generation of two-color transgenic zebrafish using the green and red fluorescent protein reporter...
TRANSCRIPT

Generation of Two-color Transgenic Zebrafish Using theGreen and Red Fluorescent Protein Reporter Genes gfpand rfp
Haiyan Wan, Jiangyan He, Bensheng Ju, Tie Yan, Toong Jin Lam, and Zhiyuan Gong*
Department of Biological Sciences, National University of Singapore, Singapore 119260
Abstract: Two tissue-specific promoters were used to express both green fluorescent protein (GFP) and red
fluorescent protein (RFP) in transgenic zebrafish embryos. One promoter (CK), derived from a cytokeratin
gene, is active specifically in skin epithelia in embryos, and the other promoter (MLC) from a muscle-specific
gene encodes a myosin light chain 2 polypeptide. When the 2 promoters drove the 2 reporter genes to express
in the same embryos, both genes were faithfully expressed in the respective tissues, skin or muscle. When the
2 fluorescent proteins were expressed in the same skin or muscle cells under the same promoter, GFP fluo-
rescence appeared earlier than RFP fluorescence in both skin and muscle tissues, probably owing to a higher
detection sensitivity of GFP. However, RFP appeared to be more stable as its fluorescence steadily increased
during development. Finally, F1 transgenic offspring were obtained expressing GFP in skin cells under the CK
promoter and RFP in muscle cells under the MLC promoter. Our study demonstrates the feasibility of
monitoring expression of multiple genes in different tissues in the same transgenic organism.
Key words: dsRed, EGFP, cytokeratin, myosin light chain, skin, muscle.
INTRODUCTION
The green fluorescent protein (gfp) gene, originally isolated
from the jellyfish Aequorea victoria, is widely used as a re-
porter gene for investigation of tissue-specific gene expres-
sion and cellular localization of proteins because the fluo-
rescence of its protein product, GFP, can be conveniently
detected in living cells (Prasher et al., 1992; Chalfie et al.,
1994; Tsien, 1998). Since the introduction of the wild-type
GFP, many mutant forms have been created and screened
for improvement of fluorescence brightness or for altered
spectra of fluorescence shifted toward red or blue (Tsien
and Prasher, 1998). So far, several mutant forms of GFP are
commercially available, and they display different spectra of
fluorescence from that of wild-type GFP. The wild-type
GFP has a maximal spectrum at 508 nm, while several mu-
tant forms have maximal spectra at 440 nm (blue), 477 nm
(cyan), and 527 nm (yellow); these mutant forms are called
blue fluorescent protein (BFP), cyan fluorescent protein
(CFP), and yellow fluorescent protein (YFP), respectively
(Clontech Catalog 2000, pp. 209–222). The availability of
these different fluorescent proteins makes it feasible to carry
out multiple labeling of different organelles or structures
within the same cells or different tissues or cells in the same
organism.
Recently, a new fluorescent protein gene, red fluorescent
protein (rfp), was isolated from a sea anemone relative (Dis-
Received March 15, 2001; accepted September 18, 2001.
*Corresponding author: telephone 65-8742860; fax 65-7792486; e-mail
Mar. Biotechnol. 4, 146–154, 2002DOI: 10.1007/s10126-001-0085-3
© 2002 Springer-Verlag New York Inc.

cosoma sp.) (Matz et al., 1999), and the fluorescent protein
has a red emission spectrum with the maximum at 583 nm.
RFP is distantly related to GFP in primary sequence, and the
fluorescence can also be easily detected without any pre-
treatment. Thus, RFP is also an ideal reporter protein for
living color and can be used for multiple labeling in con-
junction with GFP and its derivatives.
So far, GFP has been widely used for transgenic re-
search in a wide variety of species, including bacteria, yeast,
plants, Caenorhabditis elegans, Drosophila, zebrafish, frogs,
and mice (Chalfie and Kain, 1998). As an important model
organism in genetics and developmental biology, zebrafish
is particularly suitable for GFP expression and detection.
