treatment in a dose-dependent manner together with decreasedlevels of inflammatory cytokines TNF-α and IL-1β (P b 0.05) in brains of STZ rats. Acetylcholinesterase (AChE)activity was also significantly reduced by Rg5 whereas choline acetyltransferase (ChAT) activity was remarkablyincreased in the cortex and hippocampus of STZ-induced AD rats (P b 0.05). In addition, Congo red and immuno-histochemistry staining results showed that Rg5 alleviated Aβ deposition but enhanced the expressions ofinsulin-like growth factors 1 (IGF-1) and brain derived neurophic factor (BDNF) in the hippocampus and cerebralcortex (P b 0.05). Western blot analysis also demonstrated that Rg5 increased remarkably BDNF and IGF-1expressions whereas decreased significantly Aβ deposits (P b 0.05). Furthermore, it was observed that theexpressions of COX-2 and iNOS were significantly up-regulated in STZ-induced AD rats and down-regulatedstrongly (P b 0.05) by Rg5 compared with control rats. These data demonstrated that STZ-induced learningand memory impairments in rats could be improved by Rg5, which was associated with attenuatingneuroinflammatory responses. Our findings suggested that Rg5 could be a beneficial agent for the treatmentof AD.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Alzheimer's disease (AD) has been recognized as a senile neurode-generative disease affecting the life quality of patients. It has beencharacterized as progressive memory capacity damage and cognitivedysfunction in clinic. Pathological evidences support the fact thata large number of senile plaques (SP) deposit in hippocampus andcerebral cortical neurons of AD patients. A great many evidences
shan Road, Nanjing, PR China.
Road, Hefei, Anhui, PR China.
g road, Wenzhou, PR China.
[email protected] (M. Liu),
e work.
ghts reserved.
have been found to confirm that the accumulation of intracellularβ-amyloid (Aβ) may be an early event in the development of AD[1,2]. Studies have showed that the impaired memory function mayalso be associated with the cholinergic system synthase such as(choline acetyltransferase, ChAT) and hydrolytic enzyme such as(acetylcholinesterase, AChE). Furthermore, the imbalanced level ofinsulin-like growth factors 1 (IGF-1) and/or brain derived neurophicfactor (BDNF) contributes to the onset and development of cognitivedysfunction. Streptozotocin (STZ), which could reduce spatial learn-ing and memory, was generally used to induce AD rats [2]. The Aβdeposits were aggregated by the induction of STZ in the hippocam-pus and cerebral cortical neurons of AD rats. In addition, multipleenzymes or proteins including AChE, ChAT, IGF-1 and BDNF canalso be modulated pathologically by STZ.
Neuroinflammatory responses occurring in the central nervous sys-tem (CNS) are closely related to the pathways leading to neuronal celldeath in AD [3]. The neuroinflammatory switch occurs before the

318 S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
detectable deposits of Aβ and accelerates the clearance disorders of Aβ.It is widely accepted that inflammatory activation of glial cell is an im-portant mediator of pathological phenomena resulting in cognitive im-pairment and neuronal degeneration. Notably, the deposition of Aβ caninitiate a vicious cycle of Aβ generation between astrocytes andneuronswhich then leads to a chronic, sustained and progressive neuroinflam-mation [4]. Inflammatory mediators, such as interleukin (IL)-1β,tumor necrosis factor (TNF)-α, cyclooxygenase-2 (COX-2) and induciblenitric oxide synthase (iNOS) have been found to be the key cytokinesin these inflammatory responses and responsible for neuroinflamma-tion [5].
Ginseng, the root of Panax ginseng (P. ginseng, Araliaceae), recordedin Compendium of Materia Medica, is one of the well-known Chineseherbs. Ginsenosides, the main active components of ginseng, havebeen found to have anti-inflammatory and anti-oxidative effects. Theneuroprotective effects of ginsenosides were also reported in ADmouse model [6]. Ginsenoside Rg5 (Rg5) (Fig. 1) could be used as anovel therapeutic agent for treating memory loss [7]. However, theeffects of Rg5 on improving cognitive dysfunction and Aβ depositin vivo and underlying mechanism were not fully understood.
Therefore, this present study aims to evaluate effects of Rg5 onstreptozotocin (STZ)-induced memory impaired rats, and explore itsmechanism.
2. Material and methods
2.1. Reagents and chemicals
Streptozotocin was obtained from MP Biomedicals (Shanghai,Chinese). Donepezil Hydrochloride Tablets (Ebixa) were purchasedfrom H. Lundbeck A/S (Tianjin, China). Ginsenoside Rg5 (purity N 98%)was ordered from Nantong Feiyu biological technology Co., LTD(Nantong, China). Chloral hydrate was ordered from Sinopharm Chemi-cal Reagent Co., Ltd (Beijing, China). AChE and ChAT kits were purchasedfromNanjing Jiancheng Bioengineering Institute (Nanjing, China). TNF-αand IL-1β ELISA kits were purchased from MAIXIN-BIO (Fuzhou, China).Monoclonal rabbit anti-COX-2 antibody and polyclonal mouse anti-iNOS antibody, BNDF, IGF-1 and Aβ antibodies were purchased fromSanta Cruz Biotechnology (CA, USA). ECL chemiluminescence kit wasobtained from Pierce Biotechnology, Inc. (Appleton, WI. USA). Otherchemicals were purchased from commercial sources.
2.2. Animals and drug administration
Wistar rats (250 ± 20 g) were obtained from Shanghai SLAC LabAnimal Center (Shanghai, China). Rats were kept at a 12 h light/dark
Fig. 1.The chemical structure ofGinsenoside Rg5 (chemical formula: C42H70O12,molecularweight = 767.01).
cycle (room temperature 25 ± 2 °C and humidity 60–70%) for threedays before experiments with free access to water and diet. Animalexperiments were approved by the Animal Ethics Committee of JiangsuProvincial Academy of Chinese Medicine. Rats were divided into sixgroups (12 rats for each), including 0.9%NaCl saline (NS)-treated group(control group), STZ-induced experimental groups, STZ + Rg5-treatedgroups (5, 10 or 20 mg/kg) [8] and positive control donepezil (3 mg/kg)group [9]. STZ (3 mg/kg) was dissolved in artificial CSF (aCSF) and thesolution of 10 μl was intracerebroventricularly (i.c.v.) injected to eachlateral cerebral ventricle to induce AD rats according to the coordinates,that is, 0.8mmposterior to bregma, 1.5mm lateral to sagittal suture and3.6 mm ventral by Hamilton microsyringe [9,10]. The injections wereperformed at the first day and the third day. Each injection time wasmaintained at 10 min. In blank control group, the same volume ofaCSF including 147 mM NaCl, 2.9 mM KCl, 1.6 mM MgCl2, 1.7 mMCaCl2 and 2.2 mMdextrose was injected on days 1 and 3. These postop-erative ratswere injected 40 million units of penicillin every day. Differ-ent doses of Rg5, donepezil, or physiological saline were orallyadministered from the 2nd day after STZ injection [11].
