gaba in the
TRANSCRIPT

Loll ¡e,-
GABA IN THE GUINEA-PIG ENTERIC NERVOUS SYSTEM
A thesís submítted fon tVrc degtee of
Doeton of PhiLosoPhY
by
ANTHONY KRANTIS, B.Sc. (Hons)
Department of PhYsiology'
UniversitY of Adelaide'
Adelaide, South Australia' 5000.
November,1981.
ât'vru dt ,f /l- I 'f I

, To my uife Sandra
uithout utlnse l.o'oe, suPPoYtJ
enc ourü.g ement, under s tantding
and pat'ience, this uas not possibLe"

I
DECLARAT I ON
I declare that this thesis.is a record of original work and
contains no material which has been accepted for the av¡ard of any
other degree or dip'loma ìn any University.
To the best of my knowledge and belief, the thesis contains no
materìal previously published or written by another persotl, except when
due reference is made in the text of the thesis.
Anthony Kranti s.
November, 1981.

1l
ACKNOþlLEDGEMENTS
I am deeply'indebted to my supervisor Dr D. I. B. Kerr without
whose guidance, knowledge, critical discussion, keen eye and friendshìp
this v¡ould not have been possìbìe.
To Dr Barbara Dennjs I am deepìy gratefu'l for"her assistance in the
development and appl'icat'ion of the autorad'iographic procedures, and the
editing of this thesis.
Mr Graenre Langsford wÍlljngly gave of his time bo assist in the
development of some experiments and ensured a continued supply of the
necessary equipment. I am thankful for his assistance.
I thank Dr M. Roberts for his valuable advice in chemicaj matters;
Steve Robjohns for his assistance with technical proble¡ns and Greg p.ike
for his he'lpfu'l discussion and friendship.
Finaì1y, I thank Mrs Karen Janles who was so pat'ient in her assistance
with the typíng of this thesis.

TABLI OF CONTENTS
DECLARATI ON
AC KNOl^lLEDGEI{ENTS
SUMMARY
CHAPTER i: GINERAL INTRODUCTION
Historg
Bioehemistry
PLtysioLcgy and PVtnrrnacoLr.tgy of GABA
GABA in PeripheraL Neruous System
GAtsA in Enteric Neraous System
The Entev,ic Nez'oous System
Fm.cti onal 0rg øní zati on
Mye'n'berLc PLerus
Myentez.Lc Neurortes
Subrntcous PLeæus
Eætz.ins'ic Innezwation of the Enteric Pleæus
Intrinsi,a Innez'uatíon of the EnterLc PLeæus
CI1APTER II: PHARMACOLOGY OF GABA ACTIONS
IntroCa,tction
MateriaLs ond Method.s
ResuLts
Díseussion
Paqe No.
l1
4
5
7
I10
t2
L2
14
15
16
16
1B
26
28
29
32
1

CHAPTER I I I: LOCALIZATION OF
SITES BY AUTORADIOGRAPHY
Introduet¿on
Methods
Autoz,adiography
Staining
Results
t3Hl-eRgR HrGH AFFiNTTY uPTAKE
rnyenter,íe
ß-a'Lanine
on lSnl-
of
Page No.
37
40
4?
43
43
43
45
46
47
48
56
57
59
60
63
65
66
uptake of lsul proLine and tsal Leuc'Lne i'nto
Løninar preparations.
Myenteti,e LoeaLízation of l7ul-Caaa ar¿ri lSnl
in Løm)nar prep tions,
Effects of L-DABA, nípecotic acid and ACHC
GÆA uptake,
Autogn adío gz,aphi e Lo caLí zati ott o f 13 nl- cnn n
coLon m.d iLewn,
ùLscussion
CHAPTER IV: M0RPH0L0GY 0F MYTNTERIC NEUROIIIES
Introduction
Methods
ResuLts
ùLscussion
CHAPTER V: UPTAKE AND RELEASE 0F t3Hl-eRAn tsY MYTNTERIC NEUR0NES
fntroduction
Methods
rdentifùeation of lsnl-ca%A v,eLeased by stimuLation

ResuL'bs2
uptake and z'eLease of Í"Hl-qABA.
CaLeiun tlepend.enee of stirn'LLated. Í7al-cmn reLease.z
Identifica'bion of l"nl-CIBA teLeased by eLectricaL
stimulation.
Díseussion
CHAPTER VI: THE EFFECTS OF GABA ANTAGONISM ON INTESTINAL MOTILITY
fntroduction
Mqtev"LaLs and. Method,s
ResuLts
Discussíon
CHAPTËR VII: MYENTERIC PURINERGIC NERVES
Introduetíon
Iúethods
ELe ctri caL s tinuLati ott.
ResuLts
Responses of the distaL eoLon to AI'P.
Responses of t?ø iLeunt to ATP
D¿seussion
Distal coLon
-L Leutn
CHAPTER VIII: GENERAL DISCUSSION A.ND CONCLUSION
Page l,lo.
67
67
69
69
70
73
7B
79
B1
86
91
92
93
93
95
97
97
9B
r02
.r'
BIBLIOGRAPHY 108

CORRIGENDA
P8, line 10: delete 'actions'.
P10, para 2; last line should read: 'but this idea has never been subjectedto further study'.
P19, ljne 17 should read: 'half of the myenterìc neurones show'.
P27, para 2, last line: Insert reference (0lsen et aL., 1978).
P30, pa ra 2, l i ne 1 : del ete ' bi ndi ng' .
para 2, last sentence: Insert reference (Breuker & Johnston, 1975).
Chapter II. Fi9s.3,5,9,11,12, abscìssa labels should read:'-log dose g/ml ' .
Fig. 7, legend, add: 'stimulus period indicated by bar'.
Chapter III. Figs. 12,13, legends, add new second sentence: 'Broken linesdel i neate gangl i a' .
P70, para 1, last sentence should read: 'since ß-althroughout, i s a more ef fecti ve 'inhi bì tor of gGABA-uptake (Browery et aL., . ), yet only
ani ne, whiI ial than
ch was presentof neuronal
P7L, para 3, line 7, should read: 'the more usual criterion of calciumdependence fails when veratridine is used as a stimulus'.
P80, para 2, last sentence, de'lete: 'coupling'.para 3, first sentence should read: 'Following addition of b'icuculline....'
P84, para 3, last sentence, should read: '... GABA4 receptors ordesensitization of GABAB responses . . ..' .
P95, para 2:'Fig. 6a' should read'Fig. 6b' and, last l'ine'Fig. 6b' shouldread 'Fj g. 6a' .
P105, para 2, should read: ... 'antagonists are difficult to measure incentral nervous preparations .. ..' .


2
In Chapter IV the development of a relativeìy sìmp1e and rap'id
staining procedure is described, wh'ich affords good morpl'rology of neurones
in laminar preparations. This method allows deliniatio;l of nerve types?1
accumulating ["H]-GABA, as shown by autoradìography. ['H]-GABA labelled
onìy a restrjcted population of myenterìc neurones.
Chapter V describes the investigatìon of the evoked release criterìori,
using chromatographic and radiochemical analysis. Electrically evoked t3ffj-
GABA release from laminar preparations cf the guìnea-pìg ileum under various
treatments, inclr-rdjng neurona'l and gliaì hìgh af'f,inity uptake blockerso
tetrodotox'in, and calcium free medium, shon/ [3tt].-engn is released by
myenteric neurones in a transmitter-l'ike manner.
In Chapter VI current concepts of the mechan'isms of peristalsjs are
discussed, together with results of experinrents jn whjch GABA-antagonìsm
interfered wjth perista'lsis, as measured'in jsola.ted preparations of ihe
guinea-p'ig dìstal colon. The results suggest important'inrpìicatìons for
studies of intestinal motiì ity.
Acetylchol jne is the transm'itter of the exc'itatory nerves stimulated
by GABA but the transmitter released by the inhibitory nerves is not knouln,
although it has been proposed to be adenosjne 5'-triphosphate (ATp). In
Chapter VII pharmacoìogìca'l jnvestìgat'ion of this hypothesis usíng a new
class of ATP antagonist is described. Furthermcre, the involvement of
ATP in transmission from non-cholinergìc excìtatory nerves was invest'igated.
ATP is not the transrnittr:r of these nerves.
Chapter VIII is an overview of the whole thesis and discussion of
the results'in regard to the various criteria for identificatjon of a
neurotransm'itter substance. Conclusions and suggestions for future

3
research are also presented. It is concluded that GABA js a neurotrans-
mitter in the guinea-pig myenter"ic plexus.

CHAPTER I
GENERAL INTROÐUCTION

4
GENERAL INTRODUCTION
Histot y
Since its synthesis in 1883 (Schotten, lEB3) gamma-anrinobutyric acid
(GABA) has been confjrmed to be an inhibjtory neurotransmi-uter in vertebrates
(0bata et aL., L967;Obata and Takedo, 1969; Curtis and Johnston, 1974) and
invertebrates (Kufflen and Edlvards, 1958; Trigg'le and Triggle, I976). The
role of 0ABA as a major neurotransmitter in the peripheral and central
nervous systems of these tu¡o 'phyl a' 'is now general ly acceptecl (for revì evls
see: Curtis and llJatk'ins, 1965; Curt.is and Johnston, I974; Inversen et; a.L.,
L975; Roberts et aL., 1976 ) . In addi ti on to i ts docunlented i nhi bi tory ro] e
there is also evicience that GABA may functjon as an excitatory transmjtter
in at least some invertebrate and vertebrate preparations (Kerkut and lrla'lker,
1962; Gerschonfeld and Lasansky, L964; hjalker et aL., I97l;Obata, 1976)..
Until recent'ly there have been, however, surprìsingìy fevr investigations of
any GABA functions'in periphera'l tissue although GABA has been observed to
have pharmacological actjons in mammal'ian jnLestjne (HobbìEer, 1958a, b;
F'lorey anC lt1cJ-ennan , 1959; Irrouiye et aL., 196]- ; Takahashi et aL. , 1961a , b;
Bianchi et aL., 1968; Lewis et aL., L972). Evjdence has accumulated
suggesting a neurotransm'itter role for GABA in t.he mammaìjan enteric nervous
system, where appìied GABA stimulates myenteric neurones (Grafe et aL., 1979;
Krantis et a.L., 1980). GABA (10 nmol/g) and its synthezìng enzynre glutamic
acid decarbor.ylase (GAD) are reported to be localized io myenterìc neurL\nes
of the guinea-pig taerria coli. Furthernrore a small proportjon of neurones
jn the myenteric p'lexus were shown to possess a hìgh affinity uptake
mechanisrn for GABA (Jessen et aL., !97ga). In light of the ub'iqu'itos
nature of GABA withjn central and peripheral neural tissue, together with
the recent finings as to ìts pharrnacologìca'i effects and apparent neuronal
local izatic.¡n wíthin the guìnea-pig in'Lestìne, the work deta'iled in this

5
thesis sought to extend our knowledge about GABA and jts funct'ion jn the
guinea-pig enteric nervous system.
Bíochemistry
GABA is synthesised to a large extent from glutam'ic acjd by the act'ion
of the enzyme g'lutamic-ac'id decarboxyl ase (GAD) . 0ther synthet'ic pathulays
for GABA utìlising different precursors and enzymes have recently been
shown to exist (Roberts, 1981), hcwever thejr contributjon to the GABA
neurotransmitter pool rema'ins to be defermined. GAD ís pt"esent jn non-
neuronal tissue but is of low act.'irlity compared to GAD present in central
nervous tissue (Baxter, 1970) where jt is rna'inly concentrat,ed in synapto-
somal fractjons (nerve terminals) (lnJu, 1976; Barber and Saito,1976; Wood
et aL., L976).
GABA 'is degraded by GABA-transaminase (GABA-T) which in the central
nervous system i s found chì ef 'ly 'in the mi tochondria (Neal and Iversen , 1969;
van rlen Berg et aL., 1975). The transamination of GABA forms succinic semi-
aìclehyde which is then oxidized to succin'ic acid r"rhich in turn can be
oxidized by the reactions of the tricarboxyìic acid cycle (Chase and'r¡lalters,
1976; van den Berg et aL.,1975). Glutamic acid formed by these reactions
may then be converted to glutamìne by the act'icn of glutamìne synthetase
(GS) anC shunted i.o the presynapt'ic nerve terminal where it is reconverted
by phosphal,e-activated glutantinase back to glutanlate fcr subsequent conversion
to GABA (neurotransmitter pool ) or enter the trìcarboxyì jc acìd cy'lce (Kvanrme,
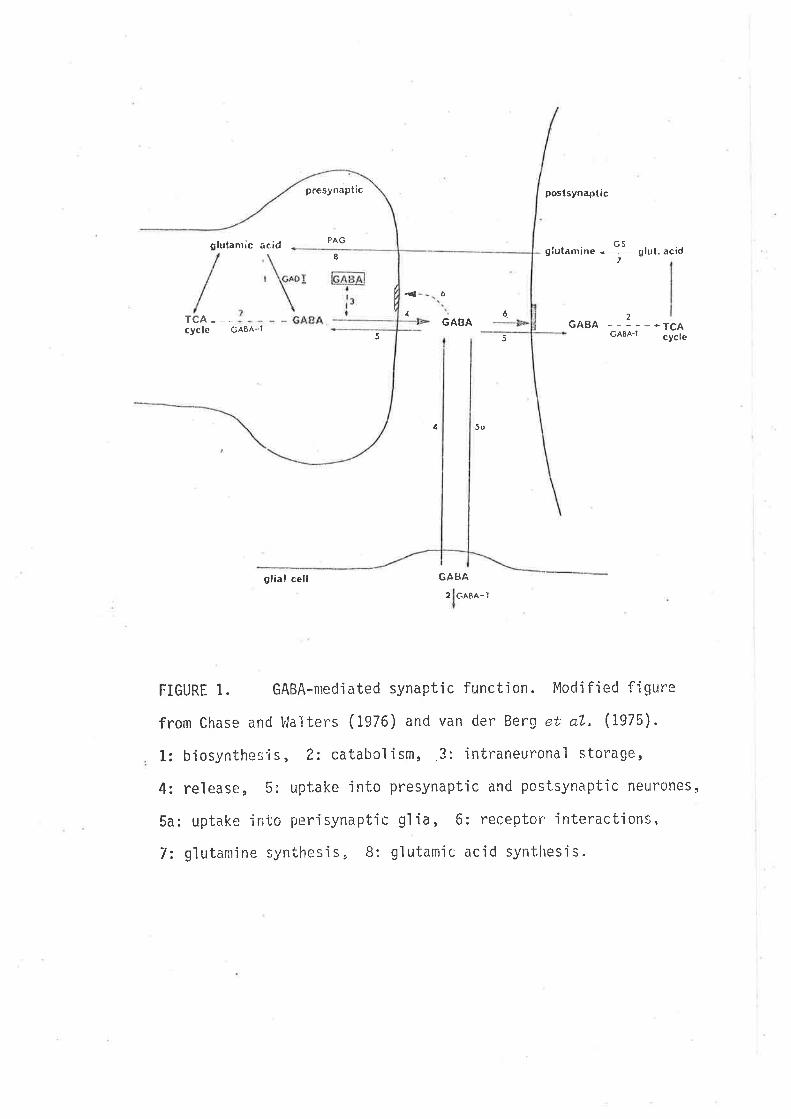
L979), Fig. 1.
Following its release at the synapse, the major mechanism of removal
of GABA from ihe synaptic regìon ìs by transport or 'uptake'u¡hjch occurs
glutamic acid
TCA-_-_cycle Gr\8Â- l
postsynapt¡c
GSglutami ne--g lut. acid7
2GABA -_--
GA8A.I-.TCA
cYcf €
pr€synapl ¡c
PAG
I
4GABA
5J
slial cell
4 5o
GABA
7 GASA-I
FIGURT 1. GABA-nred'iated synaptíc functjon. Moclified figure
from Chase and l,lalters (1976) and van der Berg et; aL, (1975).
1: biosynthesjs, 2: catabol'ism, 3: intraneuronal storage,
4: release, 5: upt.ake ìnto presynaptjc and postsynaptic netlrones,
5a: uptake into perisynaptic glia, 6z reÇeptor interactions,
7: glutamine synthesi s, 8: gl utamic acid synt'hes'is.

6
in eìther neurones or the perisynapt'ic 91ìa (Iversen and Kelly,7975;
Iversen, 1971). Hìgh and low affinity uptake systems have been descrjbed
for GABA (see Fagg attd Lane, !979) and jt has been suggested that both
transport systems play a role in GABA uptake with the loiv affinìty system
thought to be associated wjth generai nletabolite supply io cotnpartment,s
within both g'lia anC neurones (Bond, 1973). Thìs low aff'!nity uptake
mechanìsm (Km 300-1600 M) (Martin, 1976) 'is evettìy distributed in the
brain. By contras"", the high affinity uptake mechanism (Krl 0.3-40 M)
(Elliot and van Geìder, 1958; Iverselt and Neaj, 1968) has a differentìal
distrjbutjon withjn brain tissue (Hökfelt et aL., 1970) and in neurones
js related to storage'in synaptic vesicles within nerve terninals.
l"he function of gììal cell high affinity uptake of GABA'is not cìear,
however it may be related to the proposed 'cycìe' of metabolites between
neurones and gì'ia (van den Berg et aL., 1975). Another possibìl'ity ìs
that in additjon to the release triggered by the nerve impus'le, there
could be a'tonic'release of GABA from glia whereby GABA could functjon
as a modr¡lator (Tapìa, I974). This concept ìs supported by tlre observat'ion
that cells in peripheral gang'lia, where synaptic release of G/\BA'is virtuall¡t
ruled out, possess receptors for GABA (De Groat et aL., 1972) and that on
depolarizat'ion g'lia adjacent to these cells release previously accumulated
[3H] enAn (Boweryand Brown , lgl2; Minchjn ancl lversen, lg74).
Al though bgth nerve terni nal s anci g'l 'ia1 cel I s are capali'l e of hì gh
affinjty uptake of GABA, the substrate specificities of the respective
uptake sites cliffer (Schon and Kel1y, I974; Iversen and Ke]'iy, 1.975;
Martin, Lg76). The GABA analogues 2,4 dlamìnobuiyr"ic acid (DA'BA), c'is-3-
aminocyclohexanecarboxylic acjd (cis*ACHC) and n'ipecoiic acjd appear to be
more select'ive for neuronaì, compared to gìial , up'Lake (Iversen and Ke]ly,
1975; Bowery et aL,, 1976), where as ß-aianinr: is substrate specific'For

7
:]the gíiat GAIIA transpori- system (Schon and Kel ìy, I974; Schousþe, lgTg) .
/i\
Physí.oLogg and Pl'¿ar¡nacology oi GABA
There is evidence for the release of endogenous and exogenously pre-
loaded GABA from bolh vertebrate and invertebrate presynaptìc termjnals
following approprja'¿e stimulation (Jasper and Koyama, 1969; Bradfcrd, I970;
De Feudiset a.7.., 1970; Iversen et aL., I97U Curtìs et aL., 1973; Levy
et aL., 1973; Roberts, i974). Furthermore, this release displays aìl the
classjc charactel"jstics of that for a neurotransmjtter. Follow'!ng its
release, GABA act.ions on the target membrane result'in ejther hyper-
polarìazation or depolarizat'ion, both of lvh'ich are due to an ìncreased
chloride ion (Cl-) conductance.
The act'ion of GABA irl nervous tissue has been shown to result in the
hyperpolarization of postsynapt'ic membranes with a reversai potent,ial that:
is practìcally identicai to that of the inhjbitory postsynapt'ic potentìaì,
which led to ìts acceptance as a major inhibitory neurotransrnitter. It ìs
interesting to note ho'¡rever th¿rt the hyperpoìarization of the sp'inal rnotor
neurones is typicaì'ìy shorterbhan that for central neurones. 0f the
compounds known to antagon ise t.he postsynapt'ic effect of GAtlA, pi crotoxi r,
and related compounds and bicuculljne and relat.ed compounds (Curtis and
Johnston , Lg74; De Feú.Jis, Ig77) al e the most effec+"ive and uliclely used.
Membrane bjnding sturlies suggest that bicucul I ine (a convuì sant phthaleicJo
isoguinoline alkaìoid) (De Feudis,I977; Zukin et aL." I974) conrpetes for
the postsynapt'ic receptor r^rhile picrotoxin (an equìmolar mjxtr:re of
picrotoxìn'in and picrol.in) (Curtìs and Johnston, I974; Zukin et aL.,1974)
acts not entírely at the GABA*receptor, but at the associated Cl-'ionophore,
although there is some evidence for a direct receptor action as well.

I
In contrast to its observed effect upon sp'inaì motor neurones,
application of GABA onto primary afferent terminals 'in the spinal cord
incuded depoìarizatìon, simjlar to the effect of presynapt.ic inhibition
ón dorsal root potential (Levy, 1977). In tlrjs type of jnhjb'ition GABA
released at presynaptic axo-axonic synapses brings about depolarization
of presynaptic terminals causing a reduct'ion in the amplitude of the action
potential jn affer"ent terminals ant thus reduces transniiiter release. Both
presynaptic inhjbition and GABA actions at the prìmary afferents are also
blocked by the specìfic GABA antagonìst bjcuculiine and by pìcrotoxin, or
by desensitization to GABA actions. Taken together',¡lith the properties
of GABA and the hìgh concentration of GABA and ìts synthesiz'ing enzyme
(GAD) fourrd 'in the dorsal honn of the spinal cord (Fahn, ig76; Mìyata
and Otsuka, i975) th'is supports the hypothesis that GABA is a neuro-
transnritter mec'íjatìng presynatpic inhibition (Levy, Ig77; Obata , Ig76).
The inhibitory action oi GABA on vertebrate ancl invertebrate neurones,
appears to be due to the same jonic mechanism (a pronounced increase in
the permeab'il jty of the postsynaptic membrane to C1-) whjch cjamps the
membrane potential at a negative level, preventing depolarizat'ion of
the membrane by any subsequent depolarizing influence. ûABA-jnduced
depolarizations are also mediated by an increased inembrane permeabilìty
to Cl-, however the directjon of the Cl- current is reversed.
GABA ín PeriphenaL Nez"uaus System
llhile effects of GABA on mammalian central neurones are jn general
inhibitory and hyperpclarizing (Roberts, 1976; curtjs and Johnston, !914;
Krnjevic, L974), it has recent'ly been found that G/\BA has sone depolarizing
actions on peri pheral gangì ion cel I s and the'ir axoiìs (üe Groat , ).970;

9
De Groat et aL., L97L). Although the GABA induced depoìarizat'ion of
peripheral ner'/es results fronl an increased Cl- conductance, the chlorjde
equìlibrium potential (tCl) is less than for the resting membrane polent'ial
(Em), sugges'fing that the e'lectrochernjcal Cl. diffusion gradìent across
the neuronal membrane is d'irected outward whjch ìs opposjte to the'inrvardly
directed Cl- gradient medjat'ing'its central hyperpolariz'ing actions. This
difference in Cl- g.'adíent js due to an ìnwardìy directed Ci- pump which
maintains a high ICl]i jn afferent and peripheral neurones. Changes in
external sodjunr or potassiuin ion concentratìons do not signifìcantly alter
the chlorjde equil ibrium potentia'l (-33.7 or -20 nrV) of atferent neurones
(Gallagher et q.L., 1975¡ Nishi et aL., I974; Deschenes et a.L., 1976).
Stud'ies 'involving. intracellular and extracellular recording from the
superior cervical gangl'ia, and dorsal root ganglia of the nabbit, cat,
rat, bulìfrog and chick embryo showed GABA in every instance to'induce
depolarizat'ion (see Obata, I976). GABA actions 'in these cel I s showed
analogous pharmacolog'ic properties to those at central neurones 'in that
picrotoxin and bjcuculline reversibly antagonized GABA actions at low
concentraticlns. Desensitjzatjon to GABA was marked and the depoìarizat1on
decl i ned durj ng GABA appl i cati on .
Depolariz'ing effects of GABA also occur at molluscan and echinoderm
neurones (Fìorey et aL., 7975; Yarowsky and Carpenter, lg77) where GABA
may have an exc'itatory function. Furthermore, it has been suggested
that GABA may be ô.n excìtatory neurotransmitter in the inner ear of the
bulìfrog and skate (F'locP" and Lam, 1974-) and olfactory nerve of garfìsh
and pike (Roskosk'i e'i a'i.." L974). GABA also appear,s t-o have a non-
specific depoìarizing acticn on peripheral C-fjbres (Brown and Marsh,
1e7B).
Recently, 'GAB/\' receptors that are unaffected by the classical

10
'GABA' receptor antagonjsts and insensitjve to many of the GABA-mimetics
such as 3-aminopropanesuì phon'ic acìd (3-APS), i soguiv¡'ic jne anci musc'imoì ,' í ,,'i
have been described on pel"ipheral autonotrric nerve terminals and in
mammalian brajn (Bowery et aL., 1980, 1981; Bowery and Hudson, 1978).
These receptors show high affinity for baclofen ihe ß (-p-chloropheny'l)
derivat'ive of GABA (tox et aL.,1978; Curtis et aL., L974; Bowen.y et aL.,
L979; Johnston et aL., 1980) and have been termed GABAU receptors by Hì'll
anC Bowery (1981). þlhile not produc'ing axonal depo'larjzation as 'in
classical presynaptic jnhibit.ion, the functjon of 'l,l¡ese baclofen (GABAB)
receptors js nevertheless sjmilar to other axonal GABA receptors jn that
the'ir activation causes a decrease in neurotransmitter release (Bor+ery
et aL., 198û).
GABA in En.tev'ì.c Neruous Sysl;em
In pre'.,ious studies (l-lobbì ger, 1958a, b; Fi orey and McLenn$n, '1959;
Inouye et; aL,, I950; Takahashi et aL., 196la o b; B'ianch'i et aL., 1968;
Lewis eÌ; aL., 1972) that inr¡estigated the effect of GABA on the'isolated
rnammal'ian 'intesllne; GABA was ineffective when adrnjnistered alone, but 'in
the gu'inea-pig iìeum GABA (10-6 to tO-3 U¡ reciucecì the submaxirnal st'imulatory
effect.s c;f acetyìcholirre (ACh), nicotìne, histanrine (liA) and 5-hydroxy-
trypt.amine and clepressed the peristaltic reflex (Hobbiger, 1958a, b).
These effects were blocked by pjcrotoxjn. Accordjng to Inouye e.t aL.,
(1961), in producr'ng these effects GABA acted upon sjtes c'losely correlated
with tryptamine receptcrs, but this work has never been subjected to a
serious study.
In a recent and extensive pharmaco'logica'ì study o'1 the actjons of
GABA in the guinea-pig small and large 'intest'irie try Kranti s e't (rL. (1980)

11
showed that GABA causes concentration-dependent transient relaxatìons of
the distal colon, or contractions, or biphas'ic responses'(contractìons
followed by relaxatìons) of the iJuem. By usìng tetrodotoxin (TTX) wh'ich
blocks nerve conduction (Kao, 1966), ìt was determined that GABA has
little or no effect djrect upon the muscle of ùhe ileum or dìstal colon,
rather GABA st'imulates bjcuculljne sensitive receptors mediating excitaticn
of chol inerg'ic exc'itatory and enteric 'inh'ibitory neurones (the transrnitter
for which 'is presently unknown). I'Jhereas Hobbiger (1958a, b) and Inouye
et aL. (1961) observed a depression of peristalsis by GABA itself, ne'ither
the ascending excìtatory nor the descending ìnhib'itory reflex (the prìmary
reflex arcs underlyinc peristaìsis, Costa anC Furness, 1976) when e'lic'itecl
bJ, localised stretch of the colon wall were affected by bicuculline. This
apparent discrepancy'in the effect o't applied GABA upon mctility and the
induced intrinsic reflexes, 'is d'iscussed elsev¡here, a'long wì bh a more
deta'iled invesr"igation of the pharmacoiogica'l and'ionic bas'is for GABA
actior¡s jn the gu'inea-pig intestine.
Sites sirnilar to the baclofen (GABAB) receptors which are present
in mammalian central and peripheral nervous tìssue (see precedìng section)
have a.lso been shown to occur in the guinea.-pìg ileum and vas Ceferens
of mouse and guinea*pig (Bowery et aL., 1981). The actjon of GABA and
baclofen at these receptor s, depressed the twit.ch responses to electrical
stinrulation of these preparations, an action similar to that of both
adenosi ne and the opì ates (morphi ne, enkephal 'in ) (Kosterl i tz a.nd hlaterf j el cj ,
L975, North et aL., 1979) which inhibit the output of acetylcholjne from
chol inerg'ic neruones Ín the guìnea-pig.

t2
The Enteric Neruous System
The mamrnalian enteric nervous system forms an extensive network of
nervous elements totalìÍng some 107 to 108 neurones ìomprisìng up to ten
d'ifferent neurone types (see Gabe'lìa, 1976) which, together with the
different srnooth muscle and non-neuronal ìayers with which these enterjc
nerve networks are jnterposed within the dìgestive tract walì, presents
a rather complex arrangement" Therefore before any further discuss'ion
of GABA actions in the gu'inea-pig intestine, an exp'!anation of the general
structure of the intestine wall and in partìcular, the enteric nervous
systenr,'is necessary.
Func tionaL Ongani zat i on
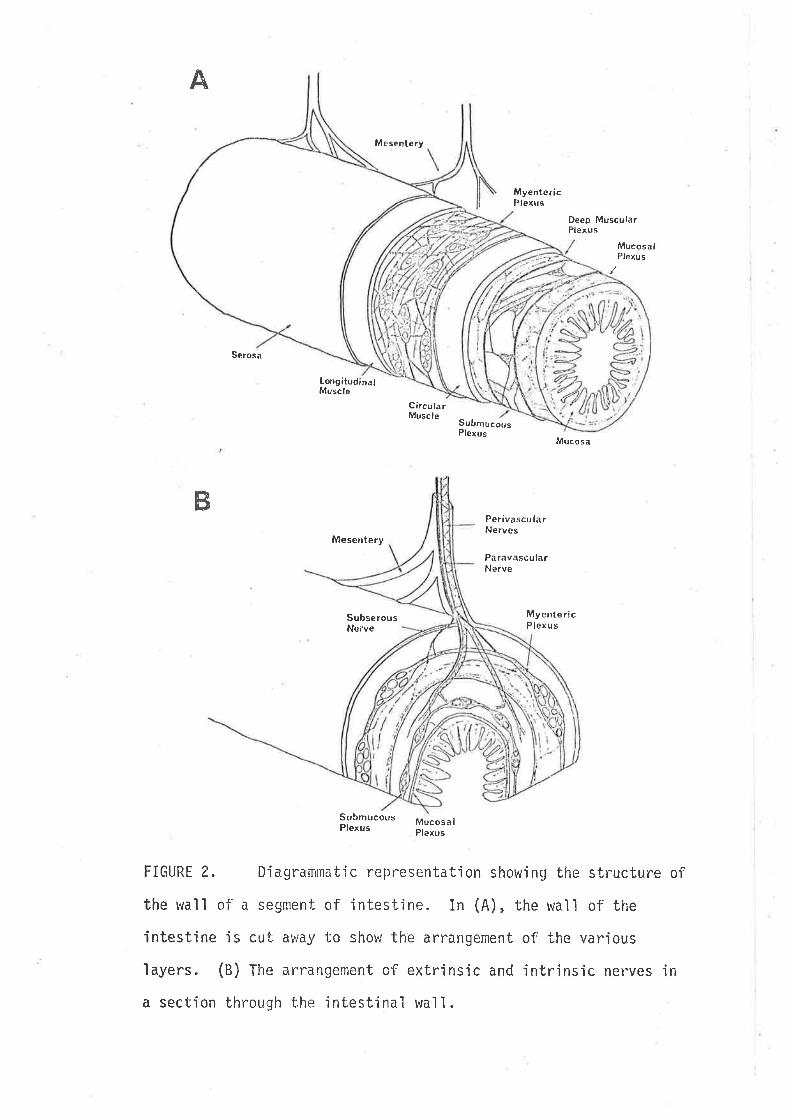
The intestinal wall comprises a number of layers: muscle, nerve
plexuses and mucosa in the general order described'in Fig. 2. This
organizatjon ìs generaììy the same throughout the length of tire gastro-
intestinal tracr-, and although thereare great changes in t.he size and
shape of the various ìayers, there js considerable cons'istency of a
given part of the traci'in a given specìes (Schabadasch, 1930; Schofieìd,
1968; Stach, 1971; Gabella, 1.976). In the small and large intestines,
fibres of the outer longitudinal muscle coat run paralìel to the axis
of the intesbine, and in the small intestine form a continous layer,
whilst in the large jntestine ìongitudinal muscle fjbres are gathered
into distinctive bands termed 'taen'ia col'i ' . The inner circular muscle
coat cons'ists of muscle fibres which are oriented perpendìcular to the
axis of the intestine. Intra-¡ruscular blood capiìlaries (84 per 10,ù00
cross-sectioned smcloth muscle cel1s, Gabeìla, I976) are contajned
A
ts
Mesentery
MyenterícPlexus
Deep MuscularPlexus
MucosalPlexus
Se¡osa
Longiludinalliluscle
Mesenlery
C ircu la rMuscle
SubmucousPloxus
Perivascu larNerves
ParavascularNerve
Mucosa
MyentericPlexus
SubserousNc;ve
SubmucousPlexus Mucosa i
Plexus
FIGURE 2. Diagrammatic representation showing the structure of
the wall of a segment of intestìne. In (A), the wall of the
intestine is cut av/ay to show the arrangement of the various
layers. (B) ffre arrangement of extrinsic and inbrinsic nerves in
a section through i:he jntestinal wall.

i3
within.the circular muscle 1ayer, runn'ing approx'imateìy parallel to
the nluscle ce'ils. Between the longitud'inal and circular muscle layers
are aggregates of gangiion cell bodies with interconnecting bundles
(fascicuìi) of axons. These aggregates and their associated connecting
fibres form the myenteric plexus (Auerbach's plexus), vrhich makes
connections wi'bh an extr insic 'innervation through the mesenterjc nerve
and a transjtìonal subserous plexus also conta'ining neurones into clistinct
gangì i a .
l'he nervous innervation of the circular muscle coat consists of
nerve bundles frorn the myenterìc pìexus which penetrate this muscle
'layer, sonìe of which then run paraì'le1 to the smooth muscle celìs, or
pass through and connect wjth the subnrucous plexus (Caial , 1911; Gabella,
l97Z). Nerve bundles frclm the myenterìc plexus also connect wìth a
nerve plexus 'musct,¡laris profundus' (Caial , 1911) which lies adjacent
to the inner (lumina'l) aspect of the circular muscie. The nerve fibres
of this 'deep muscular plexus' run parallel to the muscle cells. Dist'inct
glanglia are absent"
Located betv¡een the cjrcular muscle coat and mucosa, the subnlucous
plexus consists of two nerve ìayers compr"ised of interconnected ganglia.
These gan,lia are generally smaller than the myenterìc ganglia and\
general 1y conta'in f er^ier neurones . Extensi ons of the submucous p1 exus
pass through the 'nluscularis mucosae' to connect with the mucous pìexus.
Comprìsed of a varjety af plexuses, variously associat.ed t'iith g'lancis'in
the mucosa, these pìexuses are contjnuous with and represent an extension
of the submucous plexus" The pìexus contains nc neurone Soma.

T4
Nerve bundles are practically absent from the 'longìtudinal muscle
(Thaemert,1963; Taxi,1965; Paton and V'izi, 1969), however there are
vesiculated nerve processes of the myenteric pìexus which l'ie on'ly a few
tens of nanometres away, separated from the muscle by only a thÍn basal
lamina (Gabclla, 1972). This close apposÍtion of nerve and muscle layers
presumab'ly a1ìovrs rapid (miìl iseconds) diffus,ion of transmitter from the
nerve plexus to muscle celìs, and no doubt accounts fon the variety of
pharmacolog'ical resporrses observed in isolated longitud'inal muscle myenteric-
plexus preparations.
Myenter,í.e P1.eæus ,
The neurones comprising th'is plexus are djstributed into 3 p'ìexiform
meshworks (see Schofield, 1968). The large e'longated ganglia with inter-
connecting bundles cf unrnyelinated fibres constitute the prirnary meshwork.
t,'lithin, and continuous with this meshwork, is a secondary meshncrk whìch,
al though I acking any di scernabl e aggregates of cel I bodies comparabr'le t<l
the gangìia of the primary meshwork, contains cell bodies along the course
of the fasc'icuji. Br.rnching from,and locaìised within, the secondary meshwork
is the tertiary meshwork. There is some morphological evidence for the
presence of cell bodies wjthin this meshwork (tajal, 1911-; Gabeì'la, L979).
Reciprocal innervai'ion occurs between the nryenteric plexus and ìarge
paravascul ar nerve buriclj es t¡rhi ch ra.mi fy wi th'in the submucous pl exus , and
addjtional direct connections oqcur between the subnrucous piexus and
elements of the myenteric tertiary rneshwork. Innervatisn of the circular
muscle coats by myenteric neurones anises from this tertiary rneshwork.
in contrast to the subnrucous plexus the myenterìc plexus contains no blocd
vessel s.

15
Myenter'ic Neurones
Individiral neurones can be readily seen within the gangìion of the
primary meshwork. These neurones range in sjze dependìng on the reg'ion
of gastrointestinal tract and the species in question. In the rat caecun'r,
the myenteric neurones range between 100 um2 to ovcr 2000 um2 in cell
surface area. l'{yenteric neurones in the gu'inea-pig measure between 10-100
pm in length and can number more than 100 per ganglÍon. Morphoìogicaì
characterizationofmyenteric neurones shov¡s a broad spectrLrm of cell
types conformjng to Dogiel's (1899) classification: Type I, II ancl IIIcells, and to more recent classifications based on cell size (Gunn, 1959,
1968) and on the nunrber of processes (Schofie'ld, 1968).
The three types of ganglion cells descrjbed by Dog'ieì include:
Type I cells, characterjsed by numerous short stout dendrites of irregular
calibre (cogwheeì ìn appearance) and a síng1e slender axon which passes
into t.he intenconnect'ing fibre bundles (fascicuìi). Type II cells are
multipo1ar, however their dendrites are typìcally ìonger and usual1y extend
beyond the ganglion" These dendrites are fewer in number than for the I'ype
I neurones. Type III cells have dendrites of intermediate length whjch
appear to terminate in the same or adjacent ganglion.
In addition to these neurones, wh'ich according to Gunn's classification
are referred to collective'ly as large neurones, there are also smaller
neurones present in rela.tìve'ly large numbers; sma'ller than any of Dogiel's
cel I s.

16
Subrm,Leous PLerus
As wjth the rnyenteric plexus, the submucous plexus cons'ists of
several fine meshworks. l-loweverun'Ì'ike the myenteriô p'!exus, it js divided
into two layers. The inner plexus (nearest the mucosa) termed Meissner's
plexus, is charact.erised by a sma'l'l , regular closed pattern meshwork,
containing small ganglion cells. The outer iayer, lying against the inner
side of the circular muscìe, is termed the Shabadasch plexus or l{enle's
plexus. In most part, this plexus'is open meshed and very irregular with
the gang'lia sparseìy distributed. The ganglion cells are of the sanre type
as those of the myenterìc plexus, however the gangija of the submucous
plexus conta'in fewer cells (approx'imately B), and indeed there are fewer
cells in total wjthin this nerve plexus.
Eætnirtsic Inn.eruation of the Enteric PLeruses
A'lthough the gastrointestinal tract dÍspìays independence of action
from central control there are nonetheless rìervous connections v¡ith the
central nervous system via three major efferent pathways: the vagus nerve;
the sp'lanchnic nerves, and the peìvic nerves (Fì9. 2), through which the
central nervous system modulates gastrointest'inal activ'ity.
0n the motor side, the abdomjnal vagus accompanying bìood vessels
(paravascular), contajns mostly parasympathetíc chol'inergic (ACh)
preganglionjc nerve fÍbres of which the majority ìnnervate the stomach and
distal portìon of the large intestine. These cholinergic nerve fibres
form excitatory connections wjth both intrinsic cholìnergic and enterjc
inhjbitory neurones located in the myenteric pìexus (Danie1, 1968). There
are also non-cho1ìnergìc inhibitory f ibres (Lang'ley, 1898) and a snlall

t7
number of postgangljonìc âdrenergic fibres (Nielson et aL., 1969; Lundberg
et aL., 1976). In addìt'ion, the vagus also contains a variety of afferent
fibres (Dan'iel , 1968).
Addi ti onal exc i tatory i nput bo the 'i ntest'ine i s evi dent on sti mul ati on
of the mesenteric nerves (Bartho and Szolcsanyì, I979; Szolcsanyi and Bartho,
1979). Accordìng to ùhese authors, the excitatory responses elìcited in
this way are the result of antìdrontic stimulation of sensory nerve
processes causjng the excitation of the intrinsic cholinergìc nerves
possibly by way of true synapses" The'identit.y of the sensory neurotrans-
mitter medjating these effects is unknown although there is evidence
(Hökfelt et aL., ,1976; Franco et aL., L979a, b; Guilleme'in, I97G; Furness
and Costa, L979), precìud'ing the involvement of either substance P (SP) or
somatostati n.
Sympathetic innervation (adrenerg'ic) of the intestine is large'ly
postganglionjc,eminatjng frorn three main groups of sympathetic Aang'lia:
vertebral, pre-vertebral abdominal gangìia, terminal ganglia. Sympatlretìc
input occurs via the mesenteric nerves which emerge fro¡n t,he abdominal
pìexus of the splarrchnic nerves, the vagus, some of which may be pre-
gangì'ionic,and the pelvic nerve. These adrenergic (noradrenaline) nerve
fibres have a prejunctÍonal inhibitorry action upon the release of acetyl-
choline from 'intrjns'ic cholinergic nelrrones (Furness and Costa, Ig74;
Hirst and McKìrciy, 1974). This e'ffect is mediated through alpha-adrenergic
receptors. Howeirer t.here are also beta-adrenergic receptors present, located
on the muscle wh'ich on stimuiation cause relaxation of the intestine.
These paravascular nerves, can reacliìy be distinguished from the fine
anastomosing nerve plexus (perìvascular) rvhich supplies the arterial
innervation. The latter include vasodilator nerves invol'¿ed in the

18
intrinsic vasodilator reflexes, which release ne'ither noradrenaline (NA)
nor acetylcholine (ACh) (Biber, 1973; Fasth et a7.., L977).
Intz,insíc Inneruat'Lon of the Enteric PLeæuses
The enteric nervous system of the mamnralian'intest'ine js complex,
containìng an 'intrjnsic nervous netvrork withjn which are sensory neurones,
interneurones and motor excitatory anci inh'ibitory neurones that provide the
considerable ìntegratìve capacity necessary for the conLrol of gastro-
intestinal funct'ions including the reflex movements of gut musculature
(peristalt'ic reflex), uhere distension of the gu'l waìì initÍates enteric
reflexes causìng .ont.uctions which push the contents analward into an
area of relaxat'ion. As the contents move, the reflexes continue to be
elic'ited, contraction behjnd and relaxation in front u¡hich maintain the
propulsjon of the contents. Reflexes also include secret'ion of gastro-
intes+.inal horrnones, and the regulation of blood flow. The gut also
exhjb'its penclular non-propuìsive r¡ovements which fac'ilitate the m'ix'ing
of gut contents for digestion.
A nunrber of substances are proposed to be neurotransmitters in the
mammal ian enteric nervous system, ìnc1ud'ing dopam'int'-- (DA) , vasoactive
intestinal peptide (VIP), somatostatin, substance Po enkephaìin,
cholecystokÍnin, bornbesin, neurotensin, adenosine-5'-trjphosphate (ATP),
acetyl chol ine (ACh) , noradrenal ine (NA) , and 5-hyclroxytryptanrjne (5llT)
(Burnstock, 1972 Gabella, 1.976; Dreyfuss et aL., 1977; Costa et aL.,
1980). 0nly ACh and NA have satisfied all of the criteria for a neuro-
transmitter (Gershon, 1981) a'lthough there is considerahle evidence for
5HT having such a function (Costa and Funness, I979a; Gershon, l9B1).

19
Low frequency transmural electrical strmulatìon of the intestine
elicits a release of ACh and a concommitant contraction of both the
circular and'long'itud'inal smooth muscle coats (gulUring et aL., 1958;
Kosterlitz and l-ees, 1964), which is antagonised by muscarinìc receptor
blockade. Furbhermore, muscarinic antaEonists inhibit peristalsis
(Kosterlitz, 1968) suggesting that choljnergic motor neurones are important.
for the propulsive activity of the intestine. It has since been shown
that the final neurones in the excitatory reflex component of peristaìs'is
(Kosterlitz and Rcbinson, 1959; Furness, 1969) release ACh.
Gang'lionic nicotinic blocking agents such as hexanrethonium prevent
both the excitatory and inhibjLory components of the per.ista'ltic reflex
(eulbring et aL., 1958; l(osterf itz, 1968). There are thus cholinoreceptive
sites in the gang'lia subserving the stinruìation of both cholinergìc and
the enteric inhibitory motor neurones. In support of this Nishi and North
(1973) and North and Henderson (7975) report that hexarnethorrìum blocks
fast EPSP's in guìrrea-pig iìea1 myenteric neurones. Approxìmately one
half of nryenteric neurones recorded from show fast EPSP's and are termed
Type I cells (Nish'i and North, 1973) or S cells (Hirst et aL., 1974),
di sp'layì ng a bri ef af ter- hyperpol ari zat ion .
The actions of enteric inhibitory neurones which supply the muscle
are not pr"evented by adrenergìc bìockade, and hyperpolarization of the
intestjnal muscle stjll persists followìng degeneration of the noradrenergìc
innervation (Campbell, 1970; Burnstock, I972.). The ìderrtjty of the
inhib'itory transmitter released by these neurones has proven e'lusìve,
a'lthough a number of substances have been proposed as candrdates.
According to Burnstocl< (I972, I975) adenosìne triphosphate (ATP) js the
non-adrenergic non-cholinergic inh'ibitory transmitter, anci the work by
Burnstock and his colìeagues has been the basis of the hypothesis where

20
these nerves have come to be termed 'purìnergic'. There 'is however,
considerable evidence avaijable disput'ing the purinergic hypothes'is, the
details of which will be ciiscussed in Cltapter VII.
Recent immunohistochemical studies have demonstrated immunoreactivity
toward the hìstoìog'icaì1y localised po'lypeptìdes substance P, enkephalins,
neurotensin, somatostatin, bombesin, vasoactive jntestinal peptìde (VIP)
and gastrin/cholecystok'inill tetrapeptide, wíth'in enteric neurones (HOktelt
et aL., L975, 1976, L977; Fuxe et aL., 1977; Schultzberg et aL., 1980).
In the mammalian gut, the enkephalins and morphine cause inhibition
of neuronal acetylchol'ine release (Paton, L957; de la Lande and Porter,
1967; tlaterf i e1d et aL., 1977) and jnh'ibjtion of fin'ing of nryenteric
neurones (North and l^lilliams, 1976). 'lhese depressan'u effects of the
opiates result from the depo'larìzation of the nerve processes (Ncrth a¿
ctL,, I979), v'ia naloxone sens jtìve opiate receptors which, in the guìnea-
pig inferior mesenteric ganglion, are proposed to be located on the
chol inergic nerve terminal s (Bornsteìn and Fjelds, 1979). Thi s wou'ld
exp'lain'Lhe extensive distribution of enkephal in-l jke immirnoreactive
nerve fibres in the circular rnuscle and taenia coli (Jessen et aL., 1980)
where a direct effect of the enkephaìins on cholinergìc nerveterminals
would be possible.
Substance P (SP) neuronal processes densely innervate the myenteric
ganglia projecting both in the oral and anal directi,¡ns (Ëurness et aL.,
1980). SP has been shown to depolarize enteric neurones (Katayama and
North, 1978) and to cause contractjon of the intestine by a d'irect action
on the smoot.h muscl e (Bury and l'lashford, 1977; Sze'l j ¿t aL. , 1977 ¡, Yau,
1978. Franco e.t aL., L979a, b). There is novr evjdence that substance P
conta'ining nerve fibres are also distrìbuted f.hroughout the circular muscle

27
layer of the srlall and large'intestine, as we'!1 as in the longituciinal
layer of the taenia colj and ascending colon where jt is proposed to be
the transmitter mediat'ing non-cho'linergic contractjons of these preparations
(Franco et aL., !979a, b; Jessen et aL.,1980). Substance P is able to
initiate and increase peristalsis'in the guinea-p'ig ileunr (Holzer and Lembeck,
1978) and although there are cholinergic inputs to substance P neurones
(Franco et a'1., 1979a, b) it is unclear whether substance P nerves are in
anyway importan't for intestinal motilìty. Indeed most recently Costa et
aL. (198i) have shown that most of substance P released by electrical
stimulat'ion in fact originates from afferent fibres, not ìntrinsìc neurones,
whilst Chahl (1981) suggests that these afferent fibres make s,vnaptic
contact r,vjth the cholinergic motor neurones and that the substance P
responses as a result of electrical stjmulaticn are an a.rtefact due to
its overflow from these afferent synaptic regions.
Neurones disp'laying somatostatin immunoreactivìty occur in both the
submucosal and nryenteri c p'l exus (Furness et aL., i980 ) " Somatostat j n has
little direct effect on the smooth muscle. Rather son¡atostat'in has been
found to depolav'ize some myenteric neurones whilst hyperpoìarizing oihers
(Katayama and North, 1980). The types of nryenteric neurones affected by
sonlatostatin have not been fully invest'igated, however neurones hyper-
polarized by somaiostatin may be cholinergic-excìtatory ones sjnce somato-
statin inhibits transmission from cholinergìc neurones (Cohen et aL., L978;
Grafe et aL., 1978)" There is pharmaco'logicaì evidence (Furness and Costa,
I979) for intrinsic enteric inh'ib'itory neurones being stinrulated by somato-
statino whìch, '[aken together with the analward polarity of somatostabin
immunoreactive neLtrones, suggests that this pepbide nray have a role in the
descending 'inhibitory reflex (Furness and Costa, i980).
VIP'imtnunoreactive neurones have been iclentified within the gu'inea-p'ig

22
ileal myenteric and submucous plexuses (Schultzberg et aL., 1978). t¡lithin
the myenteric plexus these neurones run in an oral to anal dìrection,
ramifyìng to cther myenteric neurones and to the circular muscle (Furness
et aL., 1980). Furthermore, ViP relaxes smooth muscle (Furness and Costa,
1980) and i s rel eased f rom the gut fo'l l or^ri ng vagal stimul ation (Fahrenkrug
et aL., 1978). 0n this basis the non-adrenergìc, non-choliner"gic inh'ibrìtory
neurones are proposed tc be peptìdergic rather than purinergic, releasing
VIP (Costa et aL., 1980).
At present, however, there is a paucity of data to sirggest that VIP
is released by neuronal act'ivity, or that it has pharmacologicai actjons
comparable to the. non-adrenerg'ic inh'ibitorry transmitter. Incleed Cocks
and Burnslock (1979) and Mackenz'ie arrd Burrrstock (1980) have shown
pharnraco'logical'ly that exogenously admìnistered ViP does not mimick the
inhitritory response to non-adrenergic non-choj'inergic nerve stimulation
in the intestine" In addition, t^liljiams and North (1979) report that
VIP is ine'f'fective upon jleal 'long'itudinal muscle but rabher depolarjzes
myenteric neurones. Although the neurotransmitter for non-adrenerqic,
non-chol inerg'ic 'inhibition rema jns to be elucidated, the roles of these
neurones are well def ined (Furness and Costa , I973; Hjrst and frlcKirdy,
1974; Costa and turness, 1976). They are 'involved in the descendíng
inhibition, fjrst described by Bayf iss anci Starìing (1899) which produces
the reflex relaxaticn ofthe gastrointestinal tract immediate'ly in front
of substances travelling through the digestìve tube.
In addition to the intrjnsic, cholinergic excitatory innervation of
the smooth musclç coats of the intest'ine, nerve mediated contractions
resjstant to muscarinìc blockade have been reported. These 'include a
'rebound'contraction of the muscle fol'low'ing an inhibitory influence
(Bennett, 1966; Campbell and Burnstock, 1968; Furness, L97!) whjch may

23
appear as a primary response to stimulation, ìf the period of nerve
stimulation is brjef and the inherent muscle tone low. However, we
cannot prec'lude the possìbiìity that the depo'ìanjzation leading to thìs
type of contraction is in fact induced by an excita+.ory substance be'ing
released onto the muscle. For post-inh'ibitory 'rebound' this is almost
certajnly due io prostagland'in release (Burnstock et q.L", 1975).
Studies by Ambache and Freeman (1968). Ambache .e.t aL. (1970) and
Furness and Costa (1973) have shown slow contractions of the longitudinal
muscle response to transmural stimulation (5-50 Flz) urh'ich have a different
time course f rom the so cal I ed 'rebound' contnactiori descri beri above,
that could be blocked by tetrodotox'in (ffX). It has been suggested tlrat
a 5-hydroxyLryptamìne (sHT)-like substance may be mediating this r"esponse
(Costa and Furness , 1973; Costa and Furness , 1979a ) . Hot'¡ever, much more
evidence is needed to substantiate this suggestion since in the distal
colon, circu'lar nluscle is quìte insensitive to SHT (Costa and Furnessn
1979b). Moreover, whilst cholinergìc excitatory input to enteric neurones
vja nicotinìc receptors is v¡ell establìshed (Kosterlile and Lees, 1964;
Bul bring ancl Gershon, 1967; Crema et aL., i970) , there are synapt'ic
actions whene excitation is not blocked by nicot'inic gangìionic blockers
(Uingledine and Goldstein, 1976), but'is antagonised by the 5HT antagonìst
methysergide or by desensitization to 5HT (Katayama and t{orth, 1978; l'lood
and Mayer, 1979).
Sites of action for 5HT on myenterìc neurones, distinct from nicotjnicI'
receptors have beert identified (BroÇee and Johnson, 1963; Gershon,1967;
North et a'I., 1980) rvhi ch together wi th e,¡ì dence for i ts synthes ì s
(Gershon et aL., 1.965), storage (Gershon and Ross, 1966a, b) and release
(Gershon, 1965; Bulbring and Gershon, 1967) has prompted speculation on
the poss'ibility t.hat 5HT is a neurotransmittet'in the enteric nervous

24
system (Costa and Furness, i979a). However 5HT has w'idespread multiple
effects in the mammalian'intestine and the precise nature of 5HT actjons
in the mammal'ian gastrojntestinal tract remains unclear.
Contractions of the ileal 'longitudinal muscle, distÍnct from post-
ínhibitory'rebound'and the above slow excitatjon, are also elìcited by
transniural stimulat'ion (5-50 Hz). Accordjng to Franco et aL. (1979a, b)
these are related to the act'ion of substance P (SP). SP released by
intestinal nerves, or applìed exogenously, acts directly upon the muscle
to induce a contraction. Whether any ofthese substances are released
durìng the peristaltic refIex, remains to be determ'ined.
Accord'ing to l{ishi and North (1973) and Hjrst (1979) Lhe slow
depolarization potentials recorded from a small proportion of both Type
l and Type 2 myerrteric neurones are infact not mimicked by 5HT but have
the sanre'ion'ic mechanism as the potentìaì change caused by SP" IPSP's
have i:een recorded in myenter.ic neurones iWood and l4ayer, 1978) and
approximate'ly 45% of the submucous p'lexus (Hirst et a7,., t974; Hirst
and I'lcKirdy, 1975). The transmitter mediating IPSP's of enteric neurones
is at present unknown. The ionic mechanism appears to be similar tr¡ the
effects of somatostatjn and enkephaf in (l.jiIIiams and ltlorth, 1979; Nishi
and North, L973; North et a!", L979) although the hyperpolarizing mechan'ism
has not been cornpleteìJr elucjdated.
Although adrenerg'ic (alpha and beta) receptors occur throughout the
gastroìntest'inal tract, noradrenal íne aci'ing at these s'ites is in most
part released from extrinsic ay.onal processes except for the proximal
colon of the guinea-pig where intrjnsic non-;idrenergic cell bodies have
been found, located the myenteríc and submucous pìexuses (Gabe'lja, I976).
The peristence of IPSP's recorded from enteric neurones following

25
extrins'ic deuervation and subsequent loss of catecholamines (Johnson
et q.L., 1980), prec'ludes the possibility of the jntrinsic 'inhl'bitory motor
neurones being noradreriergic (Furness and Costa, 1978). Holvever, there is
considerable evidence for noradrenaline to be released from extrinsic rìerve
terrninal s which 'inrpinge on chol inergic axons and exert a presynaptìc
inhib'ition of ACh release (Kosterlitz et aL.,1970; Holman et aL., 1972;
Nishi and North, 1973; North and N'ish'i , 1974)
In addition to those compounds already described, a number of chem'ical
substances have pronounced pharmaco'logica"ì aciions on the intest'ine, inc'luding
bradykjnin, histamjne anC gastrins. Bradykin'in (Day and Varre, 1963; [-ishlock,
1966; Downi ng and, I,lorri s, 1981) el ic j t both contraction ond rel axat'ion,
and histamíne via djrect and'indirect actions upon the smooth muscle
(Danie'l , 1968; Downing and Morrì s, 198i) .
The gastrìns contract a number of mammalian gastrointestinal
preparations and st-inrulaùe gastric and pancreatic secretions (see Danìeì,
1968). At present the evidence for eìther neurotensjn or bombes'in tcl
have major pharmacoiogica'l cr physiologica'l act'ions in the mammalian
alimentary canal is limited (Costa and Furness, 1980; Schultzenberg ei
aL., I979), holever, neurotensin contracts the gu'inea-pig íìeunr through
a nerve med'iat.ed partìy chol inerg'ic process (Kitabgi and Freychet, L978,
i979) and poss'ibìy a substance P-erg'ic process (Monìer and Kìtabgi, 1980).