The transparency of zebrafish embryos and external devel-
opment make it feasible to visualize GFP throughout em-
bryo genesis and larval development. Therefore, transgenic
expression of GFP in zebrafish under tissue-specific pro-
moters has become a powerful tool to recapitulate endog-
enous gene expression programs (Higashijimas et al., 1997,
2000; Long et al., 1997, 2000) and to analyze the function of
gene promoters (Meng et al., 1997, 1999; Ju et al., 1999;
Muller et al., 1999; Chen et al., 2001). Recently, gfp under a
heat-shock-inducible promoter has been introduced into
zebrafish, and the inducible expression of GFP provides a
real opportunity to trace cell lineage at any given stage
(Halloran et al., 2000). In addition, gfp transgenic zebrafish
also have the potential use in tracing cell migration, organ
genesis, nuclear and cellular transplantation, purification of
specific cells for a cell-type-specific complementary DNA
library, establishment of in vitro cell lines, etc.
As both gfp and rfp genes are available, it is interesting
to determine the feasibility of generating 2-color transgenic
animals. In the present study, we used the zebrafish as a
model to transfer gfp and rfp driven by 2 different tissue-
specific promoters. In transgenic zebrafish embryos, both
fluorescent proteins were expressed efficiently and faithfully
according to the specificity of the promoters used. Germ-
line-transmitted F1 generation fish with GFP expression in
skin and RFP expression in muscle were obtained, confirm-
ing the tissue-specific expression of the 2 fluorescent pro-
teins under 2 different tissue-specific promoters.
MATERIALS AND METHODS
Zebrafish Maintenance
Zebrafish were purchased from a local ornamental fish farm
and maintained in our aquarium according to the Zebrafish
Book (Westerfield, 1995). Developmental stages of embryos
are presented as hour postfertilization (hpf) or day postfer-
tilization (dpf) at 28.5°C, according to Kimmel et al. (1995).
DNA Constructs
Gene promoters were isolated by linker-mediated nested
polymerase chain reaction (PCR), as previously described
by Liao et al. (1997). Two promoters were used in the
present study: one from a cytokeratin gene (krt8) for skin
specificity (Ju et al., 1999) and the other from a myosin light
chain 2 gene (mylz2) for muscle specificity (Xu et al., 1999,
2000). The gfp and rfp reporter gene constructs, pEGFP-1
and pDsRed-1, were purchased from Clontech (Palo Alto,
Calif.). The 2 gene promoters, 2.2 kb from the cytokeratin
gene (CK) and 2.0 kb from the myosin light chain 2 gene
(MLC), were inserted into the 2 reporter gene constructs in
front of the coding region at the EcoRI and BamHI sites.
The resulting constructs were named pCK-EGFP, pCK-
RFP, pMLC-EGFP, and pMLC-RFP, respectively.
Microinjection and Fluorescence Detection
Microinjection was carried out as previously described (Ju
et al., 1999). Linearized plasmid DNA was injected into
embryos of 1- to 2-cell stage at a concentration of 150 µg/ml
in 0.1 M Tris-HCl/0.25% Phenol red. When 2 different
DNA constructs were injected, the concentration of each
DNA construct was 75 µg/ml. DNA (300–500 pl) was in-
jected into each embryo. Injected embryos were kept at
28.5°C and monitored and photographed using a Zeiss Ax-
iovert 25 fluorescence microscope. GFP was observed with
the filter BP450-490 (blue light) and RFP was observed with
the filter BP546 (yellow light). Hatched embryos or fry were
anesthetized in 0.1% 2-phenoxyethanol prior to being pho-
tographed.
Screening of Transgenic F1 and Cryosection
Embryos injected with pCK-EGFP/pMLC-RFP were raised
to adults. Each adult founder was crossed with a nontrans-
genic zebrafish, and at least 100 offspring from the cross
were examined for expression of fluorescent proteins.