2.3. Behavioral testing
2.3.1. Step through testThe step through test was performed on 28 days after the adminis-
tration of STZ in a multifunction passive avoidance apparatus [12]. Theapparatus consisted of two compartments including illuminated oneand dark one. The two compartments were spaced by a black wallwith a hole in the lower middle part. The test was conducted consecu-tively for 2 days, namely, day 1 was used for training while day 2 fortest. Each rat was placed in the illuminated compartmentwhich deviat-ed from the dark compartment. After adaptation trial for 1 h, these ratsreceived a training trial. Besides the same above procedure as adapta-tion trial, the rats received the electric foot shock (about 30 V) fromthe stainless steel grid floor when they entered the dark compartment.The testing trial on day 2was the same as the procedure of training trialon day 1. The retention time, namely, from the placement in the illumi-nated compartment to the dark one,was recorded asmemory retention.The rats which did not enter the darkroom within 120 s were consid-ered to have good memory retention.
2.3.2. Morris water maze testMorris water maze, equipped with around the pool and an automatic
recording system, was used for the evaluation of Rg5 on learning andmemory ability of AD rats [13]. The diameter of morris water mazewas 160 cm while the depth was 50 cm. Camera lens was placed inthe center of a black plexiglas rectangular tank above 2 m height. Themidpoint of the quadrant pool wall was set as water point and dividedinto four quadrants. Themazewas filled with water to a depth of 25 cmand the temperature was kept at 25± 1 °C. A black platform (diameter12 cm, high 30 cm) was fixed in the fourth quadrant and placed 1 cmbelow the water surface. After administration for 28 days, the test wasperformed for six consecutive days (7:30 am to 16:00 pm). In the firstday, rats swam in freestyle 2 times (1 min/time) to adapt to the waterenvironment. From the second day, the rats were trained two times at9:00 and 16:00 every day. The latency time of each rat from the startingpoint to find hidden underwater platform and climb up the platformwas recorded within 90 s. If rats did not find the platform within 90 s,the latency was recorded as 90 s. On the sixth day, the platform wasremoved to conduct space exploration experiment, and all the ratswere placed into the water point optionally in the same way. The timedistance to find the platform location, each quadrant swimming dis-tance and the number of times crossing the platform location of ratswere recorded within 90 s. The data was recorded with the MiniSeecomputerized tracking system.

Table 1Effect of Rg5 on the latency time in Passive avoidance test in AD rats (n = 12, M(QR)).
Group n Latency (s)
Control group (NS) 12 109. 5 (106.25–123.5)STZ 12 25.5 (24.00–27.75)⁎⁎
STZ + Rg5 (5 mg/kg) 12 46.6 (44.00–48.50)STZ + Rg5 (10 mg/kg) 12 65.5 (62,25–69.50)#
STZ + Rg5 (20 mg/kg) 12 89.5 (87.00–92.50)#
STZ + Donepezil 12 103 (102–104.75)##
A nonparametric test (Kruskal Wallis H) was used in this experiment. Latencies wereexpressed as median/interquartile range.⁎P b 0.05, ⁎⁎P b 0.01 vs. control group; #P b 0.05,##P b 0.01 vs. STZ model.
319S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
2.4. Determination of ChAT and AChE activities
Six rats from each group were sacrificed by decapitation after thewater maze test [11,14]. The cerebrums were dissected quickly. Tenpercent cerebral homogenate (1:10, w/v) in cold salinewas centrifugedwith 3500 ×g for 10min at 4 °C. Supernatant was taken to measure theactivities of AChE and ChAT. The optical density (OD) value of AChEwasdetermined at 412 nm a microplate reader while ChAT at 324 nm. TheAChE activity was expressed as U/mg protein while the ChAT activityas U/g protein.
2.5. ELISA for TNF-α and IL-1β levels
The levels of inflammatory cytokines TNF-α and IL-1β in cerebralhomogenate were measured using ELISA kits according to manu-facturer's protocols [15]. The absorbance of TNF-α and IL-1β was mea-sured in a microplate reader at 450 nm. The contents of TNF-α, IL-1βwere expressed as ng/ml.
2.6. Congo red staining
Congo red staining was preformed to check Aβ deposits in the hip-pocampus and cerebral cortex of AD rats [16,11]. The remaining 6 ratsfrom each group were sacrificed by decapitation. The cerebra was dis-sected and kept at −70 °C for immunohistochemistry. The cerebratissues were cut into 5 μmsections. Sequentially, sectionswere incubat-ed with hematoxylin solution for 10 min. After being washed with PBSfor three times, sections were incubated by congo red solution (SigmaAccustain amyloid staining kit, St. Louis, MO) for 20min. An ethanol se-ries was used for the dehydration. Sections were cleared with xylenebeforemountingwith neutral gumunder a coverslip, and photographedwith Leica image analysis system.
2.7. Immunohistochemistry for BDNF and IGF-1 expressions [11]
Immunohistochemistry was performed according to the previouslydescribed method. In brief, sections were blocked with 10% horseserumat 37 °C for 30min and then incubatedwith the primary antibod-ies of BDNF (1:200) and IGF-1(1:200) overnight at 4 °C. After beingwashedwith PBS for three times, sections were incubated with second-ary antibody at 37 °C for 1 h. At the end of incubation, sections werestained by Avidin Biotin Complex for another 30 min and developedby 3,3-diaminobenzidine (DAB) chromogen for 15 min in the dark. Allsections were visualized and photographed under a light microscope(Olympus IX71, Japan). The images were analyzed with Image-ProPlus software.
