gastrointestinal physiology food …people.upei.ca/bate/motility.pdfgastrointestinal physiology...
TRANSCRIPT

DIGESTIVE PHYSIOLOGY
GASTROINTESTINAL PHYSIOLOGY Ingested foodstuff contributes to an increase in the overall weight of the animal, but does not become an integral part of the structure or metabolic activities of the animal, until it is absorbed somewhere through the digestive tract. What is not absorbed is secreted in the faeces as waste. The gastrointestinal tract constitutes a machine through which all food has to pass. The first step in the use of such a machine is to acquire, chew and swallow the food (Fig.1). Animals achieve this basic step through the process of prehension. Depending on the species and age, different animals use different modalities to ingest food. There are specific anatomical adaptations that permit an animal tingest food in a given manner (Fig. 2).
o
Some animals use the lips to prehend food while others utilize the tongue (Fig. 3). Similarly, some species require significant mastication prior to swallowing, while others, such as most ruminants, masticate very little prior to first swallowing. This strategy permits collecting a significant volume of food in the rumen, and then, they start a process of rumination. This process consists of regurgitating portions of the ingested food to thoroughly masticating it and reducing the particle size; this activity is normally conducted with the animal resting in a recumbent position.
FOOD INGESTION
• PREHENSION
• MASTICATION/CHEWING
• DEGLUTITION/SWALLOWING
PREHENSION
• MECHANISM VARIES GREATLY WITH SPECIES– HORSES USE LIPS TO MANIPULATE AND
TEETH TO RIP– CATTLE USE TONGUE TO MANIPULATE
AND LOWER TEETH TO RIP– CARNIVORES USE INCISORS TO CUT
AND CANINES TO RIP– POULTRY USE BEAK OR BILL
Figure 2. Some mechanisms of prehension
Figure 1. Stages of food ingestion
PREHENSION BY CATTLE
• Use the tongue to manipulate food into mouth
• Teeth and hard palate to press and tear
• Mobile lower teeth
Figure 3. Prehension mechanism used by cattle
Carnivores do not significantly masticate their food and they depend on a very aggressive enzymatic digestion to reduce particle size. They masticate their food enough to reduce the size in order to be able to swallow it. Furthermore, avian species do not have teeth to masticate (Fig. 4). They depend on production of mucus to coat the particles prior to swallowing, and then, they macerate the food
1

within a specialized organ that may contain hard materials, such as pebbles to help in the reduction of particle size.
MASTICATION
• VERY IMPORTANT IN HERBIVORES
• NOT IMPORTANT IN CARNIVORES
• NON EXISTING IN AVIAN SPECIES
Figure 4. Role of mastication in different species
Herbivores masticate their food not only to reduce the size of the particles but also to break the cellular wall and permit the access of digestive enzymes. It also reduces the incidence of damage to the walls of the gastrointestinal tract (GIT) (Fig. 5). Once the food has been masticated, it has to be swallowed. This process requires an initial voluntary stage which is to move the bolus towards the back of the oral cavity. Then, an involuntary reflex is activated which permits the passage of the bolus from the oral cavity to the esophagus without entering the trachea and the respiratory system (Fig. 6). During most of the time, the animal maintains an open passage between the nose or mouth to the trachea, thus permitting the passage of air into the lungs and minimizing the entrance of air into the digestive system (Figs. 7, 8). When there is food to be swallowed, a passage from the mouth to the esophagus has to be created to permit the passage of food. This is achieved by the elevation of the soft palate, which prevents the entrance of food into the nasal cavity and by the movement of the epiglottis backwards, thus blocking the
entrance to the trachea and opening the passage towards the esophagus. All these changes, which take place in a very rapid sequence are involuntary and constitute the process of deglutition (Fig. 9). During deglutition, the respiratory process is temporarily suspended.
