genetic variation and relatedness of freshwater pearl mussel
TRANSCRIPT

Karlstad University StudiesISSN 1403-8099
ISBN 91-7063-027-5
Division for Environmental SciencesDepartment of Biology
LICENTIATE THESIS
Karlstad University Studies2005:55
Amra Hadzihalilovic-Numanovic
Genetic Variation and Relatedness
of Freshwater Pearl Mussel
Margaritifera margaritifera
L. Populations
Genetic Variation and Relatedness ofFreshwater Pearl Mussel Margaritifera
margaritifera L. Populations
The two papers presented in this thesis focus on the population genetics of freshwater pearl mussel populations in Sweden, using RAPD-PCR method. Specifically, I stud-ied genetic variation within and between 5 populations in a single drainage area in south-western Sweden. This study was followed by an investigation of evolutionary relationships and genetic variation of 14 populations of freshwater pearl mussel from different drainage areas in south-central Sweden. In both studies, I found that genetic variation was larger than reported from previous studies using other techniques, and that variation was larger between than within populations. I did not found any correla-tion between geographic and genetic distance, which indicates that mussel populations have adapted to local environmental conditions in a relatively short time. I also found that genetic distance between populations from a single drainage area was greater than found in other studies, despite small geographic distances. When comparing populations from multiple drainage areas, I found that populations were highly dif-ferentiated, indicating little gene flow between them. There was no significant positive relation between genetic variation and population size or age structure but there was a significant positive relation between mean age and population size, indicating that many populations have gone through bottlenecks recently.

Karlstad University Studies
2005:55
Amra Hadzihalilovic-Numanovic
Genetic Variation and Relatedness
of Freshwater Pearl Mussel
Margaritifera margaritifera
L. Populations

Amra Hadzihalilovic-Numanovic. Genetic Variation and Relatedness of Freshwater Pearl Mussel Margaritifera margaritifera L. Populations.
Licentiate thesis
Karlstad University Studies 2005:55ISSN 1403-8099ISBN 91-7063-027-5
© The author
Distribution:Karlstad UniversityDivision for Environmental SciencesDepartment of BiologySE-651 88 KARLSTAD SWEDEN+46 54-700 10 00
www.kau.se
Printed at: Universitetstryckeriet, Karlstad 2005

Abstract
The two papers presented in this thesis focus on the population genetics of
freshwater pearl mussel populations in Sweden, using the RAPD-PCR method.
Specifically, I studied genetic variation within and between 5 populations in a
single drainage area in south-western Sweden. This study was followed by an
investigation of evolutionary relationships and genetic variation of 14
populations of freshwater pearl mussel from different drainage areas in south-
central Sweden. In both studies, I found that genetic variation was larger than
reported from previous studies using other techniques, and that variation was
larger between than within populations. I did not found any correlation
between geographic and genetic distance, which indicates that mussel
populations have adapted to local environmental conditions in a relatively short
time. I also found that genetic distance between populations from a single
drainage area was greater than found in other studies, despite small geographic
distances. When comparing populations from multiple drainage areas, I found
that populations were highly differentiated, indicating little gene flow between
them. There was no significant positive relation between genetic variation and
population size or age structure but there was a significant positive relation
between mean age and population size, indicating that many populations have
gone through bottlenecks recently.

Contents Publications 3 Introduction 4 Objectives 6 Material and methods 6 Summary of results 9 Discussion 13 References 16 Acknowledgments 20

Publications
This thesis is based on the following papers which are referred to by their
Roman numerals.
I Hadzihalilovic-Numanovic, A. and Arvidsson, B.L. 2005. Genetic variation
in freshwater pearl mussel (Margaritifera margaritifera) populations in south-
western Sweden. Manuscript.
II Hadzihalilovic-Numanovic, A., Arvidsson, B.L., Österling, E.M. and
Hultman J. 2005. Genetic variability in populations of freshwater pearl mussels
(Margaritifera margaritifera) in relation to population size, age structure and
isolation. Manuscript.
3