2.8. Western blotting analysis for the expressions of COX-2, iNOS, BNDF,IGF-1 and Aβ [17]
The brain tissue was homogenized in lysis buffer (pH = 7.5) con-taining 20 mM Tris · Cl, 100 mM NaCl, 5 mM MgCl2, 1 mM EDTA, 1%TritonX-100, 1 mM DTT and 1 mM PMSF. After being centrifuged with13,000 g at 4 °C for 15 min, the supernatant was collected. Equalamounts of supernatant protein (50 μg)were denatured in loading buff-er and heated at 100 °C for 5 min and then subjected to electrophoresisconstituent of 10% SDS-PAGE at 40 mA. Sequentially, these proteinswere transferred onto a PVDF membrane and then blocked in 5% BSAblocking buffer for more than 1 h. Membranes were incubated withprimary COX-2 (1:200), iNOS(1:200), BNDF(1:200), IGF-1(1:200) andAβ (1:200) anti-bodies overnight. After beingwashedwith PBS contain-ing 0.05% Tween-20 (PBST) for 3 times, membranes were incubatedwith the horseradish peroxidase-bound secondary antibody (1:1000)for 2 h at room temperature. Finally, the visualization of proteins wasshown by adding ECL chemoluminescence reagents (0.1 ml/cm2).
Image pro plus (IPP) software was used for the quantification ofproteins.
2.9. Statistical analysis
All data were expressed as means ± standard deviation (SD),except data of Step through test described as median/interquartilerange. In Step through test, Kruskal–Wallis H test was chosen to do non-parametric test of multiple independent samples because we havefound that data did not meet the normal distribution. In Morriswater maze test, days and treatment factors needed to consider sothat 2-way ANOVA was used. Finally, one-way ANOVA was selected toanalyze other data in our experiment. Statistical analysis was performedby analysis of variance (ANOVA) or Nonparametric Test using SPSSversion 16.0 software. Differences with a P value less than 0.05 wereconsidered statistically significant in all the cases.
3. Results
3.1. Step through test
After i.c.v. injection with STZ for 28 days, the step through test wasperformed to evaluate the improvement on long-term memory in ADrats which were caused by Rg5. As shown in Table 1, a significantshorter memory retention was observed in STZ-induced experimentalrats than those of control one [F(12,77) = 3.44, P b 0.05]. In order toevaluate the effect on the cognitive deficits of AD rats, we examinedthe improvement of Rg5 on STZ-induced ratsmediated. The oral admin-istration of Rg5 (5, 10, 20 mg/kg) enhanced significantly the memoryretention as compared with STZ group [F(12,77) = 5.61, P b 0.05].These results indicated that Rg5 could improve the learning and mem-ory ability of STZ-induced AD rats.
3.2. Morris water maze test
Morris watermaze test was conducted to evaluate the spatial recog-nition memory of AD rats. Multivariate analysis of variance was used toanalyze the data in this test due to two factors like time and treatment.As shown in Fig. 2B, there was no significant difference among differentgroups on the mean latency and on the path length at first four days.From the beginning of the fifth day, the mean latency gradually short-ened compared with the first day in the control group and donepezilpositive control group, the escape latency in the fifth and sixth dayswere significantly shorter [F(12,77) = 10.46, P b 0.05]. The escapelatency low-dose Rg5 group (5 mg/kg) was not very obvious comparedwith the first day, only when the sixth day time was significant short-ened [F(12,77) = 5.85, P b 0.05], while the high-dose groups (10 and20 mg/kg) were more effective. However, the induction of STZprolonged significantly the mean latency on days 2 and 3 while length-ened the path length on days 3 and 4, compared with the NS controlgroup [F(12,77) = 8.27, P b 0.05]. On the fifth and sixth day, a
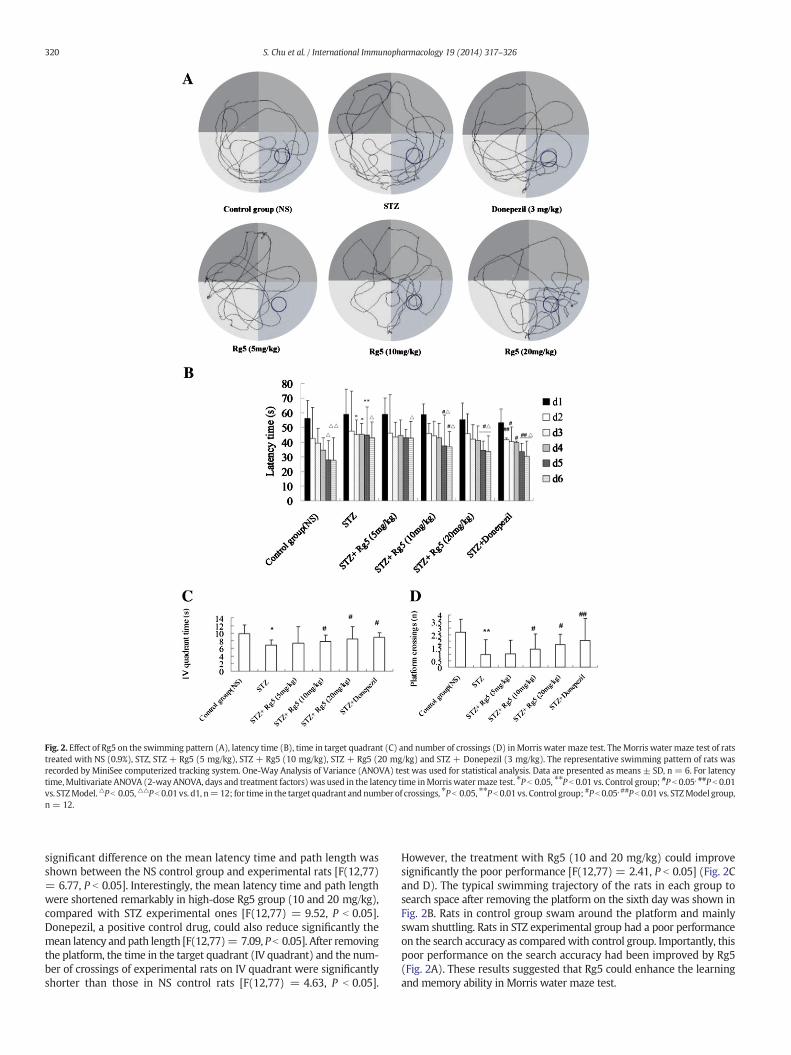
Fig. 2. Effect of Rg5 on the swimming pattern (A), latency time (B), time in target quadrant (C) and number of crossings (D) inMorris water maze test. The Morris watermaze test of ratstreated with NS (0.9%), STZ, STZ + Rg5 (5 mg/kg), STZ + Rg5 (10 mg/kg), STZ + Rg5 (20 mg/kg) and STZ + Donepezil (3 mg/kg). The representative swimming pattern of rats wasrecorded by MiniSee computerized tracking system. One-Way Analysis of Variance (ANOVA) test was used for statistical analysis. Data are presented as means ± SD, n = 6. For latencytime,Multivariate ANOVA (2-way ANOVA, days and treatment factors) was used in the latency time inMorriswatermaze test. ⁎P b 0.05, ⁎⁎P b 0.01 vs. Control group; #P b 0.05, ##P b 0.01vs. STZModel.△P b 0.05,△△P b 0.01 vs. d1, n=12; for time in the target quadrant and number of crossings, ⁎P b 0.05, ⁎⁎P b 0.01 vs. Control group; #P b 0.05, ##P b 0.01 vs. STZModel group,n = 12.