CHAPTER I I
THE PHARI4ACOLOGY OF GABA ACT I ONS

26
Ir¿tv,oduciíon
Recent'ly Kranti s et aL. (tgeO) have shown that GABA has stjrnulating
actions on enteric tleurones,as seen in the isolated guinea-pig small and
large intestine, in contrast to ìts recognjsed inhjbitory actions in
central and peripheral nervous tissue (Krnievic, 1974). These excitatory
actions were antagonised b.y the GABA antagon'ist bicuculline cr by making
the ti ssue tachyphyl act'ic to GABA. Exc'itatory actions of GABA have al so
been observed when 'recording from neurones of the guinea-pig rnyenterìc
pì exus (Grafe ei; a'1. , 1979), but apart f rom thei r demonstra.ted sensi tì v'ity
to picrotoxin,whích may suggest a h'itherto unrecognised functìon of GABA
in the intestine ,(e.g., enteric neurotransmitter), little is knov'rn of the
pharmaco'logy of these responses. However, for the study of a neurotrans-
mitter some informatjon about the site and mechanism of drug action is
necessary (Orrego , I979).
In the mamrnaliarr centrai nervous system, GABA actions a't- specifìc
receptors resul'b in an increased chloride conductance that'is responsìbìe
for both pre and postsynaptic inhibition (Curtis and Johnston, 1974;
De Feudis, 1977). l,Jhen released on to postsynaptic receptors, GABA causes
an increased chloride permeabilÍty of the target membrane resultìng in a
net influx of chloride (Cl-) ions into the cells so that the nrembrane
becomes hyperpoìa.rized. Although presynôptìc GABA actions also result
in an increased Cl- pernreability, the Cl- ions leave the cells instead
of entering then,so that the cell membrane is depolarized. GABA
depolarizing actions have also been observed in mammalian pe¡^ipheraì
nervous tissue, includìng syrnpathetic gangì'ia (Adams and Brov¡n, 1975),
C-fibres (Brown and Marsh, 1978) and sensory neurones (Gallagher et aL.,
1978). All of these GABA effects show analogous pharmacolog'ic and ionic
propertìes. In contrast, there is a class of receptors meCìafing GABA

27
effects selectively activated by baclofen (wh'ich is the g-p-chlorophenyl
derivate of GABA). This effect, no coupled with Cl- ìonophores, js knourn to
reduce transmitter release in a number of preparations (Bowery et aL., 1g8i).
Most experimental approaches to identify'ing tl're nature of GABA actions
in varjous nervous tissue have involved isolated tissue preparations, s'ince
they permit the use of known concentrat'ions of drugs and measured manìpu1at'ion
of the extracellular ion'ic environment. Picrotoxjn and bicuculline reversìbìy
antagonjse the neuronal depressant actions of GABA in various vertebrate and
invertebrate preparations (Galindo, 1969; CurtiS et aL., 1970) and hence
have been used extensÍ vely to exami ne GABA-synapt'ic transmi ss'ion (De Feudì s ,
L977). Although,there is some controversy es to the specificity of these
antagonists (Dudel , 1965; Bowery anC Brown, I9r-4; Barker et aL., tg75), itis general'ly a.greed that bicucull ine exerts its action at the GABA-r'eceptor
whilst picrotoxin acts at the associated ionophore.
A number of probìems are assoc'iaied with investigation of the ionjc
basis of GABA effects. For example, removal of Cl may result in an
altered osmolarìty, also the commonly used substituent anjons may have the
additional probìem of bindjng calcium, usually to an undefjned extent.
Furthermoren if other than mono-valent substitu'fent anions are used (e.g.,
sulphate) the ionÍc strength of the solution js difficult to determine
(4. H. Bretag, personal communication). The perrneability of the membrane
to the substituent anion may also result in equilibration of thìs anion
across the membrane, so that it can substitute for Cl- in the GABA-mediated
conductance change. Ail in all this experimental approach would thus
appear unsati sfactory. An al ternat'ive method 'invol ves the use of specì f ic
ion transport blockers (e.g., ammoniu¡'n salts and turosemide, Nicoll, I975,
1978), whìch in addition to avoiding many of the problems associated with
ionìc substitution, affords 'less intenference with cationic permeability.

28
Furosemide selectìvely blocks hyperpo'larizing and depolarizing responses
to GABA in frog spina'l cord, presumably by bìocking the ionic channels
operated by GABA (Nico'l'l , 1978).
A more detailed investìgat'ion of GAUA-induced responses in the guinea-
pig ileum using furosemide and a nevv and potent inhibjtor of chloride
transport,piretanìde (Zeuthen et aL.,1978), together with the established
GABA-antagonists bicucull ine and pìcrotoxÍn is reported l'rere.
Iutatenials and Methods
Guinea-pigs of either sex weighing between 200-400 g were stunned
by a blow to the head and bled. Segments of proxima'l 'ileum were qu'ickìy
removed and placed in a modÍfied Krebs solutjon of ihe'tollowing composition
m mol/1 : Na+ 151.0; K+ 4.6; Mg** 0.6; ca2+ z.B; cl - t34.9; HC0r" ?4.9;
HZP04- 1.3; S0O- 0.6; glucose 7.7. The solution was maintajned at 36"C
and gassed with a mixture of 95% 0, and 5% C}Z, to maintain the pH at
7.3. Segments of ileum 4 cm in length were then attached at one end to
the bottom of a 10 ml water-jacketed bath conta'iníng warmed Krebs solution,
with the other end attached by a cotton thread to a Harvard rotary trans-
ducer and allowed to equiljbrate in the organ bath for 60 min before
commencjng e'lectrical stimulation or drug treatments. Mechanjcal reac.t'ivìty
of the ileal segments to the various treatments were displayed on a Grass
Po'lygraph (Model 5D).
For experiments in which intramural nerves were stimulated and tissue
responses recorded, two platinum wire ring electrodes 4 nm apart, were
pìaced around the segments of intestíne. Pulses of 0.2 msec duration were

29
delivered at a frequency of 0.2 Hz for a 10 sec stjmulation periotl, or of
0.1 msec duration at 0.1 Hz for varyìng train lengths. The strengüh of
stimulation was adiusted Lo give responses of near maximum amplitude.
Drug volumes administered were never Inore than L% of the bath volume.
Agonists were left in contact with the tissue for periods of 20 to 60 sec
at intervals of 4 or more minutes. Antagon'ists were added at leasi 10
min before the agonist was tested. Drugs used were: acetyìcholjne chloride
(ACh), atropìne sulphate, bicuculIìne, gamma-aminobutyric ac'id (GABA),
picrotoxinin, furosemide, histamine, bradykin'in, piretanide, tetrodotox'in
(TTX). Furc¡semide was a gift from Hoechst Austral ja and p'iretairide a
gift from Dr Alan Bretag.
Stq.tísticaL AnaLysis: Results are given jn the figures as means + s.e.
means. The Student's t-test for paired and unpaired samples u,as used to
assess the sign'ificance (p.0.05) of difference between mean values of the
dose response effects. ¡\ll regression lines were calculated by bhe least
squares methoC. Regression l'ine slopes of the Clark plot (Clark, igz8)
were compared usìng the t value derived from * = (b:þ_q)'N1:7 whene bs = slope' - --rT:eof the control; N = sample size\ y2 = coefficìent of deterrnination. The
regression line of the Schild p'lot was constrained to a sìope of -1, and
the 95% confidence limits of the pA, vaìue, calcuiated according to Tallarida
et aL. (1979).
ResuLts
When GABA was appìied to the ileum in the orqan bath it produced
either contractions or else biphasic responses (transient contractions
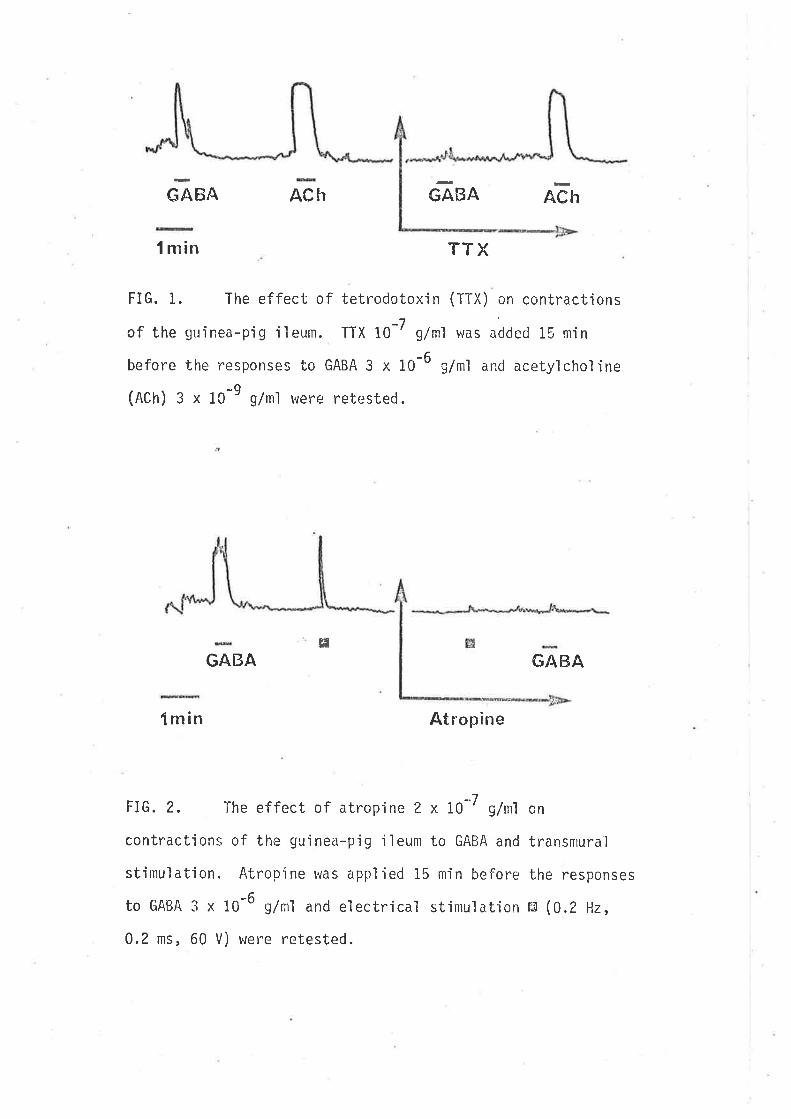
GABA ACh GABA ACh
l min TTX
FIG. 1. The effect of tetrodotoxin (TTX) on contractìons
of the guinea-pig iìeum. TTX 10-7 g/ml v¿as added 15 min
before the responses to GABA 3 x 10-6 g/ml ancl acetylcholine
(ACh) 3 x 10-9 g/ml were retested.
GABA G/\BA
fmin Atropine
FIG. 2, The effect of atropine 2 x i0-7 g/nrì on
contractions of the guinea-pjg ileum to GABA and transmural
stimulation. Atropine was applíed 15 min before the responses
to GABA 3 x 10-6 g/rll and electrical stimulation r'J (0.2 Hz,
0.2 ms, 60 V) were retested.
g

30
followed inlmediately by relaxation), as describeci by Krantis et aL. (1980).
0nly those tissues showing purely contractile responses were subsequentìy
used. The response of the ileum to appììed GABA was immediate, with the
contractions reaching nraximum wìth'in 15-20 sec. The threshol d concentration
was about I0'7 g/m1 and the maximunr contraction was observed to occur
betweerr 6 x 10-6 - 10-5 g/ml. These concentration-dependent contractions
were the result of GABA stimulation of neurones concerned with cholìnergic
innervation of the smooth muscle of the 'i'leunr, since GABA actions were
abolished by tetrodotoxin 10-7 g/n1 (a conrpound which interferes with
nerve action potent'ia'ls by block'ing sodium channeìs, Kao, 1966) (Fig" 1),
and the muscarinìc antagonist atropine 2 x LO-7 g/m1 (Fig. 2). GABA
applied in the pr.esence of prolonged application of the same concentrat'ion
of GABA did not evoke any detectable response of the ileum. This effect
is characterjstic of the responses to GABA be'ing desens'itized. The con-
tractions were antagonized in a concentration dependent manner by both
picrotoxìnin and bicuculìine, whjch usually had no effect upon tissue
activity when added to the bath although on occas'ion a transjeni contraction
of the ileum uras observecl.
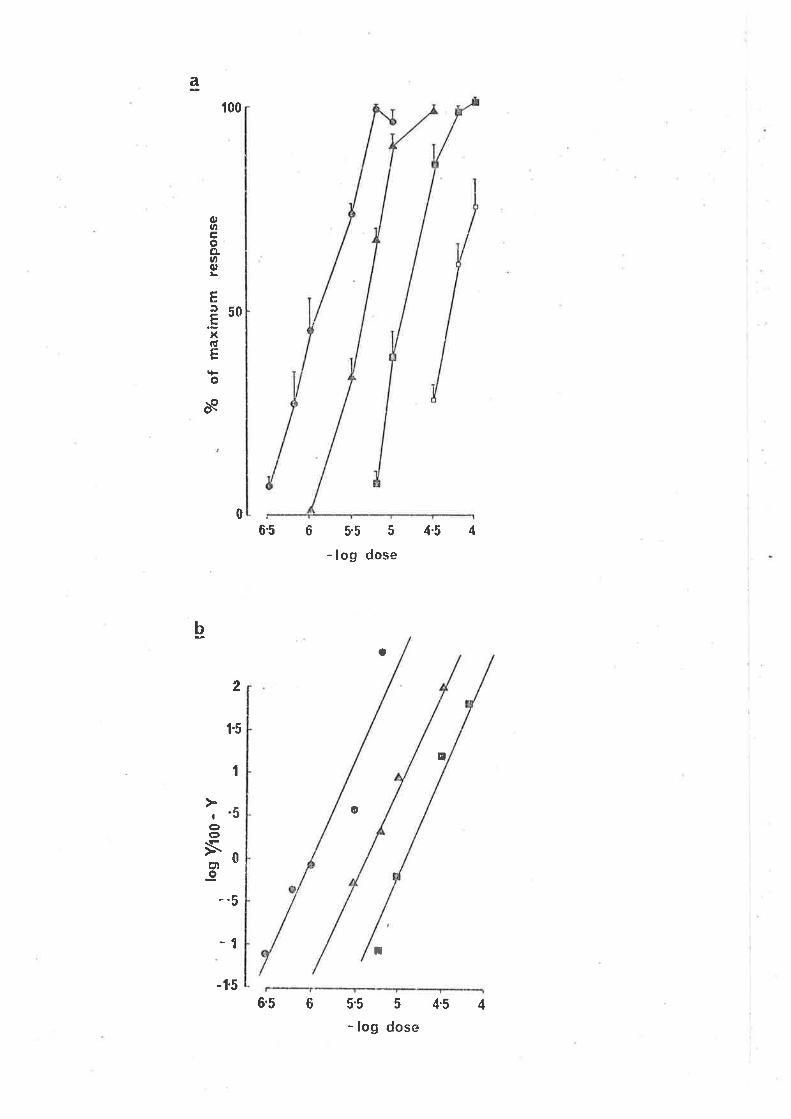
Bicuculline (a specific antagon'ist of GABA b'inding; Jchnston, 197E),
10-6, 2 x 10-6, 5 x 10-6 g/ml elicjted a parallel shjft to the rÍght
(signif icant, cl.f . -- 6, p.<0.05) in the dose response curve for GABA-induced
contractions (Fjg. 3a). In the Clark piot (Fig. 3b) bicuculline did not
alter the s'lope (not sìgnificant cl.f. = 6, PtO.05), "lut caused a parallel
d'i spì acenrent, suggest'ing o conrpeti tì ve ac'bi on of bi cucul I j ne at GABA
receptors on the myenteric neuy'ones involveci jn the ì"esponse. From the
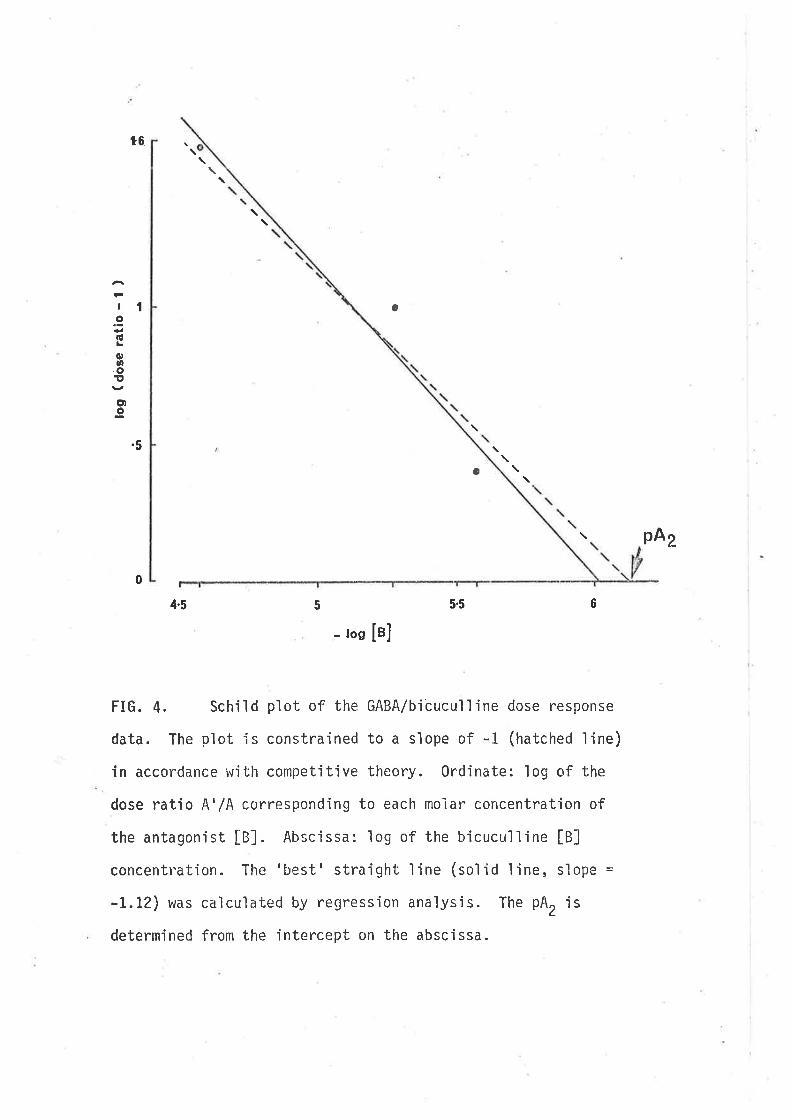
Schild plot (constrained to a slope of -1), the pA, for bicucullìne r^ias
6.12 t 1.39 (7.5 x 10-7 niol/l ) (2.8 x 10-7 g/nrl ) (Fìg. 4). Bicuculline
itself has some cholinerqjc character which conce'!vabty could be responsjble
for the observed actions on GABA-induced responses. To exclude this

,FIG. 3.
(a) The effect of bicuculline on the concentration
response curves for the contraction e'l'icìted by
GABA i n the guì nea-p'i g i 1 eum. (O Responses 'in
normal solution; (f) responses to GABA 20 min aft,er
the addition of bjcuculline, 10-6 g/ml to the bathìng
soltrtion;, (F) in the presence of bicuculline, 2 x 10-6
g/ml; (tr) 'in the presence of bicucul'line, 5 x 10-6
9/ml. Each poìnt is the nrean and standard error of
responses obra'ined in 5-8 preparat'ions.
(b) Clark plot of the bicuculline induced antagonisrn
of the GABA concentration response curve. (6)
Control responses; (^) responses in the presence
of bicuculline, 10-6 g/m1; (l) 2 x L0'6 9/ml.
3100
oangoctanq,L
È!50E.T
túç
o
ñ
5-5 5
- log dose
4.5 4
E
66.5
0
2
1.5
-.5
-1
bo
o
¡
5.5 5
- log dose
f
5
0
IIosCDo
4.s 46.5 0
-15
t6
I G
.5
3
6
- ros [s]
FIG. 4. Schiìd plot of the GABA/bicuculline dose resÐonse
data. The plot ís constrained to a s'lope of -1 (hatched fine)
in accordance with competitive theory. Ordinate: log of the
dose ratio A'/A corresponding to each moiar concentration of
the antagonjst [B]. Abscissa: 1og of the bicuculline [B]
concentration. The'Lrest'straight line (so1id line, slope =
-L.tz) was calculated by regression analys'is. The pA, is
determined from the intercept on the abscìssa.
I
.9ct
oo.ot,oo
pAa
0
5.554.5
GABA Ach GABA
Bicuculline
Ach
I min
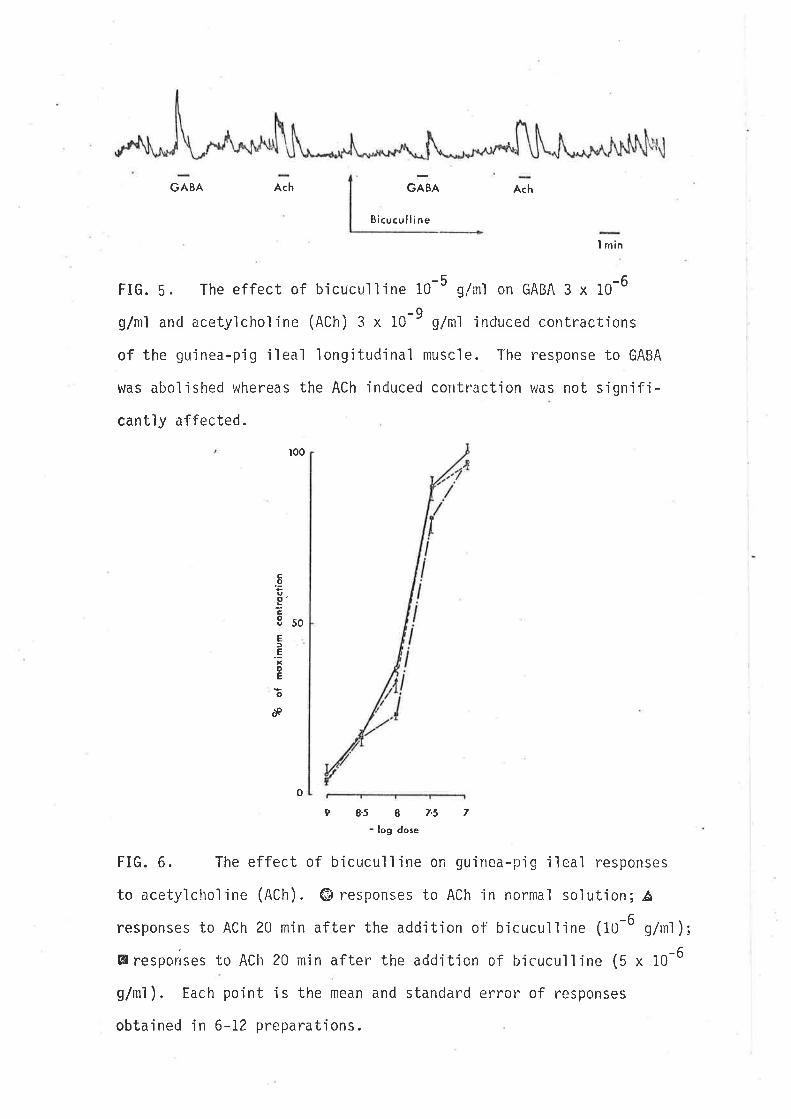
FIG" 5. The effect of bicucull'ine 10-5 g/mì on GABA 3 x 10-6
g/ml and acetylchol'ine (ACh) 3 x 1C-9 gim'l induced contract'ions
of the gui nea-pi g ì'l eal 'longi tudi nal musci e. The response to GABA
was abol'ished whereas the ACh induced contractjon vras not sign'if i-
cantjy affected.
ì00
9 8.5_ I 7.5 7
' log dose
FIG. 6. The effect of bicuculline on guinea-pjg ì1ea1 responses
to acetyìchol ine (ACh). @ responses to ACh in nornral solution; A
responses to ACh 20 min after the addition of bicuculline (tO-6 g/rnl);
øresponses to ACh 20 min after the addition cf bicucull'ine (5 x i0-6
g/ml). Each point is the mean and standard error of responses
obtained in 6-12 preparatìons.
c.9
o'a9soEt
.ExoE
o
æ
o

Bicuculline
FIG. 7 . The action of bicuculline 5 x 10-6
responses of ìongitudinal strip of guinea-pig
stimula.tjon. Field stimulation 0.1 ms, 30 V,
Bicuculline had no significant effect upon the
g/nl on
il eum to
0. 1 Hz.
twi tch
the
field
response.
HAHA
GA8¡
GAEA
8K
Ach
GÀBA
GABA
GABA
8K
A<hGABA
Pic ¡oloxinin
I min
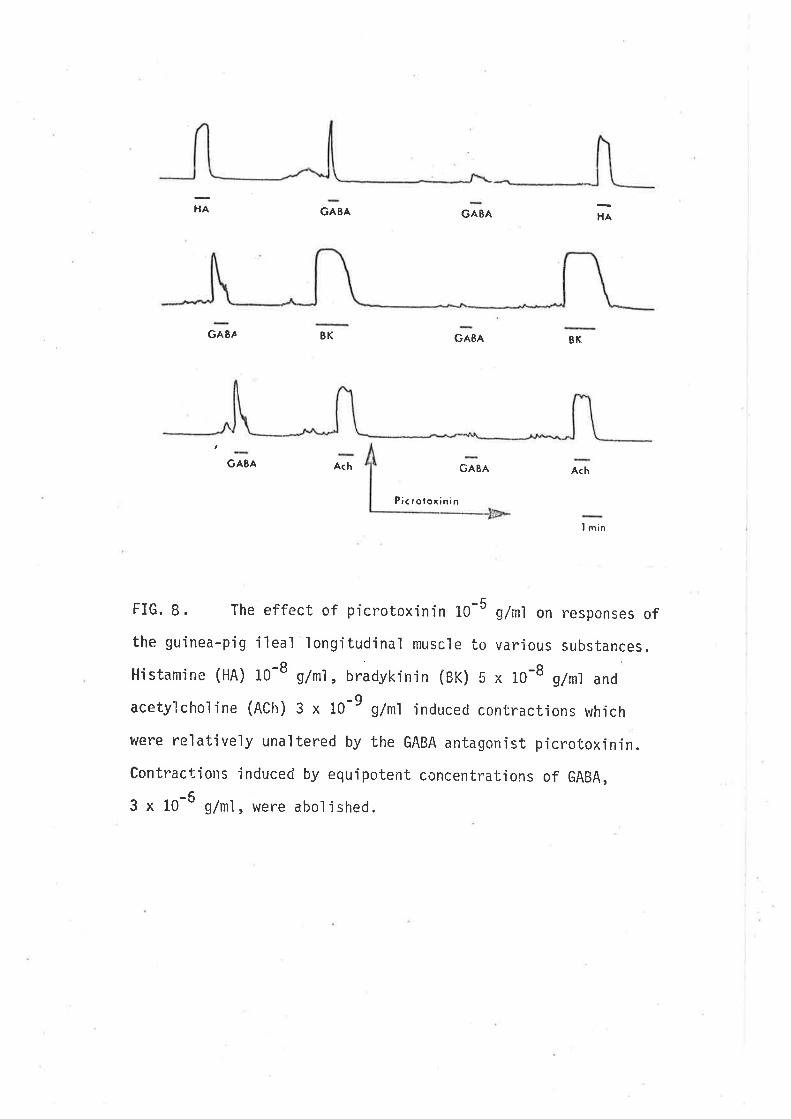
FIG. B. The effect of pÍcrotoxinin 10-5 g/ml on responses of
the guinea-pig ileal ìongitudinal muscle to various substances.
Histamine (HA) 10-8 g/mì, bradykÍnin (BK) 5 x 10-B g/mt and
acetylcholine (Ach) 3 x 10-9 g/mì induced contractions wh.ich
were relatively unaltered by the GABA antagonist picrotoxinin.
contractiorrs induced by equipotent concentrations of GABA,
3 x 10-6 g/mì , were abol i shed.

FIG. 9.
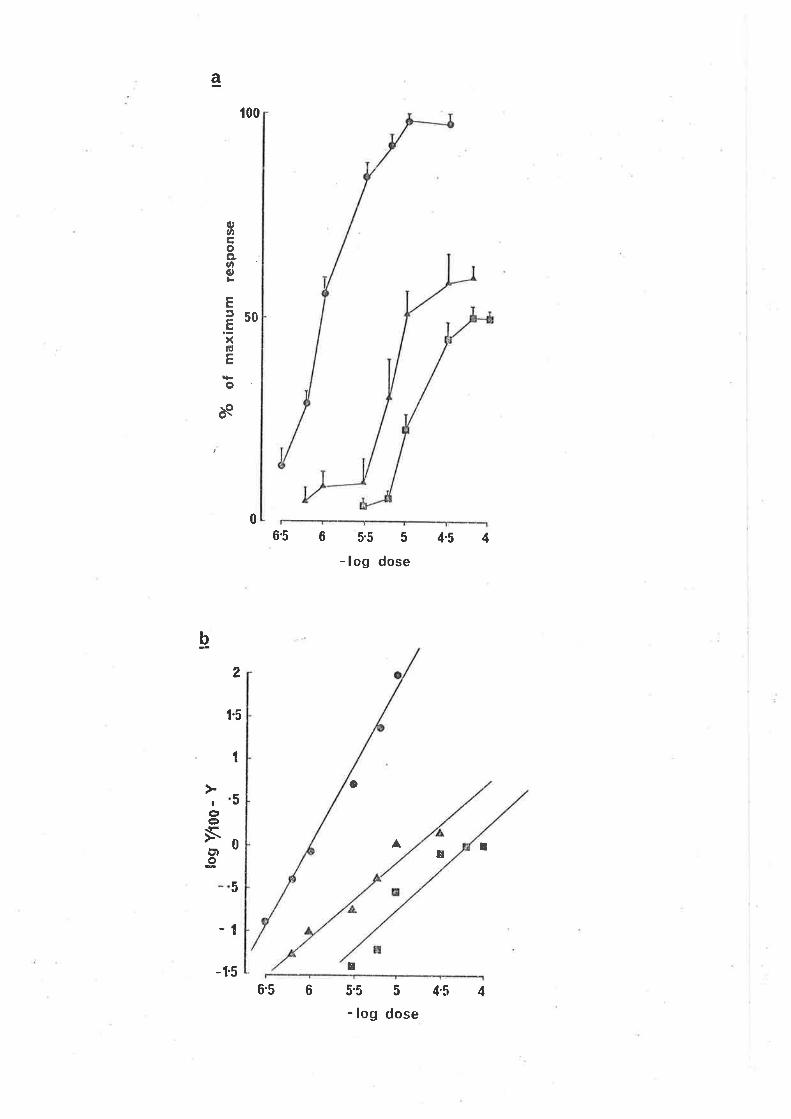
(a) The effect of pÍcrotoxinin on the concentration
dependent responses of the guinea-pig ileum to GABA.
(Q) Responses i n normal sol uti on ; (¡) responses
20 min after the addition of picrotoxin'in, 2 x 10-6
g/ml ,to the bathing solution; (¡) responses in the
presence of picrotoxinin, 4 x 10-6 g/ml. Each point
is the mean and standard error of responses obtained
in 5-8 preparations.
(b) Clark p'lot of the picrotoxinin induced antagon'ism of
GABA concentration response curve (S) contro'l responses;
(A) responses in the presence of picrotoxinin, 2 x 10-6
glml; (¡) 4 x 10-6 9/n1 .
e
100
0
oaàqoctoo
E?s0g'itgE
o
òe
5.5 5
- log dose
o
A
E
5.5 5
- log dose
4.5 4
E
66.5
b
?
I
1.5
5
-1
¡
5
0
Ioesgl-9
4.5 46.5 6
-f.5
T Ach GABA GABA AchT GABA
Pi¡etanideI min
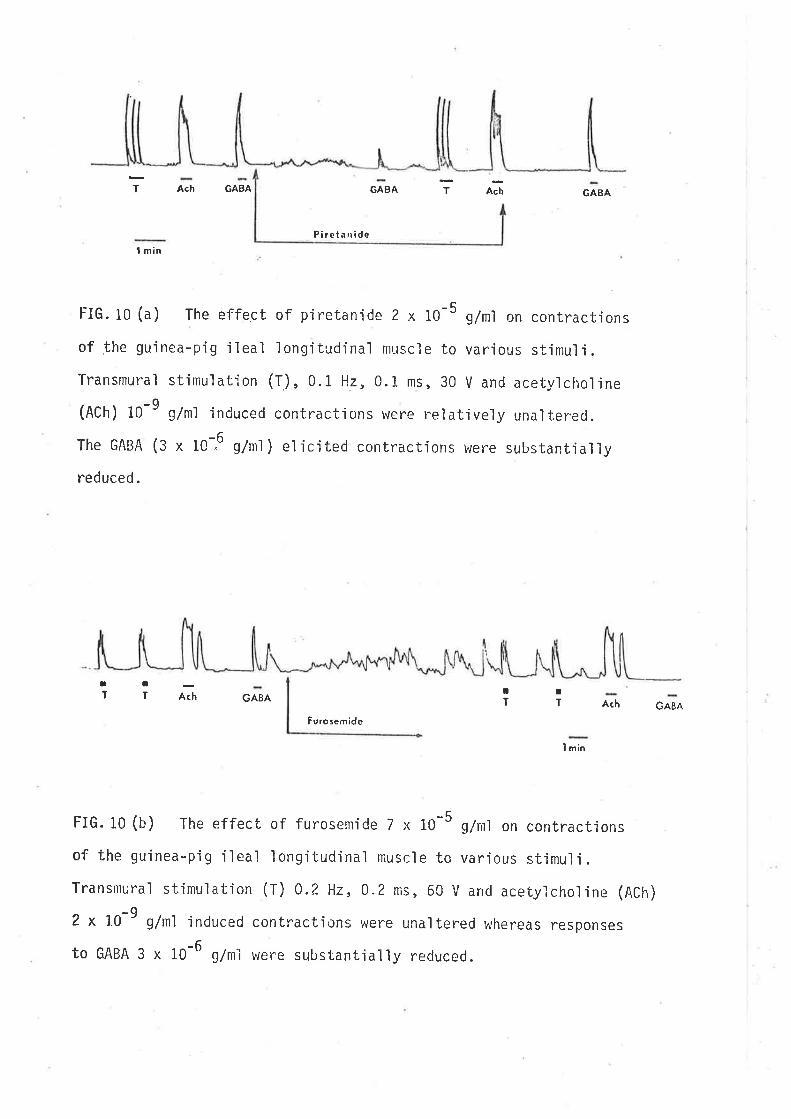
t-IG.10 (a) The effect of piretanide 2 x l0-5 g/ml on contractions
of the gu'inea-pig ileal longitudinal muscle to various stjmuli.
Transmural stimul ati on (T) , 0 .r Hz, 0. 1 ms , 30 v and acet,vl chol i ne
(Ach) 10-9 g/ml induced contrac'uions were reìativery una'lterec.
The GABA (3 x 1o--6 g/ml) elicited contractions were substantìaììy
reduced.
GABA¡T A(h GABA
Furosemide
I min
FIG.10 (b) The effect of furosenride 7 x 10-5 q/nrl on contractions
of the gujnea-pig ileaì longituclinal muscle to various stimuli.
Transmural stimulation (T) 0.2 Hz, o.z ms, 60 v and acetylcholine (Ach)
-o2 x 10' g/n1 induced contracti¡f,ns were unaltered whereas responses
to GABA 3 x 10-6 g/ml were substantìa'l'ly reduced.
IT a
TAch
tT

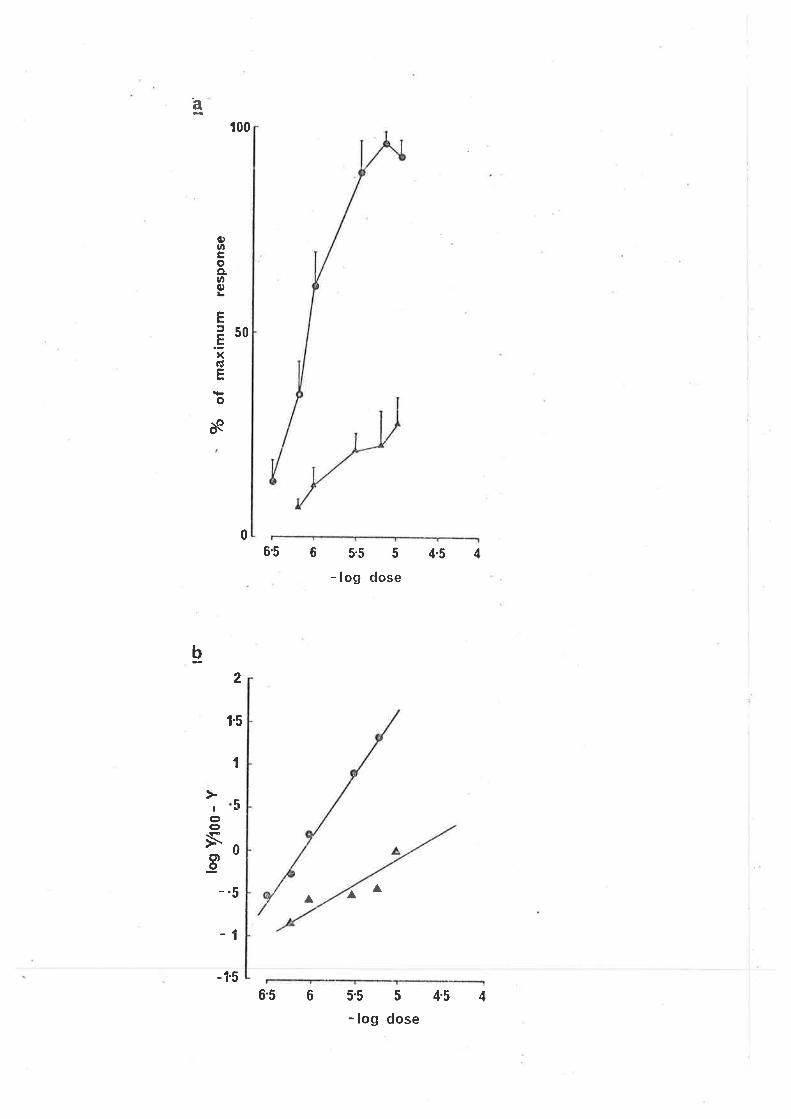
FIG. 11.
(a) The effect of piretan'ide on the concentration
response curve for GABA in the guinea-pig ileum.
C responses of the longitudinal muscle in normal
soíution; A responses 'in the presence of-^
.piretanide 5 x 10 " g/ml" Piretanide was
present in the bath 20 min before add'ing GABA.
Each point is the mean and standard error of
responses obtained from 4 preparations.
(b) Cìark plot of the piretanjde induced antagonism
of the GABA concentration response curve,
O contro'l responses, A responses in the presence
of piretanide.
a
!'vttroCt3hc,
ElsoÈ'i.!E
o
rOC'\
0
5.5 5
- log dose
4.5 466'5
þ
100
2
1.5
f
5
0
IeeSo)o
-.5
-1
^ A
5.5 5
- log dose
4'5 46.5 6
-15

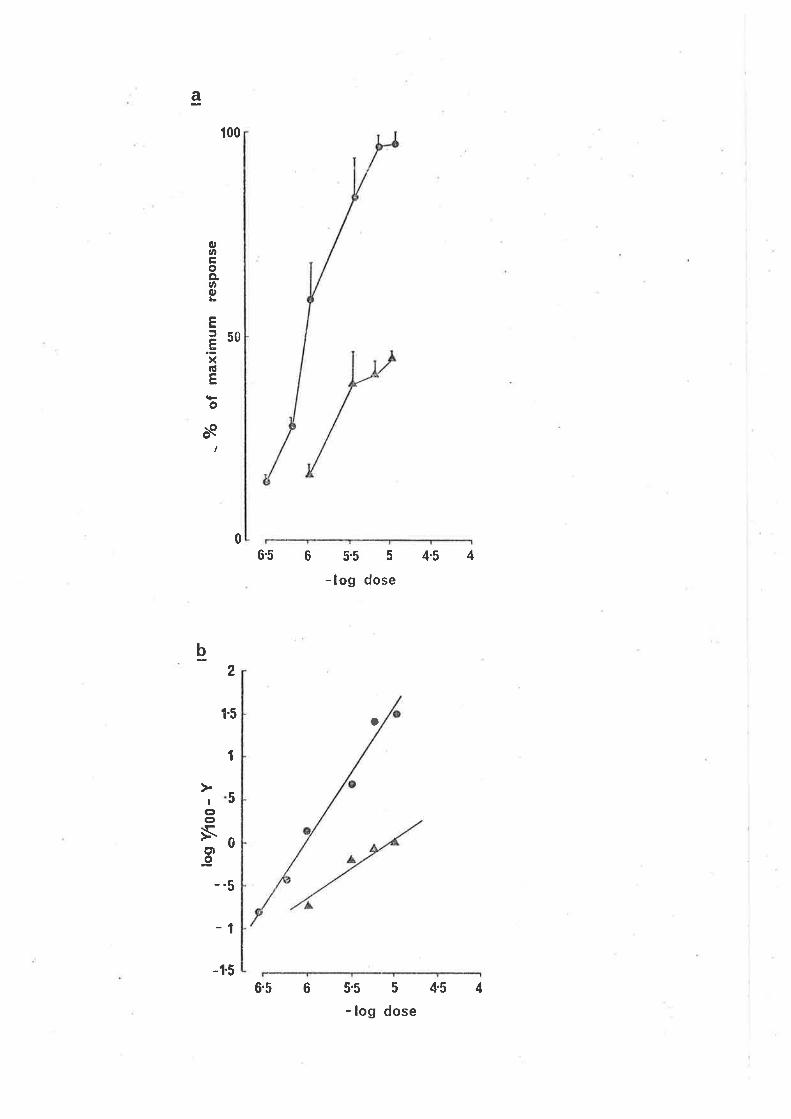
FIG. L2.
(a) The effect of furosemide on the concentration
response curve for GABA in the gu'inea-pig i1eum.
Q responses of the longitudinal muscle in normal
solution; A responses in the presence of
furosemide 2 x 10-5 g/ml. Furosemide was present
in the bath 20 min before adding GABA. Each point
is the mean and standard error of responses obtained
from 4 preparatìons.
(b) Clark plot of the furosemíde induced antagonism
of the GABA concentration response curve,
O control responses, A responses in the presence
of furosemide.
4
ooCocloob
tr?s0È'irú
E
o
-oo\
100
1.5
I
l
0
66.5 5.5 5
- log dose
4.5 4
b2
o
¡C'oSo)I
5
0
-.5
-l
5.5 5
- log dose
-156.5 6 4.5 4

31
possibility the effects of bicuculline on contractions elicited by
acetylcholine (ACh) or by stimulation of intrinsic cholinergic neurones (Fig. b)
were tested, but no sign'ificant (d.f.= 6, p>0.05) alteration of the dose
response curve for ACh in the presence of bicuculline, 10-6 or 5 x 10-6
9/ml (l- jg. 6) nor alteratjon of the chol'inergic twitch response el jc'ited
by transmura'l stimulation (Fig.7) was observed.
The specificity of picrotoxìnin for the GABA-induced contractile res-
ponses of the ileum is shown in Fig.8. Whereas GABA responses were abolished,
contractions elicited by acetylchoìine, hjstamine or bradykinin renlained
relativeìy unaffected. Picrotoxi nin, 2 x 10-6 and 4 x 10-6 g/mì produced
a shift to the ri,ght (s'ignificant, d.f. = $, p.0.05) of the dose response
curve for GABA-induced contractions in the ileum and also had the effect of
reducing the maxjmum response (F'ig. 9a). The slope of the Clark plot (Fis.
9b) was altered (significant, d.f. = g, p.0.05), thus the action of picro-
toxinin appeared non-competitive, in keep'ing w'ith the proposaì that it acts
to block GABA-induced chloride conductanceat the'ionophore (Johnston, 1978).
An interest'ing question raised by thìs study concerns the ionic
basis of these excitatory actions of GABA in the guinea-pig ileum that
are antagonised by bicuculline and p'icrotoxin'in, since the latter
antagonist would suggest that a chloride ionophore might be involved
(Curtis and ,lohnston, I974). Although excitatory, the responses to GABA
were nevertheless chlorìde-dependent as shown by the use of d'iuretic
chloride-ion channel blockers (Bretag et aL., 1980). Furosem'ide, z x 10-5
g/n1 (Nicoll, 1978) or piretanide, 2 x i0-5 g/ml (Zeuthen et aL., 1978)
substantially reduced GABA-induced responses but vlere without effect on
those elicited by ACh or transmt¡ral stimulation of the ijeum (Figs.10a,
b). The effectiveness of the channel blockers in antagonizing GABA
actions can be seen in the dose response curves (Figs. 11 and 12), where,

5¿
in both instances, there was a shift to the right (signìficant, d.f. = 6,
p<0.05) and a reduction in the maxìmum response, indicating a non-competitìve
nature for the antagonìsrn by these Cl- channel blockers.
Discussion
The present results confirm and extencl the fjndings of Krantis et aL.
(1980) tnat GABA-jnduced contractjons of the guinea-pig iìeurn are not the
result of d'irect act'ion'in the smooth rruscle, rather they result from
stirnulatory actions on cholinergic motor neurones since they are blocked
by muscarinic antagonìsn and tetrodotoxjn. These actions do not involve
pre-gangì ionic cholinergic nervous transmission, be'ing unopposed by
nicotin'ic antagonjsm (Krantis et e.L,, 1980) which further suggests that
GABA direct'ly stimulates the intrinsic cholinerg'ic motor neurones.
Excitatory actions of GABA in peripheral nervous tissue are well
documented. Intracellular and extracellular recorclings from the superior
cervi cal gang'l j on and dorsal root gangl i on of the rabbi t, cat, rat, bu'ì I f rog,
and duck embryo show jn every jnstance a GABA-induced cìepolarizat'ion
through an increase'in membrane permeability to C1- (De Groat, 1970, L972;
De Feudis, 1977). Thjs effect is due to activatjon of'true'GABA-receptors
in that it is ìnrjtated by GABA analogues and blocked by picrotoxin and
bicuculline (De Groat, L970; Bowery and Brown,1974;Obata, 1976). However,
this depolarization cioes not cause sympathetic neurones to discharge.
Rather, it is characteristic of presynaptìc inhibition where the GABA-
induced depolar"ization of nerve terminals results jn a reduced transmitter
ouiput. Transmitter release by this GABA-induced t.crminal depolarization
has never been described and is unlike'ly to be the cause of the

33
presently described persistent stimuiation of jleal cholinergic neurones.
Gràfe et al. (1979) showed the GABA induced depolarjzation of myenteric
neurones to be through a pìcrotoxin sensitive mechanism which leads to
myenteric exc'itation wjth firing of the intrinsic cholinerg'ic motor neurones.
Since the act'ion of picrotoxìnin seen here was reversed by wash'ing the
ileal segment with normal Krebs solutjon, it seems unlike'ly that the non-
competitive nature of picrotoxìnin'is caused by its irreversible binding
to the GABA-receptor. Alternatjvely, pìcrotoxinin could be interfering
with the ionic mechanisms under"lying the GABA'induced depolarization.
Indeed there 'is consiclerable ev'idence to suggest that picrotoxin (an
equimo'lar mixture, of p'icrotoxìnin and pìcrotin) (Zukin e-b aL., 1974;
Curtis and Johnston, 1974; Snyder atrd Goodman, 1980), blocks GABA
inhibitory synapses in vertebrates and invertebrates non-competitively
by actions at the Cl- ionophore (Krniev'ic, L974; Shank et aL., L974) and
not at the GABA receptor although there is some evjdence for a direct
receptor action (De Feud'is, 7977). In the present study furosenlide and
piretan'ide selectively antagonjsed GABA stimulatory actions 'in the guinea-
pig ileum. Since furosemide (Montoreano et aL., 1975; Nìcoll, 1975,1978;
Brazy and Gunn, 1976) and Piretanjde (Zeuthen et aL., 1978) Lrlock chloride
transport, and the Cl- 'ionophore (Nico1'l , 1978), it 'is very l'ikely that
picrotoxin actions in the intestjne are at the Cl- ionopho'ne that is
regulated by the GABA-receptor. Whether both furosemide and piretan'ide
exert their effects by either blockìng the jonic channels,ot" the chloride
pump remains to be rescrlved. It would be interesiing to look for a chloride
pump'in neurones of the guìnea-pig iìeal myenteric plexus, since excitation
of celìs by GABA through a chloricie mechanism necessitates a high intra-
cellular Cl- ùo produce depoìarization via an increased outward Cl-
conductance (Obata, L976; Nicoll, 1978).