Introduction
Freshwater pearl mussels Margaritifera margaritifera L. are members of the class
Bivalvia within the phylum Mollusca. They belong to the order Unionida and
family Margaritiferidae. The genus Margaritifera was first described by
Schumacher in 1816 (Mya margaritifera L. 1758 = Margaritifera margaritifera).
However, the taxonomy of the genus Margaritifera is still unclear and, therefore
also the number of living species within the genus (Araujo and Ramos 2001).
Fossil margaritiferids have been recorded from Cretaceous and Jurassic periods
in North America, Europe and East Asia (Starabogatov 1970). The anatomy of
margaritiferida also suggests this to be one of the oldest families amongst the
Mollusca. During the ice ages, populations may have been restricted to more
southerly refuges in Europe. It is likely that repopulation occurred in Europe
after the end of the last ice age (Purser 1988). Freshwater pearl mussels live on
both sides of the Atlantic in clean streams and rivers with low levels of calcium
carbonate, little organic matter in the substrata and with healthy native
salmonid populations. Population numbers are declining in all countries. The
species is nearly extinct in many areas and only undisturbed river basins
maintain populations with important juvenile recruitment (Araujo and Ramos
2001).
Most freshwater mussels share a complex life cycle because they require a fish
host during their larval stage. The millions of larva, called glochidium, normally
attach to the fish’s gills, where they encapsulate and spend several weeks
completing their development until they metamorphose to a benthic juvenile
(Reutter et al. 2001). The only way mussels can disperse is through their hosts
and information on the relationship between the mussels and their hosts is
therefore essential in any attempt to protect endangered freshwater mussels.
The freshwater pearl mussel is typically dioeciously, in common with other
freshwater bivalves. Sexual maturation occurs between seven and 15 years of
4

age (Young and Williams 1984 a), and the fertile period is continued up to 80
years (Bauer 1992). It is perhaps the longest-lived freshwater animal and the
specie’s life span is between 30 and 167 years (Hastie et al. 2000).
During this century, freshwater pearl mussels have been extirpated from many
regions of central Europe (Buddensiek 1995), and are now being studied in
many countries. Understanding of their ecology, behavior, and genetics are
crucial in order to develop appropriate conservation strategies. According to
Bogan (1993), the reasons for the decline of populations include habitat
alteration and destruction, decline and extinction of fish host populations,
commercial exploitation and introduction of alien species. Mussels are
extremely important to freshwater communities and their disappearance may be
an indication of deteriorating water and environmental quality (Reutter et al.
2001).
Efforts to protect endangered species often entail collecting information on the
distribution and maintenance of genetic variation. Populations that have
recently undergone large decreases in size are expected to lose genetic variation,
which may have important consequences for the long-term viability of a
population. The more variation, the better the chance that at least some of the
individuals will produce offspring that in turn reproduce, thereby propagating
the population. Conservation of the genetic diversity is a fundamental concern
in conservation and evolutionary biology (Montgomery 2000). One reason for
the lack of recruitment in many populations of freshwater pearl mussel may be
reduced genetic variation, which is correlated with small effective population
size (Hedrick 2000).
5

Objectives
This thesis focuses on population genetics of freshwater pearl mussels in
different populations in Sweden. In paper I, the objective was to assess levels of
genetic diversity within and between five populations of freshwater pearl
mussels in one drainage area in south-western Sweden. I predicted that genetic
variation would be larger between than within populations and also that
geographically close populations would be more genetically similar. In paper II
the main purpose was to examine genetic relationships between 14 populations
of the freshwater pearl mussel in south-central Sweden and to examine how
genetic variation was related to population size, age structure and isolation. I
hypothesize that large populations should be more genetically variable, that
genetic variation in populations with very high or low mean age should be less
variable, and that genetic differentiation should be positively related to
geographic distance.
Material and methods
Sample collections, population parameters and DNA extraction
In paper I, two whole mussels and biopsies from another 18 mussels were
taken in May 1998 in five streams from the Byälven drainage area in south-
western Sweden (localities given in abbreviation due to risk of illegal fishing).
Individuals used in paper II were collected from 14 streams in south central
Sweden during summer 1998 (5 populations, see above) and 2004 (9
populations, 15 individuals from each), and preserved in 70% ethanol. The age
structure of populations investigated in 1998 was based on 38 length measured
individuals and the average age-length curve for all 9 populations measured in
2004 (Karlsson 2004). An age-length curve, based on length of the shell
6

