gender- and age-related differences in the association between social relationship quality and trait...
TRANSCRIPT

Gender- and Age-Related Differences inthe Association Between Social
Relationship Quality and Trait Levels ofSalivary Cortisol
Alan Booth and Douglas A. Granger
Pennsylvania State University
Elizabeth A. Shirtcliff
University of Wisconsin
The majority of studies linking individual differences in the quality of socialrelationships and activity of the hypothalamic-pituitary-adrenal (HPA) axishave focused on the early development of attachment between infants andtheir caregivers. Later in development, during middle childhood and ad-olescence, the parallel HPA links to age-appropriate social relationships withpeers, parents, and siblings remain largely unspecified. This study ad-dressed this knowledge gap. Early morning saliva samples were obtainedfrom 367 children in middle childhood (ages 6–10) and 357 adolescents (Mage 5 11–16 years) on two successive days 1 year apart and assayed forcortisol. Latent state–trait modeling was employed to separate variance incortisol levels attributable to ‘‘stable trait-like’’ versus ‘‘state or situationalspecific’’ sources to minimize the high moment-to-moment variation in basaladrenocortical activity. During adolescence but not middle childhood, andfor girls but not boys, lower levels of ‘‘trait cortisol’’ were associated withpoor quality social relationships. The pattern was robust, extending to thequality of relationships with parents, siblings, and peers. Importantly, therelationship was independent of the rates of internalizing or externalizingproblem behavior. We found that isolating the variance in cortisol levels
JOURNAL OF RESEARCH ON ADOLESCENCE, 18(2), 239–260Copyright r 2008, Society for Research on AdolescenceNo Claim to original US government works
Requests for reprints should be sent to Alan Booth, Department of Sociology, 211 OswaldTower, Pennsylvania State University, University Park, PA 16802. E-mail: [email protected]

attributable to stable intrinsic sources revealed an interpretable pattern thatlinked individual differences in basal HPA activity to social relationshipsduring adolescence. Studies are needed to reveal the biosocial mechanismsinvolved in the establishment of this gender- and age-specific phenomenonand to decipher whether or not individual differences in this hormone-behavior link are adaptive.
The majority of studies linking individual differences in the qualityof social relationships and activity of the hypothalamic-pituitary-adrenal (HPA) axis have focused on attachment between infants andtheir caregivers (Ahnert, Gunnar, Lamb, & Barthel, 2004). The findings areconsidered important because they establish the significance of socialcontext as a mediator of individual differences in the set point or thresholdof reactivity related to the psychobiology of the stress response (van Bakel& Riksen-Walraven, 2004). These differences have been shown to influ-ence the developmental trajectories of key childhood outcomes related toemotion regulation, immunity, cognition, and social behavior (e.g., Luec-ken, 1998; Plotsky et al., 2005).
The literature also demonstrates that later in development, during mid-dle childhood and adolescence, social context plays an equally importantrole in child development (Parke & Buriel, 1998). However, in contrast tothe work on early childhood, the parallel HPA links to age-appropriatesocial relationships with peers, siblings, and parents remain largely un-specified during middle childhood and adolescence. That individualdifferences in HPA activity and stress-related reactivity are associated withproblem behavior during middle childhood and adolescence (Granger,Stansbury, & Henker, 1994; Klimes-Dougan, Hasstings, Granger, Usher, &Zahn-Waxler, 2001) suggests that social relationship quality may indeedcontinue to be linked to HPA activity well beyond early childhood. In thepresent study, we address this knowledge gap by exploring the nature ofthe association between HPA activity and social relationship quality withpeers, siblings, and parents during adolescence.
The direction of the association between baseline cortisol and problembehavior is undergoing a serious conceptual change. Early theorists be-lieved that the relationship is mostly positive; the link is now widelyregarded as largely negative, or as both negative and positive. The im-plications of this change for the links between cortisol and behavior dur-ing middle childhood and adolescence are largely unspecified. A priorstudy focused on the link between trait cortisol and externalizing behaviorby the authors (Shirtcliff, Granger, Booth, & Johnson, 2005) revealed anegative relationship between trait cortisol and problem behavior. Studyresults also indicated that gender needs to be taken into account when
240 BOOTH, GRANGER, AND SHIRTCLIFF

examining factors that differentiate associations between baseline cortisoland behavior. The present study expands the knowledge base by exam-ining the associations between baseline cortisol and the extent and qualityof children’s relationships with parents, siblings, and peers while takinginto account gender and stage of development.
DIRECTION OF LINK BETWEEN CORTISOL AND PROBLEMBEHAVIOR
Much of the early research on baseline cortisol’s link with antisocial orproblem behavior tended to emphasize a positive association. Gunnar andVazquez (2001), after a careful review of a substantial number of studies,tentatively concluded that low baseline cortisol (hypocortisolism), asso-ciated with early life adversity such as caretaker insensitivity, neglect, andabuse, elevates cortisol, which results in down-regulation of the set pointof the HPA axis. Although they noted a number of exceptions, they ob-served a pattern in which the low baseline cortisol production is associ-ated with significant and consistent problem behavior among adults andchildren.
Their search yielded a wide range of studies that reported a link be-tween hypocortisolism and problem behavior. The studies they cite re-ported an association between low baseline cortisol and depressedsocioeconomic status and decreased well-being (Brandstadten, Baltes-Gotz, Kirschbaum, & Hellhammer, 1991), job stress (Caplan, Cobb, &French, 1979), disruptive behavior in boys (McBurnett, Lahey, Rathouz, &Loeber, 2000), and child shyness in new social settings (Granger et al.,1994). Other studies support and elaborate on the idea that low cortisollevels are linked to problem behaviors, including conduct disorders(Oosterlaan, Geurts, Knol, & Sergeant, 2005; Pajer, Garner, Rubin, Perel, &Neal, 2001).
Studies of adults provide other examples of links between hypo-cortisolism and problem behavior. A longitudinal study of the frequency,severity, and intensity of a wide range of adverse life events was inverselyrelated to morning cortisol (Rosal, King, Yunsheng, & Reed, 2004). Therewas no indication that they checked to see whether the results differed formen and women. A study of marital role quality indicated that maritalconcerns were associated with low cortisol following waking. No genderdifferences were observed, but women were greatly underrepresented inthe sample (Barnett, Steptoe, Phil, & Gareis, 2005). A study of middle-classmothers of 2-year-old children revealed that mothers with more hours of
RELATIONSHIP QUALITY AND TRAIT CONTROL 241

