functional evolution of jumping in frogs: interspecific ... · in this setup, an array of four...
TRANSCRIPT

Functional Evolution of Jumping in Frogs: InterspecificDifferences in Take-Off and Landing
Stephen M. Reilly,1* Stephane J. Montuelle,2 Andr�e Schmidt,2,3 Cornelia Krause,2
Emily Naylor,2,4 and Richard L. Essner Jr.5
1Department of Biological Sciences, Ohio University, Athens, Ohio2Department of Biomedical Sciences, Ohio University Heritage College of Osteopathic Medicine, Athens, Ohio3Orthopedic University Hospital Friedrichsheim, Frankfurt Initiative for Regenerative Medicine, Frankfurt, Germany4Department of Biology, University of California, Riverside, California5Department of Biological Sciences, Southern Illinois University Edwardsville, Edwardsville, Illinois
ABSTRACT Ancestral frogs underwent anatomicalshifts including elongation of the hindlimbs and pelvisand reduction of the tail and vertebral column that her-alded the transition to jumping as a primary mode oflocomotion. Jumping has been hypothesized to haveevolved in a step-wise fashion with basal frogs taking-offwith synchronous hindlimb extension and crash-landingon their bodies, and then their limbs move forward. Sub-sequently, frogs began to recycle the forelimbs forwardearlier in the jump to control landing. Frogs with fore-limb landing radiated into many forms, locomotormodes, habitats, and niches with controlled landingthought to improve escape behavior. While the biology oftake-off behavior has seen considerable study, interspe-cific comparisons of take-off and landing behavior arelimited. In order to understand the evolution of jumpingand controlled landing in frogs, data are needed on themovements of the limbs and body across an array oftaxa. Here, we present the first description and compari-son of kinematics of the hindlimbs, forelimbs and bodyduring take-off and landing in relation to ground reac-tion forces in four frog species spanning the frog phylog-eny. The goal of this study is to understand whatinterspecific differences reveal about the evolution oftake-off and controlled landing in frogs. We provide thefirst comparative description of the entire process of jump-ing in frogs. Statistical comparisons identify both homolo-gous behaviors and significant differences among speciesthat are used to map patterns of trait evolution and gener-ate hypotheses regarding the functional evolution of take-off and landing in frogs. J. Morphol. 277:379–393, 2016.VC 2015 Wiley Periodicals, Inc.
KEY WORDS: anurans; Ascaphus; Bombina; Lithobates;Rhinella; forces; locomotion
INTRODUCTION
Frog jumping is a unique form of locomotion,thought to have arisen in a step-wise manner from ageneralized tetrapod trotting gait (Abourachid andGreen, 1999; Essner et al., 2010). Anatomical shiftsincluding the elongation of the hindlimbs and pelvisand reduction of the tail and vertebral column her-alded a transition to jumping as a primary mode oflocomotion with subsequent adaptations enhancing
jumping performance (Noble, 1931; Griffiths, 1963;Gans and Parsons, 1966; Zug, 1985; Emerson, 1979,1985; Carroll and Holmes, 2007; Handrigan andWassersug, 2007). The evolution of jumping involvesnot only changes in the take-off behavior but in thelanding as well. The ancestral landing behavior offrogs is hypothesized to be represented by the Leio-pelmatidae (the genera Ascaphus and Leiopelma),which crash-land on their bodies and then cycle theirlimbs forward and back to their resting position(Essner et al., 2010). Subsequently, as is found in therest of the living anurans (Lalagobatrachia), frogsappear to have made the important shift to protract-ing and adducting the forelimbs earlier in the jumpin order to use them to control landing (Peters et al.,1996; Essner et al., 2010; Gillis et al., 2014; Cox andGillis, 2015). Frogs using forelimb landing have radi-ated into many forms, locomotor modes, habitats,
Additional supporting information may be found in the onlineversion of this article.
Contract grant sponsor: the Baker Fund and the Faculty Fellow-ship Leave Program at Ohio University (SMR); Contract grantsponsor: Faculty Research Funds from Southern Illinois Universityat Edwardsville (RLE); Contract grant sponsor: the National Sci-ence Foundation (the Ohio Board of Regents, the Ohio UniversityOffice of the Vice President for Research, and the Ohio UniversityHeritage College of Osteopathic Medicine (SJM)); Grant number:MRI DBI-0922988.
*Correspondence to: Stephen M. Reilly; Department of BiologicalSciences, Ohio University, Athens, OH. E-mail: [email protected]
Author Contributions: SMR, SJM, RLE performed all of theresearch and data analysis and the majority of writing and studydesign. AS, CK, and EN assisted in experimental analyses and ENassisted in data extraction and analysis. All authors provided com-ments on the manuscript.
Received 11 September 2015; Revised 16 October 2015;Accepted 20 November 2015.
Published online 29 December 2015 inWiley Online Library (wileyonlinelibrary.com).DOI 10.1002/jmor.20504
VC 2015 WILEY PERIODICALS, INC.
JOURNAL OF MORPHOLOGY 277:379–393 (2016)

and niches (Gomes et al., 2009; Jorgensen and Reilly,2013) with controlled landing facilitating rapidescape behavior by resetting the limbs for anotherjump (Gans and Parsons, 1966; Emerson, 1978; Ess-ner et al., 2010).
The physiology, biomechanics, and muscle func-tion of the frog hindlimb during take-off have beenthe focus of numerous publications in the last 40years (reviewed in Duellman and Trueb, 1994;Marsh, 1994; Wells, 2010). However, features oflanding behavior, when noted, are usually limited torecording the impact time of the hands and whenthe animal is back to its sitting position (e.g., Peterset al., 1996). Several studies of the landing per sehave appeared more recently. The motor functionand biomechanics of the limbs during landing havebeen studied primarily in the marine toad Rhinellamarina of Hyloidea (Gillis et al., 2010; Akella andGillis, 2011; Azizi and Abbott, 2013; Griep et al.,2013; Azizi, 2014; Azizi et al., 2014; Gillis et al.,2014; Cox and Gillis, 2015; Reilly et al., 2015a). Inaddition, patterns of landing forces have beenreported for two ranoid species (Rana esculenta:Nauwelaerts and Aerts, 2003; Rana dybowski: Wanget al., 2014). In terms of comparative studies of land-ing kinematics, only one study (Essner et al., 2010)compared the relative timing of forelimb and hind-limb recovery in three taxa including Ascaphus, aleiopelmatid and two lalagobatrachians, Bombina(Bombinatoridae) and Lithobates (Ranidae).
In order to understand the evolution of jumpingand controlled landing in frogs, data are needed tocompare the movements of the forelimbs, hindlimbsand body during take-off and landing across anarray of model taxa. Comparisons across specieslead to many questions. When do the forelimbs andhindlimbs begin to recycle forward and when do theyhit the ground? What parts of the body contact theground during landing and what is the role of thefeet in landing? How do body and limb kinematicsrelate to each other and to landing forces within agiven species? More importantly, what do interspe-cific differences in the kinematics of landing acrosstaxa spanning the anuran phylogeny reveal aboutthe evolution of take-off and controlled landing infrogs?
The goal of this study is to describe take-off andlanding behaviors and their relation to ground reac-tion forces in four model frogs chosen to representkey terrestrial taxa spanning the anuran phylogeny(illustrated in Fig. 5, below). The Rocky MountainTailed frog, Ascaphus montanus, was chosen to rep-resent the ancestral belly-flopping form of landing(Essner et al., 2010). The fire-bellied toad, Bombinaorientalis, was selected as a basal lalagobatrachianspecies that represents one of the first clades to useforelimb landing (Essner et al., 2010). The bullfrog,Lithobates catesbeiana, and the marine toad, R.marina, were chosen as representatives of the twomajor groups of the Neobatrachia (Ranoidea and
Hyloidea, respectively) that typically use the fore-limbs to control landing (Nauwelaerts and Aerts,2003; Griep et al., 2013; Gillis et al., 2014; Cox andGillis, 2015). They provide comparative data formore derived jumping taxa and complement themany studies of jumping using bufonid and ranidfrogs as the primary research models (Gillis et al.,2014; Robovska-Havelkova et al., 2014).
In addition, we selected species of similar ecolog-ical modes of locomotion. Using the same ecologi-cal classifications as in Robovska-Havelkova et al.(2014), our study compares three semiaquatic spe-cies and one terrestrial species (Rhinella). We clas-sify Ascaphus as semiaquatic based on extensivefield observations that they jump on land morethan into water (Reilly et al., 2015b). We considerB. orientalis to be semiaquatic because of their ter-restrial diet (e.g., Kuzmin, 1990; Bisa et al., 2007).Therefore, by comparing ecologically similar spe-cies, the effects of ecology are minimized in ourstudy sample so that phylogenetic patterns may beobserved and discussed.
We recorded animals jumping using high-speedvideo while simultaneously recording ground reac-tion forces. Kinematic analyses were used to iden-tify and describe behavioral events in relation tothe magnitudes and timing of take-off and landingforces across the study taxa. These provide the firstcomparative descriptions of the complete jumpingbehavior of frogs. Statistical comparisons revealnovel homologous behaviors and significant differ-ences between species. Patterns of trait transitionsare mapped on a phylogeny in order to generate thefirst general hypotheses regarding the functionalevolution of take-off and landing in frogs.