34
Where GABA has been proposed to be a true excjtatory transmitter, in
the seã urchin tube foot (Florey et aL., 1975) and ap'lysia (Yarowsky and
Carpenter, I9V7) its actir¡ns did not involve any alteration in Cl- perme-
abiìity, rather they appeared to be l,la+ or K+ dependent. Therefore, the
findjng in this study that stimulation of myenteric neurones by GABA
occurred through a Cl- dependent mechan'ism is unexpected. Although the
ionic mechanism mediating GABA effects in the sea urchin tube foot and
guinea-p'ig ileum is ciifferent, GABA actjons'in these preparations are
similar¡ the results of excitatory act'ions at chol inerg'ic motor neurones.
In addition to this action of GABA on ileal cholinergÍc excitatory
neurones, the GABA-'induced excjtat'ion of the non-cholinerg'ic, non-adrenergic
inhibitory neurones (Kranti s e'b a.L., i9B0) persumably a'lso acts through a
C'l- dependent mechanism, not examined for its cletails in this study.
Bicuculline has been reported to have a variety of effecbs other
than blocking GABA actions, such as inhibition of acetylcholìnesterase
(AChE) (Svenneby and Roberts, I9731' Breuker ancl Johnston, i975) and
potentiation of acetyìcholine action (Miller and l4clennan, I974), no doubi
by this ACht inhjbìbion, and a djrect modificat'ion of nerve axon or muscle
membrane conductance (Freeman, L973; Shank et aL., I974), Both picrotoxinin
and bicucul ì ine specif ical'ly antagonì sed GAB/ì-induced resporrses in the
ileum, although, jn contnast to picrotoxinin, antagonism by bicr,rculline
was of a compet'it'ive nature. The potency of t,his antagonist has been
characterised by the Cel^ived pAr. The simjlarity of aciion for bicuculline
in centraì nervous tjssue (Curtis and Johnston, 7974), guinea-pig large
intestine (Kranti s et d.L.-, 1980) and the guinea-pig smaìl jntestine in
this study, indicates the usefulness of the guìnea-pi g in ',titro intestinal
preparation as a model fcrr investigation of GABA-receptors.
In contrast to the present resujts Bolery eb aL. (1981) arrd Ong (1981)

35
describe inhjbitory act'ions of GABA in the guinea-p'ig intestine, a
bicuculline insensjtive depression of electrìca'lly evoked contractions of
the Íleum and distal colon. These actions mediated by baclofen-sensitive
(GABAB) receptors, are not influenced by furosemide and therefore considered
by these authors to be not Cl dependent. Recently Kerr and Ong (i981)
have shown stimulated release of endogenous GAB,A from intrinsic'ileal
myenterÍc Reurones resulting in both GABAA and GABA, receptor mediated
events. The presence of both excitatory and jnhib'itory responses to GABA
in the intestine is sim'ilar to the variety of responses to other putative
transmitter substances such as dopamìne, noradrenal ine ancl S-hydroxytry-
ptamine (Snyder and Goodman, 1980). The actions of GABA in the guinea-pig
intestine could be an example of a transmitter from a sing'le neurone being
excítatory on one target cell and inhibitory on another (i(erkut and Þieech,
1966) or an example of inhibitory and exc'itatory effects of one transmitter
on a singìe cell (Kehoe, 1972).
The sirnp'lest model for the role of 'excitatory'GABA in the guinea-pig
Íntestine is for GABA to be a transmittev'of afferents, directly relaying
on the effector neurones concerned rvith the peristaìtjc neflexes, without
intervention of interneurones. A'lso, GABA could be a transmitter of
myenteric interneurones which in turn jmpínge direct'ly upon the respective
nrotor neurones. Alternative]y, GABA could be acting through 5HT, or a
relateci amjne, as 5HT receptors are also found on these same effector
neurones (Costa and Furness, 1979). llourever, since there is at present
controversy as to the role, if any, of 5HT or a related amine jn the
intrinsic excitatjon pathways, the final model for GABA actìons cannot be
decided here. Furthermore, the polysynaptic nature of various reflex
pathways within the nyenteric pìexus ís well known and it'is quite
feasible for two different interneurones, emp'loying different transrnitters,
to intervene between the sensory and motor elements of the myenteric reflexes

36
The findings of th'is study reinforce the notion that GABA may welì
have a transmitter functíon in the mammalian enteric nervous system,
where it evident'ly acts through two specific GABA receptors, the one
coupled to a Cl- dependent excitatory mechanism, the other inhibìting
transmitter output.

CHAPTER I I I
3LOCALIZATION OF I
UPTAKE SITES
GABA HiGH AFFINITY
AUTORAD I OGRAPHY
H]
BY

37
Introductiorz
Tracing suspected neurotransmitter substances e.g. monoamjnes, g'lycine
glutamate and gamma-aminobutyric ac'id (GABA) has been carried out using
various methods, includìng biochemical (Kuriyama et aL., 1966; Obata et aL.,
1970) ancl neurophysìological (Obata et aL." !967; Iversen et czL., 1971).
Autoradj<lgraphy is now recogn'ized as a powerful tool for mappÍng GABA-
ergic neurones (see Snyder et aL., 1970; Neal, 1977; Johnston and Iversen,
I97l; Höktelt and Liundahl, L972). This technique is based on the
accumulat'ion of radioactive'ly labelled, exogenousìy applied substances to
the nerve cells by spec'ific scdium dependent hjgh affinity uptake mechanisms
localized to nerve cell nlembranes. T'issue relatjveiy rich'in synapses
for a suspected transmitter, have a highly active high affinity membrane
transport system wh'ich in contrast to low affìnity processes, are of
considerable physiologìca1 importance as an inactivation mechanism for
the t.ransmitter after its actjon at the receptor (lversen, 1967;0rrego,
t979; Fagg and Lane, 1979). High affinity uptake mecharrisms for GABA
have been identjfied in the mammalian central (De Feuciis, L975; Johnston,
L978; Fagg and Lane, 1979) and peripheral nervous systems (Martin, 1976),
where GABA is localized to nervous tissue. The [3ti]-engn transported by
this mechanism is accumulated in the same pool as the endogenous GABA,
rnainly in the synaptosome fraction although uptake of [3tl]-GABA also
occurs into cell bodies of a number of brain neurones (Schon and Iversen,
1972). This uptake can be distinguished from bjnd'ing of GABA ident'ifying
receptors oll many ne'drones, sincê binding unì ike GABA. uptake occurs at
OoC and does not requ'ire the presence of sodjlim icns.
The app'lication of autoradiography for the investigation of GABA in
the central nervous system, has jnvolved several meihoCologies of isotope
administnation. Homogenates incubated in [3U]-GRBA solutions can be usecl

38
to obtain information about the local'ization of [3H]-engn to nerve
terminals (Iversen and Bloom, 1972). This allows the deterntination of the
percentage of GABA accumulating termjnals in different areas, however,
there is a distinct lack of histological informat'ion. Thìn slices of
brain tissue incubated ín uitro witfr [3H]-GABA, have been used to g'ive
anatomjcal informatìon about the localization of uptake sites (Iversen
and B'loom, 197?.1 Hökfelt and Liungdahl , 1970). Hovrever, thi s in uítro
technique generalìy results jn poor morphology, due to a partial
destruct'ion of norrnal cell bodies and swelf ing of perìneuronal eletnents
(e.S., glia) (Hökfelt and Ljungdahì , L972). The specific uptake mechanism
of GABA conta'inìng neurones may a'lso be altered by clamaging neuronaì
membrane, or botli perikarya and/or its processes during the dissection
and sl'icing procedure. Furthermore, penetration of the isotope from the
surface i s I'imi ted and coul d I ead to erronous negative resul ts (l-losl i and
Hosli, lg72)" A third method of [3H]-GABA administration ìs by ín uiuo
injections (¡iökfelt and Ljungdahl , Ig72). [3H]-enen does not readily cross
the blood brain barrier (see Rapoport, 1976), therefo,'e the 'isotope must
be administered by intraventricular or intracer"ebral iniection" Intra-
ventricular adnlinistratjon must be coupled with pretreatment with amino-
oxyacetic acid (A0AA), to prevent metabolism of the ìsotope (t^Jallach,
1961). Under these conditions GABA containing neurones are labelled
(Ljungdahl and Hökfel,t, 1973), however, there is lirnited d'iffusìon of?
["H]-GABA from the ventricular system to the nervous perychyme and only
the most superficiaì cells are labelled.
Intracerebral injection results in hìgh isotope concentration in the
tissue immediately surrounding the cannula, togetlier with a swel'lìng of
the extracellular space. Under these circumstances signìficant radio-
activity, together with good morpho'logy is confined to a narrow zone

39
peripheral to the site of iniection.
Another approach in the identjfication of GABA-con'[ain'ing neurones
is using t'issue culture, where the labelled amino äcìd can be directìy
added to the culture (Hosli and Hosli, L979). Although diffusìon of the
transmitter in culture occurs more rap'idìy than in sl'ices, thus a1lowìng
shorterincubation times, such preparations are not representative of
the brain region under investigation since normal cellular investment. is
not mainta'ined (e.g., transp'ìants are devoid of there gl ial envelope)
(Hosli and Hosli , 1978, 1979). Th'is ìs part'icular'ly relevant when tracing
GABA since thìs amino acid is also accumulated by a hìgh affìnity mechanism
to perisynapt'ic glìa1 cells (Hosli and Hosji, 1972), whìch may represent
an 'important inactl'vation pathway for GABA (Henn and Hamb¡¡rger, 1971).
Furthermore, these preparations do not allow delineation of the anatomical
arrangement of GABA neurones wjthin the CNS.
Autoradiographic evidence, Schon ancJ Ke'l'ly (Ig75) shoiv ttre [3H]-eRgR
high affin'ity transport systems in neurones and gfial ceils to djffer in
their substrate spec'ificity, a1lc',wing d'ifferentiation of the uptake sites.
Uptake into neurones is blocked by L-2,4-diam'inobutyric acid (L-DABA), and
n'ipecotic acid wh'il st ß-alanitre is a potent inh'ibitor of gl ìai uptake
(Iversen and Johnston, L97I; Iversen and Ke1ly, 1975).
Recently, Jessen et; aL. (L979) have shown high affin'ity uptake sites
tor [3H]-GABA in cultured myenteric neurones of the gu'inea-pig intestine.
In addjtìon, glutamic acid decarboxy'lase (CAD) and endogenous GABA were
also demonstrated to be present in the myenterÍc p'lexus.
Laminar preparations of thjs nerve netwot^k consistìng of the myenteric
pl exus at.tached to the l ong'itudi nal muscl e I ayer al I ow a more detai I ed

40
examination of nerve fibre ramification with'in this ìayer (Gabella and
Costa, L967; Furness, 1970). Such preparations of the intestine have
been used for the immunohistochemical (Costa et.aL., 1980; Schultzberg
et aL., 1980),fluorescenceh'istochem'ical (Furness,'1969; Costa and Furness,
7972;0lsen et aL., 1976), and autoradjographic (0iab et aL., 1976),
investigations of the distribution of various.substances within the
mammalian enteric nervous system. The generaì organizatìon of the mammalian
myenteric plexus is reminiscent of the Cl'lS rather than other autc¡nomic
gang'lia. l-lowever, the wall of the gastrointestinal tract prevents no
barrier tothe diffusion of substances from the bathing soìution to the
various intrinsic nert,e networks, lending itse'ìf to in'oítro labellìng for
autoradiography
Evidence is presented here to show the efficacy of [3H] u, un
autoradiographic marker ìn laminar preparations of myenteric plexus using2a
["H] proline and ["H] leucine, two amino aciCs known to be actively taken
up and transportecl by neurones. In aclditìon, the d'isposition of [3Hl-
GABA accumulating neurones within the guinea-pìg enteric nervous system
was deter"injned by l ight microscopic autoraciiography using specific?
inhib'itors of ["H]-GABA high affìnity uptake mechanisms. The uptake of2)
["H]-eRgn was compared with that for ["H] Ê-alanine which is substrate
specific for g'liaì cell high affín'ity uptake sjtes.
Methods
Preparatìons of myenteric plexus were obiained from freshly killed
guinea-pigs of either sex, weighing 250-400 g. Segments of small and large
intestine B-10 cm 1ong, were quickìy removed and placed into Krebs solution

41
(the composition as described in Chapter II) maintajned at (37'C) and
gassed with a lnixture of 95% 0, and 5% C}Z. Two tissue preparat'ions
were used.
1. Longìtudinal muscle with attached myenteric plexus tlissected
free from the segments of intestine and cut ìnto short lengths (laminar
preparations) (Fig. 1). Individual tissues weìle pre-incubated for 10 min
in 350 ul Krebs at 37oC.
To block non-neuronal and neuronal uptake of 3H GABA (Schon and
Kel1y, 1975), Ê-âlanjne, L-2,4-diaminobutyric acid or n'ipectoìc acid wene
added to give a final concentration of tO-3 tvt, and the tissues were
incubated for a further 5 min r^rith shakìng. Labelling of h'igh affinity
uptake sites was carried out usìng either tri tiated GABA (66 Cilnrmo'ìe;a
2,3-3H GABA) or trítiated Ê-alanine (32 Cilmmole; ß-[3-3H] alanine) at
a final concentration of 5 x 10-9 M in the presence of uptake 'inhibitors
where approprìate. in a further series of experiments, tritiated L-
proline (40 Cilmmole; 5-31-i proline) or tritiated L-leucine (52 Cilmmole;
4,5-3H leuc'ine) were aclded to gìve a final concentration of tO-B N.
TÍssues were incubrated r^¡i tl'r shakìng for 20 mìn, then washed in fresh
Krebs solution, pìaced on subbed slides and fixed wjth 4% glutaraldehyde-
phosphate buffer. GABA is a small easily diffus'ible substance and has
to be immobil jzed during the preparation steps of autorad'iograplry. To
avoid dislocation of l3l-t:l-gngn together with good tissue preservation,
gìutaraldehyde js routìne1y used as the fixatjve (Ljungdahl et aL., 1975).
Glutaraldeh.vde has the added advantage of b'indìng GABA to the tissue
affordìng high retention of [3H]-gnen during fixatjon and dehydratìon steps.
2. Short segments of rvhole intestine 2-3 cm long were treated for1
["H]-GABA autoradiography as descrjbed for the laminar preparations,
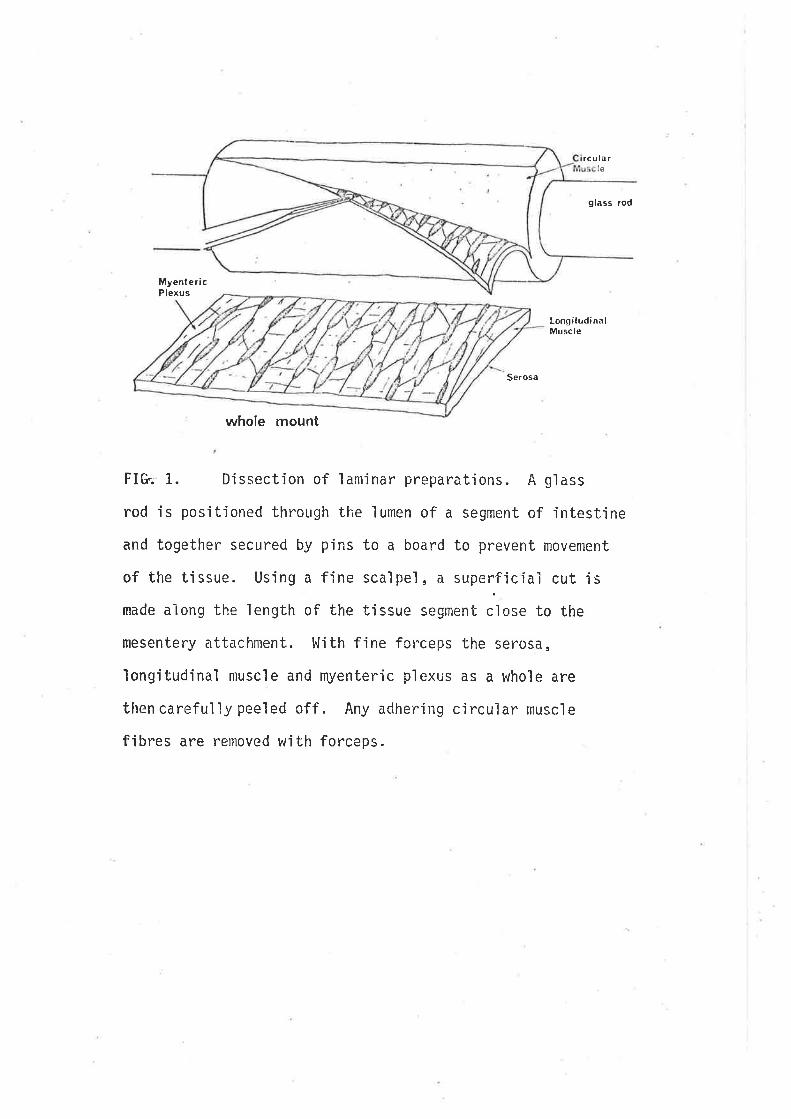
rcularle
glass rod
MyentericPlexus
Longiludi nalMuscle
Serosa
whole mount
FIt+j 1. Dissection of Iaminar preparations. A Eiass
rod is positíoned through the ìumen of a segment of intestine
and together secured by pins to a board to prevent movement
of the tissue. Using a fine sca'lpel , a super.ficÍal cut is
made along the length of the tissue segment close to the
mesentery attachment. l.lith fine forceps the serosa,
longitudinal muscle and myenteric pjexus as a whole are
then carefully pee'led off. Any adhering circular muscie
fibres are removed with forceps.

42
however, before fixat'ion a g'lass rod was posit'ioned through the lumen in
order to ma'intai n the i ntegri ty of the 'tubul ar' nature of the 'intesti nal
segment. To determine v¡hcther enzymìc degradation of GABA could alter
the uptake and distribution of 3H GABA, seiected preparations were incubated
in the presence of amino-oxyacet'ic acid (nOnn, 10-5 M), a specific inhibitor
of the GABA metabol'izing enzyme GABA-transaminase (GABA-T) (Da Vanzo et aL.,
1964). Following fixation the 'tube' preparations vjere renoved from the
gìass rod, washed, dehydraùed and embedded in paraffin. t.ongitudinal
sections (20 urn) cut perpendicular to the circular ntuscle celìs, were
pìaced on subbed slides for autoradiographic treatment.
Autoradiography
For l'ight microscopic autoradiography Ilford l(2 emul sìon was used
because of ìts adequate sensitivíty for use with tritium. For coating,
the emulsion uras melted in a water bath at 41"C and d'iluted in the ratio
of one part ernu'lsion to one part dist,jlled HrC. The siides fronr both
tissue preparations were dipped individually for i0 sec then p'laced on
a ìevel cold p'!ate to set for 15-30 min. Th'is pl ocedure ensured an
emulsion coal; of uniform thickness. The slides r^¡ere then removed to a
bench and allowed to set for a further 3 hours wher"eupon they were packed
in light tight boxes containing dessicant, seaìed, encased in a light
proof plastic bag and stored'in a refrigerator at 4"C, for 14 days.
Slides were deveìoped for 4 min in fresh undjlutecl Kodak Dig deve'loped
at 20"C. After rinsing ìn disti'lled HrO the slìdes were fixed for B mjn
in sodium th'iosulphate, r'insed in distìllecl Hrü, then icashed in gentìy
running tap water for at least 15 min. Silver grains were almost tota'lly
absent from segments of nerve tissue treated for autoradiography without

43
)tritium ["H] present in the incubating medium, suggesting chemographic
or phys'icaì artefact to be minimal.
Statning
In order to rejate the depos'ition of siìver grains associated with)
["H] uptake to the under'ly'ing cytoarchitecture tissue, preparat'ions were
stained with either cresyl vjolet or neutral red. This was carried out
either directly after wash'ing or the tissues were dried and stajned at
a later t'ime. Further morphoìogica'l characterization of the nerve cell
types accumulating [3H] amino acids,was carried out by comparing auto-
radiographs with laminar preparations of the myenteric pìexus not treated
for autoradiography, but stained by the Nitro BT method (see Chapter IV).
Compounds used ivere: ß-alan jne, amino-oxyacetic acid (A0AA) , c'is-3-
am'inocyc'lohexanecarboxyì ic acid (ACHC) , L,2-4-dianlinobutyrìc ac"id (L-DABA)
(a11 Sigma). [3H]-engn, [3H] ß-alanine, i3nl leuc'ine, [3u] pr.otìne (alt
Radiochemical Centre, Amersham).
ResuLts
3uptake o¡ ín-pooLi.ne and
"I-Leuaine into myenteríc íatnina.n preparations
Light microscopic autoradìographs showed an extensive uptake of both
3H-proline and 3H-leucine into laminar preparations of the myents,eric
(Auerbach's) plexus taken from all levels of the small intestine. No
sÍgnificant labelling was seen outside the plexus, on smooth muscle or

44
blood vessels. The most characteristic feature of the autorad'iographs
was the presence of numerous c'lusterings of silver grains, found more
particu'larly over the gangìia and primary plexuses. Many of these were
very dense and recognisably over neuronal cell bodies inciuding emergent
processes, most of which were dendrìtes, aìthough some emergent axons were
labelled for a short distance (F'igs. 2,3, 4). T'he extent of the cell
labelling became more obviours when these autoradìographs were conposed
with similar lam'inar preparatìons sta'ined by the Nitro-BT nrethod (F jg. 5).
In both, a great variety of neurones could be reccgnised, varying from
uni pol ar to mLtl ti po'l ar w'i th di sti nct j ve dendri ti c ran if i cati ons , but a
classificatjon of these cell types was not attempted.
Less dense clusterings of silver grains were also to be found
scattered over the ganglia and the primary and secondary p'lexuses. Littledetail of the underlying labelled structures could be made out, but there
was.sufficient variation in form to suggest that these, too, were labelled
cell bodies. In the tertjary plexus, at the intersections of the p'lexus,
similar accunulatìons of silver grains were commonly found.some of whìch
were rather jntenseìy 'labelled such that processes could be recognised
and the'ir form was most suggestive of labelled cell bodies (Fig. 6).
In addjtion to the accumulations of silver grains obviously related to
cellular uptake o'f these tritiated amino acids, there vras also a faint,
diffuse labelìiirg found over the ganglia and p'lexuses. This was particularly
obvious over t,he gang'lia and primary plexuses, but could also be seen over
the secondary anci tertiary pìexuses. In some a!eas the silver grains were
sufficiently nurnerous that Auerbech's pìexus was r:learìy recognisable, but
the more dense accumul at'ions due to cel I I abel ì ì ng were neverthel ess a'ìways
recognisable against the relatively faint background labellìng the nature
of which is unknown.

Autoradiographs showing uptake of Imyenteric plexus.
3Hl into guinea-pig ìleaì
FIG. 2. t3Hl leucine (10-8 M). Labelled cell bodies are
located throughout the plexus. Scale bar represents 50 pm.
FIGS. 3 and 4" [3H] proline (to-8 N). Labelled cells are
located throghout the course of the prìmarry (pp) and secondary
(Ps) meshworks. LabeT'led structures (B) can also be identified
at intersections of the tertiary plexus (Pt). Scale bar represents
50 um.
2-QFIG. 5 (a) ["H] leucine (10 " M). Dense accumulat'ions of silver
grains car¡ be seen over both the soma and emergent
axons of extragangìionic neurones (N) located within
fasciculi of the primary plexus (Pp).
(b) Light n:'icrograph of guìnea-pig Íleaì myenteric plexus
stained by the modif ied NADPH-'Jelrydr"oganase method.
A neurone (N) wjth long dendrites and singìe axon
characteristic of a Dogieì Type II neurones can be
seen located to a gangìion (G).
Scale bar represents 50 pm.
FIG. 6. t3Hl leucine (f'O-8 f,l). The secondary (Ps) and tertiary
(Pt) meshwor'ks can be seen with a diffuse cover of silver graìns.
Dense accumulations of silver grains (B) are present over inten-
sections of the tertiary pìexus (Pt). Scale bar represents 50 um.
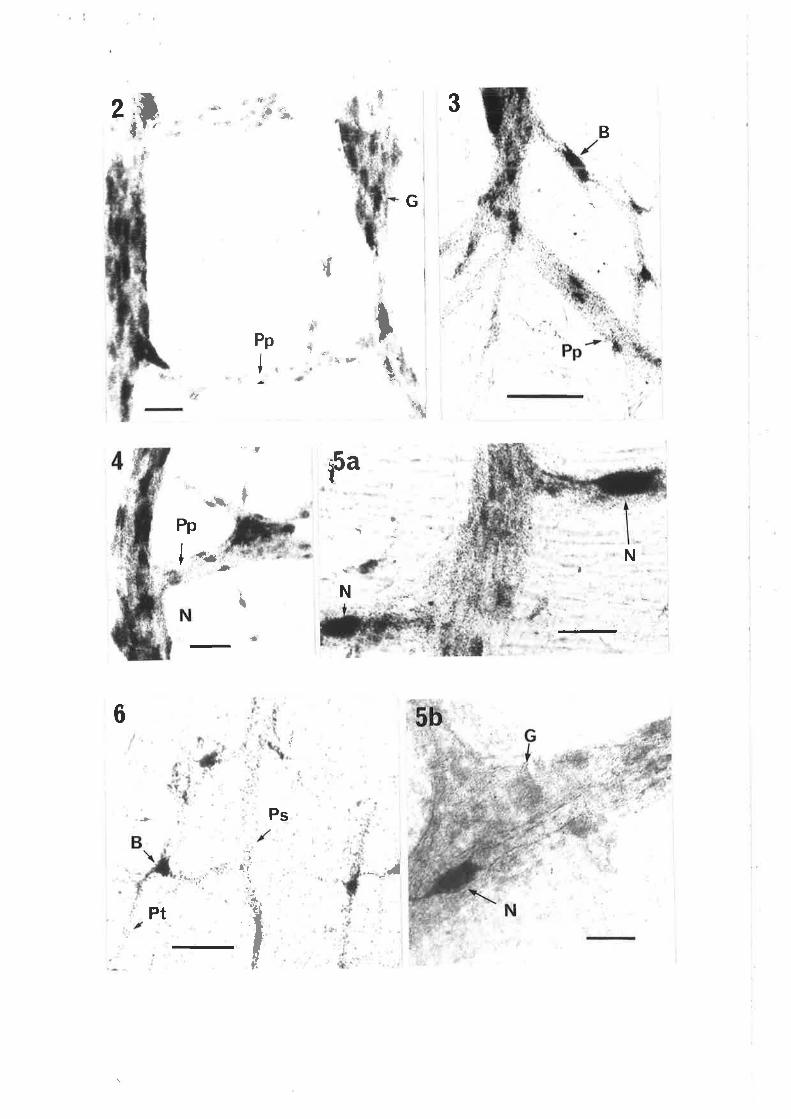
3I
rttI
ì
ì'it2B.t
G
{
lt
J
*.t
\lt
Pp
I
\
¡\' l
þtÐ,
¡\ ';,i '
-f
¡5u
N
tPp
a
\ N
-
J
6
{-
, Ps
--¿)
ri1'
-a,$¿¡
lr.
-
Pt

FIG. 7. Light microscopic autoradiograph of guinea-pig
ileal myenterÍc plexus treated with 5 x 10-9 t't [3H]-enBA in, -?the presence of 10 " M ß-alanine. The three meshworks,
primary plexus (pp), secondary plexus (ps) and tertiaryplexus (Pt) are readily identified. Dense accumulations of
silver grains (B) are c'learly visible within one ganglia (G)
and along the couì"se of the secondary and tertiary meshworks.
scale bar represent-s 50 um. counter stained with neutra'r red.
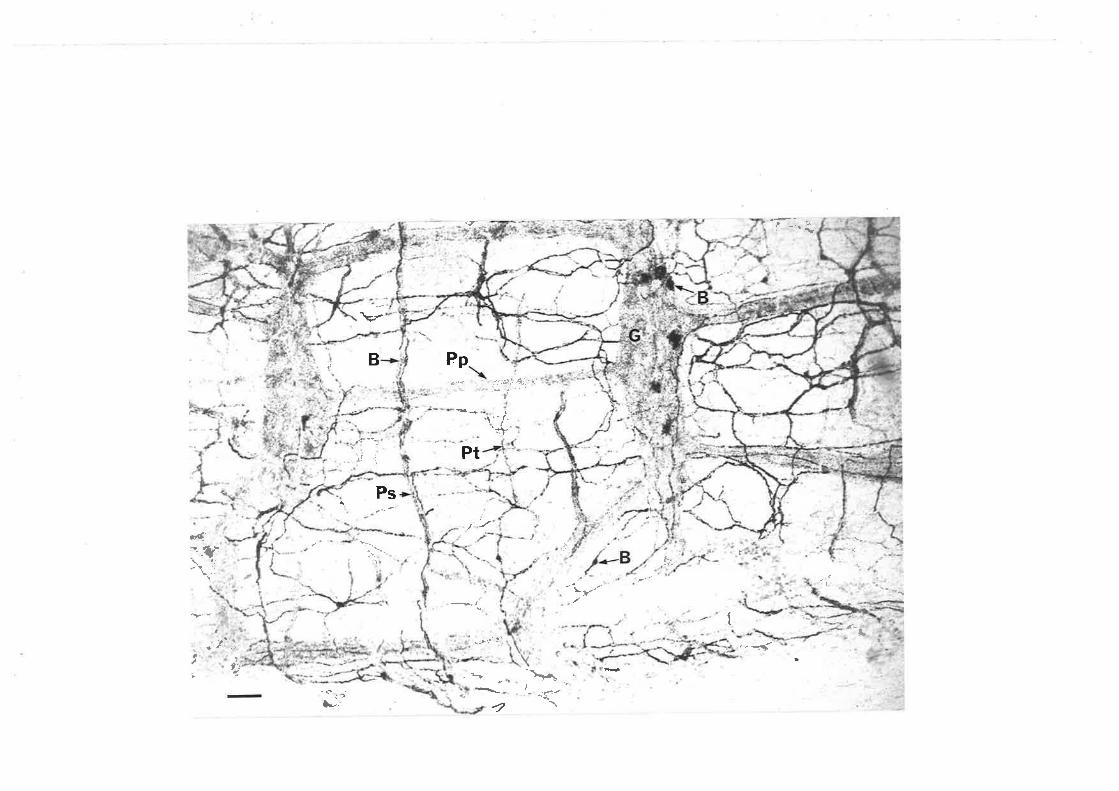
f v :c
I
\.'-
B Pp
r:-.i¡.-qr:t'f
\"/¡'rÌ'
)t. Pt
Ps
t.4':,ff a
Þ'
I
I
t: t.¿ll f ,
1
i;: -\.\'ì1:.. 4

FIG. 8. Light microscopic autoradiograph of a
myenteric p'lexus from the guinea-pig ieiunum treated
with 5 x 10-9 u [3H]-enBA in the presence of to-3 I't
ß-alanine. The distribution of [3H.]-enen uptake sites
within the ganglia (G), and meshworks (prìmary Pp,
secondary Ps, tertíary Pt) are readily'identified.
Counter stained with neutral red. Scale bar represents
50 ¡rm.

ï*t:
;úi '
fr ;,,t- -l
x'- ---4. B
I
trî.\r}
:¿'ssl¡'
-' ----,!iù..'-
{* ..
-v -/'
..{- ,t'.¿ ^ --./
Ps B;,\,-)
I
I
' t,
-''t't'. -\i
\\+
1"'j -,
t'':- -
t
+
Pt ,a - -. .. . r-¡-l\
tt\!

45
Myentenie LocaLization of tSul-c¿,n¿, ard.3n-B-aLanineinLaminaz, preparations
Autogradiographs of laminar preparation of longitudinal muscle with
attached myenteric plexus from ileum or jejunum, were siml'lar whether the
tíssues were incubated witrr [3H]-GABA 5 x 10-9 M alone or in the presence
of ß-alan'ine tO-3 tt (an inhib'itor of gl ial GABA uptake). The chara.cteristic
plexiform meshworks of the myenteric p'lexus were reveaìed by the silver
grains due to t.he 3H-GABA labelling. In most part, there was a diffuse
cover of silver grains over the gangìia and fasciculi of the primary
plexus (Pp), giving a lightly shaded appearance, whilst the s¡naller fjbre
bundles of the secondary (Ps) and tertiary (ft) meshworks were, in places,
outlined by more,dense accumulations of si'ìver grains resemb'ling dìstinct
"tracts" (Figs. 7,8). Some trajls of s'ilver grains were seen over the
ganglia and primary mesht^/ork, often as singìe fibre or fibre bundle. No
dense accumulations of siìver grains were observed'in regions where the
ìongitudinal muscle was devoid of myenteric plexus, neÍther v¡as there
labelling over adherent circular muscle fibres or of blood vessels.
In rnan¡r preparatìons very dense accunrulations of silver gra'ins were
found aggregated in patches over the gangfia, but were localized tc only
a few cells in each. In addition, similar accumulatjons were found
scattered along the course of the primary, secondaryand tertìary meshworks,
particularly at junctìons of the network. In some jnstances within the
gangìia, these very dense accumulatjons were recogniseably localised over
not onìy the so¡na but also dendrites and the emer"gent axon of individual
neurones (Figs. 9, 10, 11). The types of gang'ljonic neurones labelled
with 3H-GABA included unipoìar cells wjth an oval or'tear-drop'shaped
soma, or were nlultipolar, particularly those with a basal dendrjtic cluster
and offset nucleus (Fig. 12).

FIGS, 9, 10, 11 and 12.
Dense accumulations of silver grains can be seen over both
the soma and enrergent processes of neurones (N) located w.ithjn
four different ganglia of the guinea-pig ilea] myenteric plexus?_o
treated fcr ["H]-GABA (5 x 10-' M) autoradiography. ß-alanine-?10 " M was present in the incubation medium. Indivjdual
preparations with the exceptìon of Figure 12, were counter
stained with neutna'l red. Scale bar represents 50 um.
FIG. 13. Accunulations of silver graìns assoc'iated with
[3H]-engn uptake in the guinea-pig i'ìeal myenteric p'lexus.
Heavily labellecl tracts and bodies (B) are present in the
secondary (Ps) and tertiary (et) meshr,¡orks. [3H]-GABA finalconcentration 5 .x 10-9 M was userl. g-aian.ine 10-3 M was
present in the incubation medium. Tissue counter stained
with neutral red. Scale bar represents 50 ¡rm.
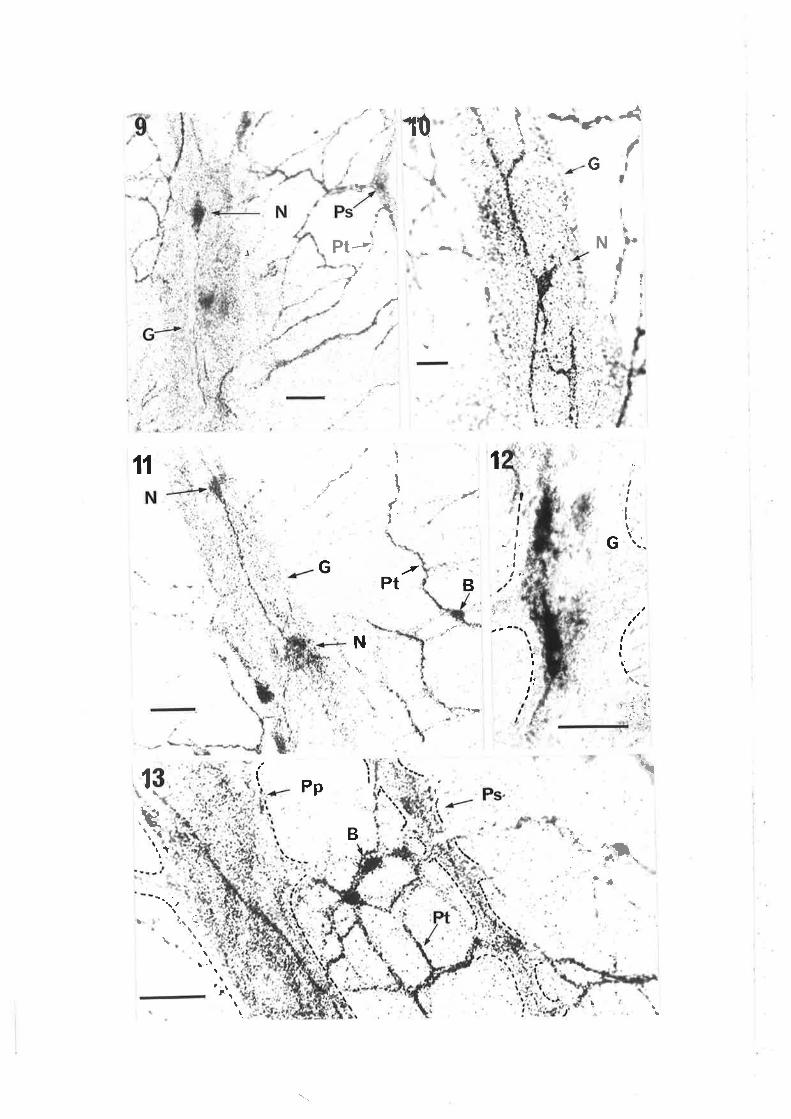
4/I
Pt'<' -'\--\/
ì^3,t1
G
ìv't\
G
1- ,;a,P¡
i.,t
.\,J\fl
\ :-ètfI
a'a
I
¡?
:t I
I
I
NfoI
¿,'Iìi
,t;'\ t
/a.;11
1 -.
tt,
'tI.t
/,
I
II)
11
f,
,
I- t.
I,,
BPtG
.,
ItIIT'
t,:. !.
NfI,tl,'
\I
tI
Pp1\
B,Et.'(
¡ìt
I
:'
I\t.I
lt

46
No specific cell types labelled by 3H-engR could be d'istinguished
within the secondary and tertiary meshworks, aìthough intense labe'llìng
of structures (mean ìength 25 pm, w'idth 15 irm) occurred over intersectjons
of the tertiary meshwork (Fig. 13) these were evidently part of the
simi 1ar, 'larger popul ati on descri bed above as bei ng ì abe'l I ed wi tn 3H-
?leucine or "H-proline (Fig. 6). In autoradiographs of ileal laminar
preparations, the distribution of the labelf ing witn [3U] ß-alanine
lacked any of the features of [3H]-enen uptake (Fìs. 14). very few silvergraìns were present, the cresyl violet counter stain predominating. The
silver grains v¡ere not obviously accumulated over any cel'l bodies, glial
or neural, nor lvas there any ìabellìng over tracts. Where silver grains
were present, they formed a light, diffuse cover over the gangìia and the
primary and secondary meshworks but not the tertiarry meshworks" This was
in sharp contrast to the accumulations of silver grains seen over neurones
of the primar,v meshwork and distinct tracts withir¡ all three meshworks in
[3H]-enan treaterl tjssue, indicating tfiat [3H] B-alanine was not taken-up
by neuronaì elements of the plexus.
ÐEffects of L*DABA, nipecotic acid and ACHC on I i(l-G{IBA uptake
In tissues treated w;th [3¡t]-GABA in the presence of [--DABA 10-3 M,
nipecotìc acjd 1c-3 f''!, or ACHC 10-3 M, there was a'rnrost a complete lack
of dense accurnulatjons of silver grains rlver t.he p'lexlts, either in gìia or
t.he meshrvork, jndica*"ing that neurones were no longei- labelled (Ëig. 15),
due to inhibiùion cf 3n-gnBn uptake. At t.he concentr ations used, nipecotic
acjd appeared to be more effective than L-LIABA or" ACHC as an inhibitor of
GABA uptake into neurones of the myenteric plexus, but quant'itative anaìys'is
of the effects of uptake inhib'ition was not attempted. Such siìver grains
as v{ere present itt preparations treated'wjth uptake inhibjtor; formed a

FIG. 14. Light microscopic autoradìograph of gu'inea-pig
ileal myenteric plexus after incubation i'n [3H] g-ulanine
(S x tO.'9 N). Silver grains are almost totally absent.
Only the ganglia (G) are readily delíneated, due in most
part to counter staining the tissue wìth cresyi violet.
Scale bar represents 50 um.
FIG. 15. Autoradiograph of a guinea-pig ììeal myenteric
plexus treated wittr [3H]-GABA 5 x 10-9 in the presence of
L-DABA to-3 tt. The gangl'ia (G) of the primary meshwork (Pp)
are readi'ly identifiable, counter stained with cresyl violet.
Onty a very diffuse cover of silver grains 'llas evident over
these structures. Scale bar represents 50 um.
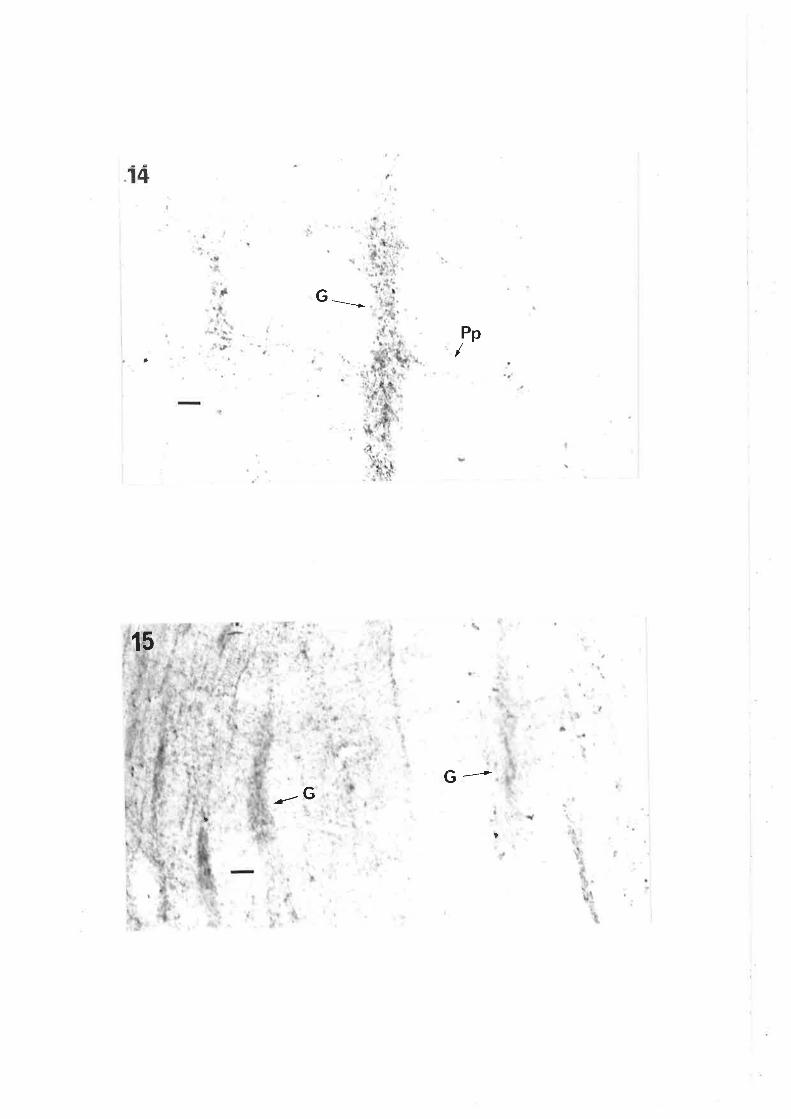
4ï
G
/Pp
G --_*
:J
15
^r- G
i
,

Light mìcroscopic autoradiographs of guinea-pig intestine
treated wittr [3tl]-cABA (s x to-9 tvl) in the presence of ß-
alanine 10-3 M. Transverse sections (20 prn) were prepared
in paraffin and counter stained with cresyl violet.
FIG. 16. Ileum.
FIG. 17. Distai colon.
The layers of the intestine wa11, serosa (s),longitud"inaì
muscle (m), myenteric plexus (mp), circular muscle (cm),
submucosa (sm), lunren (L) are identjfied by the cellular
stain. Siìver grains associated witfr [3H] uptake are
present over the myenteric plexus and circular muscle on'ly.
Ganglionic neurones (N) can be identified some of which
sfrow [3tl]-GABA labelìing. Scale bar represents 50 um.
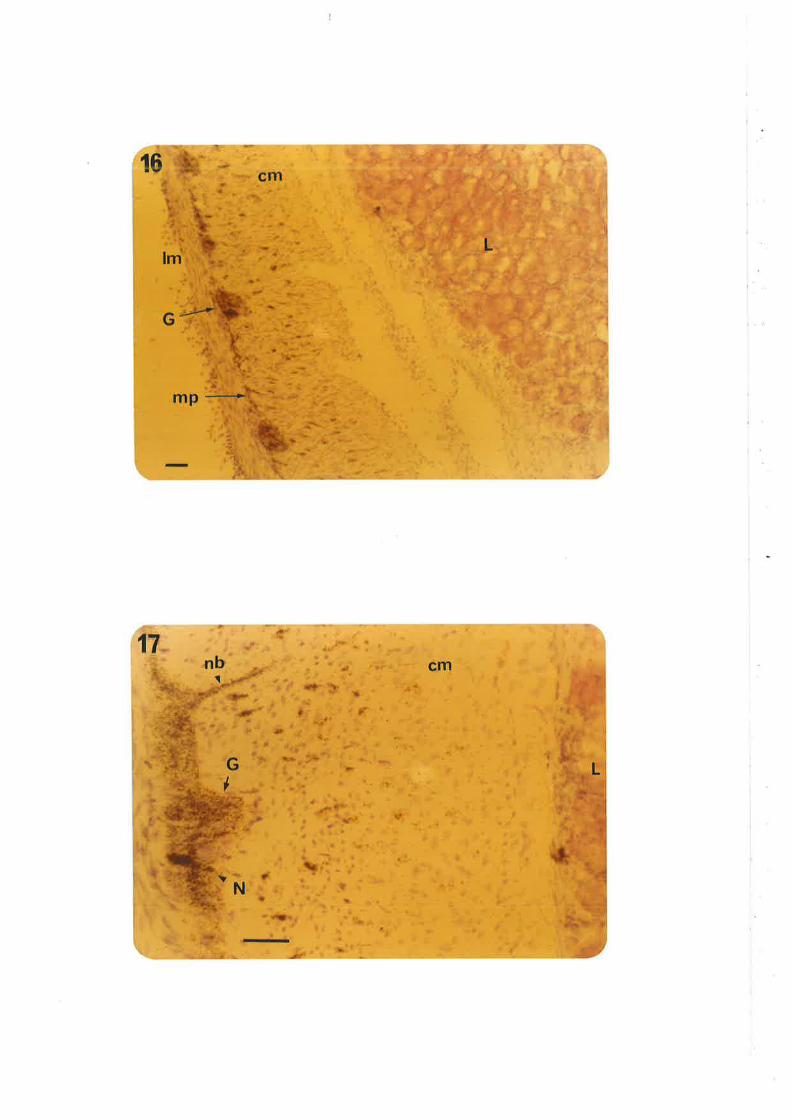

47
djffuse cover that was more obvious over the gangìia and primary meshworks
and could be distingu'ished from the cresyl violet counterstain.
Incubation of the tissue witfi [3tl] B=alan'ine'i'n the presence of L-DABA
to-3 M, which was observed to marked'ly reduce the uptake or [3H]-GABA., did
not alter the d'istrjbution of [3H] g-alanìne'in the plexuse ìn keeping lv'ith
the apparent specificity of L-DABA for neuronal hÍgh affinìty uptake sites
and an exclusive glial uptake tor [3H] ß-alanine.
Autoy.adiographie Local.ízation of 7n-c¿.nL in sections of coLon and iLewn
In light microscopìc autoradiographs of trattsverse paraffin sections
from the guìnea-p'ig ì'leum and distal colon, labelled wittr 3H-eRgA, there
was an extensjve d'istribution of label within the ganglia, and, in addition,
a widespread dìstribution of 3U-eRgR uptake sìtes in the meshworks of the
pìexus, and particularly in the circular muscle layer. By contrast,
accurnulat'ions of silver grains associated with 3H-GRAA uptake were almost
completeìy absent from the longitudinal muscle, submucosa, muscularis
mucosae or vjlli as seen in counterstained preparations (Figs. 16, L7).
A diffuse cover of siìver gra'ins occurreci over the ganglia, together
with dense accumulatjons of siìver graìns clear'ly overintraganglionic
cells includ'ing theìr dendrites (figs. 18, 19, 20). In many jnstances
reg'ions devoi d of si'l ver graì ns were outl i neC and resemb j ed unl abel I ed
cells near those labelled in the same sect'ion (Fig. 2I).
A conspìcuous feature was the accumulation of s'i1ver grains over
structures, presumably fibre bundles, extendìng from the gangl'ia and the
interconnect'ing pr ìmary meshwork into the circular muscle 'layer

Autoradiographs sholing uptake of [3Uj-eABA (5 x 10-9
the presence of ß-alanine (tO-3 t'l).
M) in
FIG. 18. Transverse paraffin sections prepared from the
guinea-pig i1eum. Counter stained with cresy'l violet.
FIG. 19. Transverse paraffin sections prepared from the
guinea-pig distal colon. counter stained wiür cresyl violet"
Cresyì violet stained cells characteristic of the different muscle
and nerve layers and lumen can be seen. s'iivergrains are present
onìy over the myenteric p'lexus and circular nruscle. in the
gangìia (G) labelled neurones (N) can be identifìed and on
occasion their emergent processes observed. Scale bars represents
50 um.
FIG. 20. Transverse paraffi¡r sections prepareci from the
guinea-pig ileum. A neurone (N) intensely labellecl wiܡ [3H]-
GABA is v'isible in the gangl ion (G). Emergent pnrlcesses can
be easi]y d': st'!nguì shed. No counter stai n . Scal e bar represents
50 um.
FIG. 2L. Transverse paraffin sect'ion prepared from the guinea-
pig ileum. counter stained with cresyl vioret. In a gang'lion
unl abel I ed cel I s (u) can be seen ìyì ng near neurone (N ) i ntense'ly2
labelled wíth ["H]-GABA. Scale bar represents S0 um.
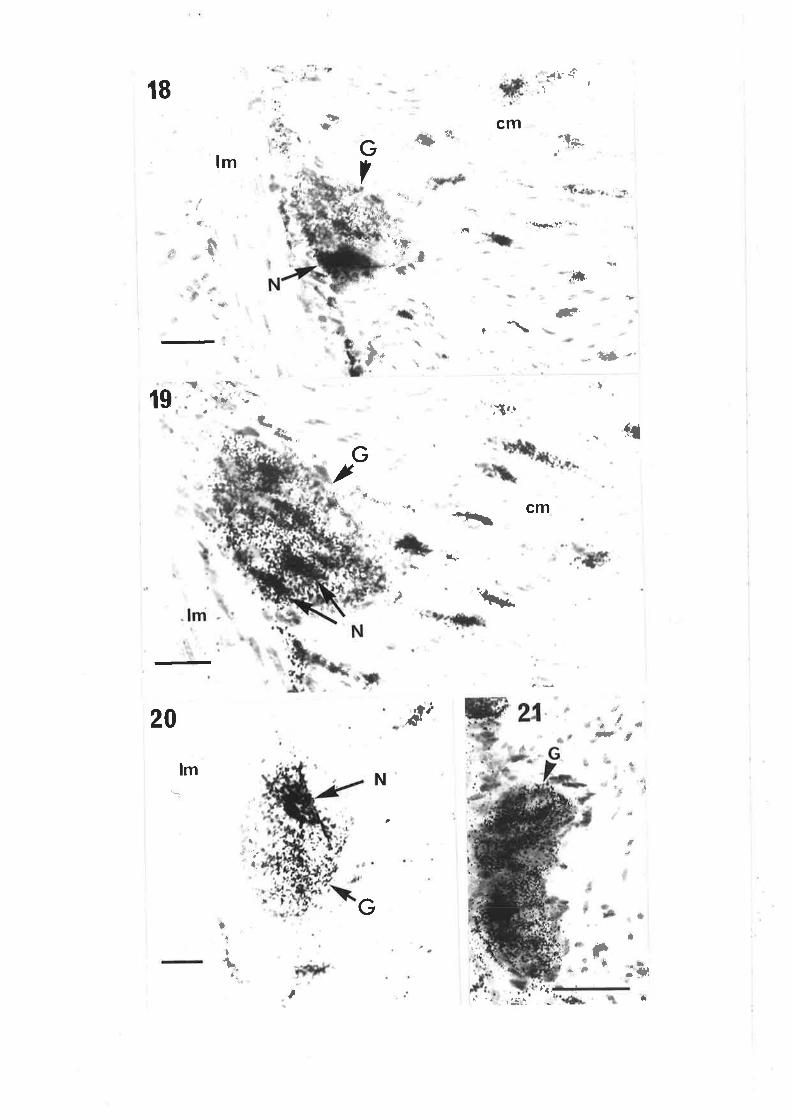
18
20
lmG7
cm**.'
'tri
4 r,; 't*þ
'tr'{
{
¿)r.*': _l-*\
$.¡t,
tç'
..ô-
I
G/
t\
,a,r
i,-t\
.-rat '.T
19(,
tr..
q
Ia#,
a
t
{¡|..
Icm
.)
- ,ö?¡
lm ¡
I
NI
I
l'¡.a
¡
a
lˡ
tIa.
GÒ
.? jþt