employment and a large number of children in the household had lowermorning cortisol values than other mothers (Adam & Gunnar, 2001).Mothers who were secure in their relationship functioning had higherlevels of morning cortisol.
Hypocortisolism appears to be unrelated to depression, however,which may be caused by an impairment of negative feedback regulation(Heim, Ehlert, & Hellhammer, 2000), a finding confirmed in a study ofadolescents (Shirtcliff et al., 2005) and confirmed in a study of maltreatedchildren conducted by Cichetti and Rogosch (2001a, 2001b).
Although not an exhaustive review of the literature, evidence continuesto accumulate indicating that low levels of cortisol are related to problembehavior or adverse circumstance for children and adults. Evidence showsfairly consistently that hypocortisolism does not apply to depression. Re-search also suggests that the finding applies to both genders, although astudy using stringent methods assessing cortisol (latent state–trait [LST]modeling) indicated that hypocortisolism is unrelated to externalizingbehavior in female middle childhood and adolescence (Shirtcliff et al.,2005). Also, studies of children have not taken into account stage of de-velopment. The transition from middle childhood to adolescence involvesextensive changes in physical, cognitive, and social development that mayaffect links between social behavior and low cortisol levels.
GENDER DIFFERENCES IN THE QUALITY OF SOCIALRELATIONSHIPS
There is uniform agreement that the way in which males and femalesrelate to others differs in fundamental ways. Research on adults revealsthat a female’s relationships with friends and kin are both more extensiveand intensive than men’s (e.g., Booth, 1972). Females have more contactwith friends and confide in them more than do men. Women have awider network of relatives than do men and spend more time with them.Within the nuclear family mothers tend to be more temporally and emo-tionally involved with their children than do fathers (Collins & Russell,1991; Crouter, Helms-Erikson, Updegraff, & McHale, 1999; Russell & Sae-bel, 1997) and wives monitor the quality of their intimate relationshipsmore closely than do men (Thompson & Walker, 1989). Similar, age-ap-propriate, gender differences are found among boys and girls (Maccoby,2003). For instance, girls receive more verbal communication frommothers than do sons and the communication is better understood byfemale offspring.
242 BOOTH, GRANGER, AND SHIRTCLIFF

These gender differences may have deep roots in an adaptive mecha-nism that increases the chances of survival. Taylor et al. (2000) modeledthese differences within such a framework. They posited that the well-known ‘‘fight or flight’’ stress response may be more characteristic of malesthan females, and that females’ biobehavioral stress responsivity may bemore accurately organized around the notion of ‘‘tend and befriend.’’Rather than relying heavily on dominating others, challenging constraints,taking risks, and escaping overwhelming challenges, females focus moreon care of their children by moving them to safety and calming them, andaffiliating with others, usually females, who will help care for their chil-dren. Supporting this notion is research that indicates that boys, whenfaced with challenging environments, are 10 times more likely to engage inrisk and dominance behavior than are girls (Moffitt, Caspi, Rutter, & Silva,2001). Females deal with challenges by managing interpersonal relation-ships in ways that require taking initiative in interpersonal relations andalliance building. Because the HPA axis is so fundamental to adapting tothe social environment and the nature of the stress-response is so differentfor males and females, it is likely that the association between quality ofsocial relationships and HPA activation will vary by gender. For example,low trait cortisol was found to be related to deviance and risky behavior inboys and not girls, which supports the differences observed by Taylor andcolleagues (Shirtcliff et al., 2005). Given the fairly consistent evidence thatcortisol has an inverse association with negative behavior, it was expectedto have a negative relationship with the quality of relations engaged in byfemale youth with siblings, parents, and peers.
DEVELOPMENTAL DIFFERENCES IN THE QUALITY OF SOCIALRELATIONSHIPS
Throughout middle childhood and early and late adolescence, children gothrough extensive physical, cognitive, self definitional, and social chang-es. These changes are related to alterations in sibling, parent–child, andpeer relationships. Physical maturation between the ages of 10 and 14appears to be associated with sons and their mothers interrupting eachother more and explaining themselves to each other less. Also, sons deferto their mothers less and family interaction becomes more rigid. In con-trast, the father–son relationship during this period is characterized bypaternal assertiveness and adolescent deference (Hill, Holmbeck, Marlow,Green, & Lynch, 1985a, 1985b; Steinberg, 1981). Relationship changes as-sociated with development have also been noted in girls. Declines in thequality of parent–daughter relations were observed from just before the
RELATIONSHIP QUALITY AND TRAIT CONTROL 243

onset of menarche to more than 12 months following its onset (Hill et al.,1985a). At the onset of menarche, mothers were viewed as less accepting,more controlling, and less influential, and mothers reported decreasedparticipation by daughters in family activities. Fathers were also seen asless influential and accepting during this period. On the basis of thesestudies lower parental acceptance was expected during adolescence thanduring middle childhood.
As offspring move into middle and late adolescence, cognitive devel-opment takes the form of thinking in a more relativistic and abstract waysuch that offspring begin to question what parents regard as rules andcustoms and see them as matters of choice and preference (Steinberg & Silk,2002). Such cognitive changes not only have a bearing on parent accep-tance, but are an integral part of developing and expanding associationswith peers. Whether these changes lower parental acceptance is unknown,but the increase in the number of peers is often associated with distancingbetween parents and children that is manifested as less time spent withparents and perhaps fewer expressions of affection (Steinberg & Silk, 2002).
Children spend a great deal of time with their siblings. McHale and Crouter(1996) indicated that middle childhood children spend more time with sib-lings than with parents. In addition, sibling relationships are often intensiveboth positively and negatively (Dunn, 1983). Some sibling conflict is to beexpected; however, prolonged adverse sibling relationship quality is detri-mental to children’s well-being and is reflected in problem behavior (Conger,Conger, & Elder, 1994; Stocker, 1994; Stocker, Burwell, & Briggs, 2002). How-ever, sibling time together diminishes as they move into adolescence.
Given the tendency for (1) a period of parent–child conflict to occur atthe beginning of adolescence, (2) an increase in the distancing of childrenfrom parents throughout adolescence, (3) a decline in sibling interactionduring adolescence, and (4) hypocortisolism associated with negative be-havior, low trait cortisol was expected to be associated with poor parent–offspring relationship quality as well as sibling relationship quality moreso during adolescence than middle childhood. Also, because peer ties aremore salient during adolescence than middle childhood, an inverse re-lationship between low cortisol and time spent with peers was expected tobe more pronounced at the later stage of development.
LST MODELING OF SALIVARY CORTISOL
A comprehensive review of 20 years of literature that cites a link betweenindividual differences in cortisol levels and social behavior reveals an ab-sence of clear findings. It is possible that the cross-study inconsistencies
244 BOOTH, GRANGER, AND SHIRTCLIFF

