from molecules to bird communities: a mistletoe story
TRANSCRIPT

i
FROM MOLECULES TO BIRD COMMUNITIES:
A MISTLETOE STORY
This thesis was submitted in fulfilment of the requirements for the degree of Doctor of
Philosophy (Veterinary Studies), Murdoch University, by:
Kathryn Renee Napier
BSc., BSc. (Hons)
School of Veterinary and Life Sciences
Murdoch University
Murdoch, Western Australia
June 2013

ii
DECLARATION
I declare that this thesis is my own account of my research and contains as its main
content of work, which has not previously been submitted for a degree at any tertiary
education institution.
Kathryn Renee Napier

iii
ABSTRACT
Worldwide, mistletoes act as a keystone resource, providing food (nectar, fruit and
foliage) and structural (nesting sites) resources to hundreds of fauna species. 75 species
of ‘showy’ Loranthaceae mistletoes are native to Australia, and are found in wooded
habitats throughout the mainland. The mistletoebird (Dicaeum hirundinaceum) is
considered the primary disperser of mistletoe fruit in south-west Western Australia
(WA). Other ‘generalist’ species, including several honeyeater species and the
silvereye (Zosterops lateralis), also regularly consume and disperse mistletoe fruits.
This thesis takes a broad, eco-physiological approach to investigate the
interactions between two Australian loranthaceous mistletoes species (Amyema miquelii
and A. preissii), their host plants and their avian consumers. This was achieved through
a combination of intensive field surveys and sampling at five sites where mistletoe was
extremely abundant, and laboratory experiments assessing various aspects of avian
digestive physiology of three frugivorous bird species; the mistletoebird (specialised
frugivore), silvereye (generalist frugivore) and singing honeyeater (Lichenostomus
virescens; generalist nectarivore).
A stable isotope approach was used to investigate the parasitic relationship
between A. miquelii and A. preissii and their eucalypt or acacia hosts (respectively).
Results demonstrate that these mistletoes regulate their water use in relation to the
supply of nitrogen available from the host.
Next, the importance of mistletoe to bird communities in south-west WA was
investigated through surveys and the use of stable isotopes. The presence of fruiting

iv
(but not flowering) mistletoe was associated with significant changes in bird community
structure. Mistletoebirds were more likely to be recorded during months when ripe
mistletoe fruit was present, and overall bird species richness was higher for these survey
months. The contribution of mistletoe fruit to the diet of mistletoebirds ranged from
33% to 55%, demonstrating that despite mistletoe fruit being low in nitrogen, it is an
important source of nutrients. Fruiting mistletoes therefore provide important food
resources to bird communities in south-west WA.
Various aspects of avian digestive physiology were compared for three species
that include mistletoe fruit in varying degrees to their diet. Mistletoebirds, silvereyes
and singing honeyeaters demonstrated similar patterns of sugar preferences with
similarly high (>97.5%) apparent assimilation efficiencies (AE*) for sucrose, glucose
and fructose and lower AE* for the pentose monosaccharide xylose (56-78%), yet
demonstrated differences in their absorption of dietary sugars. Mistletoebirds, in
contrast to the other two species, did not vary bioavailability (f) with diet concentration,
and appear to absorb xylose through both mediated and paracellular mechanisms. This
may be a result of the short, specialised intestinal tract of mistletoebirds, which
facilitates faster transit rates of mistletoe fruit compared to silvereyes and honeyeaters.
This thesis presents new insight into the parasitic relationship between
mistletoes and hosts, and the relationship between mistletoes and its avian consumers.
Mistletoebirds differ from other opportunistic mistletoe feeders in their ability to
process large numbers of mistletoe fruit quickly, while obtaining sufficient nutrients
such as nitrogen and carbohydrates from these fruits.

v
ACKNOWLEDGMENTS
Firstly, I’d like to thank my supervisors – Patricia Fleming, Todd McWhorter and
Carlos Martínez del Rio. I don’t think I could express just how much I have enjoyed
working with you over the last 6+ years. Trish, you have taught me more than I would
have ever thought possible – your brilliant advice, encouragement, and unbelievable
dedication to your students are such an inspiration. I would not be at this stage in my
research career nor have experienced such fantastic opportunities without your constant
encouragement, support, patience and friendship, and I will always be grateful.
Todd, I really appreciate the encouragement, support and advice given over the
long distances between Perth, Laramie and Adelaide. Your patience, kind words and
knowledge freely given have helped me immensely over the years. I have dearly
missed working with you in person, and hope that I may rejoin you someday in the lab.
Carlos, you gave me the opportunity of a lifetime by accepting me into your lab
group at the University of Wyoming. I thank you and Martha for the support and help
you gave in those first few weeks at 2184 m! Laramie and Wyoming now have a
special place in my heart, and the Medicine Bow Mountains are one of my favourite
places on this earth.
My deepest gratitude is also given to my sources of funding and awards, without
which this research would not have been possible: the 2010 Western Australian
Fulbright Scholarship and Gregory Schwartz Enrichment Grant (Australian-American
Fulbright Association), the Holsworth Wildlife Research Endowment, the Stuart Leslie
Bird Research Award and travel grant (Birdlife Australia and Birdlife WA), and the

vi
Jean Gilmore Postgraduate research bursary (Federation of University Women, SA).
The financial support given by Murdoch University is also very much appreciated.
Sincere thanks are also given to the staff at the Araluen Country Club and
Resort, Dr Manda Page and Jo Kuiper from the Australian Wildlife Conservancy, and
Peter Monger and family in York for allowing access to their field sites. I would also
like to thank reviewers whose comments greatly improved previous versions of the
manuscripts in Chapters 3, 5 and 6.
I would also like to thank Suzanne Mather for her assistance with bird surveys
(and your willingness to get up at 4 am!), the use of your ladder, and for worrying if I
would fall off the ladder. You’re an inspiration.
I would also like to thank the following people for their assistance with various
aspects of my research: Tony Start (Western Australian Herbarium, Department of
Environment and Conservation) for his mistletoe knowledge and comments on a draft
version of Chapter 3; Simon Cherriman, for climbing trees and tagging them for me at
Paruna; Joao Paulo Coimbra (University of Western Australia) for the provision of
avian intestinal tissues; Clare Auckland and John McCooke (Murdoch University) for
their assistance with intestinal enzyme assays; Bill Bateman for demonstrating his
sensational grasshopper catching skills and assisting with mist-netting; Shannon
Dundas, Tracey Moore and Penny Nice (Murdoch University) for their assistance with
field work; the staff at the Murdoch University Animal House, particularly Derek
Mead-Hunter, for their assistance in keeping birds in captivity; David Perry (University
of Wyoming Macromolecular Analysis Core) for HPLC analysis; and the staff at the
University of Wyoming Stable Isotopes Facility for being so very patient with me while

vii
we processed hundreds of samples. Special thanks must also go to Susan Nicolson
(University of Pretoria) for her advice and collaboration.
To my fellow research students in the dungeon and bat cave offices, past and
present, I thank you for your friendship and support. To Penny, Tracey, Narelle, Wil,
Heather, Gill, Shannon, Ivan, Chelsea, Bryony, Renata, John, Kelly, Tegan (honorary
dungeon member), Buddy and Rocket (dungeon mascots) – my time at Murdoch was a
truly enjoyable experience thanks to you wonderful, fantastic people (and animals). To
Jon and Brenna in Laramie, I thank you for your friendship and advice, and for not
laughing at me (too much) during my first bear encounter. I look forward to future
collaborations!
Last, but not least, I would like to thank my family and friends for their constant
support and love: to my parents, Jeff and Clarice Napier, Sandy Chaney, siblings David
and Alison Napier and future brother in-law Eddie Terry, friends Brett and Lois
Andrijich and Liz Snyder-Campion, and my husband Simon Aplin. Simon, your love,
our adventures, and your constant support of my academic endeavours enrich my life in
countless ways. I thank you from the bottom of my heart.

viii
DISCLAIMER
This PhD thesis consists of chapters that have been prepared as stand-alone
manuscripts. These manuscripts have either been accepted for publication (Chapters 3
and 5), submitted for consideration of publication (Chapter 6), or are being prepared for
future submission (Chapters 2 and 4). As a consequence, there may be some repetition
between chapters. To reduce unnecessary replication of references between chapters,
the references are compiled together in Chapter 8.
To maintain consistency in formatting throughout the thesis, the chapters may
differ slightly from the future published manuscripts.

ix
PUBLICATIONS ARISING FROM THIS THESIS
Napier, K. R., McWhorter, T. J., Nicolson, S. W., and Fleming, P. A. (2013). Sugar
preferences of avian nectarivores are correlated with intestinal sucrase activity.
Physiological and Biochemical Zoology 86, 499–514.
Napier, K. R., Mather, S. H., McWhorter, T. J. and Fleming, P. A. (2013). Do bird
species richness and community structure vary with mistletoe flowering and
fruiting in Western Australia? Emu In press.
OTHER PUBLICATIONS ARISING FROM THE PERIOD OF CANDIDATURE
Purchase, C., Napier, K. R., Nicolson, S. W., McWhorter, T. J. and Fleming, P. A.
(2013). Gastrointestinal and renal responses to variable water intake in
whitebellied sunbirds and New Holland honeyeaters. Journal of Experimental
Biology 216, 1537–1545.
Napier, K. R., McWhorter, T. J. and Fleming, P. A. (2012). A comparison of
pharmacokinetic methods for in vivo studies of nonmediated glucose absorption.
Physiological and Biochemical Zoology 85, 200–208.
Napier, K. R., McWhorter, T. J. and Fleming, P. A. (2008). Mechanism and rate of
glucose absorption differ between an Australian honeyeater (Meliphagidae) and
a lorikeet (Loriidae). Journal of Experimental Biology 211, 3544–3553.

x
Napier, K. R., Purchase, C., McWhorter, T. J., Nicolson, S. W. and Fleming, P. A.
(2008). The sweet life: diet sugar concentration influences paracellular glucose
absorption. Biology Letters 4, 530–533.
Fleming, P. A., Xie, S., Napier, K., McWhorter, T. J. and Nicolson, S. W. (2008).
Nectar concentration affects sugar preferences in two Australian honeyeaters and
a lorikeet. Functional Ecology 18, 223–232.

xi
TABLE OF CONTENTS
DECLARATION ....................................................................................................................................... II
ABSTRACT ............................................................................................................................................. III
ACKNOWLEDGMENTS ........................................................................................................................ V
DISCLAIMER ....................................................................................................................................... VIII
PUBLICATIONS ARISING FROM THIS THESIS ............................................................................ IX
OTHER PUBLICATIONS ARISING FROM THE PERIOD OF CANDIDATURE ........................ IX
TABLE OF CONTENTS ......................................................................................................................... XI
LIST OF FIGURES .............................................................................................................................. XVI
LIST OF TABLES ............................................................................................................................. XVIII
1 GENERAL INTRODUCTION ........................................................................................................ 1
1.1 MISTLETOE-HOST INTERACTIONS ................................................................................................... 2
1.2 MISTLETOE-ANIMAL INTERACTIONS ............................................................................................... 5
1.2.1 Provision of resources .......................................................................................................... 5
1.2.2 Pollination and dispersal ..................................................................................................... 7
1.3 PHYSIOLOGICAL ADAPTATIONS TO FRUGIVORY ............................................................................. 9
1.4 STUDY AIM ................................................................................................................................... 13
1.5 MISTLETOE STUDY SPECIES .......................................................................................................... 13
1.6 AVIAN STUDY SPECIES .................................................................................................................. 18
1.6.1 Mistletoebird (Dicaeum hirundinaceum) ........................................................................... 18
1.6.2 Silvereye (Zosterops lateralis) ............................................................................................ 19
1.6.3 Singing honeyeater (Lichenostomus virescens) .................................................................. 19
2 CARBON AND NITROGEN STABLE ISOTOPES OF AUSTRALIAN MISTLETOES AND
THEIR HOSTS......................................................................................................................................... 22
STATEMENT OF AUTHOR CONTRIBUTION ................................................................................................ 23
2.1 ABSTRACT .................................................................................................................................... 24
2.2 INTRODUCTION ............................................................................................................................. 25

xii
2.3 METHODS ..................................................................................................................................... 27
2.3.1 Field sites and sampling ..................................................................................................... 27
2.3.2 General statistics ................................................................................................................ 30
2.4 RESULTS ....................................................................................................................................... 32
2.4.1 Carbon ................................................................................................................................ 32
2.4.2 Nitrogen .............................................................................................................................. 32
2.4.3 Relationships between mistletoes and hosts ....................................................................... 36
2.5 DISCUSSION .................................................................................................................................. 39
2.5.1 δ13
C of mistletoes and hosts: water relations and effects of aridity ................................... 39
2.5.2 Relationships with nitrogen ................................................................................................ 40
2.5.3 Conclusions ........................................................................................................................ 41
3 DO BIRD SPECIES RICHNESS AND COMMUNITY STRUCTURE VARY WITH
MISTLETOE FLOWERING AND FRUITING IN WESTERN AUSTRALIA? ............................... 42
STATEMENT OF AUTHOR CONTRIBUTION ................................................................................................ 43
3.1 ABSTRACT .................................................................................................................................... 44
3.2 INTRODUCTION ............................................................................................................................. 45
3.3 METHODS ..................................................................................................................................... 49
3.3.1 Study sites ........................................................................................................................... 49
3.3.2 Fruit and flower phenology ................................................................................................ 50
3.3.3 Bird species ........................................................................................................................ 52
3.3.4 Statistical analysis .............................................................................................................. 52
3.4 RESULTS ....................................................................................................................................... 53
3.4.1 Presence of mistletoe flowers ............................................................................................. 54
3.4.2 Presence of ripe mistletoe fruit ........................................................................................... 55
3.5 DISCUSSION .................................................................................................................................. 60
3.5.1 Reliability of mistletoe resources ....................................................................................... 60
3.5.2 Abundance of mistletoe ...................................................................................................... 61
3.5.3 Degree of consumer specificity .......................................................................................... 62
3.5.4 Temporal redundancy of mistletoe ..................................................................................... 64

xiii
3.5.5 Resources (functions) not otherwise present ...................................................................... 65
3.5.6 Conclusions ........................................................................................................................ 66
4 MISTLETOEBIRDS VARY THEIR DIETARY INTAKE OF ARTHROPODS DEPENDING
ON TIME OF YEAR ............................................................................................................................... 69
STATEMENT OF AUTHOR CONTRIBUTION ................................................................................................ 70
4.1 ABSTRACT .................................................................................................................................... 71
4.2 INTRODUCTION ............................................................................................................................. 72
4.3 METHODS ..................................................................................................................................... 74
4.3.1 Study sites ........................................................................................................................... 74
4.3.2 Breath, blood and feather samples ..................................................................................... 76
4.3.3 Diet sources ........................................................................................................................ 78
4.3.4 Stable isotopes analysis ...................................................................................................... 78
4.3.5 Isotope mixing model.......................................................................................................... 79
4.3.6 General statistics:............................................................................................................... 80
4.4 RESULTS ....................................................................................................................................... 83
4.4.1 Diet sources ........................................................................................................................ 83
4.4.2 Mistletoebird tissues ........................................................................................................... 84
4.4.3 Isotope mixing model.......................................................................................................... 87
4.4.4 Comparison with other bird species ................................................................................... 87
4.5 DISCUSSION .................................................................................................................................. 92
4.5.1 Detecting diet switching of mistletoebirds .......................................................................... 92
4.5.2 Contribution of fruit vs arthropods .................................................................................... 94
4.5.3 Comparison with other bird species ................................................................................... 95
4.5.4 Arthropods and mistletoe fruit are both important sources of dietary nitrogen ................. 96
5 SUGAR PREFERENCES OF AVIAN NECTARIVORES ARE CORRELATED WITH
INTESTINAL SUCRASE ACTIVITY ................................................................................................... 98
STATEMENT OF AUTHOR CONTRIBUTION ................................................................................................ 99
5.1 ABSTRACT .................................................................................................................................. 100
5.2 INTRODUCTION ........................................................................................................................... 102