MASTICATION IN HERVIVORES
• REDUCE PARTICLE SIZE– INCREASE SURFACE AREA– BREAKS CELLULOSE MEMBRANE
• LUBRICATE FOR SWALLOWING• MIX WITH SALIVA• EXPOSE TO DIGESTIVE ENZYMES• REDUCE EXCORIATION OF GIT
Figure 5. Role of mastication in hervivores
DEGLUTITION• ONE VOLUNTARY INITIAL STAGE
– MOVEMENT OF BOLUS TO BACK OF ORAL CAVITY
• A PHARYNGEAL STAGE (INVOLUNTARY)– PASSAGE FROM ORAL CAVITY TO
ESOPHAGUS• ESOPHAGEAL STAGE (INVOLUNTARY)
– PASSAGE OF FOOD INTO THE STOMACH
Figure 6. Steps involved in deglutition
SOFT PALATE
NASOPHARYNX
EPIGLOTTIS
ESOPHAGUS
GLOTTIS
TRACHEA
Figure 7. Anatomical components involved in deglutition
2
RESPIRATION
Figure 8. Lateral view of the air pdurin
assage g respiration
DEGLUTITION
Figure 9. Sequence of anatomical changes taking place during deglutition
3
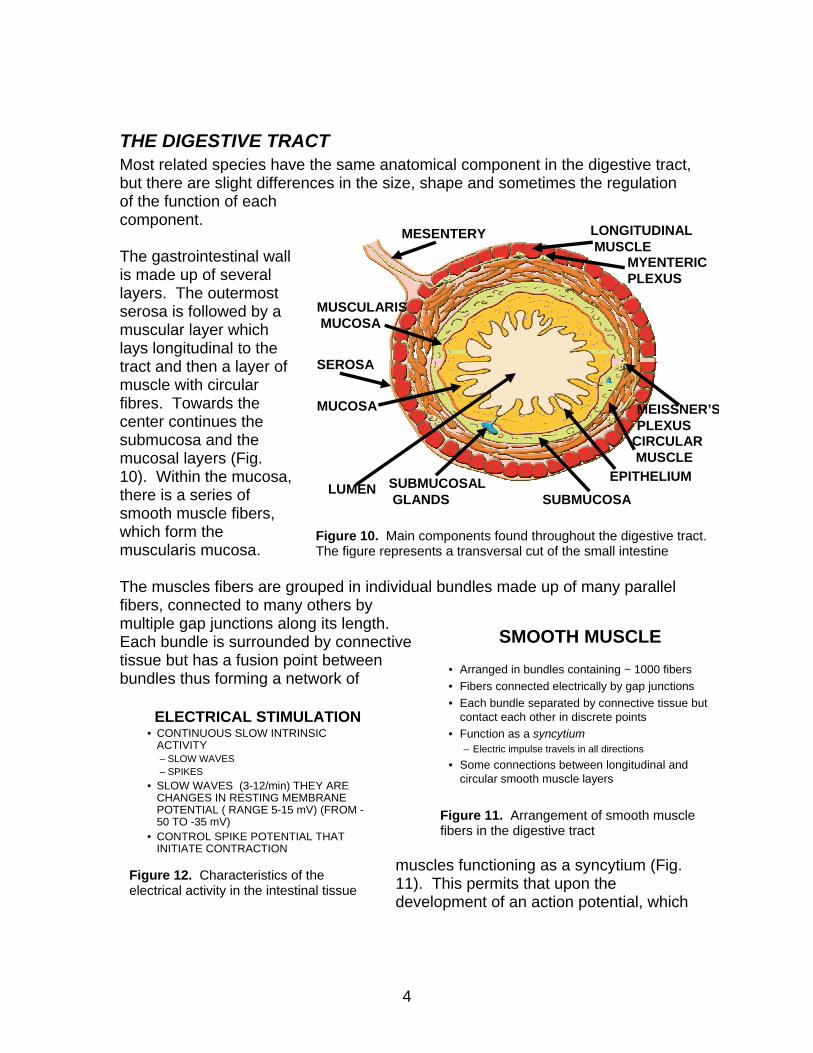
THE DIGESTIVE TRACT Most related species have the same anatomical component in the digestive tract, but there are slight differences in the size, shape and sometimes the regulation of the function of each component.
