full text (1153.5 kb)
TRANSCRIPT

Acta
Vol. 30 No. 3-4
Palaecntclogica
pp. 115-135
Polonica
VVarszav;a , 1985
EGON T. DEGENS, JOZEF KAZMIERCZAK and VENUGOPALAN ITTEKKOT
CELLULAR RESPONSE TO CaH STRESSAND ITS GEOLOGICAL IMPLICATIONS
DEGENS, E . T. , KAZMIERCZAK, J ., and ITTEKKOT, V. : Cellular responseto CaB stress and its geological implications. Acta Palaeont. Polonica, 30, 3/4,115-135, 1985. (issued 1986).
Knowledge on transport and regulation of free calcium in the living cell isused in support of the theory (Kazmierczak et al. 1985) linking the onset ofbiocalcification a t a bo u t the P r eca m b r ia n /Ca m b ria n boundary to a r is e in Ca l+
concentrations in the shelf se as to levels toxic to biota . Following this event ,fluctuating Ca l+ levels in the Phanerozoic se as a re supposed to havechallenged a v ariety of protists and invertebrates to respond by depositfngno, th in, or t h ick skeletons respectivel y . Ch anges in type and' extent of calcification, a s observed in t h e stratigraphical record , a r e in ter pr e te d to reflectthe pulsa t i n g flow of Ca 2+ ions through crust, se a, a n d b iota . Some tmpllcations of that theory to (i) the history of s ea w ater, (i i ) the global carboncycle, (ii i) s t able carbon isotope geo ch em is t r y, a n d (Iv) sedimentation of suspended clays, a re briefly discussed.
Key W 0 r d s : Cell physiology, b iomineralization, calcification, calcium geoch emistry, carbonate a n d clay sedimenta tion.
Egon T. D egens, Venugopalan ltt ek kot : SCOPE/UNEP Interna t i onal CarbonUnit , Univers ity of Hamburg , Bundesstrasse 55, D -2000 Hamburg, F ederal Repubtic of G ermany ; Joze] Kazmierczak , Zaklad Paleobiologii , Polska AkademiaNauk, A t. Z w i rk i I W lgury 93, 02-089 W arszawa , Poland. R eceiV ed : F ebrua ry 1985.
INTRODUCTION
Carbonates are extracted from modern ocean principally by the actionof organisms. The VVay how they do it is still debatable. In 1962 it wassaid that "the nature of the local mechanism of calcification is one ofthe most important unresolved problems in biochemistry" (Urist 1962).In the years inbetween, research has advanced to a point that weunderstand by now the general pattern of calcification, but the localdetails are still missing (for reviews see : Degens 1976; Krampitz andWitt 1979; Westbroek and de Jong 1983).
Fossil products of biocalcification, i.e . shells, bones, teeth, are criticalelements, for example, to reconstruct the path of organic evolution, to

116 E. T . DEG E NS, J . K AZMIERCZA K & V. ITTE KKOT
outline paleogeographic settings, to determine type and dynamics offormer ecosystems, or to time geologic events. Thus, geoscientists shouldbe interested to know why organisms deposit carbonate skeletons andwhat cellular and environmen tal mechanisms drive organisms to depositcalc ite, ar agonite or Ca-phosphate (apatite). Such a knowledge couldassist to unravel the mystery behind the firs t appearance of calcareousf ossils at the end of the Precambrian era.
Ca! + REGULATIO N
Eukaryotic calcification is under strict cellular control and part ofthe biological calcium cycle. CaH is a. ubiquitous ion at vir tually allstrategic sites of an organism. The qu estion has often been asked whycalcium was chosen in first place to occupy such a central position inthe metabolism of the eukaryotic cell (Degens 1976; Williams 1976; Carafoli and Crompton 1978; Erulkar 1981). It is the structure of thecalcium ion which can perfectly adjust to fit all kinds of physiologicalassignments. This is principally du e to high affinity of calcium foroxygen ligands in complex .biochemical molecules, a characteristicunsurpassed by any other of the common chemical elements . Work tasksfor CaH range from structural to functional and the essentials arebriefly mentioned.
Structural aspects
The molecular geometry of phospholipid membranes bears strikingr esemblance to inorganic phosphate minerals in that in membranes andminerals corrugated layers of me tal ion-phosphate coordination complexesexist. Fig. 1 (top graph) depicts a phospholipid membrane cross-sectioncomposed of double agg regates of hydrocarbons and oxygen polyhedra.To reveal the type of molecular pattern that is exposed at the ionicsurface of phospholipid, the three-dimensional structure of two typicalphosphate minerals is shown (fig. 1) ex hibiting a sheet of phosphate-cation
Fi g. 1. (top) Schematic phospholip id membrane cross- section comp osed of doubleaggrega tes of hydrocarbons and oxygen polyhedra. The surface area of two hydroca rbon chains is about 40 A2 which is identica l to the surface area occ up iedby a tetrahedron-octahedron unit . T he 40 A peri od icity observed in biologicalmembranes is a re sult of this st r uctural de sign ; (center) presenta t ion of molecularst r ucture of a sheet compose d of phosphate-cation chains cross -linked into a pl anarpattern . The cor r ugated lay ers are simplified. In rea lit y, Ca2+ is coordinatedwith 8 ox ygen s and not with 6; (bo ttom) molecula r st ructure of vivian ite Fe3(P04h ·· 8 H20. Phosphate te trahedra link single and doub le octahe dral gr oups. Note thespace geometry of t he open channels in center and bottom gr aph whic h iscontro lle d by the ty pe of me ta l io n and coordinat ion number (from Matheja and
Deg ens 1971).
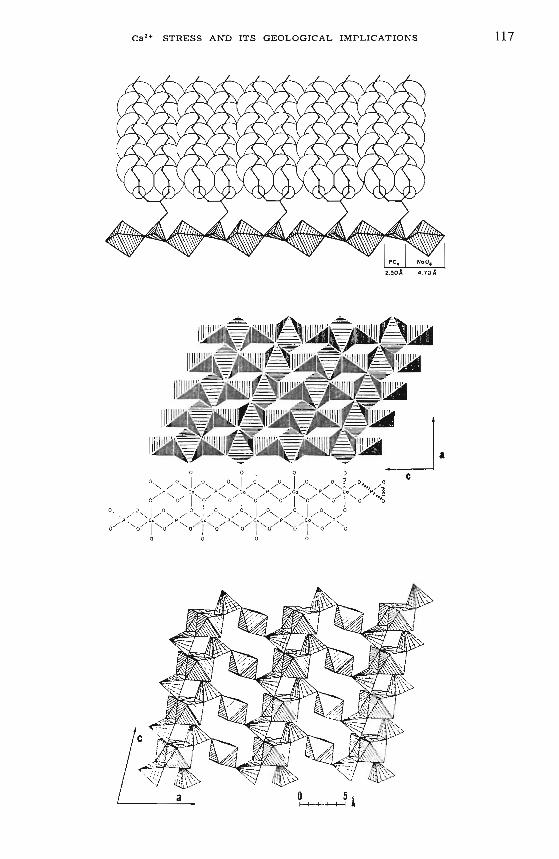
Ca2+ STRESS AND ITS GEOLOGICAL IMPLICATIONS 117