xiv
5.3 MATERIALS AND METHODS ......................................................................................................... 108
5.3.1 Birds and their maintenance ............................................................................................ 108
5.3.2 Apparent assimilation efficiency (AE*) ............................................................................ 109
5.3.3 Sugar preference trials ..................................................................................................... 111
5.3.4 Intestinal enzymes............................................................................................................. 113
5.3.5 Foraging data and Australian nectar composition .......................................................... 117
5.3.6 General statistical analysis .............................................................................................. 118
5.4 RESULTS ..................................................................................................................................... 120
5.4.1 Apparent assimilation efficiency (AE*) ............................................................................ 120
5.4.2 Sugar preferences ............................................................................................................. 120
5.4.3 Intestinal enzymes............................................................................................................. 123
5.4.4 Foraging data and Australian nectar composition .......................................................... 126
5.5 DISCUSSION ................................................................................................................................ 132
5.5.1 Are there differences in apparent assimilation efficiency between sugar types? ............. 133
5.5.2 Can we explain hexose preferences on dilute diets? ........................................................ 134
5.5.3 Can we explain sucrose preference on concentrated diets? ............................................. 135
5.5.4 Do laboratory results reflect foraging preferences in the wild? ...................................... 136
5.5.5 Conclusions ...................................................................................................................... 137
6 MISTLETOEBIRDS AND XYLOSE: AUSTRALIAN FRUGIVORES DIFFER IN THEIR
HANDLING OF DIETARY SUGARS ................................................................................................. 139
STATEMENT OF AUTHOR CONTRIBUTION .............................................................................................. 140
6.1 ABSTRACT .................................................................................................................................. 141
6.2 INTRODUCTION ........................................................................................................................... 142
6.3 MATERIALS AND METHODS ......................................................................................................... 147
6.3.1 Study species .................................................................................................................... 147
6.3.2 Apparent assimilation efficiency (AE*):........................................................................... 149
6.3.3 Pharmacokinetic experiments .......................................................................................... 151
6.3.4 Gut passage times (GPT) ................................................................................................. 152
6.3.5 Statistical analysis ............................................................................................................ 153

xv
6.4 RESULTS ..................................................................................................................................... 153
6.4.1 Apparent assimilation efficiency (AE*) ............................................................................ 153
6.4.2 Pharmacokinetic experiments .......................................................................................... 156
6.4.3 Gut passage times (GPT) ................................................................................................. 160
6.5 DISCUSSION ................................................................................................................................ 160
6.5.1 Apparent assimilation efficiency (AE*) ............................................................................ 161
6.5.2 Mechanism of sugar absorption ....................................................................................... 162
6.5.3 Metabolism of xylose ........................................................................................................ 165
6.5.4 Conclusions ...................................................................................................................... 166
7 GENERAL CONCLUSIONS ...................................................................................................... 167
7.1 MISTLETOE-HOST INTERACTIONS ............................................................................................... 167
7.2 MISTLETOE-ANIMAL INTERACTIONS ........................................................................................... 168
7.3 PHYSIOLOGICAL ADAPTATIONS TO FRUGIVORY .......................................................................... 170
7.4 FUTURE DIRECTIONS ................................................................................................................... 172
7.5 CONCLUSIONS ............................................................................................................................ 174
8 REFERENCES ............................................................................................................................. 176

xvi
LIST OF FIGURES
Figure 1.1 The location of five study sites in south-west Western Australia. ........................................... 16
Figure 1.2 Two Loranthaceae mistletoe species common in south-west Western Australia: the box (or
stalked) mistletoe Amyema miquelii and wireleaf mistletoe Amyema preissii. ............................ 17
Figure 1.3 Digestive adaptations associated with consuming a mistletoe fruit diet were investigated for
three focal frugivorous bird species. ............................................................................................ 21
Figure 2.1 Stable isotope values and percentages of carbon and nitrogen for mistletoe, infected and
uninfected host samples on acacia (black symbols) or eucalyptus (white symbols) hosts. .......... 35
Figure 2.2 Correlations between mistletoe and host a) δ13
C (‰), b) carbon content (%), c) δ15
N (‰) and
d) nitrogen content (%) ................................................................................................................ 38
Figure 3.1 Flowering and fruiting phenology of the mistletoes Amyema preissii (left hand panel) and
Amyema miquelii (right hand panel) from February 2010 to January 2011. ................................ 58
Figure 3.2 The number of bird species recorded at each site in the presence (1) or absence (0) of open
flowers (a) and ripe fruit (c), and the number of nectarivorous bird species recorded at each site
in the presence (1) and absence (0) of open flowers (b). ............................................................. 59
Figure 4.1 Median, 25th
and 75th
quartile values of δ15
N (‰) signatures of arthropods sampled at three
sites in south-west Western Australia. ......................................................................................... 82
Figure 4.2 δ13
C (a) and δ15
N (b) values (‰) of breath, whole blood and feathers for mistletoebirds at
three sites in south-west Western Australia. ................................................................................ 86
Figure 4.3 δ13
C and δ15
N values (‰) of a) whole blood and feathers from individual mistletoebirds and
b) average whole blood values for mistletoebirds, silvereyes, brown honeyeaters and yellow-
rumped thornbills at three sites in south-west Western Australia. ............................................... 89
Figure 4.4 Proportional contribution of fruit and insects to the whole blood of mistletoebirds a) and
proportional contribution of mistletoe fruit to individual mistletoe birds b) obtained from a dual-
isotope (δ13
C, δ15
N) Bayesian isotope mixing model (SIAR). ..................................................... 90
Figure 5.1 Evolutionary relationships. .................................................................................................... 116
Figure 5.2 Concentration-dependent total sugar intake. .......................................................................... 121
Figure 5.3 Concentration-dependent sugar preferences. ......................................................................... 122

xvii
Figure 5.4 Gut nominal surface area and intestinal enzymes. ................................................................. 125
Figure 5.5 Feeding observations for New Holland honeyeaters, red wattlebirds, singing honeyeaters and
silvereyes in Western Australia (Brown et al. 1997) and rainbow lorikeets in Western Australia
and the Queensland-New South Wales border region (Cannon 1984; Brown et al. 1997). ....... 127
Figure 5.6 Average nectar composition from 16 Australian plant genera (mean fructose, glucose,
sucrose). ..................................................................................................................................... 129
Figure 6.1 Apparent assimilation efficiency (AE*) of D-xylose (solid) and D-glucose (lines). ............. 155
Figure 6.2 Bioavailability (f, %) of radiolabelled L-glucose (white) and D-xylose (grey) at two diet
concentrations (0.25 and 1 mol·L-1
, solid and lines respectively). ............................................. 159

xviii
LIST OF TABLES
Table 2.1 The characteristics of the five field sites in south-west Western Australia. .............................. 29
Table 2.2 Leaf carbon and nitrogen content (%), δ13
C and δ15
N values (‰) of mistletoe, host and
uninfected host plants. ................................................................................................................. 31
Table 2.3 δ13
C and δ15
N values (‰) and carbon and nitrogen content (%) for all samples (mistletoe,
infected and uninfected hosts) were analysed as dependent variables in separate mixed-model
ANOVAs. .................................................................................................................................... 34
Table 3.1 The characteristics of the five sites surveyed in south-west Western Australia. ....................... 51
Table 3.2 Bird species recorded in 53 surveys over 5 sites in south-west Western Australia from February
2010 to January 2011. .................................................................................................................. 68
Table 4.1 The characteristics of the three field sites in south-west Western Australia. ............................ 75
Table 4.2 Stable isotope values δ13
C and δ15
N (‰) and carbon and nitrogen concentrations (%) for
mistletoebird tissues (breath, blood and feathers) and diet sources (arthropods and mistletoe
fruit). ............................................................................................................................................ 85
Table 5.1 Summary of sugar type preferences, apparent assimilation efficiency (AE*) and digestive
capacity. ..................................................................................................................................... 107
Table 5.2 Details of birds euthanased for digestive enzymes. ................................................................. 119
Table 5.3 Summary of maltase activity. .................................................................................................. 130
Table 5.4 Nectar composition of 16 Australian plant genera. ................................................................. 131
Table 6.1 Relative sugar composition (% dry mass±SEM) of mistletoe fruit viscin............................... 146
Table 6.2 Parameters used to determine bioavailability (f) of radiolabelled L-glucose and D-xylose in
mistletoebirds, silvereyes and singing honeyeaters at two diet concentrations (0.25 and 1 mol·L-1
hexose solutions). ....................................................................................................................... 158

1
1 GENERAL INTRODUCTION
Examining the interactions of plants and animals addresses some of the most
fundamental and challenging questions in ecology (Sutherland et al. 2013). The
interactions between mistletoes, their hosts, and animals are particularly interesting due
to the significant influence of mistletoes on the ecological communities they inhabit.
Mistletoes are a diverse group of shrubby, aerial, flowering plants with over
1,500 species found over a wide range of habitats and across all continents with the
exception of Antarctica (Kuijt 1969; Calder 1983; Watson 2001). Their growth form of
obligate hemiparasitism exhibits a unique strategy by combining the parasitism of a host
species with their own ability to photosynthesise (Těšitel et al. 2010). There are five
families within the order Santalales, with Loranthaceae and Viscaceae comprising the
majority (> 98%) of mistletoe species (Nickrent 2002; Mathiasen et al. 2008). Most
mistletoes rely on birds or insects for pollination and fruit dispersal, which leads to a
broad range of mistletoe-animal interactions, yet the closest interactions mistletoes form
would be their hemiparasitic relationships with their hosts, in order to obtain water and
nutrients (Miller et al. 2003).
This thesis includes five experimental chapters, investigating the relationships
between mistletoes, their hosts, and the avian community. These studies use a variety
of experimental methods to examine these relationships from a molecular to community
level. Firstly, I investigated the parasitic relationships between mistletoes and their
hosts (Chapter 2). Secondly, I investigated the importance of mistletoe as a food
resource to bird communities, and the contribution of mistletoe fruit to the diet of

2
mistletoebirds (Chapters 3 and 4). Thirdly, I examined the digestive adaptations
associated with consuming a mistletoe fruit diet for three focal avian species (Chapters
5 and 6). This introductory chapter therefore focuses on three aspects; the interactions
between mistletoes and their hosts, the interactions between mistletoes and animals, and
the physiological adaptations associated with frugivory.
1.1 MISTLETOE-HOST INTERACTIONS
One general characteristic of parasites is the deleterious relationship they have with
their hosts. The infection of commercially-valuable conifers with the dwarf mistletoe
(Arceuthobium), and subsequent reductions in growth, fruiting and seed production
(Mathiasen et al. 2008), is an example of a classic parasite-host interaction. However,
the relationship between mistletoe and host may switch from parasitism to mutualism
by the introduction of a third species, such as an avian disperser. For example, juniper
hosts infected with mistletoe attracted significantly more avian seed dispersers
compared to uninfected host trees, which resulted in higher recruitment of juniper
seedlings in stands where mistletoe was present (van Ommeren and Whitham 2002).
While some mistletoe species, like the dwarf mistletoe, can have deleterious
effects on their hosts, most mistletoes actually play a key role in ecosystems. As
mistletoes depend solely on their hosts for their water and the majority of their
nutritional requirements, it is not in the best interest of the mistletoe to impact on the
mortality of its host. While infection by mistletoes can negatively impact their hosts in
some cases, mature, healthy trees in general do not suffer major ill effects, aside from
the loss of the branch distal to a mistletoe infestation (Tennakoon and Pate 1996). The
host safeguards the mistletoe by providing a buffer against large-scale fluctuations in

3
the availability of nutrients and resources that may limit the growth and distribution of
other plants (Ehleringer and Marshall 1995). Mistletoe plants have a low annual
survival rate for a variety of reasons. Susceptibility to fire (Rowe 1983) and frost
(Smith and Wass 1979), and a high light requirement for successful germination,
establishment, and maturation, has lead to a narrow microsite tolerance in many
mistletoe species (Lamont 1982; Knutson 1983; Lamont 1983a; Sargent 1995; Yan and
Reid 1995; Hawksworth and Wiens 1996; Polhill and Wiens 1998). Mistletoes are also
generally more sensitive to water shortages in comparison to their hosts, leading to
increased mistletoe mortality during periods of drought (Watson 2011).
Loranthaceous mistletoes are dependent on their hosts for water and mineral
nutrients (Ehleringer et al. 1985; Marshall and Ehleringer 1990), and obtain these by
parasitising the xylem of their hosts (Calder 1983; Marshall et al. 1994). Despite the
lack of a connection to the phloem, xylem-tapping mistletoes can also obtain varying
amounts of carbon from the host photosynthate (Hull and Leonard 1964; Marshall and
Ehleringer 1990; Dörr 1997; Govier et al. 1967). The haustorium (a vascular
connection which attaches the mistletoe to the stem of a host plant) facilitates the one-
way flow of water and dissolved nutrients from the host xylem stream (Lamont 1983b;
Ehleringer and Marshall 1995), although the mechanisms of this movement are still not
completely understood. Transpiration rates of mistletoes are generally higher then those
of their hosts, and this is likely achieved by mistletoes keeping their stomata open, even
under dry conditions or at night, when other plants typically close their stomata to
minimise water loss (Schulze et al. 1984; Ehleringer and Marshall 1995). These high
transpiration rates also cause reduced xylem water potentials in the host branches,
which acts to reduce the net photosynthetic rate of the host (Knutson 1983; Stewart and

4
Press 1990; Mathiasen et al. 2008). Mistletoes accumulate high concentrations of
xylem-transport cations in their tissues, particularly phosphorous and potassium
(Lamont 1983b), which further facilitates the absorption of water and solutes by
promoting lower xylem water potential in mistletoe tissue compared with their hosts
(Knutson 1983; Mathiasen et al. 2008). The xylem hydraulic conductivity of host
branches distal to a mistletoe infection is likely to be decreased due to transpiration by
the mistletoe, causing the death of the end of the host branch while the mistletoe
remains living and attached to the host (Tennakoon and Pate 1996).
Parasitism of a host by mistletoes therefore inevitably effects water, nitrogen
and carbon assimilation of host plants (Wang et al. 2007). Studies of stable isotopes of
carbon and nitrogen have been instrumental in the development of the knowledge of
mistletoe biology. Conventionally expressed as a ratio of the heavy to the light isotope
(δ) in units per mil (‰), the studies of carbon (13
C to 12
C) and nitrogen (15
N to 14
N)
stable isotope ratios have facilitated our understanding of how mistletoes acquire carbon
and nitrogen from their hosts. The δ13
C of plant material also gives an indication of the
long term water use efficiency of the plant, due to the strong correlation between carbon
isotope discrimination and the ratio between carbon assimilation and transpiration (i.e.
water use efficiency) (Farquhar et al. 1982). This relationship results from the
discrimination against 13
C in favour of 12
C during the diffusion of carbon dioxide into
plant leaves and during the fixation of carbon by ribulose-1,5-biphosphate carboxylase
in the mesophyll (Toft et al. 1989).
More positive values of δ13
C (i.e. 13
C-enriched) are therefore found in more
water-efficient plants. In arid areas, the δ13
C values for mistletoes are generally more

5
negative than those of their hosts, and the greatest differences in δ13
C between
mistletoes and hosts were on hosts with low nitrogen contents (Ehleringer et al. 1985).
This suggests that a greater differential in water use efficiency is required to extract
nitrogen from nitrogen-poor hosts. I therefore used δ13
C and δ15
N to examine the
parasitic relationships of five unique mistletoe-host pairs from five sites in south-west
Western Australia (WA) in terms of the water use efficiency of mistletoes, their hosts
and uninfected host plants.
1.2 MISTLETOE-ANIMAL INTERACTIONS
1.2.1 Provision of resources
While some mistletoe species can have deleterious effects on their hosts, most
mistletoes actually play a key role in ecosystems (Watson 2001; Mathiasen et al. 2008).
Mistletoes offer reliable food and shelter resources for hundreds of fauna species,
providing resources and ecosystem services out of proportion to their abundance and
contribution to biomass (Davidson et al. 1989b; Watson 2001; Watson 2002; Mathiasen
et al. 2008). Mistletoes may therefore be considered a ‘keystone species’ (discussed in
further detail in Chapter 3). For example, mistletoe plants provide a broad range of
nutritional resources to both obligate and partially-dependent animal species, and create
a habitat that is used for nesting and roosting by a wide variety of species (Watson
2001; Mathiasen et al. 2008).
Mistletoes are used as a food source by a wide range of animals, with species
from 66 families of birds and 30 families of mammals recorded feeding on mistletoe
nectar, fruit and leaves worldwide (reviewed by Watson 2001). A wide range of bird