320 S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
significant difference on the mean latency time and path length wasshown between the NS control group and experimental rats [F(12,77)= 6.77, P b 0.05]. Interestingly, the mean latency time and path lengthwere shortened remarkably in high-dose Rg5 group (10 and 20 mg/kg),compared with STZ experimental ones [F(12,77) = 9.52, P b 0.05].Donepezil, a positive control drug, could also reduce significantly themean latency and path length [F(12,77)= 7.09, P b 0.05]. After removingthe platform, the time in the target quadrant (IV quadrant) and the num-ber of crossings of experimental rats on IV quadrant were significantlyshorter than those in NS control rats [F(12,77) = 4.63, P b 0.05].
However, the treatment with Rg5 (10 and 20 mg/kg) could improvesignificantly the poor performance [F(12,77) = 2.41, P b 0.05] (Fig. 2Cand D). The typical swimming trajectory of the rats in each group tosearch space after removing the platform on the sixth day was shown inFig. 2B. Rats in control group swam around the platform and mainlyswam shuttling. Rats in STZ experimental group had a poor performanceon the search accuracy as comparedwith control group. Importantly, thispoor performance on the search accuracy had been improved by Rg5(Fig. 2A). These results suggested that Rg5 could enhance the learningand memory ability in Morris water maze test.

Fig. 3. Effect of Rg5 on the activity of AChE (A) and ChAT (B), the levels of TNF-α and IL-1β(C) in STZ-induced AD rats. Rats were induced with STZ and then given different concen-trations of Rg5 (5, 10 and20 mg/kg) or positive drugdonepezil (3 mg/kg). One-WayAnal-ysis of Variance (ANOVA) test was used for statistical analysis. Data are presented asmeans ± SD, n = 6. ⁎⁎ P b 0.01, STZ vs. NS control group; #P b 0.05, ##P b 0.01, vs. STZ;$P b 0.05, vs. Donepezil.
321S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
3.3. Effect of Rg5 on the changes in the activities of ChAT and AChE
The activities of ChAT and AChE were measured after the learningand memory tests. The levels of AChE in the STZ group were significanthigher than those inNS control ratswhereas the levels of ChAT in exper-imental rats were lower than NS control rats [F(6,35) = 8.48, P b 0.05].However, the administration of Rg5 (5, 10 and 20 mg/kg) could decreaseremarkably STZ-induced AChE activity to 0.632± 0.012, 0.519± 0.022and 0.452 ± 0.013 U/mg [F(6,35) = 10.77, P b 0.05] (Fig. 3A and B).Moreover, Rg5 (5, 10 and 20 mg/kg) could also increase significantly
the STZ-induced ChAT activity to 71.13 ± 6.14, 88.42 ± 7.57 and98.24 ± 5.18 U/g. Our data indicated that Rg5 could improve cognitivedeficits through reducing AChE activity and increasing ChAT activity incerebral cortex and hippocampus.
3.4. Increase of Rg5 on BDNF and IGF-1 expressions in the hippocampus andcerebral cortex
In order to evaluate Rg5's effect on neurophic factors of AD rats,BDNF and IGF-1 expressions were observed in the hippocampus andcerebral cortex of AD rats by immunohistochemistry (Fig. 5). The no-ticeable immunoreactivities of BDNF and IGF-1 were visualized in thehippocampus and cerebral cortex of blank control rats (Fig. 5A and B).The percentages of positive cells in experimental rats were lower thanthat of NS control group [F(6,35) = 11.83, P b 0.05]. The reduction ofBDNF and IGF-1 deposits were prevented by the administration ofRg5 (5, 10 and 20 mg/kg) or donepezil (3 mg/kg) [F(6,35) = 8.04,P b 0.05]. Furthermore, these up-regulations on BDNF and IGF-1deposits in the hippocampus and cerebral cortex were in a dose-dependent manner (Fig. 5C and D). The results of Western blot anal-ysis also showed that Rg5 (10 and 20 mg/kg) or donepezil (3 mg/kg)could up-regulate the expressions of neurophic factors BDNF andIGF-1 in brains (Fig. 7A and B). The above findings indicated thatRg5 had a neuroprotective effect on cognitive deficits in AD rats.This neuroprotective effect was associated with the modulation ofBDNF and IGF-1 deposits.
3.5. Amelioration of Rg5 on Aβ deposition in the hippocampus and cerebralcortex
The pattern of the chronic inflammatory response is strongly associ-ated with fibrillar Aβ deposits in AD. The fibrillar Aβ-induced chronicinflammatory responses contribute to the brain disorders. To detect theAβ accumulation in the hippocampus and cerebral cortex of AD rats,congo red stainingwas preformed and presented in Fig. 4A. The Aβ accu-mulation in hippocampus and cerebral cortexwasmarkedly enhanced inAD rats compared with the NS control group [F(6,35) = 9.25, P b 0.05](Fig. 4B). However, the treatment with Rg5 (5, 10 and 20 mg/kg) ordonepezil (3 mg/kg) could attenuate significantly the accumulation ofAβ in the hippocampus and cerebral cortex of AD rats [F(6,35) = 6.92,P b 0.05]. In order to confirm the above results, western blot analysiswas also performed to evaluate the regulation of Rag on Aβ deposits.As depicted in Fig. 7A and B, a significant decrease was observed in Rg5(10 and 20 mg/kg) or donepezil (3 mg/kg) groups, compared withSTZ-inducedmodel rats [F(6,35)= 9.24, P b 0.05]. These results indicat-ed that Rg5 ameliorated the inflammatory responses in AD rats viareduction of Aβ deposits. These results showed that Rg5 alleviateddose-dependently STZ-induced inflammatory responses in the hippo-campus and cerebral cortex and this was related to the amelioration ofAβ deposits in AD rats [F(6,35) = 8.11, P b 0.05].
3.6. Effect of Rg5 on the changes in the levels of TNF-α and IL-1β
TNF-α and IL-1β have been found to play an important role inneuroinflammatory responses and cause the death of nerve cells. Toevaluate Rg5's attenuation on the levels of pro-inflammatory cytokines,ELISA was used to measure the levels of IL-1β and TNF-α in cerebralhomogenate. As shown in Fig. 3C, the levels of TNF- α and IL-1β wereincreased significantly by the treatment of STZ. Interestingly, theoral administration of ginsenoside Rg5 could significantly inhibit thelevels of pro-inflammatory cytokines IL-1β and TNF-α[F(6,35) = 9.06,P b 0.05]. These results indicated that Rg5's alleviation on cognitive def-icits was associated with its attenuation on neuroinflammatoryresponses.