may be due to the high moment-to-moment variation in adrenocorticalactivity, and the use of sample sizes may be too small to yield adequatestatistical power for the small-to-modest effects expected. As noted byKirschbaum et al. (1990), diurnal variation is controlled by collectingsamples at the same time of day. Three sources of variance in cortisol levelsremain: internal person factors (i.e., trait), external situational and/orperson–situational interaction (i.e., state), and the reliability of the bio-chemical assays used to measure cortisol (i.e., measurement error). If cor-tisol–behavior associations are tested using variance collapsed acrossthese sources, the ability to detect an effect may be compromised. Toaddress this possibility, LST methodology (Kenny & Zautra, 2001; Nessel-roade, 1988; Steyer, Eid, & Schwenkmezger 1997) is used in this study toestimate the variance in cortisol levels attributable to these sources, andrelated trait and state variance separately to social behavior. In a priorstudy the method revealed that male youth with low trait cortisol weremore likely to report higher levels of externalizing behavior problems thanthose with moderate or high trait-like levels, but individual differences instate-like cortisol (moment-to-moment situational variance) were unre-lated to externalizing behavior problems (Shirtcliff et al., 2005). Prior suc-cess and the desire to be able to compare the findings from this study withprior work led to the decision to use LST methods in modeling the linkbetween baseline cortisol and social behavior.
PURPOSE OF THE STUDY
The goal of the study was to assess the nature of the association betweenquality of social relationships and trait and state cortisol. In order to fullyexplore the issue, a broad range of relationships (siblings, parents, andpeers) were studied. Gender-related and age-specific differences in thisassociation and moderation by internalizing and externalizing problembehavior were hypothesized. The authors conclude with a discussion ofthe adaptive value of the observed associations and the social, personal,and biological mechanisms that link trait cortisol to social behavior.
METHODS
Participants
Interview data and saliva samples were obtained from 724 youths (Mage 5 13.5 years; range 5 6–16 years) and their parents as part of a lon-gitudinal study of 400 families, each of which consisted of two biological
RELATIONSHIP QUALITY AND TRAIT CONTROL 245

parents and at least two offspring (see Booth, Johnson, Granger, Crouter, &McHale, 2003). Those who provided saliva were compared with thosewho did not on a variety of demographic and social variables. Those whoprovided saliva did not differ from those who did not in any systematicway.
Participants were recruited via letters sent to families of students inseven counties in a northeastern state. Letters contained descriptions ofthe study and the request that parents return postcards to express theirinterest in the project. Eligible families included nondivorced couples withtwo pre-teens or two adolescents. The families were selected so that thegender composition of the offspring dyad was balanced with approxi-mately equal numbers of male–male pairs, female–female pairs, male–female pairs where the male was older, and female–male pairs where thefemale was older.
Families participating in the study were middle and working class andresided in rural areas, towns, and small cities. Median income for thesepredominantly dual-earner families was $35,000 for fathers and $17,200for mothers. Average educational attainment was 14.4 years. Ninety-sevenpercent of the participants were of European American origin. The re-maining 3% were Asian and Latino. Mean age for offspring was 13.72(range from 10 to 16.5 years in year 1). Parents had been married for anaverage of 17.5 years. Families were given a $100 honorarium and anadditional $25 if they provided saliva samples.
Youth were interviewed in 1997, 1998, and 1999. Separate home inter-views were conducted with both first- and second-born offspring. Duringthese interviews family members reported on their family relationshipexperiences and individual well-being. A practice saliva sample was col-lected during the interview. Participants were then instructed on how tocollect, store, and mail two weekday morning saliva samples following theinterview. Instructions were specific about the time of day in which thesamples were to be collected—before eating breakfast and conductingother early morning preparations for the day. Family member recordedthe time each sample was collected and those times were close to the timethey were expected to provide the samples. A household member (usuallythe mother) was responsible for overseeing the morning saliva collectionsfor the entire family. We are mindful that subjects could misrepresent whatthey did with respect to saliva collection, but it seems unlikely. Familieswere extraordinarily dedicated to the study. Not a single family was lostduring the 3 years of the study. They were contacted many times betweeninterviews and they were paid $100 for each interview and $25 for pro-viding saliva samples. Behavioral data were collected across 3 years andsaliva collection took place in years 2 and 3 of data collection.
246 BOOTH, GRANGER, AND SHIRTCLIFF

Measures
Salivary cortisol. Saliva was collected from each child upon arisingon two consecutive days in year 2 and then again according to the sameschedule 1 year later for a total of four samples. Children who had a feveror problematic samples (i.e., discolored) were asked to provide saliva at alater date. Participants were instructed to provide saliva before brushingteeth or eating the morning meal on each of the four mornings. FollowingDabbs (1991), subjects were instructed to chew Trident original flavor-sugarless gum to stimulate saliva flow, and salivate 10–15 mLs of salivainto a 20 mL scintillation vial over 15–20 minutes. There was sufficientvariation in the time of morning samples (M 5 7:54 a.m., SD 5 2:08) thattime of day was included as a covariate at the state cortisol level.
All assays were done following the final saliva collection and all sam-ples for each person were done in the same batch. Cortisol was assayedusing a modification of a serum-based radioimmunoassay (DiagnosticProducts Corporation, Santa Monica, CA). The test used 200 mL of saliva,and had a lower limit of sensitivity of .03 mg/dL and upper limit of 2.0 mg/dL. The intra-assay coefficient of variation (CV) for cortisol was 7.6%, witha correlation between duplicates of .99, po.0001. The inter-assay CVs,estimated across 128 separate runs for high (1.91 mg/dL), mid (.86 mg/dL)and low (.14 mg/dL) controls, were 4.98%, 5.08%, and 10.42%, respectively.All samples were tested in duplicate, and duplicate values that varied bymore than 5% error were repeated. Cortisol was normally distributed(M skew 5 .70, range .45–.88).
Social relationships. The interviews included three measures ofrelationship quality in three domains relevant to adolescents (parents,siblings, and peers). High scores represented low levels of parentacceptance, sibling intimacy, and time spent with peers. Three waves ofdata for each measure ensured that the trait dimensions of socialrelationships were captured. The mean of the 3 years was used for eachscale.
The Parental Acceptance Scale consists of the first 24 items of the par-ents’ version of the Child’s Report of Parental Behavior Inventory(Schaefer, 1965). Sample items include the following: ‘‘I am a parentwho likes to talk with my child and be with him/her much of the time’’; ‘‘Iam a parent who doesn’t pay much attention to my child’s behavior.’’Answer categories are ‘‘Not like you, Somewhat like you, A lot like you.’’ acoefficients range from .73 to .89 (Schwarz, Barton-Henry, & Pruzinsky,1985). Higher scores represent low acceptance.
RELATIONSHIP QUALITY AND TRAIT CONTROL 247