6
and insect species are known to pollinate mistletoes; many of these birds and insects
tend not to be dependent on mistletoe nectar as a primary food source, while there are
few specialised mistletoe fruit dispersers (Davidar 1983; Davidar 1985; Reid 1986;
Watson 2001). Bats are also known to visit mistletoe flowers, but their role as a
potential pollinator has not yet been confirmed (Mathiasen et al. 2007). In addition to
flowers and fruit, mistletoe leaves also provide an extremely nutritious food source to
folivorous animals such as possums, deer and gorillas, and mistletoe clumps are often
used as a foraging substrate by insectivorous species (Watson 2001).
A wide range of species have been recorded nesting in mistletoes, including
mammals, marsupials, and birds (see extensive reviews by Watson 2001; Cooney et al.
2006). In Australia, 66% of all arboreal-nesting bird species (217 species from 29 bird
families) have been reported nesting in mistletoe; the dense clumps provide structural
support for nests, a more attractive microclimate, and also a ‘hiding’ spot for the nests
of smaller passerines, with lower predation rates demonstrated for nests in mistletoe
compared to eucalypts (Cooney et al. 2006; Cooney and Watson 2008).
Mistletoes have recently been described as the Robin Hood of woodlands, by
taking nutrients from their hosts to redistribute to those in need, a more favourable
association then being compared to Dracula, taking nutrients from an unfortunate host
(Press 1998). While mistletoes provide abundant food and nesting resources, mistletoe
leaf litter (which is shed year-round) also returns valuable nutrients such as potassium,
phosphorous, and nitrogen back to the host and surrounding plant community (March
and Watson 2010). The high concentration of minerals and water present in mistletoe
leaves, coupled with the high rates of mistletoe leaf turn over, act to increase leaf litter

7
mass and the availability of mineral nutrients, and improve soil fertility by slowing the
overall rate of decomposition and providing a steady supply of nutrients (March and
Watson 2007; March and Watson 2010; Watson 2011).
1.2.2 Pollination and dispersal
With the exception of the insect pollinated root-parasitic Western Australian Christmas
tree, Nuytsia floribunda, and Atkinsonia, Atkinsonia ligustrina, the majority of
Australian loranthaceous mistletoes are pollinated by birds (Hawkeswood 1981; Watson
2011). These ornithophilous mistletoes tend to display brightly-coloured, odourless
flowers, with abundant, sugar-rich nectar (up to 60% total sugar content, primarily
glucose and fructose sugars, Reid 1986; Stiles and Freeman 1993; Baker et al. 1998).
Although they are not entirely dependent on mistletoe nectar as a primary food source, a
wide range of bird species pollinate these ornithophilous mistletoes, including
spinebills, wattlebirds, and singing, spiny-cheeked and New Holland honeyeaters
(Meliphagidae; Reid 1986; Watson 2001; Watson 2011).
Although the seeds of several lineages of mistletoe (e.g. Viscaceae) are rather
spectacularly self-dispersed by hydrostatic explosion (Hawksworth and Geils 1996),
mistletoes in general are bird dispersed. Mistletoe fruits contain chlorophyllus seeds
which are enclosed in a thick pericarp. This pericarp is made up of epicarp, mesocarp
and endocarp layers (Salle 1983). The thick mesocarp is composed of hairy cells,
spherical gel, and a mucilaginous substance known as viscin (Salle 1983; Azuma et al.
2000). Viscin tissue occupies a significant proportion of the mature fruit, and is
composed of cellulose in a mixture of both acidic and neutral polysaccharides (Salle

8
1983; Gedalovich et al. 1988; Azuma et al. 2000). The seeds voided by the bird are still
surrounded by a layer of viscin, which is unique to parasitic plants (Richardson and
Wooller 1988; Azuma et al. 2000), and functions in the dissemination of mistletoe by
birds. As the mistletoe seeds can adhere to the cloaca of the bird, wiping by the bird
may be required to remove the seeds and thus adhere them to host branch, where they
remain ‘glued’ prior to germination (Azuma and Sakamoto 2003).
The nutritional composition of the fleshy fruit pulp varies among species, but
most mistletoe fruits contain a high proportion of carbohydrates, lipids and protein
(Walsberg 1975; Godschalk 1983a; Herrera 1987; McPherson 1987; Witmer 1996;
López de Buen and Ornelas 2001; Barea 2008). While a small number of highly
specialised avian species tend to be associated with the dispersal of mistletoe (e.g.
Phainopepla nitrens in North America, Walsberg 1975; Dicaeum species in southern
Asia and Australia, Davidar 1983; Reid 1986), generalist frugivores also contribute to
the dispersal of mistletoe fruit (e.g. Colius indicus in South Africa, Godschalk 1983b;
Acanthagenys rufogularis in Australia, Reid 1989). Mistletoe specialists exhibit
specific anatomical adaptations to a mistletoe diet, which results in extremely rapid gut
passage times of mistletoe seeds compared to dietary generalists (Murphy et al. 1993).
Dietary generalists may therefore be particularly important in the establishment of new
mistletoe populations (Mathiasen et al. 2008), due to the fast gut transit time of
mistletoe specialists often precluding long dispersal distances. Several studies have
demonstrated that the ‘seed rain’ of specialist mistletoebirds (Dicaeum hirundinaceum)
is limited, as they tend to visit trees that are already infected with mistletoe, thereby
maintaining only existing populations of mistletoe (Ward and Paton 2007; Rawsthorne
et al. 2012). The establishment of new populations of mistletoes is likely to be carried

9
out by non-specialists such as the spiny-cheeked honeyeater (Aca. rufogularis), which
has been reported to visit both infected and uninfected stands within the (longer) time
taken to defecate mistletoe seeds (Rawsthorne et al. 2011). Migrating generalists, such
as silvereyes (Zosterops lateralis), have also been speculated to be dispersers of
mistletoe to islands, through the deposition of mistletoe seeds stuck to legs or feathers
(Watson 2011).
Several recent studies have examined how mistletoes may influence changes in
Australian bird communities (see studies by Turner 1991; Watson 2002; Watson et al.
2011; Watson and Herring 2012), but the interactions between mistletoes and birds in
Western Australia (WA) have not been studied in great detail. I therefore examined the
importance of mistletoe as a food resource to bird communities in south-west WA
through the use of field surveys and stable isotopes analysis. This was achieved by
conducting bird and mistletoe flowering and fruiting phenology surveys approximately
every five weeks from February 2010 and January 2011 (Chapter 3). Stable isotopes of
carbon and nitrogen measured in plant and animal tissue samples were subsequently
used to investigate the relative importance of mistletoe fruit and insects as a source of
dietary protein to mistletoebirds at three of the five study sites (Chapter 4).
1.3 PHYSIOLOGICAL ADAPTATIONS TO FRUGIVORY
Fruit pulp and nectar is the ‘reward’ offered to dispersers and pollinators; however
sugary fruits and nectar are characterised by a high water and carbohydrate content,
generally containing low levels of lipids and protein (Herrera 1987; Nicolson 1998).
Specialist frugivores and nectarivores must therefore assimilate large amounts of sugars
and process large volumes of preformed water, while conserving proteins and salts

10
(Yokota et al. 1985; Beuchat et al. 1990; Murphy et al. 1993; Witmer and van Soest
1998; McWhorter and Martinez del Rio 1999; Levey and Martinez Del Rio 2001;
McWhorter et al. 2003; Lotz and Martínez del Rio 2004). The reliance on a
predominantly frugivorous diet is associated with several specialised adaptations to
frugivory that allows these birds to meet their relatively high energy demands.
Firstly, specialised frugivores typically demonstrate a wide and short alimentary
tract (Richardson and Wooller 1988; Witmer 1998). In mistletoebirds, this allows for
the rapid processing of large numbers of mistletoe fruit, with mistletoebirds
demonstrating shorter gut retention times in comparison to non-specialised frugivores
(discussed in Chapter 6). To rely on mistletoe fruit as a main dietary source of
carbohydrates and nitrogen, these birds must be able to efficiently utilise these nutrients.
Most frugivorous birds, like specialised nectarivores, demonstrate high assimilation
efficiencies of the sugars found in fruit (Witmer 1999; Lotz and Schondube 2006).
The fruit pulp of fruits consumed by birds in generally rich in the
monosaccharides glucose and fructose (collectively termed hexoses), while the
disaccharide sucrose averages only 8% of total sugars in passerine-consumed fruits
(Baker et al. 1998). While glucose and fructose are quite easily absorbed by both
passive (i.e. paracellular) and mediated (active) mechanisms, sucrose must be
hydrolysed to its hexose components by the intestinal enzyme sucrase-isomaltase
(hereafter referred to as sucrase) before it can be absorbed (Nicolson and Fleming
2003b). The ability to assimilate sucrose varies widely between different bird species
due to the presence or absence of sucrase; frugivores in the Sturnidae-Muscicapoidea
lineage that lack this enzyme are unable to tolerate sucrose (Martínez del Rio and

11
Stevens 1989; Brugger et al. 1993; Sabat and Gonzalez 2003; Gatica et al. 2006; Brown
et al. 2012). Most frugivores and nectarivores are able to efficiently assimilate both
sucrose and hexoses (Lotz and Schondube 2006; Fleming et al. 2008; Napier et al.
2008a). However, some opportunistically frugivorous birds such as the yellow-rumped
warbler (Dendroica coronata), which does express sucrase but has high rates of food
passage and intake, appear to be constrained by the time taken in the hydrolysis of
sucrose (Afik and Karasov 1995). This may explain the presence of hexoses as the
dominant sugars in bird consumed fruits.
The significant reliance on paracellular absorption for uptake of glucose and
small water-soluble nutrients in small birds and bats is thought to compensate for a
reduction in intestinal absorptive surface area (Caviedes-Vidal et al. 2007). These
animals face strong selection pressures to digest food rapidly in order to reduce digesta
mass carried during flight, and this may be accomplished by the rapid absorption of
glucose via the paracellular pathway. Paracellular absorption provides a non-saturable
absorptive process that matches glucose absorption capacity to acute changes in dietary
nutrient concentration (Afik et al. 1997; Ferraris 2001); therefore the relative
contribution of paracellular to total glucose absorption increases with an increase in diet
concentration, presumably due to increased digesta retention time in the intestine
(McWhorter et al. 2006; Napier et al. 2008b). Some frugivorous birds and bats (as well
as specialised avian nectarivores), are known to rely extensively on the paracellular
pathway for the absorption of glucose (Karasov and Cork 1994; McWhorter et al. 2006;
Tracy et al. 2007; Napier et al. 2008b; McWhorter et al. 2010), although similar tests
have not been carried out for specialist frugivores. The mediated absorption of glucose
has been examined in one specialised frugivore, the cedar waxwing (Bombycilla

12
cedrorum), which demonstrates three times the mediated transport of glucose in
comparison to the American robin (Turdus migratorius), a seasonal frugivore (Karasov
and Levey 1990). It has been suggested that this allows cedar waxwings to maintain
efficient absorption of sugars in spite of relatively high intake and processing rates of
fruit (Witmer and van Soest 1998).
Secondly, specialised frugivores have relatively low protein requirements in
comparison to dietary generalists (Witmer 1998). Fruits typically contain low amounts
of protein, and, while frugivory is a common dietary strategy, exclusive frugivory is
considered rare (Morton 1973). For most avian frugivores, fruits are a non-exclusive
food resource that is supplemented with animal protein (such as insects) and nectar.
Even specialist frugivores (such as mistletoebirds) exhibit diet switching and consume
insects during periods when protein requirements are particularly high (e.g. during
moult and female reproduction) (Higgins et al. 2006). Protein is a vital dietary
component for birds, as it is the principle structural and catalytic component of all
tissues and birds lack the enzymes necessary for the synthesis of nine of the 20 amino
acids required (Klasing 1998). Consequently, birds must consume dietary protein to
meet their essential amino acid requirements and to supply nitrogen for the synthesis of
non-essential amino acids (Klasing 1998).
Thirdly, a large proportion of fruit mass is taken up by indigestible seeds (30–
50% dry fruit mass, see Levey and Grajal 1991 and references therein) which increases
the energy expenditure of flight and can also decrease the capacity to fill the gut with
more food due to their bulk. The ability to efficiently process these seeds is therefore of
significant importance to frugivorous birds. Some frugivores may regurgitate seeds or

13
discard seeds before ingesting the fruit pulp; while several mistletoe dispersers
regurgitate mistletoe seeds, such as the streak-necked flycatcher (Mionectes striaticollis)
(Montaño-Centellas 2013) and Australian Raven (Corvus coronoides) (Watson 2011),
Australian mistletoe specialists defecate mistletoe seeds. Frugivorous birds that avoid
regurgitating seeds have demonstrated the ability to separate fruit pulp from seeds in the
intestine, and defecate seeds quicker than fruit pulp (Levey and Grajal 1991). This
rapid passage of seeds increases capacity for fruit consumption, and therefore
assimilation of nutrients from fruit pulp (Levey and Grajal 1991).
I investigated various aspects of digestive physiology in three focal avian
frugivorous species, including the apparent assimilation efficiency of sucrose and
hexoses, sugar preferences and the capacity to hydrolyse sucrose (Chapter 5). The
mechanisms of absorption of glucose and xylose (a pentose monosaccharide that is
abundant in some species of mistletoe fruit) were examined for the three focal avian
species (Chapter 6).
1.4 STUDY AIM
This thesis takes a multi-faceted approach in the investigation of the interactions
between two Australian loranthaceous mistletoes species (Amyema miquelii and A.
preissii), their hosts, and their avian consumers in south-west Western Australia.
1.5 MISTLETOE STUDY SPECIES
Two endemic Australian Loranthaceae mistletoe species common in south-west WA
were the focal mistletoe species for this thesis. Field work was conducted at five study

14
sites in south-west WA (Fig. 1.1) – detailed site descriptions are included in Chapters 2
and 3 (Tables 2.1 and 3.1).
The box (or stalked) mistletoe, A. miquelii, (Lehm. ex Miq.) Tiegh., is found in
open forest and woodlands dominated by Eucalyptus species. It is dependent on
eucalypts as a host, although it occasionally parasitises Acacia species (Western
Australian Herbarium 1998–; Watson 2011). Amyema miquelii is the most widespread
Australian mistletoe (Western Australian Herbarium 1998–; Watson 2011). It is often
found in extremely high abundances and is a popular nesting site for a wide range of
bird species (Watson 2011). It has red flowers, grouped in threes on individual stalks,
and smooth fruits that are green when unripe and yellow upon ripening (Fig. 1.2a, b).
This mistletoe is highly distinguishable across the landscape due to the yellow-bronzed
colour of its leaves when exposed to sunlight. Three study sites contained A. miquelii
parasitising Corymbia calophylla (Forrestfield and York 2) and Eucalyptus accedens
(Paruna).
The wireleaf mistletoe, A. preissii (Miq.) Tieghem, is found in open forest,
woodland and semi-arid woodlands, and is primarily dependent on Acacia species
although it has been recorded growing on other host species (Western Australian
Herbarium 1998–; Watson 2011). It has a wide spread inland distribution across the
mainland of the continent with scattered distributions extending to the coast of WA
(Western Australian Herbarium 1998–; Watson 2011). Amyema preissii has abundant
red flowers, borne in groups of three, with pale pink fruits (Fig. 1.2c, d). It is also a
popular foraging site for a range of insectivorous bird species (Watson 2011). It is the
only mistletoe with slight, needle shaped leaves, and despite its common name, its

15
foliage is succulent and rather delicate (Watson 2011). Two study sites contained A.
preissii parasitising Ac. baileyana, Ac. podalyriifolia (Araluen) and Ac. acuminata
(York 1).

16
Figure 1.1 The location of five study sites in south-west Western Australia.
Perth is indicated by a star for reference.

17
Figure 1.2 Two Loranthaceae mistletoe species common in south-west Western Australia: the box
(or stalked) mistletoe Amyema miquelii and wireleaf mistletoe Amyema preissii.
a) scenescent flowers and yellow leaves of A. miquelii, with the mistletoe moth (Comocrus behri); b) the
yellow ripe fruits of A. miquelii; c) open flowers and needle like leaves of A. preissii and d) ripening pink
fruits of A. preissii.

18
1.6 AVIAN STUDY SPECIES
1.6.1 Mistletoebird (Dicaeum hirundinaceum)
The mistletoebird (body mass mb ~8 g) is the only Australian species of the family
Dicaeidae, a group of passerine birds found mainly in southern Asia and New Guinea
(Richardson and Wooller 1988). The interaction between the mistletoebird and
mistletoe is considered the most specific of all seed dispersal systems in Australia
(Serventy 1973). The mistletoebird is able to grasp and swallow mistletoe fruits (after
removing the exocarp by squeezing the fruit with its beak) that are relatively large in
comparison to its small body size (Liddy 1983; Richardson and Wooller 1988).
Mistletoebirds also include nectar and insects in their diet (Reid 1990). The
mistletoebird displays a short, specialised alimentary tract that is typical of specialised
frugivores, which allows the rapid processing of the large number of mistletoe fruits
that are ingested (Richardson and Wooller 1988). The mistletoebird is locally nomadic,
and its presence tends to correspond with the availability of fruiting mistletoe (Higgins
et al. 2006; Rawsthorne et al. 2012). The distribution of the mistletoebird ranges
throughout mainland Australia to several Indonesian islands in the Arafura Sea (Higgins
et al. 2006). The sexes are dichromatic, with males (Fig. 1.3a) displaying a glossy
bluish-black head, wings and upperparts with a bright red throat, chest and undertail,
and a white belly with a central dark streak. Females are grey above with a grey streak
on a white belly, with a paler red undertail. Immature birds resemble paler females, but
have a bright orange beak and gape (Fig. 1.3b).