FIGS. 22 and 23.
Light microscopic autoradiographs of transverse paraffin sectjons
(20 um) prepared from the gu'inea-pig i1eum. Accumulations of
silver gra'ins occur over the ganglia (G) and associated nerve
bundles (nb). Dense clusters of si'lver gra'ins,also occur
over the ciruclar muscle (cm). The serosa (s) and longitudinal
muscle (lmi are free or ¡3H1-GABA labelling. Scale bars
respresent 50 um.
FIG. 24. Transverse paraffin sect'ions of the guinea-pig
distal colon treated tor [3H]-GABA autoradiography. Sections
were cut oblique'ly across the width of the colon wall. Silver
gÈains occur over distjnct tracts running paraììel to the
circular muscle fibres stretchìng across the jayer between
the gangiia (C). The tissue was counter stained with neutral
red. Scale [:ars represent 50 um.
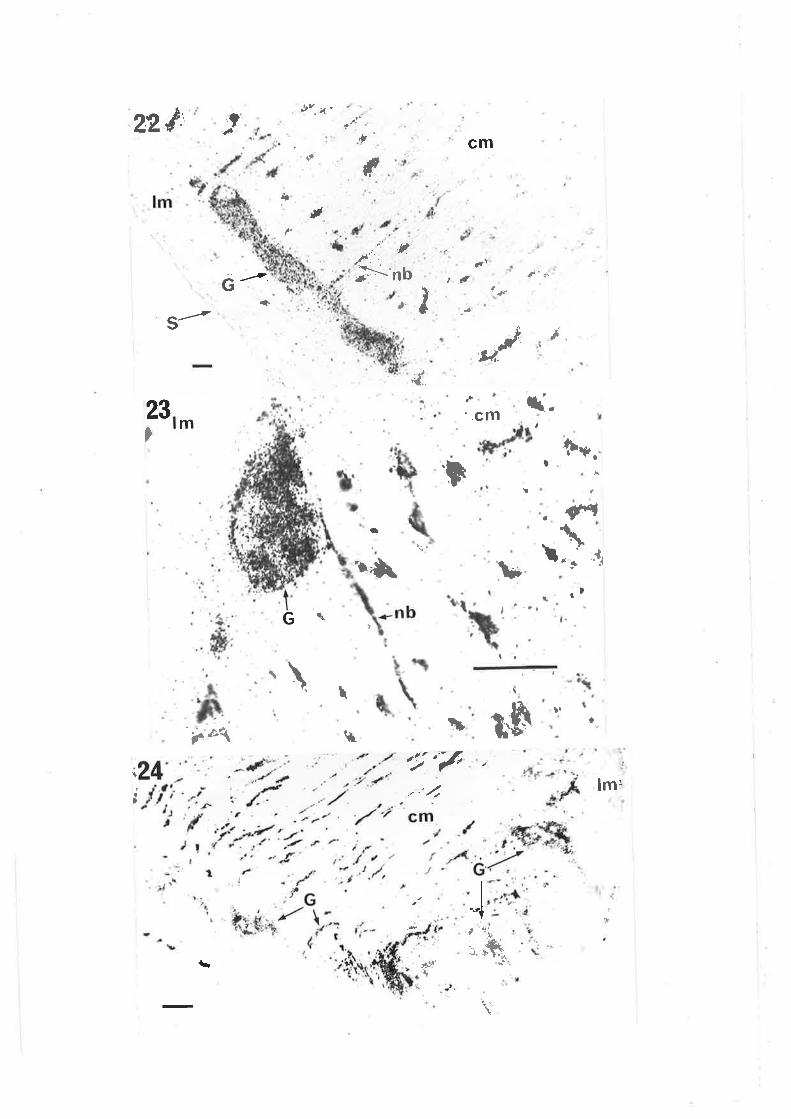
zzi: ,t+
s¿
23
\ ,* {".1
rt
a
þr, u-
{
It
cm
aat
,,
åÍ
--l ;:.---'nbG tJ
+ ,. ¡
'.cm(.
uì
-}¿\ fÞ
$
.,,t.
i
'tl.
m t
t '-t,
)
ìù tlrv
#\
t
a
G \
\ \,l
\ \
|¡t'iÊ ê1 hü"v..'¿
1
ta)
lm:
J,.J
tg-\\
,

48
(Figs.22,23). These trails of silver graìns were found only over the
outer half of the circular muscle ìayer, there being no label present
in the innermost part of this layer or the submucosal plexus. There were
also many punctate aggregations of silver grains scattered irreguìarly
within the same regìon (Fig. 24). In transverse sections these started
abrupt'ly for the most part some 50-100 pm from the myenterìc plexus ìayer,
apparently representing transversely cut labelled fíbre bundies. This
notion was reinforced by the observation of many trails of s'ilver grains
running paralìel to the circular muscle fibres in sections cut obìiquely
across the width of the colon wall (F'ig. 25).
ùLscussion
Since ß-emissions have little penelrating pouJer it, is perhaps surprisìnga
that so many ['i1]-jabelled structures could be detected in the rnyenteric
plexus by autorad'íography based cn dissected laminar preparatÌons dipped in
sensitive emulsion. Howeve¡, the basal lamina enveloping the gangìia is
sufficiently thin and possìbly may even be broken in places by the dissection,
so that the ernuls'ion can be affected by g-particles arjsÍng fnom'Eritiurn
labelled structures wit.hin gangiia and smaller d'ivision of the plexus.
l4oreover, unìike the situatíon in sympathetic and spinal gangìia, ùhe non-
neuronal elements of the plexus by no means comp'letely ensheath the
ntyenteric neurones, part'icul arly where neuronal processes ari se, so that
many nerve cel'l bodies, dendrites ancl axons are bare or in jmmecljate
contact with basaì larnina (schofieìd, 1968; Gabelìa, I97L, I972; Cook and
Burnstock, 1976a). Corrsequentìy the conditions for autoradiography ìn
dissected myenteric plexus lam'inar preparat'ions,must be more favourable
than might at fjrst. seem possible" Therefore it is most likely that

49
the dense accumulations of silver grains assoc'iated with [3H] amino acid
uptake'in the myenteric plexus represent labell ing of superfìc'ial neurones
and their processes ìn the ganglia and meshw'orks where, ìn many instances'
considerable deta'il of denclrites and other processes could be seen' This
js confirmed by the similar labellìng of neurones found in autoradiographs
from paraffjn sections of the intestjnal wall,where there can be no
quest.ion of I imitation in the penetrating power of the ß-emiss'ions'
In all the autoradjographs follow'ing'incubation with any of the
tritiated amino acicls,there was a diffuse cover of silver gra'ins over the
plexus, partìcu1arly the gang'lia and the primary and secondary plexuses'
but not the smooth muscle'itself. There was little d'ifference in the
appearance of the diffuse silver grain cover.in preparations labelled w'ith
3H-eRgR alone or in the presence of ß-alanjne, a specific 91ìaì cell upt'ake
blocker for GABA (Ivensen and Ke]1y, Ig75), which suggests that th'is
labellìng vras not assoc'iated w'ith gliaì cells, unless the substrate
spec.ificitjes of myenteric E1ìa'l cells differ from those in the central
nervous system. There is some controversy concerning glial elements of
the myenteric pìexLrs. Gabella (1969,1979) describes g'l'ia1 cells' but'
Cook and Burnstock (1976b) maintain that these should be called instead
Schwann cells, neservìng the term'gliaì cell'for the central nervous
system. Be that as i+, may, 1t should be pointed out that blood vessels
overlay, but do not penetrate the plexus (Gabella, L979; Jacobs ' 1977')
consequent'ly the variety of glial types would be restricted sjnce astrocytic
contacts on blood vessels vrould be missing' poss'ib1y wìth obl'igodendrogf ia
represented by schwann cells. Many of the cell bodjes and processes of
neurones in the gangìia are bare of'glia'and aìthough satellite nuclei
outnumber neurones several fold (Gabella, lgTg), there was no dense [3H]
ß-alan.ine labellìng wjth sììver graìns outlining gììa'l cell bodies as is
seen wjth [3H] ß-alan'ine radjoautographs in sensory gangìia and central

50
nervous system (Schon and Kelly, 1975; Hosli and Hosf i, 1980). Evidently
the diffuse cover of si'lver grains over the plexus seen'in this study,
represents amino acid uptake by some non-neuronal tissue of the plexus, or
may well be in part due to labelling of nerve terminals near, or receptors
on, neurones recejving a GABA-ergic input within the gangf ia.
Neurones with high affinity [3¡t]-GABA uptake mechanisms are to be
expected in the myenterìc plexus if there is, as has been suggested
(K.ranti s et aI,., 1980; Krantis and Kerr, 1981a) GABA-ergic control of
peristaltic actjvity. In the present study the use of selectjve 'inhibitors
of high affin'ity upt.ake of [3H]-engn jnto neural and g'ìial elements showed
that the high affinity uptake sites were maìnly within neuronal elements
of all three meshworks of the plexus.
Reciprocal connexions are made between the submucous pìexus and the
gangìia of Auerbach's plexus. These connexÍons penetrate across the width
of the circular muscle layer and give rise to a tertiary plexus innervatìng
that layer (Gabel'la, 1972; Jessen et aL., 1980). The stout branches ìeavinçt
Auerbach's plexus, between the longìtudinal and circular muscle ìayers,
were labelled by 3H-gRgR, but on'ly over the outer half to two-thirds of
the circular muscle layer. Silver tra'ils from labelled tertiary bundles
could be traced into the circular ìayer, and labelling'in the tertiary
plexus was also very evident 'in the laminar preparat'ions.
This dense labelfing of the tertiary plexus with [3H]-GABA was a most
unexpected finding in vieur cf the evìdence (Kranti s et aL., 1980) that GABA
acts upon the cholinergic motor neurones, and the non-cholinergìc non-
adrenergic inhib'itory neurones of the plexus, but not on the smooth muscle
of the intestine. From what is known of the functional organisation of
the plexus it might be expected'bhat GABA-ergic neurones would be confined

51
to ganglia or the primary plexus rather than inthesecondary or, more
particuìarìy, the tertiary plexus where very dense 3H-eRgR labelling was
found. Yet the latter labelf ing occurred over typìcaì GABA-uptake sites,
in that it was prevented by both ACHC, L-DABA and nipecotic acid, all of
which are effecr,'ive inhibitors of neuronal uptake of GABA (Schon and
Kelìy, 1975; Johnston, 1976). However, GABA receptors have also been
described on mammal'ian perÍphera'l axons (Brown and Marsh, L975) as have
receptors of the'Baclofen' type (Bowery et aL., 1981), and the heavy
label'ling of the tertiary plexus would be explained if high affinìty GABA
uptake sites were associated with either of these axonal receptor-types.
Furthermore, if, as shown by Bowery ei aL. (1979) and by Ong (1981) fronr
this laboratory, activ'ity of the myenteric plexus is subiect to tnodulation
(presumably prejunctional) by the novel GABAB (Baclofen) type of GABA
receptor, then a GABA-ergic ìnnervation responsible for such modulation
might be expected to accompany the motor fibres of the tertiary pìexus
and would possess uptake sites, possìbìy some of those here seen as heavy
3H-gRgR label'ling of the tertiary pìexus, partìcularly where found
coursing through the ci rcul ar muscl e I ayer in paraì'le'l wi th the muscl e
fibres, characteristic of nervous ramifications within the circular muscie.
Enkephalins also inh'ibit electricaì1y evoked contractions of the smooth
muscle in the guinea-pìg iìeum (Van Nueten et øL., L977; l,Jaterfjeld et aL.,
t977), by presynapt'ic actions at nerve terminals of motor nerves releasing
acetylcholìne (North et aL.,1979; Bowery et aL.,1981;0n9, 1981. The
distribution of enkephalin immunoreactive fibres wjthin the circular
muscle as seen in sections of the guinea-pig ileum (Schultzberg et aL.,
1980) bore a striking resemblance to the autoradiographs in this stLdy
a'lthough the presence of 3H-eRgR labelled f ibres to f ibre bunclles
innervat'ing in most part onìy the outer half of the muscle is unexp'lained.
A further possib'ility is that the labelled fibres may be in part GABA-ergic
afferent fibres. If so, these represent sensory endings or hitherto

trtJL
undescribed nerve cell soma located to the outer half of the circular
muscle'ìayer, where the label was found, and not sensory fìbres frorn the
submucous pl exus or" the p'lexus muscul ai re prof ond of Ca jal (1911) (deep
muscular pìexus, Schofield, 1968) previous'ly suggested by Gabel'la (I974)
to be a source of afferent input to Auerbach's plexus, since 3H-eRgA i,absent from these 'layers
Although neuronal processes, presumably axons, were labelled in
gangìia and the meshworks of the plexus by 3H-eRgR, surprisingly few
fibres labelled witfr 3H proline or 3H leucine. No doubt the 20 minute
incubation tirne with these partìcul ar tritiated am'ino acids was too
short for any significant slow transport to occur. Slow transport of
incorporated amino u.id proceeds at about 200 um/hr (Cowan et aL." Ig72)
so that anterograde transport for only some 70 um would be seen after 20
minutes incubation. Ne'ither of these amino acids effect'iveìy label by
retrograde transport, nor do they sÍgnificantly enter fibres of passage
(Cowan et aL.,1972). Thus, at most, one would expect to see cell bodies
and short, labelled processes as found here in the myenteric pìexus.
Uptake of 3H proì"ine and 3H l.u.ine into cell bodjes is seen in the
central nervous system within 5 minutes of application of the labeì, and
within 30 minutes cell t¡otlies and a few ¿endrites are strongly labelled
(Droz and Leblond, i963). The kinetics of the uptake of these amino acids
into neurones of the plexus was'not investjgated, but a varjety of cells
were visibie after only 20 minutes incubation followed by ìmnrediate fixation
and subsequent treatment for autora.d'iography. l4ost of the labelled ¿ell
bodies were in the gangìÍa, but many dense accumulations were also found
not only within the primar"y and secondary fasciculi of the nreshurork, but
also at intersections of the iertiary plexus. Although the 'inrerstitai
cells of Cajal are noþr thoLrght 'ùo be non-neuronal (Gabel1a, 1972) the

53
autoradiographs of laminar preparations contained many examples of dense
accumulatìons of silver graÍns at Íntersect'ions of the tertiary plexus.
The resulting pìctures bear a striking resemblance to the plexus and
interstjtial cells illustrated in Cajal figures 572, 573. More of these
structures were labelled witfr [3H] profine and 3H l.r.ine than with 3H-
GABA, but a significant number did label with.the latter; thejr disposìtion
and trjangular shapes, illustrated in our figures 7,8,13, may be
compared with the interstjtial cells in the Caial figures. No 'labe'lling
of these structures was seen with 3H-eRgA if neuronal uptake blockers of
GABA were present,which strongly suggests that they represent labelled
neurones.
Cell bodies have already been not'iced in the primary and secondary
p'lexuses usìng the N'itro BT sta'in (Gabel la, 1969, I979). 0f particular
interest were the intensely labelled structures w'ith'in the primary fascjcuji
of ileal segments incubated with the tritiated amino ac'ids. E'longate
extra-ganglionic neurones have been reported in the primary fasciculi
(takeo and Sugai , L974), hor'lever, cells that were morphologica'lly sim'ilar
to those hitherto described only in ganglia were also observed within the
primary fasciculi (compare FiEs. 4 and 5 w'ith F'ig. 29 of Schofield, 1968).
It is concluded that ail three divis'ions of the meshwork within the p'lexus
contain cells that are very ììkely neurones since they no longer labelled
with 3U-GABA after treatment rvith inhibitors cf neuronal high-affin'ity
GABA uptake.
There is a lack of agreement concerning the classification of the
variety of celì types found in the ga.nglia of Auerbach's plexus" Cajal
(1911) whilst descrìbing various d'ifferent cells nevertheless recognised
onìy two fundanrental types, those with short and those with long processes.
Gunn (1968) reverted to Dog'iel's classification with three subdivisions,

54
whilst, more recently, Schofield (1968) has recognised onìy two types:
Type i, multipolar with short dendritic arborizatjons, and Type II,generaì1y multipoìar with elongate processes (but often lacking dendrìtes).
Exampìes of all these could be seeh in laminar preþarations stained by the
Nitro-BT method and in the autoradiographs using [3H] proline an¿ [3H]
leucine, e.9., compare Fig.4 and Schofield (1'968) Fig. 29. By contrast
for the main part only two morpho'logica'l varjents were found to label 3H-
GABA. One of these was unipolar with the axon clearly labelled, as in
Figures 9, 10 and 11 (compare Cajal Fìgure 569), the other, more common,
had an offset nucleus in an eìongate ce'l'l body with a brushl'ike basal
dendritic arborization, as in Figure 12, c.f. Cajal (1911) (Figure 569).
Evidentìy 3H-eRgR labelled only a fraction of the total cell popuìation
capab'le of being sholn by autor adiography was wit¡r 3tl luecinu or 3H
proline. The significance of thesu 3H-eReA labellecl celì types is not
clear, however, there is great difficulty ìn correlating the structure
and function of neurons on the basis of morpholog'ical considerations a1ong,
as cðn be seen in the claims of motor or association functions for Dogiel
Type I cells, and sensory (Schofield, 1968), or cholinergic nrotor, for
Type iI cells (Hill , L927; Arnbache and Freeman, 1968).
Since more than one type of nerve cell possessed a GABA-uptake mechanism,
it is possible that GABA subserves serveral functions in the myenteric plexus.
In particular, GABA-ergìc neurones could be involved in nerve pathways
mediating reflex peristalsis since GABA has pharmacologìca'l actions at the
inhjbitory and excitatory motor nerves underlying this activity. Such a
possibif ity is supported by the recent demonstrat'ion that GABA antagonism,
markedly alters perìstaìsis in the guinea-pig co'lon as measured by faecal
pe]1et movements (0n9, 1981). Together with biochenlical and autoradio-
graphic evjdence (Jessen et aL., 1979; Hosoya et aL., 1981) for the
localization of GABA and its s.ynthesising enzyile (GAD) to mammalian myenteric

55
neurones,this evidence suggests a neurotransmi+.ter role for GABA in the
myenteric p'lexus.

CHAPTER IV
IVIORPHOLOGY OF MYENTERI C NEURONES

56
Introduction
Enteric neurones dìsp'lay varying suscept'ibil'ity to a number of
staining reactions including choì'inesterase, silver impregnation, methyìene
blue and Nissl stains, alì of which have beett util'ized in the histochemical
and histoìogicaì jnvestjgations of the morphology of the mammalian enteric
nervous system (see Schofield, 1968). The Nissl met.hods do not stain cell
processes (particuìar1y axons), and a'lthough silver.and methylene blue
methods outline cell bodies and their processes, they are relativeìy complex
and capricious in their use. Recently, the histochemical demonstration of
a major group of oxidation-reduction enzymes: the pyridine-linked
dehydrogenases wh,ich require either nicotìnam'ide adenine nucjeotÍde (NAD)
(DPN) or its phosphorylated derivative NADPH (TPN) as co-enzymes, has
been used for the investigation of the morphology of a number of animal
tissues (see Pearse, 1960).
The techn'ique (Pearse,1960) ìs based on the direct transfer of
electrons and H+ from the co-enzyme substrate to the dehyrJrogenase for
transfer to the electron acceptor (tetrazo'lium salt) reduc'ing it to an
intensely coloured insoluble formazan deposit, thus staining the cel1 blue.
More recently this technique has been appiied to the morphof igicaì
investigatìon of the myenteric p'lexus of rat caecum (Dupont et aL., 1965),
rat irrtestine (Gabella, L967), as v¡ell as guinea-pjg and human rectum
(Gabella, 1969). In these studies, the reduced form of the co-enzyme
(NADH) and a higher potentiaì tetrazolium salt (nitro BT, Pearse, 1960)
were used, resu'lt'ing in a more intensely bìue deposit of ciiformazan. The
authors describe the intra mitochondrial depos"ition of diformazan as on'ly an
indicator of the localìzation of the appropriate diaphorase (Pearse, 1960),
however ih" t..r 'd'iaphorase' may be artifactual s'ince this enzyme has been

57
identified to be a 'fipyl dehydrogenase' (White et aL., 1968).
The abi'lity of this'Lechn'ique to distinguish nerve cells from non-
neuronaj elements ìs due to the very intense pyridine-linked enzyme
activity wìthin nerve cel1s, which results in their staining'in a shorter
time than is required for non-neuronal ceils.
In the present study, a rapid, conven'ient modÍfication of Gabella's
technique has been used, with nicot'inamide adenine dinucleotide phosphate
(reduced fornr) NADPH as the co-enzyme substrate, This allows better
observation of not only the scma but also of emergent processes within
stretch nrounts of myenteric plexus of the guinea-pig i1eum.
Method
Segments of freshly excised guinea-pig proximal ileurn 10 cm 'long
were placed into Krebs solution (36"C), gassed w'ith a m'ixture of 95% 0,
and 5% C}r. blhjle still bathed'in Krebs solutjon, a g'lass rod 6 mm in
diameter was positioned through the lumen of indivjdual ileal segments
and the longitudinal muscle layer wìth attached nryenteric p'!exus was
careful'ly stripped alvay in one continuous sheet. Portjons of this'whole-
mount preparation' (stretch preparation, Gabella, i979) were treated by a
modifjcatjon of the NADII-diaphorase (dehydrogenase) h'istochemical method
of Gabella (1969).
The whole mount preparation of longitudinal muscle strip with
interposed myenteric plexus was stretched out onto a slide with the plexus
uppermost. The slide was then placed onto di"y ice and allowed to f,reeze.

58
over. Thjs was usually comp'lete wjthin 1 min. The length of the freezing
period appeared to be critical, with best results always obtained from
preparat'ions allowed onìy the minimum time to freeze over. The frozen
tissue was then carefulìy detached from the slide and placed into a vial
containing the reaction mixture, and protected from the light by wrapping
in alum'inium foil. The reaction mixture was prepared by mix'ing equa'l
volumes of Nitro BT 91 mg/m1) and Nicotinam'ide adenine dinuc'leotide
phosphate (reduced form) NADPH (1.4 mglml) dissolved in 0.9% normal
saline. These solutions were fresh'ly prepared in separate light proof
containers, and mixed together only immedìately prìor to the addition of
the frozen tissue.
Tissues were allowed t-o incubate in the staining solution wìth gentle
agitation at room temperature, for periods of between 10-20 m'in. The
degree of staining could be determined by vìsuaì observation. When the
tissue was judged sufficiently dark in colour, it was removed, carefu'lIy
washed, placed on a slide with the myenteric pìexus uppermost, and fixed
with 4% glutaraidehyde for 5 min. Fol'lowìng fixation the tissue was
washed, dehydrated and mounted for light m'icroscopìc observation.
A varjatjcn jn the time of fixation and/or concentration of fixative
did not appear to affect the localization of reaction product. Holever
there was an obvjous alteration in the intens'ity and clarity of the
staining reaction. Material fixed for a short time with lovl concentration
of fixative seemed to shol ìmproved definition and ìntensíty of sta'ining
over that f ixed for a longer time. Thì s 'tnaturation' period lasted
approx'imate'ly 10 days after which there was no further noticeable alteration
in the quality of the staining.

59
ResuLts
The interconnected ganglia of the primary pìexus (lp) were cìearly
visible in these whole-mount preparations of the myenteric plexus stained
with the modified NADPH - dehydrogenase histochemica1 technique (Fig. 1).
Many nerve cells could be seen within the ganglia, and the soma of these
celJs showed varying degrees of sta'ining from very intense to lighilystained. Large opaque nuclei were often visible. In those neurones
intensely stained by this histcchemical technique, cìear definition of
their soma and processes (which could on occasion be traced some distance
fr^om their origin) uias usually obtained, a'lthough it was not often possib'le
to distinguish between axon and dendrìtes, a lÍnlitation common to many
histologicaì procedures. But in many 'instances it was poss'ible to identify
several monphoìogica'lly different types of neurones described in a number
of studies as Type I, II and III neurones of Dogiel (1895,1899) and
variations of these types (Gunn, 1959, 1968; Gabe'lla, t97g); Fig. Z.
No obvious pattern of localization of these cells was apparerrt.
Dogiel described Type I cells as having numerous stout processes
sometimes arrattged in a cogwhee'l appearance. These cells usually possess
a single slender axon which leaves the soma and passes from the gangìion
into one of the fasciculi. Arr exarnp'le of a Type I cell stained with this
Nitro-BT method can be seen in Fig. 3. This neurone bore a strong
resemblance to a Type I celj stained by a modified Bie'lschowsky method
in Schofieìd (1968)
Multipolar cells with fewer processes bhan the Type i cells, but with
characteristic fine processes of Type II cells were often observed (Fig. 4).
l^lithin many gangìia varjous celì types could be observed in close

EXPLANATION OF FIGURES
All figures are light micrographs of the guinea-pig ileal
myenteric p'lexus stained by the modifíed NADPH-dehydrogenase
method. Scale bars represent 50 um.
FIGURE 1. 'The ganglia containing many nerve cells, and the
interconnecting fasiculi of the primary plexus (Pp) are clear'ly
visible. Nerve cells display variable stainìng'intensities.
Nerve fjbres can be seen coursing through the fasicu'li.
\
h$:"ã
--'¿)t':1.
t)
cto.
..'¿
1'On
,l
4:¿
-:\
..tï.Gr
\

FIGURE 2. ,Myenterìc neurones of varying types are easi'ly
identifiable within ganglia (G). Emergent processes can be
traced some dìstance from their origin.
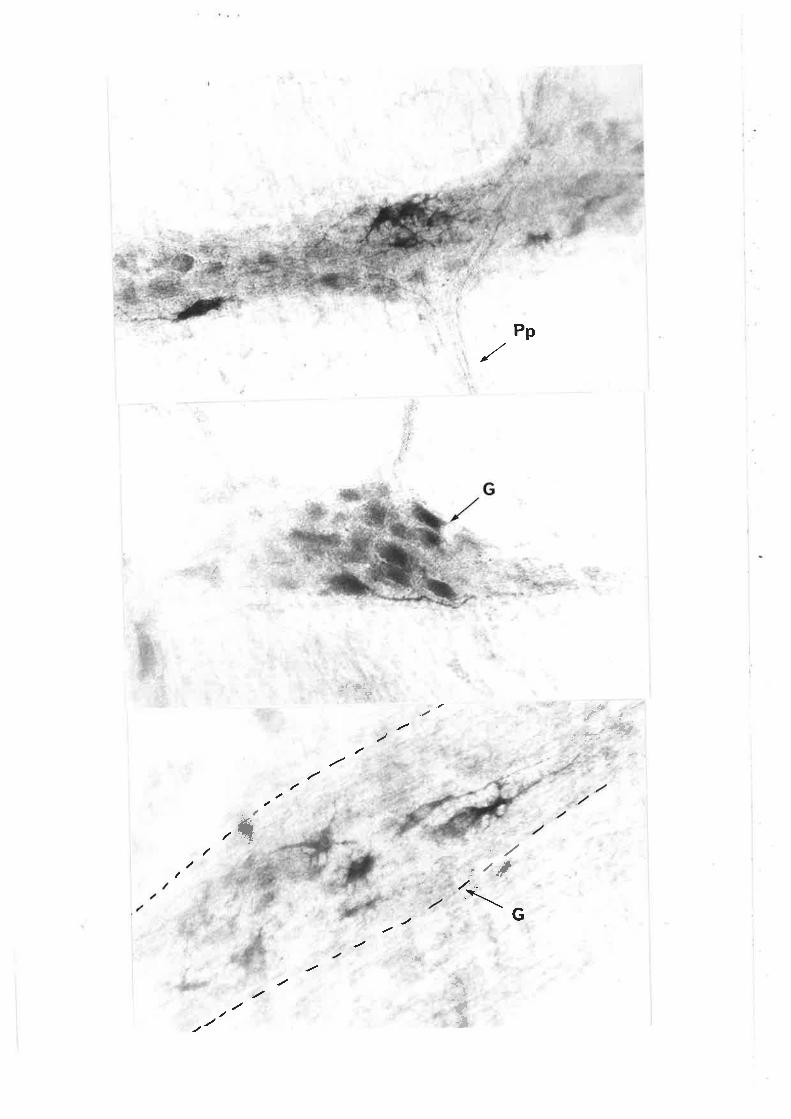
GË-
tdl
,.'' ]+ì1:;á, L ',...*/4' .'¿t
t,.
Ð\-
f z t
./
fþI
H¿"4'' i
ê{ _û'.:t r.'
I,1
d¿
Í

FIGURE 3. Dogìe1 type I nerve cells (N), with soma and
emergent processes intensely stained.
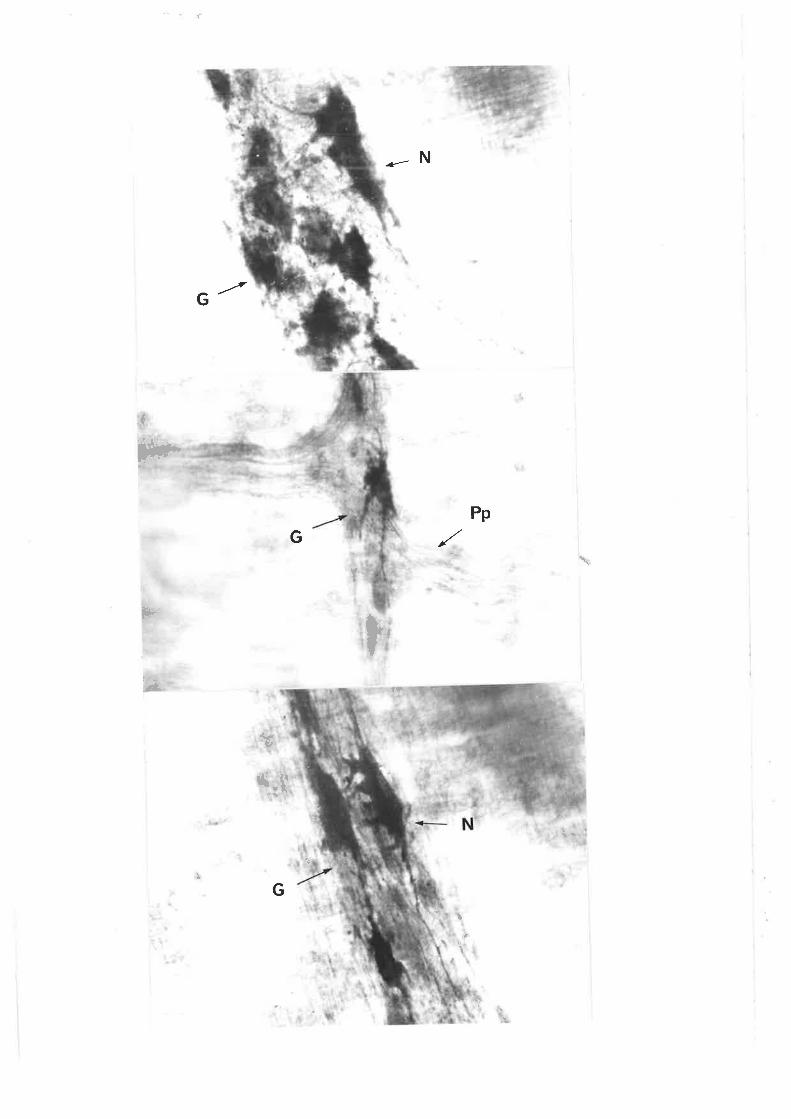
-,.,.-N
G -r'
{
st*
Fl-
Pp
,/G
f
l I
úL 'tG
E
t={
\
IG
I
\
.!-
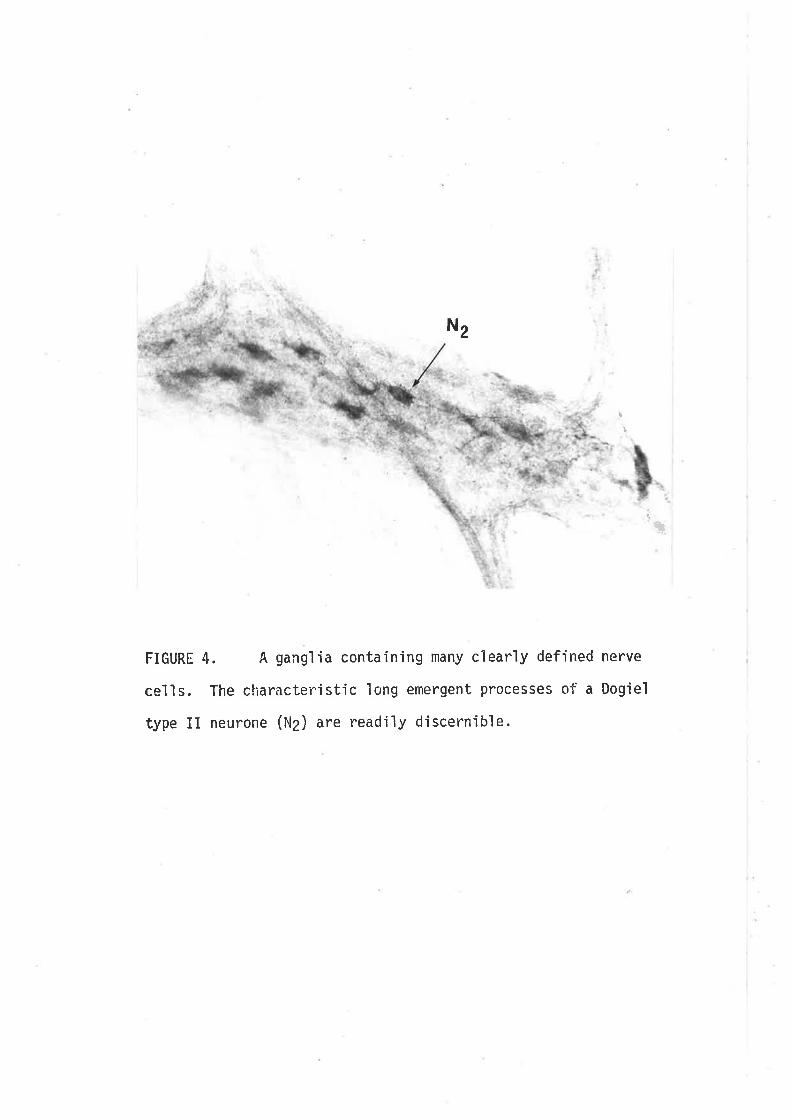
N 2
I
q
FIGURE 4. A ganglia containing many clearly defined nerve
ce1ls. The characteristic long emergent processes of a Dogie'l
type I I neurone (ltlZ ) are readi ly di scerni bl e .
It

FIGURE 5. Intensely sta'ined nerve fibres can be seen 'in
the fasiculi cours'ing between ganglia. In some instances
enpassant fibres within gang'lia can be identified.
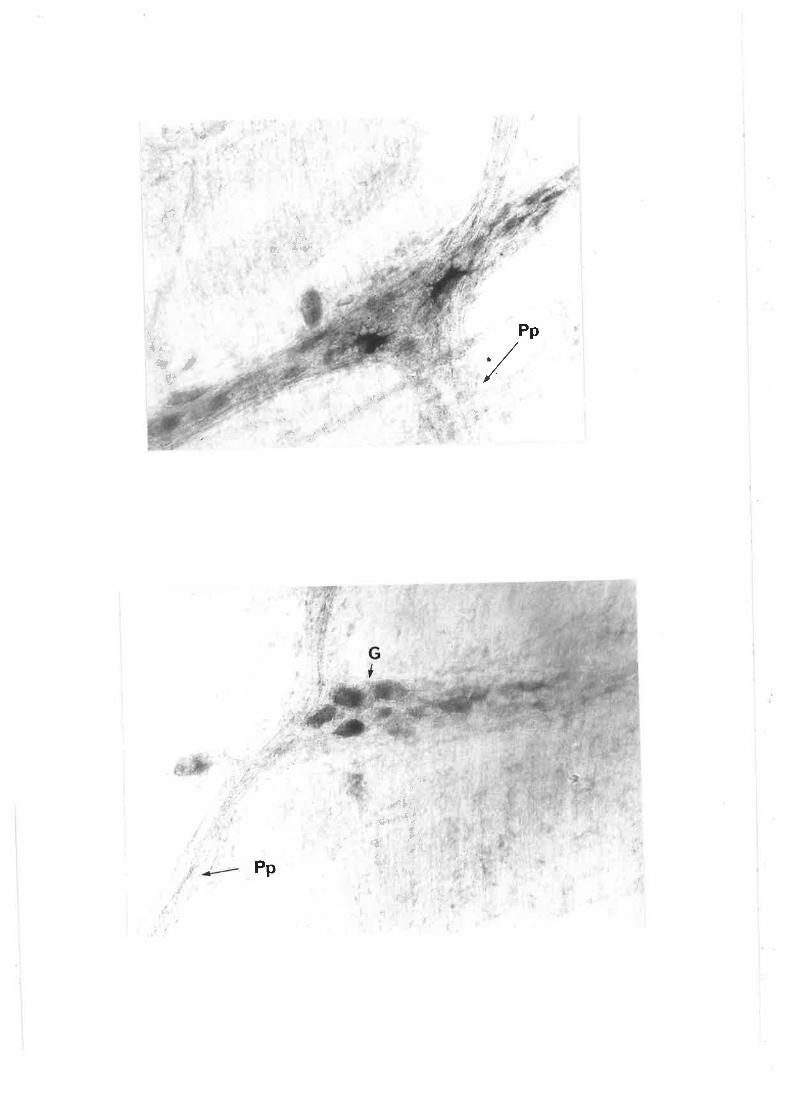
,
t'.,
aç¡I
3:ç
f
Pp t
.i ,,.
?F /
,t.l'¡î
ß
Ft.!
,;i
âfrl;,4'.
,$;'i
Pp+'
,l
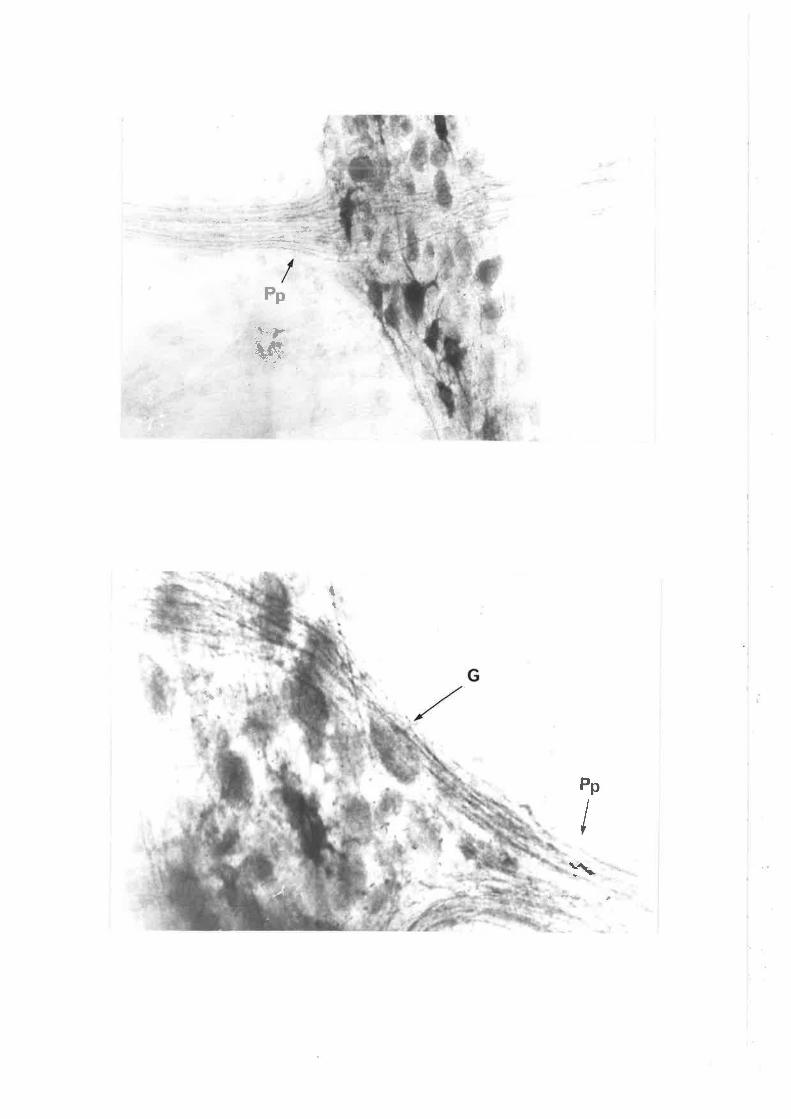
o-a-- /,
II
1-
t,'.' J.r,JI
at
o-o f H:
È r+
fY

t,
tüI a
I 'i N'¡
t
{l,
¡
fÇ,'

60
proximity to each other. In Fig.4, an e'longate neurone distingu"ished
by short emergent processes aborìsed too either end of the soma can be
identified ìying near the Type II neurone.
In addition to being able to trace the processes of indjvidua'l
neurones some distance frorn their orìgin,'it was possible to discern
enpassant fibres cours'ing through the ganglia and aìong the interconneciing
nerve bundles (fascicul'i) of the prinrary nreshv¡ork (Fig. 5).
Elements of the secondary (ps) and tertiary (Pt) rneshworks of the
plexus lvere not sta'ined by thìs method, however extragang:lionjc neurones
(Takeo and Sugai, i974; Gabella, 1979) in the interconnecting fasciculi
of the primary (pp) rnesfrwork lvere sometimes stained (Fig. 6).
Discussion
The NADH-diaphorase (dehydrogenase) histochemical technique used by
Gabella (1967, L969, 1972, L979) afforded demonstration of nerve cell
soma but with on'ly little deliniat'ion of the emergent processes. Conversely
the stainìng assoc'iated with the NADPH-dehydrogenase activity observed in
this study enab'led better observat,ion of the rnorphology of the nerve cells"
It is weìl known that nicotinamide adenine dinucleo'bide (NAD) and nicotjnamide
adenine clinucleotide phosphate (NADP), despite their s'imilarity in molecular
structure, rareìy senve as interchangeabìe hydrogen (electron) acceptors/
donors to the same pyridine linked enzymes (Lehninger! L972). For the most
part NAD-dependerrt react j ons are associ ated wi th steps i n cel l ul ar respi rat'ir.ln ,
therefore the diformazan sta'in formed under these circumstances would be
confined to the mitochondria (Pearse, 196C) iocated to the soma and terminal

61
region of the nerve cell, resulting in little or no staining of the nerve
fibres. The ¡iADP-dependent reactions provide one link by which electrons
can be transferred from catabolic to biosynthetìc pathways (Lehninger' 197?;
pearse, l960) and any diformazan deposits associated with these reactiorrs
would occur mainly in the soluble cytop'lasm afford'ing better staining of
the neurone soma and its processes.
The myenteric pìexus and individual gangì'ionic neurones, as stained
by this NADPH-delrydrogenase histochemical technique, bore a striking
resemblance to neuronal elements of the ileal myenteric plexus stained
by other methods (see Schofield, 1968; Gabella, L979). The numt¡ers cf
intenseìy stained nerve cel I s varied w'ithin ind jvidual gangl ia and bet'lveen
preparations, therefore 'it was not possible to establish the relative
numbers of nrorpho'logicaìly d'ifferent cell types. Furthermore, there
appeared to be no distribution pattern for partìcular cell types, which
is in contrast to the evidence from other studies (see Schofield, 1968).
For example the Dogìe'l Type I ce'|l ìs proposed to be located at the
periphery of the ganglion (Dog'iel, 1895, 1899), but here were found in
all locations within the gangìia. Many l'ighter stained celis were also
present and these may represent a class of nerve cells not particuìar1y
sensit'ive to th'is technique. This could reflect a ìower enzyr-natic activ'ity
for these cells, or ìnadequate cytop'lasmic levels of the electron acceptor
or the co-enzyme.
Endogenous co-enzyme levels are normally too low to allow histochenlical
dehydrogenase react'ions to occur, and many factors operate under histo-
chemical condit'ions to reduce the amount of available co-errzyrne (Pearse,
1960) therefore jt is necessary for co-enzyme levels to be increased and
maintained at high levejs throughout the incubatjon. The occurrence of

62
many intenseìy stained cells indicates the reaction mixture levels of the
co-enzyrne and electron acceptor to be appropriate and suggest instead
differential penetration by the reactants into the nerve cells, presumably
due to the short 'freeze' t'ime used.
There is a vast literature on the morpho'logy of enteric neurones
(see Schofield, 1968; Gabella, 1976), characterised by a wide divergence
of views as to the nature of these nerve cells. It may indeed be impossible
to classify neurones into distinct catergories since it now appears that
enteric neurones comprise a wide spectrum of morphoìog'ica'l types. Nonetheìess,
the NADPH-Cehydrogenase histochemical technique affcrcls researchers a rapid
and sinrple n¡ethod, for idenr-ify'ing various nerve cell types within the
intramural nerve networks of the mammal'ian intestine.

CHAPTER V
uPTAKE AND RELEASE oF t3ur GABA BY
MYENTERIC NERVES
¡

63
Introduetion
The myenteric p'lexus cf guinea-pig, cat and human intestine has been
shown to contain the 0ABA synthesising enzyme gìutam'ic acid decarboxyìase
(GAD) as well as endogenous GABA in low concentration (Jessen et aL., 1979;
Hosoya et aL., 1981; l{iki et aL., 1981). A'lso, neurones of the guinea-pig
myenteric plexus are selectively labejled by [3H]-gngn under conc.litions
favouring h'igh affin'ity uptake, as shown by autoradiography (Jessen et aL,,
1979. Krant'is and Kerr, 1981) which together with the pharmacological
actions of GABA in the 'intestine (Krant'i s et aL., 1980; Krantis and Kerr,
1981;6ng, i9S1), is strongly indicative of a transm'itter role for GABA
in the'intestine., However, even tlre demonstration of a h'igh affinìty
uptake system for GABA js at best onìy supportìt'e evidence for the
identification of GABA as a transmitter, for, even if radiojabelled GABA
js accumulatecl by neuronal elements, the question still remains as to
whether it enters a rapidly re'leasable neurotransrnitter pooì (Orrego,
Lglg). Stronger and more d'irect evidence 'in the enteric nervous system
would be provided by the demonstratjon of an evoked release of GABA from
myenteric plexus preparations, whìch'is considered to be one of the prìntary
and fundamental criteria for neurotransm'itter identìficat,ion (see 0rrego,
1979). This criterion has been 'investigated for several amino acjds in
tissue slices or synaptosome preparations preloaded with radiolabelled
amino acids, and the spontaneous and evoked efflux of radìoactìvity
monitored (Orrego, 1979). Although jnvest'igation of the evoked release)
of IJH]-6ABA from the nryenteric p'lexus js novel , other such preparat'ions
have been used to study the evoked release of raciiolabelled substances
incl ud'ing [3H] adenos'ine 5o-hyc,lroxytryptamine (Su et aL., 1971) , [3H]
5-hydroxytryptamìne (Scnuttz and Cartwright, Ig74), and [3H] acetylchoìine
(Kilbinger and hlessler, 1980).

64
In general , neurally-evoked release of amino acìd transmitters 'is
calcium dependent (Fagg ancl Lane, L979; Rubin, i9B0) and the demonstration
of a calcium dependent release of a substance'is support'ive of a transmjtter
role,although there are comp'l'ications depend'ing on the mode of stimulation
and conditjons under which the effìux study is conducted (Szerb, L979).
Nevertheless calcium dependent, electrically evokecl r-olease of [3H] eRgR
from brain slices has several times been demonstrated (Srinivasan et aL.,
1969; Katz et aL., 1969; Szerb, Lg7g),which pronrpted this attempt at a
corresponding study 'in the enteric nervous system. A problem associated
with interpretatjon of data for such evoked release of amjno acids is the
need to di stìngu'ish whi ch ti ssue el ements accumul ate and rel ease the
radiolabeljed am'ino acid. There is now considerable evidence that gliaì
cells are an irnpor"tant s'ite for amino ac'id uptake (Fagg and Lane, L979)
and glia'l cejls may be depolarized,by high concentrations of potassium or
by electrical stimulat'icn, to nelease non-t,ranstnitter anlino ac'ids (Kuffler
and Nicholls, 1966; Katz et aL., 1969; Watson, Lg74;0rrego et aL" 1976)'
Furthermore, uptake and release of radiolabelled GABA has been demonstrated
in sympathet'ic gangìia (Young et aL., !973; Boutery and Brown, 1976) where
GABA is not ccnsidered to be a transmitter but'is accumulated by gìia.
Hor,rever, the fact that [3H]-engn'is also taken-up'into glia by a h'igh
affinity uptake system is a complication that can largeiy be overcome by
t¡e use cf ß-atanine which'inhibits any GABA uptake into gììal cells
(Schon and Kelly, 1975) thus ensuring that the greatest proportion ofa
[3H]-eRgR releaserl by electrical st'imulation wjll be of neuronal origin.
That a neuraj source is involved in the release of a putative transmitter
from the tissue can be confirmed by preventìng neural actjv'ity with a
neurotoxin and by preventing up""ake of transmitters with specific jnhibitors
of neuronal high affinity uptake. FoTGABA such neuronal uptake 'inhjbitors
are L,Z-4-diarn1nobutyric acjd L-DABA) and cis-3-aminocyclohexane-carboxyf ic
acid (ACHC) (Iversen and Keìly, Lg75; Bowery et aL., 1976), whilst nipecot'ic

65
acid inh.ibjts both 91ìal and neuronal GABA-uptake (Schousboe et ã.L., 1979).
In addressing the problem as to whether GABA js a transmitter in the enterjc
nervous systetn i have now empìoyed the criterja of uptake'inhibìtion, calc'ium
dependence, ancl neurotoxin sensitìvity of release' in order to show that
[3H]-engn can be accumulated by isolated longìtudinal muscle-myenteric
plexus preparatìons of both guinea-p'ig iìeunr and colon and subsequently
released by appropriate electrical st'imulat'ion.
Methods
Guinea-pìgs of eíther sex weighing ?50'400 grams vlere stunned by a
blow to the head and bled. Segments of ileum and of the proxima'l portion
of the dista-l colon were removed and placed'in Krebs (the composition
as described in Chapter II). The solution was bubbled contìnuously with
a gas rnixture of 95% 0, ancl 5% C}Z, and ma'intained at' 36"C. Lengths of
ì ongi tudi nal muscl e coat (3 cm) wi th attached myenteri c pl exus ('l am'inar
preparations) were then djssected from the segments by the method
described in Chapter II and kept in the Krebs solution to which had been
added amino-oxyacetic acid (A0AA),2 x tO-5 N, to inhibit GABA metabolìsm
(|l|allach,1961) and ß-alanine, 10-3 M, to prevent GABA-uptake by gliaì
ceils (schon and Ke]1y, i975). This medium was used in all subsequent
mani pu1 ations.
Individuaj preparations measuring approximatr:ìy 5 mm in wjdth and
25 mnr in length when stretched out, were preincubated in glass'incubatjon
chambers for 10 mjn ìn 350 ul of the perfusion meclìum after whjch t3H]-
GABA was added to gìve a final concentration of 5 x 1C-9 GABA, and
incubation was ccntìnued for a further 20 rnin" The tissues were then

66
removed, b'lotted of excess ìncubatìng medium and suspended between pìatinurn
stimulating electrodes in a glass perfusìon chamber containìng 2 mì of the
perfusion medium maintained at 36"C. The tissue was washed repeatedìy (totai
40 ml) over an equilib,ratjon period of 55 min, to establish a basal efflux of
[3H]-engn, after which superfusjon was commenced and 2 ml fractjons collected
at 30 sec intervals. During th'is superfusion phase electrical stimulation
(Grass S48 Stimulator) was applied for 2 min periods, using 60 V and 3 ms
duration puìse at various frequencies (S to 100 Hz).
To test the'influence of Ca2+ on the stimulated efflux of tritium,
the Krebs solution was substituted by a Caz+-free perfusion med'ium conta'inìng
0.1 mM ethyìeneglycoi-bis-(g-amjno-ethy1 ether) N, N'-teira-acetic acid
(EGTA) a¡d buffered wìth Trjs (¡ mN), the rema'inittg constìtuEents being
as in the Krebs solution, this solution also conta'ined A0AA and ß-alanine.
Superfusate fractions were collected and added to 10 n¡l of a Trìton/
toluene sc'intÍllator, and the radicactivity measured by'liqu'id scintillat'ion
spectrometry us'ing a 10 minute count'ing period in a Packard trj-card
I iqu'id scintil I atic¡n spectrometer model 2002. Count'ing eff iciency (using
an external standard) vras determined at 36 1 1, r = 9. Radioactivity has
been expressed as c.p.m. rather than d.p.m. since variations in countìng
effj ci ency between sampì es vtas negl i gi bì e .
rd.enti.fication o¡ t'nl-CABA reLeased by stir,nLLation
Thin layer chromatography (TLC) on cellu'lose v¡as used to identify
[3H]-gngn in the perfusate. Perfusate samp'les, taken every 30 sec, were
collected over two niìnute perìods and pooled. The first collection was
made Curing basal efflux, iust prior to stimulatjon, and a second was

67
made during the stimulation period. Each pooled sample was concentrated
10 fold by vortex evaporat'ion and the GABA conient adjusted to = 4 mg/ml
by the addition of unlabelled GABA, as was an aliquot of standar¿ [3H]-
GABA. chromatograms were run using the system n-butanol:pyridine:Hrg
(1:1:1 by volume). The dried chroma[ogram was developed with ninhydrin
over a GABA reference to identify 'its final posit'ion on the p'late. The
remainder of the TLC plate was then divided appropriately into 1 cm strips
from origin to solvent front and the cellulose pov,rder of each strip was
scraped Ínto a scintillation vial containing 1 m1 of HrO. The samp'les
h,ere counted in 10 ml of a Triton/toìuene scintillator. Regions yieldìng
tritium counts could then be related to GABA on the ninhydrin treated
segment.s.
compounds used were: ß-alanine, amjno-oxyacetic acid (A0AA) , L,z-4-
diaminobutyrìc acid (L-DABA), nipecot'ic acìd, tetrodotoxin (TTX), (all
sigma), and veratridine (Sandoz). GAtsA (call;iochem, Sigma) and [3H]-onan
(66 Ci mmol-1; 2,3-[3H]-GABA) (Ra¿iochemical Centre, Amersham).
Scintillation fluid:33 gs PPO and I gm Dimethy'l popop dissolved jn a
Triton i(i0/toluene m'ixture (1:2 parts/volume).
ResuLts
upta.ke and. reLease of tSnl-ctp¿.
The majority of experiments were conducted on preparations from the
ileum. As a prel im'inary, longìtudinal muscle-myen-ieric plexus preparations
were incuba.ted for various periods of time r¿¡ittr i3H.]-eABA,5 x 10-9 M, in
the presence of A0AA and ß-alaninei adeauate uptake cccurred within 20 m.in