LUMEN
MUCOSA
MUSCULARISMUCOSA
MESENTERY
SUBMUCOSALGLANDS SUBMUCOSA
CIRCULARMUSCLE
LONGITUDINALMUSCLE
SEROSA
EPITHELIUM
MYENTERICPLEXUS
MEISSNER’SPLEXUS
Figure 10. Main components found throughout the digestive tract. The figure represents a transversal cut of the small intestine
The gastrointestinal wall is made up of several layers. The outermost serosa is followed by a muscular layer which lays longitudinal to the tract and then a layer of muscle with circular fibres. Towards the center continues the submucosa and the mucosal layers (Fig. 10). Within the mucosa, there is a series of smooth muscle fibers, which form the muscularis mucosa. The muscles fibers are grouped in individual bundles made up of many parallel fibers, connected to many others by multiple gap junctions along its length. Each bundle is surrounded by connective tissue but has a fusion point between bundles thus forming a network of
muscles functioning as a syncytium (Fig. 11). This permits that upon the development of an action potential, which
SMOOTH MUSCLE• Arranged in bundles containing ~ 1000 fibers• Fibers connected electrically by gap junctions• Each bundle separated by connective tissue but
contact each other in discrete points• Function as a syncytium
– Electric impulse travels in all directions• Some connections between longitudinal and
circular smooth muscle layers
Figure 11. Arrangement of smooth muscle fibers in the digestive tract
ELECTRICAL STIMULATION• CONTINUOUS SLOW INTRINSIC
ACTIVITY– SLOW WAVES– SPIKES
• SLOW WAVES (3-12/min) THEY ARE CHANGES IN RESTING MEMBRANE POTENTIAL ( RANGE 5-15 mV) (FROM -50 TO -35 mV)
• CONTROL SPIKE POTENTIAL THAT INITIATE CONTRACTION
Figure 12. Characteristics of the electrical activity in the intestinal tissue
4

is generated at some point, it can move in all directions. Furthermore, there are connections between the longitudinal and circulatory layers which permit the synchronization of movements to mix or propel the intestinal content. To control the tone and the motor activity of the gut, the membrane of the smooth muscle is continuously stimulated by intrinsic electrical activity in the form of slow waves or spikes. Variation in the frequency of the membrane potential changes creates the rhythm of the slow waves (range from 3-10/min). What ultimately changes to create the slow waves is the resting membrane potential (Fig. 12).
SPIKE POTENTIALS• LAST 10-20 msec• TRUE ACTION POTENTIAL• TRIGGERED WHEN MORE + THAN
-40mV• THE HIGHER THE SLOW WAVES RISE
MORE FREQUENT SPIKE POTENTIALS OCCUR (1-10 /sec)
• TRIGGERED BY Ca+ INFLUX THROUGH CALCIUM-SODIUM CHANNELS– SLOWER THAN SODIUM CHANNELS OF
NERVE FIBRES
Figure 13. Conditions required to trigger a spike potential
To trigger a spike the resting membrane potential of the smooth muscle has to be larger than -40 millivolts. These spikes trigger a true action potential, which in turn controls contractile activity. The frequency of the spike potential will increase when the slow wave resting membrane potential is higher (Figs. 13, 14). Therefore, the more depolarized the membrane becomes the more excitable it is and has more chances of contractile activity. The mechanism of achieving this higher membrane potential is presumed to be through influx of Ca++ through calcium-sodium channels as oppose to the faster sodium channels used by nerve fibres
MEMBRANE POTENTIAL
0 6 12 18 24 30 36TIME (sec)
MEM
BR
AN
E PO
TEN
TIA
L (m
V)
-10-20-30-40-50-60-70
Figure 14. Characteristics of different membrane potentials
FACTORS HYPERPOLARIZING MEMBRANES
(membranes potential more negative)• Norepinephrine or epinephrine on fibre
membrane• Sympathetic stimulation
– Secreting norepinephrine
Figure 15. Factors making membrane potential more negative
5

Several factors contribute to the depolarization of the fibres membranes. Physical distension (stretching) of the muscle, usually due to the presence of food in the tract; acetylcholine stimulation as well as, paraneurone
s secreting acetylcholine, and finally, sevgastrointestinal hormones (Fig. 15
sympathetic stimulation by
eral ).