118 E. T . DEGENS, J . KAZMIERCZAK & V. ITTEKKOT
Fig. 2. Graph drawn after an electrom icr ograph of a beef-heart mitochon dr ia lcr oss-s ecti on. Outer gra in s represent st r uctural prot eins. The b lack lines belowthe protein grains reveal the position of the phosphate-metal ion oxygen po lyhedrawhich obey a 40 A period icity. Glycerol and double aggr ega tes of hydrocarbonsj oined to the cryst a lline tetrahedron-octahedron sur face occupy the spac e inbetween. Structural relationships between principal m olecular un its of a phospholipid membrane a r e depicted in the center of the f igure. For graphical rea sonsonly a cross-section through a single layer is shown and the hydrocarbon ch ainsare only partly drawn. The w ay Ca 2+ is fixed to carboxylate groups a t the
internal face of membranes is schemat ical1y presented.
chains crosslinked in a planar pattern. Note the open channels betweenthe tetrahedra-octahedra assemblages. The ionic surface of phospholipidmembranes is overlain by a protein layer which is coordinated by thesharing of oxygens to the ionic surface. Fig. 2 shows an electromicrograph of a beef heart mitochondrial cross-section next to which structuraldetails of a phospholipid bilayer are schematically drawn.
It is by now well established that the dispositions and functions ofmembrane proteins are the critical element in calcium transport andregulation. Binding of CaH ions to specific sites at the internal face ofmembran es al ters conformation of some membrane constituents, thusallowing transfer or passage of hydrophilic substances across the lipidbarri er. Membrane transport of a sugar carrier such as glucose is thusmedi ated by means of change of membrane permeability (Elbrink and

Ca2+ STRESS AND ITS GEOLOGICAL IMPLICATIONS 119
Bihler 1975). This transport is characterized by Michaelis-Menten saturation kinetics, chemical specificity and restructuring of substrate andmembrane components rather than by simple diffusion processes. Thecytoplasmatic calcium level is affected by CaH and Na " exchange inthe endoplasmic reticulum and by excitation-contraction coupling in thesarcoplasmic reticulum system which supplies CaH for muscle activities.Furthermore, rapid mitochondrial oxidation may deplete the regulatoryCaH pool which in turn is recharged by means of a feedback systemfrom storage sites inside the cell or through some form of energydependent "ion pump" system (Takeichi and Okada 1972; Elbrink andBihler 1975; Knudsen and Horwitz 1977; Rasmussen 1981; Llinas 1982).The close association of Na + and CaH in the "ion pump" system couldbe a consequence of similar ionic radii (close to 1.0 A) and high affinityof both ions to oxygen in octahedral coordination.
The way CaH is fixed to carboxylate groups exposed at the hydrophobic site of th p membrane protein is schematically presented infig. 2. CaH can open or close off communication channels through therestructuring of the membrane fabric. The interchangeable nature ofmetal ions causes membranes to act as dynamic molecular sieves. Poresize and sieving qualities are variable and depend on metal ions regulatedby enzymes. Assuming ATP traps a met al ion, such as CaH , aperiodicpulsation of the lattice will take place. Such oscillations are knownfrom mitochondria (Chance and Yoshioka 1965). The surface lattice ofmembranes acts on the phospholipid molecules in a manner similar toa zone melting and refining process. It can be considered a two-dimensional coding device for bonded proteins, whereby CaH serves as amessenger. Permeability is achieved by placing the Ca2+ at openinggates of the membrane structure where it acts as modulating factorfor material flux, ene r gy transfer and excitability (Matheja and Degens1971).
Functional aspects
Ca2 + serves as "an important cellular r egulator. Its message is relayedby a series of high-affinity calcium binding proteins of which calmodulin,troponin C (TN-C), and parvalbumin are thr ee common sp ecies (e.g.,Collins et aL 1973; Drabikowski et al. 1974; Kendrick-Jones 1974; Tuftyand Kr:etsinger 1975; Kretsinger 1976; Cheung 1980, 1982).
It was su ggested that many proteins involved in calcium transport andregulation are homologous and would contain one or several "EFhands" . The ubiquity of EF-hands may simply reflect a thermodynamicallypreferable conformation. Fig. 3 illustrates the basic structural eleme ntsof the EF-hand. Comparison of EF-hand amino acid sequences in severalhigh-affinity calcium binding proteins reveals the significance of the
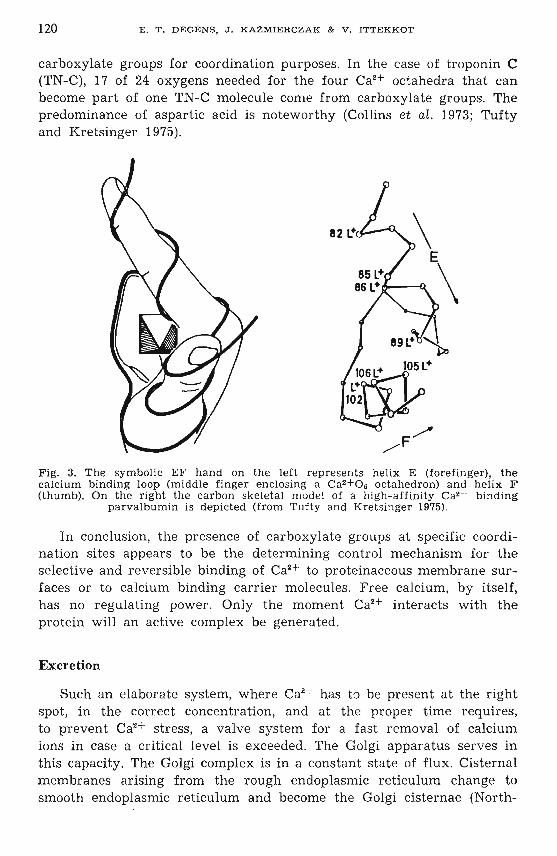
120 E . T. DEGENS, J. KAZMIERCZAK & V . IT T E K K OT
carboxylate groups for coordination purposes. In the case of troponin C(TN-C), 17 of 24 oxygens needed for the four CaH octahedra that canbecome part of one TN-C molecule come from carboxylate groups. Thepredominance of aspartic acid is noteworthy (Collins et al. 1973; Tuftyand Kretsinger 1975).
Fig. 3. The symbolic EF h and on the left represents helix E (forefinger), thecalcium binding loop (middle finger enclosing a Ca2 +06 octahedron) and helix F(thumb). On the right the carbon skeletal m odel of a hi gh-affinity CaH binding
parvalbumin is depicted (from Tufty and Kret singer 1975).
In conclusion, the presence of carboxylate groups at specific coordination sites appears to be the determining control mechanism for theselective and reversible binding of CaH to proteinaceous membrane surfaces or to calcium binding carrier molecules. Free calcium, by itself,has no regulating power. Only the moment CaH interacts with theprotein will an active complex be generated.
Excretion
Such an elaborate system, where CaH has to be present at the rightspot, in the correct concentration, and at the proper time requires,to prevent CaH stress, a valve system for a fast removal of calciumions in case a critical level is exceeded. The Golgi apparatus serves inthis capacity. The Golgi complex is in a constant state of flux. Cisternalmembranes arising from the rough endoplasmic reticulum change tosmooth endoplasmic reticulum and become the Golgi cisternae (Nor th-