MATERIAL AND METHODSStudy Species and Taxonomy
We follow Frost et al. (2006) in using the taxon name Leio-pelmatidae to include the sister taxa Ascaphus and Leiopelma.A. montanus (Mittleman and Myers, 1949) (n 5 3; snout-ventlength [SVL] 5 4.33 6 0.01 cm; Mass 5 6.2 6 0.1 g) were col-lected from Beauty Creek, Coeur d’Alene National Forest,Idaho (47836.390N; 116840.130W), under permits from the U.S.National Forest Service and Idaho Fish and Game (Permit #130502). The remaining species were obtained commerciallyand included B. orientalis (Boulenger, 1890) (n 5 3; 4.27 6
0.01 cm; 4.9 6 0.4g), L. catesbeiana (Shaw, 1802) (n 5 3;14.70 6 0.23 cm; 317.0 6 13.7 g), and R. marina (Linnaeus,1758) (n 5 3; 13.61 6 0.12 cm; 255.0 6 17.9 g).
Experimental Setups
Frogs were induced to take-off and land on force plates whilebeing recorded with high-speed video. Due to differences in sizeand mass among the species, two force plate setups were used.For the two smaller species (Ascaphus and Bombina), groundreaction forces were recorded at 5000 Hz as they jumped betweentwo 0.15 3 0.15 m force plates (AMTI MC3A-100) using LabVIEW2013 (National Instruments). An AOS X-PRI high-speed camera(AOS Technologies AG, Switzerland) was synchronized at 250frames per second (fps) in lateral view. For the two larger species(Lithobates and Rhinella), ground reaction forces were recorded
380 S.M. REILLY ET AL.
Journal of Morphology

at 500 Hz as they took-off and landed on a single 1.1 3 0.33m forceplate (Bertec Quadfit) synchronized with an Oqus 31 high-speedcamera recording at 200 fps in lateral view (Qualisys Motion Cap-ture Systems, Sweden). In this setup, an array of four synchron-ized Oqus 31 high-speed cameras (200 fps) were calibrated torecord the 3D position of an infrared reflective landmark glued onthe occiput and sacrum of the Lithobates and Rhinella subjects.Videos, landmark position, and forces of Rhinella and Lithobateswere recorded using Qualisys Track Manager (Qualisys MotionCapture Systems, Sweden). Prior to any analyses, the 3D positionof the infrared reflective landmarks was filtered using a movingaverage over five frames.
Force plates had rough surfaces for traction and were zeroedat the beginning of each recording session. During a recordingsession, frogs were placed on a starting position on the forceplate with their snouts pointing down the longitudinal axis ofthe single or double plate setup. They either jumped spontane-ously after several seconds or were induced to jump byapproach of the hand or sometimes by gently tapping the poste-rior of the body. Because of differences in thermal preferences(Essner et al., 2010), recording sessions were carried out at158C for Ascaphus and at 20–228C for the three other species.From numerous recordings reviewed, we selected five to sevenjump sequences per individual (from take-off through landing)that (1) were parallel with the long axis of the force plate and(2) where the hindlimbs were used in synchrony, so that oppos-ing mediolateral forces cancel each other. All analyses includethe selected take-offs and their subsequent landings.
Take-Off Kinematic and Force Variables
The review of video recordings of take-offs revealed keybehavioral events universally observed in all four frog speciesand others that were unique. The timing of each of these eventswas digitized for each jump using VirtualDub (http://www.vir-tualdub.org/index.html). The jump began with the onset ofhindlimb extension (start hindlimb extension, Time 0). Thiswas recorded as the frame before any noticeable movementoccurred between the knee and the body and was coordinatedwith the time that vertical and fore-aft forces began to changefrom body weight (BW) and zero, respectively. Prior to the onsetof visible hindlimb extension movements, slight but obviouspostural or head movements or tensing of the body wereobserved indicating that the frogs were about to jump (time tostart moving). Time to hands-up was noted as the framewhen the last finger left the ground. Take-off time was thetime when the last toe left the ground, which correspondedwith both the vertical and fore-aft forces reached zero.
Take-off velocity was calculated as the velocity of the headat the instant of take-off. In Rhinella and Lithobates, headvelocity was calculated in Qualisys Track Manager (QualisysMotion Capture System, Gothenburg, Sweden) from the filteredfirst derivative of the 3D displacement of the occiput landmarkover time. For Ascaphus and Bombina, take-off velocity wascalculated from the displacement of the head (snout) in lateralview digitized over five frames centered on the frame in whichtake-off occurred (Marsh and John-Alder, 1994).
Take-off angle was computed as the angle between the sac-rum, the tip of the last toe to leave the ground and the horizontalat the instant of take-off. The position of the sacrum was used toestimate the position of the center of mass. Jump distance wasthe longitudinal distance traveled by the occiput landmark inLithobates and Rhinella, and snout in Ascaphus and Bombinafrom the onset of take-off to toes-down (described below).
The maximum vertical and maximum fore-aft (accelera-tive) forces during take-off were extracted using SPIKE2(Cambridge Electronic Design, UK). To correct for differences inmass across species, force magnitudes were converted to BWs.The Peak Resultant Force angle was computed as the angleof the resultant force of the maximum vertical and fore-aftforces with the horizontal. Force timing variables (onset, offset,time to maximum) were extracted and Take-off duration was
defined as the difference in time between the onset and offsetof accelerative fore-aft forces.
Landing Kinematics and Force Variables
The review of video recordings of numerous landings revealeda series of key behavioral events that can be used to describeand compare landing behavior across the four species. Becausethese represent novel observations they are presented in theresults section. In short, landing started with substrate contact(hands-hit in the lalagobatrachians, body-hit in Ascaphus) andended with a stereotypical behavior of putting the longest toe-down which returned the frog to a resting position. Accordingly,behavioral landing period was defined as the time fromsubstrate contact (time zero, which corresponded to the onset offore-aft (braking) ground reaction forces) to toe-down. We thenidentified and digitized the times of homologous behavioralevents (e.g., body and feet impact times, hindlimb flexion eventsdescribed in the results section).
The onset and offset times and key force events characteriz-ing vertical and fore-aft ground reaction forces were digitizedfrom the force record of each trial. Review of the force profilesshowed that ground reaction forces started at impact, increasedto exhibit patterns of landing forces and then returned to staticlevels; that is, vertical forces returning to BW and fore-aftforces returning to zero (indicating the end of forward move-ment). The difference between the time of substrate contact (5the onset of braking fore-aft forces) and the time that the fore-aft forces return to 0 (henceforth, TFA0) was termed the forcelanding period. Initial comparisons showed that the relation-ship between behavioral landing period and force landingperiod differed across the species. In other words, the end ofthe behavioral landing (toe-down) did not necessarily corre-spond with the end of landing forces (TFA0) in all species.
In terms of force events, in Ascaphus we recorded the timingand magnitudes of peak body landing forces. In addition, thetime when the fore-aft force initially leveled off to low brakingforces after the main pulse of landing forces was digitized tomeasure the duration of the primary impact forces (Fig. 3). Inthe species using their hands to contact the substrate (i.e.,Bombina, Lithobates, and Rhinella), we digitized the timingand peak magnitudes of the forelimb and body landing forces.In Bombina and Lithobates, peak forelimb forces occurred atthe instant the body-hits, whereas Rhinella exhibited independ-ent forelimb and body peaks (Fig. 3). As in the take-off varia-bles, all landing force magnitudes were converted to BWs.
Data Normalization
Several studies have already reported significant effects ofbody size and jump distance in frogs, both intraspecifically andinterspecifically (e.g., Emerson, 1978; Zug, 1985; Marsh, 1994;Choi et al., 2000; Cox and Gillis, 2015). In our study sample,there was a threefold variation in SVL between the small andlarge species and all of the force, velocity, and angle variableswere significantly correlated with SVL except for one variable(maximum vertical force of forelimb landing). In addition, thesevariables were also significantly correlated with jump distanceexcept for the same variable. Therefore, to remove the effects ofbody size and jump distance, nontiming variables (except maxi-mum vertical force of forelimb landing) were adjusted by scalingthem to body-lengths jumped (jump distance in body lengths).After scaling, no positive correlations were found between thesevariables and SVL or body-lengths jumped. This normalizationallows us to compare variables across species as if they were alljumping one body length. As found in many other studies, phylo-genetic differences in morphology, physiology, and jumpingbehavior are likely to be even more important than size whencomparing distantly related species (Gomes et al., 2009; Jorgen-sen and Reilly, 2013).
Both take-off and force landing duration (TFA0) were also cor-related with jump distance. Thus, to allow the comparison of
381TAKE-OFF AND LANDING IN FROGS
Journal of Morphology
behavioral events to force dissipation patterns across species,take-off timings were scaled to take-off duration and landing tim-ings were scaled to percent of force landing period (TFA0). Table1 presents raw and adjusted take-off variables. Table 2 presentsraw and adjusted landing variables. All intraspecific and inter-specific statistical comparisons were done on the adjusted varia-bles (except maximum vertical force of forelimb landing).
Statistical Analyses
All statistical analyses were conducted using SPSS 13.0(IBM). Normality and homoscedasticity of all adjusted variableswere verified prior to analysis. Kinematic, timing, and forcesvariables were analyzed using a multivariate analysis of var-iance (MANOVA) with univariate F-tests to test for specieseffects (fixed factor) with individual effects (random factor)nested within species to account for repeated measures. Take-off and landing data sets were analyzed separately (Tables 1and 2). When significant differences between species weredetected by the MANOVA, Bonferroni’s post hoc tests wereused to determine significant differences between species.
Student’s t-tests were used to investigate the temporal corre-spondence between key kinematic and force events within spe-cies. In these comparisons, significance (P<0.05) indicates thatthe two events occurred at different times. Alternatively, a non-significant t-test (P>0.05) indicates that events occurredsimultaneously.