Among 18 founders examined, 1 produced embryos ex-
pressing both fluorescent proteins. Some of the transgenic
fry were fixed in 4% paraformaldehyde in phosphate-
buffered saline overnight at 4°C and sectioned at 15-µm
thickness by a cryostat prior to the examination of GFP and
RFP expression as described above.
Two-color Transgenic Zebrafish 147
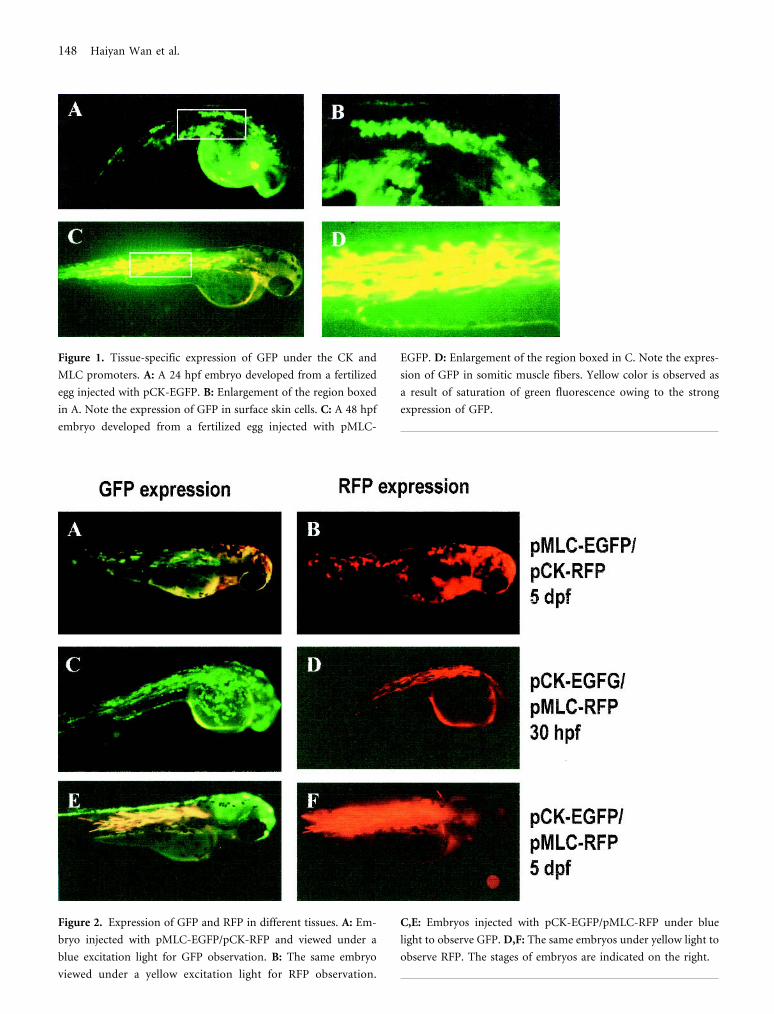
Figure 1. Tissue-specific expression of GFP under the CK and
MLC promoters. A: A 24 hpf embryo developed from a fertilized
egg injected with pCK-EGFP. B: Enlargement of the region boxed
in A. Note the expression of GFP in surface skin cells. C: A 48 hpf
embryo developed from a fertilized egg injected with pMLC-
EGFP. D: Enlargement of the region boxed in C. Note the expres-
sion of GFP in somitic muscle fibers. Yellow color is observed as
a result of saturation of green fluorescence owing to the strong
expression of GFP.
Figure 2. Expression of GFP and RFP in different tissues. A: Em-
bryo injected with pMLC-EGFP/pCK-RFP and viewed under a
blue excitation light for GFP observation. B: The same embryo
viewed under a yellow excitation light for RFP observation.
C,E: Embryos injected with pCK-EGFP/pMLC-RFP under blue
light to observe GFP. D,F: The same embryos under yellow light to
observe RFP. The stages of embryos are indicated on the right.