C a2+ STRESS A ND ITS G EOLOGICAL IMPLICATIO NS 121
cote 1971). Those cisternae then break down to ve sicles which can fusewith and extend the plasmalemma. Therefore, the Golgi apparatus maybe viewed as a m embrane tr ansformer from the endoplasmic reticulumto the cell m embrane. During this irreversible process proteins aresorted, refined , and de liver ed to multipl e destinations.
Among the numerous functions the Gol gi exercises in the eukaryoticcell (Rothmann 1981) its responsibili ty for ex cretion , including ther emo va l of excess CaH from wit hin the cell to the outside environment,has a direct bearing on the mechanism of calcification. A high-affinitycalcium binding protein se rves as a carrier for surplus CaH. The proteinbec om es solubilized by the addition of glucuronic acid or a similaracting sugar (Degens 1976). The newl y assembled glycopro tein complexis now r eady for shipment (excreti on ) to the outside enviro nment.
CALCIFICATION
Everything would be alr ight, if the aquatic habita t wo u ld follow theconcept: "The solution to pollution is diluti on ". Yet , in a wide va rietyof invertebrates , protein sheets positioned at or close to the ce ll wallsinteract with the CaH -charged complex and carbonate dep osi tion isinitiated. The mechanism is as follows.
The outer protein sheet of a silk- type nature develops hydrogenbonding to the hydrophilic side of the glycopr otein complex followingthe r emoval of the glucuronic acid by means of a proteas e. The nowfreel y ex posed hydrophobic side composed principall y of carboxyla tegroups derived from aspartic acid will a tract additional calcium whichbecomes structured in the form of CaH0
6 or CaHO g coordination polyhedra. CaC03 deposition is mediated by carbonic anhydrase whichfunc tions as a kind of "che mical hom eostat" by r eleasing calcium fromcellula r depots and by activating car bonate from the aque ous m edium,The role of carbonic anhydr ase in calcification does not rest on it sability to hydrate CO 2 , but to re move carbonic acid fr om the site ofcalcifica tion (Goreau 1959):
Ca 2 + + 2 HC03 -+ Ca(HCO)3 -+ CaC03 + H2C03.
Ex pre ssed differentl y , the substr ate of ca r bonic anhydrase is the neutralH 2C0 3 mol ecule (Koenig and Brown 1972). As long as calcium is n otthe limiting fa ctor , the rate of for mation of calcium carbonate willdep end on the rate by which carbonic acid is carried off from theca lcification site. Sin ce the over all crystallisation of CaC0 3 will liberate2- 4 kcal/mole, there is no energy problem in CaC0 3 dep osi tion . Inthe pr esence of carbonic anhydrase inhibitors, carbonate formation slowsdown and eventua lly ceases (Goreau 1963) .

122 E. T . DEGENS, J. KAZ MIERCZA K & V . ITTEKKOT
The structural relationships that ar ise during calcification are brieflydescribed. The attachment of the Ca2 + carrier to a protein template willactivate the mineralizing subs trate and calcium carbonate crystals arelaid down in epitaxial order. Crystals are nucleated on stereochemicallyanalogous faces when proteins are adsorbed onto a rigid substrate (Addadiand Weiner 1985). Crystal growth te rminates upon secretion of a newlyformed Ca2+ carrier protein on the gro wing crystal surface, whereb ya kind of organic blanket is formed . Subsequen tly, the protein templatebecomes attached to the hydrophilic sid e of the Ca2+ carrier and asandwich str uct ure is produced (fig . 4). The cycle can start again.Thousands of layers can be generated during the life time of an organism(e.g. in a mollusc she ll) . Fig. 5 depicts a vertical cross-section of alternatinglayers of the two typ es of proteins.
F ig. 4. Sch ema t ic represen ta t ion of st r uc tu ra l re lat ion ship between layers ofs i lk - type templa te (ban ded) , h igh-a ff in ity CaH b indi ng prote in (d otted) a nd CaC03
(bl ock- sh a ped ) (f rom Degen s 1976).
F ig. 5. Sc hemati c repr esen ta t ion of ver t ica l cross- sect ion show ing a lt e rna t inglayer s of h igh affinity CaH bin d in g p r ote in s (hexag on s) a n d sil k - ty pe template(plea te d sheet con fo r mation ) ; t he CaC03 ha s been omi tted for gr aphical rea son s
(f rom Degen s 1976).