ligament (Hendelberg 1960) was constructed for each population (Karlsson
2004). Subsequently, the length of at least 250 individuals was measured in
selected sampling plots in the field in 2004, and used to determine the age
structure of each population. Population sizes were obtained from censuses
conducted by the county authorities, using a standardized method (Eriksson et
al. 1998). Estimated population sizes varied between 1500 and 54 000 mussels,
and the age structure varied highly between populations. The geographic
distances between populations was measured on maps by connecting streams
with the shortest possible water route for dispersing host fish.
In both paper I and II DNA was extracted from approximately 25 mg of foot
tissue, following the NucleoSpin C+T protocol for the isolation of genomic
DNA from cells and tissue (Macherey-Nagel). The DNA was then quantified
and stored at –20 C for up to 6 months, and thawed at room temperature
immediately prior to running RAPD-PCR.
RAPD-PCR analysis and electrophoresis
PCR reactions were performed in a PTC-100 thermocycler (Programmable
Thermal Controller, MJ Research, Inc.), using Operon primers (Operon
Technologies Inc.). Twenty Operon set-C, P and A were evaluated. Sixty
primers were arbitrarily used and tested on approximately twenty individuals to
determine which primers generate clear bands. Of these, in the paper I was
chosen five (OPA 18, OPC13, OPC 14, OPP 12 AND OPV 10) and in the
paper II three were chosen (OPC13, OPC14 and OPV 10). Each amplification
reaction was performed using a single primer and repeated at least once to
verify band autosimilarity and repeatability (Perez et al. 1998).
Amplification products were then separated by gel elektrophoresis. Five µl of
the PCR products was run on 2 % agarose gel stained with ethidium bromide.
7

After electrophoresis the gels were viewed under a UV trans-illuminator and
then photographed using black and white film. Gels were scored using a Gel
Pro Analyzer.
Data analyses
Due to failure to obtain high-quality DNA from more than 10 individuals in
one of the Byälven drainage area populations, only 10 randomly selected
individuals from each of the other populations were included to ensure equal
sample size in the analyses. Each band in the RAPDs profiles was treated as an
independent locus with two alleles, presence or absence of a band (Lynch and
Milligan 1994). Amplification products were scored manually, 1 for presence
and 0 for absence of homologous bands. Since RAPD markers are dominant,
we assumed that each band represented the phenotype at a single bi-allelic locus
(Williams et al. 1990). Relationships among RAPD phenotypes were studied
following the recommendations of Lynch and Milligan (1994).
Population genetic parameters such as expected heterozygosities (or gene
diversity), genetic differentiation, and genetic distance were calculated using
corrections for bias in estimating the allele frequencies. Genetic diversity was
also measured as the percentage of polymorphic bands. The percentage of
polymorphic RAPD loci was calculated for each individual, each population, as
well as the mean and overall value for all populations and each primer.
RAPD marker frequencies were used to calculate the within population gene
diversities (Hs), the total gene diversity (Ht) and the coefficient of genetic
differentiation among populations (Gst) according to the formula by Nei
(1978). The unbiased genetic identity and the genetic distance were estimated
according to Nei (1978).
8