The Sibling Intimacy Scale developed by Blyth, Hill, and Theil (1982)includes eight items that assess the degree to which offspring perceive thattheir relationship with their sister/brother is a source of emotional close-ness, support, acceptance, and satisfaction (e.g., How much do you shareyour inner feelings or secrets with your sister/brother? How much do you go toyour sister/brother for advice/support?). Cronbach’s a was .85. Items are ratedon a 5-point scale, ranging from ‘‘Almost Always’’ to ‘‘Never’’ and av-eraged. Higher scale scores reflect lower perceived intimacy. The scale wasstable and reliable across 3 years of assessment, a5 .86 (a5 .87 in girls anda5 .74 in boys).
Time spent with peers was calculated from the time-use data collectedduring seven nightly phone interviews. Involvement was measured byoffspring’s reports of the activities (from the daily time-use data) in whichthey participated with same- and opposite-sex peers (non-family mem-bers). The number of minutes spent with friends in each social context wassummed across the seven phone calls. Although collecting time-use data islabor intensive, these data do have some important advantages. Ratherthan relying on participants to remember the frequency and duration ofevents over an extended period of time, which increases the likelihood ofmemory and recall biases (Huston & Robins, 1982), youth are only re-quired to report on their activities for the day that they are telephoned.Gathering this type of specific information over a series of 7 days providesa more accurate description of youth’s time with peers than paper-and-pencil measures in which individuals rate their general level of involve-ment or companionship (Huston & Robins, 1982). Peer relationships werestable across years and thus were averaged, a5 .69 (a5 .76 in girls anda5 .74 in boys). Higher scores represent less time spent with peers.
Age (in years) was used as the primary index of development. Whetherthe inclusion of pubertal status, measured using the Petersen PubertalScale (Petersen, Crockett, Richards, & Boxer, 1988) further explained vari-ance in cortisol measured was explored. Consistent with Rosmalen andcolleagues (2005), pubertal status did not add explanatory power beyondage.
RESULTS
LST Model of Salivary Cortisol
LST was used to examine the association between trait cortisol and be-havior. Figure 1 presents a conceptual overview of the basic LST model.The analyses first examined whether decomposing salivary cortisol intomeasurement error, state- and trait-related variance was viable, and then
248 BOOTH, GRANGER, AND SHIRTCLIFF