148 Haiyan Wan et al.

RESULTS
Tissue Specificity of CK and MLC Promoters
The 2 promoters used in the present study have been pre-
viously characterized in transient transgenic zebrafish em-
bryos (Ju et al., 1999) or by direct injection of naked DNA
into adult muscles (Xu et al., 1999). When pCK-EGFP was
injected into zebrafish embryos, GFP was specifically ex-
pressed in the surface skin cells in early embryos (Figure 1,
A and B), consistent with our earlier observation (Ju et al.,
1999). In embryos injected with pMLC-EGFP, GFP expres-
sion was specifically detected in trunk skeletal muscles (Fig-
ure 1, C and D). These expression patterns have been con-
firmed in stable transgenic zebrafish lines by expression of
GFP under the 2 promoters (our unpublished observa-
tions). Therefore, the 2 promoters used in the present study
were highly tissue-specific.
Two Fluorescent Proteins Expressed in TwoDifferent Tissues
To determine whether the 2 fluorescent proteins driven by
different tissue-specific promoters can be expressed cor-
rectly in targeted tissues, pMLC-EGFP/pCK-RFP and pCK-
EGFP/pMLC-RFP were respectively coinjected into ze-
brafish embryos. As shown in Figure 2, both fluorescent
proteins were presented faithfully in the expressing tissues
according to the respective promoters.
In embryos injected with pMLC-EGFP/pCK-RFP, GFP
and RFP were correctly expressed in muscle and skin cells,
respectively (Figure 2, A and B). RFP was first detected in
skin cells at 16 hpf, while GFP was first expressed in muscle
fibers of somites at 22 hpf. It is interesting to note that
strong expression of RFP can be detected as orange color
under the excitation light for GFP (Figure 2, A); however,
no GFP fluorescence can be observed under the excitation
light for RFP (B).
In embryos injected with pCK-EGFP/pMLC-RFP
(Figure 2, C–F), GFP was first detected at the superficial
layer of injected embryos at 6 hpf (shield stage), and the
expression continued in the outmost layer of skin cells dur-
ing embryogenesis. This is in contrast to the earliest detec-
tion of RFP expression at 16 hpf in embryos injected with
pMLC-EGFP/pCK-RFP. In muscle cells, RFP was first ob-
served at 30 hpf, also later than the earliest detection of GFP
(22 hpf) under the same MLC promoter. Thus, GFP is
detected earlier than RFP in the same tissue under the same
promoter.
Unexpectedly in the embryos injected with pCK-EGFP/
pMLC-RFP, GFP was also observed in muscle fibers at the
same stage as RFP detection. It is apparent that all GFP-
expressing muscle fibers also express RFP (Figure 2, C).
This phenomenon is likely due to the formation of a het-
eroconcatemer of the 2 injected plasmid DNAs, and thus
the muscle-specific elements from pMLC-RFP affected the
expression of gfp, as previously reported by Muller et al.
(1997). During embryogenesis, RFP accumulation increased
steadily in muscle fibers. In comparison, GFP fluorescence
in muscle fibers became relatively weak and seemed to dis-
appear after 72 hpf (Figure 2, E).
Despite the observation of “ectopic” expression of GFP
in muscles of the embryos injected with pCK-EGFP/pMLC-
RFP, we never observed the expression of RFP in the
muscles of the embryos injected with the reciprocal com-
bination, pCK-RFP/pMLC-EGFP. Neither was “ectopic”
expression of GFP or RFP observed in skin cells from em-
bryos injected with either combination, pCK-EGFP/pMLC-
RFP or pCK-RFP/pMLC-EGFP.
Two Fluorescent Proteins Expressed in IdenticalCells or Tissues
To investigate whether there is any interference between the
2 fluorescent proteins if they are expressed in identical cells,
coinjection of pCK-EGFP/pCK-RFP or pMLC-EGFP/
pMLC-RFP was carried out. In both cases, GFP and RFP
were correctly expressed in the same tissues and presented
in identical sets of cells (Figure 3). However, as observed
from injection experiments with 2 heterogenous promoter
constructs, GFP generally appears about 10 hours earlier
than RFP. The first detection of GFP in skin and muscle was
around 5 hpf and 22 hpf, respectively; in comparison, the
appearance of RFP in the 2 tissues was 16 hpf and 30 hpf,
respectively. The timing of GFP and RFP appearance in
embryos injected with different combinations of DNA con-
structs is summarized in Figure 4.