Ca2+ STRESS A ND ITS GEOLOGICAL IMPLICATIONS 123
The key role CaH exercises in cellular regulation is in the lastinstance the reason behind calcification. Biocalcification has been probably forced onto the evolving eukaryotic cell as a consequence of thebuild-up of CaH in the ancient sea (Kazmierczak et a]. 1985). It wasprincipally invented not for storage of calcium, protection against predators or environmental hazards, but is a byproduct of excretion of excessCa 2+ from within the cell. That the or ganisms took later advantage of it,is a different story.
BIOGEO CHEMICAL IMPLICATIONS
CaC03 forms principally along two routes: (i) a chemical one:
Ca2+ + 2HC03i~ CaC03 + CO 2 + H20
and (ii) an enzymic one which instead of CO 2 and H20 yields H2C03
Ca2 + + 2 HC03 ~ Ca(HC03h ~ CaC03 + H2C0 3 •
The carbonic anhydrase mediated CaC03 deposition is restricted to theeukaryotic cell and came into existance with the onset of biologicalcalcification at about the Precambrian/Cambrian boundary. It is emphasized that microbially induced carbonate deposition of the type associated,for example, with stromatolites of Precambrian or Phanerozoic age isstrictly chemically controlled. This has interesting consequences for carbonisotope geochemistry (Degens et al. in press) because a chemically precipitated CaC03 will form in isotopic equilibrium making the solid phase 4per mil heavier than the starting bicarbonate. Expressed differently,a common 5 13C value of -1 per mil for Precambrian limestones (Schidlowski 1982) would signal a - 5 per mil for the former marine bicarbonate pool. In contrast, the majority of skeletal carbonates have a 5 13Cidentical to its sea water bicarbonate source. Only a few species existwhere some contribution of isotopically light respiratory CO 2 is indicated.The approximately 2 per mil drop in 5 13C at about the Vendian timecould principally be related to a switch from chemical to enzymatic carbonate deposition.
As mentioned above, the first calcification events during Vendian areconsidered the r esponse of the eukaryotic cell to higher CaH levels in theancient sea. The question comes up why has the CaH pressure not beenfelt by the organisms earlier in time? We believe that the answer restsin the "pe culiar " chemistry of the Precambrian sea. It was suggested(Degens et al. 1984; Kempe and Degens 1985) that not rock saltbut soda was the major solute in the earl y ocean. A general chemistrysimilar to that presently found in lakes from volcanic r egions is envisioned.Lake Van , the largest sod a lake on Earth is regarded the prototype of

124 E. T . DEGENS, J. KAZMIERCZAK & V. ITTEKKOT
a primordial sea (Kempe 1977) . Soda Jakes contain little CaH , becauseresidence time of calcium is extremely short, due to its fast removalfrom the water column in the form of aragonite, calcite or dolomitedepending on Mg/Ca ratio. Soda lakes are highly alkaline with a pHin the range of 9.5 to 10.5. This has interesting consequence for the fateof CO 2 in the early atmosphere. In a matter of less than 100 millionyears, all carbon .in the crust, if released in the form of CO 2 , would beconsumed by weathering and could readily be accomodated as dissolvedcarbonates in a soda ocean (Kempe and Degens 1985). Only a moderateamount of CO 2 would remain airborn and PC02 would be at most a fewtimes higher than today.
In the course of the Precambrian, the initial alkaline sea becametitrated with acid volatiles and pH dropped down to about 8 (fig . 6).
9pH
8
10/'-fl-I-I-II111--------/
/ Soda OceanI
IIII,,Soda
100%....u:¥.:...L.:>....<...:~~.<.+'-"-':..>...L¥"""'-':...L..1~""'-"-.,..'"'"-"~¥>...£-.'~~..c....>....'-¥c.>..L..>o..L.>.f
f Initial Equilibration
100 %,-.-T""""T-.-T""""T-.-......-:=:-=--..,.......,.-.,...-,.,...,...-rT,...-T"""""T--.-T""""T--r::::::::!I'~-=~
Halite
4 3 2Ga B.P
1 oFig. 6. Evolution of ocean chem ist r y during geologic time (pH-curve deduced fromtotal a lka n ity and pH-rela ti on of present-day soda lak es (from Kempe and
Degens 1985).
This marked the time when water chemistry shifted from soda to sodiumchloride dominance, and the dissolved carbonate system changed fromsodium to calcium-magnesium regulation. CaH started to increase andwhen a critical point was r eached where the eukaryotic cell had toswitch on the Golgi for the excretion of excess CaH , skeletal minerals
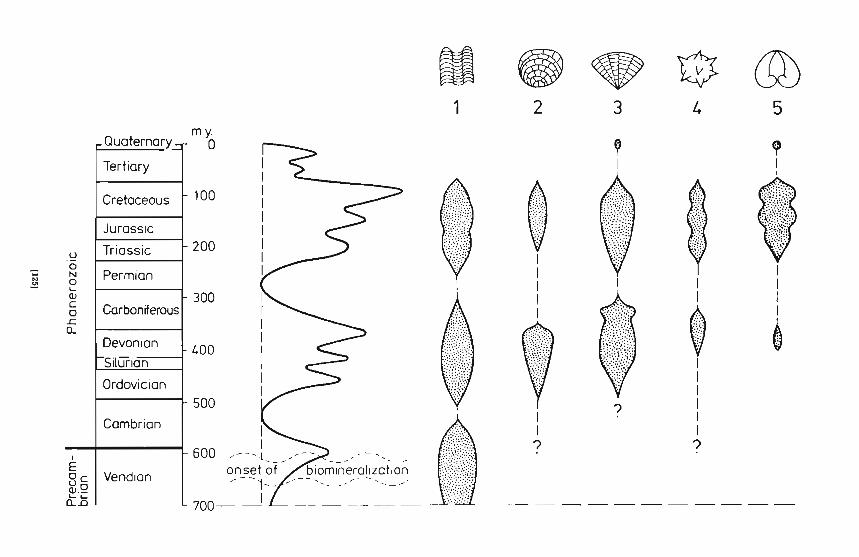
Fig. 7. Curve of rel at ive abunda nc e of ma ri ne ca r bona te sedi men ts during thePhan erozoic (compiled fr om va ri ous sou rces) compared w ith strat igraphic di stribution and fr equ en cy of selected groups of organisms and biosedimentary structures conside re d as good indicato rs of a ca lcium in toxi ca ted environ ment. 1 ca lcareous st ro matoli tes in general , 2 - st ro mato poro id stromato li tes: subti da l, insi tu calcifi ed coccoid cya no ba cter ia l mats typical part icularly for lower Palaeozoicca r bona te platforms. 3 - scl e rosponges: siliceous sponges (Dem ospongia e) withaddi t iona l ba sal ca lca reous skeleton ; typi cal fossil representative s are favo sit idsand chaetet ids , 4 - ca lci sphe r ic str uc tu res : mos tly post mortem ca lcifi ed phytop lan kt ers like ac r ita rc hs or vo lvocacea n algae or calcified rest ing cysts ofdi noflage lla tes abundant in protect ed areas of pa st carbonate pl a tf orms, 5 thi ck-shell ed molluscs (e.g., megalodontid and ru dist bivalves or nerineid ga st ro -
pods) (from Kazmierczak et al. 1985)
.. il .$@1 2 3 4 5
IIII
o
,II
II
entIIIIII
?
?I
?II
?
t100
300
500
200
m.y.o
600
400
Quaternary'--=Tert iary
Cretaceous
Jurassic
. ~Triassic
0N Perm ian0'-QJC Carboniferousos:Q..
Devonian
-Silurian
Ordovic ian
Cambr ian
I
E°c VendianU oQJ.-.... '-Q...o
...~