Fig. 4. Effect of Rg5 on Aβ deposition in the hippocampus and cerebral cortex in AD rats by Congo red staining. (A) The representative Aβ deposition expression in the hippocampus andcerebral cortex; (B) the activity of Aβ deposition expression in the hippocampus; (C) the activity of Aβ deposition expression in the cerebral cortex. Donepezil was chosen for the positivecontrol. Rats treatedwith NS, STZ, STZ+Rg5 (5 mg/kg), STZ+Rg5 (10 mg/kg), STZ+Rg5 (20 mg/kg) and STZ+ donepezil (3 mg/kg). One-WayAnalysis of Variance (ANOVA) test wasused for statistical analysis. Data are presented as means ± SD, n = 6. ⁎⁎ P b 0.01, STZ vs. NS control group; #P b 0.05, ##P b 0.01, vs. STZ; $P b 0.05, vs. Donepezil.
Fig. 5. Effect of Rg5 on the BDNF and IGF-1 expressions in the hippocampus and cerebral cortex of AD rats. (A) the representative BDNF expression in the hippocampus and cerebral cortexby immunohistochemistry; (B) the representative IGF-1 expression in the hippocampus and cerebral cortex by immunohistochemistry; (C) the relative expression ratio of BDNF (%);(D) the relative expression ratio of IGF-1(%). Donepezil (3 mg/kg) was chosen for the positive control. Data are presented as means ± SD, n = 6. One-Way Analysis of Variance(ANOVA) test was used for statistical analysis. ⁎⁎P b 0.01, STZ vs. NS control group; #P b 0.05, ##P b 0.01, vs. STZ; $P b 0.05, vs. Donepezil.
322 S. Chu et al. / International Immunopharmacology 19 (2014) 317–326

Fig. 6. Effect of Rg5 on the protein contents of COX-2 and iNOS in different groups. (A) the representative expression of COX-2 and iNOS in different groups; (B) the changes of COX-2 andiNOS protein contents. Rats treated with NS, STZ, STZ + Rg5 (5 mg/kg), STZ + Rg5 (10 mg/kg), STZ + Rg5 (20 mg/kg) and STZ + donepezil (3 mg/kg). One-Way Analysis of Variance(ANOVA) test was used for statistical analysis. Data are presented as means ± SD, n = 6. ⁎⁎ P b 0.01, STZ vs. NS control group; #P b 0.05, ##P b 0.01, vs. STZ; $P b 0.05, vs. Donepezil.
323S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
3.7. Effects of Rg5 on levels of COX-2 and iNOS in AD rats by westernblotting
COX-2 and iNOS, two important inflammatory cytokines [51], havebeen found to contribute to the neuroinflammatory responses in
Fig. 7. Effect of Rg5 on the protein expressions of BNDF, IGF-1 and Aβ by western blot analysisdensity. The homogenates of brains of rats treatedwith STZ, STZ+Rg5 (10 mg/kg), STZ+Rg5 (ysis of Variance (ANOVA) test was used for statistical analysis. Data are presented as means ±
the pathogenesis of AD. The western blotting analysis showed thatthe expressions of COX-2 and iNOS could not be detected in controlgroup (Fig. 6A and B). However, the inductive effect of STZ couldremarkably up-regulate these expressions in experimental group[F(6,35) = 5.39, P b 0.05]. More importantly, the oral administration
(A). The relative expressions of these brands to β-actin were calculated according to the20 mg/kg) and STZ+ donepezil (3 mg/kg)were used for this experiment. One-Way Anal-SD, n = 6. ⁎P b 0.05, ⁎⁎ P b 0.01, vs. STZ group; $P b 0.05, vs. Donepezil.

324 S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
of Rg5 could down-regulate markedly STZ-induced protein expressionsin a dose-dependent manner [F(6,35) = 4.01, P b 0.05] (Fig. 6A and B).The above findings also indicated that the attenuation of Rg5 onneuroinflammatory responses was associated with its alleviation oncognitive deficits.
4. Discussion
Alzheimer's disease (AD), a multifactorial neurodegenerative dis-ease, has been characterized by progressive learning and memory im-pairments and neuro-pathological features including Aβ plaque. Thisprogressive lesion has been confirmed to be associated with the imbal-ance of the generation and metabolism of Aβ and neuroinflammatoryresponses [18]. Clinical and experimental studies have confirmed thatneuroinflammatory responses might contribute to Aβ deposition inbrain tissue and lead to cognitive dysfunction in AD [18]. In this study,we found that Rg5 could improve cognitive dysfunction, attenuate Aβdeposition, increase BDNF and IGF-1 expression levels, as well as ame-liorate neuroinflammatory responses in STZ-induced AD rats. Thisstudy indicated that Rg5 had a beneficial effect on STZ-induced cogni-tive dysfunction and this efficacy was derived from its attenuation onneuroinflammatory responses, at least in part.
The inductive effect of STZ can delay persistent Aβ metabolism andaccelerate the formation of inflammatory microenvironment in AD[11]. For STZ-induced AD model rat, studies showed that intraventricu-lar injection of STZ can cause AD-like neurodegenerative changes innormal rats, such as mitochondrial dysfunction, neuronal death, learn-ing and memory and cognitive dysfunction [19]. The mechanism maybe associated with the weakness or destruction of the insulin signaltransduction, phosphorylation and destruction of its intrinsic tyro-sine kinase activity of the insulin receptor, and increase of the phos-photyrosine phosphatase activity. However, another AD animal model,such as Scopolamine-induced mice, can not completely simulate thevarious features of AD. Scopolamine is an anticholinergic drug whichcan block the binding sites of the acetylcholine receptors in cerebralcortex and then causes the cholinergic system dysfunction. Studiesrevealed that cholinergic dysfunction induced by scopolamine can bereversed. However, AD is a chronic, progressive and irreversible neuro-logical disease. In this study, the treatment of STZ could result in thepoor performance in the step through test and Morris water mazetest. STZ also increased remarkably the secretion and expression ofinflammatory cytokines leading to poor cognitive dysfunction.