Although RFP was detected later than GFP, the inten-
sity of its fluorescence increased steadily during develop-
ment. By 72 hpf, only RFP fluorescence was detected in skin
cells in embryos injected with pCK-EGFP/pCKRFP (Figure
3, C and D). Similarly, under the MLC promoter, RFP
fluorescence became predominant after 5 dpf and GFP fluo-
rescence was greatly overshadowed, even under the optimal
excitation light for GFP (Figure 3, G and H).
Two-color Transgenic Zebrafish 149

Two-color Transgenic F1
To examine the expression of GFP and RFP in stable trans-
genic zebrafish, embryos injected with pCK-EGFP/pMLC-
RFP were raised to adulthood and screened for transgenic
offspring. One founder was identified to produce embryos
expressing both GFP and RFP. The transmission of trans-
gene to F1 is 10.6% (n = 85), indicating transgene mosa-
icism in the founder. In F2 offspring, standard Mendelian
ratio was observed, indicating stable integration of trans-
gene in germline. The transgenic offspring displayed green
fluorescence in skin and red fluorescence in muscle (Figure
5, A and B). Cross section of transgenic offspring confirmed
that GFP expression was restricted to the superficial layer of
skin cells and RFP was expressed specifically in skeletal
muscle (Figure 5, C and D). Similar to the observation in
the transient expression assay, RFP in cross sections under
a blue filter can also be viewed as orange (Figure 5, C).
Since the 2 fluorescent proteins were always expressed
in the same individuals, the 2 gene constructs were likely
Figure 3. Expression of GFP and RFP in the same tissue. A,C:
Embryos injected with pCK-EGFP/pCK-RFP and viewed under a
blue excitation light for GFP observation. B,D: The same embryos
viewed under a yellow excitation light for RFP observation. E,G:
Embryos injected with pMLC-EGFP/pMLC-RFP under blue light
to observe GFP. F,H: The same embryos under yellow light to
observe RFP. The stages of embryos are indicated on the right. The
yellow color of skin cells in C is due to the saturation effect of
fluorescence and correlates to cells with the highest intensity of red
fluorescence.
150 Haiyan Wan et al.

integrated into the same chromosome locus. More likely,
the 2 constructs had been concatenated and integrated. Un-
like the expression in transient transgenic embryos, where
weak GFP expression was also detected in muscle (Figure 2,
C), no GFP expression was detected in muscles from stable
transgenic embryos and fry.
DISCUSSION
Using the gfp reporter gene under zebrafish fish gene pro-
moters, faithful expression of the transgene has been re-
peatedly demonstrated (Higashijimas et al., 1997, 2000;
Long et al., 1997, 2000; Meng et al., 1997, 1999; Ju et al.,
1999; Muller et al., 1999). In the present study, using 2
different fluorescent reporter genes, we have further dem-
onstrated the faithful expression of the 2 transgenes simul-
taneously in the same fish and thus proved the feasibility of
generating 2-color transgenic animals. Multicolor trans-
genic animals should be useful in examination and tracing
of the development of 2 or more tissues and organs simul-
taneously in the same animal, thus providing a valuable tool
for a better and closer comparison. In particular, the com-
Figure 5. Expression of GFP and RFP in a transgenic F1 fry (5 dpf)
from a founder injected with pCK-EGFP/pMLC-RFP. The same
embryo is show for GFP expression (A) and for RFP expression
(B). A transgenic F1 fry of the same stage was cross-sectioned
through the trunk for observation of GFP expression (C) and RFP
expression (D). The same section was used in C and D under
different filters.