RESULTSInterspecific Comparisons of Take-OffBehavior
Take-off behavior and ground reaction force pro-files for each species are illustrated in Figure 1 andsupporting information Video S1-4. Statistical com-
parisons of take-off kinematic and force variablesare presented in Table 1. In our experimental set-ups, the frogs averaged jump distances of about2.5–5 body lengths (BL). In terms of behavioralevents during take-off, Ascaphus started movingsignificantly earlier than the other species. Time tohands-up ranged from 41 to 55% of take-off dura-tion, with only the extremes of the range being dif-ferent (i.e., Rhinella was significantly later thanLithobates).
Two major behavioral differences were evident intake-off videos (Fig. 1, supporting information Vid-eos S1-4). First, the onset of forelimb recovery dif-fered across the four species. After hands-up,Ascaphus usually extended its elbows andadpressed its humeri and often the entire forelimbsposteriorly to its sides. The forelimbs remained inthat position and did not start to move forward untilwell into the landing phase. Occasionally, the fore-limbs hung more laterally along the side of thebody. However, in the other three species, forelimbrecovery began during take-off (supporting informa-tion Videos S1-4). Rhinella started forelimb recov-ery earlier than the other species and all threespecies extended their hands under the neck bytake-off and under the snout by mid-flight.
The second major behavioral difference duringtake-off is that Ascaphus maintained full hindlimbextension throughout the flight phase and earlylanding, while the other three species started
TABLE 1. Timing, force, and performance variables during the take-off phase of frog jumping
Ascaphus Bombina Lithobates Rhinella
Jump distance (cm) 21.5 6 0.7 14.4 6 0.6 38.1 6 3.5 31.8 6 2.8Jump distance (BL) 4.97 6 0.16 3.39 6 0.15 2.59 6 0.23 2.34 6 0.22Take-off (TO) duration (ms) 108 6 3 120 6 3 182 6 18 176 6 6
Adj. Take-off duration (ms/BL J) 22.3 6 1.0 36.5 6 2.1 74.2 6 9.6 85.9 6 8.5Time started moving (% TO duration) 282.4 6 10.5 230.8 6 3.2 236.6 6 10.4 235.9 6 4.7Time to hands-up (% TO duration) 49.7 6 1.9 49.9 6 3.1 41.3 6 3.3 55.5 6 3.2
Time to maximum V force (% TO duration) 72.9 6 1.1 64.7 6 0.9 70.3 6 4.7 61.3 6 3.9Time to maximum FA force (% TO duration) 73.9 6 1.0 66.5 6 1.4 74.9 6 5.4 67.7 6 4.6Maximum V force (BW) 2.17 6 0.08 1.76 6 0.09 1.64 6 0.11 1.62 6 0.12
Adj. Maximum V force (BW/BL J) 0.44 6 0.01 0.53 6 0.03 0.67 6 0.06 0.75 6 0.06
Maximum FA (accelerative) force (BW) 2.81 6 0.10 1.75 6 0.11 0.88 6 0.07 0.96 6 0.07Adj. Max. FA (accelerative) force (BW/BL J) 0.58 6 0.02 0.52 6 0.03 0.36 6 0.04 0.46 6 0.06
Take-off angle (8) 27.8 1 0.8 31.3 1 1.0 52.8 1 0.8 56.3 1 0.7Adj. Take-off angle (8/BL J) 5.6 6 0.1 9.5 6 0.5 21.8 6 1.9 27.5 6 2.6
Peak Resultant force angle (8) 37.8 1 0.9 35.5 1 2.0 61.7 1 1.6 59.4 1 1.5Adj. Peak resultant force angle (8/BL J) 7.7 6 0.2 13.8 6 0.8 25.6 6 2.4 28.5 6 2.4
Take-off velocity (m/s) 0.88 6 0.01 1.03 6 0.02 1.16 6 0.07 1.09 6 0.07Adj. Take-off velocity (m/s/BL J) 0.17 6 0.01 0.31 6 0.01 0.47 6 0.05 0.49 6 0.02
Species means (6SEMs) are reported with underlining indicating means that are not statistically different based on the results of MAN-OVA on adjusted variables. Abbreviations: TO, take-off; BL, Body lengths; V, vertical; FA, fore-aft; BL J, body lengths jumped; BW, Bodyweights.
382 S.M. REILLY ET AL.
Journal of Morphology

hindlimb recovery immediately after take-off withhindlimb flexion well underway by midflight.
The shape and timing of take-off force profileswere similar across all four species (Fig. 1; Table1). Across the species studied, the vertical andfore-aft forces reached their maxima between 61and 75% of take-off duration (Table 1), but thesetimings were not significantly different among(MANOVA) or within species (t-tests: V vs. FAforces). In terms of magnitude, maximum verticalforces trended from low forces in Ascaphus and thehighest forces in Rhinella with Bombina and Lith-obates falling in between (Table 1). The fore-aftforces were greater than the vertical forces inAscaphus and were matched in magnitude inBombina. In contrast, in the neobatrachians, maxi-mum vertical forces were relatively greater thanthe maximum fore-aft forces (Fig. 1; Table 1).Accordingly, jumps of Lithobates and Rhinella werecharacterized by steeper force resultant anglesthan those of Bombina, which was significantlysteeper than Ascaphus. This pattern is paralleledby the take-off angle which was steeper in the Neo-batrachians than in the basal species. Finally, theadjusted take-off velocity shows that velocity waslow in Ascaphus, moderate in Bombina, and thehighest in the neobatrachian species (Lithobates
and Rhinella, Table 1). In summary, take-off wasslow and flat in Ascaphus, intermediate in Bomb-ina, and steep and fast in the neobatrachians.
Landing Behavior in Frogs
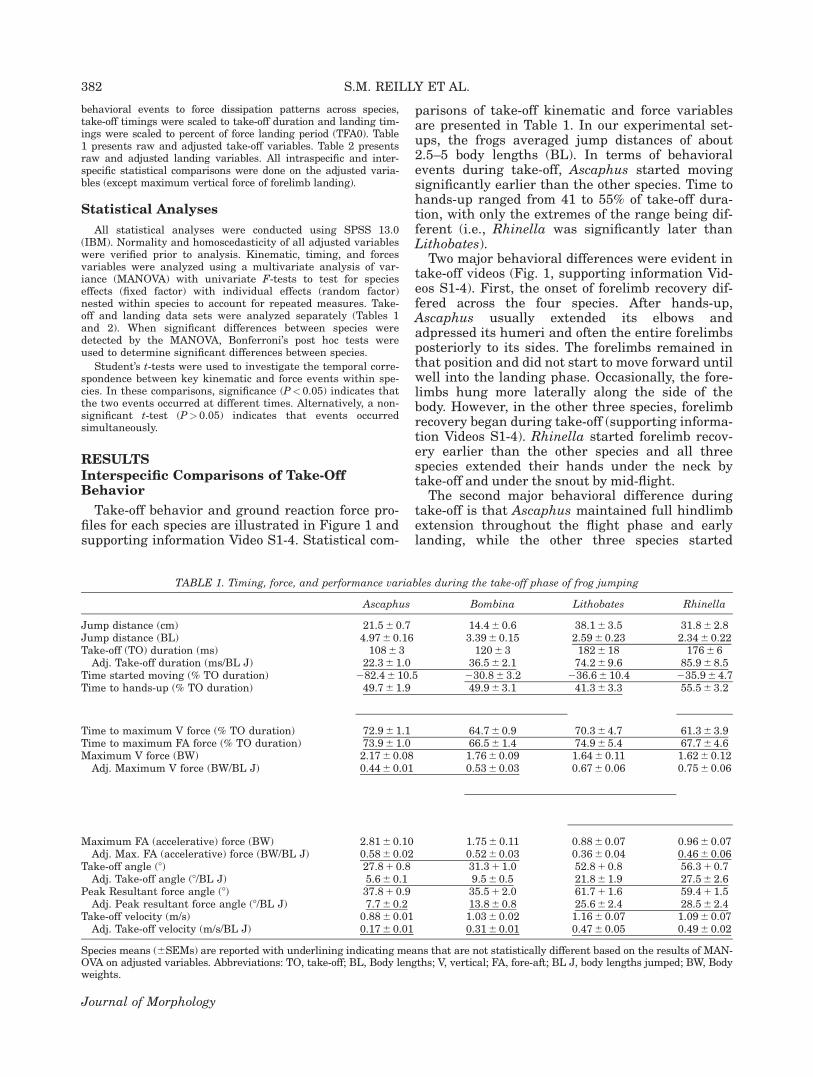
Key events describing the kinematic phases ofthe behavioral landing phase in our four modelfrogs are illustrated in Figure 2 and supportinginformation Videos S1-4. Statistical comparisonsof landing kinematic variables are presented inTable 2. Key impact events of when the hands andbody-hit the substrate were used to define the fore-limb and body landing phases. The forelimb land-ing phase is defined from hand-hit to body-hit.This phase does not exist in leiopelmatids asreported previously (Essner et al., 2010). The bodylanding phase is defined as the time from body-hit until the end of braking landing forces at TFA0.