P. ginseng has been used for the clinical treatment of various dis-eases, such as AD. Particularly, its active compounds have been usedfor a variety of diseases such as cancer, inflammation, stress and agingin Asian countries. In addition, they have also been found to be benefi-cial for the alleviation of AD [19,20]. Ginsenoside Rg5, one of thesecompounds, has been confirmed to protect scopolamine-inducedmem-ory deficit inmice [19,21]. In the present study, the alleviation on cogni-tive dysfunction and attenuation on neuroinflammatory responses inSTZ-induced AD ratsmediated by Rg5were examined. Our data showedthat Rg5 (5, 10 and 20 mg/kg)might enhance the learning andmemoryability, increase ChAT activity and decrease AChE activity, regulateinflammatory cytokines TNF-α and IL-1β levels, attenuate Aβ deposi-tion, as well as increase BDNF and IGF-1 expressions in the hippocam-pus and cerebral cortex of STZ-induced AD rats. Furthermore, theprotein expressions of COX-2 and iNOS were decreased obviouslycompared with control group. Our findings had showed for the firsttime that Rg5 could ameliorate STZ-induced cognitive dysfunctionin vivo. Importantly, this alleviation might be associated with the ame-lioration on neuroinflammatory responses.
Aβ is a peptide of 36–43 amino acids that is formed by a large trans-membrane glycoprotein expressed on the cell, such as amyloid precur-sor protein (APP). Aβ may activate inflammatory and neurotoxicprocess, including the excessive generation of free radical and oxidativedamage among intracellular proteins and other macromolecules
[18,22]. Evidence had supported the fact that there was a close correla-tion between Aβ deposit and the neurodegenerative process of AD[23,24]. In our experiment, i.c.v. administration of STZ significantlyinduced Aβ deposit in the hippocampus and cerebral cortex and thencaused impairments in memory function and cognitive ability in rats.A single acute i.c.v. administration of STZ inducedmarkedly the amnesiceffects in rats, evidenced as deficiencies in step through test and watermaze test (Table 1). However, the administration of Rg5 showed bene-ficial effect on impaired memory in our present study. Rg5 couldimprove cognitive function, namely, prolonging significantly memoryretention in the step through test and shortening the latency time inthe water maze performance as compared with that of the STZ rats(Table 1 and Fig. 2B). In addition, Congo red staining results showedthat the diminution of Aβ deposit was obvious in Rg5-treated rats ascompared with experimental ones (Fig. 4).
Accumulating studies suggest that neural systems have beeninvolved in learning and influencingmemory function. Themain neuro-transmitter acetylcholine (AChE) in central cholinergic pathway hasbeen shown as themain path of learning andmemory. AChEplays a piv-otal role in cognitive activities such as spontaneous movement andexploratory behavior in the memory and attention of learning, andworking. Because there is a serious degenerative change in brain cholin-ergic system of AD patients, the content of AChE can be decreased upto 70%–80%. Studies confirmed that the most effective treatment wasto improve the patient's cognitive and memory disorders by enhancingcholinergic function [25,28]. AChE was so unstable and prone tohydrolization. The imbalance of cholinergic system synthase (cholineacetyltransferase, ChAT) and hydrolytic enzyme like AChE may resultin neuroinflammatory responses of AD [29,30]. Many studies indica-ted that the lesions of central nervous system in AD were caused bythe degeneration of cholinergic nervous system [31]. Previous experi-mental study showed that the enhanced hippocampal AChE contentand ChAT activity could alleviate the learning and memory disorder inAD rats [32].
The inflammation leading to microglial activation and neuronaldamage in AD is mediated by the production of pro-inflammatorymolecules [33]. The activated microglia produce pro-inflammatorycytokines such as IL-1, IL-6 and TNF-α [34]. Generally, TNF-α elicitsthe neuroprotection against cerebral ischemia [35]. Our results demon-strated that Rg5 could alleviate cognitive impairment and Aβ deposit inthe hippocampus and cerebral cortex of STZ-induced AD rats throughthe increase of ChAT activity and the decrease of AChE activity. Mean-while, the expressions of TNF-α and IL-1βwere increased by STZ. Nota-bly, these expressions were significantly inhibited by the treatment ofginsenoside Rg5.
Aβ deposit can inhibit the expressions of neurotrophic factors,such as brain derived neurotrophic factor (BDNF) and insulin-likegrowth factor-1 (IGF-1) [36,37]. BDNF has been found to be a regulatorin the synaptic plasticity contributing to the development of memoryand cognitive function [38,39]. It plays a role in several events constitut-ing the pathological cascade in AD. The reduction of neuroprotectivefactor IGF-1 level in brain has been considered as an important factorin the development of cognitive impairment and Aβ deposit [40]. In re-cent years, many studies have shown that the lack of BDNF and IGF-1induces and accelerates the accumulation of soluble Aβ oligomers[41]. In our experiment, the data showed that expressions of BDNFand IGF-1 in the hippocampus and cerebral cortex of AD rats weredecreased significantly by the lateral i.c.v. injection of STZ (Fig. 4A, B,C and D). Immunohistochemistry analysis demonstrated that Rg5 up-regulated remarkably the expressions of BDNF and IGF-1 in the brainof AD rats.
Both cyclooxygenase-2 (COX-2) and nitric oxide synthase (iNOS)are inducible enzymes. COX-2 plays an active role in the exaggerated in-flammation while iNOS takes an important part in oxidative stress [42].COX-2 and iNOSmediated the similar pathological processes [43]. Theyinteract with each other to aggravate their damaging effects [44,45].

325S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
As a key enzyme in the synthesis of prostaglandins (PGs), COX-2 is oneof the important mediators of inflammation [46]. The stimulation ofCOX-2 can lead to the dramatic and transient synthesis of PGE2, whichhas pathophysiological effects [47]. The expression of iNOS wasincreased after stimulation, and it plays a key role in oxidativestress [48]. In the present study, we further provided evidence thatRg5 suppressed the activation of COX-2 and iNOS. Therefore, the inhibi-tion of cytokines and chemokines can provide a novel and rational ther-apeutic approach to the inflammation-induced progression of AD[49,50].
5. Conclusion
In conclusion, the present study that Rg5 could alleviate cognitivedysfunction in STZ-induced AD rats through regulating cholinergic sys-tem, attenuating Aβ deposition, as well as increasing of neurotrophicfactors' (BDNF, IGF-1) expressions. Importantly, Rg5 also showed goodamelioration on STZ-induced neuroinflammatory responses. These re-sults that Rg5's alleviation on STZ-induced cognitive dysfunction wasassociated with its function on attenuating neuroinflammatory re-sponses. It might provide insights into Rg5's beneficial effect on thetreatment of AD.