Figure 4. Summary of the timing
of appearance of GFP and RFP in
embryos injected with different
combinations of DNA constructs.
Green bars indicate GFP expression
and red bars indicate RFP
expression. The injected DNA
constructs and expressing tissues
are indicated on the left and right,
respectively. The hours after
fertilization are indicated at both
the top and bottom.
Two-color Transgenic Zebrafish 151

parison is valuable for 2 adjacent tissues or cell types. For
example, expression of the 2 fluorescent proteins can be
targeted to endocrine and exocrine cells of pancreas under
2 suitable promoters, and thus a useful transgenic model
could be created for detailed study of pancreas development
and the interaction of different types of cells in the pan-
creas.
When the 2 fluorescent proteins were expressed in the
same tissues, GFP generally appeared several hours earlier
than RFP. This was not likely to be due to the timing of
transcription since the same promoter was used. It seems
that GFP is more sensitive for detection than RFP and re-
quires a lower concentration for visualization. The gfp DNA
used in the present study was a mutant form, encoding a
GFP variant containing a critical amino acid substitution in
the chromophore region, and the codons have also been
modified based on preferred human codon usage (Cormack
et al., 1996). The resultant GFP is called enhanced GFP, or
EGFP, which is 35-fold stronger in fluorescence intensity
than the wild-type GFP in human cells and presumably in
other vertebrate cells as well (Yang et al., 1996).
At later stages, however, it seemed that the fluorescence
intensity of RFP in both muscle and skin cells was stronger
than that of GFP. It is thus likely that RFP is more stable
than GFP and that a higher steady level of RFP can be
reached within the cells. Under our conditions, GFP could
only be detected under its excitation light (blue) but not
under the excitation light for RFP (yellow); while RFP,
when it was expressed at a high level, could be detected as
orange under the blue light and frequently interfered with
the detection of GFP. However, strong expression of GFP
seemed to have no effect on detection of RFP. Despite the
potential interference of strong RFP with GFP detection, the
simultaneous visualization of both GFP and RFP under the
same excitation light could be beneficial under certain cir-
cumstances as no switch of light or double exposure is
required for real-time observation.
The reporter genes are commonly used to determine
temporal and spatial patterns of gene expression. For tem-
poral expression, based on our unpublished data in stable
gfp transgenic zebrafish lines, detection of GFP fluorescence
is generally 2 to 3 hours later than detection of gfp messen-
ger RNA in developing embryos because of the delay of the
translation event and accumulation of sufficient GFP for
detection. According to the current study, the appearance of
RFP fluorescence is delayed by about 10 hours compared
with that of GFP fluorescence, and thus it is likely that the
detection of RFP is 12 to 13 hours later than the detection
of its mRNA. Therefore, gfp reporter gene should be a better
choice than rfp if the timing of gene activation is critical in
the analysis.
Although stable transgenic animals have been made for
many species, the fate of exogenously introduced DNA and
how it is integrated into the host chromosomes remain
unclear. It has been reported that the foreign DNA, once
injected into fertilized eggs, forms DNA concatemers and
undergoes a rapid amplification and subsequent degrada-
tion (Flytzanis et al., 1985; Stuart et al., 1988; Chong and
Vielkind, 1989). Because of this, as previously demon-
strated, analysis of DNA cis-elements can be achieved by
coinjection of a promoter-reporter gene construct and a
testing DNA cis-element. Conceivably, the cis-element will
be ligated to the promoter in vivo, thus eliminating the
laborious process of constructing a proper test DNA plas-
mid in vitro. This approach has proved useful in zebrafish
(Muller et al., 1997, 1999). In our present study, as shown
in Figure 2, C, indeed muscle expression of GFP was ob-
served due to the presumable concatemerization of a
muscle element from a heterologous DNA construct. How-
ever, the expression of GFP in this way was relatively weak,
and probably only effective in early embryos in which a
large amount of amplified exogenous DNA remains avail-
able. Consistent with this, the muscle expression of GFP in
embryos injected with pCK-EGFP/pMLC-RFP became
weak and even undetectable at late embryonic stages when
most of the exogenously DNA was presumably degraded.