19
1.6.2 Silvereye (Zosterops lateralis)
The silvereye (mb~11 g), is a small generalist frugivore that feeds on fruit, nectar and
insects (Wilkinson 1931; Thomas 1980; Richardson and Wooller 1986). The silvereye
does not have a specialised intestinal tract, but in line with its ingestion of insects, has a
more muscular gizzard than specialist nectarivores (Richardson and Wooller 1986).
The silvereye has been recorded feeding on both fruits and flowers of A. preissii and A.
miquelii (see Reid 1986 and references therein), and defecating mistletoe seeds resulting
in successful establishment when transferred to suitable host branches (Keast 1958).
The silvereye is widespread throughout eastern, southern and western Australia, New
Zealand and many outlying islands, and is rather complex in its geographical variation –
nine subspecies are recognised in the region (Australia, New Zealand, Antarctica and
surrounding islands; Higgins et al. 2006). Zosterops lateralis chloronotus is recognised
in western South Australia and WA, where it is usually considered resident through
most of its range in WA, but is a seasonal visitor or nomadic in low rainfall areas of the
WA wheatbelt (Higgins et al. 2006). Plumage is usually indistinguishable between the
sexes, with Z. l. chloronotus (Fig. 1.3c) displaying a conspicuous ring of white feathers
around the eye, yellowish olive upperparts, with the head, back rump and uppertail
coverts duller and paler, and grey belly and undertail (Higgins et al. 2006).
1.6.3 Singing honeyeater (Lichenostomus virescens)
The singing honeyeater (mb ~20–35 g) is widely distributed throughout mainland
Australia, and is generally present in open shrublands and low woodlands that are
dominated by acacias. It is considered resident or sedentary throughout its range, with

20
four subspecies recognised (Higgins et al. 2001). The nominant L. v. virescens is
recognised in south-west and southern Australia (Higgins et al. 2001). There is no
sexual dimorphism in plumage. Birds have a distinctive black streak through the eye
bordered with a yellow streak below the eye, a white throat and white to grey underbody
with brown to grey streaks (Fig. 1.3d; Higgins et al. 2001). The singing honeyeater is a
considered a nectarivore that also ingests a relatively high proportion of insects and fruit
in its diet (Collins and Morellini 1979; Richardson and Wooller 1986). Like silvereyes,
singing honeyeaters also have a more muscular gizzard than specialist nectarivores
(Richardson and Wooller 1986). Singing honeyeaters have been recorded feeding on
the flowers and fruit of several mistletoe species, including A. miquelii and A. preissii,
though its role as a potential disseminating agent of mistletoe has not been investigated
(Reid 1986).

21
Figure 1.3 Digestive adaptations associated with consuming a mistletoe fruit diet were investigated
for three focal frugivorous bird species.
a) male mistletoebird, b) juvenile mistletoebird, c) silvereye, and d) singing honeyeater.
a) b)
c) d)

22
2 CARBON AND NITROGEN STABLE ISOTOPES OF AUSTRALIAN
MISTLETOES AND THEIR HOSTS
Kathryn R. Napier1,2
, Todd J. McWhorter3, Carlos Martínez del Rio
2, Patricia A.
Fleming1
1 School of Veterinary and Life Sciences, Murdoch University, Murdoch, WA 6150,
AUSTRALIA
2 Department of Zoology and Physiology, University of Wyoming, Laramie, WY 82071, USA
3 School of Animal and Veterinary Sciences, University of Adelaide, Roseworthy Campus, SA
5371, AUSTRALIA

23
STATEMENT OF AUTHOR CONTRIBUTION
K.R. Napier: Initiated the research, performed the leaf sampling and prepared
samples for stable isotopes analysis, entered and analysed the data, and wrote the
manuscript.
T.J. McWhorter: Assisted with the design and initiation of the research and
provided editorial comments to versions of the manuscript from draft to final versions.
C.M. del Rio: Provided training and assistance in stable isotopes analysis,
including preparation of samples and statistical analysis, and provided editorial
comments to final versions of the manuscript.
P.A. Fleming: Assisted with the design and initiation of the research, advised
and assisted K.R. Napier with statistical analyses, and provided editorial comments to
versions of the manuscript from draft to final versions.

24
2.1 ABSTRACT
Australia has 75 species of native loranthaceaous mistletoes. To date, very few data
have been published on the δ13
C values of Australian mistletoes, while mistletoe δ15
N
has not been investigated in Australian mistletoes to date. This paper presents δ13
C,
δ15
N and nitrogen and carbon content (%) data on two loranthaceous mistletoes
(Amyema preissii (Miq.) Tieghem and A. miquelii (Lehm. ex Miq.) Tiegh.) on five
unique mistletoe-host pairs from five sites in south-west Western Australia. We also
investigated the effects of nitrogen-fixing hosts (Acacia species) vs. non nitrogen-fixing
hosts (Eucalyptus species), aridity and season on these data. Australian loranthaceous
mistletoes demonstrate a relationship with δ13
C and δ15
N that is consistent with those
found in previous studies. Unlike other mistletoes studied to date, proportionally more
nitrogen was taken up by mistletoes parasitising nitrogen-fixing (and therefore nitrogen
rich) acacia hosts, compared to mistletoes on nitrogen-poor eucalypts. Australian
mistletoes regulate their water use in relation to the supply of nitrogen available from
the host.

25
2.2 INTRODUCTION
Loranthaceous mistletoes are predominantly arboreal hemiparasites, which parasitise
the xylem stream of their hosts (Calder 1983; Marshall et al. 1994) via the haustorium
(a vascular connection which attaches to a host plant, Lamont 1983b; Ehleringer and
Marshall 1995). Xylem-tapping mistletoes are dependent on their hosts for water
(Ehleringer et al. 1985) and dissolved nutrients, including nitrogen (Marshall and
Ehleringer 1990; Schulze et al. 1991; Marshall et al. 1994; Wang et al. 2007), but do
not rely extensively on organic solutes from the host phloem (Hull and Leonard 1964).
Studies of carbon (δ13
C) and nitrogen (δ15
N) stable isotope ratios have aided and greatly
developed our understanding of how mistletoes obtain nutrients like carbon and
nitrogen from their hosts. While hemiparasitic plants obtain virtually all water and
mineral nutrients from their host plants, their reliance on their hosts for carbon varies
widely, partly due to the photosynthetic capacity of the mistletoe (Press 1995).
High transpiration rates of mistletoes, as well as high resistances in the
hydrolytic pathway between host and mistletoe (especially at the haustorial interface,
Davidson and Pate 1992), are the principal mechanisms maintaining a water potential
gradient by which water and dissolved solutes flow towards the parasite (Davidson et
al. 1989a). A strong correlation between carbon isotope discrimination and the ratio
between carbon assimilation and transpiration (i.e. water use efficiency, Farquhar et al.
1982) first led to the generalisation that more positive (i.e. 13
C-enriched) values of δ13
C
are found in more water-efficient plants. Ehleringer et al. (1985) reported that δ13
C
values for mistletoes were generally more negative than those of their hosts in samples
taken from arid or semi-arid regions of Europe, Africa, North America and Australia,
suggesting mistletoes are more profligate in their water use (Bannister and Strong

26
2001). In more mesic environments, however, where water is not limiting, both
mistletoe and host may transpire freely and gain an adequate supply of water and
xylem-transported nutrients (Panvini and Eickmeier 1993; Bannister and Strong 2001),
leading to smaller differences in δ13
C between mistletoes and hosts.
Ehleringer et al. (1985) also demonstrated that differences in δ13
C between
mistletoe and host were least for pairs where the host has high nitrogen content. Large
differences in δ13
C between mistletoes and hosts appear to be a consequence of high
leaf transpiration rates in mistletoes, necessary for the parasites to extract sufficient
nitrogen from the xylem of a nitrogen-poor host. Small differences in δ13
C are
associated with lower water conductance rates that are adequate to extract sufficient
amounts of nitrogen from a nitrogen-rich host. Therefore, nitrogen content (Ehleringer
et al. 1986; Bannister 1989) and δ15
N (Schulze et al. 1991; Wang et al. 2007;
Tennakoon et al. 2011) of mistletoes and hosts are usually strongly correlated.
The differences in δ13
C may also be related to the degree of heterophic carbon
gain (i.e. the proportion of carbon in mistletoes obtained from its host) by mistletoes.
Smaller differences in δ13
C are expected if mistletoes obtained higher amounts of
carbon from their hosts (Bannister and Strong 2001). Heterotrophic carbon gain ranges
from 5 to ~80% worldwide (see review by Bell and Adams 55 2011), but Australian
mistletoes studied to date appear to source only 5 to 21% of their carbon from the host
(Marshall et al. 1994).
Australia has 75 species of native loranthaceaous mistletoes which are found in
wooded habitats throughout the mainland (Barlow 1984; Barlow 1992; Watson 2011).
Very few data have been published on the δ13
C values of Australian Loranthaceae

27
mistletoes to date (Marshall et al. 1994; Miller et al. 2003). This paper presents δ13
C
and δ15
N stable isotope ratios and nitrogen and carbon content (%) data on six
mistletoe-host pairs and uninfected host plants from five sites in south-west Western
Australia (WA). We investigated the differences between δ13
C, δ15
N, nitrogen and
carbon content (%) between mistletoes and their hosts (where hosts are either nitrogen-
fixing acacias or non-nitrogen-fixing eucalypts), and determine the effects of aridity and
season on these data.
2.3 METHODS
2.3.1 Field sites and sampling
The study was conducted at five sites (100 x 200 m) in south-west WA where mistletoe
was extremely abundant (see Table 2.1 for site details). Two sites contained Amyema
preissii (Miq.) Tieghem parasitising acacia hosts, while three contained A. miquelii
(Lehm. ex Miq.) Tiegh. parasitising eucalypt hosts. Mistletoes and their host plants as
well as uninfected host plants from the same sites (nearby plants that had no visible
signs of previous or current mistletoe infestation) were sampled at each location in May
(autumn) and October (spring) 2010 and March (summer) 2011. A total of 212
mistletoe-host pairs and 85 uninfected host plants were sampled; all had similar
exposure in the canopy. Host leaves were collected proximally to a mistletoe
infestation (Bannister and Strong 2001). Approximately 15-20 fully-expanded leaves
from each plant were collected and combined for isotopic analysis.
Leaf samples were dried at 60 °C for 72 hours, then ground to a fine powder and
homogenised. Prepared samples were analysed at the University of Wyoming Stable

28
Isotope Facility, USA. Total carbon and nitrogen content (%) and δ13
C and δ15
N
compositions (‰) were determined using a Finnigan Delta Plus XP continuous flow
inlet Stable Isotope Ratio Mass Spectrometer online with a Costech EA 1108 Elemental
analyser or CE Elantech 2500 CHN Elemental analyser. δ13
C values are expressed with
respect to Vienna Pee Dee Belemnite (V-PDB) and δ15
N values with respect to
atmospheric molecular nitrogen.

29
Table 2.1 The characteristics of the five field sites in south-west Western Australia.
Rainfall range is shown for ‘winter’ (May to September inclusive). Weather data obtained for 1981–2010 (Bureau of Meteorology 2011) from the nearest meteorological station to
the study areas: Araluen and Forrestfield – Gosnells City (32°02′S, 115°58′E); Paruna – Pearce RAAF (31°40′S, 116°1′E); York 1 and York 2 – York (31°53′S, 116°46′E).
Site Mistletoe species Host species Host
system
Aridity
scale
Site description Days with Temp. max.
>35°C, min <2°C; Avg.
annual rainfall (winter
rainfall range)
Nitrogen fixing hosts
Araluen 1
Araluen 2
Araluen Country Club,
Roleystone, WA
(32°08′S, 116°05′E)
Amyema preissii
A. preissii
Acacia baileyana
Ac.podalyriifolia
Acacia
Acacia
Mesic
Mesic
A heavily watered suburban garden
with both non-native and native plants
present
35, 1 day
795 mm (83–165 mm)
York 1 Private farming
property, York, WA
(31°51′S, 116°44′E)
A. preissii Ac. acuminata Acacia Semi-arid A fragmented, semi-arid acacia
woodland subject to grazing
45, 47 days
401 mm (44–71 mm)
Non–fixing hosts
Paruna Paruna Sanctuary,
Avon Valley, WA
(31°41′S, 116°7′E)
A. miquelii Eucalyptus accedens Eucalyptus Intermediate Australian Wildlife Conservancy,
pristine warm temperate eucalypt
woodland
31, 3 days
669 mm (70–133 mm)
Forrestfield Forrestfield, WA
(32°0′S, 116°1′E)
A. miquelii Corymbia calophylla Eucalyptus Mesic Roadside suburban reserve in a
fragmented low eucalypt woodland
35, 1 day
795 mm (83–165 mm)
York 2 Private farming
property, York, WA
(31°50′S, 116°44′E)
A. miquelii C. calophylla Eucalyptus Semi-arid A fragmented, semi–arid eucalypt
woodland, subject to grazing
45, 47 days
401 mm (44–71 mm)

30
2.3.2 General statistics
δ13
C, δ15
N, C and N content (%) and Δδ13
C values for all leaf samples were analysed as
dependent variables in separate mixed-model (MM) ANOVAs with plant sample
(mistletoe, host or uninfected host leaves), host taxon (acacia or eucalypt), and degree of
aridity (i.e. mesic, intermediate and semi-arid) as categorical factors, and season
(autumn, spring, and summer) as a random factor (see Table 2.2). Linear regression
was used to assess the relationships between mistletoe and host δ15
N/ δ13
C,
nitrogen/carbon content (%), and differences in mistletoe and host δ13
C values (Δδ13
C,
‰). Data are reported as mean ± 1 SD throughout. Statistical analyses were performed
with Statistica (StatSoft Inc 2007). Statistical significance was accepted for α<0.05.

31
Table 2.2 Leaf carbon and nitrogen content (%), δ13
C and δ15
N values (‰) of mistletoe, host and uninfected host plants.
Data are averaged over three collection periods (May (Autumn) and October (Spring) 2010, March (Summer) 2012), with the apparent heterotrophy of mistletoes calculated from
equation 2.1 and n referring to number of mistletoe and host pairs. Data are presented as mean ± 1 SD.
%C %N δ13C (‰) δ15N (‰) Site Aridity Mistletoe Host Mistletoe Host Mistletoe Host Mistletoe Host Uninfected
host Mistletoe Host Uninfected
host n
Nitrogen-fixing hosts
Araluen 1 Mesic Amyema preissii Acacia baileyana 44.06±1.95 52.31±4.38 3.93±0.84 2.41±0.38 -30.84±0.99 -29.78±1.72 -31.14±1.05 -0.55±1.15 -0.24±0.97 -0.62±0.51 36
Araluen 2 Mesic A. preissii Ac. podalyriifolia 44.16±2.20 53.05±1.49 3.49±0.80 2.22±0.33 -30.37±0.98 -28.89±1.39 -29.21±0.55 -0.69±0.83 -0.57±1.17 -0.78±0.68 50
York 1 Arid A. preissii Ac. acuminata 46.86±2.06 51.42±1.23 2.88±0.56 2.45±0.20 -29.17±0.98 -27.72±0.77 -27.63±0.95 1.52±0.61 1.17±0.70 1.36±0.93 37
Non-fixing hosts
York 2 Arid A. miquelii Corymbia calophylla 50.75±1.11 49.49±0.86 1.04±0.19 1.09±0.11 -29.15±0.90 -26.55±0.98 -27.39±1.22 3.20±1.19 1.79±1.02 1.47±0.95 24
Forrestfield Semi-arid A. miquelii C. calophylla 51.19±1.34 51.25±1.22 0.80±0.20 1.06±0.15 -30.40±0.78 -28.27±0.57 -28.76±0.84 -3.44±0.57 -3.64±0.28 -3.42±4.53 39
Paruna Semi-arid A. miquelii Eucalyptus accedens 50.15±4.21 51.79±1.57 0.65±0.16 0.84±0.07 -31.20±0.60 -28.27±0.87 -28.82±0.59 -1.82±1.50 -1.47±0.90 -2.03±0.79 26

32
2.4 RESULTS
δ13
C, δ15
N, C and N content (%) values of mistletoes, infected hosts and uninfected
hosts are summarised in Table 2.2.
2.4.1 Carbon
δ13
C values (‰) and carbon content (%) of leaves differed significantly between
mistletoes, hosts, and uninfected host plants (Table 2.3), with host plants and uninfected
hosts significantly enriched in δ13
C compared to mistletoe leaves (Fig. 2.1a) and
mistletoes having a lower carbon content than the host plants (Fig. 2.1c). Host genus
had a significant effect on δ13
C values and carbon content (%) (Table 2.3) of plant
samples (i.e. mistletoe and infected and uninfected host leaves), with eucalypt infected
hosts and uninfected host plants enriched in δ13
C compared to samples taken from
acacia plants (infected host and uninfected hosts, Fig. 2.1a), and mistletoes on eucalypt
hosts demonstrating a higher percentage of carbon compared to mistletoes on acacia
hosts (Fig. 2.1c). Samples (including mistletoe and infected and uninfected host leaves)
collected from semi-arid sites were enriched in comparison to mesic and intermediate
sites, but degree of aridity had no effect on carbon % (Table 2.3). δ13
C and carbon %
values did not differ significantly with season (Table 2.3).
2.4.2 Nitrogen
δ15
N and nitrogen content (%) differed significantly between mistletoes, infected and
uninfected host plants (Table 2.3), with samples taken from acacia plants (mistletoe and
infected and uninfected hosts) significantly enriched in δ15
N and with higher nitrogen

33
contents compared to samples taken from eucalypt plants (mistletoe and infected and
uninfected hosts) (Figs. 2.1b, d). Nitrogen content from mistletoes on acacia hosts was
higher compared to infected and uninfected acacia hosts (Fig. 2.1d). Degree of aridity
had a significant effect on δ15
N values and nitrogen content (%) (Table 2.3), with
samples from semi-arid sites enriched in δ15
N and with higher N content by comparison
with mesic and intermediate sites. Season also had a significant effect on δ15
N values
and nitrogen content (%) (Table 2.3), with δ15
N values more enriched in autumn
samples compared to spring and summer samples, while nitrogen content was highest in
spring and lowest in summer.