126 E . T . DEGENS. J. K A Z MIERC ZAK & V . ITTEKKOT
started to form and an almost spasmodic evolution of the skeletonizedbiota began.
Massive marine limestone deposits observed at certain times duringthe Phanerozoic are in all likelihood linked to an effective recharge ofCaH to the sea through riverine and hydrothermal channels and itsconcentration along shelf margins (Kazmierczak et al. 1985). This causeda number of shallow water organisms to calcify (which otherwise didn't)and others to lay down thicker skeletal deposits. In fig. 7, the relativeabundance of five fossil groups indicative for CaH stressed marine environments are illustrated: (1) calcareous stromatolites, (2) stromatoporoid stromatolites, (3) sclerosponges, (4) calcispheres, and (5) thickshelled molluscs. All exhibit the same distribution pattern and goodcorrelation with maximal extensions of Phanerozoic carbonate facies.The Phanerozoic limestone facies developed mostly as so-called carbonate
. platforms or ramps which are considered comparatively stable off-shoreshallow marine habitats in an epicontinental sea comparing with on-shoresettings. Of particular interest are calcispheres, a heterogenous group ofphytoplankters (volvocaceans, acritarchs, dinoflagellates) mineralized notin vivo but onl y post mortem (Kazmierczak 1976). They occurred atCaH peaks and concentrated excessive calcium in their extracellularexcretes (chelants) which after cell death calcified immediately closethe sediment/water interface. Fig. 8 depicts the same forms of acritarchsin an unmineralized state from calcium depleted black shale facies andin CaCO J encrusted "calcispher ized" state from carbonate facies.
CaH stress exercised by the environment on the evolving organismseventually reached a stage that ev en unicellular eukaryotes started tocalcify. The em ergence of calcareous nannophytoplankton and of plankticforaminifers during the Jurassic may be viewed as a response to thischallenge. In the Cenozoic, areal reduction of carbonate platforms leftthe burden of CaH removal to the pelagic plankton community. Thisis precisely the s.euation we are left today.
The fact that even today many normally calcifying organisms stopCaCOJ deposition at times of en vironmental stress (e.g. Lutz and Rhoads1977) or during active stages when CaH is in high demand (e.g. Hemleben et al. 1979) may serve as a reminder that calcific ation ranks inlow priority for the metabolism of the organism and is just the cellularresponse to an environmentally induced CaH stress.
NON-CALCIFYING ORGANISMS
One aspect of cellular Ca2+ regulation has not been mentioned so far:What do non-calcifying organisms do when subjected to a CaH stress?Th e answe r is : the same! Th ey secret e glycoproteins, coordinate CaH

CaH S T RESS AND ITS G EOLOGICAL IMPLICATIONS 127
Fi g. 8. Examples of variously preserved ac r it a rch phytoplankters (Baltisphaeridium grou p) depending on basically d ifferent ca lci um availabili ty in t he ir life andbur ia l envi r onment; (top) organ ic- wa lled specimen from calcium depleted en v ironment (Chagrin S hale, Upper Devonian , Oh io; cou rtesy Dr. E. R. Wican de r);bar scale = 10 11m; (bottom) heavily calcified specimen with poo rly preservedorgani c wall from calcium str ess environment (F r asn ian carbonate platform, Upper
Devonian , southern P olan d); bar scal e = 50 11m.
and excrete a high affinity Ca-binding protein , except that there isno proteinaceous cell wall template to hold back the dissolved complex .
In a city lake, the Hamburg Alster, an about gO-fold supersaturationwith r esp ect to calcite ex isted at the time of sa mpling. We examine d

128 F.. T. D EGENS, J . KA ZMIERCZAK & V. ITTEKKOT
the molecular nature of the excretion products released to the environment during a non-calcareous algal bloom (Michaelis et al. 1976). Of thetotal proteinaceous material excreted by the plankton about 90% consisted of aspartic acid attesting to the universiality of this amino acidfor CaH -fixation. In such a situation rough ly half of the daily photosynthesized organic matter ended up as dissolved organic carbon whichbecause of its high functionality would readily combine with clayminerals forming macroflocs a few hundred microns in size. Thesesettled almost instantaneously to the sediment-water interface, yieldingan organic-rich mud.
Similar interaction occur in the open sea and a variety of organicexcretion products have been reported (Anderson and Zeutschel 1970;Thomas 1971; Hellebust 1974; Williams 1975; Fogg 1983). It is noteworthythat clay minerals on their own would almost never reach the seafloor should they only rely on Stoke's Law. The physical settling velocities of clays are in the order of a few em per day. Convection in theeuphoti c zone measuring a few meter per day would thus keep clays insuspension. However, when aggregated to macroflocs (Kranck and Milligan 1980; Honjo 1982), or packaged in the form of fecal pellets, after beingeaten by zooplankters (Schrader 1971; Honjo 1976; Roth et al. 1975;Honjo and Roman 1978), clay aggregates will settle at rates of a fewhundred meters per day (fig. 9). Material collected in sediment trapsin mid-water at depths of several thousend meters demonstrate thatclay-sized fractions are removed from the surface layer via macroflocsor fecal pellets. There are strong seasonal and interannual variationswhich are directly related to the pulses of planktic activity in the sea(Deuser and Ross 1980; Deuser et al. 1981, 1983; Honjo 1982).
Interestingly, this transport mechanism affects not only the verticaltransport of particles. Horizontally transported particles in mid-waterare also found to be translated into a vertical flux (Honjo 1982). It hasalso been suggested that sinking macroaggregates will scavange particlessuspended in the water column (Deuser et al. 1983) increasing the abiogenic material flux to deep ocean (Honjo et al. 1982).
In ancient environments devoid of such mechanisms before the adventof zooplankton and organic matter secreting planktic algae, sedimentation processes must have been significantly different and more inagreement with Stoke's Law. Higher residence times for particles willbe the result and as a consequence, more intense particle-seawater interactions should occur. In such an ocean, dissolution or ion-exchangeof si licate minerals with seawater column (Tsunogai et al. 1973) wouldhave been an efficient source for dissolved Ca2+. Conditions similar tothis were probably repeated also in the Phanerozoic during periods oflarge scale extinctions of marine planktic biota. At those times, theterrestrial detritus content in the ocean would have been enormous

F ig. 9. Settl ing rates per day of materi a ls in various size fraction in t he marine envi r on ment (fr om Degen s and Ittekkot 1984).
(129)
2 Ac ta P al aeontologica Polonic a Nr 3-4/85