The gene flow between populations was calculated from the estimated Gst
values (Slatkin and Barton, 1989). Cluster analysis was performed using Nei’s
(1978) genetic distance based on the unweighted pair group method with an
arithmetic mean metric (UPGMA). All the above analyses were performed
using POP GENE software (Vers. 1.32, http:/www.ualberta.ca/~fueh/
download.htm). The relation between genetic variability and population
parameters was analyzed by parametric correlation (Pearson correlation), and
the relation between geographic and genetic distance was analyzed by Mantel
test. All analyses were done in SAS (version 9.1).
Summary of results
Paper I
The 50 mussels comprised 49 different phenotypes. No RAPD fragments were
found to be unique to a particular population. Primers produced 5 to 13
polymorphic bands per primer and a total of 47 polymorphic bands across all
50 samples. The number of polymorphic bands displayed per population
ranged from 27 to 34. Most of these bands (82.5 %) were polymorphic among
populations.
Total genetic diversity varied between populations, ranging from 0.15 to 0.21,
with an overall level of 0.23 for all individuals. Mean gene diversity within
populations was 0.18. Most of the genetic variation was found to be between
populations rather than within populations. Average estimates of genetically
effective immigration (Nm) were 1.9. All populations received more than one
immigrant per generation. The mean genetic identity between populations was
94%. A hierarchical cluster analysis of evolutionary relationships among the five
populations indicated that population B and D were the most closely
associated, whereas population E was somewhat more distant (Fig. 1a).
9
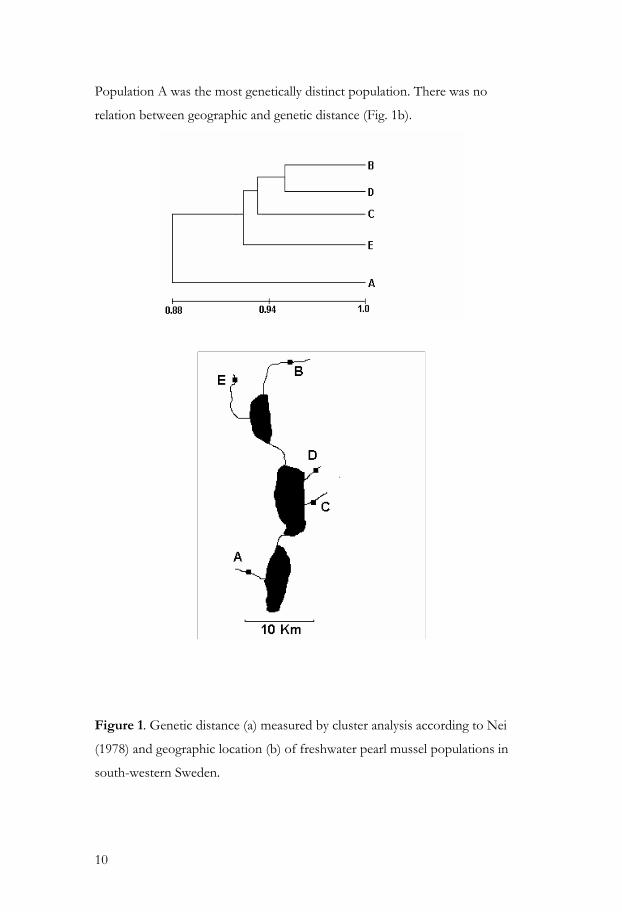
Population A was the most genetically distinct population. There was no
relation between geographic and genetic distance (Fig. 1b).
Figure 1. Genetic distance (a) measured by cluster analysis according to Nei
(1978) and geographic location (b) of freshwater pearl mussel populations in
south-western Sweden.
10
Paper II
All primers produced 100 % polymorphic bands. From the 140 mussels, 133
different RAPD phenotypes were produced. Allele frequencies varied
significantly across populations for all but 6 alleles. Only three unique bands
were found, all in Bratteforsån. The number of polymorphic loci displayed per
population ranged from 11 to 33, and the percentage of polymorphic bands
varied from 28 in Billan to 85 % in Bratteforsån, with an average of 50 % for all
populations. The mean gene diversity for individual populations ranged
between 0.11 and 0.25, with a mean value across populations of 0.17. The
overall gene diversity for all individuals was 0.30. Mean genetic differentiation
across all populations was 0.43, ranging between 0.06 and 1.00 for individual
loci. Genetic differentiation between pairs of populations varied from 0.06 to
0.46.
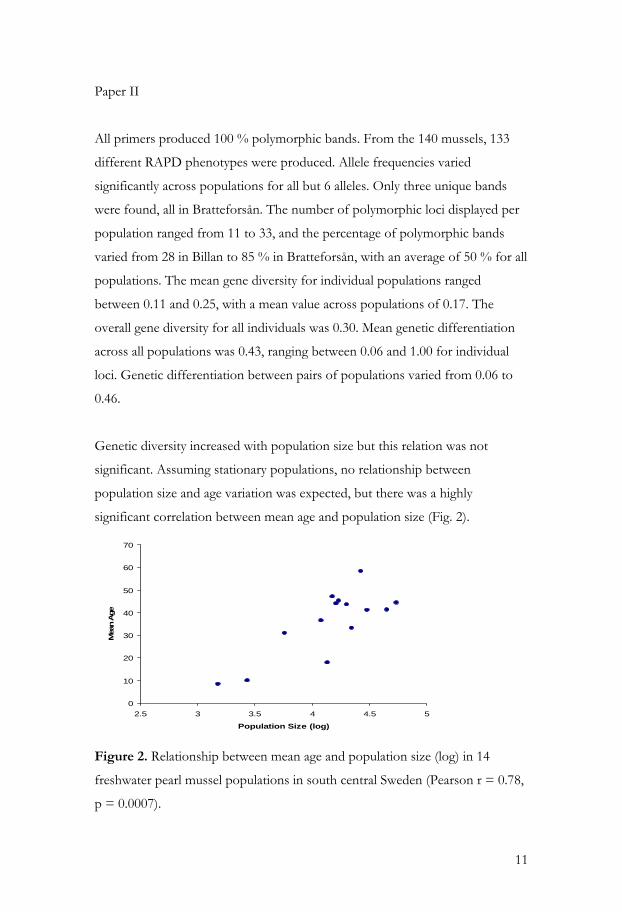
Genetic diversity increased with population size but this relation was not
significant. Assuming stationary populations, no relationship between
population size and age variation was expected, but there was a highly
significant correlation between mean age and population size (Fig. 2).
0
10
20
30
40
50
60
70
2.5 3 3.5 4 4.5 5
Population Size (log)
Mea
n Age
Figure 2. Relationship between mean age and population size (log) in 14
freshwater pearl mussel populations in south central Sweden (Pearson r = 0.78,
p = 0.0007).
11