FIG. 1 (a) The frequency dependent evoked'release of [3H]-
GABA f rom gui nea-p'i g i 1 eaì 'l ongi tudi nal muscl elmyenteri c pl exus
preparation. Each point represents the radioactivity of a 2 ml
fraction collected during a 30 sec period. Abscissa: bars
represent electrical stimulation (2 min) at 10 H2,20 Hz (3 msec
duration, 60 V) respectively. A0AA 2 x 10-5 M and ß-alanine
tO-3 U were present throughout the experìment.
aFIG. 1 (b) "H-GABA efflux during superfusion and electrical
stimulation of longitudÍnal muscle-myenteric pìexus preparations.
r guinea-p'ig distal colon; 6 guinea-pig iìeum. Each point represents
the radioactiv'ity in a 2 ml fraction coìiected during a 30 sec
perìod. Abscissa: bar represents ? nin electrical stimulation
period QA Hz, 3 msec duration, 60 V). A0AA 2 x 10-5 M and ß-
alanine 10-3 M were present throughout the experiment.
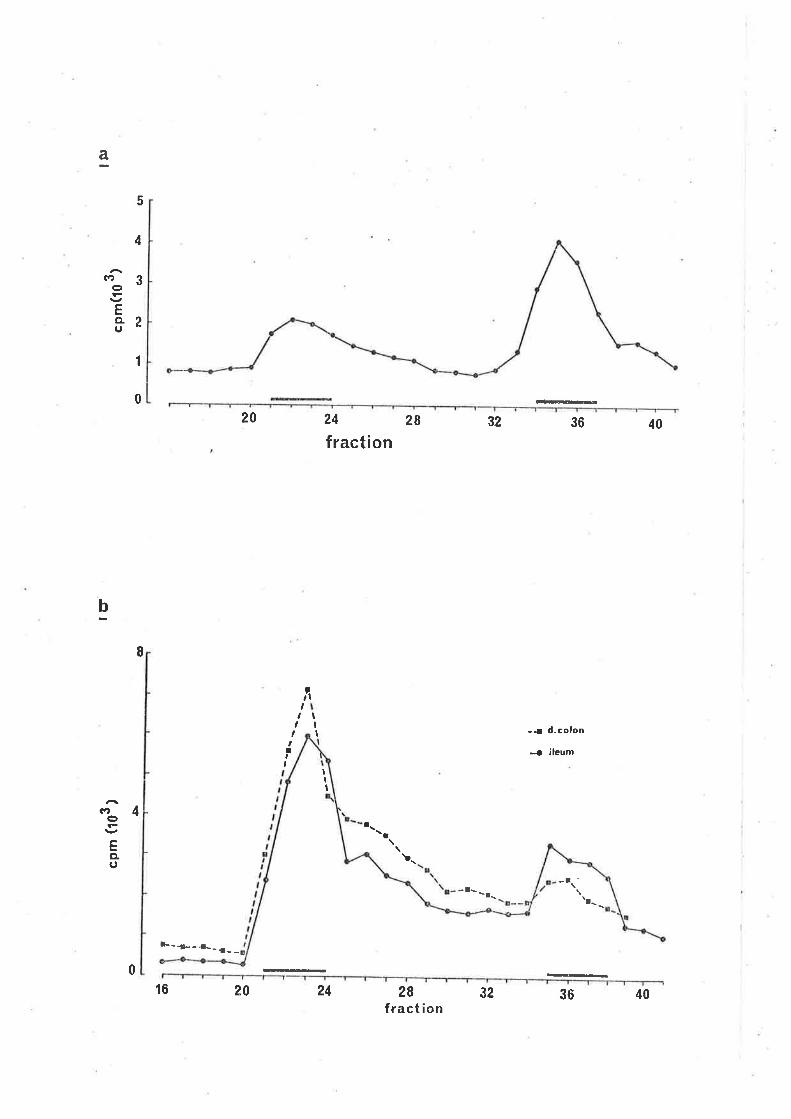
a
5
4
3(v:,
?
I
o
Eo-(J
0 k
403620 24
f raction28 32
þ
I
¡tt¡
,I,I
b\
4Gâo
ECL(J
,Itt
--¡ d.colon
.-¡ ileum
tIt
a\\l
-t\ -¡--a
'¡-
ß-{--.1--a__
0 _-%
28f ract ion
16 20 24 32 36 40

FIG. ?. The effect of tetrodotoxin (TTX) on the stimulated
efflux of [3H]-GABA from guÍnea-pig ileal ìongituciinal mr¡scle-
nlyenterìc plexus preparations. Adiacent segments of ileum were
incubated under the same conditions and then arranged (at random)
in a perfusion bath under the foìlow'ing concfitions and order.
0 normal perfusion medium; 0 normal medium r^iith TTX tO-7 l'l;
E normal medìum; D normal medium with TTX fO-7 l,l. Each poìnt
represents the radioactivity in a 2 m'l fract'ion collected over
a 30 sec pe¡'iod. Abscissa: bar represents a 2 min electrjcal
stimulation period (2C Hz, 3 msec duration, 60 V). A0AA 2 x 10-5
M and ß-alanine 10-3 M were present throughout the experiment.
FIG. 3. The effect of high affinity uptake inhibition on the
basal and stimulated efflux of [3H]-CABA from guinea-p'ig ìong'itudinal
muscle-myenteric pìexus preparations. @ normaì incubation and
perfusion medium EAvrith L-DABA 10-3 present. 0 with nipecotic_a
acid 10-' M present. Each point represents the radioactivity in
a 2 ml fraction collected during a 30 sec period. Abscissa: bar
represents a 2 min electrical stjmulation peniod (20 Hz, 3 msec
duration, 60 V). A0AA 2 x 10-5 M and ß-alanjne 1.0-3 M were
present'ehroughout the experiment.
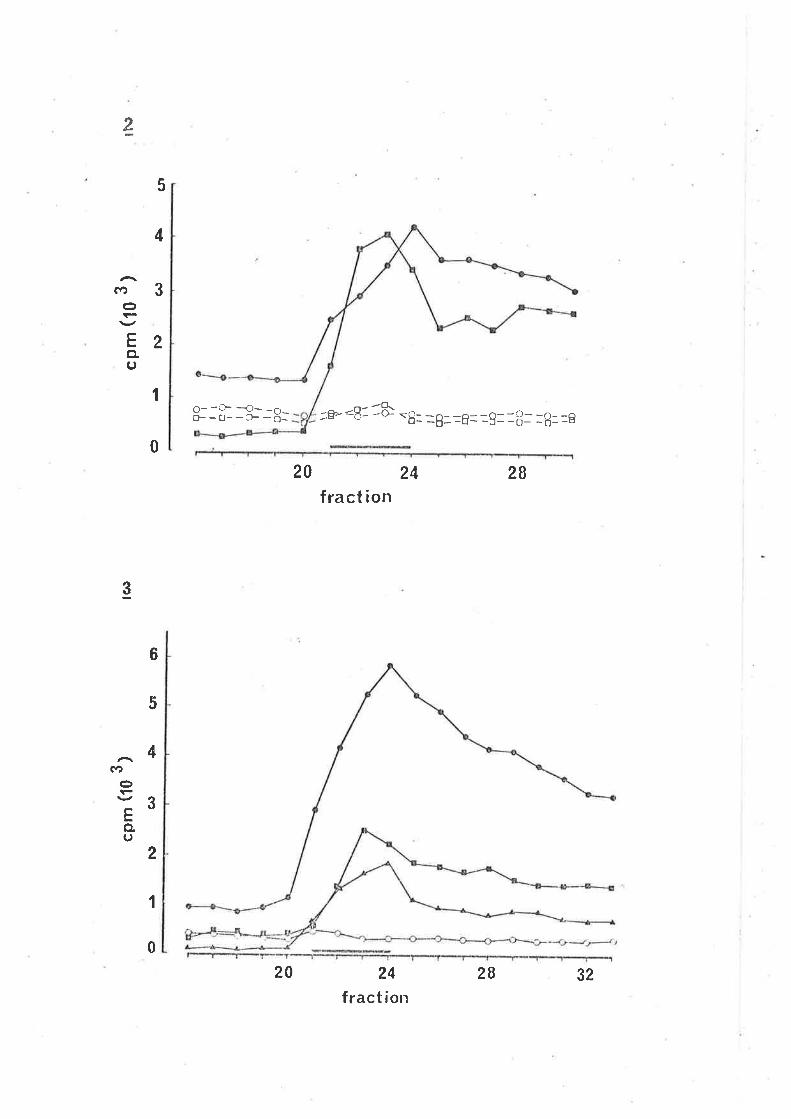
z
(Ðo
Ect,u
3
5
4
2
I
0
o- -'its -{- -6-Þ--CJ---È-c- :È- -}'-lb =B: =B- :B= :B: :Br :g: :Ê
20 24 28
3
f ract ion
2A 24
f ract io¡r
1,)o
Eo.L)
6
5
4
2
1
0
3
2B 32

6B
of incubation, which was adoptecl as the standard tirne for loading the t'issue
with [5H]-GABA. The efflux of tritium from the perfused preparatjon was
followed over the washing period. There were two components in the
unstimulated efflux of tritium: an injtial, rapìd washout-phase with high
tritium content last.ing some 30-40 minutes, followed by a slower relatìvely
constant effiux level n here termed the basal efflux. Once this basal
efflux was achieved, generally r^rìthin one hour, electrical field st'imulation
of the preparatiorr induced an immed'iate increased release of trit'ium (Fjg.
1). The stimulated efflux of tritium reached a maximum wjthìn two minutes
of stimulated aI 20 Hz. These condit'ions were found to be optimal and
used in all subsequent manipuìations. 0n cessation of electrical stjmulation
the release of radioactiv'ity again declined towards the basal efflux level.
Succeeding periods of stimulation at 20 Hz were always less effective in
provok'ing f urther triti urn rel ease.
prepar"ations from the distal colon treated jn the same way dìspìayeC
simiìar efflux characteristics (Fig. i), but were not invest'igated further"
Neither KCI , 56 mFl, nor veratridrine, 5 X tO-S N v,/ere very effective
in stirnulatìng [3U]-'Ongn efflux from the myenteric plexus, beyond and
approxima'ue doubling of counts with KCI depolarization and an almost
undetectable jncrease with veratrid'ine stinlulation. Consequently we used
eìectrical st'imulaticn as the preferred method for evoking [3ä]-CRgR
release from the plexus preparations.
Tetrodotoxin (TTX) (tO-0 M) which woulrj blocl< all neural actìvity vthen
added to the perfusion medium, was effective in vitrual'ly abolishing the
stimulated rr:lease of [3H:J-enen from ileal prqparations (Fjg. 2). These
tests were run on dupìicat.e, separ'ate, tìssues with and wjthout TTX, since
djff iculties v\,ere encounte'red 'in lvash jng-orrt the TTX rapidly enough to
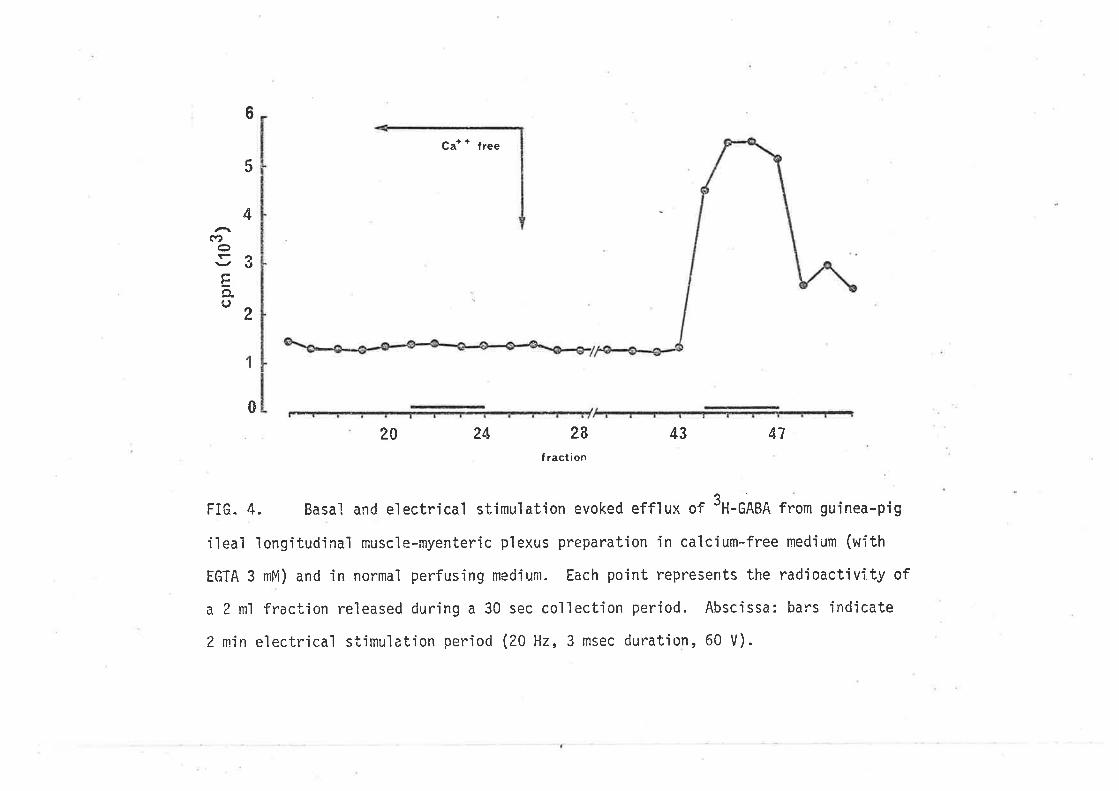
6
La lree
îaê
o.()
5
4
2a,
2
1
0
47
FIG.4. Basal and electrical stimulation evoked eff'lux ot 3H-GRAA from guinea-pig
ilea"l ìongituci'inal muscle-myenteric plexus preparation in calcium-free medium (with
EGTA 3 mM) and in normal perfusing medium. Each point represents the radioactivity of
a 2 m'l fraction reieased during a 30 sec collection period. Abscissa: bars indicate
2 nin electrical sijmulation period (20 Hz, 3 msec duration, 60 V).
20 24 2B
f ract i on
43
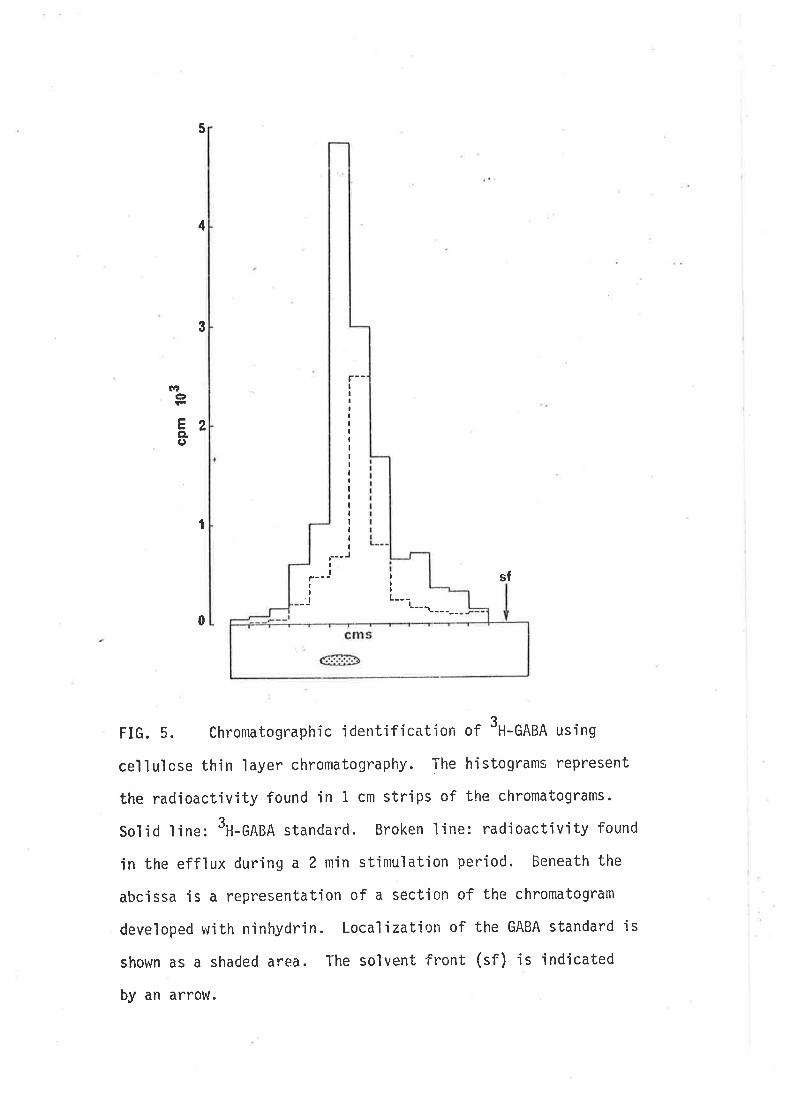
5
4
3
sf
FIG. 5. Chromatographic identification of 3H-eRgR using
cellulcse thin ìayer chromatography. Th. histograms represent
the radioactivity found in 1 cm strips of the chromatograms.
Sotid ìine: 3n-enBR standarcl. Broken line: radioactjvity found
in the efflux during a 2 min stinrulation period. Beneath the
abcissa is a representation of a section of the chromatogram
deve'loped with ninhydrin. Local ization of the GABA standard is
shown as a shaded area. The solvent front (sf) is indicated
by an arrow.
c'i¡
2
o
ECLIJ
1
0
'I
ItIt---
I
f-I
_l
r--III!IIIIIIIIIIIIIIIII
.J
r-'II

69
permit denlonstration of stimulate¿ [3H]-GABA efflux from the washed TTX
treated tissues.
When the ileal tissue was treated with the neüronal GABA-uptake
inhibìtors nipecotic acjd (10-3 M) or L-DABA (tO-g M), before and during
loading witfr [3H]-GABA, the subsequent electrical'ly evokecl efflux of
tritium was reduced, w'ith L-DABA beìng less effective than nipecotic acid
(Fig. 3).
2
CaLcium dependerrce of stirruLated H-GABA reLease
in a further series of experìments, Lhe ileal tissue was preloaded1t+
with [tH]-GABA and then perfused urjth Ca''-free mediurn contajnìng EGTA,
0.1 ml'l. Spontaneous efflux of tritium was unchanged under these cond'itiotrs,
but electrical stimulation no 'longer evoked any additional efflux. Upon
rep'lacìng the perfusion medium with that contcrining a normal Ca2+ concent-
ration, electr''ical stimulation aga'in evokeci an appreciable efflux of
tritium (Fig. 4).
ïd.entifícation o¡ 3u-caal. r'eLeased by eT,ectt'i,ca'/' stírm'Llation
0n TLC plates, the major portion of radioactjvity released by electrical
stimulatjon co-chromatoqraphed wjth standard GABA ¿.nC [3H]-GAIIA, all seen to
have a RO va'lue of 0.45 (Fìg. 5). The trit'ium assoc'iated with the basal
efflux, taken just prìor to stinrulation, rân with a simj'iar R' but, amounted
only to some 5% of the peak stimulated output (not shown). There was no
other major peak of trjtium presented in chromatograms from either the basal
or stimulated efflux.

70
ùLscussion
Spontaneous output of trjt'ium fel'l throughout each experiment, f irs'l
rapidìy then more sìowly. Carry-over of [3H]-ensn from the incubating
medium, as well as washout from non-neural sources, would account for the
bulk of the initial, rapjcl, high-1eve1 efflux.fronl the tjssues during the
early wash'ing perìod. Desp'ite the presence of A0AA in the media some of
thi s t¡iti urn probably consi sted of both [3H]-engn and tnetabol i tes i ncl ud'ing
tritiated water (Gardner and Richards, 1981). The possibi'lìty can be
djscounted that significant [3H]-engn could be releasecl from some non-
neural source âs c1 lêsult of smooth muscle contractions during the
stimu1ation pe¡iodn for the efflux was greatly reduced by jnhibìting ner-lral
[3H]-enBn uptake. It is also unl'ikely that release of [3H]-eRgA tro*
gìia'l cells could contribute significantty to the observed efflux, whether
spontaneous or stimulated, s'ince ß-alanine which was present throughout
and is a more effective inhibitor of GABA-uptake than gììaì (Bowery et aL.,çJ
Lg76; SchousÙe et aL., 1969), yet onìy the st'inrulated efflux \^,as modjf ied
by ihi s i nh'i bi ior.
Autorad'iograLrhic ev'idence shor,/ed that most of the [3H]-engn accumulated
by iìea1'long'iturl'inal-muscle myenterìc-p'lexus preparations was incorporated
into neurones of the plexus under conditiotrs of high affin'ity uptake
(Krantis and Kerr, 1931). It can therefore be assumed that similar
accunrulation of [3U]-GABA haci occurred jn the plexus of the ileum and
colon in the present s'budy. in the autoradìographs there was clear
evidence of cell bodies labelled vriihin ganglia of Auerbach's plexus
together with profuse labelling of processes in'lhe prilnary, secotrdary
and tertìary meshwork of the p'ler.us (Krantis and Kerr, 1981). Furthermoìe'
L-DABA and njpecot'ic acid which are inh'ib'itors of neuronal GABA uptake'
prevented th'is labelling in the autoradiographs (see Chapter III) and

7T
greatly reduced the stjmulated efflux of [3H]-UABA relative to that fro¡¡
untreated tissues. Such labelled neurones and processes are, then, the
most likely source of the [3H]-engn release from the plexus by e'lectrical
stimulation in thjs study
A0AA was present throughout these experiments in order to preventa
metabol'ism of ["H]-GABA after uptake (l,la1'lach, 1961). However, it has
been suggested that A0AA, 'in the concentrations custornarily emp'loyed for
this purpose, is not entireìy effective in blocking GABA catabolìsm
(Gardner and Richards, 1981). Nevertheless the bulk of the radioactivity
measured cluring electrica'l stimulation of the tissue was [3H]-GABA that
co-chromatographed with stanclancl GABA. This neural [3H]-Cngn was reaciily
mobiljzed but rorrO be partiaìly depletecl by st'imulation at 20 Hz for as
little as two minutes, no dclubt represent'ing exhaustion of a l'imited
releasable store. Both tetrodotoxin and calcium-free perfus'ion were able
to stop the stimulated efflux of [3H]-GABA, agaìn confìrming its neural
origin. Frequencìes of stimulatìon about 2Ct Hz were less effective in)
evoking ["H]-GABA release, vrhich would suggest the existence of GABA
autoreceptors capable of depress'irrg outpul, of transmitter by a prejunctional
inhibitory feedback mechanism. Alternatively the release may have been
depressed by some other, co-re'leased, transmítter.
Somewhat surprisingly, it uras found that neither veratrid'ine nor KCI
were as effect.ive as el ectr jcal st'imulation 'in provoking [3H]-GABA effl ux
from ileal pìexus preparatìons, although both have been utilìzed ìn the
studying of such efflux from other tissues (Szer'b, 1979; M'inchin, I979;
Srjnivarson eb aL., 1969). tlh'il st veratridine stimulation distinguishes
neuronal from gì ial sources of transmìtter efflux (Bowery and t'leal , 1978)
the more usual criterion is used as a stimulus. 0n the contrary, veratridine
causes an increased output of transm'itter in the absence of Ca2+ (Minchin,

72
1979; Szerb, i979). Sjnce we d'id not explore the influence of Ca2+ on
this efflux in the irrtestine we have no explanatiott for the relative'ly
small [3H]-engn efflux evoked by veratridine or KCI in the present study,
partìcularly since KCI might be expected to release [3H]-gnen from both
gl ia1 and neuronal sources (Bowery and Neal ' 1978).
There are conf 'l i cti ng reports concernì ng ca'lcì um dependence of
transmitter ouiput when using electrical stjmulation. Although Szerb
(1979) showed such dependence, Srinivasan et aL. (1969) found a slightly
increased [3H]-engn ef'Flux from electrically stimulated brain slices in
the presence of iow CaZ+ without chelating agent, which is to be expecterj
since there would likely lre an electrjcaììy-hyperexcìtable phase during
progressive deplet'ion of membrane Ca2+ under these condjtions. In the
present study this was avo'ided by the combinecl use of EGTA and no CaZ+
in the med'ium, thus more rapidly depleting Ca2+ from the rnembranes. There
is the possibil ìty that such treatment, particular]y 'if proìonged, mìght
damage the t'issue and prevent transnlitter output, but evidently th'is djd
not occur for there was a prompt reiurn of'bhe stimulated 3H-GABA et"flux
upon restor"ing the Ca2+ content in the perfusion medium to nor¡nal. tlectrical
stimulation thus remains a valid method for eliciting putat'ive transrnitter
output from the functional myenteric plexus, as has a'lready been sholvn in
the ileum for cholinergìc transmissjon uslng [3tt]-choline (Szerb , L976;
Kilbinger and Wessìer, 1980) and for [3H]-S-¡rydroxytryptamine (Schulz and
CartvrrÍght, Lg74; Jonaka j t et aL., IgTg) as wel I as foi [3it]-u¿unos'ine
triphosphate from taenja coli fo the grrinea-pig (Su et aL., lgTI). t3Hl-
GABA released fron the nryenteric plexus is of neuraj origin, the re'ieas*
2+being Ca'' dependent a.nd tetrodotoxjn sensjt'ive, indicative of GABA
being a transmìtter in the gu'inea-pig enteric nervous system.

CHAPTER VI
THE EFFECTS OF GABA ANTAGON I SIVI ON I NTESTI NAt- MOTI LITY

73
fntroductí'on
The extensjve ramificat'ions of GABA-ergic neuronal elements w'ithin
the myenteric plexus and their innervation of the circular muscle layer
suggests that GABA releasecl fronr such neurones could be'involved in some
physiologicaì function of the guinea-pig'intestine. Several funct'ions
are carried out by this organ includjng secret'ion, absorptìon, digest'ive
m.ixing (segment.ation) and propt¡lsive activ'ity (pe¡istalsis)' The
pharmacologicaì act'ions of appl'îed GABA result in the stimulation of the
final motor nerves mediating the respectìve inhibìtory and excitatory
reflexes underìying perìstalsis, jnclicating that GABA-ergic mechanjsms
may very well be,involved in the regulatjon of gastrointestinal motìlity.
There have, hclvever, So far been fev¡ investjgations of any GABA-ergìc
neuronal .invol vement 'in peri s+.al s j s beyond the observat'ion that app'l i ed
GABA inmed.iately depresses peristalsis'in the isolated mammalian ijeum
(Hobbiger, 1958a, b; Inouye et aL., 1960). Although this effect does nct
in itself djrectjy l'mpficate any GABA-ergic mechanism jn the control of
peristalsis such an involvement may very well occur.
Investigation of peristals'is and its unclerly'ing mechanisms is diffìcult
since the intninsic ne,-\r,ous controi of the gastrointestìnal tract'is very
complex and not conrpletely understood (see chapter i). Baylìss and Starl'ing
(1g9g) gave the first cleta'iled account of nerve-mediated 'reflexes' under-
lying the passage of substances +.hrough the jntestitre, wh'ich bhey called
,peristalsjs'. Frorn'bhe'ir experìments on anaesthetìzeC dogs, rabbits and
cats , Bayì .iss and Starf ing (1899, 1900 , 1901) showed 'tocal st'imLrl at'ion of
the intest.ine wall to el jcit 'reflexes" a wave of contractjon, (excita-tion)
above and relxation i¡ front of the po'int of stimulation and t^raves o'F
contractions which travel in an aboral direction. Furtherrnore, they
surmised that these e.ffects were coordinated and involved local neuronal

74
mechanisms resid'ing w'ith'in the Auert¡ach's (myenterìc) plexus. The init'ial
observatjons of Bayliss and Starling were elaborated upon by Trendeìenburg
(1917) who devised the now classical experìmental preparations whereby
per.i staì t-ic movements i n the i ntesti ne of the gu inea-pì g or rabbi t were
elicìted by elevat.ion of intraluminal pressure, wh'ich in turn caused radìal
stretch. Sensory receptors located in the wall o't the intestine subsequeniì.y
detect the distens.ion and trigger contractjon or relaxation of the respectjve
intestjnal reg-ions. Trendelenburg d'istingu'ished two phases of peristals'is;
a preparatory and enrpty'ing phase. The preparatory phase, which results
in the relaxat'ion of the intestine is not itself able to coordinate the
onr,¡ard movement of i ntesti nal contents , rather, i t abol i shes acti vi ty 'in
the regìon immediately in front of the cont.ents, and onward movement
resuits from contract'ion of the intestine wall behind the contents. The
involvement of chol iner.gic motorneurones in the 'excitatory reflex' !{as
determjned by Bayì'iss and starìing (1899) and Trendelenburg (I9t7 ), and
since confirmed by more recent studies (Hirst and McKirdy, t974; Costa
anci Furness, lg76). At.ropine and hyosc'ine blocked propu'lsion of the
intest.inal contents by disrupting the excitatory reflex hourever the
inhìbitory reflex remained intact.
The existence o.F cholinergic interneurones within the neural pathways
for,peristalsjs,was first reported by Feldb'urg and Lin (19a9) and Paton
and Zairnis (lgag). They showed the enrptying plrase to be inhibìted by the
nicotin-ic gangl jon blocking aç1ents, tubocurarìne and hexamethonium.
Us.ing methods different from the classical Trendelenburg method for
elic'iting 'peristalsis', Hukuhara et aL. (1958) proved that local
mechanical or chemicai st.'imul atiorr of the intestinal mucosa of denervated,
dog small intestine induced perìstaltic movements which were abolished by
hexamethon'ium. Furthermore, crema et aL. (1970) and Frigo and Lecchinì
(19i0) showed propulsive movemerrt.s of cat or guÍnea-pìg colon induced by

75
a solid bolus to be aboìished by hexamethonium. The inhjbitory reflex has
al so been shown to be po]ysynaptic, 'invol v'ing chol inerEic interneurones
which in tunn stimulate the intrins'ic inh'it¡itory motor neurones leading to
the inh1bitjon eljcited by this pathway (Kosterlitz, 1968)' The transmitter
released at the neuro-effector junctions for this inh'ibitory reflex, is
stiìl unknown. (see'intrinsic'innervation"'chapter I)'
Accorcling to Kosterl itz et aL. (tgso) tne 'emptyìng phase' descrjbed
by Bayliss and starfing (1899) does not'involve a coordination of
contractions wìth.in the two muscre ìayers of the intestine wall. Rather'
contractions of either muscle'is dependent upon the degree of radial
stretch, w'ith a greater distention bejng requìred to trigger contractions
of the ci r"cul ar muscl e. The contracti ons of the I ong'itud'inal and ci rcul ar
muscle ìayers usually follow each other. However, high concentrations of
acety'lchol.ine, carbamylchof ine, or histamine block the longitudinal musc'le'
without affecting the circular muscle Kosteri'itz (1968) and Hukuhara and
Fukuda (1965) suggestecl that shortening of the long nruscle was a consequence
of the dì1atìon of the circular muscle'layer, however Kosterlitz and l'latt
(1965) pointed out that during the intervening period between circular
muscle contract'ing the longitudinal muscle remains 'extended', which
suggests th'is explanat'ion ís inadequate' tlectrophysìologicaì evidence
(Kottegoda, 1970), shovrs that jn transmuraììy stimuiated segments of
i ntesti ne the c'i rcul ar muscl e are j nhì bi ted lvhi I st the l ong'i tudi nal
muscle hras exci+.ed anci vice Versa, suggest'ing a reciprocal rather than
sequential excitation and jnhibitjon of both rnuscle coaüs during peristalsis'
However, as Hìrst (1979) pointed out, transnrural sb'inrulation does not allovr
any observatìon of tempora'f separat'ion be"uv¡een excitatjon and'inhibition'
Thus these results cannot be int.erpretated. whether contractjons of the
tongituclinal muscle during the preparatory pherse 'is due to some neural
activation ìs also unclear. Kosterlitz (1968) described the contractions

76
to be abolished by TTX, atropine and morphine, however, Hirs'L et aL.
(1975) when recordíng several centimetres from ihe point of distension
found no electrophysio'logiç¿1 correlates for neural activity during
longitudìnal muscie contraction in isolated segments of intestine.
InterestinglV, Feìdburg and Lin (1949) found local anaesthetjcs did not
b'lock contract'ions of the long musc'le during the preparatory phase.
Krantis ei aL, (1980) have shown that GABA induces relaxations of the
guínea-pig iìeal and colon longitudinal muscle coat which are blocked by
TTX indicating that GABA has no direct effect upon the muscle. Adrenergìc
blockade of longitudinal muscle strips devoid of an-v attached nerve plexus
did not prevent the GABA-jnduced responses, suggesting they were mediateci
by the non-aclren..gi., non-cholinergìc inhibitory neurones, intrÍns'ic
to the íntestine. Furthermore, when recording nrechanicaì activity from
both muscle coats together, the action of GABA was simultaneous and the
same. These findings suggest an intrinsic inhibÍtory innervation of the
Euinea-pig longìtudinal muscle. According to Taxi (1965) and Gabella
(7972) the icngitudinal muscle does not receive any direct nervous
innervation. However the close proxim'ity (nanomet,*es) of vesiculated
axonal processes on the surface of the myenteric plexus tcl the muscle
would allow ihe innervation of the nruscle'layer by diffusion of a neuro-
transmit+*er substance released from these neuronaì processes. The findings
of Kranti's et aL. (1980) equally disprove the idea of recìprocaì or
sequentìaì relatiorrships between the two muscle coats although it is
possible that some integrative facility wì I-,hin the m.l,enteric p]exus could
give rise to physl'c'log'ical events which could lead tc cr:ordìnation of
the muscle coats in these ways.
The exact site of the sensory receptors mediating the coup'!ing between
radial stretch and muscular activity is st-ill unknor.,.:n. Bulbring et aL. (ig58)

77
showed that removal of the mucous membrane abolishes the reflex contraction
of gui nea-pi g i 1ea'l c'ircul ar muscl e. Further.more, they demonstrated the
mucous membrane to be fnnervated by neurones aris'ing from the gangl'ia of
the submucous plexus, corrobcrated by the rnorpholog'icaì description of
Schofield (1963) for nervous innervation of the intestine wall. However,
Ginzel (1959a, b) showed t.hat coagulat'ion of the mucot¡s metnbrane by sì1ver
nitrate does not abolish the peristaltic reflex, and although an ileal
segment sheds most of its mucous membrane whilst in an organ bath preparation,
the reflex response to d'istensjon remains (Diament, 1961). This apparent
difference in results could be expla'ined on the basis of the different
stimuli used to elicit the respective reflex cont'ractions, or by sensory
receptors being also located within the remain'ing'layer of the'inbest-ine
wall. Interestingly Frigo and Lecch'ini (1970) showed propulsion of a
bolus dicl not occur in intestinal segments devoid of the submucosal and
mucosa1 layers, and Hirst (1975) showed removal of the submucous plexus
disrupts the radjal stretch ìndurced descendjng excitation but not the
inhibition. However, according to Costa and Furness (1976) app]ying a
ìocalised stretch to the intestine wal'l by means of a weìght elicits a
relaxation anal (descend'ing inh'ibit'ion) and contract'ion oral (ascend'ing
excitation) to the pojnt of distensjon wh'ich are unaffected by the
removal of the submi¡cous pl exus and mucosa '
The controversy surrounding the importance of the three intrinsic
nerve pathways; descend'inE inhibitory, descenrling excitatory and ascendìng
excitatory, in the peristaltic reflex may mereìy ref'lect bhe nature of the
respective sensory receptors and the'ir locat'ion ulithin the wall of tl¡e
intestine. Therefore a'lthough peristalsis can readiiy be demonstrated'in
isolated preparations of the intest'ine and the local responses recorded,
anaìysis js made djfficult by the complicateci nature of the underlyjrìg nerve
pathways v¡hich are not 'fu'l'ly elucidated and show differing susceptibiì ity

78
to spec'if ic mocles of stimul ation '
If GABA-ergic mechanisms are involved jn the control of peristals'is"
it would best be demonstrated by GABA antagonism'in the myenteric plexus
usìng GABA tachyphyrax.is, or bicucu'lrine. pe.istarsìs as init'iated by
faecal peì'lets would approximate as neariy as possible to the natural
situation. Therefore modificat'ion of peristals'is by such antagonism as
measured by faecai pelìet propulsion jn the isolated guinea-p'ig d'ista'l
colon was used in the following study'
Materi.aLs ar¿d Methods
Guinea-pigs of e'ither sex weighing 250-400 g were stunned by a blow
to the head and bled. segments of the distal colon were renovecl and
placeC in Krebs solutio¡ of the fol1owíng composit'ion [ml4]: Na+ 151'0;
K+ 4.7; ¡rg2+ a.6; ca}+ 2.8; cl- 143.7; HrPLo- 1'3; s0o2- o'o; g'lucose 7'7'
The solut'ion was bubbled with a gas mjxture of 95% 0, and 5% c0, and
ma'i nta i ned at 35 o
C .
Two experimental arrangements were used to exam'ine the effects of GABA
antagon.ism on intestjnal motif ity. Freshly excised segments clf distai ccrlcn
contain.ing faeca.r perìeLs were placed'into warmed Krebs sorution and tne sub-
sequent number of peilets expelled over 2 mirl intervals was recorded in the
presence and absence of GABA ant.agonists. In a further series of e;tperìments
similar segments cf distal colon, etnpty of faecal peì1ets' r!'ere arranged as
described by Costa and Furness (Lg76) for recording the passage of faecal
pellets aìong ìsolatecl segments of intestine. Pellets of Faeces, previousìy
dried and coateci with polyurethane, were placed into the lunlen at the oral end of

79
the segment at 10 min intervals, and their passage along a measured port.ion
of the segment was timed. Act'ivity of circular muscle durìng peristalsis
was recorded by Grass FT03C transducers, using ? g/cn springs connected by
cìips to the serosa of the colon wall.
Drug volufies were never more than l% of the bath volume.
were: bicuculline (K & K Ltd.); gamma-aminobutyrjc acid (CABA)
5-hydroxytryptam'ine (Si gma ) .
Drugs used
(Si gma ) ,
ResuLùs
As prevìous1y described (KrantiS et aL., 1980), upon the addition of_Ã
GABA 3.10 " g/mì to the bath an immediate transient relaxatjcn of the
colon was observed (Fig. 1). After equiìÍbrium in the presence of lhis
concentration of GABA, a further 3.10-5 g/ml cf GABA rqas adcled to
the bath without effect, confirming that desensitìzation of GABA. had
occurred. Moti'l'ity, as measured by the rate of faecal pe11et expu'ision
from the freshly excised cclon, was s'ign'if icantly reduced by such
desensitization to GABA 3 x 10-5 g/nrl or by treatment with bicuculline-Â -Ã5 x 10 " g/ml,3.7 x 10-'g/mì (Fig. 2). 0n occasion, pellet expuìsion
entìrely ceased ciuring GABA tachyphylaxis. The reiducticn in the rate of
pe11et expuìsion lvas readily reversed by washing uiith Krebs solutjon.
The mean propulsive speeci of the coated faecal Fel'leis as tjmed
along isolated segments of distal colon Iay in the raflge 0.5-.1.88 nrn/sec,
comparab'le to that observed by Costa and Furness (1976), and uras reduced
on average by some 60% fa11ow'ing the establishment of GABA-tachyphylaxjs,
Agai n, in some 'instances the passage of the pel 1et compl ete ly sioppe,J

FIG. 1. Recording of circular muscle activìty from two points
in a segment of djstal colon during the addìtion of GABA.
Recording 0 is from a point 20 mm oral to the recording point A.
-qGABA 3 x 10 " g/ml when first adminjstered induced a relaxation
of the distal colon. A second concentration of GABA 3 x 10-5
g/nl, was aCded to the bath'ing soìut'ion u¡ithout washing out the
previously app'lied GABA. The development of tachyphylaxis to
applied {ìABA is evident from the lack of effect of the chalienge
concentration of GABA"
FIG. 2. The effect of GABA antagonism on þìellet expulsion from
the guinea-pÍg distaì coion. Ordinate, percentage of control
expuìsion rate. Abcissa, histogram columns (rnean and s.e. of 4-6
preparations) (C) contro'l; (B) in the presence of bicuculline
5 x 10-6 g1ml ; (81) bicuculline 3.7 x 10-5 g/m1 ; (,G) in the presence
of a desensitiz'ing concentration of GABA, 3 x 10-5 g/m1 .
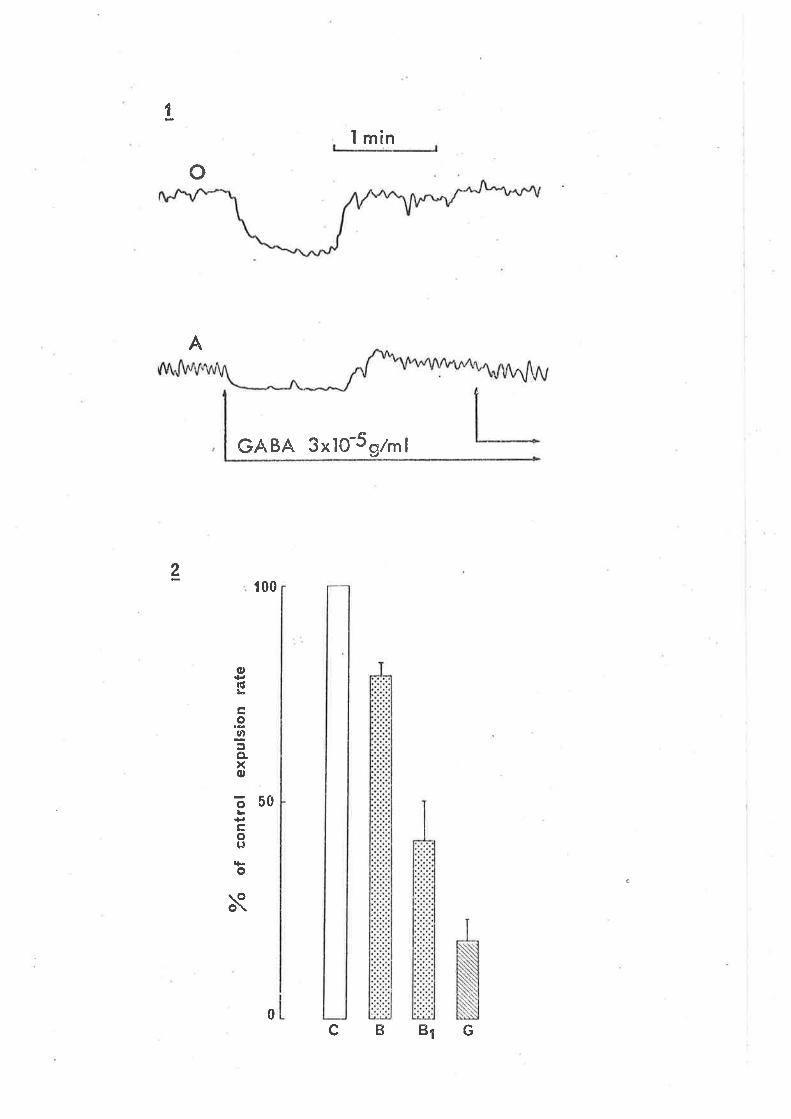
!I min
GA BA 3xtg_Sg/m I
o
A
2t00
G
or1lL
tr.9!CLxo
ã50L
Êo(J
o
\oo\
0CBBlG
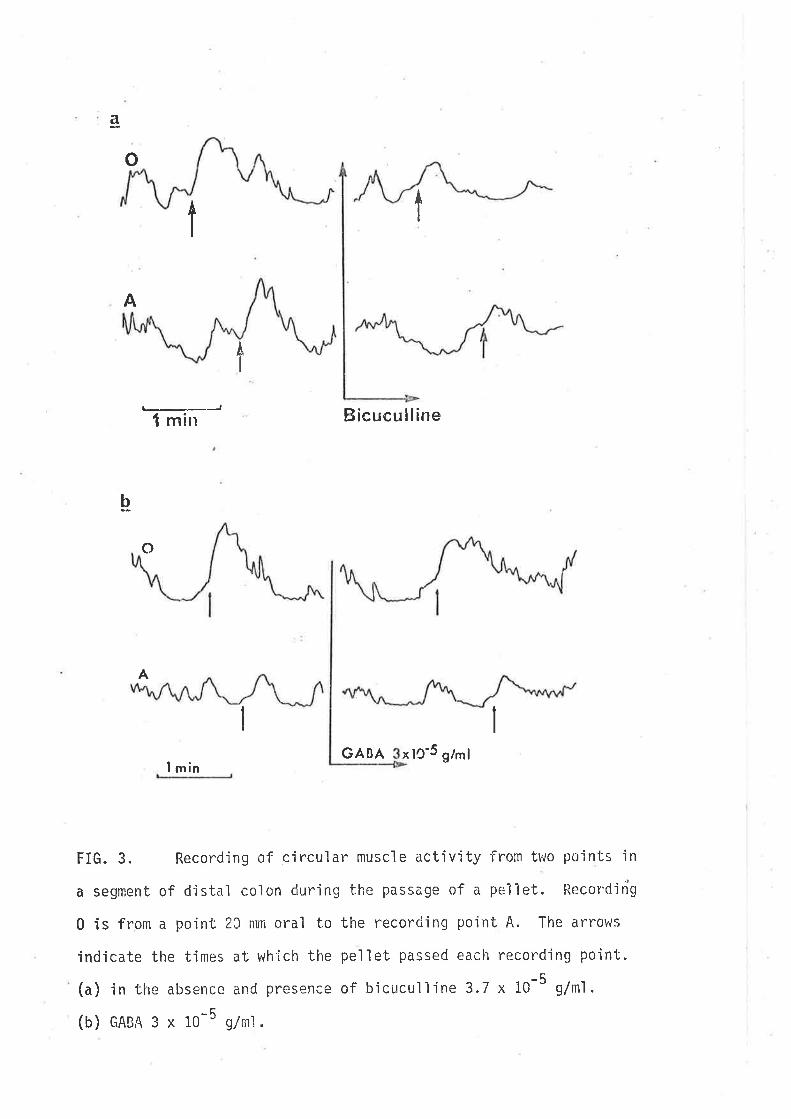
4
o
A
þ
t
t
I min
o
I min
Bicuculline
GABA x l0-5 g/ml
A
FIG" 3. Recording of circular musc'le activity from two points in
a segment of dist,rl colon during the passage of a pejlet. Recorclirig
0 is from a point 20 nm oral t.o the recorci'ing pojnt A. The arrows
indicate the times at whjch the pe'llet passed each recording poìnt.
(a) in the absence and presence of bjcuculljne 3.7 x 10-5 9/ml.
(b) GABA 3 x 10-5 9/ml.
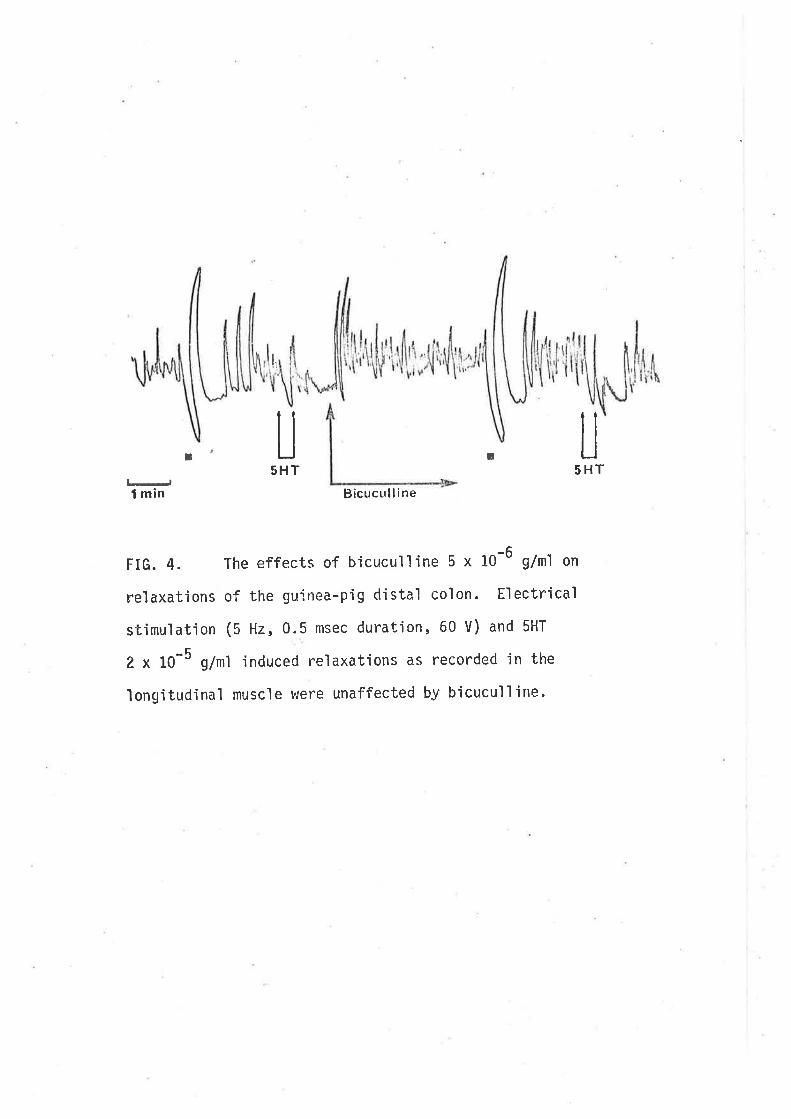
h
tJ5HT
l min Bicucu lline
F'IG. 4. The effects of bicuculline 5 x 10-6 g/ml on
relaxations of the guÍnea-pig dista'l colon. Electrical
stimulatjon (5 Hz, 0.5 msec duration, 60 V) and 5HT
2 x 10-5 g/ml induced reìaxations as recorded in the
longitudinal muscle were unaffected by bicuculline.
IJ5HT
I I

BO
during GABA tachyphylaxìs. Repeated washing of the t'issue with normal
Krebs solution reversed the effect of tachypliylaxis, and the slowing of
propuìsion was readily reproduc'ible upon re-instituting GABA-tachyphy'laxìs
in the same t'issue.
Bicuculline 3.7 x tr0-5 g/ml, also reduced the ntean propulsive speed
of the coated pel'lets by some 50%, and this action, too' was read'iìy
rever"sed follolving washout from the bath. The reduc.tion in propuls'ive
speed was dose-dependent and reproclucible on the sante tissue. The passage
of the pellet was never stopped in the presence of b'icuculline coupling.
During the cnset of GABA antagon'isnt folìowing addition of bicucull'ine
to the batl¡ phasic activity, recorded from the circular muscle,l.las briefly
dampened, but the overa'll 'tone'of the circular muscle was not signifìcant'ly
altered and normal activìty was restored wjthìn a few minutes. In other
preparations where GABA tachyphylaxis was employed, a'l'l recorded mechanical
activity in the circular muscle fjrst ceased, often with a transient
relaxatjon jnmedjateìy following addition of GABA 3 x 10-5 g/ml to the
bath, and then sìoviiy recovered during the onset of tachyphylaxis. The
subsequent chalietige dose of 3 x 10-5 g/rn'l r^ias lvithout effect. Circular
muscle act'ivir;y Curing peristalsis was modified both by GABA tachyphylaxis
and by application of bicuculf ine, there beìng most often a recluction ìn
the arrrpl jtude o1' the.contraction of c'ircular muscle behind the pellet,
fol'lor,ríng ìts passage beneath the recording poìnt (Fig. 3). 0n'l.y
occasionally was there any s'ignificant alteration o'f the relaxation
preceedìng the passage of the pe1'let. In a separate experiment segmenis
of distal colon vrere ðrranged as descrjbed in Chapter iI (Methods) so
that the effects o't electrical and pharmaccl'iogicaì stjinulation could be
analyzeC. In the presence of atropi ne, 2 x I0-7 g/in1, transmural siimulaiion
(5 l{2,0.5 nls,60 V) and 5HT,2 x 1.0-5 g/ml ìrrduced relaxations of the distal

81
-6colon wh'ich were unaffected by bicuculline 5 x 10 e/n1 (Fi g. 4) .
ùiscussion
Evidently GABA-ergic mechanisms are jn some way concerned wjth
maint,aining normal peristalsis, since'it has now been shown that the rate
of propulsion of faecal peì1ets in the colon is slowed, and the amplitude
of the contraction behìnd the peìlet is most often reduced during GABA
antagonism by tachyphy'laxis or b'icuculline. The reduction in propuls'ive
activìty by bicucullìne does not appear to be due to any possible act'ions
on cholinergic components of peristalsis for b'icuculf ine, at the concent-
rations used in thjs study, does not alter ac;etylcholine induced responses
in the colon or ileum (see Chapter II). Furthermore, tetrodotoxin blocks
alì GABA actions'in the jntestine (Krantis et aL., 1980), so that the
reduced mot'ility w'ith GABA-antagonism canttot be due to any dìrect motor
or inhibitory actions of GABA on the smooth muscle of the intestine.
Rather, applied GABA, act'ing thr ough bicuculline-sensitive receptors,
stimulates intrinsic neurones of the myenterìc plexus (K.rantis et øL.,
1980; Krantis and Kerr, 1981b), and ev'idently antagonìsm of the actions of
these neurones is responsible for the alterat'ions in mot'i'liiy reported
here. There is thus the strong possìbiì'ity thoü GABA-ergic interneurones
or sensory neurones are involr.'ed in normal peristalsis.
Peristalsìs is usually described jn tet'ms of an ascend'ing excitaticn
and a descendi ng 'inh'ibi ti on resul ti ng f rom a i ocal stimul us , the Law of
the Intest'ine (Bayìiss and Starìing, iB99). 0f these, there is generaì
agreement concern'inE the descend'ing inhibition, but there remains some
controversy as to the nature of the 'excitation'phase contribub'ing to

83
descend'ing exc'itation local'ised to the submucous mucosal layers (Hirst,
lgTg) and that for the ascending exc'itation most probably localised to the
myenteric plexus (Costa and Furness, 1976)
According to Krantis et aL. (1980) GABA antagonìsm' by GABA tachyphylaxìs
or bicuculline, does not alter the ascending exc'itatory or the descending
inhibitory refìexes elicited by a ìocalised stretch of the 'intest'inal wal'l ,
and yet the contraction of the circuiar muscle behind the pellet was
reduced by GABA antagonism'in the present study. In th'is regard, it should
be emphasizecl as Frigo and Lecchini (1970) have already pointed out that a
faecal pel'let is a different and evidently more physioìogical stìmulus for
elicitìng peristalsis than is localised stretch of the kind used by Cost'a
and Furness (1976), as well as by Krantis et a.L. (1980). It thus appears
that radial stretch of the intestine wall by a pe'llet or ìntramural
balloon preferentjally elicits a reflex excitation of the intestine
(Kosterliiz, 1968; Frigo and Lecch'ini, 1970) by a differerrt neuronal
pathway to t,hat stimulated by localised stretch of the intest'ine wall.
A comparab'le depression of the descending'inhibition was seldom seen
in this study, although the GABA receptors on the inhibitory neurones
would suggest that it could occur. Fajlure to observe any such depress'ion
may reflect an inappropriate experimental des'ign for its demonstration 'in
the present experiments. Therefore from the pr"esent results it appears
that the contraction of the colon immediately behind the faecal. pel'let,
eliciled by radial stretch of the intestinal waì1,'invqlves some GABA-
ergic relay, probably 'interneurona'l , the a.ntagon'ism of which is sufficient
to interfere with peristalsis and slow or stop pe1ìet propulsion.
It is generally accepted that the pathv,rays mediating excitatory and
i nhi bi tory peri stal t'ic ref I exes i rivol ve chol i nergi c i nterneurones act'ing