34
Table 2.3 δ13
C and δ15
N values (‰) and carbon and nitrogen content (%) for all samples (mistletoe, infected and uninfected hosts) were analysed as dependent variables
in separate mixed-model ANOVAs.
Categorical factors included plant sample, host genus and degree of aridity. Season was set as a random factor. Significant variables are indicated in bold.
C (%) N (%) δ13C (‰) δ15N (‰)
Plant sample (mistletoe, host, uninfected host) F2,502=127, P<0.001 F2,503=103, P<0.001 F2,503=170, P<0.001 F2,503=6.70, P=0.001
Host genus (acacia or eucalypt) F1,271=35.9, P<0.001 F1,46=1030, P<0.001 F1,185=21.1, P<0.001 F1,51=82.7, P<0.001
Degree of aridity (mesic, intermediate and semi-arid) F2,268=0.23, P=0.794 F1,45=6.58, P=0.003 F2,181=107, P<0.001 F2,50=211, P<0.001
Season (autumn, spring, summer) F2,501=1.78, P=0.170 F2,501=6.85, P=0.001 F2,501=2.55, P=0.079 F2,501=6.38, P=0.002

35
C (
%)
40
42
44
46
48
50
52
54
56
58
60
13C
(‰)
-32
-31
-30
-29
-28
-27
-26Acacia
Eucalypt
N (
%)
0
1
2
3
4
51
5N
(‰
)
-6
-4
-2
0
2
4a) b)
c) d)
mistletoe host uninfected host mistletoe host uninfected host
mistletoe host uninfected host mistletoe host uninfected host
Figure 2.1 Stable isotope values and percentages of carbon and nitrogen for mistletoe, infected and
uninfected host samples on acacia (black symbols) or eucalyptus (white symbols) hosts.
a) δ13
C values (‰), b) δ15
N values (‰), c) C content (%), and d) N content (%). Data are presented as
mean ± 1 SD.

36
2.4.3 Relationships between mistletoes and hosts
The δ13
C values of mistletoes are significantly correlated with those of their hosts
(linear regression: F1,210=91.9; P<0.001, R2=0.304; Fig. 2.2a), even though there is
considerable variation around the regression line. Mistletoes were generally more
depleted in δ13
C than their hosts (Fig. 2.1a, Table 2.2), but for 18 individual samples
(out of 212), mistletoes were more positive than their hosts (Fig. 2.2a). The relationship
between mistletoe and host carbon content (linear regression: F1,207=32.9; P<0.001,
R2=0.137) significantly deviates from a 1:1 ratio (95% confidence interval of the slope:
-0.48 to -0.99; Fig. 2.2b), indicating that mistletoes do not obtain all of their carbon
from their hosts. Differences in δ13
C between mistletoes and hosts were significantly
affected by host taxon (F1,190=54.7; P<0.001) and degree of aridity (F2,189=3.50;
P=0.031), but not season (F2,205=1.16; P=0.317); differences were greater on nitrogen-
poor eucalypt hosts compared to nitrogen-rich acacia hosts (Fig. 2.2e), and were greater
at mesic and semi-arid sites compared to intermediate sites.
Both δ15
N and nitrogen content (%) of mistletoe leaves are highly correlated
with those of their hosts. The relationship between mistletoe and host δ15
N (F1,210=862;
P<0.001, R2=0.804) was not significantly different from 1 (95% confidence interval of
the slope: 0.96 to 1.10; Fig. 2.2c). The relationship between mistletoe and host nitrogen
content (F1,210=612.65; P<0.001, R2=0.745) deviates from a 1:1 ratio (95% confidence
interval of the slope: 1.59 to 1.87; Fig. 2.2d). Mistletoes on nitrogen-rich acacia hosts
tend to have higher leaf nitrogen concentrations than their hosts (F1,121=4.25; P=0.041,
R2=0.03; relationship deviates from a 1:1 ratio with the 95% confidence interval of the
slope from 0.019 to 0.923; Fig. 2.2d), compared to mistletoes on nitrogen-poor eucalypt

37
hosts (F1,87=45.3; P<0.001, R2=0.342; 95% confidence interval of the slope: 0.613 to
1.13; Fig. 2.2d; slope differs significantly from mistletoes on acacia hosts F2,208=9.1;
P<0.001 ). The differences between mistletoe and host δ13
C (Δδ13
C) were greatest
when host leaf nitrogen content was low (linear regression: F1,4=24.45; P=0.008,
R2=0.859; Fig 2.2e).

38
Col 19 vs Col 21
Col 19 vs Col 21: 2.4100
Col 19 vs Col 21: 2.2200
Col 19 vs Col 21: 2.4500
Col 19 vs Col 21: 1.0900
Col 19 vs Col 21: 1.0600
Col 19 vs Col 21: 0.8400
Plot 1 Regr
Col 21 vs Col 23
Col 21 vs Col 23: -1.0440
Col 21 vs Col 23: -1.4880
Col 21 vs Col 23: -1.4260
Col 21 vs Col 23: -2.6020
Col 21 vs Col 23: -2.1320
Col 21 vs Col 23: -2.2940
13C host
-36 -34 -32 -30 -28 -26 -24 -22
13C
mis
tle
toe
-36
-34
-32
-30
-28
-26
-24
-22
Host C %
38 40 42 44 46 48 50 52 54 56 58
Mis
tle
toe
C %
38
40
42
44
46
48
50
52
54
56
58a) b)
F1,210
=91.91; P<0.001, R2=0.304
Host N (%)
0.5 1.0 1.5 2.0 2.5 3.0
13C
mis
ttle
toe
-ho
st, ‰
-4
-3
-2
-1
0
1
c)
Host 15
N (‰)
-6 -4 -2 0 2 4 6
Mis
tle
toe
1
5N
(‰
)
-6
-4
-2
0
2
4
6 d)
Host N (%)
0 1 2 3 4 5 6
Mis
tle
toe
N (
%)
0
1
2
3
4
5
6
Araluen 1 (A, mesic)
Araluen 2 (A, mesic)
Forrestfield (E, semi-arid)
Paruna (E, semi-arid)
York 1 (A, arid)
York 2 (E, arid)
e)
F1,210
=862, P<0.001, R2=0.804
F1,4
=24.5, P=0.008, R2=0.859
F1,210
=612, P<0.001, R2=0.342
F1,207
=32.86; P<0.001, R2=0.137
F1,121
=4.25, P=0.04, R2=0.03
F1,87
=45.3, P<0.001, R2=0.745
Figure 2.2 Correlations between mistletoe and host a) δ13
C (‰), b) carbon content (%), c) δ15
N (‰)
and d) nitrogen content (%)
Dashed lines represent a 1:1 correspondence between mistletoe and host nitrogen. e) relationship
between average differences in mistletoe and host δ13
values (Δδ13
C, ‰) with standard deviations as error
bars. Grey symbols indicate Amyema miquelii on nitrogen-poor eucalypt hosts (E), white symbols
indicate A. preissii on nitrogen-rich acacia hosts (A).

39
2.5 DISCUSSION
We found significant effects of plant sample (mistletoe, uninfected and infected hosts),
host taxon (eucalypt, acacia), aridity and season on δ13
C and δ15
N values and on carbon
and nitrogen content. These patterns reflect water and nitrogen relations in these plants
which reveal some differences in the ecology of Australian mistletoes compared with
other mistletoes previously described. The differences between mistletoe and host δ13
C
were greatest at semi-arid, and surprisingly also at mesic sites on nitrogen-poor eucalypt
hosts, indicating lower water use economy by mistletoes in both of these situations.
2.5.1 δ13
C of mistletoes and hosts: water relations and effects of aridity
The range of δ13
C values reported in this study are consistent with previously published
results, with values of approximately -28 to -30‰ reported for Australian mistletoes and
approximately -26‰ for their hosts (Marshall et al. 1994; Miller et al. 2003). As dry
conditions reduce stomatal conductance, which leads to reduced discrimination against
δ13
C, water-stressed plants in more arid environments (such as York in this study) tend
to have more positive δ13
C values than more mesic environments (such as Araluen and
Forrestfield) (Farquhar et al. 1989; Stewart et al. 1995). Agricultural land-use
practices, which alter the exposure of plants to sunlight, may also influence plant δ13
C
values, resulting in more enriched δ13
C values compared to forested areas due to water
use efficiency mechanisms and the removal of the canopy effect which tends to lower
the δ13
C values of plants (Hobson 2007).
Given that mistletoes need to maintain a water potential gradient in order to
receive water and nutrients from their hosts, host water status has been hypothesised to

40
be an important determinant of mistletoe growth (Miller et al. 2003). However, like
Miller et al. (2003), we found no differences between δ13
C leaf values between infected
and uninfected hosts, suggesting that infection with mistletoe is not leading to increased
water stress on these hosts. Average mistletoe δ13
C values were lower than their hosts
at all sites, supporting the relationship observed between mistletoe and host δ13
C
observed in semi-arid and arid locations world wide (Ehleringer et al. 1985; Schulze et
al. 1991; Marshall et al. 1994). The significant differences between mistletoe and host
δ13
C values appear to be related to both nitrogen availability (between nitrogen-rich
acacia and nitrogen-poor eucalypt hosts, Fig. 2.2e), and differences in water availability
between semi-arid and intermediate sites.
2.5.2 Relationships with nitrogen
Like other mistletoe species studied to date (Schulze et al. 1991; Bannister and Strong
2001; Wang et al. 2007; Tennakoon et al. 2011), the two species of Australian
loranthaceous mistletoes in this study also acquire nitrogen from their hosts, confirmed
by strong correlations between mistletoe and host leaf δ15
N values (Fig 2.3c). Nitrogen
contents of mistletoes and their hosts are usually also strongly correlated, with a slope
that deviates from a one-to-one relationship. Previous studies report lower nitrogen
concentrations in mistletoes compared to nitrogen-rich hosts, but higher nitrogen
concentrations in mistletoes compared to nitrogen-poor hosts (Ehleringer et al. 1986;
Bannister 1989; Schulze et al. 1991; Bannister and Strong 2001). However, this was
not the case in the present study, with mistletoes having lower concentrations of
nitrogen in comparison to their nitrogen-poor eucalypt hosts, but more nitrogen than
their hosts for mistletoes parasitising nitrogen-rich acacia hosts (Fig. 2.2d).

41
The relationship between the differences in mistletoe and host δ13
C and host leaf
nitrogen content is similar to those predicted and found by Ehleringer et al. (1985) and
Schulze et al. (1991) - the difference in δ13
C is least when host nitrogen content is high
(i.e. acacia hosts, Fig. 2.2c). These differences in water use efficiency between
mistletoe and host are therefore related to the nutritional quality of the host - the most
negative differences in δ13
C suggest that a greater differential in water use efficiency is
required to extract nitrogen from nitrogen-poor eucalypt hosts.
2.5.3 Conclusions
Two species of Australian loranthaceous mistletoes demonstrate a relationship with
δ13
C and δ15
N that is consistent with those found in previous studies. Unlike other
mistletoes studied to date, proportionally more nitrogen was taken up by mistletoes on
nitrogen-rich acacia hosts, compared to mistletoes on nitrogen-poor eucalypt hosts. A
greater differential in water use efficiency is therefore required for mistletoes on
nitrogen-poor eucalypt hosts to extract nitrogen, resulting in more negative differences
in δ13
C in comparison to mistletoes on acacia hosts. Differences in water use efficiency
between Australian mistletoes and hosts are therefore related to the nutritional quality of
the host, with Australian mistletoes regulating their water use in relation to the supply of
nitrogen available from the host.

42
3 DO BIRD SPECIES RICHNESS AND COMMUNITY STRUCTURE VARY
WITH MISTLETOE FLOWERING AND FRUITING IN WESTERN
AUSTRALIA?
Kathryn R. Napier1, Suzanne H. Mather
2, Todd J. McWhorter
3, Patricia A. Fleming
1
1 School of Veterinary and Life Sciences, Murdoch University, Murdoch, WA 6150,
AUSTRALIA
2 3 Hardy Road, Nedlands, WA 6009, AUSTRALIA
3 School of Animal and Veterinary Sciences, University of Adelaide, Roseworthy Campus, SA
5371, AUSTRALIA
This chapter was accepted for publication in the journal Emu on March 25, 2013
(published online early, http://dx.doi.org/10.1071/MU12098).

43
STATEMENT OF AUTHOR CONTRIBUTION
K.R. Napier: Initiated the research, performed the mistletoe phenology and
bird surveys, collected and entered data, analysed the data, and wrote the manuscript.
S.H. Mather: Provided training and assistance in the conduction of bird
surveys, and provided editorial comments to a draft version of the manuscript.
T.J. McWhorter: Assisted with the design and initiation of the research and
provided editorial comments to versions of the manuscript from draft to publication.
P.A. Fleming: Assisted with the design and initiation of the research, advised
and assisted K.R. Napier with statistical analyses, and provided editorial comments to
versions of the manuscript from draft to publication.

44
3.1 ABSTRACT
Worldwide, mistletoes act as a keystone resource, providing food (nectar, fruit and
foliage) and structural (nesting sites) resources to hundreds of fauna species. In
Australia, loranthaceous mistletoes depend on birds for pollination and dispersal, and
provide important nectar and fruit resources to a large number of nectarivorous and
frugivorous bird species. We investigated whether bird species richness and community
structure varies with flowering and fruiting of two common mistletoe species (family
Loranthaceae, Amyema preissii and A. miquelii) through monthly surveys for one year
at five sites in south-west Western Australia. Flowering and fruiting periods were
distinct and differed both amongst sites and between mistletoe species. Nectar and ripe
fruit were available for up to 5 and 6–7 months (A. miquelii and A. preissii respectively)
at individual sites, but were available every month of the year across all sites. The
presence of fruiting, but not flowering, mistletoe was associated with changes in bird
community structure. Mistletoebirds (Dicaeum hirundinaceum) were significantly more
likely to be recorded during months when ripe mistletoe fruit was present and the
overall bird species richness was higher for these survey months. Mistletoes provide
important resources, but further investigation is required to assess keystone species
status in south-west WA.