In all landings a stereotypical two phase behav-ior of hindlimb recovery was observed across allfour species. First, the hindlimbs folded up tomaximal flexion (hindlimb flexion phase) andthen the feet, followed by the toes, were loweredonto the substrate at the end of the landing (thefoot placement phase). The hindlimb flexionphase started at the onset of hindlimb flexion
TABLE 2. Timing and force variables during frog landing
Ascaphus Bombina Lithobates Rhinella
Behavioral landing versus force landing phasesBehavioral landing period (ms) 571 6 35 148 6 9 147 6 9 225 6 13Behavioral landing period (ms/BL J) 115 6 6 44 6 3 60 6 6 109 6 11
Force landing period (ms to TFA0) 117 6 4 69 6 4 144 6 7 221 6 10Force landing period (ms to TFA0)/BL J 23.7 6 1 21 6 2 59 6 6 100 6 9
Ratio of force/behavioral landing periods (%) 21.4 6 1.1 49.0 6 4.2 99.0 6 1.6 100.1 6 3.3Ratio of force/behavioral landing (%/BL J) 4.4 6 0.3 15.3 6 1.8 41.0 6 3.7 47.7 6 4.0Timing variables (in % time to FA0)Time to body-hit (end of FL Landing phase) 0 20.7 6 1.0 35.8 6 3.3 52.9 6 2.9Time to maximum V force of FL Landing na 20.7 6 1.0 35.8 6 3.3 28.6 6 2.0Time to maximun FA during of FL Landing na 20.7 6 1.0 35.8 6 3.3 26.4 6 2.5Time to FA asymptote to BW in Ascaphus 36.2 6 1.8Time to feet slap na 40.5 6 2.8 53.2 6 0.8 naTime to maximum V force of body landing 10.1 6 0.4 41.0 6 1.9 51.8 6 3.6 78.3 6 1.6Time to max. FA (braking) force of body landing 12.5 6 0.8 44.8 6 2.2 54.4 6 2.9 80.8 6 6.1Time to feet start down/end HL Ext. phase 351.6 6 31.0 146.6 6 13.9 74.9 6 3.4 44.0 6 2.9Time to foot touch down 455.7 6 28.3 213.1 6 16.8 95.4 6 1.5 55.7 6 3.5Time to toe touch down 493.6 6 29.4 229.6 6 17.2 101.3 6 1.7 101.5 6 3.3Force variablesForelimb landingMaximum V force of FL Landing (BW)* na 1.92 6 0.13 1.38 6 0.07 1.99 6 0.15Max. FA (braking) force of FL Landing (BW) na 21.46 6 0.43 20.67 6 0.09 20.99 6 0.06
Adj. Max. FA force of FL Landing (BW/BL J) na 20.44 6 0.04 20.28 6 0.05 20.47 6 0.05Body landingMaximum V force body landing (BW) 5.87 6 0.40 4.27 6 0.18 3.07 6 0.20 1.50 6 0.05
Adj. Max. V force body landing (BW/BL J) 1.21 6 0.09 1.28 6 0.06 1.26 6 0.13 0.72 6 0.076Maximum FA (braking) force body landing (BW) 23.17 6 0.16 22.75 6 0.08 21.64 6 0.14 20.96 6 0.07
Adj. Max. FA force body landing (BW/BL J) 20.66 6 0.05 20.83 6 0.05 20.68 6 0.09 20.46 6 0.06
Species means (6 SEMs) are reported with underlining indicating means that are not statistically different based on the results ofMANOVA on adjusted variables. Abbreviations: TFA0, time that the fore-aft force retuns to zero; V, vertical, FA; fore-aft; BL J,body lengths jumped; BW, Body weights; FL, forelimb; HL, Hindlimb.*This was the only variable not correlated with jump distance or body size and was not adjusted.
383TAKE-OFF AND LANDING IN FROGS
Journal of Morphology

and ended when the hindlimbs were fully flexed.The onset of hindlimb flexion started afterTFA0 in Ascaphus (as reported previously; Essneret al., 2010) and soon after take-off in the othertaxa. During hindlimb flexion, the knees andankles came to full flexion before the hip, to bringthe hindlimbs to a partially folded position withthe knees still not fully tucked to the side of thebody. Near the end of hindlimb flexion, there wasan obvious anterodorsal rotation at the hip thatelevated the folded hindlimb and foot to bring the
thigh against the side of the body. This is thefinal portion of hip flexion which brings the hin-dlimbs to their point of maximum flexion with thethigh, shank, and foot held above the substrateand against the body (5 the end of the hindlimbflexion phase).
The foot placement phase starts at the endof the hindlimb flexion phase. After maximumhindlimb flexion and anterodorsal hip rotation,the hip slowly rotated dorsally to place the soleof the foot on the substrate with the toes elevated
Fig. 1. Representative frame sequences illustrating take-off kinematics and forces for four species of frogs scaled to take-off dura-tion. Kinematic frames illustrate key launch events and the traces illustrate ground reaction forces (GRFs) during the launch inBWs. Note the sustained posterior extension of the limbs in Ascaphus compared to the onset of forelimb and hindlimb flexion priorto take-off in the other species. Video sequences of jumping for all for species are presented in supporting information Videos S1-4.
384 S.M. REILLY ET AL.
Journal of Morphology
(foot-down). Subsequently, the toes were low-ered to the substrate with the longest toe (fourth)brought down last (toe-down, i.e., end of thebehavioral landing period). After foot-down, thefrogs often moved slightly, stood higher on theirforelimbs and/or tucked their feet in medially, butby the time to toe-down they appeared to be in anormal sitting position ready for another jump. Inaddition, if the eyes were closed during the jump,they reopened at this time (last 2 Rhinella framesin Fig. 2).
Interspecific Comparisons of LandingBehavior
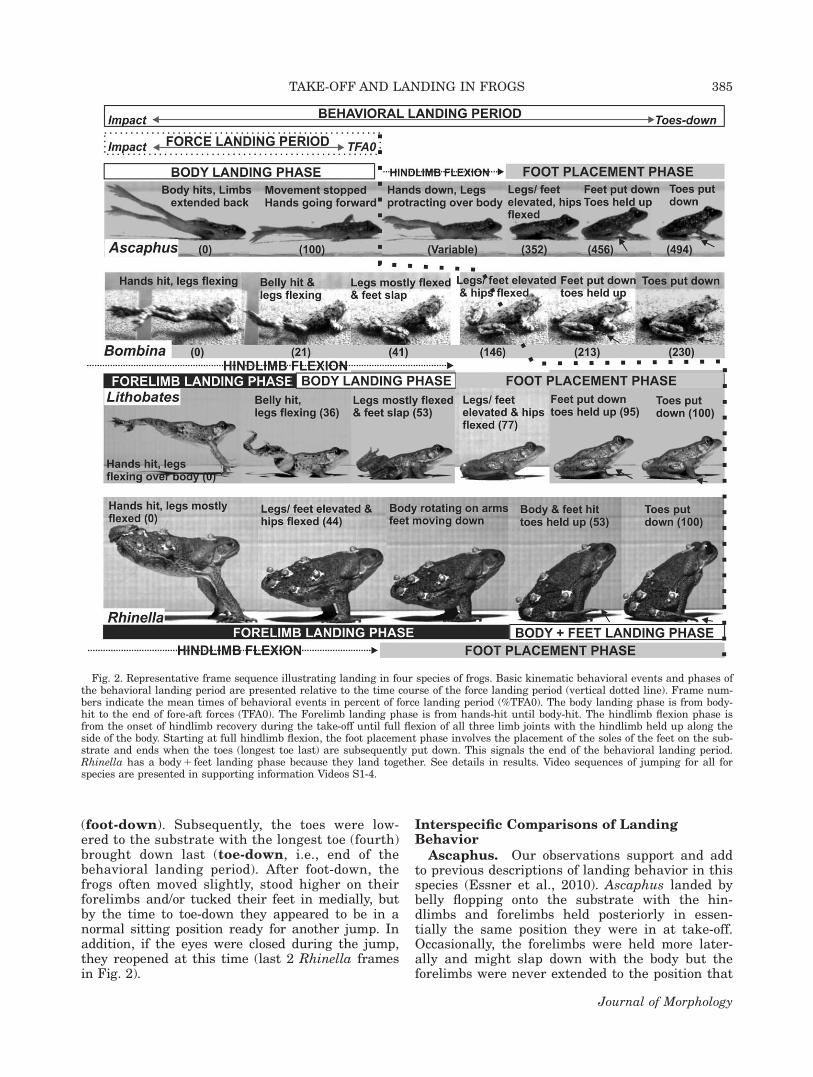
Ascaphus. Our observations support and addto previous descriptions of landing behavior in thisspecies (Essner et al., 2010). Ascaphus landed bybelly flopping onto the substrate with the hin-dlimbs and forelimbs held posteriorly in essen-tially the same position they were in at take-off.Occasionally, the forelimbs were held more later-ally and might slap down with the body but theforelimbs were never extended to the position that
Fig. 2. Representative frame sequence illustrating landing in four species of frogs. Basic kinematic behavioral events and phases ofthe behavioral landing period are presented relative to the time course of the force landing period (vertical dotted line). Frame num-bers indicate the mean times of behavioral events in percent of force landing period (%TFA0). The body landing phase is from body-hit to the end of fore-aft forces (TFA0). The Forelimb landing phase is from hands-hit until body-hit. The hindlimb flexion phase isfrom the onset of hindlimb recovery during the take-off until full flexion of all three limb joints with the hindlimb held up along theside of the body. Starting at full hindlimb flexion, the foot placement phase involves the placement of the soles of the feet on the sub-strate and ends when the toes (longest toe last) are subsequently put down. This signals the end of the behavioral landing period.Rhinella has a body 1 feet landing phase because they land together. See details in results. Video sequences of jumping for all forspecies are presented in supporting information Videos S1-4.
385TAKE-OFF AND LANDING IN FROGS
Journal of Morphology

could have any effect on controlling landing. Dur-ing landing, the toes and/or thighs or head some-times hit the substrate first or with the body.During impact, one or both hindlimbs occasionallyrecoiled forward, rotating dorsolateral at the hipbut they returned to their fully extended positionprior to the onset of hindlimb recovery. However, ifthe hindlimbs were flexed in any way from impactforces, they re-extended posteriorly prior to thebeginning of limb flexion.