There was also a marginally non-significant correlation between gene diversity
and mean age (p = 0.07).
The lowest mean value of D was between Billan and Dalsälven (0.01). This
result shows that the populations in Billan and Dalsälven are closely related.
The highest D-values (0.33) were between Torgilsrudsälven and Rällsälven, and
Teåkersälven, respectively. Evolutionary relationships among the 14
populations explored by a hierarchical cluster analysis also showed that
populations Billan and Dalsälven were the most closely associated (Fig. 3).
Figure 3. Genetic relationship between 14 populations of freshwater pearl
mussel in south-central Sweden.
12

The mean genetic distance between individuals for all populations was 0.18.
The estimated number of migrants per generation between populations (Nm)
varied highly between populations. Immigration rate varied between 7.55
individual per generation between Billan and Dalsälven, and 0.55 individuals
per generation between Älgån and Teåkersälven, with an average genetically
effective immigration rate of 0.65. The geographic distance varied from 4.5 km
to 1487 km. A Mantel test showed no significant correlation between genetic
and geographic distances or between the number of immigrants and geographic
distance.
Discussion
Genetic variation
In both papers I and II, we found that genetic variation was larger between
populations than within populations, indicating that freshwater pearl mussel
populations are highly structured today despite high gene flow between 5
populations from Byälven drainage (paper I). However, indirect measures of
gene flow are mean values of the number immigrants from the founding event
of the population. Gene flow estimates therefore contain information that is
relevant to the history of species rather than reveal present gene flow (Bohonak
1999). The gene diversity for all individuals and the percentage of polymorphic
bands in this study are higher than reported in earlier studies of the freshwater
pearl mussel and other threatened mussels (Machodrom 2003, Curole 2004).
The average genetic distance (paper I) among freshwater pearl mussel was
greater in our study, than in previous studies, despite quite small geographic
distances (Chesney et al. 1993, Machodrom 2003). The geographically closest
populations were not the most genetically similar as we had predicted. One
important assumption underlying this prediction is that all populations were
founded after the last glacial period by host fish that originated from a single
lineage (but see Garcia-Martin 1999). In paper II we found a relatively high
13

genetic similarity among geographically separated populations which suggests
that there may be a complicated postglacial history of the populations, with
temporal connections between different drainage systems when the brown
trout colonized the area
Population parameters
Though decreasing populations are expected to lose genetic variation
(Montgomery et al. 2000, Frankham 1996), we did not find any significant
relationship between population size and genetic variation. A positive
relationship is expected to be found with effective population size rather than
the observed population size. Because age-structure varies in our populations,
the effective size may not be correlated with the observed size and this may
explain the lack of a significant relationship when sample size is small. The lack
of a correlation may also be caused by differences in genetic variation found
immediately after settlement after the glacial period if several lineages of brown
trout colonized south-central Sweden (Garcia-Martin 1999). We found a
significant correlation between population size and average age of the
population. One possible explanation could be that populations with a low
average age might have gone through a bottleneck, and are presently increasing.
We also predicted a positive correlation between average age and genetic
variation but this relation was however not quite significant.
Future management
Knowledge about the ecology and habitat requirements, adaptive differences
between populations and genetic variability in individuals and populations are
necessary to develop conservation strategies for the endangered freshwater
pearl mussel. Many freshwater pearl mussel populations have disappeared and
the remaining populations are regarded as decreasing. In most cases the causes
are not known and further studies are needed. Several studies suggest that the
14