Furthermore, we never observed muscle expression of RFP
in embryos injected with pCK-RFP/pMLC-EGFP because
more accumulated RFP is required for fluorescence visual-
ization. The “ectopic” expression of GFP and RFP was never
observed in skin cells, probably because the skin-specific CK
promoter is not as strong as the muscle-specific MLC pro-
moter. The lack of muscle expression was further confirmed
in stable transgenic F1 embryos in which only one or a few
copies of transgenes are integrated. Thus, our observation
indicated that a reliable analysis of DNA cis-element re-
mains the approach to introduce in vitro ligated DNA con-
structs.
In summary, in the present study, we have generated
2-color transgenic zebrafish using gfp and rfp reporter
genes, and both genes were correctly expressed in the tar-
geted tissues according to the specificity of the promoters
used. The 2-color transgenic model should facilitate com-
parative studies of development of multiple tissues or or-
gans and differentiation of different cell types in the same
individual.
152 Haiyan Wan et al.

ACKNOWLEDGMENTS
This work was supported by an academic research grant
from National University of Singapore (NUS). H.W. was
supported by a postgraduate scholarship from NUS.
REFERENCES
Chalfie, M., Tu, Y., Euskirchen, G., Ward, W.W., and Prasher,
D.C. (1994). Green fluorescent protein as a marker for gene ex-
pression. Science 263:802–805.
Chalfie, M., and Kain, S. (1998). Green Fluorescent Protein. Prop-
erties, Applications, and Protocols. New York, N.Y.: Wiley-Liss.
Chen, Y.H., Lee, W.C., Liu, C.F., and Tsai, H.J. (2001). Molecular
structure, dynamic expression, and promoter analysis of zebrafish
(Danio rerio) myf-5 gene. Genesis 29:22–35.
Chong, S.S.C., and Vielkind, J.R. (1989). Expression and fate of
CAT reporter gene microinjected into fertilized medaka (Oryzias
latipes) eggs in the form of plasmid DNA, recombinant phage
particles and its DNA. Theoret Appl Genet 78:369–380.
Cormack, B.P., Valdivia, R.H., and Falkow, S. (1996). FACS-
optimized mutants of the green fluorescent protien (GFP). Gene
173:33–38.
Flytzanis, C.N., McMahon, A.P., Hough-Evans, B.R., Katula, K.S.,
Britten, R.J., and Davidson, E.H. (1985). Introduction of cloned
DNA into sea urchin egg cytoplasm: replication and persistence
during embryogenesis. Dev Biol 108:431–442.
Halloran, M.C., Sato-Maeda, M., Warren, J.T., Su, F., Lele, Z.,
Krone, P.H., Kuwada, J.Y., and Shoji, W. (2000). Laser-induced
gene expression in specific cells of transgenic zebrafish. Develop-
ment 127:1953–1960.
Higashijimas, S., Okamoto, H., Ueno, N., Hotta, Y., and Eguchi,
G. (1997). High-frequency generation of transgenic zebrafish
which reliably express GFP in whole muscles or the whole body by
using promoters of zebrafish origin. Dev Biol 192:289–299.
Higashijimas, S., Hotta, Y., and Okamoto, H. (2000). Visualization
of cranial motor neurons in live transgenic zebrafish expressing
green fluorescent protein under the control of the islet-1 pro-
moter/enhancer. J Neurosci 20:206–218.
Ju, B., Xu, Y., He, J., Liao, J., Yan, T., Hew, C.L., Lam, T.J., and
Gong, Z. (1999). Faithful expression of green fluorescent protein
(GFP) in transgenic zebrafish embryos under control of zebrafish
gene promoters. Dev Genet 25:158–167.
Kimmel, C.B., Ballard, W.W., Kimmel, S.R., Ulimann, B., and
Schilling, T.F. (1995). Stages of embryonic development of the
zebrafish. Dev Dyn 203:253–310.