The onsets of forelimb and hindlimb recoverywere highly variable in Ascaphus (thus were notdigitized) and the forelimbs generally recoveredwell before the legs, as illustrated in Figure 2.Both forelimb and hindlimb recoveries began latein the force landing period or after TFA0. The endof the hindlimb flexion phase, the time to foot-down, and toe-down were well after TFA0 (at 352,456, and 494% of TFA0, respectively; Table 2).Thus, hindlimb recovery occurred after landingforces had ended and took about four times longerthan the force landing period. In some landingsequences, limb recovery occurred in a trot-likefashion with one forelimb or hindlimb or diagonallimb couplet recovering slightly ahead of the otherlimb or limb couplet.
Bombina and Lithobates. These two specieshad very similar landing behavior in terms of keykinematic events, but differed in their relativetimings. Both species typically landed on theirextended forelimbs which collapsed at theshoulder and elbow joints while the hindlimbs con-tinued to flex anterodorsally over the level of thepelvis (Fig. 2). In a few trials, Bombina landedwith the belly hitting shortly after the hands.Bombina and Lithobates shared what we termedthe “feet slap” behavior. In the body landingphase, the belly hit first, followed by the pubis,proximal thighs, and the flexing hindlimbs andfeet slapping laterally in rapid succession. Thus,they land with a “hands–belly–feet slap” sequence(Figs. 2 and 3). After feet-slap, the feet and hin-dlimbs bounced and anterodorsal hip rotation (thelast part of late hindlimb flexion) occurred. Thismoved the hindlimbs to maximum flexion, withthe flexed limb held off of the substrate and tightto the body, signaling the end of the hindlimb flex-ion phase. Feet-slap is not present in Ascaphusbecause the hindlimbs remain fully extendedthroughout the body landing phase or longer, orRhinella because the hindlimbs come to full flex-ion during the forelimb landing phase.
Although similar in landing sequence, Bombinaand Lithobates differ in the timing of the behav-ioral landing period in relation to force landingperiod. In Bombina, similar to Ascaphus, maxi-mum hindlimb flexion, foot-down, and toe-downoccurred well after forces abated (147, 213, and230% of TFA0, respectively). In Lithobates, thetime to toe-down was not statistically different
from TFA0 (101.3 vs. 100%, respectively), andthus, the behavioral landing period was concur-rent with the force landing period and the frog isready to jump when the forces have returned tonormal.
Rhinella. In Rhinella, the force and behaviorallanding periods were also concurrent as time to toe-down was not significantly different from TFA0(101.5 vs. 100%, respectively). A novel feature oflanding behavior in Rhinella is that the hindlimbswere almost completely folded when the hands-hitthe substrate. This observation is in accordancewith previous reports of hopping behavior in R.marina (Azizi et al., 2014; Gillis et al., 2014; Reillyet al., 2015a). In toads, the body rotated on the flex-ing humeri for about 4/5ths of the forelimb landingphase as the humeri became adpressed against thetrunk. They then teetered on the forelimbs as thebody rotated further down to drop the feet, pubis,and posterior abdomen onto the substrate together.The feet tended to hit after the body, but this wasnot a significant difference (55.7 vs. 52.9% of TFA0,respectively). The feet were able to land with thebody because the hindlimb flexion phase was com-pleted at about 83% of the forelimb landing phase.Thus, the foot placement phase began while thebody was still aloft (Fig. 2). Accordingly, the bodylanding phase in Rhinella was actually a“body 1 foot” landing phase and the toes were heldup for a long time (about the last 45% of the forcelanding phase) before the toads settle back into asitting position.
Comparisons of the Landing Phases With theLanding Force Profiles
Patterns of forces in relation to kinematicphases are presented in Figure 3 with comparisonsof mean timing and force variables in Table 2.Within each species, the times to peak verticaland fore-aft forces during landing were not signifi-cantly different (t-tests, P’s>0.05) but the relativetiming of peak forces within the force landingphase differed. Ascaphus lands on its body, thus,the body landing phase was coincident with theforce landing phase. Ground reaction forces inAscaphus peaked quickly after impact at 10 and12% of TFAO (vertical and fore-aft forces, respec-tively) followed by a long period of highly variablefluctuations in forces as the frogs slid and bouncedbefore coming to a halt (Fig. 3). After the large ini-tial landing braking pulse, the fore-aft forces lev-eled off to low levels at 36.2% of TFA0 indicatingthat the main pulse of landing forces lasted forabout a third of the force landing phase.
As Bombina and Lithobates landed with the“hands–belly–feet-slap” sequence, both vertical andfore-aft components of the ground reaction forceincreased quickly through the forelimb landingphase and peak in the middle of the body landing
386 S.M. REILLY ET AL.
Journal of Morphology
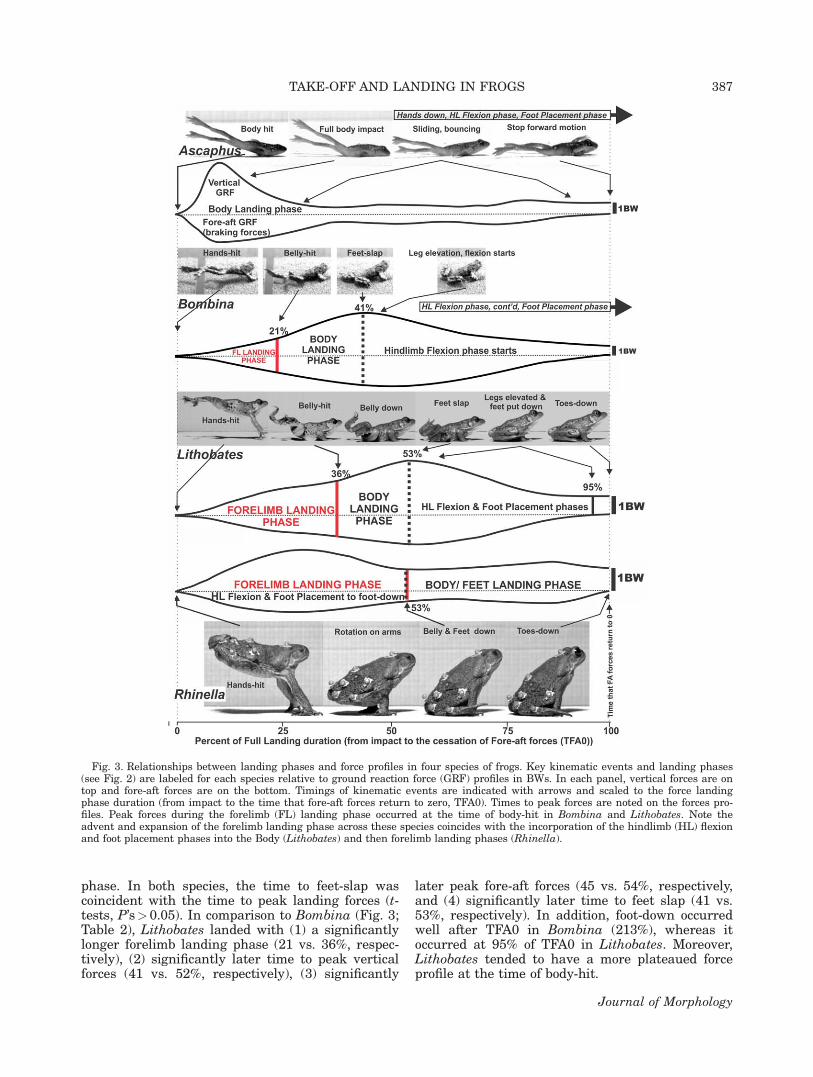
phase. In both species, the time to feet-slap wascoincident with the time to peak landing forces (t-tests, P’s>0.05). In comparison to Bombina (Fig. 3;Table 2), Lithobates landed with (1) a significantlylonger forelimb landing phase (21 vs. 36%, respec-tively), (2) significantly later time to peak verticalforces (41 vs. 52%, respectively), (3) significantly
later peak fore-aft forces (45 vs. 54%, respectively,and (4) significantly later time to feet slap (41 vs.53%, respectively). In addition, foot-down occurredwell after TFA0 in Bombina (213%), whereas itoccurred at 95% of TFA0 in Lithobates. Moreover,Lithobates tended to have a more plateaued forceprofile at the time of body-hit.
Fig. 3. Relationships between landing phases and force profiles in four species of frogs. Key kinematic events and landing phases(see Fig. 2) are labeled for each species relative to ground reaction force (GRF) profiles in BWs. In each panel, vertical forces are ontop and fore-aft forces are on the bottom. Timings of kinematic events are indicated with arrows and scaled to the force landingphase duration (from impact to the time that fore-aft forces return to zero, TFA0). Times to peak forces are noted on the forces pro-files. Peak forces during the forelimb (FL) landing phase occurred at the time of body-hit in Bombina and Lithobates. Note theadvent and expansion of the forelimb landing phase across these species coincides with the incorporation of the hindlimb (HL) flexionand foot placement phases into the Body (Lithobates) and then forelimb landing phases (Rhinella).
387TAKE-OFF AND LANDING IN FROGS
Journal of Morphology

Landing behavior in Rhinella was fundamen-tally different from the other species (Figs. 2 and3), and this was reflected in the pattern of landingforces. In Rhinella, both peak vertical and fore-aftforces occurred during the hand landing phase (29and 26% of TFA0, respectively; Table 2). Groundreaction forces then abated significantly as thetoads teetered on their arms. Subsequently, a sec-ond lesser pulse of landing forces occurred later inthe “body 1 feet” landing phase (Fig. 3). The body-1 feet landing phase started midway through theforce landing period (53%) and body 1 feet impactforces peaked at 78 and 81% of the force landingduration in the vertical and fore-aft direction,respectively (Fig. 3; Table 2).