45
3.2 INTRODUCTION
Mistletoes are a polyphyletic group of shrubby, aerial hemiparasitic flowering plants
with over 1,500 species found over a wide range of habitats and across all continents
with the exception of Antarctica (Kuijt 1969; Calder 1983; Watson 2001). A diversity
of ‘showy’ mistletoes (family Loranthaceae) is native to Australia, with 75 species
currently recognised (Barlow 1984; Barlow 1992; Watson 2011). Watson et al. (2001)
first proposed that mistletoes act as a keystone resource (sensu Power et al. 1996) in
forests and woodlands worldwide, due to the pervasive effects they have on these
habitats through the provision of nutritional and nesting resources (confirmed by
Watson and Herring 2012). Peres (2000) identified four criteria used to define a
keystone plant: reliability and abundance of resources, degree of consumer specificity
and temporal redundancy. Kotliar (2000) further proposed that keystone species should
perform functions not otherwise carried out. In this study, we address whether two
species of loranthaceous mistletoes in south-west Western Australia (WA) meet the
keystone criteria established by these authors.
In Australia, mistletoe nectars and fruits are consumed by at least 50 bird
species, including several honeyeater species (Keast 1958; Reid 1987; Turner 1991;
Brown et al. 1997). In addition to providing a nutritious food source, many mistletoe
species also have extended flowering and fruiting phenologies that minimise
competition with other plant species, but is also important for sustaining populations of
their avian pollinators and dispersers. Australian mistletoes continually draw upon their
host’s water and nutrient resources and consequently are able to flower and set fruit
during dry seasons when little other nectar or fruit is available in the landscape (Paton

46
and Ford 1977; Reid 1986; Watson 2001). In addition to flowering and fruiting at
different times to most other plants, discontinuous ripening (both within a species and
amongst species within communities) extends flowering and fruiting periods (Reid
1986; Hawksworth and Wiens 1996; Watson 2001). For example, most temperate
southern Australian mistletoes flower during summer, contrasting with largely spring
and autumn flowering for the majority of ornithophilous plant species present in these
areas (Reid 1986; Watson 2011). In more arid areas, where mistletoes are a critical
food source to nectarivorous species, some mistletoe species flower during winter,
others during summer, while other species exhibit all year round flowering (Reid 1986;
Watson 2011).
Mistletoes also provide the most reliable (and sometimes the only) source of
fruit for their avian dispersers. Further, their semi-succulent leaves have a high
concentration of nutrients and are consumed by herbivores (Watson 2001). Their dense,
multibranched structure means that many mistletoe species offer important nest and
foraging sites for animals in an otherwise open canopy (see review by Watson 2001).
In Australia, bird species from more than 60 families (across 16 orders) and several
mammalian families nest in mistletoe, and 66% (of 330 species) of Australian arboreal
nesting bird species have been recorded using mistletoes as nest sites (Cooney et al.
2006). Many insectivorous species (e.g. thornbills, whistlers) also use mistletoe clumps
as a foraging substrate, as mistletoe often has abundant and distinctive insect
assemblages (Turner 1991; Watson 2001; Start 2011; Watson 2011); however Burns et
al. (2011) found no difference in insect assemblages. Mistletoes therefore reliably
provide important food and shelter resources for hundreds of fauna species, providing
resources and ecosystem services out of proportion to their abundance and contribution

47
to biomass (Davidson et al. 1989b; Watson 2001; Watson 2002; Mathiasen et al. 2008;
Watson and Herring 2012).
Loranthaceous mistletoes provide birds with important nectar and fruit resources
(Kuijt 1969; Calder 1983; Davidar 1985; Reid 1986; Ladley et al. 1997; Robertson et
al. 1999). With two exceptions, the root-parasitic Western Australian Christmas tree,
Nuytsia floribunda, and Atkinsonia, Atkinsonia ligustrina (Hawkeswood 1981; Watson
2011), all Australian loranthaceous mistletoes are pollinated by birds. These mistletoes
tend to display brightly-coloured, odourless flowers, with abundant, sugar-rich nectar
(up to 60% total sugar content, primarily glucose and fructose sugars, Reid 1986; Stiles
and Freeman 1993; Baker et al. 1998), characteristics typically associated with
ornithophilous pollination. Although they are not dependent on mistletoe nectar as a
primary food source, a wide range of bird species pollinate these mistletoes, including
several species of honeyeaters (Reid 1986; Watson 2001; Watson 2011). The degree of
consumer specificity between mistletoes and their pollinators has led several authors to
suggest that long term negative consequences for both interacting organisms and
perhaps indeed the entire ecosystem may ensue if this balance were disrupted (Reid et
al. 1995; Watson 2001).
Birds are also responsible for seed dispersal for nearly all Australian
loranthaceous mistletoes (with the exception of N. floribunda, which is wind dispersed
(Watson 2011)). Mature mistletoe fruits are often brightly coloured, fairly large and
sweet and act as a food source for obligate and opportunistic bird species worldwide
(see review by Watson 2001). The composition of the fleshy fruit pulp surrounded by a
layer of viscin varies among species, but most mistletoe fruits contain a high proportion

48
of carbohydrates, lipids and protein (López de Buen and Ornelas 2001; Watson 2001;
Barea 2008).
In addition to supporting generalist feeders, Australian mistletoes also support
two mistletoe fruit specialists. The mistletoebird, Dicaeum hirundinaceum, is found
throughout mainland Australia (Keast 1958; Blakers et al. 1984) and is locally nomadic,
its presence corresponding with the availability of fruiting mistletoe (Rawsthorne et al.
2012). The rare painted honeyeater, Grantiella picta, found across the eastern inland
side of the continent (Reid 1986), is considered the original (Australian) ‘mistletoe bird’
as the mistletoebird did not colonise Australia until possibly as recently as the Holocene
(~12,000 YBP, Reid 1991). The diversification and radiation of mistletoes across
Australia has therefore depended upon species such as the painted honeyeater and its
ancestors (Watson 2011).
Despite their importance as a food resource, there have been few studies
examining how mistletoe may influence changes in Australian bird communities (see
studies by Turner 1991; Watson 2002; Watson et al. 2011; Watson and Herring 2012).
Additionally, we have extremely limited information on mistletoe fruiting and flowering
phenology in south-west WA. The aim of this study was to investigate if bird species
richness and community structure varies with flowering and fruiting of two
Loranthaceae mistletoe species common in south-west WA, the wireleaf mistletoe
(Amyema preissii) and box mistletoe (A. miquelii). This study examined how flowering
and fruiting phenology compared with year-round bird community surveys. We
predicted that: 1) there would be a greater number of bird species present (i.e. greater
species richness) at our study sites when mistletoe flowers or fruit were available, and

49
2) bird community structure at our study sites would reflect variability in available
resources, and 3) mistletoebirds would only be present in an area when ripe mistletoe
fruit was available. We then discuss the findings of this study in relation to the
keystone criteria identified by Peres (2000) and Kotliar (2000).
3.3 METHODS
3.3.1 Study sites
The study was conducted at five sites (100 x 200 m) in south-west WA where mistletoe
was extremely abundant, from February 2010 to January 2011 (Table 3.1). Two sites
contained A. preissii parasitising Acacia spp. hosts, whilst three contained A. miquelii
parasitising Eucalyptus and Corymbia hosts. All sites were dominated by mistletoe
with few other sources of fruit or nectar present (Table 3.1), with the exception of
Araluen. Araluen is surrounded by a dense urban matrix, including flowering and
fruiting plants in extensive adjacent gardens; and while Forrestfield was also surrounded
by suburban matrix, the area is more rural with larger property sizes and more native
vegetation (Table 3.1). One of the sites (York 2) contained both species of mistletoe,
however A. preissii was present in extremely low densities on 2 individual trees only.
In south-west WA, flowering and fruiting records suggested that both mistletoe species
have narrow flowering and fruiting periods (A.N. Start, Western Australian Herbarium,
WA Department of Environment and Conservation, pers. comm.). Flowering (January
to March) and fruiting (April to July) periods of A. preissii differ from those of A.
miquelii in south-west WA, which has flowering records from March to April and
fruiting records from June, November and December (A.N. Start, pers. comm.).

50
South-west WA experiences hot summers and cool wet winters. York (in the
highly fragmented agricultural wheatbelt of WA) is situated further inland than the other
sites, and experiences a more extreme temperature range and lower rainfall compared to
the other sites (Table 3.1).
3.3.2 Fruit and flower phenology
The flowering and fruiting phenologies of A. preissii and A. miquelii were measured by
counts of flowers (categorised as ‘bud’, ‘open’ or ‘senescent’) and fruit (‘immature’,
‘unripe’, ‘ripe’ or ‘bare’) at approximately 5 week intervals. At each site, 30 cm
sections of tagged branches (measured proximally from the tip of the branch) of up to
21 randomly-selected mistletoe plants were monitored for each survey period, as per
Barea and Watson (2007). These branches ranged in height from 0.5–15m high. Each
tagged branch was monitored every 5 weeks, with counts made of flowers (buds
through senescence) and fruit (unripe through to stalks where fruit had been removed
(‘bare’)). As the total number of flowers and fruits varied between branches, an index
of relative fruit and flower abundance was calculated (expressed as the proportion of
fruit and flowers per 30 cm of branch, summing to 1) which were averaged across all
mistletoe plants for each site/host. Mistletoes surveyed included A. preissii parasitising
Acacia baileyana (n=21), Ac. podalyriifolia (n=21) and Ac. acuminata (n=21), and A.
miquelii parasitising Eucalyptus accedens (n=14) and Corymbia calophylla (n=16,
n=18; Table 3.1).

51
Table 3.1 The characteristics of the five sites surveyed in south-west Western Australia.
The range of all mistletoe plant heights are presented underneath species names, with heights of tagged mistletoe plants underlined. Dates exclude the month of July for all sites.
Rainfall range is shown for ‘winter’ (May to September inclusive). Weather data obtained for 1981-2010 (Bureau of Meteorology 2011) from the nearest meteorological station to
the study areas: Araluen and Forrestfield – Gosnells City (32°02′S, 115°58′E); Paruna – Pearce RAAF (31°40′S, 116°1′E); York 1 and York 2 – York (31°53′S, 116°46′E).
Site Total time recorded:
dates
Mistletoe
species
Host species Site description Vegetation Days with Temp. max. >35°C, min
<2°C; Avg. annual rainfall (winter
rainfall range)
1 ‘Araluen’ Araluen Country
Club, Roleystone,
WA
(32°08′S, 116°05′E)
11 months:
Feb 2010–Jan 2011
Amyema preissii,
0.5–2m
Acacia baileyana
Ac. podalyriifolia
(non WA natives)
A heavily watered suburban
garden with both non–native
and native plants present
Nerium oleander, Corymbia calophylla,
Eucalyptus wandoo, E. forestiana, as well
as various Eucalyptus, Grevillea and
Callistemon species and one species of
Rubus also present
35, 1 day
795 mm (83–165 mm)
2 ‘York 1’ Private farming
property, York, WA
(31°51′S, 116°44′E)
11 months:
Feb 2010–Jan 2011
A. preissii
0.5–2–5m
Ac. acuminata A fragmented, semi arid acacia
woodland subject to grazing
Dominated by Ac. acuminata 45, 47 days
401 mm (44–71 mm)
3 ‘Paruna’ Paruna Sanctuary,
Avon Valley, WA
(31°41′S, 116°7′E)
11 months:
Feb 2010–Jan 2011
A. miquelii
10–15–25m
E. accedens
E. wandoo
C. calophylla
Australian Wildlife
Conservancy, pristine warm
temperate eucalypt woodland
E. accedens and E. wandoo, with scattered
C. calophylla
31, 3 days
669 mm (70–133 mm)
4 ‘Forrestfield’ Forrestfield, WA
(32°0′S, 116°1′E)
11 months:
Feb 2010–Jan 2011
A. miquelii
2–4–20m
C. calophylla
E. wandoo
Roadside suburban reserve in
a fragmented low eucalypt
woodland
Dominated by C. calophylla with scattered
E. wandoo, with Hovea pungens, Goodenia
fasciculata and Banksia spp present
35, 1 day
795 mm (83–165 mm)
5 ‘York 2’ Private farming
property, York, WA
(31°50′S, 116°44′E)
9 months:
April 2010–Jan 2011
A. miquelii
2–4–20m
C. calophylla
E. wandoo
A fragmented, semi-arid
eucalypt woodland, subject to
grazing
Dominated by C. calophylla and E. wandoo.
Gastrolobium and Banksia spp. also
present, as well as Ac. acuminata
(parasitised with very low densities of Am.
preissii).
45, 47 days
401 mm (44–71 mm)

52
3.3.3 Bird species
Monthly surveys of bird species presence/absence at each site were carried out using a
standardised search method (Watson 2003). We did not attempt to estimate bird species
abundance due to different detectabilities of species. At least three 20-minute surveys
were conducted back-to-back, with the stopping rule (after three surveys being reached)
that the number of species seen in a single sampling period of 20 minutes was less than
or equal to the number of species seen in two previous subsequent sampling periods
(Watson 2003; Watson 2004b). Surveys commenced within 90 minutes after sunrise
(0503 to 0718).
3.3.4 Statistical analysis
Differences in fruiting and flowering periods were compared among sites (and host
species in the case of Araluen) and month by two-way ANOVA, with Tukey-Kramer
post hoc tests for unequal sample sizes as required. Proportions of ripe fruit and open
flowers were arcsine squareroot transformed prior to analysis to meet requirements of
parametric statistics.
1) Bird species richness (total number of bird species recorded) was compared
among sites and for months when flowers or fruit were present (‘open’ and ‘ripe’) or
absent (‘bud’, ‘senescent’ and ‘immature’, ‘unripe’, ‘bare’) by two-way ANOVA, with
Tukey-Kramer post hoc tests for unequal sample sizes as required.
2) Bird community structure: Bird data, classified as presence (1) or absence (0)
for each bird species for each monthly survey, were analysed by multidimensional

53
scaling (MDS) in the program PAST 2.08b (Hammer et al. 2001). Bird community
structure was then compared via two-way analysis of similarity (ANOSIM, Bray-Curtis
similarity matrices; PAST 2.08b), using site (1–5) and either flowering (comparing
months with or without ‘open’ flowers) or fruiting (comparing months with or without
‘ripe’ fruit available) as independent factors. The ANOSIM test statistic (R) contrasts
the differences among groups with variation within groups, a large positive R (up to 1)
signifying dissimilarity between groups; significance is calculated via permutation
(Clarke 1993). Similarity Percentage (SIMPER) was then used to assess which
individual species were primarily responsible for observed differences (Clarke 1993)
and then subsequently for feeding guilds (frugivore, nectarivore, insectivore, granivore
and omnivore, see Table 3.2).
3) Presence/absence of bird species: To examine the relationship of bird species
and mistletoe flower presence, contingency tables were constructed for each month of
the presence/absence of individual bird species compared to the presence/absence of
mistletoe flowers or fruit. These contingency tables were analysed for significance via
Fisher’s exact probability using STATISTICA (StatSoft Inc 2007), followed by a
Bonferroni correction.
Statistical significance was set to α<0.05. Results are presented as mean ± 1 SD.
3.4 RESULTS
41 bird species were recorded over 53 surveys at the five sites, including 2 frugivorous
(mistletoebird and silvereye) and 6 nectarivorous species (brown, New Holland, singing
and white-cheeked honeyeaters, red wattlebird and western spinebill, Table 3.2).