survival rates of freshwater pearl mussels during the early post-parasitic phase
are probably crucial and the key issue linked with lack of juvenile recruitment in
most populations (Buddensiek 1995). During their long post-parasitic phase,
pearl mussels depend on a well-oxygenated and stable substrate. Thus, the best
strategy for managing the mussels should probably focus on protecting the
breeding adults and restoring the juvenile habitat (Cosgrove and Hastie 2001).
It is also necessary to restore spawning grounds for brown trout, the host of the
mussel. Releasing artificially infected host fish is a widely accepted strategy
(Bauer 1988). Transplantation of mussels from river to river has been
conducted but is relatively unsuccessful. Translocation should be seen as a last
resort and not first option for river managers (Cosgrove and Hastie 2001).The
implication of this is that introducing new individuals may not be successful in
the long term. Actions should be concentrated on maintaining and enhancing
all current populations. Restocking of salmonids from stocks not originating in
the catchments into which they are released can also be very detrimental, since
the introduced stocks can prove unsuitable for glochidial attachment (Young
and Williams 1984 b). This possibility may only work in rivers where the fish
can live and reproduce without problems (Araujo and Ramos 2001). The best
method is the reintroduction and maintenance of stocks of natural fish hosts in
its former habitat.
15

References
Araujo, R. and Ramos, A. (2001). Action plan for Margaritifera margaritifera.
Council of Europe. T-PVS (2000) 10. Strasbourg, 38 pp.
Bauer, G. (1988). Threats to the freshwater pearl mussel (Margaritifera
margaritifera) in central Europe. Biological Conservation 45, 239-252.
Bauer, G. (1992). Variation in the life span and size of the Freshwater Pearl
Mussel. Journal of Animal Ecology 61, 425-436.
Bogan, A.E. (1993).Freshwater bivalve extinctions (Mollusca: Unionida): a
search for causes. American Zoologist 33, 599-609.
Bohonak, A.J. (1999). Dispersal, gene flow and population structure. Quarterly
Review of Biology 74, 21-45
Buddensiek, V. (1995). The culture of juvenile freshwater pearl mussels
Margaritifera margaritifera L. in cages: A contribution to conservation
programmes and the knowledge of habitat requirements. Biological
Conservation 74, 35-40.
Chesney, H.C.G., Oliver, P.G. and Davis, G.M. (1993). Margaritifera durrovensis
Phillips, 1928: Taxonomic status, ecology and conservation. Journal of
Conchology 34, 267-299.
Cosgrove, P.J. and Hastie, L.C. (2001) Conservation of threatened freshwater
pearl mussel populations: river management, mussel translocation and conflict
resolution. Biological Conservation 99, 183-190.
16

Curole, J.P., Foltz, D.W. and Brown, K.M. (2004) Extensive allozyme
monomorphism in a threatened species of freshwater mussel, Margaritifera
hembeli Conrad (Bivalvia: Margaritiferidae). Conservation Genetic 5, 271-278.
Eriksson, M., Henriksson, L. and Söderberg, H. (1998). Flodpärlmusslan i
Sverige. Naturvårdsverket, Stockholm. Rapport 4887. (In swedish with english
summary).
Frankham, R. (1996). Relationship of genetic variation to population size in
wildlife. Conservation Biology 10, 1500-1508.
Garcia-Martin, J.-L., Utter, F.M. and Pla, C. 1999. Postglacial colonization of
brown trout in Europe based on distribution of allozyme variants. Heredity 82:
46-56.
Hastie, L.C., Young, M.R., Boon, P.J., Cosgrove, P.J. and Henninger, B. (2000).
Sizes, densities and age structures of Scottish Margaritifera margaritifera (L.)
populations. Aquatic conservations: Marine and Freshwater Ecosystems 10,
229-247.
Hedrick, P.W. (2000). Genetics of populations. Jones & Bartlett, Boston.
Hendelberg, J. (1960). The freshwater pearl mussel Margaritifera margaritifera
(L.). Report Inst. Freshwater Research Drottingholm. 41: 149-171.
Karlsson, J. (2004). Age determination, growth rate and age structure of
freshwater pearl mussel (Margaritifera margaritifera) in Västra Götaland and
Örebro Counties. Honorary M.Sc Thesis, Karlstad University.
17