modeled the association between social behavior and trait cortisol, whilecontrolling for age in years and time of day. Other potential control mea-sures, including smoking, drug (e.g., anabolic steroids), and oral contra-ceptive use, menstrual cycle status, and day count, did not significantlyinfluence cortisol.
Factor invariance (FI) was used to test whether factor loadings were thesame or different between groups. Constraining the variance to be equalwithin each year did not cause model misspecification, so these con-straints were included in the final model.
Indices of practical fit were used to make judgments about model fit,including the normed fit index (NFI; Bentler & Bonett, 1980), Tucker-Lewisindex (TLI or NNFI; Tucker & Lewis, 1973), the comparative fit index (CFI;Bentler, 1990) and the root mean square error of approximation (RMSEA;Cudeck & Browne, 1992). As revealed in Table 1, conventional evaluationsfor practical fit (Hu & Bentler, 1995) suggest that the fit of each model wasgood.
Dup11
Dup12
Dup21
Dup22
Dup31
Dup32
Dup41
Dup42
Day 1Year 2
Day 2Year 2
Day 3Year 3
Day 4Year 3
Trait Cortisol
Measurem
ent Error/ U
nreliability
State/ Situational Influences
Trait / Consistency
Time of Day
FIGURE 1 Basic latent state–trait model for salivary cortisol. Cortisol was measured insaliva on four mornings across 1 year. The manifest variables were the duplicate assays ofeach saliva sample (Dup.1 & Dup.2). These derived latent estimates of measurement error/unreliability and state cortisol. These four latent factors then derived a second-order latenttrait cortisol variable. State cortisol is the portion of variance unique to situational influenceson each cortisol measure; trait cortisol comprises the portion of variance in cortisol that wasconsistent across time. Time of day was controlled at the state cortisol level. Separate modelswere computed for males and females.
RELATIONSHIP QUALITY AND TRAIT CONTROL 249
In order to test for moderating effects of gender, a multiple groupstrategy was used (Li et al., 2001; Rigdon, Schumacker, & Wothke, 1998). FIprocedures across gender revealed that factor loadings were not statisti-cally different in males and females. Cortisol intercepts were significantlyhigher in females and males, but this difference may be of little practicalsignificance. Strong FI was maintained so that boys and girls had com-parable coefficients (see Table 2 Model A comparisons). Constraining thevariance estimates caused model misfit. State cortisol variance was al-lowed to vary from year to year, but other variance estimates were heldconstant across measurement occasions.
For boys and girls, respectively, variance estimates were 52% and 73%for state cortisol, 19% and 46% for trait cortisol, and 1% and 1.5% for errorunreliability estimates of cortisol. Girls had significantly more error andstate variance than boys, but there were no gender differences in trait-cortisol estimates (see Model B comparisons in Table 2). On average, traitcortisol comprised 31.4% of the total variance in cortisol.
Cortisol and Social Relationship Quality
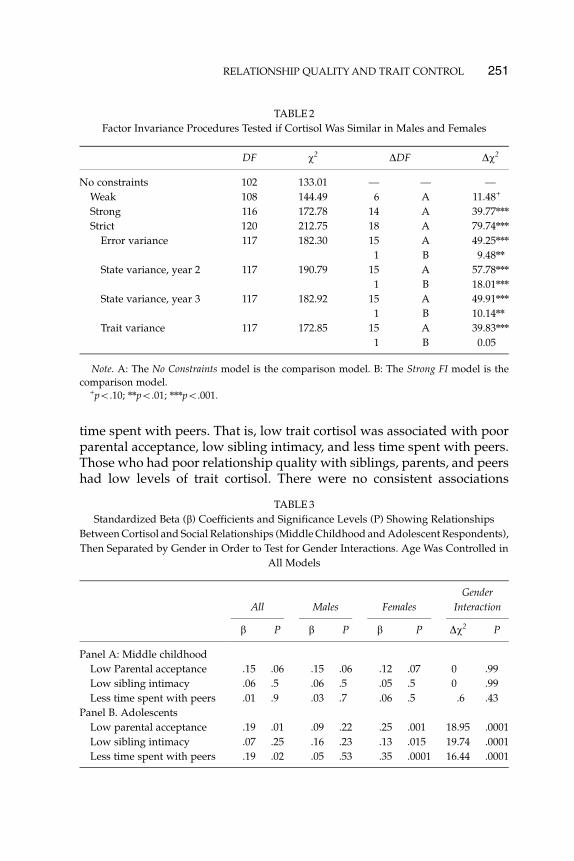
The links between trait cortisol and social behavior differed by stage ofdevelopment. As indicated in Table 3, Panel A, there was no associationbetween trait cortisol and social behavior among individuals in middlechildhood. Among adolescents, however, the associations between traitcortisol and social relationship quality were highly gender-specific amongadolescents (Table 3, Panel B). For girls, the effects were consistent inmagnitude and direction for parental acceptance, sibling intimacy, and
TABLE 1
Chi-Square Goodness of Fit and Indices of Practical Fit for the Basic and Behavioral Models
DF w2 NFI NNFI CFI RMSEA
Basic model 116 185.783nn .984 .993 .994 .027
Low parental acceptance 160 199.04n .988 .997 .998 .025
Low sibling intimacy 160 204.36nn .987 .996 .997 .027
Less time spent with peers 160 205.82nn .986 .996 .997 .027
Note. CFI, comparative fit index; RMSEA, root mean square error of approximation; NFI,
normed fit index.npo.05; nnpo.01.
250 BOOTH, GRANGER, AND SHIRTCLIFF
time spent with peers. That is, low trait cortisol was associated with poorparental acceptance, low sibling intimacy, and less time spent with peers.Those who had poor relationship quality with siblings, parents, and peershad low levels of trait cortisol. There were no consistent associations
TABLE 2
Factor Invariance Procedures Tested if Cortisol Was Similar in Males and Females
DF w2 DDF Dw2
No constraints 102 133.01 — — —
Weak 108 144.49 6 A 11.48+
Strong 116 172.78 14 A 39.77nnn
Strict 120 212.75 18 A 79.74nnn
Error variance 117 182.30 15 A 49.25nnn
1 B 9.48nn
State variance, year 2 117 190.79 15 A 57.78nnn
1 B 18.01nnn
State variance, year 3 117 182.92 15 A 49.91nnn
1 B 10.14nn
Trait variance 117 172.85 15 A 39.83nnn
1 B 0.05
Note. A: The No Constraints model is the comparison model. B: The Strong FI model is the
comparison model.+po.10; nnpo.01; nnnpo.001.
TABLE 3
Standardized Beta (b) Coefficients and Significance Levels (P) Showing Relationships
Between Cortisol and Social Relationships (Middle Childhood and Adolescent Respondents),
Then Separated by Gender in Order to Test for Gender Interactions. Age Was Controlled in
All Models
All Males Females
Gender
Interaction
b P b P b P Dw2 P
Panel A: Middle childhood
Low Parental acceptance � .15 .06 � .15 .06 � .12 .07 0 .99
Low sibling intimacy .06 .5 .06 .5 .05 .5 0 .99
Less time spent with peers .01 .9 � .03 .7 .06 .5 .6 .43
Panel B. Adolescents
Low parental acceptance � .19 .01 � .09 .22 � .25 .001 18.95 .0001
Low sibling intimacy � .07 .25 .16 .23 � .13 .015 19.74 .0001
Less time spent with peers � .19 .02 .05 .53 � .35 .0001 16.44 .0001
RELATIONSHIP QUALITY AND TRAIT CONTROL 251
between state cortisol and the three indicators or poor quality socialaffiliations.
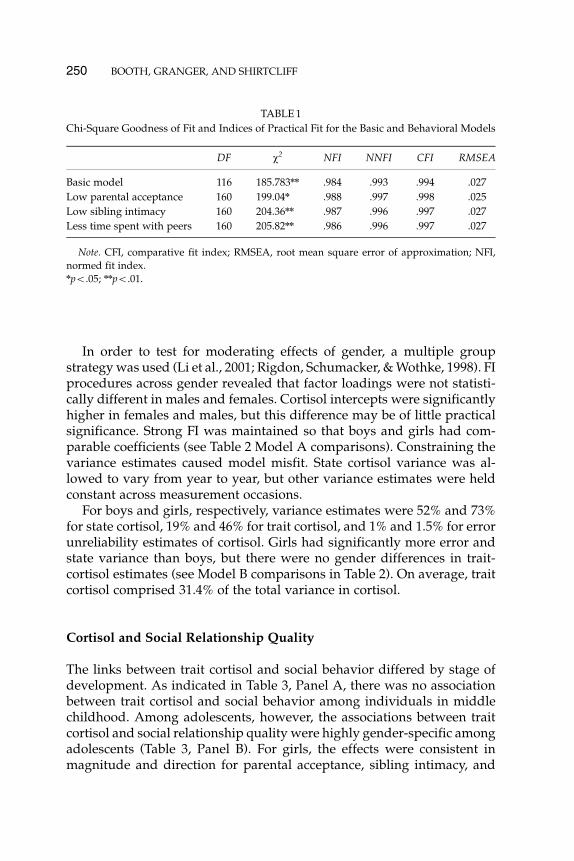
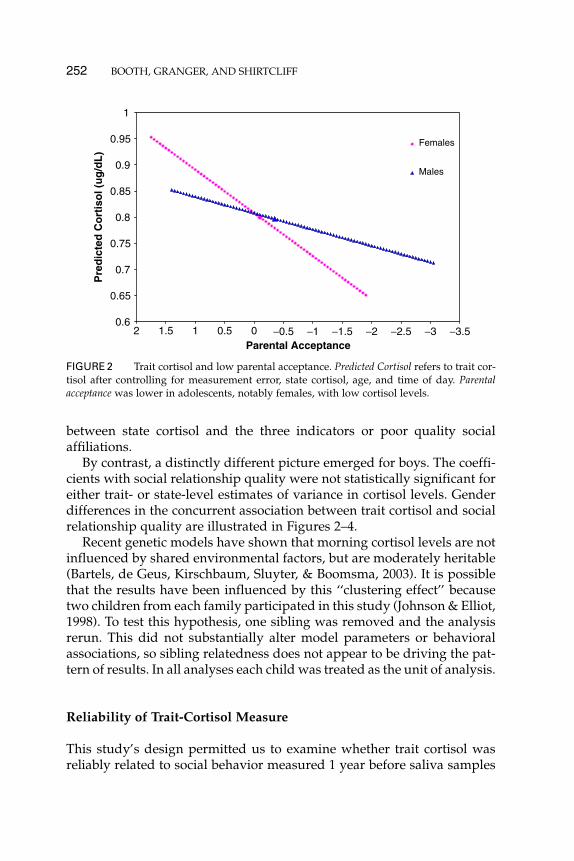
By contrast, a distinctly different picture emerged for boys. The coeffi-cients with social relationship quality were not statistically significant foreither trait- or state-level estimates of variance in cortisol levels. Genderdifferences in the concurrent association between trait cortisol and socialrelationship quality are illustrated in Figures 2–4.
Recent genetic models have shown that morning cortisol levels are notinfluenced by shared environmental factors, but are moderately heritable(Bartels, de Geus, Kirschbaum, Sluyter, & Boomsma, 2003). It is possiblethat the results have been influenced by this ‘‘clustering effect’’ becausetwo children from each family participated in this study (Johnson & Elliot,1998). To test this hypothesis, one sibling was removed and the analysisrerun. This did not substantially alter model parameters or behavioralassociations, so sibling relatedness does not appear to be driving the pat-tern of results. In all analyses each child was treated as the unit of analysis.
Reliability of Trait-Cortisol Measure
This study’s design permitted us to examine whether trait cortisol wasreliably related to social behavior measured 1 year before saliva samples
0.6
0.65
0.7
0.75
0.8
0.85
0.9
0.95
1
−1−0.5 −1.5 −2.5 −3.5−3−200.511.52Parental Acceptance
Pre
dic
ted
Co
rtis
ol (
ug
/dL
)
Females
Males
FIGURE 2 Trait cortisol and low parental acceptance. Predicted Cortisol refers to trait cor-tisol after controlling for measurement error, state cortisol, age, and time of day. Parentalacceptance was lower in adolescents, notably females, with low cortisol levels.
252 BOOTH, GRANGER, AND SHIRTCLIFF
0.6
0.65
0.7
0.75
0.8
0.85
0.9
0.95
1
8131823283338Sibling Intimacy
Pre
dic
ted
Co
rtis
ol (
ug
/dL
)
Females
Males
FIGURE 3 Trait cortisol and low sibling intimacy. Females with low trait cortisol are lessintimate with their siblings, but this was not significant for males.
0.65
0.7
0.75
0.8
0.85
0.9
0.95
1
1.05
1.1
050100150200250300350400450500550650 600Time Spent With Peers
Pre
dic
ted
Co
rtis
ol (
ug
/dL
)
Females
Males
FIGURE 4 Trait cortisol and time spent with peers. Trait cortisol levels were lower inadolescents who spent less time with peers. This effect was driven by females.
RELATIONSHIP QUALITY AND TRAIT CONTROL 253