54
3.4.1 Presence of mistletoe flowers
Flowers (and therefore nectar) of both mistletoe species were available for 2 to 3
months at each site, spanning the Australian summer and autumn months (from
December to February; A. preissii, Fig. 3.1a, c, e; from December to May; A. miquelii,
Fig. 3.1b, d, f), with peak abundances occurring at different times at each site (Fig. 3.1).
A significant site by month interaction (F48,1014=59.66, P<0.001) showed that flowering
periods differed among sites and between mistletoe species. We note, however, that
differences between hosts at the same site did not affect flowering or fruiting phenology
(A. preissii was monitored on hosts Ac. baileyana and Ac. podalyriifolia at Araluen, see
Fig. 3.1c, e).
1) Bird species richness: There were no significant differences in bird species
richness (i.e. number of species recorded) between months when mistletoe was
flowering or not (flowers present: 11.1±4.1, n=13 monthly surveys; flowers absent:
9.8±4.0 bird species, n=40 surveys; F1,43<0.01, P=0.975), or differences among sites
(F4,43=0.57, P=0.686). However, a significant interaction term (site x presence of
flowers: F4,43=6.49, P<0.001, Fig. 3.2a), showed that Araluen had significantly higher
bird species richness compared to all other sites (with the exception of York 1) when
flowers were not present (outside the summer months, Tukey-Kramer post hoc tests).
There were also no significant differences in bird species richness when the presence of
nectarivorous species only (n=6 species) was compared for months when mistletoe was
flowering vs months when flowers were absent (flowers present: 1.9±1.8, n=13 monthly
surveys; flowers absent: 1.7±1.3 bird species, n=40 surveys; F1,43=1.11, P=0.297).
There was a significant difference among sites (F4,43=20.71, P<0.001), and a significant

55
interaction term (site x presence of flowers: F4,43=2.80, P=0.037, Fig. 3.2b)
demonstrated that even when flowers were not present, Araluen had significantly more
nectarivorous species compared to all other sites (with the exception of York 2, Tukey-
Kramer post hoc tests).
2) Bird community structure: Bird community structure also varied among sites
(two-way ANOSIM; site: R=0.75, P<0.001), but the presence of flowering mistletoe
(comparing months with or without ‘open’ flowers) did not have a significant effect on
bird community structure (flowering: R=0.08, P=0.208).
3) Presence/absence of bird species: The grey butcherbird (Cracticus torquatus)
was the only bird species that was more likely to be recorded during months when
mistletoe was flowering (Fisher’s exact test: P=0.042), while the western gerygone was
less likely to be recorded during these months (P=0.042). However, as these
differences were not significant after Bonferroni correction, these results will not be
discussed further. During surveys, four nectarivorous honeyeater species (brown, New
Holland, singing, and white-cheeked honeyeaters) were directly observed feeding on
flowers of both mistletoe species.
3.4.2 Presence of ripe mistletoe fruit
Ripe fruit appeared some 2–7 months after flowering, and was available for 2 to 5
months, with peak abundances occurring in winter (June and August: A. preissii, Fig.
3.1a, c, e) and spring/summer (November and January: A. miquelii, Fig. 3.1b, d, f). A
significant interaction term (site x month: F48,1014=26.66, P<0.001) demonstrated that
fruiting periods differed among sites and between mistletoe species.

56
1) Bird species richness: There were significantly more bird species present (i.e.
greater species richness) when surveys were carried out when ripe mistletoe fruit was
present (fruit present: 12.06±4.27, n=16 monthly surveys; fruit absent: 9.24±3.65, n=37
surveys; F1,43=7.94, P=0.007). There was also a significant difference among sites
(F4,43=13.60, P<0.001) and a significant interaction term (site x month: F4,43=3.77,
P=0.010; Fig. 3.2c) with the two sites surrounded by urban matrix showing signficiantly
higher (Araluen) or lower (Forrestfield) bird species richness (Fig. 3.2c).
2) Bird community structure: Bird community structure was significantly
different for months when ripe mistletoe fruit was present, compared with months when
fruit was absent or unripe (two-way ANOSIM; fruiting: R=0.28, P=0.002), with
significant differences also present among sites (site: R=0.88, P<0.001). The
mistletoebird made the greatest contribution of all species (n=41) to the distinction
between bird community structure for fruiting and non-fruiting months (6.3%,
SIMPER). When analysed for the five feeding guilds (SIMPER, standardised by the
number of species within each guild), the two frugivore species contributed an average
of 32.7% to the distinction, compared with 25.2% for six nectarivore species, 17.6% for
five omnivorous species, 14.9% for seven granivore species and 9.6% for 21 insectivore
species.
3) Presence/absence of bird species: The mistletoebird was the only species that
was significantly more likely to be recorded during months when ripe mistletoe fruit
was present (Fisher’s exact test: P<0.001, significant after Bonferroni correction),
although mistletoebirds were also recorded at three sites (York 1, Paruna, York 2)
during months when no ripe mistletoe fruits were recorded (2, 1 and 5 months for these

57
sites, respectively). The mistletoebird and a parrot, the Australian ringneck, Barnardius
zonarius, were the only bird species directly observed consuming mistletoe fruit during
our surveys (the Australian ringneck feeding on A. preissii; mistletoebirds feeding on
both mistletoe species). Mistletoebirds were frequently observed probing the green
unripe fruits of A. miquelii and pale pink fruits of A. preissii and then either rejecting (A.
miquelii and A. preissii) or occasionally ingesting the unripe fruit (A. preissii only).
Many other species were observed perching within mistletoe clumps, including the
yellow-rumped thornbill, red-capped robin, rufous whistler, grey fantail, varied sittella,
weebill, striated pardalote, western spinebill and silvereye.

58
d) Forrestfield: A. miquelii on C. calophylla
Pro
port
ion p
er
30
cm
bra
nch
0.0
0.2
0.4
0.6
0.8
1.0
f) York 2: A. miquelii on C. calophylla
Feb Mar Apr May June Aug Sept Oct Nov Dec Jan
0.0
0.2
0.4
0.6
0.8
1.0
b) Paruna: A. miquelii on E. accedens
0.0
0.2
0.4
0.6
0.8
1.0
a) York 1: A. preissii on Ac. acuminata
0.0
0.2
0.4
0.6
0.8
1.0Fl. bud
Fl. open
Fl. senescent
Fr. immature
Fr. unripe
Fr. ripe
Bare
c) Araluen: A. preissii on Ac. baileyana
0.0
0.2
0.4
0.6
0.8
1.0
e) Araluen:A. preissii on Ac. podalyrifolia
Feb Mar Apr May June Aug Sept Oct Nov Dec Jan
0.0
0.2
0.4
0.6
0.8
1.0
Figure 3.1 Flowering and fruiting phenology of the mistletoes Amyema preissii (left hand panel)
and Amyema miquelii (right hand panel) from February 2010 to January 2011.
A. preissii: two locations on 3 host species; York 1: Acacia acuminata (a) n=21; Araluen: Ac. baileyana
(c) n=21, Ac. podalyrifolia (e) n=21. A. miquelii: three locations on 2 host species; Paruna: Eucalyptus
accedens (b) n=14; Forrestfield: Corymbia calophylla (d) n=16; York 2: C. calophylla (e) n=18. Values
are expressed as the average proportion of flowers and fruits per 30cm of branch. Lines denote flowers,
solids denote fruits. Course lines denote flowers in ‘bud’, crossed lines denote ‘open’ and fine lines
denote ‘senescent’. Light grey denotes ‘immature’ fruits, grey denote ‘unripe’, black denote ‘ripe’ and
white denote ‘bare’. Arrows indicate peak abundances of flowers (grey) and fruit (black).

59
Open flower absence (0) or presence (1)0 1
Num
ber
of sp
ecie
s
2
4
6
8
10
12
14
16
18
Araluen (n=8, 3)
York 1 (n=8, 3)
Paruna (n=8, 3)
Forrestfield (n=9, 2)
York 2 (n=7, 2)
a) All bird species
Ripe fruit absence (0) or presence (1)
0 1
2
4
6
8
10
12
14
16
18
Araluen (n=7, 4)
York 1 (n=8, 3)
Paruna (n=9, 2)
Forrestfield (n=7, 4)
York 2 (n=6, 3)
Open flower absence (0) or presence (1)0 1
0
1
2
3
4
5
6
Araluen (n=8, 3)
York 1 (n=8, 3)
Paruna (n=8, 3)
Forrestfield (n=9, 2)
York 2 (n=7, 2)
b) Nectarivorous species
c) All bird species
Figure 3.2 The number of bird species recorded at each site in the presence (1) or absence (0) of
open flowers (a) and ripe fruit (c), and the number of nectarivorous bird species recorded at each
site in the presence (1) and absence (0) of open flowers (b).
Numbers of surveys/months contributing to each data point for absent and present, respectively, are
shown in parentheses. Values are expressed as mean ± 1 SD.

60
3.5 DISCUSSION
In this study, we recorded 1) greater bird species richness and 2) altered bird community
structure in the presence of fruiting mistletoe, with 3) a higher chance of sighting
mistletoebirds for months when ripe fruit was present. Can mistletoes in south-west
WA therefore be described as a keystone resource? We discuss the findings of this
study and literature records in relation to the keystone criterion identified in the
introduction.
3.5.1 Reliability of mistletoe resources
The wire leaf mistletoe A. preissii and box mistletoe A. miquelii are important resources
for bird communities in south-west WA due to the wide distribution of these plants as
well as their extended flowering and fruiting times. A. preissii and A. miquelii each had
flowers or fruit available for half the year (A. preissii 6–7 months and A. miquelii 5
months) and across sites timing of flowering and fruiting was offset so that resources
were available over all months of the year. We predicted that there would be a greater
number of bird species present at our study sites when mistletoe flowers or fruit were
available, and that bird community structure would reflect available resources. The
presence of ripe mistletoe fruit was correlated with significantly higher bird species
richness and altered community structure. Importantly, the patterns of fruit presence did
not coincide across our field sites (maximum 100 km apart), and yet the presence of
bird species was associated with site-specific timing of fruit presence. Therefore, while
we are not able to entirely discount ecosystem-wide effects such as bird detectability
(Field et al. 2002), spring and winter migration (see review on partial migration by
Chan 2001) and rainfall driven surges attracting locally nomadic species (as well as

61
promoting mistletoe recruiting and fruiting, Reid 1987; Yan and Reid 1995), the
findings of the present study are consistent with those of previous studies that have
positively linked increased species richness to mistletoe density (Turner 1991; Bennetts
et al. 1996), and manipulative studies comparing the avifauna of two adjacent woodland
remnants where one site had been manually cleared of mistletoe (Watson 2002; Watson
and Herring 2012). Reid (1986) reported that mistletoe is one of the few reliable
sources of fruit in eucalypt forests of south-eastern Australia, and while we only
recorded fruiting phenology over one year, anecdotal observations (K. Napier, pers.
obs.) suggest that mistletoe fruit is predictably available at each site from year to year,
and is therefore extremely reliable. Mistletoes in south-west WA appear to fulfil the
criterion of resource reliability, as they appear to be predictably available every year to
sustain consumers such as the mistletoebird.
3.5.2 Abundance of mistletoe
Crude measures of mistletoe abundance at each site in the present study indicated that
mistletoe is super-abundant in the 2 ha search areas at each, but resource patch density
(sensu Peres 2000) was not measured in the present study. With this caveat, mistletoe
appears to be super-abundant at the sites presented in this study, typical of fragmented
landscapes in south-western WA, and fulfilling the criterion of high abundancy (sensu
Peres 2000). However, the attribute of resource abundance as a criterion for keystone
species status is seen as secondary to the redundancy, reliability and specificity of a
given resource (Peres 2000).

62
3.5.3 Degree of consumer specificity
Resources may range from being extremely generalised, if they are consumed by at least
half of the species in a bird community, to extremely specialised, if they are consumed
by 5% or less of the species (Peres 2000). Mistletoes promote biodiversity by providing
plentiful resources such as nutrient-rich fruit, nectar and leaf litter, as well as sheltered
nesting sites and foliar arthropods (Watson 2001; Cooney et al. 2006; Watson et al.
2011). While the influence of these resources on the structure of the avian community
can be difficult to elucidate (Watson et al. 2011), a number of studies have documented
the link between the presence of aerial mistletoes and greater avian species richness
(e.g. Turner 1991; Bennetts et al. 1996; Watson 2002; Watson and Herring 2012). The
present study clearly indicates temporal effects in bird species presence correlated with
the availability of ripe mistletoe fruit. We had predicted that frugivorous birds would be
more likely to be present when ripe mistletoe fruit was available. Not surprisingly,
presence of the only mistletoe fruit specialist found in WA, the mistletoebird, was
associated with the presence of fruiting mistletoe, and mistletoebirds were observed
feeding on both species of mistletoe fruit. Australian ringneck parrots are opportunistic
feeders (Higgins 1999) and were observed ingesting A. preissii fruits and have been
previously recorded (Forde 1986) feeding on the fruits of A. quandang and Lysiana
exocarpi (both species are found in WA, but are generally restricted to the southern
edge of the Great Victoria Desert and the Nullarbor Plain; Western Australian
Herbarium 1998–; Watson 2011). Several other bird species (nectarivorous,
granivorous and insectivorous) that are known to regularly consume or opportunistically
supplement their diets with mistletoe fruit, including the singing honeyeater, red
wattlebird, silvereye, and yellow-rumped thornbill (see Reid 1986 and references

63
therein), were recorded in the present study, although none were sighted feeding on the
fruit. Although these individual species did not show significant patterns in
presence/absence on their own, the overall pattern was a significant increase in species
richness and changes in the structure of the bird community for months with mistletoe
fruit present compared with months when fruit was absent.
Many nectarivorous, as well as insectivorous and generalist species feed on the
nectar of Australian mistletoes (see Watson 2001). In the present study, although four
nectarivorous honeyeater species were observed feeding on the nectar of both mistletoe
species, we did not find that the presence of these species at our study sites was linked
with the presence of mistletoe flowers. Many insectivorous species were also observed
perching in mistletoe clumps, which may reflect enhanced foraging opportunities
presented by the abundance of insects associated with mistletoes (Turner 1991; Burns et
al. 2011; Watson 2011; Watson et al. 2011).
Findings from the literature therefore suggest support for the criterion of
consumer specificity proposed by Peres (2000), although in the present study, only the
presence of a single species (the mistletoebird) was positively correlated with fruiting
mistletoe; mistletoe, in this study, would therefore be classified as an extremely
specialised resource, and fails to meet this criterion as it is not consumed by a large
proportion of the bird assemblage with which they coexist (i.e. extremely generalised
resource). This criterion requires further investigation.

64
3.5.4 Temporal redundancy of mistletoe
Under the keystone criterion of temporal redundancy density (sensu Peres 2000), a
resource may be considered entirely indispensable if it is available during periods of
overall resource scarcity. Mistletoes in the fragmented landscapes assessed in this study
may then be considered a ‘low redundancy’ resource as they were often the only source
of fleshy fruit and nectar available (with the exception of Araluen, where blackberry
Rubus sp. was present, see Table 3.1), and would therefore be considered entirely
indispensable. Temporal and spatial fluctuations within and among habitats due to both
individual movements and population processes occur in most bird communities
(Malizia 2001). The distribution and abundance of food resources, in particular,
influences the movements of many birds (Levey 1988). The foraging efficiency of
nectarivores and frugivores is also affected by the temporal pattern of flower and fruit
availability: if nectar and fruits are temporally and spatially predictable, animals may
retain this information and visit plants with available resources without random,
undirected searching (Wright 2005). Unlike A. quandang (surveyed at Middleback
Station, South Australia, Reid 1990), which exhibits continuous ripe fruit production
due to an overlap in successive annual fruit crops, the fruiting and flowering within A.
preissii and A. miquelii at each site was fairly distinct. However, across all five sites
examined in the present study, nectar and/or ripe fruit was available for every month of
the year due to staggered flowering and fruiting of these two species, as well as
geographic variation in timing. While the study by Reid (1990) showed that A.
quandang was able to sustain permanent populations of mistletoebirds due to the
continuous fruit availability, it appears that the distinct, but staggered fruiting
phenology of A. preissii and A. miquelii in south-west WA (and South Australia, Yan

65
1993) supports more locally-nomadic movements of mistletoebirds. We note that
mistletoebirds were present at three sites during months when no ripe mistletoe fruits
were recorded, and that this may be attributed to slight differences in fruiting and
flowering phenology that were not captured by our survey methods (e.g. ripe fruit
present on mistletoe plants that were not monitored during surveys), or mistletoebirds
feeding on the unripe fruits of A. preissii. With the caveat that fruiting and flowering
phenology was recorded for only one year, mistletoe appears to be a temporally reliable
source of fruit and nectar (see also Yan 1993). Through the provision of fruit and nectar
resources, these mistletoes act to sustain nomadic populations of mistletoebirds and
assist to sustain permanent populations of nectarivorous birds in the local area
throughout the year.
3.5.5 Resources (functions) not otherwise present
About 18 million ha or 86.5% of the agricultural region of Western Australia has been
cleared, and in the wheatbelt, the percentage is estimated to be 93% (DEP 1997). For
example, 22 districts in the wheatbelt have less than 10% native vegetation cover
(Shepherd et al. 2001), and the few patches of remaining native vegetation exist in
fragmented and isolated patches. Similar environmental disturbances have occurred in
agricultural landscapes across Australia. The quality of food resources in these
fragmented landscapes is an important consideration in terms of sustenance for fauna
species. For example, Norton et al. (1995) predicted that in extremely fragmented
habitats (such as the heavily cleared wheatbelt area), mistletoes would eventually
become extinct due to regional declines in key avian pollinators and dispersers
(Saunders 1993), as has been recorded in the wheatbelt area north of the town of

66
Kellerberrin (Norton et al. 1995). Loranthaceous mistletoes naturally have a patchy
distribution across the landscape, due to both the patterns of bird dispersal (Reid and
Lange 1988; Reid et al. 1995) and narrow microsite tolerances coupled with host
specificity (Knutson 1983; Yan and Reid 1995). The distribution of Australian
mistletoes in fragmented habitats is likely dependent on mistletoe distribution prior to
fragmentation and the impact of fragmentation on the avian pollinators and dispersers
(Norton et al. 1995). Mistletoes have become more abundant in fragmented habitats in
southeast Australia (Reid et al. 1994; Watson 2001), and tend to be either super-
abundant or absent in fragmented areas of southwest WA (Norton et al. 1995). The
presence of these plants (and the resources they provide) in fragmented, otherwise
resource-poor habitats, may therefore counteract the detrimental effects caused by
habitat fragmentation (Kelly et al. 2000; Watson 2002) and may further support their
recognition as important bird resources. Mistletoes therefore play a unique role in the
fragmented landscapes examined in the present study in their provision of vital food
resources to mistletoebirds.
3.5.6 Conclusions
Our findings of increased species richness and changes in the structure of the bird
community demonstrate that A. preissii and A. miquelii may provide important food and
shelter resources for bird species in fragmented south-west Western Australian
woodlands. However, while mistletoes produce highly reliable, low redundancy fruit
resources that play a unique role in fragmented landscapes in south-west WA, we failed
to find evidence that mistletoe nectar and fruit are consumed by a wide range of bird
species. Instead, the only bird species reliant on these food resources is the specialist

67
mistletoebird. The potential for the keystone status of mistletoes in south-west WA
requires further investigation, with comprehensive experimental ‘mistletoe removal’
tests such as those performed by Watson (2002) and Watson and Herring (2012)
recommended.