DISCUSSIONTake-Off Behavior
Phylogenetic patterns in the take-off.Observed differences in behavior and forcesdynamics in relation to take-off events provide evi-dence to form several hypotheses about the func-tional patterns of evolution of take-off behavior infrogs. These patterns are illustrated within a phy-logenetic context in Figure 4.
At the base of the phylogeny, three functionaltraits of take-off behavior are considered to beancestral because they are unique to Ascaphus andchange to different states in other species. Twofocus on hindlimb and forelimb recovery (white andgrey bars, respectively). In Ascaphus, hindlimbswere held fully extended during the flight phaseand the forelimbs were held to their sides duringtake-off. In fact, both sets of limbs remainedextended posteriorly through most of the force land-ing period (Fig. 2; Table 2) and the onset of bothforelimb and hindlimb recovery started late in theforce landing period and ended well after it. Thus,there was no limb recovery during take-off andflight phases in Ascaphus and it appears that limbswere actively extended posteriorly. In the otherthree species, the hindlimbs began to flex and foldup after take-off and the forelimbs began to extendanteriorly shortly after hands-up. Thus, onset oflimb recovery in the lalagobatrachians appears tohave shifted from the landing phase to the take-offphase.
The third trait (black bars), described by a suiteof functional characteristics, contrasts the slowand shallow take-off trajectory of Ascaphus, to theintermediate velocity and angle of Bombina, to thesteeper and faster take-off in Lithobates andRhinella. Previous studies also reported shallowertake-off angles and slower take-off velocities inBombina compared to ranid frogs (Choi and Park,1996; Choi et al., 2000). This does not appear to bea size-related phenomenon, because we scaled forsize and because a number of neobatrachian frogsin the size range of Ascaphus and Bombina have
been found to have relatively high take-off angles(Marsh and John-Alder, 1994; Choi et al., 2003).Steeper take-off angles result from a relativedecrease in fore-aft forces. The ratio of peak fore-aft to vertical forces varied from 1.3 in Ascaphus,to 1 in Bombina to 0.52 and 0.59 in Lithobatesand Rhinella, respectively, reflecting a shift to amore vertically oriented take-off force. The shift tosteeper, faster take-offs appears to be related torelatively longer hindlimb lengths and relativelygreater hindlimb muscle masses that have beenshown in ranids and other neobatrachians com-pared to Bombina (Zug, 1985; Emerson, 1978;Choi and Park, 1996; Choi et al., 2003). Longerhindlimbs and bigger muscles provide neobatra-chians with longer out-lever forelimbs and greatertotal muscle force against gravity to produce afaster, steeper take-off (Choi and Park, 1996; Choiet al., 2003). Our force data for Bombina and Lith-obates mirror these studies.
Gans and Parsons (1966) hypothesized that thesynchronous hindlimb extension which character-izes jumping may have been derived from lungefeeding. In this scenario, jumping evolved as away of expanding the foraging radius. If true, thenone might expect basal taxa to exhibit relativelyhigher fore-aft forces and lower take-off anglesduring the launch, when compared with morederived jumping taxa. Our results are consistentwith this idea.
Comparisons to other take-off studies.Because our goal was to provide a detailed descrip-tion and comparison of jumping behavior in frogs,a concern was whether individuals were perform-ing normal jumping locomotion within the confinesof the force plate dimensions we used. In ourstudy, take-off variables (distance, angle, velocity,durations, and peak forces) were comparable toother studies of jumping in these species (Asca-phus, Essner et al., 2010; Bombina, Choi andPark, 1996; Choi et al., 2000, 2003; Lithobates,Choi and Park, 1996; Choi et al., 2000, 2003; Rob-erts and Marsh, 2003; Rhinella, Choi et al., 2000;Gillis and Biewener, 2000; Reilly et al., 2015b).Lithobates jump distances were shorter than pre-viously reported mean jump distances (Zug, 1985),but they were still within the range of distancesobserved in trained professional bullfrogs jumpingin Calaveras County (Astley et al., 2013).
Take-off is a stereotyped behavior in frogs andmany performance, physiological, and energeticaspects of it have been related to jump velocity ordistance (e.g., Hirano and Rome, 1984; Lutz andRome, 1994; Marsh, 1994; Wilson et al., 2000; Wil-son, 2001; James and Wilson, 2008; Cox and Gillis,2015). Many of these studies measured the onsetand offset of take-off but few have reported othermovements during take-off of which we have iden-tified two: the time to start moving and the timeof hands-up.
388 S.M. REILLY ET AL.
Journal of Morphology

Several studies have reported take-off behaviorand noted the presence of body movement prior tothe onset of leg flexion in frogs. Peters et al.(1996) described a “beginning of movement point”in Lithobates pipiens where head movements pre-ceded the onset of hindlimb flexion by 31 ms or22% of take-off duration. Gillis and Biewener(2000) used the “onset of toad movement” as thestart of the take-off in R. marina but stated that itdid not necessarily coincide with the onset of hind-limb extension. Here, we found that time to startmoving occurs earlier than the onset of hindlimbflexion in all four species. Thus, frogs appear to bemoving in preparation for jumping before theonset of hindlimb extension.
The time to hands-up has also been quantifiedin two other ranid frogs (Peters et al., 1996; Wanget al. 2014) which were at 29% (L. pipiens) and63% (R. dybowski) of take-off duration, which isconsistent with the 41–55% range we observed.Recently, Wang et al. (2014) showed in R. dybow-ski that the forelimbs contribute to raising or lev-ering BW during take-off prior to hands-up. In ourstudy, the later time to hands-up in Rhinella(55%) compared to Lithobates (41%) may suggestsome increased contribution of the forelimbs totake-off in toads. Akella and Gillis (2011) alsofound pre-take-off activity in the anconeus muscleof the shoulder but concluded that they were likelyunimportant in augmenting force production dur-ing take-off. Clearly more muscle activity andforce data are needed in both the hindlimbs andforelimbs to fully understand their function duringtake-off in frogs.
Landing Behavior
Limb recovery in frogs is a critical behavioressential to prepare for subsequent jumps. Accord-ingly, the ability to reset the limbs quickly is a keyadaptation for predator avoidance in frogs. Ourdata show that the roles of the forelimbs and hin-dlimbs vary during frog landing behavior.
Forelimb landing and the dissipation oflanding forces. Early forelimb recovery is criti-cal for the arms-first landings that most frogs do.Accordingly, frogs that land on their forelimbshave adopted the opposite behavior of Ascaphus(that hold the limbs posteriorly) and start forelimbrecovery between hands-up and take-off.
Differences in force patterns show that there aresignificant shifts in the duration and timing offorce propagation across the four species. Landingforces in the belly-flop landing of Ascaphus peakquickly and return to near static levels by 36.2%of TFA0. Thus, most of their primary landingforces occurred significantly earlier than in theother species and entirely on the body (Table 2).The acquisition of the forelimb landing phase inBombina, Lithobates, and Rhinella, delays bodycontact and peak body forces to later in the land-ing phase (Table 2). As forelimb landing durationincreases from Bombina to Lithobates to Rhinella,peak fore-aft body forces decrease significantly(Table 2). In addition, the ratio of peak take-offforces to peak landing forces decreases from2.70 in Ascaphus to 1.21 in Rhinella. Therefore, alonger forelimb landing phase and the resultingdelay of body landing appear to decrease peak
Fig. 4. Phylogenetic patterns of take-off characteristics based on behavioral and functional differences among four species of frogs.Tree topology is from Frost et al. (2006). Shaded bars represent the different states of behavioral and functional traits: hindlimbmovements (white), forelimb movements (gray), and take-off characteristics (black). The phylogenetic interpretation is based on theassumption that characters found in Ascaphus are plesiomorphic to all other frogs.
389TAKE-OFF AND LANDING IN FROGS
Journal of Morphology

braking forces of the body, thus, enhancing thecontrol and safety of landing.
Rhinella has shifted the primary force absorbingfunction from the body to the arms, which appearsto significantly decrease peak vertical and brakingforces in the body landing phase (Table 2). InBombina and Lithobates, before the body-hits, theforelimbs take on about �44% of the peak verticalforces incurred during the body landing phase(Forelimb peak divided by body peak, Table 2).Whereas, in Rhinella’s forelimb-dominated land-ing, this is reversed with the peak vertical forcesof the forelimb landing about �31% greater thanthose of body landing (Fig. 2; Table 2). Thus,forelimb-dominated landing behavior results in asignificant decrease in the vertical forces absorbedby the body during landing.
Hindlimb function in the landing. Ourstudy shows that the hindlimbs have little contri-bution to the absorption of landing forces. Basedon Ascaphus (and other leiopelmatids; Essneret al., 2010) early frogs likely had the ability tolaunch but lacked the ability to control landingsand crash-landed on their bellies. Repositioningthe limbs may be of less importance to basal anu-rans because they rarely jump in nature and it iseven less common for them to immediately jumpagain (Reilly et al., 2015b). Our data show thatAscaphus actively holds its hindlimbs extendedposterodorsally until well after landing forces havesubsided and then the slow process of hindlimbrecovery and foot placement begins, whereas theother three species begin hindlimb recovery duringthe aerial phase. Bombina and Lithobates slaptheir hindlimbs and feet laterally at the time thatpeak body landing forces occur and the feet areplaced on the substrate at the time landing forcesabated. Rhinella completes hindlimb recovery byfolding the hindlimbs and feet anterodorsally priorto impact with the forelimbs and subsequently thefeet hit the ground with the body well after peakforces have occurred. Thus, our data show thathindlimb function during landing is to fold thehindlimbs out of the way of the body and have nocontribution to the dissipation of landing forces. InBombina and Lithobates, a lot of hindlimb flexionoccurs after body-hit and the hindlimbs and feetvisibly slap and bounce on the substrate, so itappears that these species may be taking advant-age of impact momentum to aid in the final flexionof the limbs. This is in contrast to Ascaphus whichdo not use bouncing momentum for hindlimb flex-ion because the hindlimbs are held posteriorlyduring impact.