he counterparts of this process are the
se are
egulation of all contractile and s
the s
of
muscle
d
).
y the
FACTORS DEPOLARIZING MEMBRANES(membrane potential less negative)
• STRETCHING• ACETYLCHOLINE• PARASYMPATHETIC STIMULATION
– MEDIATED THROUGH ACETYLCHOLINE• SPECIFIC GASTROINTESTINAL
HORMONES
Figure 16. Factors making membrane potential less negative
• Lies in wall of gut• Extend from esophagus to anus• As many neurones as the spinal cord• Made of two plexuses
– Myenteric or Auerbach’s plexus• Situated between long and circ muscle layer
– Submucosal or Meissner’s plexus• Situated in the sub mucosal layer
Figure 17. Characteristics of the enteric nervous system
ENTERIC NERVOUS SYSTEM
Tfactors that contribute to the hyper-polarization of the membranes. Thethe effects of nerepinephrine or epinephrineon the membranes and sympathetic stimulation whose nerves secrete norepinephrine (Fig. 16).
ENTERIC NERVOUS SYSTEM
• MYENTERIC PLEXUS CONTROLS GASTROINTESTINAL MOVEMENT
• SUBMUCOSAL PLEXUS CONTROLS SECRETIONS AND BLOOD FLOW
• INNERVATED BY SENSORY NEURONES FROM THE EPITHELIUM
• CONNECTED TO PARASYMPATHETIC AND SYMPATHETIC SYSTEM
Figure 18. Role of the enteric nervous system
R ecretory activity is supported by an independent nervous system imbedded in the wall of gut. This system is called the enteric nervousystem and it extends the length of the GIT starting in the esophagus. The total numberneurones is as large as those found in the spinal cord. The enteric nervous system is made up of two plexuses. One located between the longitudinal and circulatory layer called the Myenteric or Auerbach’s plexus. The other is founin the sub mucosal layer and is called the sub mucosal or Meissner’s plexus (Figs. 10, 17, 18These two are interconnected in order to
6
synchronize GIT function. Motility of the gut is primarily controlled bmyenteric plexus, while thesecretory activity and blood flow are controlled mainly
LONGITUDINAL CIRCULAR SUBMUCOSA
VILLUS
Figure 19. Innervations of the intestinal muscle by the enteric nervous system, sympathetic and parasympathetic

by the sub mucosal plexus. The enteric system is capable of controlling gastrointestinal independent of other body functions. However, on top of being connected between themselves, the two plexuses are further innervatedsympathetic and parasympathetic systemThis innervation permits overall inhistimulation of the gastrointestinal activities (Fig. 19).
by the s.
bition or
he mechanisms through which the ty
. It
d
imulation
in
lthough most of the activities of the ture,
h
st
s
d
he sub mucosal plexus in turn controls ity
large number of neurotransmitters (Fig. ,
determined.
Tmyenteric plexus control motor activithroughout the gut are multiple (Fig. 20)can increase the tonic contraction of the intestinal wall, as well as, the intensity anthe rate with which the rhythmical contractions take place. Finally, stof the myenteric system can result in faster conduction of excitatory waves along the intestinal wall and, as a result an increaseperistaltic movement. Amyenteric system are excitatory in nathere are also some inhibitory elements. These are believed to be mediated througthe release of peptides with vasoactive intestinal polypeptide (VIP) being the moplausible candidate. The action of these peptides appear to be focused in inhibitingsphincter activity (pyloric and ileocecal), thumodifying the rate of passage of food into the respectively.