84
through nicotinic-chol inerg'ic receptors (Akubue, 1966; Burnsto ck ni ol,.,
1966; Kosterlitzo 1968) yet the stimulating effects of GABA on the intest'ine
are not blocked by the nicotinic antagonjst pento'liniu¡n (Krantìs et aL.,
1980), so that GABA actions ìn the intestine can not be relayed through
chol Ínerg'ic jnterneurones.
Hobbiger (1958b) and Inouye et aL. (1960) have shown that GABA
antagonìzes the contracti'le responses of the guinea-pig i'leum to nicotine
and 5-hydroxytryptamine stimulation of the cholinergic motor neurones.
However, it seems unlikely that interactions between GABA and 5HT could
explain the present results which are based on GABA antagonsim since itwas found that SHT actions in the guìnea-pig d'istaì colon are unaltered
by GABA antagonism cf the kind shown here to slow peristalsis.
Preiunct'ional GABA (GABAB) receptors that are bicuculline jnsensitive
and stimulated by Bac'lofen have been iclentifjed jn the gu'inea-pìg'intestine
(Bowery et aL., 1981; Ong, 1981). These receptors nrediate a reduction in
cholinergic nerve activity of the ijeum and colon and it'is possible
that activation of these receptors may contribute to the GABA-ergic
controj of peristalsis. These receptors cannot be involved in the slowing
of peristalsis by bicuculline. However, such an involvement cannot be
excluded for the effect of applied GABA since GABA would act at both
GABAA (bicucuiline sensitive) and GABAB rer:ep+*or populations. Indeed
antagon'ism by desensitjzation to GABA, in contrast to bicuculìine,
complete'ly stopped peì'lei propul sion (s imilar to the f indings of Ong (1981)
who also observed that antagonìsm of GABAA .l_!l.ltlq receptors slowed
pel'let propulsion in the djstal colon, anci that these effects were additive
resulting 'in a cessat-ion of propulsjve activity).
Many poss'ible transmitterso or modulatons, ane now being proposed 'f,or

85
the manrmalian enteric nervous system, among which we would include GABA
as an excitatory transmitter in the guinea-pig intestine where it
evidently plays a key role in the control of peristalsis, through
interactions at an excitatory and an inhibitory receptor population.
L
I
1
I
I
l
I
I
I

CHAPTER ViI
MYTNTERIC PURINERGIC NIRVES

B6
fntroduction
The term ' puri nerg.ic , was f i rst use,J by Burnstock ( i971) to descri be
a thìrd autonomic nerve component which is nejther adrerrerg'ic nor chol'inergìc
but is thought by him to release as its principa] active substance a purine
nucleotÌde. The existence of a third class of ar¡tonomic nerves was suggested
as early as 1E98, when Langley reported that vagal stimulation produced
relaxa-tion of the stomach. it was subsequently shown that this 'relaxatìon'
coul d best be reveal ed f ol 'l owì ng chol 'inergi c bl ockade (McSwi ney and Robson ,
ß?g). since adrenergic blockacle did not prevent the jnhjbitory response
of the gu.inea-pig stomach to vagal stimulation (Greef e'o aL." t962) wh'ils+"
el ectr.ical ly i nduced hyperpol ari zat'ions or j nhi b'i tory iuncti on potenti al s
(l¿p,s) in guìnea-p'ig taen'ia coli smooth muscle cel'ls pers'isted ìn the
presence of both atrop'ine and gaunethid'ine (Burnstock e'b aL., 1963a, b; 1964),
it was proposed that the responses were clue to the release of a non-
chol inergic non-adrenerg'ic substance. In add'ition, these responses t^,ere
prevented by 'low concentrat'ions of the neurotoxin tetrodotoxin (ffX1
(oulbring ancl Tomi'ba, i967), or cold storage of the tissue at 4oc (Kuriyama
et aL., !967), which destroys nerves but leaves smooth muscle viable,
suggesting a neuronal scurce for this non-aclrenergic non-chofinergic
substance. In ackJitìon, the effects of ganglion stimujatrts and blockers
on isolatecl mammalian gut segments (Burnstocl: et aL., 1966), indìcate
that the neurones'involvecl are intrins'ic to the gut, with the'ir soma
located to the Auerbach's p'lextts (myenteric pìexus)' More recent studies
(reviewed in Burnstock, lg72) have since conf irmetl th'is hypothes'is.
There is now considerable evidence for the function of these non-
adrenerg.ic non-cho1ìnergic inhibitory nerves in the mammalian gastro-
intestinal tract (see Burnstock, lg72), including mediation of the reflex
relaxation of sph'incters (oesophogastric, itlterna'ì anai ); receptìve

87
relaxation of the stomach facilitating increased stomach size with food
intake, the descending ìnhìbitory component of peristalsis and the
mesenterjc vasodilator reflex. Nerve-rnediated jnhibitory responses that
involve neither cholinerg'ic nor adrenerg'ic neurones have also been described
in the mammalian urinary tract, ìung, eye and other organs (Appazelìer, 1976;
Burnstock and Be'll, L974 Burnstock, 1972, 1975, 1978). Holever, theìr roles
remain to be debermined.
Aìthough v;rrious st¡bstances have been expìored as possible candidates
for the non-adrenergic non-cholinergÍc neurotransmitter in the tNS, none
of the following are like'ly: catecholamines, cyc'lic-AMP, histam'ine (llA)'
5-hydroxytryptamine (5HT) , vasoactive 'intestinal pepbide (VIP), glyc'ine'
gutamic acid, gamma-aminobu'l-yr"ic acid (GABA), prostaglandirts, bradykinin
(BK) and substance P (see Burnstock, !975 and Chapter I).
Acc<¡rdjng to Burnstock (1975), onìy ATP conles nearest to fulfilling
the criteria for this transm'itter. A neurotransmitter role for ATP was
earlier suggested at central and peripheral endings of sensory fibres
(Hol ton and Hol ton , 1954) , but never confi rmed. Recentìy adenosi ne-5' -
triphosphate (ATP) has been ìmplicated 'in neuronal activity, working
through mechanisms other than 'its established bìoenenget,jc actions.
Nucl eoti des are founrl i n al I subcel I ul ar f racti ons of manlmal i an central
neurones (Abood et aL.., 1968). Furthermore, adenine nucleotides are
rel eased af ter el ectri cal st'imul ati otr of superf used bra'in ti ssue (pul I and
McIlwain, L972) antl from synaptosornes prepared from cerebral cortical
tissues following e'lecirìcal or potassium stimulation (Kuroda. and McIìwa'in,
lg74). But no pronounced actions of iontophoretically applieC ATP are
evident at central neurones (Curt'i s et aL., I96L; ['.rnievic and Phillis,
i963) and it is they'efore far from certrlin that ATP is the transmit'ter
for any of these neuroÍìes" Indeed it is known that ATP forms part of the

88
noradrenaline (NA) chromogranìn complex for NA storage in the adrenal
medulla, and synrpathetic nerve termina'lS, and jt'is also present with
acetycholine where jt is thought to be involved in maintaining intracellular
osmotic pressure (Ribeiro, 1978).
0n the other hand, adenine nucleotides do have postjunctional effects
at smooth muscle of the mammalian gastrointestinal trace (Burnstock, L97?,
1978) as wel'l as presynaptic actions at the motor axon term'inal (Gìnsborg
and H'irst, 1972; Ribjero and t'lal ker, 1975) and at both adrenergìc
(Hedqvì st anrl Fredhcltn, I976; Enero and Sai dman, 7977) and chol'inerg'ic
(Vizi ancl Knoll, 1976) nerve terminals localised in the gastrointestinal
wall.
The adenine nucleot'ides, more specifically ATP, have been explored as
the transm'itten of the intrinsic non-adrenergìc, non-cho'liner g'ic inhjbjtot"y
nerves of the gastrointest'inal vralì (Furness, 1969a, b; Bianchi et aL."
1968). ATp when appi'iecl to gastrointes'üinal preparat'iotts acts directìy
upon the smooth muscle (tsurnstock et aL.,1970) to cause hyperpolarization
of the muscle cells (Axelsson and Holmberg, 1969; Malchikova and Poskonova'
lglt). These actions of ATP are characteristic of the transmìtter released
by the non-adrenergi c non-chol i nerg'ic 'inhi b'itory nerves (see Burnstock ,
LgTZ). Adenosine diphosphate (ADP) and adenosjne also have similar actions,
but ATP is the most Potent.
The in'itial evidence for ATP or a related nucleotide as a poss'ib'le
inhjbitory neurotrans¡nitter came from the studìes by Burnstock et aL. (1970)
and Satchell and Burnstock (1971) who observed elevated nucleoside levels
in the venous efflux during stimulation of non-adrenerg'ic non-chcììnergic
inhibitory fibres to the stomach in both toads and guinea-pigs. Mone
direct evidence for the release of ATP'from these nerves conles from a

B9
study by Su et aL. (I97L), where chemical and eiectrical stimulation of
enteric neurones caused a marked increased in the release of (3H) trom
taenia coli jncubated with (3tl) u¿.nosine, which v¡as blocked by TTX but
not by gaunethidjne. However the ar-rthors po'int out that they could not
be certain whether the neurones that release [3H] u¿enos'ine were non-
adrenergic non-cho1 inergic inhibitory neurones.
In contrast to its inhibitory action, ATP also has direct exc'ítatory
actions on the smooth muscle of a number of mammalian organs (Burnstock,
Lg72)'including the gu'inea-pig small intestine. These exc'itatory effects
were gener'aììy mot e prevalent in preparations with jo'¡ tone (e.g. rabbit
stomach and gu'inea-pig iìeum) and tìssues often showed biphasic effects
(brìef contract'ion fol lcwed by relaxation) w'ith appì ied ATP. In the guinea-
pig co'lon the inhibitory mode predominates, whereas the ileum responds with
relaxation ai lower concentrations but with contraction at hjgher concent-
raticlns of ATF. Evidence has been presented that adenosirre 5'-triphosphate
(ATP) is an excitatory transmjtter in the guinea-píg (Burnsiock ei aL.,
1978), rabbit (Down'ie and Dean, 1977) and rat bladder (Brorrln et aL., 1979).
l-towever at present it is unclear whether the excitatory action of ATP in
the intestine is in anyway related to a separate class of non-cholÍnergic
exc'it.atory "purìnergic" neurones, since many of the effects are depressed
by prostaglandin synthesis inhibitors (Brown and Burnstock. 1981). However
a dual actjon for ATP on smooth muscle would not be an uncornmon phenomencn,
being established as normal functional modes for both acetylcholine (ACh)
and noradrenal i ne (NA) (Ah'lquì st, 1948) . The exi stence c¡f two di st j nct
popu'lations of postsynaptic receptors for ATP has been investigated 'in
the gujnea-pig urìnary bladder and taenai coji (Burnstock et aL.,1972a, b),
yet no conclus'!ons could be drawn from thejr observal"ions.
The abi'l'ity of ATP to desensjtize tissue (tachyphylaxis) (l-lolman and

90
Hughes, 1965) has been used to determine the involvement of ATP jn the
non-chol'inergic excitatory and non-choìinergic non-adt^energic inhjbitory
responses of nr¡merous preparations (Burnstock, 1972, 1978; Ohga and
Taneike, 1977; Bartlett et aL., 1979), but w'ith considerable disparity in
the results amongst. studies using similar or different t.jssues. In addition
to the fajlure of some smcoth muscle to deve'lop tachyphyìaxìs to ATP, the
phenomenon of desensitization does not allow researchers to distinguish
specific sjtes of drug actìons. Specific antagonists for the respectìve
postsynapt'ic actions of ATP are presently not avaÍlable, although several
compounds have been, useci as such agents. These include high concentratjons
of quinidine (Burnstock et aL.,1970), imidazoline (Rikintaru et aL.,1971b;
Tomita and Watanabe, L973; Satchell et aL., 1973), methylxanthines (Ally
and Nakatsu, L976;0klvuasaba et aL., 1977) and the isatogens (see Burnstock,
1972), but only the latter is jn any way specific, and at that, non-
competìt'ive. Indeed the methylxanthines are now considered to be only
relatively spec'ific against aderros'ine actjons'in reduc'ing transmitter
output (BurnsLcck, i979) . The non-spec'if ic and wide ranging pharmacol ogica'l
actions together with the considerable loss of smooth nluscle tone that
vitiates the use of these substances, has not allowed any reliable
determination of the nature cf the sites at which ATP acts.
The lack of a specìfic ATP receptor antagon'ist has prevented any
definjtive assessment of the possible role of purine nucleot'ides as
transmitters in a number of tissues proposed to receive'purinergìc'nerve
i nnervati on .
Recent'ly, Bohme et aL. (I972) described a class of anthraquinone-
sulphonjc acid derivatives which can be lookecl upon as ATP analogs,
where the l-amjno and 3'sulphoníc groups are close in pos'ition to the
y-phosphate and 6-amino of the purìne jn the an'bi-conformation. These
Fig. I
ONH 2
o NH
so ONa2 so ONa
2
{ //
NH
N
ct
N _\
NH
SO ONa2
N
Cibacron bl¡¡e 3GA

91
compounds have a specific affinìty for the "dinucleotjde fold", and th'is
capacity to act as an affin'ity 'label'for ATP pronpted the investigation
of one of these derivatives (CA3G-A) Reactive Blue 2 (C.I. 6I2LL) (F'ig. 1)
as an antagonist of ATP actions in the gu'inea-pig intestìne. Furthermore
an investigatìon of the involvement of ATP in the non-cholìnergic non-
adrenergic relaxation of the cj'istal colon and in the non-cholìnergic
excitation of the ileum was carried out.
Methods
Guinea-pìgs of either sex weighing 200-400 grams v¡ere stunned by a
blow to the head and bled. Portions of the proximal part of the ileum or
distal colon were quickly removed and placed into warmed modified Krebs
solution (37') (see Chapter Il,rnethods) gassed with a mixture of 95% 0,
and 5% COr. All subsequent treatments were carried out in a similar
sol uti on .
Segments 4 cm in length' were carefully denervated of their
mesenteric attachments anC prepared as a convent'ional isolated smooth
muscl e preparati on. Indi vi dual ti ssues v'/ere at'ranged i nsi de a g'lass
organ bath (10 m'l) wjth one end attached t,o the bottom of the bath and
the other end attached by a cotton thread to a rotary or force transducer.
Changes in the lengbh of the intestinal segments were recorded usìng a
Grass polygraph (Model 5D). Indjvjdual tissues lvere allowed to equilibrate
for 40-60 m'in prior to the addition of the test agonìst. Where an
antagon'ist was added during the course of the experiment an approprìate
equiìibration tjrne was alloled before acidittg the agonist.

92
ELec trícaL StírruLation
Transmural stimulat'icn of the intrinsic tlerves was by a ring electrode
consist1ng of ivro platìnum wires 3 mm apart, posìtioned so that the t'issue
with.in the ring was in contact with the exposed pìatinum wires. Pulses
were delivered from a Grass 548 stimulator using tnaximaj i'oltage. The
parameters for the different nerve st'imulations are as fojlows:
Non-adrenerg'ic inhibitory nerves: 2'I0 llz, 0.5 msec durat'ion, 10 sec
stimulation period. Non-cho'ì'inergic excitatory nerves: 10 - 50 l-lz, 0.2 msec
duration, 10 sec stimulation period. Chol'inerg'ic excitatory nerves (twitch):
0.1 H2,0.1 msec durat'ion, of varying train length. Where it t^¡as necessary
to elimina'le cholinerg'ic events, atropine 2 x 10-7 g/ml was kept in the
bath to block muscarjnÍc receptors.
Dtwgs
The vol ¡me of drug added was never more than I% sf the total bath
volume. Drugs used [Vere: adenosine; adenosine 5'-tt'iphosphate, am'ino-
phytljne, atrop'ine su'lphate, braCyk'inin, cibacron blue 3G-A' gamma-
aminobutyric acicl (GABA), histamine, noradrenaline bitatrate, papavefline,
tetrodotoxjn (TTX).
Ana,Lysis
Responses are expressed as nrm response, or 7i of rnaxinlum control
response. Statist'ical signif icance was establ'ished using Student's t-test
for paired and unpaìred samples. Evaluation of dose response effects was
by a Clark Plot (Clark, 1928). L'ines of best fit were estimated by

93
regressjon analysis (ìeast squares rnethoct). Regression line slopes of the
clark pìot were compared using the method descrjbed in chapter II.
ResuLts
Responses of the DisbaL CoLon to ATP
Since it was desjred to investigate the relaxatiotl pey se, the
experiments clescribed here were carried out wìth atr opjne (Z x lO-7 g/ml )
present in the bathing soluiion throughout. ATP produced concentration*
dependent relaxations of isolated segments of distal colon" The relaxation
was max'imum vlithin 20 secs and the response was sustained for the duration
of ATP conta.ct (¡O sec)" tlhere Al'P was left in the bathing medium fon
ìonger than 30 sec the tone of the tissue usually returned to its rest'ing
value over a per''iod of B-10 min. A'lP induced rejaxat'ions were unaltered
in the presence of tetrodotoxin (TTX) L0-7 g/m1. The threshold concentration
for the ATP induced relaxation v,ras about I0'7 g/ml and max'imum relaxat.ion
occurred with 10-3 g/m'l (Fig. 2a).
A number of preparations showed markcd tachyphylaxis to ATP (Uifrictr
et aL., 1954; Kim et ai.., 1968; Burnstock et aL., 1970). In this study,
attempts to desensitize the t'issue to ATP failed to show any tachyphyìaxis
which allowed the use c'f cunrulatjve dose-response curves for the actions
of ATP in the distal colon. This experimental design shortened the'
exposure time for the colon segments to the antagonist CB3G-4, thereby
minÍmising the possÍbì'lity of loss of tone, that has plagued the use of
other antagonists in such studies.

FIGURE 2.
(a) The effect of CB3G-A on the concentration response curves
for the relaxation elicited by ATP in the guinea-pig distal
colon. (Ð responses in normal solution, À responses to ATP
20 min after the addition of CB3G-A 7.2 x 10-5 g/m'l to the
bathing solution, ü in the presence of CB3G-A 1.5 x 10
g/nl. Each point is the mean and standard error crf
responses in 4-8 preparations. Atropine 2 x I0-7 g/nl
was presen+" in the bath throughout the experiment.
-5
(b) Clark plot of the ATP concentration nesponse curve
Ocontrol responses, A responses in the presence of
CB3G-A L"2 x 10-5 g/ml ¡ 1".5 x 10-5 g/ml .
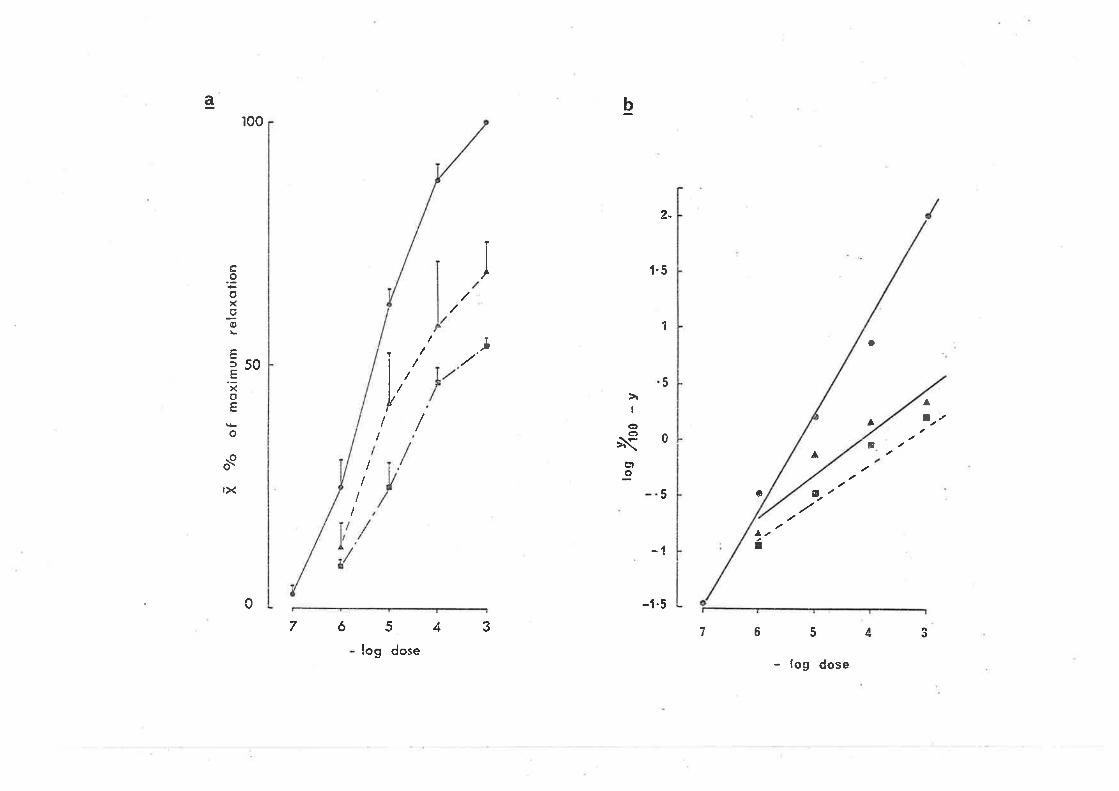
Þer00
2,
1.5
5
-,|
-f .5
50
c.9oxq(¡)
ÊI
E'io
o
-o
rx
u
1
5
0
.T o
iI
A
II
ôoNo'I
A!
IA
II
II
76 5
log dose
43
o
7 6
9t'
5 4
0
J
- log dose
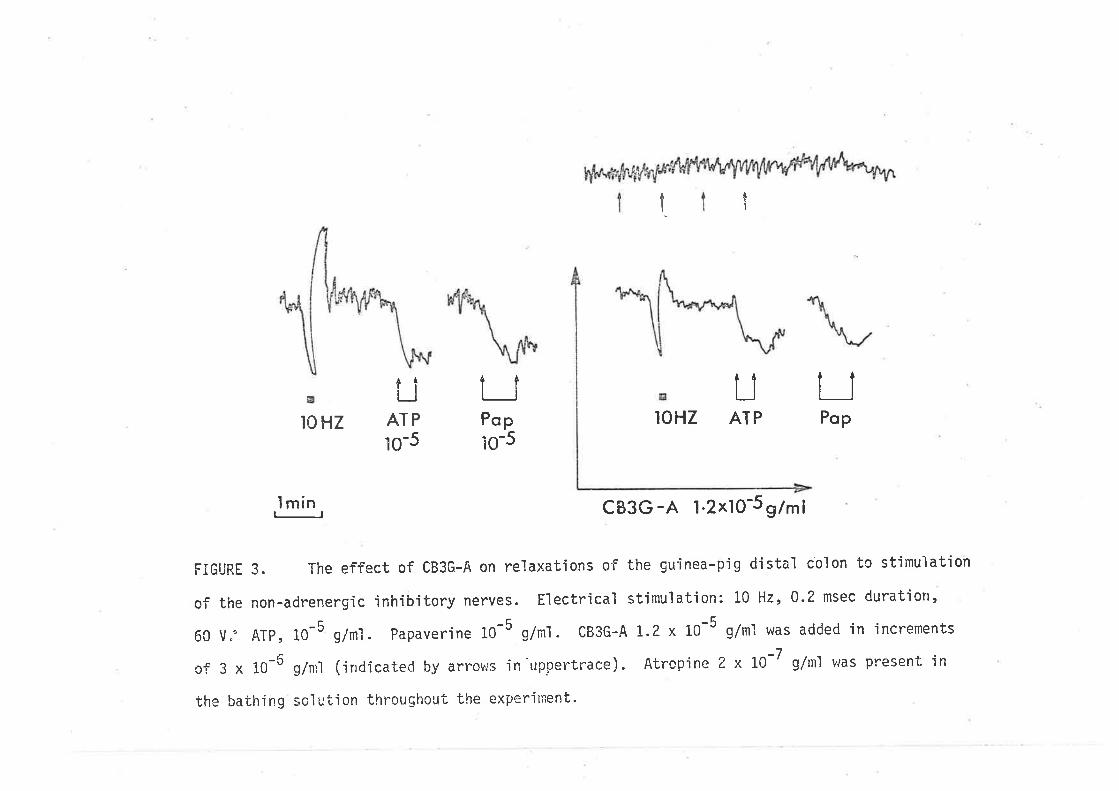
1
¡ì
iiloHz ATP
l0-5
L-JPoP
¡o-5
IJ I-JIOHZ AT P Pop
CB3G_A t.2xt0_5s/mlI min
FIGURE 3. The effect of CB3G-A on relaxations of t,he Euinea-pig distal colon to stimulation
of the non-adrenerEic inhibitory nerves. Electrica.l stimulation: 10 Hz, 0-? msec duration,
6c V.' ATp, 10-5 g/ml . papaverine 10-5 -olml . CB3G-A 1.2 x 10-5 g/ml was added in increments
of 3 x 10-5 g/nrl (indícated by ar rcvrs in'uppertrace). Atrcpì ne Z x 10-7 g/rnl vras present in
the bathing solut.ion throughoui the experiment'

94
In general there'is difficulty assocìa'bed w'ith investigat'ion of
distai colon relaxat'ion, since most putative antagonists of the relaxation
cause considerable loss of snrooth muscle tone (McFarland ei a1.., L97I;
Spedding et aL.,1975). A rapid drop in 'tone' during the administraticn
of CB3G-A up to 5 x 10-5 g/m1 could be avoided by raising the concentration
of CB3G-A in increntents of 3 x 10-6 g/ml added every 1-5 min, depending on
the i ndi v'idual preparat'ion . Thi s avoi ded the necessi ty to 're-tone ' the
tissue and thereby'l'imited the number of drugs present in the bath'ing
solutjon. An equilibration period up to 10 mjn was usually aìlowed jn
the presence of CB3G-A before cornmencjng t.he addition of ATP.
The effect of two concentrations (1.2 and 1.5 x 10-5 S/ml) of CB3G-A
on the cumulative dose-response curve for ATP are shov¡n in F'ig. 2a. Both
concentrations of the antagonìst brought about a non-parallel shift of the
curve to the right, and ciepression of the maximum response to ATP indicatìng
the antagonism by CB3G-A to be of the non-competitive type. The relaxatjon
elicited by ATP in ihe presence of atropine (2 x LO-7 g/m1) was sjmilar
to that for transmural stimulat'ion at a low frequency (10 Hz) (Fig.3)
which according to a number of studies (Furness and Costa, 7972; Bennett
et aL., 1966; Burnstock et aL., 1970) is an effective stimulus for the
enteric i nhi b j tory nerves . A'l thorrgh CB3G-A !.2 x 10-5 g/ml reduced by
40% the ATP (tO-S g/ml) ìnduced relaxation, it was withcuI effect against
the relaxation due to electrical stimulation. or the smooth muscle relaxantç
papaverine (10-" g/ml). The so called 'rebouncl' contraction whìch 'is
observed following cessation of electrical stìmulation (Burnstock et aL.,
1970) was also reduced by more than 50%.
At the two concentratìons 1.2 and 1"5 x 10-5 g/ml of CB3G-A whjch
caused significant shjfts in the ATP rlose-response curves, the voltage-
response curve for the electrically jnduceci relaxation vlas not s'ignificantìy
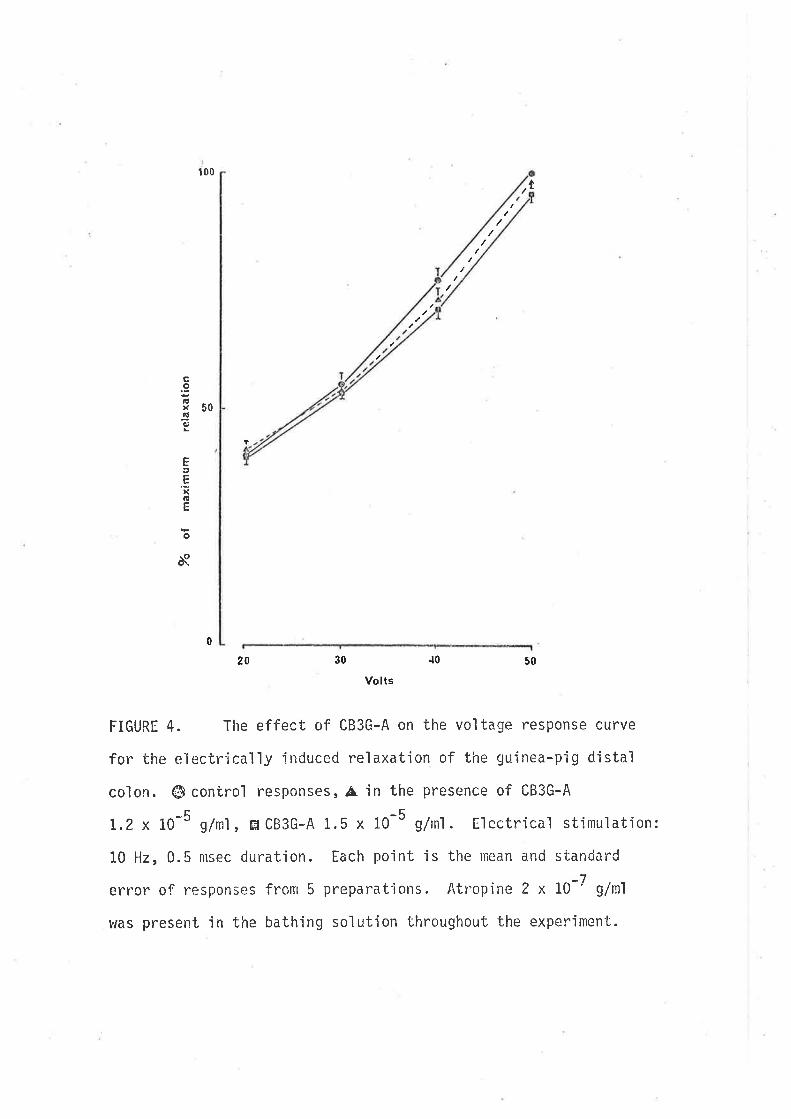
1 00
c.9
9sosÁiL
E5ExtlE
o
èe
1
0
20 30 50
Volts
FIGURE 4. The effect of CB3G-A on the voltage response curve
for the electricaì'ly induced reìaxation of the guirlea-pig dìstai
colon. @ control responses, A in the presence of CB3G-A
-Ã -6L.2 x L0-' g/m'l , H CB3G-A 1.5 x 10 " g/ml . Electrical stimulation:
10 l-12,0.5 msec duration. Each point is tlre mean and standard
error of responses froril 5 preparatjons. Atropine 2 x 10-7 g/ml
r^¡as present in the bathing so'lutíon throughout the experiment.
.t0
t
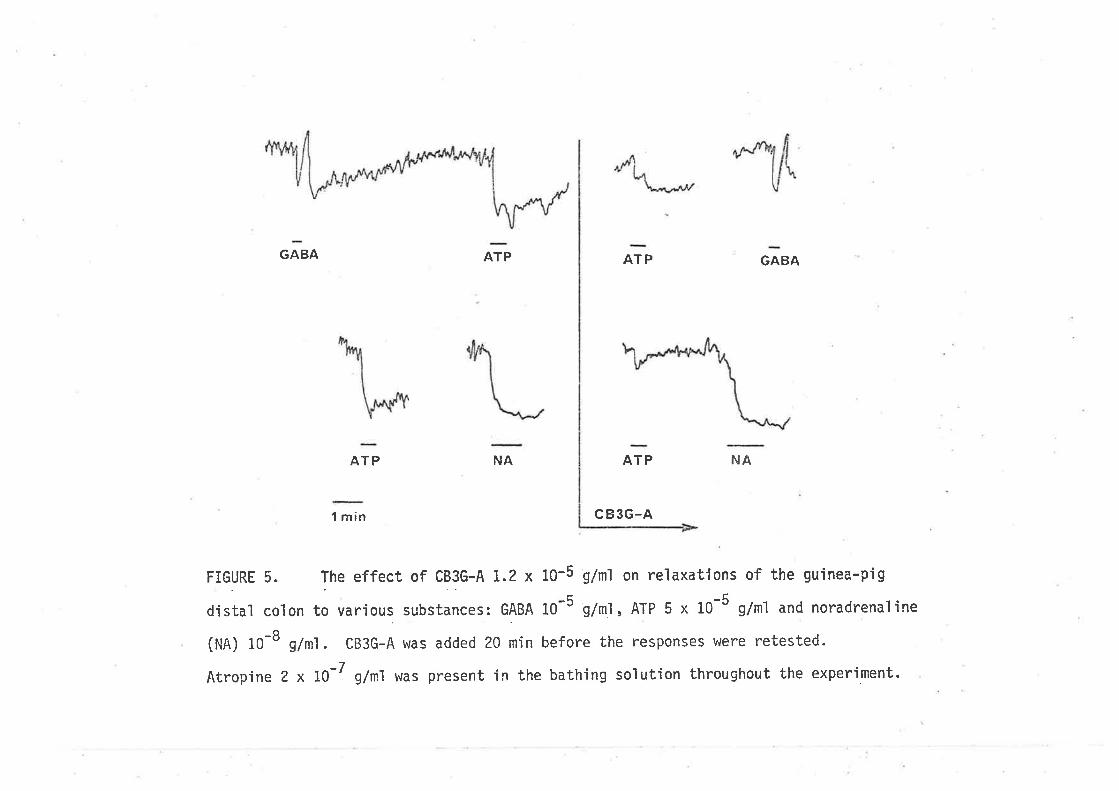
GABA ATP ATP GABA
ATP NA ATP NA
l rnin CB3G-A
FIGURE 5. The effect of CB3G-A 1.t * 1g-5 g/m'l on relaxations of the guinea-pig
distaì colon to various substances: GABA 10'5 g/mì, ATP 5 x 10-5 g/m'l and noradrenaline
(NA) 10-8 g/ml. CB3G-A was added 20 min before the responses r¡,ere retested.
Atropine 2 x L0'7 g/n1 was present in the bathing solution throughout the experiment.
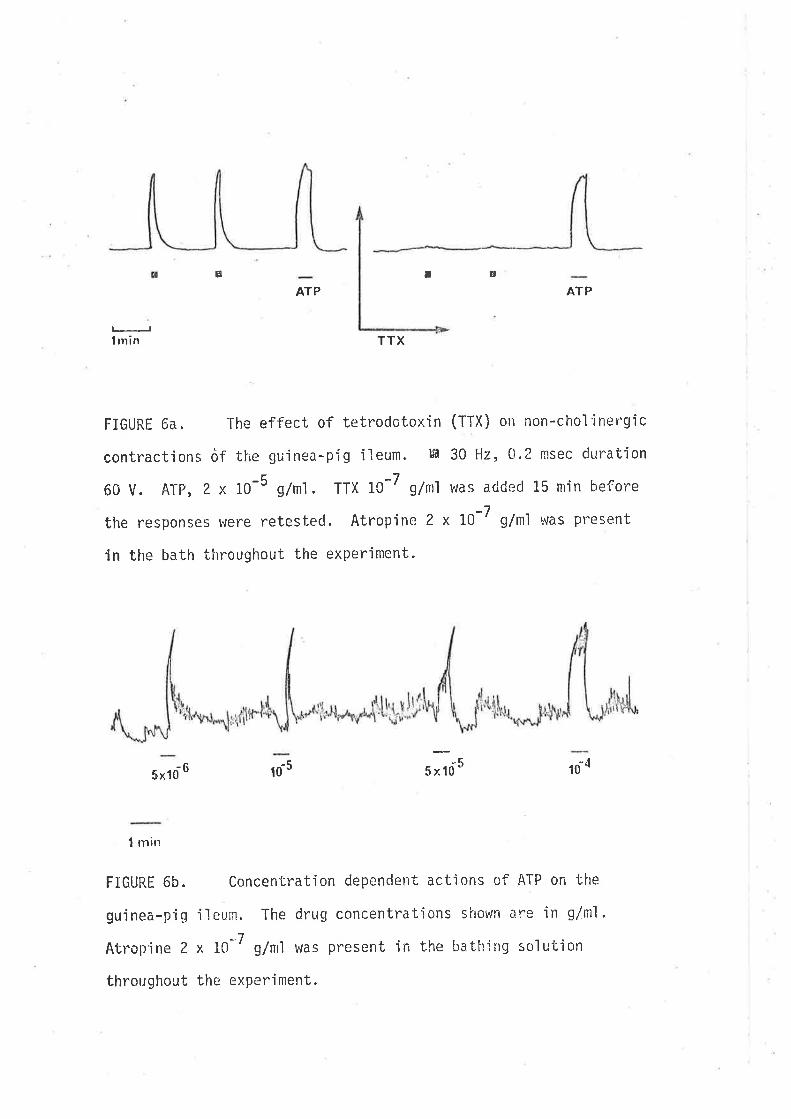
E , E
ATP ATP
10'4
lmin TTX
FIGURE 6a. The effect of tetrodotoxin (TTX) olr non-cholinergic
contractions óf the gu'inea-pig ileum. m 30 Hz, 0.2 msec duratíon
60 v. ATp, z x 10-5 g/ml. TTX 10-7 g/ml was added 15 min hefore
the respot'ìSes t^Jere retestecl. Atropi ne 2. x 10-7 g/rnl ttas present
in the bath throughout the experiment-
5x10 6 -c,10" u"tOu
1 nrin
FIGUP.E 6b. Concentration dependetrt actions of ATP on the
guinea-pig 'iìeunr. The drug concentratjons sirot¡lrì are in g/m] .
Atropine 2 x 1C-7 g/m'l was present in the bathjng solution
throughout the experinlent.
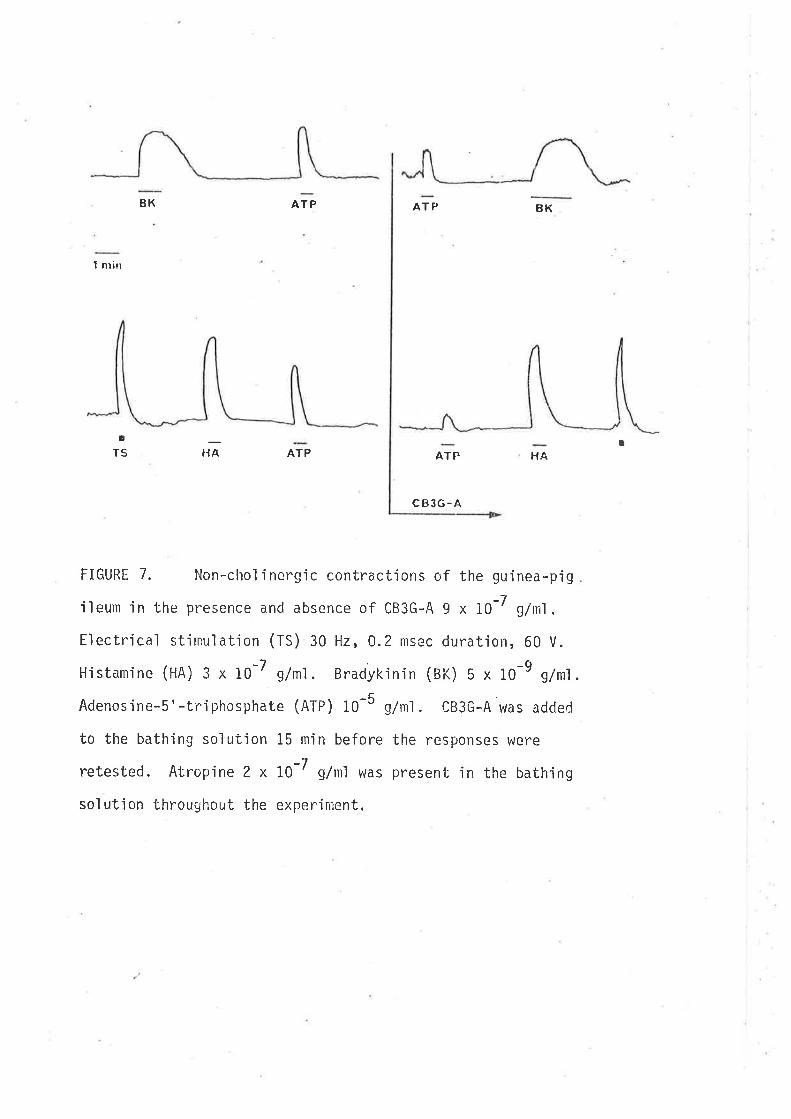
BK ATP
ATP
ATP
ATP
C B3G-A
I nlin
BK
HAHA¡
TSI
FIGURË 7, Non-cholinerg'ic contractions of the guinea-pig
ileum in the presence and absence of CB3G-A 9 x 10-7 g/nl,
Eìectrjcal stirnul ation (TS) 30 Hz , 0.2 msec duration, 60 V.
Histanrine (HR) 3 x I0-7 g/n1. Bradykinín (BK) 5 x 10-9 g/nrt.
Adenosine-5'-trjphosphate (ATP) 10-5 g/mì. CB3G-A was added
to the bathing solution 15 min before the responses were
retested. Atropine 2 x 10-7 g/rnl was present in the bathing
solutjon throughout the experiment.

FIGURE 8.
(a) The effect of CB3G-A on the concentration response curve
for the ATP induced contractions of the guinea-pig ileum.
C responses in normal solution, A responses to ATP 20 min
after the addition of CB3G-A 9 x 10-7 g/ml to the bathing
solution. Each po'int is the mean and standard error of
responses in 6-8 preparat'ions. Atropine 2 x 10-7 glml
was present in the bath throughout the experiment.
(b) Clark plot of the ATP concentration response curve
O control respones, A responses in the presence of CB3G-A
-79 x 10 ' glnl.
a
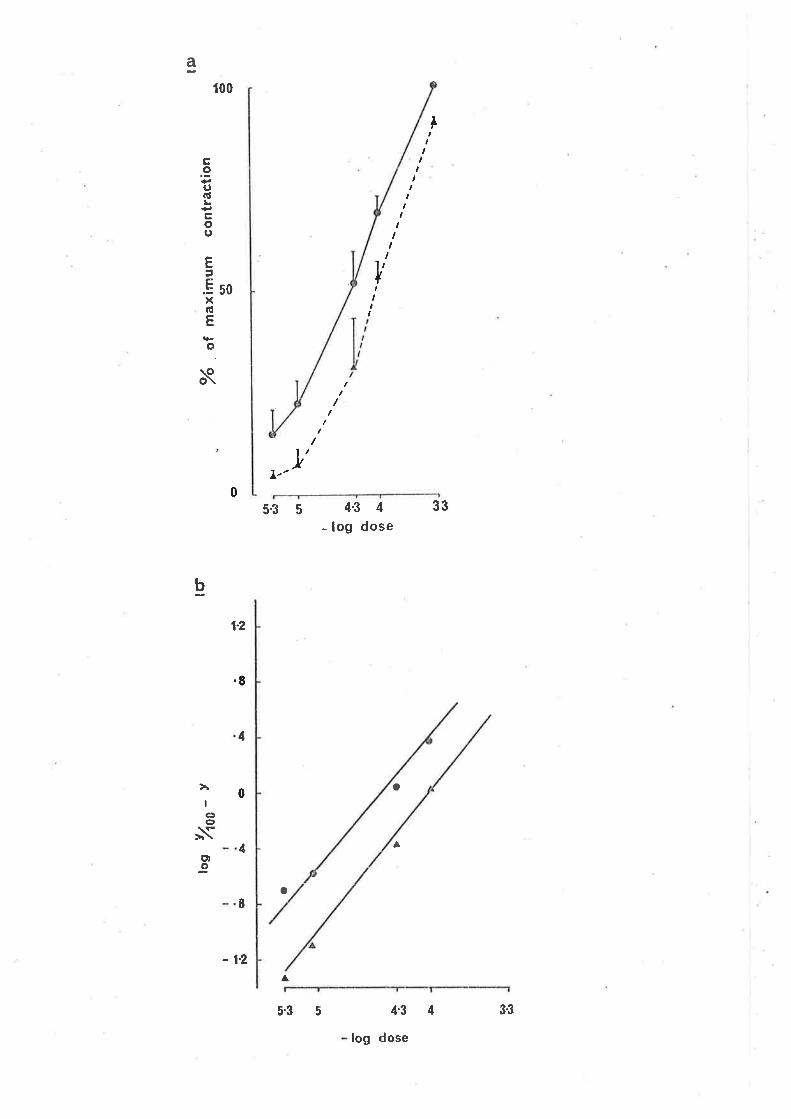
19,
log
hoo
-v o o
t67o
af
max
itnum
con
trac
tion
('l oo o
N€
Àco
I ño
õ
(t C¡Ð (rr
ctst
(rr
d¡ (rt
I õ (oè d)
0- 3o o
ó- (a o- 3+ oq¡
5
---
--¡-
.
@ 9¡
cp C^)

e5
affected (FiS. 4). Noradrenaline (NA) (receptors for which are located to
the longitudìna'T muscle of the colon) and GABA-induced relaxations by
stimulatìon of intrinsic 'inhibitory neurones (Kranti s e'ü aL., 1980) of
the distal colon were unaffected by CB3G-4, !.2 x 10-5 g/ml (Fig. 5).
Responses of the ILeun to AIP
Responses of the'ileum to ATP, in the presence or absence of atropìne
-12 x 10-' g/mr,, were found to vary Cepending on the'tone'of the preparat'ion
(see Cocks and Burnstock, !979; Campbelì, i966; Burnstock et aL., 1970).
In ileal segments of 'medjum tone','in wlrich either relaxation or contraction
could be easiìy observed, the type of response elicjted by ATP varied with
the concentration used (F'!g. 6a). At low concentrations on'ly relaxations
occumed; biphas'ic responses were el icited at concentrations around
5 x 10-5 g/n1 , whilst only contractions were e,rident at h'igher concentratjorrs
of ATP. These responses were the result of ATP actions dlrectly upon the
muscle since the responses were unaltered in the presetlce of TTX 10-7 g/ml
(Fis. 6b).
In the presence of atrcpine (2 x ß-7 g/ml), 'i'lea] segments of 'lovl
tone' were contracted by ATP (5 y. 1rl-6 - 5 x. 1.0-4 g/ml ). At a concentration
lower than tl,¡ab used to antagcn'ise relaxatiû$s ùf the distal colon to A'lP'
CB3G-A 9 x 10-7 g/ml recluceci the contraction of the ileum to ATP but did
not significantly affect the response to transniural st'inrilation (30 llz)
or to the action of e'ither brad.ykinin (BK) 5 x 1.0-9 g/ml or histam'ine (HA)
3 x 10-7 g/nr1 both of which are proposed tc contrac'b the ileal longitudinal
nruscle d'irectly (Danìe'l , 1968; Bolton , I979) (Fig. 7). The concentrat.jon-
response curve for A.TP (Fig.Ba) but not that for BK or HA (Figs. 9a, b)
was shifted to the righb. A'lthough this acûion of CB3G-A was specifìc

FIGURE 9a and b.
The effect of CB3G-A on the concentration response curves
for contractions of the guinea-pig ileum by histamine (HA)
(a) and bradykinin (BK) (b). Q responses in normal solution,
Aresponses 20 min after the addition of CB3G-A 9 x 10-7 g/m1
to the bathing solution. Each point is the mean and standard
error of responses in 6 preparations.
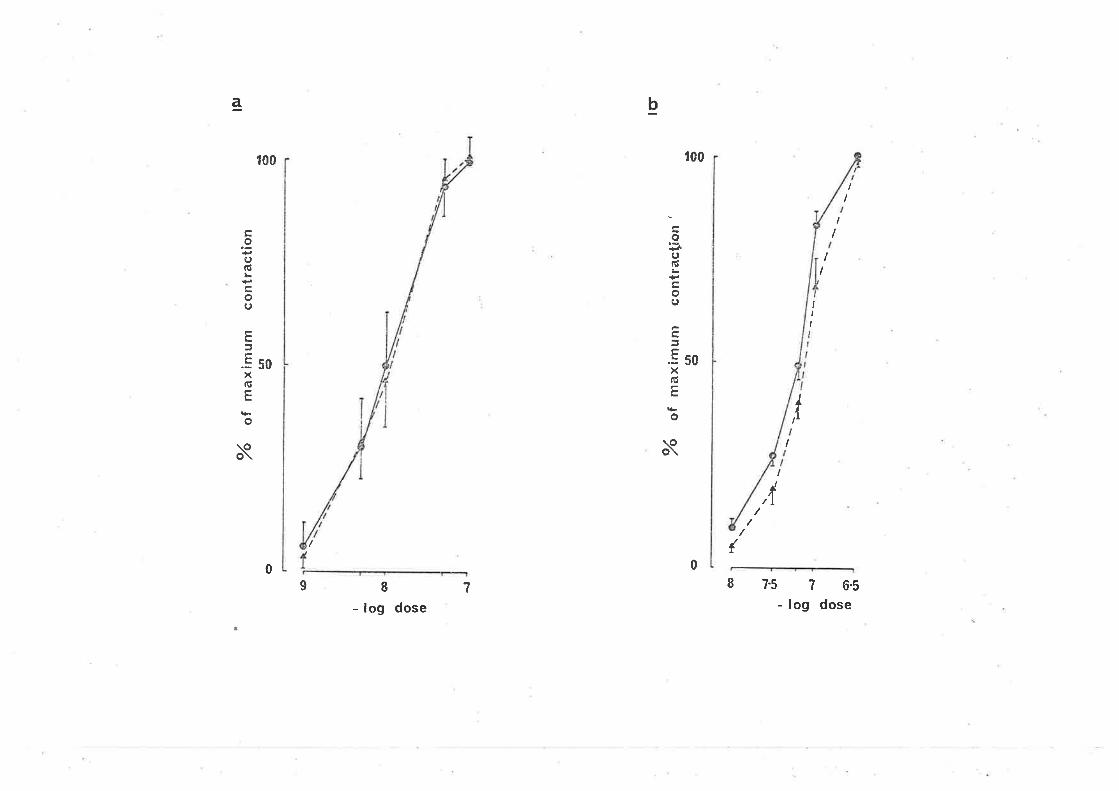
þIA
'
lcF
o%
of
max
irnum
con
trac
tion
('I <Ð
o o
tê
I ã rOco
o. o ø o
\¡ ð
o
O./ 7o
of
max
imum
con
trac
tion
'Ç
t êo o
\
+ glI ö GI
{o. o ,a oq
C'I
+-
Ê>
-
100
50
.9u(ltL
Éou
Eã.Exl!Ë
o
ñ
0
t0 15 3t
HZ
FIGURE 10. The frequency response curve for the non-cholinergÍc
excitatory nerves in the presellce and absence of cB3G-4.
Electrical stimulationz 0.? msec duration, 60 V. @ control responses'
D in the presence of CB3G-A 9 x 10-7 9/ml . Ëach poirrt 'is the mean
and standard error of responses frorn 6-10 preparations. Atropine
Z x L0-7 g/ml was in the bathing soìution throughout the experiment.
1lAden, Aden
ì min
AminoPhYlline CB3G-A
FIGURE 11. The affect am1nophylline 7 x I0-9 g/rnl and
CB3G-A 9 x 10-7 g/ml on the inhibjt1on of the cholinergic
twi tch response el i ci ted by el ectri cal st'imtll at j on of the
gujnea-pig ì'lèum. Electrical stjmulation:0.1 Hz, l msec
duration, supramaximal voltage. Adenos'ine was addefl at a
final concentration of 10-6 g/ml.
t__JGABAto-5
GABA
I minL---.1
C B3G-A
ó " l0-ó g/ml
FIGURE 12, The effect of ÇB3G-A on the bjphas'ic response
of the gu'inea-pig ì'leum to GABA. CB3G-A 6 x 10-6 g/ml was
added 15 nrin before ret,esting the response to GABA 10-5 g/ml'

96
for ATP induced responses, the non-paraìle1 shift in the C'lark plot (F'ig.
Bb) (signìficant, d.f. = $, P>0.C5) ind'icates this antagonism to be of the
non-competìtive type" At concentrations greater than 9 x 10-7 g/ml the
action of CB3G-A became non-spec'if ic. The lack of antagon'ism by CB3G-A
-79 x 10-' g/ml of contracticns 'induced by transmural stimulatìon of the non-
cholÍnergic non-adrenergic exc'itatory nerves, suggests that ATP is not
involveC in the response mediated by these nerves. This is further
supported by the lack of effect of CB3G-A on the frequency-response curve
for the relaxation induced by electrical stimulation of these nerves (FiS.
10) despite the fact that the dose-response curve for ATP js shifted to
the right by CB3G-A ai the same concentration.
ATP and rel ated substances mod j fy adrenerg'ic and chol'inergic neuro-
transmission by inhib'itìng release of the respective neurotransmitter
(Burnstock, t972; Bartlett et aL., I979; Hedqv'ist and Fredhoim, 1976;
Sawynok and Jhamandas, 1976) w-ith an order of potency of, adenosjne>AMP>
ADP>ATP.
In order to determine whether CB3G-A is an antagonist of these
prejunct'ional purìne actjons, the effect of CB3G-A 9 x 10-7 g/m'l was
compared w'ith that of aminophyìline 7 x 10-5 g/ml (a spec'ific antagonjst of
adenosine, Bartlett. eb aL., 1979) in counteracting the effects of adenosine
upon the electricaìly induced cholinergic twitch contractions in the ileum
(Fig. 11). Adenosine 5 x 10-7 g/ml reversibly inhibited the transmural'ly
stimulated contractions, aRd the 'inhibition by adenosine was prevented by
ami nophyl I 'ine. llowever, 'in the presence of CB3G-A the adenos i ne- i nðuced
reduction in the responses to ùransmural stinlulation was barely altered,
and CB3G-A had no effect on the control responses alone. Followìng washcuts
of amjnophyìline and Cll3G.-A, the'ileal contractions quickly recovered to
thei r control amp'l i tude .