Lynch, M. and Milligan, B.G. (1994) Analysis of population genetic structure
with Rapd markers. Molecular Ecology 3, 91-99.
Marchordom, A., Araujo, R., Erpenbeck, D. and Ramos, M.A. (2003)
Phylogeography and conservation genetics of the endangered European
Margaritiferidae (Bivalvia: Unionidae).Biology Journal of the Linnean Society,
78, 235-252.
Montgomery, M.E., Woodworth, L.M., Nurthen, R.K., Gilligan, D.M., Briscoe,
D.A. and Frankham, R. (2000) Relationships between population size and loss
of genetic diversity:comparisons of experimental results with theoretical
predictions. Conservation Genetics 1, 33-43.
Nei, M. (1978). Estimation of average heterozygosity and genetic distance from
a small number of individuals. Genetics 89, 583-590
Perez, T., Albornoz, J. and Dominguez. A. (1998). An evaluation of RAPD
fragment reproducibility and nature. Molecular ecology 7, 1347-1357.
Purser GJ (1988). Factors affecting the distribution of the freshwater pearl
mussel (Margaritifera margaritifera) (L)) in Britain. Unpublished PhD Thesis,
University of Aberdeen.
Reutter, D.S., Patrick, F. and Charters, D.A. (2001) Enviromental
considerations for construction of bridges and protected freshwater mussel
species, a case study. ICOET 2001 Procedings, pp 46-50.
Slatkin, M. and Barton, N. (1989). A comparison of the three indirect methods
for estimating average levels of gene flow. Evolution 43, 1349-1368
18

Starobogatov, Y.A. (1970). Fauna of mollusks and zoogeography of continental
water bodies. 372 pp.Nauka, Leningrad. (In Russian).
Young, M. and Williams, J. (1984a). The reproductive biology of the freshwater
pearl mussel Margaritifera margaritifera (Linn.) in Scotland. I. Field studies. Archiv
fur Hydrobiologie 99, 405-422.
Young, M. and Williams, J. (1984b). The reproductive biology of the freshwater
pearl mussel Margaritifera margaritifera (Linn.) in Scotland. II. Laboratory studies.
Archiv fur Hydrobiologie 100, 29-143.
Williams J.G.K., Kubelik A.R., Livak J.K. Rafalski J.A. and Tingey S.V. (1990).
DNA polymorphisms amplified by arbitrary primers are useful as genetic
markers. Nucleic Acids Research. 18, 6531-6535.
19

Acknowledgements
There are many people to thank for encouraging me to finish this thesis. I
would like to give special thanks to Birgitta McEwan for supporting my efforts
to begin this study. To Prof. Larry Greenberg for critically reading the
manuscript in various stages of completion and giving valuable comments. To
Jan Nilsson for manuscript reading and good advice. To Martin Österling and
Jens Hultman for collecting mussels used in analyses and for giving me
information on age structures. I would also thank all my colleagues at the
biology department.
Of course, most thanks to my supervisor, Björn Arvidsson, for always taking
time and for helping me in many ways to finish this thesis. All my friends (you
know who you are!) for supporting me. Last but not least I would like to thank
my family, Mensur, Amar and Kerim, for putting up with me…
20

Karlstad University StudiesISSN 1403-8099
ISBN 91-7063-027-5
Division for Environmental SciencesDepartment of Biology
LICENTIATE THESIS
Karlstad University Studies2005:55
Amra Hadzihalilovic-Numanovic
Genetic Variation and Relatedness
of Freshwater Pearl Mussel
Margaritifera margaritifera
L. Populations
Genetic Variation and Relatedness ofFreshwater Pearl Mussel Margaritifera
margaritifera L. Populations
The two papers presented in this thesis focus on the population genetics of freshwater pearl mussel populations in Sweden, using RAPD-PCR method. Specifically, I stud-ied genetic variation within and between 5 populations in a single drainage area in south-western Sweden. This study was followed by an investigation of evolutionary relationships and genetic variation of 14 populations of freshwater pearl mussel from different drainage areas in south-central Sweden. In both studies, I found that genetic variation was larger than reported from previous studies using other techniques, and that variation was larger between than within populations. I did not found any correla-tion between geographic and genetic distance, which indicates that mussel populations have adapted to local environmental conditions in a relatively short time. I also found that genetic distance between populations from a single drainage area was greater than found in other studies, despite small geographic distances. When comparing populations from multiple drainage areas, I found that populations were highly dif-ferentiated, indicating little gene flow between them. There was no significant positive relation between genetic variation and population size or age structure but there was a significant positive relation between mean age and population size, indicating that many populations have gone through bottlenecks recently.