• CONTROL MOTOR ACTIVITY ALONG THE GUT– INCREASE TONIC CONTRACTION OR “TONE” OF
GUT WALL– INCREASE INTENSITY OF RHYTHMICAL
CONTRACTIONS– INCREASE RHYTHM– INCREASE VELOCITY THUS MORE PERISTALSIS
• HAS SOME INHIBITORY NEURONES– SECRETE VIP
• THEY CONTROL SPHINCTER ACTIVITY
Figure 20. Role of the myenteric plexus
SUBMUCOSAL PLEXUS
• CONTROL FUNCTION OF SMALL SECTIONS OF THE INTESTINE
• SIGNAL FROM EPITHELIUM TO SMPCONTROL– LOCAL SECRETION– LOCAL ABSORPTION– LOCAL CONTRACTION
• REGULATE FOLDING OF GASTROINTESTINAL MUCOSA
Figure 21. Role of the submucosal plexus
MYENTERIC PLEXUS
uodenum and cecum
TNEUROTRANSMITTERS
• Acetylcholine– Mainly excites GI activity
• Norepinephrine– Usually inhibits GI activity
• Adenosine triphosphate (ATP)• Serotonin• Vasoactive intestinal polypeptide (VIP)• Somatostatin (SS)• Leu-enkephalin• Met-enkephalin• Bombesin
Figure 22. Neurotransmitters involved in the regulation of gastrointestinal activity
secretory, absorptive and contractile activin very small discrete sections of the GIT (Fig. 21). Usually a signal is generated bysensors in the epithelium of the gut and thisis conveyed to the sub mucosal where the appropriate response is prepared and implemented. A22) have been identified as participating inthe regulation of the gastrointestinal activitybut their specific roles are yet to be
7

The overall regulation of gastrointestinal function is based on reflexes controlled
second type of reflexes transmit information nd require
the
he third type of reflex involves a larger
h
n
es
n,
s hinted earlier, many hormones influence
of l
denum
by the enteric, sympathetic and parasympathetic systems. These are (Fig. 23): Reflexes which are integrated within the enteric system and are responsible for secretion, as well as, food mixing and peristalsis of the GIT.
GASTROINTESTINAL REFLEXES• INTEGRATED TOTALLY WITHIN THE
ENTERIC SYSTEM AND CONTROL– SECRETION, PERISTALSIS, LOCAL MIXING
• GUT-SYMPATHETIC GANGLIA-GUT– GASTROCOLIC REFLEX
• PROMOTES COLON EVACUATION– ENTEROGASTRIC REFLEX
• PREVENTS STOMACH MOTILITY AND EMPTYING
– COLONOILEAL REFLEX• INHIBITS ILEAL EMPTYING
• GUT-SPINAL CORD/BRAIN-GIT– USES VAGUS NERVE– FROM STOMACH AND DUODENUM
• CONTROL GASTRIC MOTOR AND SECRETORYACTIVITY
– PAIN REFLEXES• INHIBIT GASTROINTESTINAL ACTIVITY
– DEFECATION REFLEX• PROMOTES COLONIC RECTAL AND
ABDOMINAL CONTRACTION
Figure 23. Reflexes of the gastrointestinal tract
A through long distances in the GIT athe participation of the gut as the starting point, integration in the sympathetic ganglia and response by another section of the gut. Among these, we find the gastrocolic reflex which is responsible for the evacuation of the colon. The enterogastric reflex involves the participation of the small intestine and colon sending signals which reduces stomach motility and its secretion. The colonoileal reflex signals from the colon to reduce or prevent emptying the contents of the ileum into the colon. Ttrajectory from the gut to the spinal cord and/or brain and back to the GIT. Througthis type of reflex the duodenum, based on the rate at which it receives food, is capableof controlling gastric motor and secretory activity. In addition, reflexes that have paiinvolved use these mechanisms to reduce overall gastrointestinal activity. Finally, faecaccumulation in the colon trigger the defecationreflex via signals sent to the spinal cord and back with the resulting contraction of the colorectum and abdomen in order to defecate.