Frogs and many other jumping vertebrates areknown to exhibit a proximal to distal sequence ofjoint extension during take-off, with the hipextending first, followed by the knee and ankle atapproximately the same time. This sequence mayallow sufficient time for elastic energy storage
(Biewener, 2003; Nauwelaerts and Aerts, 2003;Wang et al., 2014). We note here that limb recov-ery took place in the opposite sequence (distal toproximal) in all species examined. In Bombina,Lithobates, and Rhinella, ankle and knee flexionbegan immediately after take-off, followed later byhip flexion; whereas, these events took place afterTFA0 in Ascaphus. Recent work by Schnyer et al.(2014) indicates that the release of elastic energystored in the extended hindlimbs following take-offmay contribute to initial limb recovery in Rhinella.Because Rhinella completes the hindlimb flexionphase during flight prior to impact on the handsthe rest of hindlimb recovery is hypothesized torely solely on muscle power. The same may betrue for the other species exhibiting aerial recov-ery. Ascaphus, on the other hand, is an enigma,since it appears to be overriding any elastic recoilwhich could aid in recovery, by actively keepingthe hindlimbs extended until after TFA0.
Shifting the timing of hindlimb recoveryand the foot placement phase. Our studyshows a transition from the essentially separatedlimb flexion and foot placement phases of Asca-phus, to the onset of flexion soon after take-off andearlier recovery phases in Bombina, to the coordi-nated recovery and force phases in Lithobates.Frogs with forelimb landing gain the advantage oflower landing forces and a significant ecologicaladvantage in that the limbs are reset and ready tojump again about two (Bombina) to four (Litho-bates) times faster than Ascaphus (Table 2).Behavioral and force landing durations were simi-lar in Lithobates but the Behavioral duration wastwice as long as the force period in Bombina.Bombina, with less developed forelimb landing,landed harder and waited longer for limb recoveryto be completed before it can jump again, whereasLithobates achieves limb recovery by the timeforces dissipate and thus, is ready to jump earlier.
It is interesting to note that the significantlyearlier limb recovery and controlled placement ofthe feet in Rhinella appears to be related to theirability to use a bounding gait instead of repeatedjumps (Reilly et al., 2015a). To bound, toads startthe foot placement phase early in the forelimblanding phase to extend the hindlimb ventrallybefore the feet touch the ground. Because the hin-dlimbs are already extending toward the end ofthe forelimb landing phase they are in a positionto either land normally with the body and stop orto extend further to keep the body aloft thus gen-erating a bounding step.
Phylogenetic patterns in the landing. Keyfindings describing interspecific changes in land-ing behavior are illustrated in three charactertransitions in Figure 5. Landing starts withground contact and it has already been shownthat Ascaphus belly-flops and that forelimb-basedlanding behavior appears to have arisen in the
390 S.M. REILLY ET AL.
Journal of Morphology

common ancestor of the lalagobatrachians (Essneret al., 2010). Our study adds that (1) in Ascaphus,the body absorbs most of the landing forces, (2)the forelimbs–belly–feet slap sequence in Bombinaand Lithobates allows the forelimbs to take onsome of the landing forces, and (3) in Rhinella,forelimbs absorb most of the landing forces (blackbars). The transition to a longer forelimb landingphase in Lithobates (compared to Bombina)appears to result in slower dissipation of landingforces that decreases the magnitudes of forcesabsorbed by the forelimbs and the body.
Paralleling the shift from belly flopping to fore-limb landing is the onset of both forelimb andhindlimb recovery (white bars). We show that limbrecovery starts and ends after landing forces abatein Ascaphus but begins during take-off in theother species.
In all species, we found that landing behaviorended with the stereotypical behavior where lateanterodorsal hip rotation (the end of hindlimbflexion) is followed by careful placement of thefoot, and then the longest toe, on the ground.This trait has four character states (grey bars)based on the differences between the behaviorallanding duration and the force landing duration.In Ascaphus, the foot placement phase ended atabout five times the duration of the force landingperiod, in Bombina, it lasts about twice as long,and in the neobatrachians, toe-down was coordi-nated with the end of force landing period. In
Rhinella, hindlimb flexion phase and foot place-ment precede body-hit resulting in a unique fea-ture: the body and feet hit together (stipled bar).This novel feature in Rhinella may be related totheir more terrestrial locomotor ecology and theirability to use bounding locomotion (Reilly et al.,2015a).
Comparison to other landing studies.Details of landing behavior across frogs have seenlittle study. Emerson and de Jongh (1980) ana-lyzed axial muscle activity in relation to kinemat-ics in three species of frogs. However, these wererecorded from take-off to hand-hit, so there is nocomparable landing information. Peters et al.(1996) described hopping kinematics in L. pipiens.The landing events reported were hand-hit andthe “end of movement” designated as the pointwhen the frog had settled in to a position compa-rable to the prelaunch posture. Gillis and Biew-ener (2000) added the time of body-hit in theirstudy of hindlimb muscle function during jumpingand swimming in Rhinella. The end of landingwas defined as when “the body completely comesto rest and stops moving,” mirroring our definitionbecause the penultimate video frame of their Fig-ure 1 shows the foot and body down with the lon-gest toe held up and the last frame shows the toe-down 56 ms later. Nauwelaerts and Aerts (2003)analysis of jumping and landing forces in R. escu-lenta includes the time of body-hit and the end oflanding was defined behaviorally as when the
Fig. 5. Phylogenetic patterns in landing characteristics in based on behavioral and functional differences among four species offrogs. Tree topology is from Frost et al. (2006). Shaded bars represent the different states of behavioral and functional traits: dissipa-tion of landing forces (black), forelimb and hindlimb recovery (white), and foot placement phase (grey). Rhinella is unique in that thebody and feet hit together (stippled). The phylogenetic interpretation is based on the assumption that characters found in Ascaphusare plesiomorphic to all other frogs.
391TAKE-OFF AND LANDING IN FROGS
Journal of Morphology

“frog is back in launch position” based on videosand as TFA0 based on forces. Similarly to ourobservations, they stated that end of behaviorallanding was after TFA0, although no data werepresented on how long after TFA0 the frog is backin launch position. Unfortunately, without toe-down information, we cannot deduce whether R.esculenta is similar to Lithobates and Rhinellawith toe-down occurring when forces have dissi-pated, or if the foot placement phase takes longer,similarly to Bombina. In their Figure 4 videosequence, the belly hits first, followed by the pubisand thighs and the feet appear to slap laterallylike Bombina and Lithobates. Feet-slap in R. escu-lenta also appears to be coincident with or justafter peak vertical force of the body-hit as wefound in Lithobates (Fig. 2). Nauwelaerts andAerts (2003) found two types of force landing pro-files associated with a shift in the timing of body-hit. Their Type II is the same stepped profile wecommonly observed in Bombina and Lithobates(Fig. 3) with the forelimbs taking on initial forceswith the body taking the rest of the forces once ithits. Their Type I profile was purely parabolicwith the body hitting after the peak forces andboth patterns occurred at the same jump distan-ces. Because no kinematic data were available onthese two types of landing, we are unable to deter-mine if there is a significantly different delay ofbody-hit relative to peak forces in the Type I pro-files. We observed a few profiles similar to theirType I in Lithobates (�2%) and none in Bombina,and thus, we suggest that their Type II jumps rep-resent the most common pattern of landing forces.
Future Considerations
Our comparison of jumping and landing behav-ior in four model species spanning the anuranphylogeny has revealed important new insightsabout the step-wise evolution of landing functionin frogs. However, much more work is needed.Type I landings described by Nauwelaerts andAerts (2003) suggest the possibility of a lesser con-tribution of the forelimbs during landing and laterfoot placement timing in R. esculenta. This showsthat we have only just begun to appreciate intra-specific variation in the landing behavior in frogs.Until now, the kinematics of forelimb landing havenot been studied outside of toads (reviewed in Gil-lis et al. 2014; Cox and Gillis, 2015) and theincreasing contribution of the forelimbs in Litho-bates compared to Bombina suggests that theremay be an evolutionary gradient in the contribu-tion of the forelimbs in controlling landing. Stud-ies of other basal lalagobatrachians and the sistertaxa to the Ranidae are needed to fill in the gapsand test the generality of the hypotheses we pres-ent above. Much of the Hyloidea also remain to bestudied with almost all the work on this group
having focused on the two largest and mostderived radiations (Hylidae and Bufonidae). Inaddition, since semiaquatic frogs often escape intothe water and it would be interesting to seewhether the functional patterns of landing on landwe have described here are retained when theyland in water. The recent discovery of multiplecases of convergent evolution of the sagittal-hingepelvic morphotype across the Neobratrachia(Reilly and Jorgensen, 2011; Jorgensen and Reilly,2013) provides interesting comparative frame-works to test whether landing behaviors we foundare also convergent in these taxa. Furthermore,there is little information on the contribution ofthe anuran pelvic girdles to jumping behavior andthe preponderance of landing differences in thisstudy suggests that the pelvis might play a role inlanding.
Akella and Gillis (2011) were correct in saying“that hopping is not always about the legs.” Wehave shown that hindlimb function during take-offis far more stereotyped across species than thekinematics and force patterns of the forelimbs andbody during landing. Thus, we would add thatjumping is not always about the take-off, becausemost of the interspecific differences we found werein the landing behavior of the forelimbs and body.