130 E . T. DEGE NS, J . KAZMIERCZAK & V . ITTEKKOT
because of the absence of efficient and rapid transfer mechanisms suchas fecal pellets and macroaggregates. Only particles with high specificdensity, like volcanic and cosmic dust may be expected to sink rapidlytowards the seabottom leading to geochemical anomalies.
Macroaggregate formation induced by calcium specific organicmatter will also lead to the clustering of cells. It has been found thatpolysaccharide-rich excr etion products of coccolithophorids are significant in the formation of cell macroaggregates (Ittekkot et at. 1984). It isconceivable that such macroaggregate formation could have been instrumental in the early steps of multicellular evolution (Kempe et at. 1986) .
CONCLUSIONS
CaH in the sea is derived from the principal sources: (i) rivermerun-off, and (ii) hydrothermal solutions emanating from the oceaniccrust. For most of the Precambrian removal of CaH was principallyaccomplished by carbonate precipitation in tidal and supratidal environments (e.g., in the form of stromatolites, dolostones) or in thevicinity of rifts. Seawater chemistry at that time, that is a "soda ocean",forced Ca2+ to precipitate close its marine inlets.
With the advent of a NaCI dominated ocean, somewhere during theProterozoic, CaH level in sea water could rise and the dissolved carbonate chemistry became governed by CaH - MgH mineral equilibria.Organisms responded to this CaH challenge later in the Precambrian bythe formation of multicellular aggregates and even tually in Vendiantime through skeletonization.
The biocalcification event changed carbonate deposition from a chemical to a predominantly enzymic process . In accordance, the extractionof CaH from the Phanerozoic sea became the domain of organisms.
Changes in scale of calcification as seen in weight and morphologyof shell structures are in all likelihood intimately connected with tectonicevents which supplied CaH directly through hydrothermal channels orindirectly via erosion to the marine environment.
Non-calcifying marine protists, e.g. the nannoplankton, respond toCaH and other metal ion stress by secretion of complexing moleculessuch as polysaccharides or glycoproteins. Such complexing agents interact with suspended particles and lead to macrofloc formation. Sedimentation of clay-sized suspensions in the open marine habitat is thusmediated by the activity of planktic organisms. Thus, in the finalanalysis, marine limestones and shales are products of organic-inorg anicinteractions.

Ca· 2 STRESS A ND ITS G EOLOGI C AL IMPLICATIONS
ACKNOWLEDGEMENTS
131
The w ork ha s been suppor ted by the Bundesministerium flir F or schung undTechnologie, Bonn , F .RG., and the Shell Grants Committee , London, U.K. On eof us (J . K.) gratefully ac knowle dges a fellow ship of the Alexander-von-Humboldt F oundation .
REFERENCES
ADDADI , L a nd WEINER, S. Interaction between acidi c proteins a nd crystals :ster eochemi ca l requirements in biomineralizati on . - Proc, Ntitl: Acad SciUSA , 82, 4110-4114.
ANDERSON, G. C. and ZEUTSCHEL, R. P . 1970. Release of di ssolved or ga nicmatter by marine phytoplankton in coast al of fs hor e a r eas of the Northea stPacific Ocean. - Limnol. Oc eanogr., 15, 402--407.
CARAFOLI, E. and CROMPTON, M. 1978. The regulati on of intracellular ca lciu m. Cu rrent Topics in Membrane Transp ort, 9, 151-216.
CHANCE, B. and YOSHIOKA, T. 1966. Sust ained osc ill a tion of ion ic consti tue n tsof mit och ondria. - Archiv Biochem. B iophy s., 117, 451--464.
CHEUNG, W. Y. 1980. Calmodulin pl ays a p ivot al r ole in cellular regulat ion. Science, 207, 19-27.
- 1982. Calmodulin. - Sci. Amer., 247/6, 48-56.COLLINS, J. H. , POTTER, J . D., HORN, M. C. J. , WILSHIRE, G. and JACK
MAN, N. 1973. The amino acid sequence of r abbit skeleta l muscle troponin C:Gene replication and hom ology w ith calcium-binding protein s from car pa nd hare muscle. - FEBS Letters, 36, 268-272.
DEGENS, E. T. 1976. Molecular mechanisms on carbonate, phosphate and sil icadeposition in the living cell. - Topics in Current Chemi stry, 64, 3-112.an d ITTEKKOT, V. 1984. A new look a t clay-organi c in teractions. - Mitt.Geol.-Paliiont. I nst. Univ. H amburg, 56, 229-248.KAZMIERCZAK, J . an d ITTEKKOT, V. Biom in eralizat ion a nd the ca rbonisoto pe r ecord. - (TMPM T schermak's Min eral. Petrogr. Mitt. (in press).KEMPE, S. an d SPITZY, A. 1984. CO 2 : A biogeoch emical portra it . In: O. H utzin ger (ed .), H andbook of Envir onmental Ch emistry, 127-215, Springer-Verlag,Berlin.
DEUSER, W. G. and RO SS, E. H. 1980. Seasonal ch ange in the flux of organicca r bon to the deep Sargas so Sea. - Nature, 283, 364-365.
- , - an d ANDERSON, R F . 1981. Seasonality in the supply of sediment to thedeep Sargasso Sea an d implication s for the r apid transpo r t of matter to thedeep ocean. - D eep Sea Res ., 28A, 495-505.BREWER, P. G., JICKELLS, T. D. and COMMEAU, R. D. 1983. Biologicalcontrol of the remov al of abiogenic particle s fro m the surface ocean.Science, 219, 388-391.
DRABIKOWSKI, W., BARYLKO, B., DABROWSKA, R , NO WAK, E. an d S ZP ACENKO, A. 1974. Studies on t he propertie s of TN-C component of t ro po ni nan d on its effects on the interact ion between the con stituen t s of t hi nfilame n t. In : W. Drabik ow sk i, H. Strz el ecka-Gola sze ws ka an d E. Ca rafoli(eds.), Ca lcium Binding Prot ein s, 69-107, El sev ier, Amsterdam and P WN P oli sh Sci entific Publ., Warszawa.