Acknowledgments
We sincerely thank financial support for this work from Visit-ing scholar at Teachers College Professional Development Project(2012) (FX2012026) and the National Natural Science of China(81202906).
References
[1] Li W, Yu J, Liu Y, Huang X, Abumaria N, Zhu Y, et al. Elevation of brain magnesiumprevents and reverses cognitive deficits and synaptic loss in Alzheimer's diseasemouse model. J Neurosci 2013;33:8423–41.
[2] Hampel H. Amyloid-β and cognition in aging and Alzheimer's disease: molecularand neurophysiological mechanisms. J Alzheimers Dis 2013;33:S79–86.
[3] Salkovic-Petrisic M, Knezovic A, Hoyer S, Riederer P. What have we learned fromthe streptozotocin-induced animal model of sporadic Alzheimer's disease, aboutthe therapeutic strategies in Alzheimer's research. J Neural Transm 2013;120:233–52.
[4] Joo SS, Yoo YM, Ahn BW, Nam SY, Kim YB, Hwang KW, et al. Prevention ofinflammation-mediated neurotoxicity by Rg3 and its role in microglial activation.Biol Pharm Bull 2008;31:1392–6.
[5] Li C, Zhao R, Gao K, Wei Z, Yin MY, Lau LT, et al. Astrocytes: implications forneuroinflammatory pathogenesis of Alzheimer's disease. Curr Alzheimer Res2011;8:67–80.
[6] Blanco M, Rodríguez-Yáñez M, Sobrino T, Leira R, Castillo J. Platelets, inflammation,and atherothrombotic neurovascular disease: the role of endothelial dysfunction.Cerebrovasc Dis 2005;20:32–9.
[7] Fang F, Chen X, Huang T, Lue LF, Luddy JS, Yan SS. Multi-faced neuroprotective ef-fects of Ginsenoside Rg1 in an Alzheimer mouse model. Biochim Biophys Acta1822;2012:286–92.
[8] Kim EJ, Jung IH, Van Le TK, Jeong JJ, Kim NJ, Kim DH. Ginsenosides Rg5 and Rh3 pro-tect scopolamine-induced memory deficits in mice. J Ethnopharmacol 2013;146:294–9.
[9] Gibbs RB, Chipman AM, Nelson D. Donepezil plus estradiol treatment enhanceslearning and delay-dependent memory performance by young ovariectomizedrats with partial loss of septal cholinergic neurons. Horm Behav 2011;59:503–11.
[10] Tota S, Kamat PK, Saxena G, Hanif K, Najmi AK, Nath C. Central angiotensinconverting enzyme facilitates memory impairment in intracerebroventricularstreptozotocin treated rats. Behav Brain Res 2012;226:317–30.
[11] Liu JP, Feng L, Zhang MH, Ma DY, Wang SY, Gu J, et al. Neuroprotective effect ofLiuwei Dihuang decoction on cognition deficits of diabetic encephalopathy instreptozotocin-induced diabetic rat. J Ethnopharmacol 2013;150:371–81.
[12] Fu AL, Dong ZH, Sun MJ. Protective effect of N-acetyl-L-cysteine on amyloid beta-peptide-induced learning andmemory deficits in mice. Brain Res 2006;1109:201–6.
[13] Choi JY, Cho EJ, Lee HS, Lee JM, Yoon YH, Lee S. Tartary buckwheat improves cogni-tion and memory function in an in vivo amyloid-β-induced Alzheimer model. FoodChem Toxicol 2013;53:105–11.
[14] Liu J, Feng L, Ma D, Zhang M, Gu J, Wang S, Fu Q, Song Y, Lan Z, Qu R, Ma S. Neuro-protective effect of paeonol on cognition deficits of diabetic encephalopathy instreptozotocin-induced diabetic rat. Neurosci Lett 2013(549):63–8.
[15] Basta-Kaim A, Szczęsny E, Leśkiewicz M, Gombik K, Slusarczyk J, BudziszewskaB, et al. Maternal immune activation leads to age-related behavioral and
immunological changes in male rat offspring — the effect of antipsychoticdrugs. Pharmacol Rep 2012;64:1400–10.
[16] Wang H, He J, Zhang R, Zhu S, Wang J, Kong L, et al. Sensorimotor gating and mem-ory deficits in an APP/PS1 double transgenic mouse model of Alzheimer's disease.Behav Brain Res 2012;233:237–43.
[17] Lee DH, Kim NR, Lim BS, Lee YK, Yang HC. Effects of TEGDMA and HEMA on the ex-pression of COX-2 and iNOS in cultured murine macrophage cells. Dent Mater2009;25:240–6.
[18] Pathan AR, Viswanad B, Sonkusare SK, Ramarao P. Chronic administration of pioglit-azone attenuates intracerebroventricular streptozotocin induced-memory impair-ment in rats. Life Sci 2006;79:2209–16.
[19] Grunblatt E, Salkovic-Petrisic M, Osmanovic J, Riederer P, Hoyer S. Brain insulin sys-tem dysfunction in streptozotocin intracerebroventricularly treated rats generateshyperphosphorylated tau protein. J Neurochem 2007;101:757–70.
[20] KimWY, Kim JM, Han SB, Lee SK, Kim ND, ParkMK, et al. Steaming of ginseng at hightemperature enhances biological activity. J Nat Prod 2000;63:1702–4.
[21] Kwon SW, Han SB, Park IH, Kim JM, ParkMK, Park JH. Liquid chromatographic deter-mination of less polar ginsenosides in processed ginseng. J Chromatogr A2001;921:335–9.
[22] Bae EA, Kim EJ, Park JS, Kim HS, Ryu JH, Kim DH. Ginsenosides Rg3 and Rh2 inhib-it the activation of AP-1 and protein kinase A pathway in lipopolysaccharide/interferon-gamma-stimulated BV-2 microglial cells. Planta Med 2006;72:627–33.
[23] Ciaramella A, Salani F, Bizzoni F, Orfei MD, Langella R, Angelucci F, et al. The stimu-lation of dendritic cells by amyloid beta 1–42 reduces BDNF production inAlzheimer's disease patients. Brain Behav Immun 2013;32:29–32.
[24] Mattson MP. Pathways towards and away from Alzheimer's disease. Nature2004;430:631–9.
[25] Mochizuki A, Tamaoka A, Shimohata A, Komatsuzaki Y, Shoji S. Abeta42-positivenon-pyramidal neurons around amyloid plaques in Alzheimer's disease. Lancet2000;355:42–3.
[26] Resende R, Marques SC, Ferreiro E, Simões I, Oliveira CR, Pereira CM. Effect ofα-synuclein on amyloid β-induced toxicity: relevance to Lewy body variant ofAlzheimer disease. Neurochem Res 2013;38:797–806.