were first collected. Lower levels of parental acceptance, sibling intimacy,and peer involvement measured 1 year before the first cortisol measurewere significantly related to lower levels of trait cortisol. This suggests thattrait cortisol can be reliably assessed using the LST model. This also meansthat if measures of cortisol and behavior were available over a longerperiod of time, reciprocal influences between the two could be assessed.
CONCLUSION AND DISCUSSION
An analysis of middle childhood and adolescent offspring residing withtwo biological parents in working- and middle-class homes revealed thatadolescent girls, but not boys in either age category, who had lower levelsof trait cortisol also had weak relationships in three interpersonal domainslinked specifically to the quality of social relationships with parents, sib-lings, and peers. Coupled with findings from an earlier study linking lowtrait cortisol with externalizing behavior problems in boys (Shirtcliff et al.,2005), the present study extends understanding of biosocial relationshipsthat involve the HPA axis in important ways. The findings underscore thebelief that isolation of the variance in cortisol levels attributable to stableintrinsic individual differences reveals interpretable patterns of biosocialrelationships in children.
It is not surprising that hypocortisolism is associated with poor socialrelationships to a greater extent in adolescence than in middle childhoodbecause the former are at a stage in which offspring are already distancingthemselves from parents and siblings and increasing affiliations withpeers. It is less clear why the association is observed among females butnot males. The same genre of finding applies to the Shirtcliff et al. (2005)finding that the link between low trait cortisol and externalizing behaviorwas found in males and not females. Perhaps hypocortisolism is linked toeach gender’s specialization—that is, flight or fight behavior among malesand tend and befriend behavior among females. Each gender’s behavioralspecialty becomes harsher. That is, the male’s behavior become more ag-gressive and the female’s behavior become less supportive. Under whatcircumstances would hypocortisolism and negative behavior be adaptive?
For an answer, it is useful to turn to Sapolsky’s (2005) review of the linkbetween social rank and cortisol levels among primates. While his em-phasis was on high cortisol production associated with stress rather thanhypocortisolism, his analysis is useful in interpreting the findings from thepresent study. Sapolsky pointed out that early research suggested that lowstatus was associated with high cortisol production. Later studies sug-gested that high status individuals produced high levels of cortisol. In a
254 BOOTH, GRANGER, AND SHIRTCLIFF

review of these studies Sapolsky (2005) demonstrated that the link be-tween rank and HPA activation depended on the social organization indifferent populations and species. For example, low ranked individuals inegalitarian societies where there is abundant social support do not pro-duce high levels of cortisol relative to subordinate individuals in hierar-chical despotic societies. On the other hand, highly ranked individuals inunstable societies, who constantly have to defend their status, have highlevels of cortisol relative to those highly ranked in stable societies.Throughout human history there have been long periods in which peoplehave lived in unforgiving and harsh environments (Diamond, 1999, 2005).If the response to these environments were protracted periods of elevatedcortisol, which then erodes health, the human species might have becomeextinct. If the harsh environment were sensed perinatally and the stresssystem were down-regulated, low cortisol might have kept them healthybecause the immune system would not have been as compromised asnormally occurs during the typical response to stress. It is well known thatelevated cortisol levels challenge the immune system and increase theprobability of disease and death. The externalizing behavior and theproblematic social behavior associated with hypocortisolism would befunctional in terms of protecting oneself against conspecies and loweringobligations to share food and other resources with kin (including childrenwhen resources are especially low), thereby conserving resources thatenhance the chances of survival by the reproductive pair (Booth, 1984).While this adaptive mechanism is cast in terms of populations living inresource-poor or unstable societies for extended periods, it may also applyto individuals in dysfunctional or extremely disadvantaged families.
The study would have benefited from a more diverse sample withrespect to family structure, race/ethnicity, income, and varying experi-ences with early parent–child relationship quality. Therefore, we recom-mend that the reader use caution in generalizing these findings to otherpopulations.
In addition, greater consistency in the measures of affiliation wouldstrengthen the study. Time spent with peers is quite different from thechild’s report of closeness, acceptance, and satisfaction with siblings—onemeasures time and the other affect. Our use of parental acceptance is basedon the idea that parents are less accepting of children who have poor socialskills or are engaged in antisocial behavior. In support of this idea is thenegative correlation (r 5 � .25, po.001) between children’s report of theirexternalizing behavior and parental acceptance. While the measures aremore diverse and less direct than we would prefer, we believe that the factthey all operate in the same way with respect to trait cortisol supports theoverall conclusions of the study.
RELATIONSHIP QUALITY AND TRAIT CONTROL 255

Information on the quality of care early in life might help us sort out theorigins of low trait cortisol. It would also be helpful to have informationabout whether people’s reaction to the behavior associated with low cor-tisol leads to behavior that further reinforces low levels of trait cortisol.Longitudinal data that extend over a longer period of time would allow usto assess whether trait cortisol changes, and if so, which factors (e.g., animproved or worse environment) bring about change. Such data wouldalso allow us to assess whether activities associated with HPA hypo-arousal in youth lead to more serious problem behavior later in life.
In conclusion, the nature of this study’s design leaves us uncertain as towhether low trait-like cortisol is a cause or consequence of poorsocial relationship quality and virtually in the dark regarding specificmechanisms involved. Nevertheless, these findings are noteworthy be-cause the robust pattern observed here supports an accumulatingliterature, suggesting that during late childhood and early adolescencelow levels of HPA axis activation (salivary cortisol) are linked to negativesocial behavior. This pattern of biobehavioral relationships is genderspecific.
Two main themes emerge with broader implications for the study ofadolescent development. First, our gender-linked findings underscoreZahn-Waxler, Crick, Shirtcliff, and Woods’ (2006) conclusion that all toooften developmental science has ignored the possibility of gender-specificor gender-relevant models and that future models need to allow fordifferent parameters for males and females when biological and environ-mental factors implicated in psychological adjustment or psychopathol-ogy are studied. Second, the findings highlight the value in future studiesof implementing research designs that specifically test the nature of theinterplay between biology and social context. More often than not, devel-opmentally oriented studies that have integrated salivary biomarkers de-scribe interesting associations but fall short of answering questionsneeded to refine and revise our theoretical models.
Recently, Boyce (2006) addressed this issue and defined a set of fivealternative patterns of biology-context interactions. Biological processesmay amplify the pathogenicity of context, amplify the protectiveness ofcontext, control sensitivity to context, select the symptomatic expression ofcontextual pathogenesis, or control exposures to contextual pathogenicity.Although biological processes are noted first, Boyce noted that this is asimple convention and is not meant to imply that biological factors are theactive component of the interaction—these relationships could be stated inreverse. We speculate that the present study’s findings, when viewedthrough this theoretical lens, will generate specific testable hypotheses,and that the next generation of studies will make major steps forward in
256 BOOTH, GRANGER, AND SHIRTCLIFF