132 E. T . DEGENS, J. KA ZMIERCZAK & V. ITTE K KOT
ELBRINK, J . a nd BIHLER. 1. 1975. Membrane tra n sport : Its r el a t ion to cellu la rmetabolic rates. - Sci en ce, 188, 1177-1184.
E RU LKAR, D. 1980. The ver satile r ole of ca lci um in b iologica l sys te ms. - I nter disciplinary Sci. Rev ., 6, 322-332.
F OGG, G . E. 1983. The ec olog ical sign if ica n ce of extracell u la r product s of phytoplankton p ho to syn thesi s. - Botanica Marina, 26, 3-14.
GOREAU, T. F . 1959. The phy siology of skele ton fo r mation in corals. 1. A methodfo r m easur ing the r a te of ca lciu m deposi ti on by corals u n der differentcondi t ions . - BioI. Bull. , 116, 59-75.1963. Ca lcium ca r bo nate deposit ion by coralli ne a lgae and corals in rel a t ionto the ir r ole s as reef b u ilders. - Ann . N.Y. A cad. Sci. , 109, 127-167.
HELLE BUST , J . A. 1974. Ex tracellular p ro duc ts. I n : W. D. P. Stewart (ed.), AlgalP hysiology a nd Biochemistry, 838-863, Blackwell , London.
HEMLEBEN CH., BE , A., SPINDLER, M. a nd ANDERS ON, O. R. 1979. "Dissolut ion" effects induced by she ll re sorp t ion duri ng gametogenesis in H ast iger inapel ag ica (d' Or bigny) . - J. F or am. Res., 9, 118-124.
HON J O, S. 1976. Coccoli th s : Production , t ra nspor ta t ion a nd sedi mentation. -r-' Mar.M icr opaleont ., 1, 65- 79.1982. Seasonali ty a nd interacti on of b iogenic a nd lithogenic particu la tefl ux at P ana m a Basin . - Science, 218, 883- 884.a nd ROMAN, M. R. 1978. Marine copepo d fe ca l pellets: Production , preserv atio n a nd sed im en ta tion . - J. Mar. Res., 36, 516-518.SP ENC ER, D. a nd FARRINGTON, J . W. 1982. De ep a dvect ive transp ort oflitho gen ic p ar ticles in Panama Basin . - Sci ence, 216, 516-518.
ITTEK KOT, V.. DEGENS, E . T. an d HONJ O, S. 1984. S ea sonality in the flux esof suga rs. a m ino acids a nd amino suga rs : P anama Ba sin . - Deep Sea Res.•31, 1071-1083.
K AZMIERCZAK, J . 1976. Volvoca cean n at ure of some no n- radiosphaeri d cal cispheres and para thura mminid "Fora minifera". - Act a Palaeon t. Poloni ca , 21,245- 258.
ITTEKKOT, V. and D EG ENS , E. T . 1985. Biocalcifi cati on through ti me : Envi ro nmen ta l ch all enge a n d ce ll ular resp onse. - PalCion t. Z t schr. , 59, 15-33.
K EMPE, S. 1977. Hydrographie , Warven-Chronolog ie und or ga nische Geochemiedes Va n Sees, Ost-Ttirkei. - M itt. Geol. Paliiont. Inst. U n iv . Hamburg ,H , 125- 228.
. a nd DEGENS, E. T. An ear ly soda ocean? - Che rn. Geol. , 53, 95-108.KA ZMIERCZAK, J . a nd DEGENS , E. T. 1986. The soda ocean con cept a ndit s bear ing on b iot ic a nd cr us tal evo lution . - Fifth Int. Symp. Bio m inera l.The Universit y of T exa s a t Ar lington , May 18-23, 1986, Abstra ct s.
KENDR ICK- J ON ES, J. 1974. Role of myosi n light chains in ca lc ium regula tion . Nature, 249, 631-{j34.
KNUDSO N, K. A. and HORWITZ, A. F . 1977. Tand em events in myoblast fusio n . Del: . BioI. , 58, 328- 339.
KOENIG , S . H . a nd BROW N, R. D. 1972. H 2CO J as substra te for ca r bonic a n hy drasein the dehydration of HCO~ - - Pr oc. Nail: Acad. Sci. U SA , 69. 24222425.
KRAMPITZ, G . a n d WITT, W. 1979. Bi ochemical a spec ts of biomine ralization . Topi cs in Curren t Chemist ry , 78, 57- 144.
K RANCK, K. and MILLI G AN, T . 1980. Macrofl ocs : Product ion of m arine sn owin the lab ora to ry. - Mar. Ecol. Progr. Ser ., 3, 19-24.
KRETSINGER, R. H . 1976. Cal ciu m bindi ng prote in s. - A nn ual Rev. Biochem.,
45, 239- 262.

Ca +' STRESS A ND ITS G EOLOGICAL IMPLI CATIONS 133
LLINAS, R. R. 1982. Calcium in syna pt ic transmi ssion. - Sci. Amer. , 247, 4,38--47.
LUTZ, R. A. a nd RHOADS, D. C. 1977. Anaerobiosis a nd a theory of growth lineformation. - Science , 198, 1222-1227.
MATHEJA, J. and DEGENS, E. T . 1971. Structural Mole cular Biology of Phosphates, 180 pp. Gustav Fischer, Stuttgart.
MICHAELIS, W. , MOPPER, K., GARRASI, C. and DEGENS, E. T . 1976. OrganischeSubstanzen im Sediment und Wa sser der H amburger Al ster. - Mitt . Ceol.Paleiont. Inst . Univ. Hamburg , 46, 173-188.
NORTHCOTE, D. H. 1971. The G olg i apparatus. - Endeavour, 30, 26-33.RASMUSSEN, H. 1981. Calcium and cA MP as Synarch ic Me ssengers, 370 pp . Wiley
Interscience, New York.ROTH, P. , MULLIN, M. M. and BERGER, W. H. 1975. Coccolith se di m en ta tion
by fecal pellets: laboratory experimen ts a nd field ob servation s. - Bull.Ceo!. Soc. Amer., 86, 1079-1084.
ROTH MANN, J . E. 1981. The Golgi a ppa ra t us: Two orga nell s in tandem. Science, 213, 1212-1220.
SCHIDLOWSKI, M. 1982. Content and isot opic composit ion of reduced carbonin sediments. In: H. D. Holland and M. S ch idlowsk i (ed s.), Min eral Dep osit sand Evolution of the Biosphere, 103-122, Springer-Verlag, Berlin.
SCHRADER, H. J. 1971. Fecal pellets in sed im en ta t ion of pelagic d ia toms. Science, 174, 55-57.
TAKEICHI, M. a nd OKADA, T. S. 1972. Roles of m agnesium and ca lciu m ions incell-to- substrate adhesion. - Exp. Cell. Re s., 74, 51-60.
THOMAS, J . P . 1971. Relea se of di ssolved organic matter from natural p opulati on sof m arine phytoplankton . - Mar. BioI ., 11, 311-323.
TSUNOGAI, S ., YAMAHATA, H. , KUDO, S. a n d SAITO, O. 1973. Calci u m in thePacific Ocean. - Deep Sea Res. , 20, 717-726.
TUFTY, R. M. an d KRETSINGER, R. H. 1975. Trop oni n a nd parvalbumin ca lciu mbinding reg ion s predicted in myosin ligh t chain a nd T4 ly sozy m e. - Science,187, 167-169.
URIST, M. R. 1962. Induced syst e m a t ic hypersensit iv ity. Selye 's t heory. - Ib idem,137, 120-121.
WESTBROEK, P. an d de J ONG, E. W. (ed s.). 1983. Biomineralizat ion a nd Biologic al Met a l Ac cumulation , 533 pp. D. Reid el, Dordrecht.
WILLIAMS, P . J . Ie B. 1975. Biologi cal and chemica l a spec ts of d issolved or gan icm a terial in sea w a ter. I n : J. P. R iley a nd G . Skirrow (eds.), ChemicalOceanography, 2, 301-363, Academ ic Press, London.
WILLIAMS, R. J. P. 1976. Calcium chemi st ry a nd it s relat ion to biologica lfunction . - Symp. Exp. B i ol. , 30, 1-17.
EGON T . D E G E N S • .JOZE F KA Z MI E R C ZA K , VENUGO P A L A N ITTEKKOT
GEOLOGICZNE KONSEKWENCJE REAKCJI KOMORKI NA STRES WAPNIOWY
St r eszczenie
CaH jest jednym z najwai.n iej szych [onow w proce sa ch zyciow ych, pelniacyrn,
m ie dzy innym i, ro zn orakie fun kcje jako regulator wewnatrzkornorkowy. Prze
cietne stezen ie Ca2+ w cytopla zmie wynosi 10- s-10-7 M i jest zw yk le 0 4-5 r ze-