[27] Craig LA, Hong NS, McDonald RJ. Revisiting the cholinergic hypothesis in the devel-opment of Alzheimer's disease. Neurosci Biobehav Rev 2011;35:1397–409.
[28] Inestrosa NC, Sagal JP, Colombres M. Acetylcholinesterase interaction withAlzheimer amyloid beta. Subcell Biochem 2005;38:299–317.
[29] Martorana A, Di Lorenzo F, Esposito Z, Lo Giudice T, Bernardi G, Caltagirone C, et al.Dopamine D -agonist rotigotine effects on cortical excitability and central choliner-gic transmission in Alzheimer's disease patients. Neuropharmacology 2013;64:108–13.
[30] Koga T, Bellier JP, Kimura H, Tooyama I. Immunoreactivity for Choline Acetyl-transferase of Peripheral-Type (pChAT) in the trigeminal ganglion neurons ofthe non-human primate Macaca fascicularis. Acta Histochem Cytochem 2013;46:59–64.
[31] Jamal M, Ameno K, Miki T, Tanaka N, Ohkubo E, Kinoshita H. Effects of systemic nic-otine, alcohol or their combination on cholinergic markers in the frontal cortex andhippocampus of rat. Neurochem Res 2010;35:1064–70.
[32] Wang Q, Yu X, Li L, Zheng J. Inhibition of amyloid-β aggregation in Alzheimer Dis-ease. Curr Pharm Des 2013 [PMID:23713775, Epub ahead of print].
[33] Gao Y, Li C, Yin J, Shen J, Wang H, Wu Y, et al. Fucoidan, a sulfated polysaccharidefrom brown algae, improves cognitive impairment induced by infusion of Aβ pep-tide in rats. Environ Toxicol Pharmacol 2012;33:304–11.
[34] Ho GJ, Drego R, Hakimian E, Masliah E. Mechanisms of cell signaling and inflam-mation in Alzheimer's disease. Curr Drug Targets Inflamm Allergy 2005;4:247–56.
[35] Kleinig TJ, Vink R. Suppression of inflammation in ischemic and hemorrhagic stroke:therapeutic options. Curr Opin Neurol 2009;22:294–301.
[36] Zheng Z, Sabirzhanov B, Keifer J. Oligomeric amyloid-{beta} inhibits the proteolyticconversion of brain-derived neurotrophic factor (BDNF), AMPA receptor trafficking,and classical conditioning. J Biol Chem 2010;285:34708–17.
[37] Zhang B, Tang XC, Zhang HY. Alternations of central insulin-like growth factor-1 sen-sitivity in APP/PS1 transgenic mice and neuronal models. J Neurosci Res 2013;91:717–25.
[38] Adlard PA, Perreau VM, Cotman CW. The exercise-induced expression of BDNFwith-in the hippocampus varies across life-span. Neurobiol Aging 2005;26:511–20.
[39] Xu B, Gottschalk W, Chow A, Wilson RI, Schnell E, Zang K, et al. The role of brain-derived neurotrophic factor receptors in the mature hippocampus: modulation oflong-term potentiation through a presynaptic mechanism involving TrkB. J Neurosci2000;20:6888–97.
[40] Sakata K, Mastin JR, Duke SM, Vail MG, Overacre AE, Dong BE, et al. Effects of antide-pressant treatment on mice lacking brain-derived neurotrophic factor expressionthrough promoter IV. Eur J Neurosci 2013;37:1863–74.
[41] Witty CF, Gardella LP, Perez MC, Daniel JM. Short-term estradiol administrationin aging ovariectomized rats provides lasting benefits for memory and the hip-pocampus: a role for insulin-like growth factor-I. Endocrinology 2013;154:842–52.
[42] Jimenez S, Torres M, Vizuete M, Sanchez-Varo R, Sanchez-Mejias E, Trujillo-EstradaL, et al. Age-dependent accumulation of soluble amyloid beta (Abeta) oligomers re-verses the neuroprotective effect of soluble amyloid precursor protein-alpha(sAPP(alpha)) by modulating phosphatidylinositol 3-kinase (PI3K)/Akt-GSK-3betapathway in Alzheimer mouse model. J Biol Chem 2011;286:18414–25.
[43] Zhao HH, Di J, Liu WS, Liu HL, Lai H, Lü. YL. Involvement of GSK3 and PP2A inginsenoside Rb1's attenuation of aluminum-induced tau hyperphosphorylation.Behav Brain Res 2013;241:228–34.

326 S. Chu et al. / International Immunopharmacology 19 (2014) 317–326
[44] Li YN, Wu YL, Jia ZH, Qi JS. Interaction between COX-2 and iNOS aggravatesvascular lesion and antagonistic effect of ginsenoside. J Ethnopharmacol2008;119:305–11.
[45] Kiritoshi S, Nishikawa T, Sonoda K, Kukidome D, Senokuchi T, Matsuo T, et al. Reac-tive oxygen species frommitochondria induce cyclooxygenase-2 gene expression inhuman mesangial cells: potential role in diabetic nephropathy. Diabetes 2003;52:2570–7.
[46] Kim SF, Huri DA, Snyder SH. Inducible nitric oxide synthase binds, S-nitrosylates, andactivates cyclooxygenase-2. Science 2005;310:1966–70.
[47] LintonMF, Fazio S. Cyclooxygenase-2 and inflammation in atherosclerosis. Curr OpinPharmacol 2004;4:116–23.
[48] St-OngeM, FlamandN, Biarc J, Picard S, Bouchard L, Dussault AA, et al. Characterizationof prostaglandin E2 generation through the cyclooxygenase (COX)-2 pathway inhuman neutrophils. Biochim Biophys Acta 2007;1771:1235–45.
[49] Qi JS, Li YN, Zhang BS, Niu T, Liang JH. Peroxynitrite mediates high glucose-inducedosteoblast apoptosis. J Endocrinol Invest 2008;31:314–20.
[50] Batool S, Nawaz MS, Greig NH, Rehan M, Kamal MA. Molecular interaction study ofN1-p-fluorobenzyl-cymserine with TNF-α, p38 kinase and JNK kinase. AntiinflammAntiallergy Agents Med Chem 2013;12:129–35.
[51] Chen H, Sohn J, Zhang L, Tian J, Chen S, Bjeldanes LF. Anti-inflammatory effects ofchicanine onmurinemacrophage by down-regulating LPS-induced inflammatory cy-tokines in IκBα/MAPK/ERK signaling pathways. Eur J Pharmacol 2013;724C:168–74.