97
GABA has l¡een shown to stimu'late the intrinsic cholinerg'ic excitatory
and ncin-chof inergic non-adrenergìc inhib'itory neurones of the ileum (Krantìs
et aL., 1980). Fjg. 12 shows that this action of GABA (10-5 g/nll) on a
segment of guìnea-pig ìleum was unaltered b¡r CB3G-A (6 x 10-6 g/ml).
DLscussíon
DistaL CoLon
The results of the present study have demonstrated inhibitory actìons
of ATP on i,he guin.u-p'ig clistal colon, consjstent with repor^ts that this
compound relaxes snlooth muscle jn a trumber of preparations (Burnstock, 1972,
1975). These relaxations were not due to ATP, or a related nucleotìde,
act'ing indirectly througlr adrenergic or non-chol inerg'ic non-adrenergÌc
inhibitory nerves, since TTX aboljshed the inhibitory responses to jnt.ra-
mural nerve stímulatjon and CB3G-A antagonjsed ATP actions v¡'ithout affectittg
noradrena I j ne (NA) j nduced rel axati ons . A'l though there was para'l I el j srn
between the effects of, exogenously app'lied ATP and stìmulation of non-
adrenergic inhibitory neurones jn the distal colon, onìy the A-iP induced
rel axati ons \,{ere antaç¡on'ised by CB-?G-4.
A variety of conrpounds have been shov¡n to antagonjse the actions <lf
ATP and rela'bed nucleotides on gut muscle and other tissues, jncludìng
qu'inid'ine (Burnstock ei a7., 1970), imidazolines (Satchell et aL., 7973;
R'ikimaru el; ci/,., 7971), 2-2'-pyridy]isatgen (Spedcjing et aL., 1975;
Rattan and Goyaì, i9B0) and methy'lxanthines (0kwuasaba eb aL.,1977).
However, at the concentrations used, all of the putat'ive ATP antagon'is+;s so far
tested have been found to be non-specific (Burnstock, L975; Speddìng et a,7.,r

98
1975', Rikinlaru et aL., IgTL; Dean and Downie, 1978; Nickerson' 1970). The
effects of the anthraqu'inone sulphonic acid derivat'ive CB3G-A indjcate thai
this substance (within a narrovr concentratiorr range), 'is a spec'ific, altirough
non-competitìve, antagonist ofATP actjons ìn. the gu'inea-píg large intestine.
In addjtion to its abi'lìty to relax certajnsmoothmuscle, ATP has been
shown to cause the contraction of smooth muscle in a number of ntammalian
tissue (BurnsLock et aL., 1972; Downie and Dean, 1977; Brown et aL.' !979;
Br¡rnstock et aL. , 1.978; Mori tokí et aL., 1979; Westfal 1 et aL", 1978) . The
abiljty of ATP to reversibly contract ileal preparations of low'tone' in
this study, jn the presence of atropine and TTX, suggests a direct acticn of
ATp upon the muscle through a non-cho'l'inergic mechanism, and not through
chol'inergic neurdnes as is the case'in the canine intestine (i'{orthway and
Burks, 1980) and rat ileunr (Sakai et aL." 1979). S'ince the ìleum is supp'lied
by non-chof inerg'ic non-adrenerg'ic inhib jtorV and nort-cho'linergíc exc'iiatcry
nerves (Furness and Costa, 1973), there is the possibility that ATP coujci be
the contractile substance nrerljatìng non-chol'inergic excìtatory transmission.
J- Leurn
Evidence advanced'ny other studies to suggest that A-lP is the trans-
mitter of i;lrese excitatcr¡r nerves jncludes: the blockade of responses to
both ATp anct transmural uerve stimulatÍon by a number of agents used in
the investigation of purinergìc tratlsmissjon (Satchel'l eb aL", 1973;
Burnstock et aL., Ig7?; Weetnlan and Turner, I974) and the paraìlel.
depressìon of both ATP and transmural nerve stjnluiation indttceci responses
by the development cf tachyphylaxis to ATP (Burnst.ock et aL.' I97?-).
Attempts to desensitjze jleal preparat'ions of this study to ATP were wìth
nlixed results ancl therefore it was not possible to draw any conclusions
using th'is nrethod. i-lowever, CB3G-A which specif ical ly anatognised

99
contractions due to ATP was wjthout effect on non-chol'inergic responses
to transmural stimulat'ion, which is not cons'istent with ATP be'ing the
transmitter for excitatory responses within the guinea-oig ileum. Indeed
there'is now good evidence to suggest that the transrnitter for these
nerves is substance P (Furness et aL., 1980).
The specjfìcity of CB3G-A for ATP act'ions,evident from its lack of
ef fect toward contracti ons of the i I eunrinduced by brad.yki ni n (BK) and
histamine (HA), suggests a unique site and mode of act'ion for ATP and
th'is antagonist in the small intestine. In fact a related compoun<l,
Procicn Red l1E3B antagoni ses on'ly contract'i1e responses to ATP (Kennedy
et aL., 1981). A heterogeneous popri'latìon of nucleotìde receptors have
been proposeO tor the mammal ian gut (Burnstoc k et aL., Ig70; Bartl ett e|;
aL." L979) distinguishable by their resultant actions and their differing
affinit'ies for the r¡arious adenine nucleotides. These receptors include
those aìread-v ment'ioned, r,¡hich are more sensitive to adencsine and involved
in prejunctional rnodulation of adrenergic an,j chol inergic nerve activìty,
and those located on the srnooth muscle and characterised by their greater
susceptibì1ìty to stimulation by ATP. In addition to the differentiation
of these receptors'in terms of potency orders for their purine agon'ists,
tlre nrethylxanthines have been shown to be specific antagonists at the
purinergìc receptors (P1) media'l'ing adenosine preiunctional actions
(Burnstock, I97B; Bartl ett et aL., L979). in th'is stud.y, CB3G-A was
without effect upon the chol iner'g'ic nerve rnediated contractions, and cl'id
not prevent the niethylxanthine sensitive adenosjne inhibition of the
cholinergic twitch response, jndicating that ûB3G-A does not antagonise
the P1 receptor or the mode of action of adenosine in the guinea-pig ileum.
AccordingtoBurnstock (1979), Alp acts on smcoth muscle through
spec'ific receptors. However, the effects of ATP at the mammalian smooth

100
muscle are of two types; re'laxatjon and/or contraction (see Burnsiock,1972).
Similar dual effects of ATP on the ileum were observed'in this study. The
ready reversal of the antagonism by CB3G-4, follclwing its washout from the
bathing medium, jndicates that this compound does not exert its effects by
irreversible bind'ing to ATP receptors.
Prostaglandins can exert contractile or relaxant effects, or modulate
the contraction of snlooth muscle (Bolton, 1979) and. recent'ly it has been
found that ,\TP j nteracts vri th prostagl and'ins (Dean and Downi e , 1978;
Burnst.ock et aL., !978; Mor"itoki et aL., 1979). Tl're rebound contr'acl'ions
of smooth nluscle following cessation of electrical stimulatjon of the
intramural non-adrenergic inh'ibibory nerves or appìied ATP may a'lso involve
prostagìandirrs (Burnstock et aL., 1975; Cocks and Burnstock, 19i9). The
f indjngs that C83G-A marked'ly reduced the 'rebound contraction', but not 'uhe
relaxation of the colon foj'lowing electrical stjnrulatjon of the inhibitory
nerves, suggests the action cf CB3G-A may be through its interference wit'h
an ATP stirnulated prostagìand'in format'ion in smooth muscle. Prociott Red
HE3B is raLher spec'ificalìy directed against this effect (Kennedy et aL.,
1981). There js a further possibility, that CB3G-A interferes with the
stimulus response coupling in some otlrer way. It has t¡een shown that
CB3G-A and related conrpounds bincl specificajìy to a nuniber of enzymes
ccrntaìn'ing the d jnucleoticle fold (Bohme et aL., I97?i lrl'ilson , 1976) and
therefore the act'!on of CB3G-A ma.y be w'ith some enzyme involved ìn the
smooth muscle responses to applied ATP.
In conclusjon, CB3G-A (withirì a narrow concentratjon range) appears
to be a useful antagon'ist for ATP actjons'in the guiriea-pig ìarge and
smal I i ntes+.'ine. However i ts usef ul ness f or the i nvesti gatì on of ATP
actions on mammalian smooth muscle may depend upon the t'issue under study,
since Choo (19Si) reports that CB3G-A has djfferent ef,'tects towards ATP

ì:ì
101
actions on the rat and the gu'inea-pig bladder. The pharmacologic ev
presented here ind'icates that ATP has multiple effects on intest'inaj smo
muscle, but'is unlikeìy to be the primary transmitter of either the non-
adrenergic 'inhíbitory or non-choì inergic excitatory nerves of the gu'inea-
pig intestine. Nevertheless, the ability of ATP to mim'ick responses to
stimulation of these nerves'in thìS, and other stucijes, raiSes some
i nteresti ng questi ons as to 'its f unct'ional rol e j n these tì ssues . In vi ew
of the consjderable evidence that ATP'is local'ised at, and released from,
nerve endings, inclucling those of peripheral chol'inergìc and adrenergic
nerves (Bernei s, 1970; Dowdal I et aL., 7974; Su el; aL., I97l; Sì'linsky and
Hubbard " L973; l-ledqvist and Fredholn¡, 1976) where ìt probably reaches
effect'ive contrac,tions (niUiero and l,lal ker, 1973; Ginsborg and Hirst,
L972; Jawynok and Jhamandas, 1976; I^Jakade and Wakade, 1978) it is poss'ibìe
that A-lP rnay have a synergistìc, feedback, regu'latory role in transmitter
release followìng its degradation to AMP or adenosine.

,CHAPTER VIII
GENERAL DISCUSSiON AND CONCLUSION

ñ2
GENERAL DISCUSS ION AND CONCLUSIOI'i
The amino acid .y-aminobutyric acjd (GABA) occurs throughout the
brain ancl Sp.inal cord of vertebrates where it. is recognised as an
inhib'itory neurotransmitter (Curtis and Johnston, 1974) ' There is
also ev.idence that GABA may have a transmitter function'in autonomic
ganglia (Qbata, 1976)" The a'im of thìs thesis vras to exp'ìore t'he
hypothesis that GABA is a neurotransmjtter of enteric neulones'
That GABA may have a treurotransmitter function ìn the gu'inea-pig
intestine appears'likeìy since a number of the necessary crjtetia as
defjned by wernran (1966) and 0rrego (1979) have now i:een satjsfied.
In Chapter II the pharmacoìogy of GABA act'ions in isolated preparat'ions
of the gu1nea-pìg ileum ìs descrjbed. According to the classical crit'eria,
interaction of the suspected neurotransmìtter at postsynaptic sites should
mim'ick the effects of tþe natural neurotransmi tter. ['urthermore '
pharmacoìogical agents which mod'ify the operation of the na'Lural neuro-
tra¡smitter shoulcl simj'lar1y affect the act'ion of the putative neuro-
transmjtter. In the'ileunl, GABA caused stimulat'ion of intrinsic inhibìtory
and exc j tatory rnotor neurones by ì nteractì ons at spec'if j c bi cucul I j ne
sensitive (GABAA) receptors. investìgation of the ionic mechanisms for
GABA induced contracrions shov.r these GABAA receptors are coupled with
chloricle (cl-)'!oncphores. 0ther studies (Bowery et a/.", 1981;0n9, 1981)
shor^l GABA also depresses enteric excìtabor,v neurones throuçth interact'ions
at baclofe¡ sensit'ive GABAB receptcrs. These receptors wh'ich co not
operate through Cl" channels are proposed t'o be 'pre-.junctjonal' and
their actjvation, reduces transmjtter output presunialliy due to an altered1J-
Ca¿* permeability (Llowery et aL., 1981)'

103
A common pharmaco'ìogy is shared between applied ûABA and the endogenous'ly
released GABA as seen with the use of ethylenedjamine (EDA) (Kerr and Ong,
1g81). Bicucull'ine which blocks GABAO med'iated stÍmulation of cholinergìc
neurones, reduced the tDA evokerl cholinerg'ic contract'ion of the guinea-pìg
ileurn. Furtheìn'ìore, applied EDA also caused a depression of the electricaì'ly
evokecl twitch Ì"esponse, presumably by the action of endogencrus GABA (released
by EDA stjmulation of GABA-ergìc neurones, Forster et aL., 1981) a.t GABAT
receptors since this depress'ion was blocked by desens'itìzat'ion of the ileum
to bacl ofen.
The neuronal elements jnvolved in this GABA-ergÍc transntission should
contain the neces,sary precrtrsors and synthetìc enzymes for the synthesis
of the proposed neurotransmìtter (i.e., GABA). In acldition, the putative
neurotransmitter shoujd be present in nerve terminals. There is nov¡
cons'i derabl e bi ochenri ca I ev'idence that GABA and i ts synthes i zi ng enzyme
gìutam.ic acid decarboxylase (GAD) are present in myenterjc neurones of
the guinea-piil (¿essen et aL., 1979), cat (Hosoya et aL.,1981) and human
(Micki ei c-l,, 1981) intestine. However, specifjc localization of these
substances to nerve terminals has not yet been cc¡nfinmed. Such a study
uti j ising electrontnjcroscopy would be particrrlarìy t¡seful '
Accordìng to bJernian (1966), Fagg and Lane (1979) and Qrrego (1979)'
the denronstration of a synaptic release of the puta.tirre transtnitter, in
an amount proprrrtì onaì to the magnì tude of stinrul a t.i on appl ì ed, 'i s of
consiclerable'importance for transmitter identjficaticn. Th'is process
must also show a true caiciurn dependerrcy" In addit'iovt there musU ex'ist
a mechanisn vlhereby the proposed neurotranstnitter is jnactìvated'fo'l'lowing
jts action at i.lre postsyna¡rt'!c site. However, due to the lack of sensitive
assay procedures for a vlicle range of anljno acids irrcluding GAßA, and the
technjcal diffjculties involved ìn their use, many stuclies have used

104
exogenously app'l'ied radiolabelled substances to determine the uptake,
I ocal i zati on and rel ease of pu'bat'ive transm j tter^ s'.tbstances . Where i t i s
established to be a neurotransmitter, GABA is removeC from the synaptic
cleftb:,a high affinity transport mechanism present in both neurones and
glia (Iversen and Kelly, 1975). GABA taken up by th'is transport mechanisrn
is eventual'ly returned to the presynaptic neurotransmjtter poo1. Evidence
for such a transport nlechanjsm in the myenteric p'lexus of the gu'inea-pig
was presented in Chapter III. H'igh affinity [3H]-GABA uptake sites are
present throughor.rt the meshworks of th'is nerve pìexus, however, jn contrast
to the CNS, extrasynaptic uptake (e.g., glia) of [3H]-engn does not appear
to be a major sjte for removal of [3H]-GABA fronr m:¡enteric synapses.
MyenterÍc neurones accumulatìng [3H]-gngn in this way could also be induced
to release the racl'iolabelled amino acid followjng electrical stinlulatìon,
as described jn Chapter V. [3H]-engn is released in a 'transnlitter-l'ike'
manner, being prevented by treatrnent w'ith tetroclotox'in (TTX), neuronal
high affinity uptake'inhjb'itors and calcium free medjunr. Recently,
Taniyama et a7,. (1981) reported comparabìe uptake and rejease of [3H]-
GABA by the cat colon myenteric p'lexus with remarkable coincìdence of the
stimulus fretluency used to elicit maximum evoked efflux of [3H]-gRgR,
Since GABA satisfies all of the primary criteria as describeC by
Werman (1966) and Crrego (1979) to'identify it as a neurotrans¡nitter,
it seems reasor¡able to conclude that GABA is a neurot.ransmitter of
myenteric neuì"ones in the guinea-pig 'intestine" Although other regions
of the gu'inea-pig gastrointestinal tract have not been examined, there
is a good lixelihood t,hat GABA is also a neurotransmjttt:r'in these,
as welI as in the gastrointestinal tract of other rnanmalìan species
including the rat, cat and human.
In the ma¡nmalian myenteric plexus 35% of the nellrones have been

105
shown histochemically to Lre ejther am'ine handling or positìve for VIP,
somatostatitt, enkepha'lìn anci substance P (Furness and Costa, i980).
Cholinerg'ic neurones are the only intrinsic neurones for which both the
transmjtter and to sorne extent funcLions are known, howetrer, specif ic
histochenlical methods for the localization of choljnergic neurones are
still unproven and as yet have not been app'lìed tr¡ the intestine. Many
of the as yet unidentjfied neurones may, therefore, be choljnergic since
acetylchol'ine is a ti"anslnitter of motor and'interneurones, but there is
still room for nrore neuronal types to be discovered. Amongst these we
could include GABA-er"gic neurones. Unfortunately, autoradiogr"aphy does
not allow quantitat'ive anaìysis of cells accumulating the radiolabelled
substances therefore it is not possible to estimate the proportion of
myentepic neurones that are GABI\-ergic. Preliminary experìments iust
completed in th'is labora'bory conf it'nr the presence of these GAPrA-erg'ic
myente¡ic neuro¡es. [3H]-ci s-3-aminocycl ohexanecarboyl'ic ac'i'J (ACHC)
an¿ [3H] 1,2-4-diarnìnobutyric acid (L-DABA) which are specifically
accumulated by GABA-erg'ic neurones were localÍsed to cells of ntyerrteric
gangl i a.
Although the enteric nervous system (ENS) is relat'ive1y complex,
it is stìl1 simp-ler than the brajn and as such presents a s'imp1e and
readily accessible nlodei for the study of neurotransmissiort, particularl.y
as it survives and functjons v¡el1 ín uítt'o far long periocis. In this
regard it has proved invaluable for the ìsolatiort of the enkephalins
(see Kosterl jtz , !979). receptors for whiclr cìFÊ prês€nt 'in the mamrnal ian
gastrointestirral tract. The chemical specificity of GABA-receptors iz
uitz,o have not been fully 'Cefined. Dose response curves fcr GABA'
GABA,agonists anci antagon'ists are difficujt to measure jn both periphera'l
and central nervous preparations (De Feudis, !977). l'lowever, the ENS
may afford a preparation more amenable to pharmaco'logìcal study, where

106
as shown in this stucly it was possible to determ'ine the relative potency
of the competìtive GABA antagon'ist bicuculline. Studjes on GABA-ergic
activit'ies of GABA-analogues could be carried out using the mammalian
intestine, jncl uding cl.eterm'inat'ion of the conformational requ'irements
of GABAO-agonists and the nature of the baciofen action. Indeed, recent
studies by J.Ong in this laboratory (see Ong, 1981; Kerr and Ong, 1981)
have confirmed the specifjcÍ Ly of various GABA-analogues inc'lud'ing,
muscinrcl and 3-amino-1, propanesu'lphon'ic ac'id for the myenberic GABA
receptors.
Postsynapt'ic membrane macronrolecules involved in the act'iorr of
transnritters can ,be
assayed ín ui'bro using radioactive l'igand binding
techniques (feck et aL." !976). Receptor s'ites for GABA have been
identified by binding of radioactive GABA itself (0lsen et aL., 1978),
the GABA-agonist muscjmol (Beaumont et aL., 1978), and the GABA-ant.agonìst
bicucul I i ne-methi odi de (1'lohl er and 0kada , 1977) . Such receptor" b'ind'ing
technìques wouìd provide a rnore conrp'lete characterizat'ion of the enteric
GABA receotor types.
It has been suggested tliat benzodiazepines and barbiturates exert
the effects by facil i'tat'ing GABA-ergic transmission (Guidott'i , 1978,
1980), however, the exact nature of there actions is still controrrersial.
Whether or not. these substances exert an effect in the mammalian inbestine
needs to be determined. Recent'ìy Doble and Turnbull (1981) described a
ìack of e'tfect of benzodiazepines on b jcucul l.ine insensit.ive (GABAB)
receptors in the fiejd stjmulated guìnea-pig vas deferens
As shown in Cha¡;ter VII ATP does not appear to be the transmjtter of
intrinsic excitator"y and Ínhibitory neurones in the guinea-pig large and
small 'intestine. However, investigations of t.he transmitter mediat'ing

I07
non-cholinergic non-adrenergic inhibìtion may be aided by GABA stimulatìon
of these neurones, and the presence of GABAB receptors to the excitatory
but not the 'inhjbitory neurones (J. Ong persona'l communicatìon). A number
of quest'ions relating to this ar.rangement of GABA receptors within the
myenteric plexus remain to be answered. In partìcular, what role do GABA-
ergic neurones and related receptors have in intestinal motìlity which'is
most obvjously affected by GABA-antagonism? Nonetheless GABA is most
certainly a neurotransmitter in the guinea-pig irrtestine where it exerts
its effects at two populations of GABA-receptors.
a

108
BIBLI OGRAPHY
ABOOD, L. G., KT'RAHASI, K., GRUNER, Ë. ANd DEL CERRO, M. P. (1968)'
Nucjeotide content, caìcìum accumulation, and phosphate metaboljsm
insubcellular fractions of rat brain. Biochim. Bioph.ys. Acta, 153,
531-544.
ADAMS, P. R. and BRQIN, D. A. (1975). Actions of GABA. on sympathetic
ganglion cells. J. Phys'ioì. (Lond')' Zqq'85-120'
AHLQUIST, R. P. (1948). A study of adrenotrclpic receptors
Phys'io1 . , 153; 586-60C '
Anrerr. J .
AKUBUE, P. I. (1966). The site of action of drugs on the isolated taenia
coli from the guìnea-p'ig. Brjt. J. Pharmacol . chemother.,2-1, 347-365.
ALLY, A. I. and IIAKATSU, K. (1976). Adenosjne inhibition of isolated
rabbit ileum and antagonism by theophylf ine. J. Pharmacol. txp. Ther.'
199,208-215.
AMBAÛHE, N. anci FREEÍ'IA.N, M. A. (1968). Atropine-resistant longitud'inal
muscle spasms due to excitation of non*cholinerg'ic neurons in
Auerbach's plexus. J. Physiol. (Lond'), 199' 705-727'
AMBACHE, N., VERt'ltY, H. and ZAR, M. A" (197Ù). Evidence for the release
of tlvo atrop'ine-resistant spaslTìogens from Auerbach's plexus' J'
Phys'io1 . (Lond .) , 207 ' 761-78?.
AppENZELLAR, 0. (i976 ) . The autonomi c nervous system. New York, [i'l sevi er
Pubt ishing Co. Inc., 183-204.

109
AXELSSON, J . and HCLI'îBERG, B. (1969 ) . The eff ects of extracel 1u1a11y
applìed ATP and related compounds on the electrical and mechanical
actjvity of the smooth muscle of taenia colj of the guinea-p'ig.
Acta Physio'l . Scanci . , L5, 149-156
BARBER, R. and SAiTO, K. (1975). Light m'icroscopic visualization of GAD
and GABA-T in immunocytochemical preparatìons of rodent CNS. IN:
GABA in Nervous SJ,stem Function. Kroc Foundation Series. Vol. 5,
113-132. (f¿s. Chase, T. N. and Tower, D. B.) Raven Press, N.Y.
BARKER, J. 1., NiC0LL, R. A. and PADJEN, A. (1975). Stud'ies on cottvulsants
in the 'isolated frog spina'l cord. I. Antagon'ism of amino acid responses.
J. Physiol., 245, 5?I-536.
BARTHO, t-. and SZOLCSANYI, J. (1979).
on the guinea-pig Ìso'lated ileum.
3!5., 78-81.
The site of action of Capsaicin
l{aunyn-Schmiedebergs Arch. Pharmacol . ,
BARTLETT, V., STEI,,IART, R. R. and NAKATSU, K. (1979). Evidence for two
adenine derivative rcceptors jn rat ileum which are not involved in
the non adrenergic, non chol'inergic response. Can. J. Physioì.
Pharmacol ., 57, i130-1L37 "
BAYLISS, I^l, lrl. and STARLING, [. H" (1899). The movements and jnnervation
of the srnall intestine. "1. Physìo1 . (Lond.),.?4, 99-143.
BAYLISS, hI.M. and STARLING, E. H. (1900). The movements and the'innervation
of tlre large intestirre. J. Physiol . (Lond.), ?.6, 107-118.

110
BAYLISS, l,l. M. and STARLING, E. H. (1901). The movements and innervat'ion
of thesmall intestine. J. Phys'io'l . (Lond.), ?6, 125-138.
BAXTER, C. (1970).
Neurochemì stry.
York.
The nature of y-aminobutyric acid. IN: Handbook of
Vol. 3, 289-353. (Ed. Lajtha, A) Plenum Press, Nevv
BENNETT, M. R. (1966). Rebound excitation of tlre smooth nruscle cells of
the guìnea-pig taenia-col'i after st'imulatjon of intennural 'inhibitory
nerves. (J. Physiol. (Lond. ), 185 , I24-13I.
BENNETT, 14. R., BURNST0CK, G. and H0LIr1AN, M. E. (1966). Transmjssion frrrm
i ntramural 'inh j bi tory nerves to the s¡nooth muscl e of the gu'inea-pi g taeni a
coli. J" Phys'ioì ., IB2, 541-558.
BERNEIS, K. H., PLETSCHER, A. and DA PRADA, M. (i970). Phase separation
in solutìons of noradrenaline and adenos'i¡ie-triphosphate: influence of
bivalent cations and drugs" Brit. J. Pharmacol., 39, 382-39?.
BIANCHi , C. , BEANI , L. , FRIGO, G. M. and CRtl'lA, A. (1968) . Further
evidence for the presence of non-adrenergic inhibitory structures in
the guinea-pig colon. Eur. J. Pharnacol., 4, 51-61.
BIBER, B. (1973). Vasodilator mechanisms 'in the small intestine. Acta
Physiol . Scand. , SupPl . 401, L-31.
BOHME, H. J., KOPPERSCIILAGER, G., SCI-IULTZ, ,J. and HOFMANN, E. (1-97?-).
Affinity chromatography of phosphofructokinase using Cibacron blue
l-3G-4. J. Chromatogr., 69, 209'2L4.

111
BoLTON, T. B. (1979). Mechan'isms of a.ction of transmitters and other
substances on smooth muscle. Physiol. Reviews'99. (3)' 606-718.
BSND, p. A. (1973). The uptake of y-[3H]-aminobutyrìc acid by s'lices from
various regions of rat brain and the effect of ljthium. J. Neurochem.,
20, 511-517.
BQRNSTEIN, J. C. and FIEUDS, H. L. (1979). MorphÍne presynaptica'lìy
jnhitrjts a gangl'ionic cholinergic synapse, Neuroscience [-etters, 15,
77 -82.
Bot^ltRy, N. G. and BROt^lN, D. A. (L972). Garnma-arninobutyric acid uptake
by sympatheti c gang'l 'ia . Nature New Bi ol . , 238 ' 89-91 .
BQWERY, N. G. and BRQWN, D" A. (1974). Depoìariz'ing actions of Y-amino-
buty¡ic acid and related compounds on rat superior cervical ganglia
i.n u'i.tro, Bri t. J . Pharntacol . o 50 ' 205-218.
BOl^lERy, N. G. and HUDSON, A. L. (1979). y-aminobutyric acid reduces the
evoked release [3H]-noradrenaline from sympathetjc nerve term'inals.
Br it. J. Pharnlacol . , 66, 108P.
B0hJERY, N. G. and NEAL, M. J. (1978). Differential e'tfect of veratridjne
and potassium depolariz-ation on [3H]-engn re'lease from neurones and
gl ia. J. Physiol. (Lond.), ?8, sBP-
BOI¡JERY, N. G. , DOBLE, A.N HILL' D' R.' HUDSON, A. L. , SHAÌ^J' J. S. ANd
TURNBULL, M. J. (1979). Baclofen: a selective agon'ist for a novel
type of GABA receptor. Proc. Brit. Pharm. Soc.' 444P-

LL2
BOI^JERY, N. G., HILL, D. R., HUDSON, A. 1., DOtsLE, 4., MIDDLEI.4ISS, D. N.,
SHAI¡I, J. and TURNBULL, Í''1. (1980). (-) Bactofen decreases neurotransmitter
release in the mammalian CNS by action at a novel GABA receptor. Nature,
283, 92-94.
BOI^¡ERY, N. G., DOBLE, 4., HILL, D.R., HUDSON, A" 1., SHAW, J. S., TURNBULL,
M. J. and IIARR.INGTON, R. (198i). Bicucull ine-insensjtive GABA receptors
on perìphera'l ' autonomic nerve termina'ls. Eut". J. Pharmacol ., 7I, 53-70.
BQIJERY, N. G., JONES, G. P. and NEAL, M. J. (1976). Selective inhibition
of neuronal GABA uptake by cis-1,3-aminocyclolrexatre carboxy'lic acid.
Nature (Lond.), 4!., 2Bl-284.
BRADFORD, H. F. (1970). tvletabol ic response of synaptosomes to el ectrical
stimulation: release of amjno acids. Brain Research, L9, 239*247.
BRAZY, P. C. ancl GUI'ltl, R. B. (1976). Furosemide Ínhibition of chloride
transport in human red blood cells. J. Gen. Physì0]., 68, 533-599.
BRETAG, A. H., DAll, S. R., KERR, D. I. B. and MUSKWA, A.G. (1980).
M.yoton'ia as a side-effect of diuretjc actjon. Brit. J. Pharmacol .,
7L,467-47L.
BREUCKER, E" L. M. and JOHNSTON, G. A. R. (1975). Inh'ibition of acetyì-
chol jnestera.se by bicuculline and related alk.rloicls. J. Neurochem.,
25,903-904.
BROl^lN, C., BURNSiOCK, G" and C0CKS, T. (1979). Effects of adenosine 5'-
triphosphate (ATP) and ß-y-nrethylene A'iP on the rat urinary bladder.
Brit. J. Pharmacol., 65, 97-102.

113
BR¡1¡N, D. A. and MARSH, S. (1978). Axonal GABA-receptors 'in mammalian
peripheral nerve brunks. Brain Research, 156, 187-igi.
BROI^JNLEE, G. and JOllllSON, E. S. (1963). The site of 5-hydroxytryptam'ine
receptor on the intramural nervous p'lexus of the guinea-p'ig iso'lated
ileum. Brit. J. Pharmacol ., 27, 306-322.
BULBRING, E. and GERSHQN, M. D. (1967). 5-hydroxytryptam'ine on the
peristaltic reflex. Brit. J. Phartnacoì., 13, 444-457.
BULBRING, E. and T0MITA, T. (1967). Properties o'f the inhibjtory potential
of srnooth muscle as observed in the response to fie'ld stimula.tion of the
guinea-p'ig taenia col'i . J. Physio'l . (Lond.), 189-, 299-316.
BULBRING, E., CREMA, A. and SAXBY, 0. (1958a). A method for recording
perista'lsis 'in isolated intestine. Brit. .1. Pharmacol ., 1!, 440-443.
BULBRING, E., LIN, R. C. Y. and SCHOFIEL.D, G. (1958b). An jnvestigation
of the peristaltic refIex in relation to anatornical observations.
Quart. J. Expt'l . Physiol . , 43, 26'37 .
BURNSTgCK, G. (1971). Neural nomenclature. Nature (Lond.), ?.29,282-283.
BURNSTOCI(, G. (1972). Purinergic nerves. Pharmacol. Revjews,24 (3),
509-581..
BURNST¡CK, G. (1975). Purinergic transnrission. iN: l-landbook of Psycho-
phar'macology. Vol . 5, 131-194. (f¿s. Iversen, L. 1., Iversen, S. D.
and Snyder, S. H.) Plenum Press, New York.

114
BURNST0Cr,., G. (1978). A basis for distinguishing two types of purinergìc
receptor. iN: cell Membrane Receptors for Drugs and Hormclnes: A
Multidiscipl inary Approach. pp 107-118. (Eds. Straub, R. |lJ. and
Bolis, L.) Raven Fress, New York.
BURNSTgCK, G. (i9i9). Past and current evjdence for the purinergic nerve
hypothesìs. IN: Physiological and Reguìator"y Functions of Adenosine
and Adenine Nucleot,ides. pp 3-34. (f¿s. Baer,.H. P. and Drummond,
C. I.) Raven Press, New York.
BURNSTOCK, G. and Bell, C. (1974). Peripheraì autonon'ic transnrissjon.
iN: 'ihe Peripheral Nervous Systenr. pp 2?7-3?7. (td. Hubbard, J' I')
Plenum Press, Net/ York.
BURNSTQCK' G", CAMPBELL, G., BENNETT, M. and HOLMANN, tq. E. (1963a).
The effects of clrugs on the transmissiott from autonolnic nerves to
smocth muscie of the gu'inea-pìg taenia coli" Bjochem. Pharmacol.,
12, Suppl., 134.
BURNSTQCK, ü., CAMPBELL, G., BENNETT, M. and HQLMANN, M. i. (1963b).
Inhibition of,;he smooth muscle of the taenia coli. Nature (Lond.)'
20a,581-582.
BURNSTOCK, G., í,AMPBELL, G. , BENNTTT, M. ANd HOI.IT1ANN, M. E. (i964).
Innet'vatjon of 'fhe guinea-pjg taenìa co'l'i : are there intrinsic
inhibitory net'ves v¡hich are distjnct front sympathetic nerves?
Int. J. l'.leuropharmacol .,3, 163-166.
BURNSTOCK, G., CAMPBELL, G. and RAND, M. .1. (1966). The Ínhibitory
innervation of the guinea-p'ig caecum. J. Physio'l . (Lond' ), 182, 504-526

115
BURNSTQCK, G., CAMPBELL, G., SATCHELL, D. G. and SMYTHE, A. (1970).
Evjdence that adenosine triphosphate or a related nucieotide 'is the
transmitter substance released by non-adrenergic jnhibìtory nerves
in the gut. Brit. J. Pharmacol. Chemother., 40' 668-688'
tsURNSTOCK, G., SATCHELL, D. G. and SMYTHE, A. (197?a)" A contparison of
the excitatory and inhibitory effects of non-adrenergic, non-cho'lìnergic
nerve stimulation and exogenously app'lìed ATP on a variety of smooth
muscle preparatìons from d'ifferent vertebrate species. Brit. Ll.
Pharmacol ., 46, 234-242.
BURNSTOCK, G., DUMSDAY, B. and SMYTHE, A. (1972b). Atropine resistant
excitation of the urinary bladder: The possìbif ity of transnlission via
nerves releasing a purine nucleotide. Brit. J. Pharmacol.,44' 451-461.
BURNST0CK, G., COCKS, T., PADDLE, B. and STASZ[tlISKi-BARCZAK, J. (1975).
Evjdence that prostaglandin 'is responsible for the rebound contraction
follc¡wjng stjmulation of norì-adrenergic; (purinergic) inhìbjt'ory nerves
Ëur. J. Pharmacol., 31, 360-362.
BURNSTOCK, G., CQCKS, T., CROi,llE, R. and KASAKQW, L. (1978). Puninergic
'innervaiìon of the guinea-pjg urìnary trladder. Brit. J. Pharmacol .,
63, L25-t28.
BURY, R. hl. and MASHFORD, M. L. (1977). A pharnracological jnvestigation
of synthetjc substance-P ctr the jsolated guìnea-pìg ileum. Clin. exp'
Pharmacol . Physìol ., -1, 45-?-461.
CAJAL, S. R. Y" (1911). Histolog'ie du Systemé Nerveux de l'Homne et
des Verùébrés. Vol. II. Paris, Maloine.

CAMPBELL, G. (i966).
gu'inea-P'ig caecum.
116
Nerve-med'iatecl exci tati on of the taen'ia of the
J. Physiol , (L.ond.), lB5, 148-159.
CAÍ{PBELL, G. (1970). Autonornic nervous supply to effector tissue. IN:
Smooth Muscìe. pp 451-495. (f¿s. Bu'lbring, E., Bradin9, A. F., Jones,
A. l^I. and Tomita, T.) Londcn: Arnold.
CAMP[IELL, G. and BURNSTOCK, G. (1968). The comparìtive phys'ioìogy of
gastroi ntes'bi nal moti I 'i ty. iN : l-landbook cf Phys i o1 ogy , Sect. 6 :
Al imentary Canal Vol . IV , ?-213-2266. Ameri can Phys'io'logì ca1 Soci ety,
l¡lashi ngton , D. C.
CHAHL, l_. A. (19S1). Evidence that the cholinergìc ccntractile response
of the guìnea-p-ig ileum to capsaìcin results frorn substance P release.
Pr"oc. Aust. Physiol . Pharmacol . Soc." L2. (2) ' 113P.
CHASE, T. N. and IIALTERS, J. R. (1976). Pharmacology approaches to the
manipulation of GABA mediated synaptic fttnctìon itr man. IN: GABA in
Nervous System Function. pp 497-513" (f¿s. Roberts, E., Chase, T. N.
and Tower, D. B. ) Raven Pt'ess, Nelv York.
CH00, L. K. (1981). The effect of reactjve blue' an antagonist of ATP,
on the isolated urinary bladders of grlinea-pig and rat. J. Pharm.
Pharmacol., 33, 248'254.
CLARK, A. J. (i928). The antagonism of acetyicholine by atropine.
J. Physiol . (Lond.) , 61, 547-556.
CoCKS, T. and BijRNSTOCl.., G. (1979). Effects of neunonal polypeptìdes on
intestjnal smooth muscle; a comparison witlr non-adrenergic' non-
chol inergìc ncrve stimulal'ion and ATP. Eur. J" Pharmacol . , !1, 25I-?-59.

Lt7
COHEN, M.1., ROSING, E., l,.llLEY, K. S. and SLATËR, I. H. (1978).
Somatostatin inhibits adrenergic and cholinergic neurotransmission
in smooth muscle. Life Science, 23, 1659-1664.
C00K, R. D. and BURNST0CK, G; (1976a).
pl exus i n the gui nea-p'ig i ntest'ine.
5, t7t-194.
C00K, R. D. and BURNST0CK, G. (1976b).
pl exus 'in the gui nea-pì g i ntesii ne.
J. Neurocytology, 5o 195-246.
The ul trastructure of Auerbach's
I . Neuronal el ements. J. Neurocytol ogy,
The ul trastructure of Auerbach's
I I . Non-neuronal el ements.
COSTA, l'1. a.nd FURNESS, J. B. (1972). Slow contraction o'f, the gu'inea-pìE
proximal colon'in response to the stimula.tjon of an unidentified type
of nerve. Brit. J. Pharmacol . , 45, 151-152.
CSSTA, M.and FURNESS, ,1. B. (1976). The per"ist.aìtic reflex: an analysis
of nerve pathways and their pharnracoìogy. Naunyn-Schmiedeberg's Arch.
Pharmacol ., 294, 47'60.
CSSTA, M. and FURNESS, J. B. (tgZga). 0n lhe poss'ibility that an indoleamine
is a neurotransmitter in the gastro'intestirtal tract. Biochem. Pharmacol .,
28, 565-571.
CQSTA, M. ancl FURN[SS, J. B. (1979b). The sites of actjon of S-h¡rfl¡c'¡y-
tryptam'ine 'in nerve-muscle preparatjons frorn the guinea-pìg smaì1
ìntestìne and colon. Brit. J. Pharrnacol., 65, 237-?48.
CQSTAn M., BUtFA, R., FURNESS, J. B. and S0LCIA, E. (1980). Immunohjsto-
chemical local izat'ion of polypepticies 'in peripheral autonomic nerves
usìng whole mount preparations. Histochemistry, 61, 157-165.

118
CQSTA, M., FURNESS, J.8., BUFFA, R. and SAIDo S. I. (1980). D'istribution
of enteric nerve ceil bodies and axons showing immunoreactivity for
vasoactive intestinal polypeptide in the guinea-p'ig ìntest'ine.
Neuroscience, 5: 587-596.
cQsTA, M., FURNESS, J.8., LLEWELLYN-SMITH, I. J. and cUtLLQ, A.c. (1981).
Projections of substance P-containìng neurones wjthin the guinea-pig
small intestine. Neuroscience, 6 (3), 4Il-424"
COl^lAN, l,J. f'l., GOTTLIEB, D. I., HENDRICKSON, A. E", PRiCE' J. L. and
1J66LSEY, T" A. (1972). The autorad'iographic denronstration of axonal
cc¡nnectjons in the central nervous system. Brain Research, 37, 2l-5L.
CREI'IA, 4., FRIG0, G. M. and LECCHINI, S" (1970). A pharmacological analysìs
of the peristaltìc reflex in the isolated colon of the guinea-p'ig or
cat. Brit. J. Pharmacol., 34, 334-345.
CURTIS, D.R. and JOHNSTON, G. A. R. (1974). Amino-acjd transnritters in
the mantmalian central nervoLls system. Ergebn. Phys.iol ., 69, 97-188.
CURTIS, D. R" and I^IATKINS, J. C. (1965). The pha'r'nraco'lcgy of anrino acids
to gamma-aminobutyric acid. Pharmacol. Rev -, 17 (4) ' 347-391.
CURTIS, D. P.., DUGGAN, A. lll., FELIX, D. and JQ}iNSTÛN, G. A. R. (1970).
GABA, bicuculline and central inhibition. ll.r+;ttre (Lond.), ry-,r222-t224.
CURTIS, D. R., FELIX, D. R", GAI'IE, C. i. A' and l4cCUl-L,CICK, R. M. (1.973)
Tetanus toxin and t.he synaptic release of GABA. Braitl Research, 51,
362-385.

119
CURTIS, D. R., GAME, C. J.4., J0HNSTON, G. A. R. and I'1cCULLOCH, R. M.
(1974). Central effects of ß-(p-ch'lorophenyl )-y-aminot:utyric acid.
Brain Research, 70, 493-499.
CURTIS, D. R., PHILLIS, J. l¡1. and WATKINS, J. C. (196i). Cholìnergic and
non-cholinerg"ic transm'iss'ion 'in the manmai'ian spinal cor"d. J. Physiol . o
158, 296-323
DANIEL, E. E. (1968). Pharmaco]ogy of the gas't,ro'intestinal tract. IN:
Handbook of Phys'io1ogy, Sect. 6 Al imentar,y Canal, Vol . IV, Motì1ity,
2267-2324.
DA VANZO, J" P., IqATHEWS, R.. J., YOUNG, c. A. and I^IINGERS0N, t. (tg6+).
Studies on the mechanism of actjon of amino-oxyacetjc acid. Tox.
Appl . Pharrnaco'l ,,6,396-401"
DAY, M" and v¡tNE, J. R. (1963)" An analysis of the direct and indirect
actÍ clns of drugs on the i sol ated gui nea-p'ig i I eum. Bri t. J . Pharmacol . ,
20, 150-170.
DEAN, D. M, and D0liNIE, L'l. (1978). Interaction of prostagland'in and
aclenosine 5'-triphosphate in the non cholìnergic neurotransmÍssion in
rabbjt detrusor. Prostagìandjns, 16, ?45-251"
DE FEUDIS, F. V. (1975). Amino acids as central neurotransmitters.
Ann. Rev. Pharmacol., 15, 105-130.
DE FEUDIS, F. V. (1977). GABA-receptors in the vertebrate nervous system.
Prog. Neurobiol., 9, 123-145.

r20
DE FEUDIS, F. V., DELGADQ, J. M. R. and RQTH, R. H. (1970). Content,
synthes j s and col I ectabi ì ity of aminci ac'ids in various structures of
the brains of rhesus monkeys. Bra'in Research, 18, 15-23.
DE GROAT, hl. C. (1970). The actions of y-aminobutyric acid and rejated
amino acids in mammalian autononric ganglìa; J. Pharmacol. Exp. Ther.,
t72,384-396.
DE GROAT, I^t. C., LALLEY, P. M. and BLOCK, M. (1971) . The effect of
bjcucull.ine and GABA on the superior cervical gangl'ion of the cat.
Brain Research, 25, 665-668.
Dt GR0AT, I,l" C., LALLEY, P. ¡vl" and SAUI{, þl. R. (1972). Depolarizatìon
of dorsal root ganglia jn the cat by GABA and related amjno acids:
antagoni sm by pi crotoxi n and bi cucul I j ne. Brai n Research , 44-, ?73-277 .
DE l".A LAI',IDE, I. S. and PORTER, R" B. (19õ7) " Factor"s influencing the action
of nrorphine on acetyìcholjne release in the guinea-pìg ìntestìne. Brit.
J. Pharmacol. Chemother., 29 (2), 158-i67'
DESCHENES, lu1. , FËLTZ, P. and r-AMOUR, Y. (1976). A model for an estimate
ín uiuo of the ion'ic basis of presynaptic inhibition: an intracellular
analysis of the GABA-induced depoìarizatjon in rat clorsal root gang'l'ia.
Brain Research, 118, 486-493.
DIAB, I. M., DiNERSTEIN, R" J. I^IATANABE, M. and RÜTH, L. J. (1976).
[3H] *orph'ine local jzaticln in myenteric neurones" Sciettce, 193, 689-69i.
DIAMENT, M. 1., K0STERLiTZ, H. l¡l. and I'lcKENZLI-, J. (1961). Role of the
mucous nlembrane'in the perista'ltic reflex in the isolat.ed'ileum of the
guinea-pig. Nature (Lond. ), 190, 1205-12A6.

127
DINGLEDINE, R. and G0LDSTEIN, A. (1976). Effect of synaptic transnrjssion
bloclcade on morphine act'ion in the guìnea-p'ig myenteric plexus.
J. Pharmacol. Exp. Ther., 196, 97'106-
DoBLE, A. and TURNBULL, M. J. (1981). Lack of effect of benzodiazepines
on bicucull ine-insensjtive GAB-receptors iri the field-stinlulated
gu'inea-pig vas deferens preparation. J. Pharm. Pharmacoì ., 33: 267-268
DOGIEL, A. S. (1895).
den Säugetieren.
Zur Frage über die Ganglien der Darmgeflechte bei
Anat. Anz., 10, 517-528.
D9GI:E¡-, A. S. (1q99)" Uber den Bau der Gangljen in den Geflechten des
Darmes und der gal'lenbl ase des Menschen und der Satrgeth'iere.
Arch. Anat. Phys'iol ., !, i30-159.
DQl{ilALL, t'1. J., BQYNE, A. F. and i{HITTAKER, V. P. (I974). Adenosine
triphosphate. A const'ituent of chol ìnergic synapt'ic vesicles.
Biochem. J., 140 (1), L-I?.
DOl,lNIE, J. W. and DEAN, D. M. (1977). The ccntribut'ion of chol inerg'ic
postgangl ionic ner¡rotransmjssion to ct¡n'uract'ions of rabbit detrusor.
J. Pharmacol. Exp. Ther.. 203, 417-425.
D0trll{ING, D. A. and MORRIS, J. S.
stomach by bradykinin. Arch.
(1981). Relaxation of guinea-pig
Int. Pharnracodyn., 249, 229-234.
DREYFUS, C. F., SHERMAN, D. L. and GERSHON, l'1" D. (1977). Uptake of
serotonin by intrins'ic neruones of the myenterìc plexus grown in
organotypic tjssue culture. Brain Research, i?B, 109-123.

122
DRSZ, B. and LEBLQND, C. P. (1963). Axona.l rn'igration of proteins in the
central nervous system and peripheral nerves as shcwn by radioautography.
J. Comp. Neurol ., I2I'- 325-346.
DUDEL, J. (1965). Presynaptìc and postsynaptìc effect of inhib'itory drugs
on the crayfish neuromuscular junction. Pf1ügers Arch. geS. Phys'iol. '283, 104-118"
DUPONT, J. R., JERVIS, H. R. and SPRINZ, H. (1965).
the rat cecum 'in relation to the germ free state.
r25, 11-18.
Auerbach's pl exus of
J. Comp. Neuroì . ,
ELI-I6TT, K. A. C. And VAN GELDER, N. M. (i978). 0cclusjon and metaboljsm
of "¡,-aminobutyric acid by braìn tjssue. J. Neurochem., 3, 28-40.
tf'lER.g, M. A. and SAIDI4AN, B. A. (!977). Possìble feedback jnhib'ition of
noradrelral'ine release by puritte compounds. Naunyn Schmiedeberg's
Arch. Pharmacol . , 297, 39-46.
FAGG, G. E. and LANE, J. D. (i979). The uptake and release of puta'tit'e
amino-acid neurotransmittei"s. Neuroscience, 4, 10i5-1036.
FAHII¡, S. (1976). Regionaì distribution stuC'ies of GABA and other
putati ve neurotransm.itters and thei r enzJ¿mes ' IN: GABA i n Nervous
System Funct'ion. pp 169-186. (Ecis. Roberts, t., Chase, T. N. and
Tower, D, B. ) Raven Press, Ne'¿,1 York.
FAHRENKRUG, J., I-{AGLUND, U., JODAI., M., LUNGRTN, 0., OLBE, L. ANd
SCHAFFAI.iTZKY DE MUCKADËLL, 0. B. (1978). Nervous release of vasoact.jve
jntestinal po'lypeptìde ìn the gastrointestinal tract of cats: possìb1e
physio'logical ìrnp1ìcations. J. Physjol . (Lond .), ?W, 291-305.

123
FARBER, E., srERl{BERG, l,l. H. and DUNLAP, c. E. (1956). Histochemjcal
localization of specifjc oxidative enzymes; betrazolium stajns for
diphosphopyridìne nucleot'ide diaphorase and trìphosphopyridine
nucleotide diaphorase. J. Histochem. Cytocheil.', 4, 234-265.
FASTH, S., HULTEN,1., JOHNSON, B. J. ANd ZËITLIN, I. J. (T977). ChANgCS
in the kinin system in cat colon durìng the atropine-res'istant response
to pelvic stimulation. J. Phys'io'i . (Lond.), 265, b6p.
FELDBERG, llJ. and LIN, R. C. Y. (1949). The action of'local anaesthetics
and D-tubocurarine on the isolated intestíne of the rabbit and guÍnea-
pig. Brit. J. Pharmacol.,4,33-44.
FrsHLOCK, D. J. (1966).
and large intestine.
Effects of bradykinjn on the human jsolated small
Nature, 2L2, 1533-i535.
FL0CK, A. and LAItl" D. (1974). Neurotr'ansmitter synthesis in 'inner ean
and lateral 'line sense organs. Nature (Loncl. ), Z4g, !42-145.
FLOREY, t. and McLENNAN, H. (1959). The effects of factor I and of
garnma-aminobutyric acid on smooth rnuscl e preparations. J. ph¡,sioì .
(Lond. ), 145, 66-75.
FL0REY, E., cAHiLL, Iut. A. and R.ATHMAYER, M. (1975)" Ëxcitatory act jons
of GABA and of acetylchol ine jn sea urch'in tube feet. Conrp. Biochern.
Physiol.,5lc, 5-12.
FORSTER, p., LLOYD, H. G. E., MORGAN, p. F., PARKER, M., PERKINS, 1,1. N.
and STONE, T. t,J. (1981). Ethylenedianiine acts upon GABA receptors and
uptake s'ites. Proc . of B. P. S. , lst-3rd Apri 1 , ZT4p .