134 E. T . DEGENS, J. KAZMIERCZAK & V. ITTEKKOT
d6w wielkosci nizsze od stezen zewnqtrzkom6rkowych. Wysokie st ezenia jon6w
wapnia w sqs iedztwie kom6rki powodujq zwiekszony naplyw Ca 2+ do cytoplazmy,
zagrazajacy normalnemu fun kcjonowaniu organizrnu. Do usuwania toksycznego
nadmiaru Ca 2+ z kom6rki sluza roznego rodzaju "pompy jonowe" oraz nosniki
bialkowe (tzw. kalcyproteiny). Biokalcyfik acje mozna rozpatrywac jako [edna
z form detoksykacji wapniowej kom6rek. Rozni sie ona od innych spo sob6w usu
wania nadmiaru Ca 2+ z kom6rek tym, ze w tym przypadku wyprowadzany na
nosniku bialkowym (glikoproteinowym) wapn jest przy w spoludziale enzym6w
(weglanowe] anhydrazy lub alkalicznej fosfatazy) neutralizowany do postaci slabo
rozpuszczalnej w wodzie soli, kt6rej ostateczna forma mineralogiczna zalezy od
specyficznosci wzornika (template) bialkowego, na kt6rym epitaksjalnie narastaia
krysztaly.
Om6wione w pracy dokladniej mechanizmy transportu i regulacji Ca 2+ w ko
m6rce posluzyly do poparc ia hipotezy Ka imierczaka, Ittekkota i Degensa (1985),
laczace] powstanie pierwszych struktur szkieletowych na przelomie prekambru
i kambru ze wzrostem stezen Ca2+ w szelfowych srodowiskach morskich do po
ziomu toksycznego dla zasiedlajacych je organizm6w. Zgodnie z ta hipoteza, w ciq
gu fanerozoiku koncentracja Ca2+ w morzach szelfowych ulegala wielkoskalowym
fluktuacjom, odpowiada jqcy m cyklom regresywno-trun sgresywnyrn, sterowanym
aktywnoscia stref ryftowych. Rezultatem tych fluktuacji byloby okresowe, masowe
pojawianie sie w prawie w szystkich grupach organizrnow morskich form opa
trzonych szczegolnie masywnyrni, wapiennymi lub Ca-fosforanowymi strukturami
szkieletowymi.
CaH w dzisie js zych morzach pochodzi z dw6ch gl6wnych zrodel : (1) produkt6w
wietrzenia ladowego, znoszonych wodami r zecznymi i (2) roztwor6w hydrotermal
nych , wydobywajacych sie ze skor upy oceanicznej w strefach ryftowych. Przez
wiekszosc prekambru usuwnnie Ca 2+ ze sr odow iska morskiego nastepowalo przede
wszystkim przez chemiczne wytracanie weglanow w strefach plywowych i nad
plywowych (np . w formie st romatoli t6w), lub w poblizu ryft6w. Sklad chemiczny
w ody morskiej w tym czasie zbl izony byl do skladu dzisiejszych przyryftowych
i wulkanicznych jezior sodowych, kt6rych wysokie pH (>10) pozwala utrzymac
w roztworze tylko znikome ilosci Ca2+. Z prze isciem od oceanu sodowego do
chlorkowego (halitowego), co mi alo miej sce w ciagu proterozoiku, poziom Ca2+w wodzie morskiej st opn iowo wzrastal, szczeg6lnie w zbiornikach epikontynen
talnych, zas ilanych wraz z postepuiaca kratonizacjq litosfery coraz wiekszymi
ilosciami CaH pochodzacego z wietrzenia kontynent6w. Reakcjq organizrnow na
wzrastajqcy stres wapniowy byla prawie jednoczesna biokalcyfikacja w ielu grup
bezkregowcow w wendzie i wczesnym kambrze. "Wymuszenie" przez srodowiskc
wapiennych st r uktur szk ieletowych na organi zrnach nie bylo procesem ograni
czonym do pogranicza prekambru i kambru. W ciagu fanerozoiku szereg grup orga
nizm6w bezszk ieletowych odpowiedzialo na stres wapniowy swoich srodow isk zycia
wyksztalceniem wapiennych szk ielet6w (np. w dewonie otwornice bentosowe, zas
na przelomie triasu i jury planktonowe otwornice i CZE:SC nannofitoplankter6w).
Biokalcyfikacja zmienila generalnie charakter wytracania weglanow w morzu

Ca+' STRESS AND ITS GEOLOGICAL IMPLICATIONS 135
z chemicznego na enzymatyczny, a ekstrakcja Ca 2+ w morzach fanerozoic znych
stala sie prawie wylacznie domena organizrnow.
Wiele pierwotniak6w i glon6w planktonowych reaguje na stres wapniowy sro
dowiska obfit ym wydzielaniem wielocukrow czy glikoprotein kompleksujqcych
Ca z+. Obecnosc tych substancji w wodzie prowadzi z kolei do makroflokulacji
drobnych cza stek osadu zawieszonych w wodzie, kt6re bez Iaczenia sie w wieksze
agregaty nie moglyby opadac na dna. Sedymentacja zawiesiny ilastej jest wiec
zwiazana z intensywnoscia wydalnicza planktonu, zalezna bezposrednio ad ste
zenia jon6w wapnia (a takze innych metali) w srodowisku,
Praca byla czesciowo finansowana przez Polska Akadernie Nauk w ramach
problemu MR II 6.