ACKNOWLEDGMENTS
We thank Susan Williams and Mike Jorgensenfor discussions about patterns of frog jumpingbehavior. Funding was provided by 1) the BakerFund and the Faculty Fellowship Leave Programat Ohio University (SMR), 2) Faculty researchfunds from Southern Illinois University atEdwardsville (RLE), and 3) the National ScienceFoundation (MRI DBI-0922988), the Ohio Board ofRegents, the Ohio University Office of the VicePresident for Research and the Ohio UniversityHeritage College of Osteopathic Medicine (SJM).All procedures were approved by the Ohio Univer-sity Institutional Animal Care and Use Committee.
LITERATURE CITED
Abourachid A, Green DM. 1999. Origins of the frog-kick?Alternate-leg swimming in primitive frogs, families Leiopel-matidae and Ascaphidae. J Herpetol 33:657–663.
Akella T, Gillis GB. 2011. Hopping isn’t always about the legs:Forelimb muscle activity patterns during toad locomotion.J Exp Zool Part A 315:1–11.
Astley HC, Abbott EM, Azizi E, Marsh RL, Roberts TJ. 2013.Chasing maximal performance: A cautionary tale from thecelebrated jumping frogs of Calaveras County. J Exp Biol216:3947–3953.
Azizi E. 2014. Locomotor function shapes the passive mechani-cal properties and operating lengths of muscle. Proc. R. Soc.B 281:20132914.
Azizi E, Abbott EM. 2013. Anticipatory motor patterns limitmuscle stretch during landing in toads. Biol Lett 9:20121045.
392 S.M. REILLY ET AL.
Journal of Morphology

Azizi E, Larson NP, Abbott EM, Danos N. 2014. Reduce torquesand stick the landing: Limb posture during landing in toads.J Exp Biol 217:3742–3747.
Biewener AA. 2003. Animal Locomotion. Oxford: Oxford Uni-versity Press.
Bisa R, Sfenthourakis S, Fraguedakis-Tsolis S, ChondropoulosB. 2007. Population density and food analysis of Bombinavariegata and Rana graeca in mountainous riverine ecosys-tems of northern Pindos (Greece). J Biol Res 8:129–137.
Carroll RL, Holmes RB. 2007. Evolution of the appendicularskeleton of amphibians. In Hall BK, editor. Fins into Limbs.Chicago: University of Chicago Press. pp. 185–224.
Choi I, Park K. 1996. Variations in take-off velocity of anuranamphibians: Relation to morphology, muscle contractile functionand enzyme activity. Comp Biochem Physiol 113A:393–400.
Choi IH, Shim JH, Lee YS, Ricklefs RE. 2000. Scaling of jump-ing performance in anuran amphibians. J Herp 34:222–227.
Choi IM, Shim JH, Ricklefs RE. 2003. Morphometric relation-ships of take-off speed in anuran amphibians. J Exp ZoolPart A 299:99–102.
Cox S, Gillis G. 2015. Forelimb kinematics during hopping andlanding in toads (Bufo marinus). J Exp Biol 218:3051–3058.
Duellman WE, Trueb L. 1994. Biology of Amphibians. Balti-more: The Johns Hopkins University Press. 670 p.
Emerson SB. 1978. Allometry and jumping in frogs: helping thetwain to meet. Evolution 32:551–564.
Emerson SB. 1979. The iliosacral articulation in frogs: Formand function. Biol J Linn Soc 11:153–168.
Emerson SB. 1985. Jumping and leaping. In: Hildebrand M,Bramble DM, Leim KF, Wake DB, editors. Functional Verte-brate Anatomy. Cambridge: Belknap Press. pp. 58–72.
Emerson SB, De Jongh HJ. 1980. Muscle activity at the ilio-sacral articulation of frogs. J Morphol 166:129–144.
Essner RLJ, Suffian DJ, Bishop PJ, Reilly SM. 2010. Landingin basal frogs: Evidence of saltational patterns in the evolu-tion of anuran locomotion. Die Naturwissenschaften 97:935–939.
Frost DR, Grant T, Faivovich J, Bain RH, Haas A, HaddadCFB, de S�a RO, Channing A, Wilkinson M, Donnellan SC,Raxworthy CJ, Campbell JA, Blotto BL, Moler P, Drewes RC,Nussbaum RA, Lynch JD, Green DM, Wheeler WC. 2006.The Amphibian Tree of Life. Bull Am Mus Nat Hist Soc 297:1–370.
Gans C, Parsons TS. 1966. On the origin of the jumping mecha-nism in frogs. Evolution 20:92–99.
Gillis GB, Biewener AA. 2000. Hindlimb extensor muscle func-tion during jumping and swimming in the toad (Bufo mari-nus). J Exp Biol 203:3547–3563.
Gillis GB, Akella T, Gunaratne R. 2010. Do toads have a jumpon how far they hop? Pre-landing activity timing and inten-sity in forelimb muscles of hopping Bufo marinus. Biol Lett 6:486–489.
Gillis GB, Ekstrom L, Azizi E. 2014. Biomechanics and controlof landing in toads. Integr Comp Biol 54:1136–1147.
Gomes FR, Rezende EL, Gizante MB, Navas CA. 2009. The evo-lution of jumping performance in anurans: Morphological cor-relates and ecological implications. J Evol Biol 22:1088–1097..
Griep S, Schilling N, Marshall P, Amling M, Hahme LM, HaasA. 2013. Pectoral girdle movements and the role of the gleno-humeral joint during landing in the toad, Rhinella marina(Linnaeus, 1758). Zoomorph 132:325–338.
Griffiths I. 1963. The Phylogeny of the Salientia. Biol Rev 38:241–292..
Handrigan GR, Wassersug RJ. 2007. The anuran Bauplan: Areview of the adaptive, developmental, and genetic underpin-
nings of frog and tadpole morphology. Biol Rev Camb PhilosSoc 82:1–25.
Hirano M, Rome LC. 1984. Jumping performance in frogs as afunction of muscle temperature. J Exp Biol 108:429–439.
James RS, Wilson RS. 2008. Explosive jumping: Extreme morpho-logical and physiological specializations of Australian rocketfrogs (Litoria nasuta). Physiol Biochem Zool 81:176–185.
Jorgensen ME, Reilly SM. 2013. Phylogenetic patterns of skele-tal morphometrics and pelvic traits in relation to locomotormode in frogs. J Evol Biol 26:929–943.
Kuzmin SL. 1990. Trophic niche overlap in syntopic postmeta-morphic amphibians of the Carpathian mountains (Ukraine:Soviet Union). Herpetozoa 3:13–24.
Lutz GJ, Rome LC. 1994. Built for jumping: The design of thefrog muscular system. Science 263:370–372.
Marsh RL. 1994. Jumping ability of anuran amphibians. AdvVet Sci Comp Med 38:51–111.
Marsh RL, John-Alder HB. 1994. Jumping performance of hylidfrogs measured eith high-speed cine film. J Exp Biol 188:131–141.
Nauwelaerts S, Aerts P. 2003. Takeoff and landing forces injumping frogs. J Exp Biol 209:66–77..
Noble GK. 1931. The Biology of the Amphibia. New York:McGraw-Hill.
Peters SE, Kamel LT, Bashor DP. 1996. Hopping and swimmingin the leopard frog, Rana pipiens: 1. Step cycles and kinemat-ics. J Morphol 230:1–16.
Reilly SM, Jorgensen ME. 2011. The evolution of jumping infrogs: Morphological evidence for the basal anuran locomotorcondition and the radiation of locomotor systems in crowngroup anurans. J Morphol 272:149–168.
Reilly SM, Montuelle SJ, Essner R Jr, Schmidt A, Halsey L,Jorgensen M, Naylor E. 2015a. Conquering the world in leapsand bounds: Hopping locomotion in toads is actually bound-ing. Funct Ecol, 29:1308–1316.
Reilly SM, Essner RJ, Wren S, Easton L, Bishop PJ. 2015b.Movement patterns in leiopelmatid frogs: Insights into thelocomotor repertoire of basal anurans. Behav Proc 121:43–53.
Roberts TJ, Marsh RL. 2003. Probing the limits to muscle-powered accelerations: Lessons from jumping bullfrogs. J ExpBiol 206:2567–2580.
Robovska-Havelkova P, Aerts P, Rocek Z, Prikryl T, Fabre AC,Herrel A. 2014. Do all frogs swim alike? The effect of ecologi-cal specialization on swimming kinematics in frogs. J ExpBiol 217:3637–3644.
Schnyer A, Gallardo M, Cox S, Gillis G. 2014. Indirect evidencefor elastic energy playing a role in limb recovery during toadhopping. Biol Lett 10:20140418.
Wang Z, Ji A, Endlein T, Samuel D, Yao N, Wang Z, Dai Z.2014. The role of fore-and hindlimbs during jumping in theDybowski’s frog (Rana dybowskii). J Exp Zool Part A 321:324–333.
Wells KD. 2010. The Ecology and Behaviour of Amphibians.Chicago: University of Chicago Press.
Wilson RS. 2001. Geographic variation in thermal sensitivity ofjumping performance in the frog Limnodynastes peronii.J Exp Biol 204:4227–4236.
Wilson RS, Franklin CE, James RS. 2000. Allometric scalingrelationships of jumping performance in the striped marshfrog Limnodynastes peronii. J Expl Biol 203:1937–1946.
Zug GR. 1985. Anuran locomotion: Structure and function. I.Preliminary observations on relation between jumping andosteometrics of appendicular and postaxial skeleton. Copeia1972:613–622.
Zug GR. 1985. Anuran locomotion: Fatigue and jumping per-formance. Herp 41:188–194.
393TAKE-OFF AND LANDING IN FROGS
Journal of Morphology