124
FOX, S., KRNJEVIó, K., MORRIS, M. E., PUIL, I. ANd lnlERMAN, R. (1978).
Action of baclofen an mammal'ian synaptic transmjssion. Neuroscìence,
3,495-515.
FRANCO, R., C0STA, M. and FURNESS, J. B. (1979a). Ev'idence for the release
of endogenous substance P from intestinal nerves. Naunyn Schml'edeberg's
Arch. Pharmacol . , 306, 185-201.
FRANCO, R., COSTA, t4. and FURNESS, J. B. (1979b). Evidence that axons
containing substance P in the guìnea-p'ig iìeum are of intrinsic origin.
Naunyn-Schmiedeberg's Arch. Pharmacol ., 307, 57-63.
FREEMAN, 4.. R. (1973). Electrophysiological analysis of the actions of
strychnine, bicuculline and pìcrotoxin on the axonal membrane.
J. lleurobioì., 4, 567-582.
FRIG0, G" M. and LECCHINI, S" (1970). An inrproved method for studying
the peristaltic reflex in the isolated colon. Brit. *1. Pharmacol.,
39, 346-356.
FURNESS, J. B" (1969a). An electrophysÍoiogícal study of the innervation
of the snrooih muscle of the colon. J. Ph"vsjoì. (Lond. ), 205, 549-562.
FURNESS, J. B. (1969b). The presence of inhibitory fibres in the colon
after sympathetic cl.enervation. Eur. J. Pharniacol",6, 349-352.
FURNESS, J. B. (1970). The origin and distribut'ion of adrenerg'ic nerve
fibres in the gurinea-pig colon. Histochemie, 2L,295-306.

r25
FURNESS, J. B. (1971). Secondary excitation of intestinal smoo't,h muscle.
Bri t. J . Pharmacol . , 41, 2L3-2'¿6 .
FURNESS, J. B. and COSTA, M. (1973). The nervous release and the action
of substances v¡hich affect intest'inal muscle through neither adreno-
receptors nor cholinoreceptors. Phil. Trans. R. Soc., 265, 123-133.
FURNESS, J. B. and C0STA, M" (1974). The adrenergic innervation of the
gastroìntestjnal tract. Ergebrr. Physiol. Biol. Cheni. Exp. Pharmakol.,
69,1-51.
FURNËSS, J. B. and COSTA, M. (1978). Djstribution of intrinsjc nerve
cell bodies unfl uron, which take up aromatic amines andtheir precursors
in the srnall intestine of the guinea-pig. Cell Tissue Res., 189' 527-543.
FURNESS, J. B. and COSTA, M. (1979). Actjons of somatostat'in on excìtatory
ancl jnhìbitory nerves in the intestine. Eur. J. Pharmaccll., 56, 69-74.
FURNESS, J.8., C0STA, M., FRANCO, R. and LLEWELLYN-SMiTI-|, I. J. (1980).
Neuronal peptides jn the intestjne: distribution and poss'ible func'bjons.
IN: Neural Pept.ides and Commun'icat,ion. pp 601-617. (tcls. Costa, E.
and Trabucchi, M. ) i{aven Press, NevJ York.
FUXE, K", HörrEtt, T., sAID, S. I" and F1UTT, V. (L977). Vasoactive
jntestinal po'lypeptide and the nenrous system. Immuno-histochemical
eviclence for local Ízation in central and periphera'l neurones,
partìcularly intracortjcal neurones of the cerebral cortex. Neuroscience
Letters, _5, 24I-246,

cABEt-LA, G. (1967 ) .
and adul t rats.
126
Neurone number ìn the myenteric pìexus in new-born
Experientia, ¿1 (1), 52-53.
GABELLA, G. (1969). Detectjon of nerve cell.s by a histochemical technique.
Experientia, 25 (2), 2LB-219.
GABEI-LA, G. (19ii) . Gl ial cel I s jn the myenteric p'lexus . Z. Naturforsch,
26b, 244-245.
GABtLI-A, G. (I972). Innervation of the intestinal muscular coat,
J . Neurocytol ogy, _]. , 34I-362.
GABELLA, G. (i974). Specia'l muscle cells and their innervation ìn the
mammal ian small intestine. Cell Tissue Res., I53, 63-77.
GABËLLA, G" (i976). Structure of the Autonomjc Nervous System. Chapman
and Hali, London.
GABtLLA, G. (1979). l.nnervation of the gastrointestinal tract. Int.
Rev. Cyto'l .,590 129-193.
GABELLA, G. and C0STA, M. (i969). Adrenergic innervation c¡f the intestjnal
smooth musculature. txperientia, 25 (4), 395-396.
GALINDO, A. (1969).
nervous s¡,rstem.
GABA-picrotoxin interactjon in the mammalian centrai
Brain Res. , L4, 7b3-767. u
GALLAGIIER, J. P., HiGASI-iI, H. and HISHI, S. (1975). The ionic requ'irenients
for the production cf the GABA depolarjzatic¡n at cat primary afferent
neurones. Fedn . Proc . , 34, 418 ( 1121P ) .

t27
GARDNER, C. R. and RICHARDS, M. H. (1981). Presence of radiolabelled
metabolites in reiease studies using [3H] "¡-amìnobubyric acid"
J. Neurochem'istry, 36 (4), 1590-1593.
GERSCHQNFELD, ll. M. and LASANSKY, A. (i964). Actìon of gìutamic acid and
other naturalì.y occurring amìno-acids on snail central neurones.
Int. J. Neuropharnlacol ., 3, 301-314.
GERSHQN, M. D. (1.96i). Effects of tetroclot,ox'in on innervated smooth muscle
preparatiorrs. Brit. J. Pharmacol ., ?9, ?59-279,
GERSHflN, M. D. (1,981). The enteric nervous system. Ann. Rev. Neurosci."
4, 227 -2.72.
GERSH0N, M. D. and RQSS, L. L. (1966a). Radioisotopìc studies of the
bindjng, exchange and distribution of 5-hydroxytrypl:amìne synthes'!zed
from its re.<!ioactive precursoy'. J. Phys iol . (Lond.), _1t0, 45L'-476.
GERSHON, M. D" and R0SS, L. L. (1966b). Localization of sjtes of 5-
hydroxytryptarni ne storage and metabc¡l i stn by rad j oautography. J.
Physiol . (t-ond.), 186, 477-492.
GERSHON, M. D." DRAKONTIDES, A. B. and R0SS, L. L. (1965). Serotonin:
Synthes'is ancj rejease from the myentert'c plexus of the mouse'int,estine.
Sci ertce , I49 , '197 -I99,
GINSBQRG, B. L. and HIRST, G. D. S. (1972). The effect of adenosine on
the release of the transmitber from the phrenic nerve of the cat.
J . Phys i o'l . (Lond ") , 224 , 629-645 .

t28
GINZEL, K. H. (1959a).
peristaìtic ref1ex?
Are mucosal nerve fibres essentia'l for the
Nature (Lond.), 184, 1235-1236.
GINZEL-, K. H. (1959b). Investigations concerning the jnìtiatjon of the
peristaltic reflex in the guinea-pig 'i1eum. J. Physiol. (Lond.), 148,
75-76.
GRAFE, P.o GALVAN, M. and ltlAYER, L. J. (1979). Actions of GABA on guinea-
p'ig myenteric neurones. Pflügers Arch. Suppl to Vol . lB3.' I75.
GRAFE, P., MAYER, L. J. and l^l00D, J. D. (1979). Evjdence that substance
P cloes not rnediate slow synaptjc excitatjon within the myenterìc
pl exus. Nature (Lond .) , ?-19, 720-72I.
GREEF, K., KASPERAT, H. and OSSI¡JALD, t^J . (L962). Paradoxe wirkungen der
eleltvischen Vagusreìzang am isolierten Magen-und Herzvot hofpräparat
des l',leenschr¿¡ei nchens sowj e deren Bee'inf I ussung durch Gangì i en bl ocker
Sympathicolytica, Reserpin und Cocajn. Naunyn-schnliedeberg's Arch.
Pharmakol . Exp. Pathol ", ?9, 528-545.
GUIDOTTI, A" (1978) . Synapt'ic mechani sms in the action of benzodìazep'ines.
IN: Psychopharmacology - A Generatjon of Progress. pp L349-I357.
(E¿s. Lipton, M.4., D'imascio, A. and Killam, K. [:.) Raven Ppess, llew
York.
GUIDOTTI, A. (1980). Pharmacologica'l and biochemjcaj evidence for an
interactinn between the benzodiazep'ine and GABA receptors. IN:
Receptors for lleurotransmitters and Pept'ide Hormones. pp 271-275.
(E¿s. pepeu, G. , Kuhar, M. J. and ENNA, S. J. ) Raven Press, llew York.

129
GUILLEMIN, R. (I976). Somatostatin inhibits the release of acetylcholine
induced electricall.y irr the myenteric p'lexus. Endocrinoìogy, 99,
1653- 1 6s4 .
GUNN, M. (1959). Cell types in the myenteric plexus of the cat. J
Neurol., f]1, 83-i00.
Comp
GUt'lN, M. (1968). Histo]ogical and hjstochemical observatjcns on the
myenteric and submucous plexuses of mammals. J. Anat.,102 (2),223-239.
HEDQVIST, R. and FREDI-|O["M, B. B. (1976). Effects of adenosine on
adrenergic neurotransmissjon: prejunctjonal inhibitìon and post-
iunctional enhancement. Naunyn Schmiedeberg's Arch. Pha.rrnacol ., 293,
2I7 -223.
HENN, F. and I-IAMBERGER, A. (I971) . Gl ial cel I f unction: uptake of trans-
mitter substances. Proc. Nat. Acad. Sci. US., 68, ?-686-?691.
HILL, C. J. (1927). A contributiorr to our knowledge of the enteric plexuses.
Phil. l"rans" Roy. Soc. l-ondon, Ser. 8., 215, 355-387.
HILL, D. R. and BOl,,lERY, l,¡. G. (1981). 3H-baclofen and
bicucul l'ine-insensi tÍve GABAU si tes in rat bra'in.
L49-I52.
HIRST, G. D. S. and f'lcKiRDY, H. C. (L9-14).
descending ìnhibition ìn guinea-pig sma'11
238, L29-r44"
JH-GABA bi nd to
Nature , 290,
A nervous mechanism fon
intestine. J. Physiol . (L,ond.),
HIRST, G. D. S" (i979). Mechanisms 01= perìsta1sis. Brit. Irled. Buìl .,
91 (3) , ?.63^268.

130
HIRST, G. D. S. and McKIRDY, H. C. (1975). Synaptic potentjals rect¡rded
from neurones of the submucous pìexus of the guinea-pig small intestine"
J. Physio'l . (Lond .) , ?49, 369-385.
HIRST, G. D. S., HOLMANN, M. E. and SPENCE, I. (1974). Two types of
neurones in the myenteric plexus of duodenum in the guìnea-p'ig. J
Phys'io1 . (Lond .) , 236, 303-326.
tllRST, G. D. S., H0LMANN, M" Ë. and McKIRDY, H. C. (1975). Two descendÍng
nerve pathways activated by dìstension of guinea-pig small intestine.
J. Phys'io1 . (Lond.), 244, LL3-127.
HOBBIGIR, F. (1958a). Effects of y-aminobutyric acjd ctr the iso]ated
mammalian ileum. Jap. J. Physjol .,,1-{2, I47-L64.
HSBUIGER, F. (1958b). Antagorrisrn by y-arnìnobutyric acjd to the actions
of 5-hydroxytryptamine and nicotine on isolated organs. J. Phys'iol.'
r44" 349-360.
Hgffftt, t. and LJUNGDAI-IL, A" (i970). Cellular localization of labelled
gamma-am'inobutyric acìd [3H]-enBA in rat cerebellar cortex: An
autoradiography study. Brain Res., ??, 391-396.
Hoff Elt, T. and LJUNGDAHL , A. (1972) . Il'l : Stud'ies of Neurotransmi tters
at the Synaptic Level. Advances ìn Biochemical Psychopharmacology,
Vol.6, pp 1-36. (Eds. Costa, E., Iversen, L. L. and Paoletti, R.)
Raven Press, New York.

131
nörrrir, T., ELDE, R., J0HANSSON,0., LUFT, T. and ARIMURA, A. (1975).
Imnrunohistochem'ical ev'idence for the presence of somatostat'in a
powerful inhjbitory peptide in some primary sensory neurones.
Neurosci . Letters , 1, 23L-235.
HöKFELT, T., TLDT, R., JOIIANSSON, 0., LUFT, R:, NILSSOI!, G" ANd ARIMURA'
A. (1976). Immunohistochemical evidence for separate populatjons of
somatostatin-containìng and substance P-contaìning primary afferent
neurones in the rat. Neuroscience,1,131-136.
HöKFELT, T., JONSSON, G. and LJUNGDAHL, A. (1970). Reg'ional uptake anci
Isutrcellular localization of ["H] gamma-aminobutyrìc acid (GABA) 'in ra't.
brain slices. Life Science, 9,203-212-
HöKFILT, T., JOHANSSON, O., KELI.ERTH, J.0., LJUNGDAHL,4., NILSSON, G.,
NyGARDS, A. and PERNOW, B. (L977). Inmr¡nohistochemicaj djstribut'ion
of substance P. IN: Substance P. Nobel Syrnposium, Vo. 37, pp 117-145.
(EOs. Von Euler, U. S. and Pernow, B.) Raven Press, Nelv York.
HOLMANN, M. E. , HIRST, G. D. S. and SPENCE, I. (.7972) . Prel imjnary
studi es of the neurones of Auerbach ' s pl exus Lrs'ing i ntracel I ul ar
microelectrodes. Aust. J. Exp. Biol . Iuled. Scì., 50, 795-801.
H0LMANN, M. E. and HUGHES, J. R. (1965).
muscl e. Aust. J . Exp. Bi ol . l4ed. Sci
InhiLrition of iniestinal smooth
, 43, 277-?go,
HOLTON, F. A. and HOLTON, P. (1954). The caÞ'ilìa.rry dilator substances in
dry powders of sp'inal roots: a poss'ible role of adenosine triphosphate
in clremical transmìssion from nerve enciings. J. Physiol . (Lond.), !26-,
t24-140.

L32
HQLZER, P. and LEMBECK, F. (1978). Effects of neuropept'ides on the
e'tficiency of the peristalùic reflex. Naunyn-Schnliedeberg's Arch.
Pharmacol ., 307, 257'264.
HOSLI, L. and HOSLI, E. (1972). Autorad'iographìc localization of the
uptake of gìyc'ine jn cultures of rat medulla oblongata. Brain Res.,
45,6t2-6r6.
HSSLI, L. and HOSLI, E. (1978). Action and uptake of neurotransmitters
in Cl,lS tissue culture. Rev. Physio'l . Bjochem. Pharmaco'l ., 81, 136-188.
HoSLI, L. and HOILI, E. (1979). Autoradiographìc studies on the cellular
local'ization of GABA and ß-alanine uptake by neurones and glìa in
tissue culture. IN: GABA-Biochemistry and CNS Function. pp 205-218.
(fas. Mandel, P. and De Feudis, F. V.) Plenum Press, New York.
H¡SLI, E. and HQSLi, L. (1980). Cellular localization of the uptake ofa
["H:j ß-alanjne in cultures of the rat centra] nervous system.
Neuroscience, 5, I45-I52.
HUKUI1ARA, T. and FUK.UDA, H. ( i965 ) . The nroti 1 i ty of the j sol ateci
guinea-p'ig smaìl intestine. Jap. J. Physio'l ,, 15, 125*139.
HUf..UHARA, T. , YAMAGAMI , M. and NAIGYAMA, S . (1958) . 0n the i ntestìnal
intrinsic reflexes. Jap. J- Physiol., 8, 9-20.
INQUYE, 4., FUKIJYA, M., TSUÛHIYA, K. and TSt,lJI0f.A, T. (1960). Studies
on the effects of y-am'inobutyrìc acid on the isolated guinea-pig'ileum.
Jap.J. Phys'iol .,10, t67-I82.

IVERSEN, L. L. (1967).
sympathet'ic nerves .
133
The uptake and storage of noradrenaline in
Cambridge Unìversjty Press, Cambridge.
IVERSEN, L. l-. and BL00M, F. t. (Ig72). Studjes of the uptake ot [3H]
GABA and [3tl] glycjne in slices and homogenates of rat brain and
spinal cord by electron mìcroscopic autoradiography. Bra'in Res.,
41, 131-143.
IVERSEN, L. L. a.nd JOllNSTOt{, G. A. R. (1971). GABA uptake in rat central
nervous system: cornparison of uptake in slices and homegenates and
the effects of sorne inhibitors. J. Neurochem., 18, 1939-1950.
IVERSEN, L. L. and KILLY, J. S" (1975). Uptake and metabol'ism of
y-am'inobutyric acjd by neurones and glial cells. Biochern. Pharmacol .,
24, 933-938.
IVERSEN, L. L" and NËAL, M. J. (1968). The uptake ot [3H] GABA by s'lices
of rat cerebral cortex. J. Neurochem., 15, 1141-1149.
IVERSEN, L. 1., IVERSF:N, S. D. and SI'{YDER, S" H. (19i5). Handbook of
Psychclpharmacoi ogy " Vo j . 4. Ami no Ac j d Neurotransm'itters . Pl enum
Pres s , Ner,,/ York .
IVERSËN , L. L. , MITCI-IELL, J . F. and SRINIVASAV, V. (I971) . The rel ease
cf garnrna-aminobutyrjc acid during ìnhibition in the cat vìsual cortex.
J. Physìo1 . (Lond.), ?]?,519-534
JACgBS, J. M. (I977). Fenetration o't systemica'l1y- jniected horseradish
peroxidase'into gang'ìja and nerves of the autonorn'ic nervous system.
J. Neurocytol.. 6, 607-618.

134
.]ASPER, H. H. and KoYAMA, I. (1969). Rate of release of amino acids from
the cerebral cortex in the cat as affected by brainstent and thalanl'ic
stirnulation. Can. J. Physìo1. Pharmacol -, 47 (10) ' 889-905.
JESSEN, K. R., MIRSKY, R., DENNISON, M. E. and BURNSTOCK, G. (1979).
GABA may be a neurotransnljtter in the vertebrate peripheral nerves.
Nature, 28I, 71-74.
JESSEN, K. R., SAFFREY, M. J., VAN NOORDEN, S.' BLOOM' S. R", POLAK, J. M.
and BURNST0CK, G. (1980). Immurroh j stochemìcal studies of the enteric
nervous sys'eem in tjssue culture and in s'itu: local'izatiotr of vaso-
active intestinal pept'ide (VIP), substance-P and enkephalin immuno-
reacti ve nerves i n the gu'inea-pi g gut. Neurosc'ience , 5, 1717'7735.
JQHNS$N, S. M., KATAYAMA, Y. and l'lORTH, R. A. (1980). Slow synapt'ic
po'i,e¡t'ial s in neurones of bhe myenteric pl exus. J. Physio'l . (Lond. ) ,
301, 505-5i6.
JQHNSTQN, G. A. R. (1976). Physìoìogícaì pharmacoìogy of GABA and 'its
ant.agon'ists in the vertebrate nervous system. Il'l: GABA in Nervous
System Function. pp 395-41.i. (Eds. Roberts,8., Chase, T. N. and
Tower, D. B.) Raven Press, llevt York.
J0I1NSTON, G. A. R. (1978).
neurotransnritters. Ann
Neuropharmacology of amino acid inhìbitory
Rev. Pharmacol. Tox., 18, 269-289.
JQHNSTQN, G. A. R. and IVERSEN, L" L. (1971). Gìycine uptake in rat
central nervous systenr slices and hornogenates: evidence for different
uptake systenrs ìn spìna1 cord and cerebraj cortex. J. Neurochem., 18,
1951-1961.

135
J0NAKAIT, G. M., TAMiR, H., GINTZLER, A. R. and GERSHON, M. D. (1979).
?Release of ["n] serotonin and its binding proteìn from enteric neurones.
Brain Res., L74, 55-69.
f..40, C" Y. (1966). Te'brodotoxìn, saxitoxin and their s'ignificance in
the study of excitation phenontena. Pharmacol. Rev., _13' 997-1049.
KATAYAI4A, Y. arrd NORTH, R. A. (1978). Does substance P mediate slow
synaptic excitat'ion within the myenteric plexus? Nature (Lond.), ?l!,387-388.
KATAYAFIA, Y. and,NORTH, P.. A. (1980) . The action of sotnatostat jn on
neurones of the myenteric plexus of the gu'inea-pìg ilerrm. J. Physioì.
(Lond.), 301, 315-323.
KATZ, R. I., CHASE, T. N. and K$PIN, I. J. (i969). Effect of ions on
stirnulus-'induced release of amino acids from mammalian brain slices.
J. Neurochem. , 16, 961-967 .
KEHQE, J. (I972). Ionic mechan'istns of ttlo-component chcl'inergìc
inhib'ition in Aplysia neurones. J " Phys'io1 . , 225 ' 85-114.
KENNEDY, J.4., KERR, D. I. 8., KRANTIS, A. and 0KADA, l'. (1981).
Antagonism o-1 the contractile effect.s of ATP ìn guinea-pig ìleum
by tri azinyl dyes. Proc. Aust. l'leurosci . Soc. , 1' BgP.
KERKUT, G. A. and MEECH, R. W. (1966).
of H and D cells in the snail brain.
819-832.
The i nternal chl ori de concentrat,'ion
Cornp . Bi oc hem . Phys'i ol . , 1,9 ,

136
KERKUT, G. A. and l,lALKER, R. J. (i962).
of Hel i x nerve cel I s . Comp. Bi ochern
The specìfic chem'ical sens'it'ivìty
Physi 01 ., L, 277-288.
KERR, D. I. B. and 0NG, J. (1981). GABA actions in guinea-p'ig ileum
through release of endogenous GABA by ethylenediamine. Proc. Aust.
Soc. Clin. Exp. Pharmacol " (Dec., Adelaide) o 3P.
KILBINGER" H. and I,JESSLER, I. (i980). Inh'ib'it'ion by acetyìcholine of
the stimuiatjon-evoked release of [3H] acetylcholine from the gu'inea-
pig myenteric plexus. Neuroscience, 5, 1331-1340.
KIM, T. S., SHULMAN, J. and LEVINE, R. A. (1968). Relaxant effect of
cyc'lic adenosine-3', 5'-monophosphate on the jsolated rabbìt ileum.
J. Pharmacol . Exp. Ther., J.ffi, 36-42.
KQSTtRLITZ, H. l,ll. (1967). Intrinsic intestjnal reflexes. Anrer. J. dig.
Dis., 12, 245-254.
KQSTERLITZ, H. l,J. (i968). Intrjnsìc and extr insic nervous cont,rol of
motiIity of the stomach and the intestines. IN: Handbook of Ph.vsiolog¡',
Sect. 6, Vol" IV, pp 1579-i627, Anerican Plrysìological Socìety.
KOSTERLITZ, H. I^J. and LEES' G. M.
intrins'ic intestinal reflexes.
(1964). Pharmacolog'ica.l anaìysis of
Pharmacol. Rev., 16, 301-339.
KQSTERLITZ, H. W. and ROBINSON, J" A. (1959). Reflex corrtractions 'of
the longitudina'l muscl e coat of the isolated guìnea-pig 'iìeum. J'
Physiol . (Lond.), !re_, 369-379.

t37
KQSTERLITZ, H. þ'l. and WATERFIELD, A. A. (i975). In uítto models in the
study of structure-actjvity relat'ionsh'ips of narcotic analgesìcs.
Ann. Rev. Pharmaco'l ., 75, 29-47.
KQSTERLITZ, H. l.rl. ancl I{ATT, A. J. (1965). Adrenergic receptors in the
guinea-p'ig 'il eum. J. Physi o1 . , I77 , 1lP.
KOSTERLITZ, H. 1,'l., LYD0N, R. J. and WATT, A. J" (1970). The effects of
aclrenalìne, noradrenaline and isoprenal jne on inhibjt'ory and ß-
adrenoceptors in the'longìtudinal muscle of the guinea-pig'ileum.
Brit. J. Pharmacol., 39, 398-413.
K0STERLITZ, H. l¡l., PIRIE, V. l''|. and ROBIll50N, J. A. (1956). The mechan'isrn
of the pe¡istaìtic reflex in the isolated guinea-p'ig ileum. J. Physìol.
(Lond.), 133,6Bi-694.
KQTTEGQDA, S. R. (1970). Perjstalsis of the small jntestine. IN:
Smooth Muscle. pp 525-541. (¡¿s. Buibrìng, E., Brading, A. F.,
Jones, A. ['l. and Tomi ta , T. ) Arno'l d, London.
KRANTIS, A. and KERR, D. I. B. (1981a). Auborad'iographic localization of
[3H1 gur,na-aminobutyric ac'id in the myenteric plexus of the guìnea-pìg
snral I intestine. Neurosci . l-etters , 23 ' 263"268.
KRANTIS, A. and KERR, D. I. B. (1981b). GABA jnduced excitatory responses
in the guinea-pig small intestjne are antagon"ized by bicucullìne,
p'icrotoxinjn and chloride ion blockers. Naunyn Schmiedeberg's Arch.
Pharmacol . (in press).

138
KRANTIS,4., COSTA, M., FURNESS, J. B. and ORBACH, J. (1980). Y-aminobutyric
acid stimulates intrinsic inh'ib'itory and excitatory nerves in the guinea-
pig intestine. Eur. J. Pharmacol ., 67, 461-468.
KRNJEVIó, K. (1974). Chemical nature of synaptic transmìssion in
vertebrates. Physìoì. Rev., 54' 418-540.
KRNJEVIó, K. and PHILLIS, J. l,J.
cerebral cortical neurones.
(1963). Act'ions of certain amjnes on
Brit. J" Pharmacol ., 2A, 47I-490.
f..UFtLËR, S. l^l. and EDI,JARDS, C. (i958). Mechanisnr of gamma-amìnobutyric
acid (GABA) aqd its relation to synaptìc inhibition. J. Neurophys'iol.,
2i, 589-610.
KUFFLER, S. hl. and NICHQLLS, J. G. (1966). The physìo1ogy of neurog'l'ia1
cells. Ergebn. PhYsiol., 57, 1-90.
KURIYAMA, K., HARBER, B.SISKENS, B. and ROBERTS, E. (1966). The v-
aminobuty¡ic acid system jn rabbit cerebellum. Proc. Natl. Acad. Scì.,
55,846-852.
KURIYAMA, H., 0SA, T. and TOIDA, T. (1967). Nervous factors ìnfìuenciitg
the membrane activ'ity of intest'inal smooth tnuscle. J. Phys'iol . (t-ond. )'
l9r,257-270.
KURODA, Y. and McILI^JAIN, H. (i974). Uptake and release of [3H] adenine
clerivatjves at beds of mamnralian cortjcal synaptosomes in a superfusjon
system. J. Neurochem., 22, 691-699.

139
KVAMME, E. (1979). Reguìation of glutaminase and ìts possible implicat'iorr
for GABA metabol ism. Ill: GABA-Bìochemistry anci CllS Functjons. Vol .
L23, pp 111-138. (fOs. Marrdeì, P. and De Feudis, F. V. ) Aclvances in
Experimenta'l Med'ic'ine and B"ioìogy. Plenum Pness, New York.
LANGLEY, J. N. (1898). The inhjbitory fibres'in the vagus for the end
of the oesophagus and the stomach. J. Phys'io1. (Lond.), 23 , 407-4L4
LEI-INiNGER, A. L. (1972) . Bi ochem j stry: The mol ecul ar basi s of cel I
structure and function. Worth Publ'ishers, New York.
LEVY, R. A. (L977,). The role of GABA'in primary afferent depolarization.
Prog. Neurob'ioì., 9, 21L-267.
I-EVY, l,l. 8., REDBURN, D. A. and COTMAN, C. hj. (1973). Stimulus coup'led
secretjon of y-anìinobutyric acid fro¡n rat brain synaptosomes.
Sc i er¡ce , 181 , 676-678 .
LEl,lIS, C. P., I'ÍcMARTIN, C., ROSENTHAL, S. R. and YATES, C. (1972).
Isolation and identification of pharmacologicalìy active amino ac'icis
in skin and their structure-activìty relat'ionship on the guinea-pig
ileum. Brit. J. Pharmacol ., 45, I04-t1.7.
LJUNTDAHL, A. , Uöffe tt, t. and HALASZ, N. (1.975). Atternpts to local ize
amino acid neurotransmitter suspects v¡jth autoradìography. Proc. 10th
FEBS Meet'ing Parìs. Bio'lo'igcal Membranes: Neurochernistry, 4!, 95-102.
LUNDBERG, J., AHLMAN, H., DALSTROM, A. and KEWENTER, J" (1976). Nerve
fibres in the human abdorninal vagus. Gastroenterology, 70, 47'2-474.

140
MACKTNZIE, I. and BURNSTOCK, G. (i980). Ev'idence against vasoactive
'intesti nal po'lypeptì de bei ng the non-adrenergi c , non-cho'l i nerg'ic
inhibitory transmitten released from nerves supplying the smocrth
muscle of the muscle of the guìnea-pig taenìa coli. Eur. J. Pharmacol.,
67, 255-264.
MALCHIKOVA, t.. S. and FOSKONOVA, M. A. (1971). Study of t'he mechanism of
the inhibitory ìnfluence of adenosine triphosphate on tire smooth muscles
of the gu'inea-pig. J" Evol . Biochem. Phys'ioi., .2, 294-302.
MARTIN, D. L. (1976). Carrier-mediated transport anC removal of GABA fronr
synapti c reg'ions . IN: GABA i r¡ Nervous System Funct'ion. pp 347-386 .
(E¿s. Roberts, E., Chase, T. N" and Tower, D. B. ) Raven Press, I'lew York"
MIHICH, E., CLARK, D. A. and PHILLIPS, F. S. (1954). Effects of aderrosine
anal ogs on 'i sol ated 'intest'ine and uterus . J . Pharmacol . Exp. Ther. ,
111_, 335-342 .
MILLER, J. J. anC McLEI'lliAN, H. (1974). The action of bicuculline upon
acetylchol ìne-i¡ciuced excitations of central neurones. Neuropharmacoic;gy,
13,785-787.
MIKI, Y., TANIYAþIA, l(., TANAKA, C. and TOBE, T. (1981), Localization of
GABA and GAD in Auerbach's plexus jn human colon. Proc. Ëighth Int.
Cong" Pharm., 1564P.
I4INCHIN, M. C. l,I. (1979). Sorne properties of protoveratríne-inCuced
.¡-aminobutyate release from rat braìn slices. Biochem" Soc. Trans.,
7, 139-140.

141
MINCHIN, M. C. and IVERSEN, L. L. (Lg74). Release of [3H] gut*u-
aminobutyric acid from g'lial cells in rat dorsal root ganglia. J
Neurochem., 23, 533-540.
MIYATA, Y. and 0TSUKA, M. (19i5). Quant'itative histochemistry of y-
aminobutyric acid in cat spìna1 c<¡rd wjth speciaì reference to
presynaptic inhibitjon. J. Neurochem., 25, 239-244.
uöHlfH, U. antl 0KAD0, T. (Ig77). GABA receptor binding with 3H (+)
bicuc.ulljne-methiodile in rat CNS. Nature (Lond.), 267,65-67.
MONT0REANO, R., RABIT0, C. A. and VILI-AMIL, ful. t. (1975). Effect of
furosemide on the permeabjl'ity to Cl of the isolated skin of Leptodaciyles
ocellatus. Experìentia, _31, 1305-ß47.
MORITûKI, H., TAKEI, M., KASAI, T., MATSUI4URA, Y. and ISHiDA, Y. (1979).
Possible involvement of prostaglandins in the act'ion of ATP on guinea-
pig uterus. J. Pharmacol . Exp. Ther., ?-I'I (1), 104-1.11.
McFARLAND, S.4., GUYT0N, C.A. and PFAFFMAN, M. A. (1971). Effects of
theophyìline on the taenia coli. Arch. 'int. Pharmacodyn., 192, 179-IB7 "
I4cSWINEY, B. A. ancl ROBSON, J. H. (1929). The response of smooth muscle
to stimulation of the vagus nerve" J. Phys'io1 . (Lond.), 68, 1,24-t3I.
NEAL, M. J. (1971). The uptake of [14c] giy.ine by slices of mantmalian
spinal cord. J. Physiol. (Lond.), ?15, 103-117.
NtAL, M. J. and IVERSEN, L. L. (i969). Subcellular distribution of
endogenous and [3H:] v-urirrobutyric ac'icl 'in rat cerebral cortex.
J. Neurochem., 16, I245-L252.

L42
NICKSRSSN, M. (1970). IN: The Pharmacological Basis of Therapeutjcs.
(f¿s. Goodman, L. S. and Gilman, A.) Maxmillan, New York.
NICSLL, p.. A. (1978). The blockade of GABA medjatêd response ìn the frog
spinal cord by ammonìum ions and furosemide" J. Physiol., 283, |ZI-I3Z"
NIELSEN, K. 1., OI^JMAN, C. and SAN'ì'INI, M. (1969). Anastomos'ing adrenerg'ic
nerves from the syrnpathetic trunk to the vagus at the cervical level jn
the cat. Brain Res., 12' 1-9.
NISI-II, S. and NORTH, R. A. (1973). lntracellular recording from the
myenteric plexus of the guinea'pig 'i1eum. J. Physio'l . ([.ond "), 23I,
47L-49t.
NISHI, S., MINQTA, S. and IGRCZI{AR, A. G. (I974). Prirnary afferents
neurones: the ionic mechanism of GABA-r¡erliated depolarization.
l{europharmacologY, 13, ?15-219.
NQRTH, R. A. and NISHI, S. (1974). Properties of the gangiion cells of
the myenteric pìexus of the gu'inea-pìg ìieum determjned by intra-
cel I u j ar recor.di ng. IN: Proc . 4th Int. Symp. Gastro j ntestì nal Mot'i1 ì t'y,
pp 667-676. Mìtcheì.l, Vancouver.
NORTH, R. A. and HENDERSQN, G. (1975). Actjon of morph'ine on guìnea-p'ig
myenteric plexus neurones studied by 'intracelIular recording. Life
Science, 17, 63-66.
NoRTH, R.4., HINDERSON, G., IGTAYAI''IA, Y" and JOHNS0N, S. M. (1980)'
Electnophysiolog'ica1 evidence of presynapb'ic inhibition of acetyl-
choline release by 5-hydroxytryptamine in the enteric nervous system.
Neurosciences 5, 581-586.

143
NORTH, R.4., KATAYAMA, Y. and t,.llLLIAMS, J.T. (i9i9). 0n mechanism
and site of action of enkephalin on single myenteric neurones. Brain
Res", L65, 67-77.
NORTH, R. A" and IIILLIAMS, J. T. (1976). Enkephalin 'inhibits f ixing of
myenteric neurones. Nature (Lond.), 264, 460-461.
NgRTHWAY, M. and BURKS, T. (1980). Stimulabjon of cholinergìc nerves in
dog 'intestine by ader¡ine nucleotides. Eur. J. Phartnacol ., 65 (1),
1 1- 19.
0BATA, K. (1976)., Exciiatory effects of GABA. IN: GABA jn Nervous System
Function, pp 283-286. (fas. Roberts, E., Chase, T. N. and Tower, D. B.)
Raven Press, New York.
CBATA, l(. ancl TAKEDA, K. (1969). Release of GABA into the fourth ventricle
i nduced by stirnu'l at'ion of the cat cerebel I um. J . Neurochem. , ,16 ,
1043-1447.
0BATA, K., IT0, M.,OCHI, R. and SAT0, N. (1967). Pharrnacologicaì
piopertÌes of the postsynaptic inhibition of Purk'inje cell axons and
the action of 'y-amìnobutyric acid on Deìter's neurones. Exp. Brain
Res. , 4, 43-57 .
0BATA, K., OTSUf';q, M. and TANAKA, Y. (i970). Deternrination of gamma-
amìnobutyrjc acid in sing'le nerve celIs of cat central nervous system.
J. Neuroclrem., 17, 697-698.

144
OHGA, A. and TANEIKE, T. (1977). Dissim'i'larity between the responses to
adenosine triphosphate or its related compounds and non-adrenergic
inhibitory nerve st'imujation in the longìtudinal smooth muscle of pig
stomach. Brit. J. Pharmacol., 60, 22I-23I.
OKbiUASABA, F. K., HAMILTON, J. T. and C00K, M. A. (1977). Relaxations of
guinea-pig fundic str ip by adenosine, adenìne nucleotides ancl electrical
stimulation: antagon'ism by theophy'ìline and desensitization to adenosine
and its der ivat'ives. Eur. J. Pharmacol",46, 181-198.
OLSEN, R. l,.I., BAN, [1. and MILLËR, R. (1976). Studies on the neuropharmaco'ìogica'l
activity of b'i,cuculline and related compounds. Brain Res., 10?, 283-299.
0LSEN, R. 1.1., TICKU, M. K., VAN NESS, P. C. and GREENLEE, D. (1978).
Effects of drugs on y-aminobutyric acicl receptors, uptake, reìease and
synthesì s 'í,n uitro. Brain Res. , I39, 277-294.
0NG, J. (1931). Baclofen: Some effects in guinea-pig snraìl and ìarge
intestine. Proc. Aust. Phys'io1 . Pharmacol . Soc., 12 (2), 114P.
ORREGO, F. (1979)" Criteria for identification of centrai neurotransmjtters
and their appìication to studies with some nerve tissue pneparations in
uitro. Neuroscience, 4, 1037-1057.
ORREGO, F., MiRANDA, R. and SALDATE, C. (1976). Electricaìly induced
release of labelled taurine, cx and ß-alanine, gìycine, gìutamate and
ot.her amiiro acids from rat neocortical sl ices in utitt,o. Neurosc'ience,
l, 325-332.

L45
PATON, l^l. D. M. (1957). The actìon of morphine and related substances on
contractjon and on acety'lchol jne output of coaxia:ìly stìmulated guìnea-
pig ileum. Brit. J. Phar^macol . Chemother., 12, IL9-L22.
PATON, l,J. D. M. and VIZi, E. S. (1969). The inhibitory action of noradrenal jne
and adrenal'ine on acetylcholjne output by guìnea-pig longitudinal muscle
stri p. Bri t. J . Pliarmacol . , 35 , 10-28
PATON, l^J. D" M. and ZAIMIS, E. J. (1949). The pharmacological actions of
po'lymethyl ene b'istrimethyl ammonì um sal ts. Brit. J. Pharmacoi . , 4,
381-400.
PEARSË, A. G. E. (1960) . Histochem'istry, Theoret'ical ancl Applied. 2nd
Edition. Churchill . London.
PECK, E. J., Jr., SCHAEFFER, J. M. and CLARK, J. ll. (1976). In pursuit of
the GAtsA recepotr. IN: GABA in Nervous System Function. pp 3i9-336.
(fAs. Roberts,8., Chase, T. N. and Tower, D. B.) Raven Press, Netat York.
PULL, I. and l4cILl,lAiN, H" (1972). Adenine derivatjves as neurohumoraj
agents jn the brain: The guantities liberated on excitation of super-
perfused cerebra'l tissues. Biochem. J., 130, 975-98i.
RAPQPQRT, S. I. (i976). Blood-brain Barrier jn Physiology and Medicine.
Raven Press, New York.
RATTAN, S. and GQYAL, R. (1980). Evidence against purìnergic (-) nerves
in the vagul pathwaSr to the opossum lower esophageil-sphìncter.
Gastroenter oìogy , --/?, B9B-904.

146
RIBIERO, J. A. and I^IALKËR, .1. (1975). The effects of adenosine triphosphate
and adenosine d'iphosphate on transmiss'ion at the rat and frog neuro-
muscular junctions. Brit. J. Pharmacol ., 54, 2L3-2I8.
RIKIMARU,4., FUKUSHI, Y. and SUZUKI, T. (1971). The effects of irn'idazole
and phentolamine on the relaxant responses of guìnea-p'ig taen'ia col i
to transmural sùimulation and to adenos'ine triphosphate. Tohoku J.
Exp. t'îed., 105, 1.99-200.
R0BERTS, P. J. (I974). The release of amino acjds with proposed neuro-
transmitter function trom the cuneate and gracìle nucle'i of the rat
ín uiuo. tsrain Res., 67, 419-428.
ROBERTS, E., CHASE, T. N. and TOWER, D. B. (i976). GABA in Nervous
System Function. Raven Press, New York.
ROSKOSKI, R., RYAN, L. D. and DIECKt, F. J. D. (1974). y-aminobutyric
acid synthesized jn the o'lfactory nerve. Nature, 25I, 526-529.
RUBIN, R. P. (1970). The rc¡le of calcium in the release of neurotransmitter
substances and hornones. Pharmacol. Rev., 22 (3), 389-428.
SAf'ÁI , K. K. , HUMSON, D. L. and SHAPI R0 , R. ( 1979 ) . The effects of
adenosine and adenine nucleotides on the guinea-p'ig vas deferens:
evidence for existence of purinergic receptors. Life Sciences, 24"
1299- 1308.
SATCHËLL, D. G. and BURNSI-CCK, G. (197i). Qr¡antiia'itve stud'ies of the
release of purine conìpounds fol1owìng stimulation of non-adrenergìc
inhibitory nerves 'in the stomach. B'iochem. Pharmacol ., 20, 1694-1.697.

t47
SATCHELL, D., BURNSTOCK, G. and DANN, P. (i973). Antagonism of the effects
of pu¡inergic nerve stinrulation and exogenously applied ATP on the
guinea-p'ig taenia colì by 2-substituted jnidazolines and related
compounds. Eur. J. Pharmacol ,, 23, 264-269.
SAbJYNOK, .1. and JHAMANDAS, K. H. (i976). Inhib'ition of acetylcholine
rel ease from chol 'i nerç¡i c nerves by adenos'ine , adeni ne nucl eot'i des and
morphine: antagoni sm by theophyl I ine. J. Pharmacol . Ëxp. Ther . , I97-,
379-390.
SCHABADASCII , A. ( 1930) . Intrarnural e Nervengef I echte des Darmt"ohrs .
Z. Zellforsch Mìkroskop. Anat., 19, 320"385.
SCHOFIELD, G. C. (1968). Anatomy of muscular and neural tissues ìn the
alìmentary canaì. IN: Handbook of Phys'iology, SecÙ'ion 6, Vol . IV,
pp I579-16?7 . American Phys'iological Soc'iety.
SCHON, F. anci IVERSEN, L. L. (Ig72). Selective accumu]at'ion ot f 3Hl g,qAR
by sateìlite cells in rat cerebellar ín tíua. Brajn Res., 4?-, 503-507.
scHQN, F. and KEI-I.Y, J. S. (Ig74). The characterì zaij on of I3Hl eRsR
uptake i nto the satel 1 i te g'l i al cel I s of the sensor-v gang'l 'i a. Bra'in
Res., 66, 289-300.
SCHQN, F. and KELLY, J. S. (1975). Selective uptake of 3H ß-alanine by
gl ia: assoc'iation w'ith the 91ial uptake system for GABA. Brain Res. ,
86, 243-257.
SCH0TTEN, C. (1883). Veber die Oxydation des Piperìdins. Ber d'b chem.
Ges., 16, 643-649.

i48
SCH0USBE, A. (1979). Effects of GABA-analogues on the h'igh-affìnity
uptake of GABA in astrocytes jn primary cultures. IN: GABA-Biochemistry
and CNS Function. Adv. Exp. Med. Biol., Vol. 123,219-237.
SCHOUSBE, 4., TilORBEK, P., HERTZ, L. and KROGSGAARD-LARSEN, P. (1979).
Ëffects of GABA anaiogs of restricted conformatioh on GABA transporl
in astrocytes and brain cortex slices and on GABA r'eceptor bindìng.
J. Neurochem., 33 (i), 181-i89.
SCHULTZBERG, I'f ., HOKF'ËLT, T., NILSSON, G., TERENIUS, 1., REHFELD, J. F.,
BP.oWN, M., ELDt, R. , G0LDSTEII'1, M. and SAID, S. (1980)" Distributi<¡n of
pepti de- and catechol ami ne-conta'ini ng neurones 'in the gastro'intest j nal
tract of rat and guìnea-píg: jnmunohistochemical siudies with ant'isera to
s¡bstance P, vasoact'ive'intestinal poìypeptide, enkephalins, somatostatin,
gastrin/cholecystokin'in, neurotensìn and dopamìne B-hydroxyl ase.
Neuroscjence, 5 " 689-744,
SCHULTZ, R. and CARTWRiGIIT, C. (1974). Effect of morphìne on serotonin
release from myenteric pìexus of the guinea-p'ig. J. Pharmacol. Exp.
Ther., 190 (3), 420-430.
SllANK, R. P., P0NG, 5. F., FREEMAN, A. R. and GRAHAM, L. T. (19i4).
Bicuculline and pÍcrotoxin as antagonists of 'y-aminobutyrate and
neuromuscular inhibition of the lobster. Brain Res., 72, 77-78'
SILINSKY, t. M. and HUIIBARD, J. I. (1973). Release of ATP frorn rat'motor
nerve termi nal s . Natut'e , 2t+3 , 404-405,
\
SNYDER, S. H. and GOODI",AN, R" R. (1.980) . Mul ti pì e neurotransmi tter receptors
J. Neurochem., 35, (1), 5-15.

149
SNYDER, S. H., KUHAR, M. J., GREEN, A. I", C$YLE, J. T. and SHASKAN' E. G.
(1970). Uptake and subcellular localizaticn of neurotransmitters'in the
brain. Int. Rev. Neurobiol., 13, I27-I58.
SPEDDING, M. , Sl,,lEETf",AN, A. J. and WEETMAN , D. F. (1975) . Antagoni sm of
adenosine 5'-triphospha.te induced re1axation by 2-2' -pyridylisatogen in
the taenja of the guinea-pig caecum. Brit. J" Pharmacol ., -53, 305-310.
SRINIVASAN, V., NEAL, M. J. anrl MITCHELL, J. F. (l.g0g). The effect of
electrical st'imulation and h'igh potassium concentrations on the efflux)
of ['H] y-aminobutyric acid from brain slices. J. Neurochem., 19,
1235-7244.
STACH, bl. (197i). ÜUer ¿ie in der Djckdarmwand aszenclievenden Nerven des
Plexus pe]vinus und dje Grenze der vagalen und sakralparasympathischen
Innervatjon. Z, Mikrosk-anat. Forsch., Leipzig, 84., 65-90.
su, c., BEVA.N, J. and tsURIìSTOCK, G. (1971). [3H] adenosine: release rluring
stimulation of enteric nerves. Science' 173, 337-339
SVENNEBY, G. and ROBËRTS, E. (1973). Bicuculline and N-methylbjcuculline-
compet'itive inh'ibjtors of brain acety'lcholìnesterase. J. Neurochem.,
2t, t025-i026.
szÉli, J. , MOLiNA, E., ZAPPIA, L. and BERTACINNi, G. (.1977). Act'ion of
some natural polypeptides on the longitud'inaì nluscle of the guìnea-
pig ileurn. Eur. J. Pharnlacol ., 43, 285-287.
SZER.B, J. C. (1976). Storage and release of label'led acetylchol'ine ìn the
myenteri c pi exus of the gui nea-pì g i 1eunr. Can . J. Physi o'l . Phar¡racol . ,
54 , L2-22,

150
2+SZERB, J. C. ( 1979) . Rel ati onsh'ip betrveen Ca -dependent and inCependent
release of t3Hl GABA evoked by h'igh K.+, \,eratridìne or electrical
stimula'['ion from rat cort'ical slices. J. I'leurochem.,32, 1565-1573.
SZ6LCSANYI, J.and BARTHO. L. (1979). New t,vpe of nerve-medjated choljnergic
contracti on of the gu'inea-pì g smal l i ntest'ine ancl i ts sel ecti ve bl ockade
by capsa'i ci n. Naunyn Schmj edeberg' s Arch. Pharmacol . ' 305 ' 83-90.
TAKAHASHI , H. , UCHIKURA, K. , TAKAHASHI, ll. and IKEDA, 0' (1.961a).
Relat'ionshjp betvreen pharmacologìcai actions on the mammalian ileum and
chem'i cal structure of y-ami nobutyrí c aci d. Jap. J . Physi ol . , _1_1.'
?29-237.
TAKAHASHI " l-1. , IKEDA, 0. , ITAYA, T. and KOSHIN0, F" (1961b). Antj -
stinlulant actions of y-amìnobutyric acid and its derivatjves on the
gu'inea-p'ig ileum. Jap.J. Physìol ., 11, 476-186.
TAKES, 0. ancl SUGAI, N. (1974). Morpholoç¡y of extragangl jon'ic flourescent
neurones in the myentenìc plexus of the small jntestine of the rat.
J. Comp. Neurol . , l-58, 109-120.
TALLARiDA, R. ,1., COl^lAN, A. and MARTIN, I'1. A. (1979). pAt and receptor
differentiatiorr: a statjstical ana'lysis of competÌtive antagonìsm.
Life. Sci. , 25, 637-654.
TAPIA, R. (1974) . The role of y-aminobutyric acid nretaiiol'ism jn l.he
regul at'ion of cerebral exci tab'i'l i ty. IN: Neurohutnoral Codìng of Brai n
Funct'ion. pp 3-26. (Eds. Myers, R. D. and DrLlckr:r-Colin, R.. R. ) Plenum
Press, New Ycrk.

151
TAXI, J. (1965). Contribution a I'etude des connexions des neurones
moteurs du systéme nerveux autonome. Amer. Sc'i . t{at. Zool . ' 12th Ser. 7,
413-67 4 .
THAEMERT, J. C. (1963). The ultra-structure ancl disposjtion of vesiculated
nerve processes 'in smooth muscle. J. Cell. Bjol ., Ui, 36I-377.
TQMiTA, T. and I^IATANABE, H. (1973) . A compari son of the effects of
adenoi s i ne tri phophate wi th noradrenal i ne and wi th the i n hi bì tory
potential of the guinea-pig taenaì col'i. J. Physiol. , 231 ' 767-I77.
TRENDELENBURG, P., (1917). Physioìogische L¡nd pharmacko'logische versuche
über die Dünndarmperistaltik. Naunyn Schmiedeberg's Arch. exp. Path.
Pharmak., 81 , 55-L29.
TRIGGLE, C. R. and TRIGGLE, D. J. (1976). Structure and function of the
synapse. IN: Chemical Fharmacology of the Synapse. pp 1-128.
(Eds. Trjggle, D. J. and Triggle, C. R.) Acadenjc Press, London"
VAN DTN BERG, C. J., MATHESON, D. F., RONDA, G., RETJNIERSE, G. L.4.,
BLOKHUIS, G. G. D., KR00N, M. C., CLARKE, D" D. and GARFtrNKtL, D. (1975).
A model of g'lutamate rnetabol i sm i n brai n: A biochelnì cal analysi s of
heterogenous structure. IN: Metabolìc Compartlnentation and Neuro-
transmiss'ion. pp 515-543. (Eds. Berl, S., Clarke, D. D. and
Schrreider, D,) Plenum Press, New York.
VAN NUETEN, J", VAN REE, J. M. and VANHOUTTE, P. M. (1977). Inhibjtion
by met-enkephalin of peristalt'ic act'ivity in ihe guinea-pig iIeum,
and 'its reversal by nalaxone. tur. J. Pharmacol ,, fi, 34i-342,

752
VIZI , E. S" ancl KNCLL, J. (1976). The inhibìtory effect of adenos'ine and
related nucletrtìdes on the release of acetylcholine. Neuroscience,1,
391 -398.
IIAKADE, A. R. and WAKADE, T. D. (1978). Inhibition of ttoradrenaline
release by ac.lenosìne. J. Physioì. (Lond .), 282, 35-49.
IIALKER, R. J. , CROS.SMAN, A. R. , bJOODRUFF, G. N. and KERKUT, G. N. (1971).
The effect of bicucr"¡lline on the gamma-am'inobut.yric acjd (GABA) receptors
of neurones of perip'laneta Amerjcana and helix aspersa. Brain Res",
33,75-82.
WALLACH, D.P. (196i). Studies on the GABA pathway - I: The ìnhibitjon
of y-anri nobutyri c aci d-o ketogl ut.ar j c aci d transami nase i.rt uil;v,o and
in uiuo by U-7524 (amino-oxyacetic acid). Biochem; Pharmacol., 5o
323-331.
hJATERFItLD, A" A., SMOCKUM, R. þJ. J., HUGHES, S. J. ' KOSTERLITZ, H. l^'l. and
HENDERSON, G. (1977). rn uitro pharmacology of the opioid peptides,
enkephaf ins and endorphins. Eur" J. Pharmacol., 43, 107-116.
hJATSON, l.l. E. (I974). Physiology of Neurogl'ia. Physìol . F.ev. , 54,, 245-?.71.
hIEETMAN, D. F. and TURNER, N. (1974). The effects of inhjbitors of adenosine
5'-triphosphate biosynthes'is on the responses of the guìnea-pig isolated
bladder and some other preparatìons to nerve stir¡ulat'ion. Arch. "Int.
Pharmacodyn. Ther., 210, 12-26.
WERMAN, R. (1966). A review - criter"ia for jdentification of a central
nervous systeni transmjtter. Comp. Biochem. Physiol ., -18, 745-766.

153
1¡ËSTFALL, T. C. (I977), Local regulat'ion of adrenergic neurott ansmission.
Physiol. Rev. , 57 , 659-728.
!\lHITE, A. , HANDLER, P. and SMITH, E. L. (1968). Princ'ip'les of b'iochemistry.
McGraw-Hill, New York.
t^lILLIAMS, J. T" and N0RTH, A. (1979). Vasoactive intestinal poìypeptìde
excites neurones of the nryenteric p'lexus. Brain Res. , 175, 174-177 .
¡r1ILSSN, J. E. (i976). Applicatjon of blue dextran and cibacron blue
F3GA in purification and structural s'bud'ies of nucleotjde-requ'iring
enzyrnes. Biochem. Biophys. Res. Commun., 72- (3) ' 816-823.
l^i00D, J. D. and MAYER, C. J. (1978). Intracellular study of electnical
act j v'ity of Auerbach' s pl exus 'in guì nea-pì g sma'l l i ntesti ne. Pf 1 ügers
Arch. Eur. J. Physiol ., i73, 265-275.
l^l00D, J. D. and MAYER, C. J. (I979) . Serotonergi c act.'ivat j on of toni c-type
enterj c neurones in gui nea-p'ig smal I bovlel . J. Neurophys io1 . ' 4?'
582-593.
¡¡00D, J. G., McLAUGHLIN, G. antt VAUGHN, J. E. (1976). Inrmunocytr¡chenrjcal
I ocal i zati on of GAD i n el ectron m'i croscop'ic preparati ons of rodent
CNS. IN: GABA in Nervous System Function. Kroc Foundation Series,
Vol. 5, pp 133-148. (Eds. Roberts, E., Chase, T. N" and Tower, D. B.)
Raven Press, New York.

154
l,lu, J.-Y. (1976). Purìfication, characterization and kinetic studies of
GAD and GABA-T fnom rnouse bra'in. IN: GABA in Nervous System Function.
Kroc Foundatjon Series, Vcl. 5, pp 7-55. (Eds. Roberts, E., Chase, T. N.
arrd Tower, D" B.) Raven Press, New York.
YAROIISKY, P. J. and CARPENTTR, D. 0. (1972). GABA mediated excitatory
responses on aplysia neurones. L'ife Sciences, 20, 1441-1445.
YAU, t^J. M. (1978) . Effect of substance P on i ntest'inal muscl e.
Gastroenterol goy, 7 4, 228-23I.
Y0UNG, J. A. C., BROhlN, D. 4., KELLY, J. S. and SCllON, F. (1973).
Autoradiographic local'ization of [3H] y-anrinobutyric acid accumulat'ion
in periphera'ì autonom'ic gangl'ia. Brain Res., 63,479-486.
ZEUTHEN, T., RAI']OS, M. and ELLORY, J. C. (i978). Inh'ibition of active
chloride transport by p'iretamide. Nature, 273, 678-680"
ZUKIN, S. R., YOUNG, A. B. and SNYDER, S. H. (1974). Gamma-arninobutyrìc
acid binding to receptor s'iles ìn :at central nervous system.
Proc. Nat. Acad. Scì. (U.S.A.),21,4802-4807.
i