ENDOCRINE CONTROL OF MOTILITY
• Cholecystokinin– Secreted by I cells of duodenum and jejunum
mucosa– Response to FA, monoglycerides in intestinal
content– Increase contractility of gallbladder
• Secretion of bile into small intestine• Emulsifying fat
– Inhibits stomach motility– Slow down transit
• Gastric inhibitory polypeptide– Secreted by mucosal of upper intestine– Response to fat and protein principally– Slow down stomach emptying– Allow proper digestion of intestinal content
Figure 24. Hormones influencing motility of different component of the GIT
Adifferent aspects of motility of the GIT and some influence both, motility and secretion the GIT (Fig. 24). Cholecystokinin is a powerfucompound secreted by I cells of the duoand jejunum which, in response to the presence of fat digestion by-products fatty acids and monoglycerides, triggers contraction and emptying of the gallbladder thus making available bile salts in the small intestine in order to emulsify the fats coming from the
8

stomach. At the same time, cholecystokinin inhibits stomach motility, threducing the rate of stomach emptying into the duodenum and permitting bedigestion. A similar effect is seen when fat and proteins are being emptied into the duodenum and upper jejunum. To this end, the mucosa releases a compound known as gastric inhibitory polypeptide which slows down the rastomach emptying.
us tter
te of
arger ration
t id
of
FUNCTIONAL MOVEMENTS WITHIN THE GIT
Figure 25. Deposition of food in layers in the stomach
The stomach carries out a significant amount of movement in order to mix its content and to separate physically large particles into smaller portions. As the food enters the stomach it starts forming layers, with the deepest layers being the smaller particles, ready to be emptied into the duodenum, and those close to the esophagus being the lparticles which need significant disintegbefore moving to lower layers (Fig. 25). Mosof the digestion is done chemically by acand/or enzymes but this process is significantly supported by the movementsthe stomach which is performed in a
9
PERISTALSIS
Figure 26. Sequential movement of a primate's stomach that permit mixing of ingesta and advance of digested materials towards the duodenum

repetitive manner following each time a similar pattern. This pattern changesfrom species to species, and depends on the specific anatomical designorgan. Figure 26 depicts the pattern of movement of a human stomach. The stomach has significant movement to mix and initially digest the ingesta. In oto properly digest the food and discard what is indigestible, the GIT has to mthe intestinal contents along the tract, but at the same time, provide opportunity for proper digestion. Two types of movements within the GIT; the propulsive movements and the mixing movements are used to achieve this (Fig. 27).
of the
rder ove
he
FUNCTIONAL MOVEMENTS OF THE GIT
• PROPULSIVE MOVEMENTS– MOVE CONTENTS ALONG THE TRACT– RATE PERMITTING DIGESTION
• MIXING MOVEMENTS– MIX CONTENT
• FACILITATE ENZYMATIC DIGESTION– INCREASE EXPOSURE TO EPITHELIAL
WALL• FACILITATE DIGESTION AND ABSORPTION
Propulsive movements push contents along ttract and these are implemented by a coordinated effort of longitudinal and circular muscles of the gut (Fig. 28). The rate at which the contents are moved influences the degree of digestion taking place, with a longer retention translating in a more thorough digestion process. Propulsive movements take place in waves in which consecutive sections of the intestine reduce their diameter forcing the intestinal content to move in the direction of the anus. Mixing movements do not displace food along the tract. They consist in the constriction of relatively close sections of the intestine forcing a change in position of the content, as well as to facilitate
phydisinthem(Fig
MIXING MOVEMENTS (Segmentation)PERISTALSIS
Figure 29. Pattern of segmentation or mixing peristaltic movements of the GIT
Figumove
10
Figure 27. Propulsive and mixing movements of the gut permit mixing anddisplacement of the digesta through the GIT
xpose
sical separation and tegration of particles to e better to digestive enzymes
. 29).
PROPULSIVE MOVEMENTS
PERISTALSIS
re 28. Pattern of propulsive peristaltic ments of the GIT