Liao, J., Chan, C.H., and Gong, Z. (1997). An alternative linker-
mediated polymerase chain reaction method using a dideoxy-
nucleotide to reduce amplification background. Anal Biochem 253:
137–139.
Long, Q., Meng, A., Wang, H., Jessen, J.R., Farrell, M.J., and Lin,
S. (1997). GATA-1 expression pattern can be recapitulated in liv-
ing transgenic zebrafish using GFP reporter gene. Development
124:4105–4111.
Long, Q., Huang, H., Shafizadeh, E., Liu, N., and Lin, S. (2000).
Stimulation of erythropoiesis by inhibiting a new hematopoietic
death receptor in transgenic zebrafish. Nat Cell Biol 2:549–552.
Matz, M.V., Fradkov, A.F., Labas, Y.A., Savitsky, A.P., Zaraisky,
A.G., Markelov, M.L., and Lukyanov, S.A. (1999). Fluorescent
proteins from nonbioluminescent Anthozoa species. Nature Bio-
tech 17:969–973.
Meng, A., Tang, H., Ong, B.A., Farrell, M.J., and Lin, S. (1997).
Promoter analysis in living zebrafish embryos identifies a cis-
acting motif required for neuronal expression of GATA-2. Proc
Natl Acad Sci USA 94:6267–6272.
Meng, A., Tang, H., Yuan, B., Ong, B.A., Long, Q., and Lin, S.
(1999). Positive and negative cis-acting elements are required for
hematopoietic expression of zebrafish GATA-1. Blood 93:500–
508.
Muller, F., Williams, D.W., Kobolak, J., Gauvry, L., Goldspink, G.,
Orban, L., and Maclean, N. (1997). Activator effect of coinjected
enhancers on the muscle-specific expression of promoters in ze-
brafish embryos. Mol Reprod Dev 47:404–412.
Muller, F., Chang, B., Albert, S., Fischer, N., Tora, L., and Strahle,
U. (1999). Intronic enhancers control expression of zebrafish sonic
hedgehog in floor plate and notochord. Development 126:2103–
2116.
Prasher, D.C., Eckenrode, G., Ward, W.W., Prendergrast, E.G.,
and Cornmier, M.J. (1992). Primary structure of the Aequorea
victoria green-fluorescent protein. Gene 111:229–233.
Stuart, G.W., McMurray, J.V., and Westerfield, M. (1988). Repli-
cation, integration and stable germ-line transmission of foreign
sequences injected into early zebrafish embryos. Development 103:
403–412.
Tsien, R.Y. (1998). The green fluorescent protein. Annu Rev Bio-
chem 67:509–544.
Two-color Transgenic Zebrafish 153

Tsien, R.Y., and Prasher, D.C. (1998). In: Green Fluorescent Pro-
tein: Properties, Applications, and Protocols, Chalfie, M., and Kain,
S. (eds.). New York, N.Y.: Wiley-Liss, 97–118.
Westerfield, M. (1995). The Zebrafish Book: A Guide for the Labo-
ratory Use of Zebrafish (Danio rerio). University of Oregon.
Xu, Y., He, J., Tian, H.L., Chan, C.H., Liao, J., Yan, T., Lam, T.J.,
and Gong, Z. (1999). Fast skeletal muscle-specific expression of a
zebrafish myosin light chain 2 gene and characterization of its
promoter by direct injection into skeletal muscle. DNA Cell Biol
18:85–95.
Xu, Y., He, J., Wang, X., Lim, T.M., and Gong, Z. (2000). Asyn-
chronous activation of 10 muscle-specific protein (MSP) genes
during zebrafish somitogenesis. Dev Dyn 219:201–215.
Yang, T.T., Kain, S.R., Kitts, P., Kondepudi, A., Yang, M.M., and
Youvan, D.C. (1996). Dual color microscopic imagery of cells
expressing the green fluorescent protein and a red-shifted variant.
Gene 173:19–23.
154 Haiyan Wan et al.