translating our observations to address their implications for adolescentdevelopment.
ACKNOWLEDGMENTS
This research was supported in part by the Population Research Institute,Behavioral Endocrinology Laboratory, and the Children, Youth and Fam-ilies Consortium at Pennsylvania State University as well as the W. T.Grant Foundation (Grant 9617796). The Population Research Institute hascore support from the National Institute of Child Health and HumanDevelopment (Grant R24 HD41025). Collection of data on family processand child psychosocial functioning was supported by two grants from theNational Institute of Child Health and Human Development (HD 32336and HD 29409) to Dr. Ann C. Crouter and Dr. Susan McHale. We aredeeply indebted to Drs. Crouter and McHale for comments on an earlierdraft of the paper.
REFERENCES
Adam, E., & Gunnar, M. (2001). Relationship functioning and home and work demandspredict individual differences in diurnal cortisol patterns in women. Psychoneuroendocri-nology, 26, 189–208.
Ahnert, L., Gunnar, M., Lamb, R., & Barthel, M. (2004). Transition to child care: Associationwith infant–mother attachment, infant negative emotion, cortisol elevations. Child Devel-opment, 75, 639–650.
Barnett, R., Steptoe, A., Phil, D., & Gareis, K. (2005). Marital-role quality and stress-relatedpsychobiological indicators. Annuals of Behavioral Medicine, 30, 36–43.
Bartels, M., de Geus, E. J., Kirschbaum, C., Sluyter, F., & Boomsma, D. I. (2003). Heritability ofdaytime cortisol levels in children. Behavioral Genetics, 33, 421–433.
Bentler, P. M. (1990). Comparative fit indexes in structural models. Psycholological Bulletin,107, 238–246.
Bentler, P. M., & Bonett, D. G. (1980). Significance tests and goodness of fit in the analysis ofcovariance structures. Psychological Bulletin, 88, 588–606.
Blyth, D., Hill, J., & Theil, K. (1982). Early adolescent’s significant others: Grade and genderdifferences in perceived relationships with familial and nonfamilial adults and youngpeople. Journal of Youth and Adolescence, 11, 425–450.
Booth, A. (1972). Sex and social participation. American Sociological Review, 37, 183–192.Booth, A. (1984). Response to scarcity. The Sociological Quarterly, 25, 113–124.Booth, A., Johnson, D. R., Granger, D. A., Crouter, A. C., & McHale, S. (2003). Testosterone and
child and adolescent adjustment: The moderating role of parent–child relationships.Developmental Psychology, 39, 85–98.
Boyce, W. T. (2006). Symphonic causation and the origins of childhood psychopathology. InD. Cicchetti & D. Cohen (Eds.), Developmental psychopathology, Vol. 2. Developmentalneuroscience (pp. 797–817). New York: John Wiley & Sons.
RELATIONSHIP QUALITY AND TRAIT CONTROL 257

Brandstadten, J., Baltes-Gotz, R., Kirschbaum, D., & Hellhammer, D. (1991). Developmentaland personality correlates of adrenocortical activity as indexed by salivary cortisol. Ob-servation in the age range of 35–65 years. Journal of Psychosomatic Research, 35, 173–185.
Caplan, R., Cobb, S., & French, J. (1979). White collar work load and cortisol: Disruption of acircadian rhythm by job stress? Journal of Psychosomatic Research, 23, 181–192.
Cichetti, D., & Rogosch, F. (2001a). Diverse patterns of neuroendocrine activity in maltreatedchildren. Development and Psychopathology, 13, 677–693.
Cichetti, D., & Rogosch, F. (2001b). The impact of child maltreatment and psychopathology onneuroendocrine functioning. Development and Psychopathology, 13, 783–804.
Collins, W., & Russell, G. (1991). Mother–child and father–child relationships in middlechildhood and adolescence: A developmental analysis. Developmental Review, 11, 99–136.
Conger, K., Conger, R., & Elder, G. (1994). Sibling relationships during hard times. In R.Conger & G. Elder (Eds.), Families in troubled times: Adapting to change in rural America(pp. 325–352). New York: Aldine.
Crouter, A., Helms-Erikson, H., Updegraff, K., & McHale, S. (1999). Conditions underlyingparents’ knowledge about children’s daily lives in middle childhood. Child Development,70, 246–259.
Cudeck, R., & Browne, M. W. (1992). Constructing a covariance matrix that yields a specifiedminimizer and a specified minimum discrepancy function value. Psychometrika, 57,357–369.
Dabbs, J. M., Jr. (1991). Salivary testosterone measurements: Collecting, storing, mailing sa-liva samples. Physiology and Behavior, 49, 8115–8117.
Diamond, J. (1999). Guns, germs, and steel: The fate of human societies. New York: Norton.Diamond, J. (2005). How societies choose to fail or succeed. New York: Norton.Dunn, J. (1983). Sibling relationship in early childhood. Child Development, 54, 787–811.Granger, D., Stansbury, K., & Henker, B. (1994). Preschoolers’ behavioral and neuroendocrine
responses to social challenge. Merrill-Palmer Quarterly, 40, 20–41.Gunnar, M., & Vazquez, D. (2001). Low cortisol and a flattening of expected daytime rhythm:
Potential indices of risk in human development. Development and Psychopathology, 13,515–538.
Heim, C., Ehlert, U., & Hellhammer, D. (2000). The potential role of hypocortisolism in thepathophysiology of stress-related bodily disorders. Psychoneuroendocrinology, 25, 1–35.
Hill, J., Holmbeck, G., Marlow, L., Green, T., & Lynch, M. (1985a). Menarcheal status andparent–child relations in families of seventh-grade girls. Journal of Youth and Adolescence,14, 301–316.
Hill, J., Holmbeck, G., Marlow, L., Green, T., & Lynch, M. (1985b). Parental status and parent–child relations in families of seventh grade boys. Journal of Early Adolescence, 5, 311–344.
Hu, I. T., & Bentler, P. M. (1995). Evaluating model fit. In R. H. Hoyle (Ed.), Structural equationmodeling (pp. 45–73). Thousand Oaks, CA: Sage.
Huston, T. L., & Robins, E. (1982). Conceptual and methodological issues in studying closerelationships. Journal of Marriage and the Family, 43, 901–925.
Johnson, D., & Elliot, L. (1998). Sampling design effects: Do they affect the analyses of datafrom the National Survey of Families and Households? Journal of Marriage and the Family,60, 993–1001.
Kenny, D. A., & Zautra, A. (2001). Trait–state models for longitudinal data. In L. M. Collins &A. G. Sayer (Eds.), New methods for the analysis of change (pp. 243–263). Washington, DC:American Psychological Association.
Kirschbaum, C., Steyer, R., Eid, M., Patalla, U., Schwenkmezger, P., & Hellhammer, D. (1990).Cortisol and behavior: 2. Application of a latent state–trait model to salivary cortisol.Psychoneuroendocrinology, 15, 297–307.
258 BOOTH, GRANGER, AND SHIRTCLIFF

Klimes-Dougan, B., Hasstings, P., Granger, D., Usher, B., & Zahn-Waxler, C. (2001).Adrencortical activity in at risks and normally developing adolescents: Individualdifferences in salivary cortisol response to social challenges, basal levels, and circadianrhythms. Development and Psychopathology, 13, 695–720.
Li, F., Duncan, S., Duncan, T., Yang-Wallentin, F., Acock, A., & Hops, H. (2001). Interactionmodels in latent growth curves. In G. Marcoulides & R. Schumacker (Eds.), Newdevelopments and techniques in structural equation modeling (pp. 561–614). Hillsdale, NJ:Erlbaum.
Luecken, L. J. (1998). Childhood attachment and loss experiences affect adult cardiovascularand cortisol function. Psychosomatic Medicine, 60, 765–772.
Maccoby, E. (2003). The gender of child and parent as factors in family dynamics. In A.Crouter & A. Booth (Eds.), Children’s influence on family dynamics: The neglected side of familyrelations (pp. 191–206). Mahwah, NJ: Lawrence Erlbaum.
McBurnett, K. M., Lahey, B. B., Rathouz, P. J., & Loeber, R. (2000). Low salivary cortisol andpersistent aggression in boys referred for disruptive behavior. Archives of General Psychi-atry, 57, 38–43.
McHale, S., & Crouter, A. (1996). The family contexts of children’s sibling relationships. In G.H. Brody (Ed.), Sibling relationships: Their causes and consequences (pp. 173–195). Norwood,NJ: Ablex.
Moffitt, T. E., Caspi, A., Rutter, M., & Silva, P. A. (2001). Sex differences in antisocial behaviour:Conduct disorder, delinquency, and violence in the Dunedin Longitudinal Study. Cambridge:Cambridge University Press.
Nesselroade, J. R. (1988). Some implications of the trait–state distinction for the study ofdevelopment over the life span: The case of personality. In P. B. Baltes, D. L. Featherman, &R. M. Lerner (Eds.), Life-span development and behavior. Hillsdale, NJ: LawrenceErlbaum.
Oosterlaan, J., Geurts, H., Knol, D., & Sergeant, J. (2005). Low basal salivary cortisol isassociated with teacher-reported symptoms of conduct disorder. Psychiatry Research, 134,1–10.
Pajer, K., Garner, W., Rubin, R., Perel, J., & Neal, S. (2001). Decreased cortisol levels in ad-olescent girls with conduct disorder. Archives of General Psychiatry, 58, 297–302.
Parke, R., & Buriel, R. (1998). Socialization in the family: Ethnic and ecological perspectives.In W. Damon (Series Ed.) & N. Eisenberg (Volume Ed.), Handbook of child psychology: Socialand personality development (pp. 463–552). New York: Wiley.
Petersen, A., Crockett, L., Richards, M., & Boxer, A. (1988). A self-report measure of pubertalstatus: Reliability, validity, and initial norms. Journal of Youth and Adolescence, 17, 117–133.
Plotsky, P. M., Thrivikraman, K. V., Nemeroff, C. B., Caldji, C., Sharma, S., & Meaney, M. J.(2005). Long-term consequences of neonatal rearing on central corticotropin-releasingfactor systems in adult male rat offspring. Neuropsychopharmacology, 30, 2192–2204.
Rigdon, E. E., Schumacker, R. E., & Wothke, W. (1998). A comparative review of interactionand nonlinear modeling. In R. E. Schumacker & G. A. Marcoulides (Eds.), Interaction andnon-linear effects in structural equation (pp. 1–16). Hillsdale, NJ: Lawrence Erlbaum.
Rosal, M., King, J., Yunsheng, M., & Reed, G. (2004). Stress, social support, and cortisol:Inverse associations. Behavioral Medicine, 30, 11–21.
Rosmalen, J. G. M., Oldehinkel, A. J., Ormel, J., de Winter, A. F., Buitelaar, J. K., & Verhulst, F.C. (2005). Determinants of salivary cortisol levels in 10–12 year old children; a population-based study of individual differences. Psychoneuroendocrinology, 30, 483–495.
Russell, A., & Saebel, J. (1997). Mother–son, mother–daughter, father–son, and father–daugh-ter: Are they distinct relationships? Developmental Review, 17, 111–147.
Sapolsky, R. (2005). The influence o f social hierarchy on primate health. Science, 308, 648–652.
RELATIONSHIP QUALITY AND TRAIT CONTROL 259

Schaefer, E. (1965). Children’s reports of parental behavior: An inventory. Child Development,36, 413–424.
Schwarz, J., Barton-Henry, M., & Pruzinsky, T. (1985). Assessing child-rearing behavior: Acomparison of ratings made by mother, father, child, and sibling on the CRPBI. ChildDevelopment, 56, 462–479.
Shirtcliff, E. A., Granger, D. A., Booth, A., & Johnson, D. (2005). Low salivary cortisol levelsand externalizing behavior problems. Development and Psychopathology, 17, 167–184.
Steinberg, L. (1981). Transformations in family relations at puberty. Developmental Psychology,17, 833–840.
Steinberg, L., & Silk, J. (2002). Parenting adolescents. In M. Borstein (Ed.), Handbook of par-enting: Vol. 1. Children’s parenting (2nd ed., pp. 103–133). Mahwah, NJ: Lawrence Erlbaum.
Steyer, R., Eid, M., & Schwenkmezger, P. (1997). Modeling the intraindividual change: Truechange as a latent variable. Methods of Psychological Research Online, 2, 1–33.
Stocker, C. (1994). Children’s perceptions of relationships with sibling, friends, and mothers:Compensatory processes and links with adjustment. Journal of Child Psychology andPsychiatry, 35, 1447–1459.
Stocker, C., Burwell, R., & Briggs, M. (2002). Sibling conflict in middle childhood predictschildren’s adjustment in early adolescence. Journal of Family Psychology, 16, 50–57.
Taylor, S., Klein, L. C., Lewis, B. P., Gruenewald, T. L., Gurung, R. A. R., & Updegraff, J. A.(2000). Biobehavioral response to stress in females: Tend and befriend, not fight-or-flight.Psychological Review, 107, 411–429.
Thompson, L., & Walker, A. (1989). Gender in families: Women and men in marriage, work,and parenthood. Journal of Marriage and Family, 511, 845–871.
Tucker, L., & Lewis, C. (1973). A reliability coefficient for maximum likelihood factor analysis.Psychometrika, 38, 1–10.
van Bakel, H. J., & Riksen-Walraven, J. M. (2004). Stress reactivity in 15-month-old infants:Links with infant temperament, cognitive competence, and attachment security. Devel-opmental Psychobiology, 44, 157–167.
Zahn-Waxler, C., Crick, N., Shirtcliff, E. A., & Woods, K. (2006). The origins and developmentof psychopathology in females and males. In D. Cicchetti & D. Cohen (Eds.), Developmentalpsychopathology, Vol 1. Theory and method (pp. 76–138). New York: John Wiley & Sons.
260 BOOTH, GRANGER, AND SHIRTCLIFF