68
Table 3.2 Bird species recorded in 53 surveys over 5 sites in south-west Western Australia from
February 2010 to January 2011.
Surveys were conducted approximately every 5 weeks from February 2010 to January 2011 (11 surveys
in total for Araluen, York 1, Paruna and Forrestfield; 9 surveys in total for York 2 from April 2010 to
January 2011). The total number of observations was n=511. F refers to frugivore, I (insectivore), N
(nectarivore), G (granivore), and O (omnivore). Nomenclature follows Christidis and Boles (2008).
Number of surveys present
Mistletoe present at site: Amyema preissii Amyema miquelii
Scientific name: Common name: Feeding guild:
Araluen York 1 Paruna Forrestfield York 2
Columbidae Phaps chalcoptera Common bronzewing G 11 2 Ocyphaps lophotes Crested pigeon G 1 Cacatuidae Calyptorhynchus banksii Red-tailed black-cockatoo G 1 3 Calyptorhynchus latirostris Carnaby's black-cockatoo G 3 Eolophus roseicapillus Galah G 7 1 3 5 Psittacidae Barnardius zonarius Australian ringneck G 11 7 9 11 8 Purpureicephalus spurious Red-capped parrot G 3 6 Cuculidae Cacomantis pallidus Palid cuckoo I 1 Cacomantis flabelliformis Fan-tailed cuckoo I 1 Climacteridae Climacteris rufa Rufous treecreeper I 8 Maluridae Malurus splendens Splendid fairy-wren I 8 1 2 Acanthizidae Smicrornis brevirostris Weebill I 11 7 2 Gerygone fusca Western gerygone I 10 5 8 7 Acanthiza chrysorrhoa Yellow-rumped thornbill I 11 9 2 Acanthiza apicalis Inland thornbill I 7 Pardalotidae Pardalotus striatus Striated pardalote I 8 9 7 Meliphagidae Acanthorhynchus superciliosus Western spinebill N 6 6 Lichenostomus virescens Singing honeyeater N 4 9 Anthochaera carunculata Red wattlebird N 11 1 2 Lichmera indistincta Brown honeyeater N 4 4 7 8 8 Phylidonyris novaehollandiae New Holland honeyeater N 11 1 Phylidonyris niger White-cheeked honeyeater N 8 5 Pomatostomidae Pomatostomus superciliosus White-browed babbler O 4 Neosittidae Daphoenositta chrysoptera Varied sittella I 4 Campephagidae Coracina novaehollandiae Black-faced Cuckoo-shrike I 1 2 1 Pachycephalidae Pachycephala rufiventris Rufous whistler I 3 3 2 3 3 Colluricincla harmonica Grey shrike-thrush O 4 6 1 Artamidae Artamus cinereus Black-faced Woodswallow I 2 Cracticus torquatus Grey butcherbird O 2 2 Cracticus tibicen Australian magpie I 10 3 2 Strepera versicolor Grey currawong O 2 Rhipiduridae Rhipidura albiscapa Grey fantail I 5 9 9 3 Rhipidura leucophrys Willie wagtail I 8 9 Corvidae Corvus coronoides Australian raven O 6 4 7 1 4 Monarchidae Grallina cyanoleuca Magpie-lark I 2 Petroicidae Petroica boodang Scarlet robin I 3 2 Petroica goodenovii Red-capped robin I 9 5 Timaliidae Zosterops lateralis Silvereye F 5 1 5 5 Hirundinidae Petrochelidon nigricans Tree martin I 2 5 Nectariniidae

69
4 MISTLETOEBIRDS VARY THEIR DIETARY INTAKE OF
ARTHROPODS DEPENDING ON TIME OF YEAR
Kathryn R. Napier1,2
, Todd J. McWhorter3, Carlos Martínez del Rio
2, Patricia A.
Fleming1
1 School of Veterinary and Life Sciences, Murdoch University, Murdoch, WA 6150,
AUSTRALIA
2 Department of Zoology and Physiology, University of Wyoming, Laramie, WY 82071, USA
3 School of Animal and Veterinary Sciences, University of Adelaide, Roseworthy Campus, SA
5371, AUSTRALIA

70
STATEMENT OF AUTHOR CONTRIBUTION
K.R. Napier: Initiated the research, mist-netted and sampled mistletoebirds and
sampled dietary sources, prepared samples for stable isotopes analysis, entered and
analysed the data, and wrote the manuscript.
T.J. McWhorter: Assisted with the design and initiation of the research and
provided editorial comments to versions of the manuscript from draft to final versions.
C.M. del Rio: Provided training and assistance in the conduction of stable
isotopes analysis, including preparation of samples and statistical analysis, and provided
editorial comments to final versions of the manuscript.
P.A. Fleming: Assisted with the design and initiation of the research, assisted
K.R. Napier with mist-netting and bird sampling, advised and assisted K.R. Napier with
statistical analyses, and provided editorial comments to versions of the manuscript from
draft to final versions.

71
4.1 ABSTRACT
Mistletoebirds (Dicaeum hirundinaceum, family Dicaeidae) inhabit a wide variety of
wooded and forested habitats across Australia. Movements of mistletoebirds are poorly
described, but they appear to be locally nomadic and follow fruiting mistletoe around
the landscape. Mistletoebirds are the primary dispersers of mistletoe seeds and are
mistletoe fruit specialists, with a specialised alimentary system adapted for the ingestion
and quick passage of mistletoe fruits. They are also known to ingest other fruits, nectar,
and invertebrates. We used stable isotopes of carbon (δ13
C) in breath, blood and
feathers and nitrogen (δ15
N) in blood and feathers from mistletoebirds to detect potential
changes in diet through time and to investigate the proportional contribution of
arthropods vs. fruit to their diet. Mistletoebirds were mist-netted and sampled and diet
sources (i.e. arthropods and mistletoe fruit) sampled at three sites in south-west Western
Australia (WA) when ripe mistletoe fruit was available at each site. Sampling occurred
during the austral autumn, winter and summer months. We found that mistletoebirds
appear to change their diet over time, as indicated by the significant differences in δ13
C
values of breath and feathers (tissues that have significantly different turnover rates)
from birds at two of the three sites. We also found that the contribution of arthropods to
the diet of mistletoebirds varies depending on the time of year, or between sites. The
contribution of arthropods to the diet of mistletoebirds ranged from 45% to 67%, and
could be associated with increased protein requirements during breeding and moulting
or differences in availability of food sources between the sites occupied at these times.

72
4.2 INTRODUCTION
Mistletoebirds (Dicaeum hirundinaceum, family Dicaeidae) inhabit a wide variety of
wooded and forested habitats across Australia. Movements of mistletoebirds are poorly
described, and vary with descriptions ranging from largely resident with some local
dispersion to dispersive over large distances, but are usually associated with the
presence of fruiting mistletoe (Higgins et al. 2006). Of 190 recoveries, 98.9% of
banded birds were recovered <10 km from their original banding location (Higgins et
al. 2006). In south-west Western Australia (WA), mistletoebirds are generally only
present in an area when ripe mistletoe fruit is available (Napier et al. 2013). These birds
are the primary dispersers of mistletoe seeds (Higgins et al. 2006) and are mistletoe
specialists, with a specialised alimentary system adapted for the ingestion and quick
passage of mistletoe fruits (Richardson and Wooller 1988), but are also known to ingest
other fruits, nectar, and invertebrates (Reid 1990; Higgins et al. 2006).
Exclusive frugivory in birds is fairly rare, as frugivorous birds may face
nutritional limitations due to the low protein content in most fruits (Morton 1973).
Many frugivorous birds supplement their diet with insects as a means to source
adequate levels of dietary protein, which is essential to meet amino acid requirements
(Klasing 1998). Observational studies of foraging behaviour, food ingested and
analysis of faeces have established that mistletoebirds primarily feed on mistletoe fruit,
but supplement their diet with insects and spiders during the breeding season and when
fruit is scarce (Higgins et al. 2006). These observational studies, however, are unable to
determine the nutritional importance of these different dietary components, as they do
not measure the assimilation of nutrients into the tissues of consumers (Barea and
Herrera 2009). Stable isotopes analysis allows the quantification of the origin of

73
assimilated nutrients (Hobson 1999), which gives an estimation of their dietary
importance (Herrera et al. 2001a; Barea and Herrera 2009), making it a powerful tool in
the reconstruction of diets and estimation of trophic position.
Barea and Herrera (2009) recently used a stable isotope approach to determine
the extent to which mistletoebirds and painted honeyeaters (Grantiella picta, the only
other mistletoe fruit specialist in Australia) in south–east Australia obtain assimilated
nitrogen from mistletoe fruit versus arthropods, and reported that both mistletoebirds
and painted honeyeaters obtain approximately half of their nitrogen from mistletoe fruit
(mean proportional contribution of mistletoe fruit±SE, mistletoebirds: 0.5±0.14; painted
honeyeaters: 0.4±0.04). Herrera et al. (2009) also used a similar stable isotope
approach and estimated that a tropical mistletoe fruit specialist, the yellow-throated
euphonia (Euphonia hirundinacea), obtained most of their assimilated nitrogen from
various fruits (76-100%), which increased with increasing fruit availability at different
times of the year.
We followed a similar, yet slightly more comprehensive approach to Barea and
Herrera (2009) and Herrera et al. (2009) to determine if we could detect changes in the
diets of mistletoebirds over time by sampling birds at three locations in south-west WA
at different times of the year (i.e. when peak abundances of mistletoe fruit were
available at each site), and by also sampling several tissues from each individual bird.
We used stable isotopes of carbon (δ13
C) in breath, blood and feathers and nitrogen
(δ15
N) in blood and feathers from mistletoebirds to firstly detect potential changes in
diet and to provide an estimate of the timeframe of these dietary changes, and secondly,
investigate the proportional contribution of arthropods vs. fruit to the diet of

74
mistletoebirds. We also present δ13
C and δ15
N values for other bird species captured
and sampled along with mistletoebirds, to provide a comparison with birds from other
feeding guilds.
4.3 METHODS
4.3.1 Study sites
The study was conducted at three sites in south-west WA where mistletoe was abundant
(see Table 4.1 for details). Two sites contained Amyema preissii (family Loranthaceae)
parasitising acacia hosts, while the other site contained A. miquelii parasitising a
eucalypt host.

75
Table 4.1 The characteristics of the three field sites in south-west Western Australia.
A, J, M, F, U indicate adult, juvenile, male, female and unknown sex. Fruiting and flowering phenology surveys determined when ripe mistletoe fruit was available, with the average
proportion of ripe fruit available per 30 cm branch presented in parentheses (Napier et al. 2013). Rainfall range is shown for ‘winter’ (May to September inclusive). Weather data
were obtained for 1981-2010 and for the months of sampling (Bureau of Meteorology 2011) from the nearest meteorological station to the study areas: Araluen– Gosnells City
(32°02′S, 115°58′E); York 1 and York 2 –York (31°53′S, 116°46′E).
Site Number of
birds sampled
Mistletoe
species
Host species Site description Sample period
(proportion of fruit
available)
Ripe mistletoe
fruit available
Days with Temp. max. >35°C, min <2°C
Avg. annual rainfall (winter rainfall range)
Total rainfall during sampling; avg. max. Temp
York 1 Private farming
property, York, WA
(31°51′S, 116°44′E)
A (M=2; F=1)
J (U=2)
Amyema
preissii
Acacia
acuminata
A fragmented, semi arid acacia
woodland subject to grazing
April 2010
(0.38)
Apr–June 2010
45, 47 days
401 mm (44–71 mm)
2.6 mm; 27°C
Araluen
Araluen Country Club,
Roleystone, WA
(32°08′S, 116°05′E)
A (M=1; F=1) A. preissii
Ac. baileyana
Ac. podalyriifolia
A heavily watered suburban
garden with both non-native
and native plants present
June 2010
(0.37)
(0.13)
May–Sept 2010 35, 1 day
795 mm (83–165 mm)
60.5 mm; 19.7°C
York 2 Private farming
property, York, WA
(31°50′S, 116°44′E)
A (M=3)
J (M=1)
A. miquelii Corymbia
calophylla
A fragmented, semi-arid
eucalypt woodland, subject to
grazing
Dec 2010–Jan
2011
(0.21–0.38)
Nov 2010–Jan
2011
45, 47 days
401 mm (44–71 mm)
Dec: 20.5 mm; 31.5°C, Jan: 28.8 mm; 35°C

76
4.3.2 Breath, blood and feather samples
11 individual mistletoebirds (8 adults: 6 males, 2 females; and 3 juveniles: 1 male, 2
undetermined sex), were captured by mist-nets placed in front of fruiting mistletoe from
March 2010 to January 2011 – the sampling period was determined by the availability
of ripe mistletoe fruit, and therefore differed at each site (see Table 4.1). One individual
was re-captured >2 weeks after initial sampling at the same site and was re-sampled;
data were therefore averaged for this individual. At least 300 trap hours were conducted
at each site. Three other avian species were also captured at at least two of the sites: the
nectarivorous brown honeyeater (Lichmera indistincta, n=10), the frugivorous silvereye
(Zosterops lateralis, n=26) and insectivorous yellow-rumped thornbill (Acanthiza
chrysorrhoa, n=25). None of these species are sexually dimorphic in their plumage, so
sex was not determined.
We selected three tissues (breath, whole blood and feathers) that can be sampled
both quickly and non-destructively, and that also have significantly different turnover
rates so that we were able to detect potential changes in diet over time. Tissues that are
metabolically active reflect the average diet of an individual, and vary according to the
turnover rate of the tissue. δ13
C obtained from breath samples reflects the diet
consumed several hours since the last meal, with a half-life of approximately 4.4 hours
in small birds (Podlesak et al. 2005). The half-life of carbon and nitrogen in whole
blood in small birds is approximately 4–6 and 11 days respectively (Hobson and
Bairlein 2003; Pearson et al. 2003). Feathers, in contrast, are metabolically inert and
therefore their isotopic composition reflects the diet consumed during feather formation
(Chamberlain et al. 1997).

77
Upon capture, birds were held in a calico bag and carried to a central sampling
station. Birds were then weighed and banded. Breath samples were collected as
described by Carleton et al. (2006). Birds were placed into a 100 ml cylindrical tube,
which was fitted with two one-way stopcock valves. The tube was gently flushed with
~500 ml of CO2-free air over a period of 30 s. The tube was then sealed, and exhaled
CO2 accumulated for two min. A 30ml air sample was extracted and injected into pre-
evacuated gastight vials (Exetainer, Labco Ltd, Buckinghamshire, UK) until a positive
pressure was achieved. Approximately 50 µl of whole blood was collected from the
brachial vein with heparinised capillary tubes by venepuncture. Blood samples were
dried in sample vials in the field initially, and then upon return to the laboratory, at
60°C for 72 h. Samples were then ground to a powder, homogenised and stored at 4°C
until stable isotopes analysis. Body feathers (chest) were obtained from the calico
holding bag or during handling. Feathers were cleaned in 90% EtOH, finely cut, and
stored in paper envelopes until stable isotopes analysis. It was noted that mistletoe
seeds (free from arthropods) were excreted in the calico bags of 7 of 12 mistletoebird
captures. Birds were then released at the site of capture.
All field work and experimental procedures were approved by the Murdoch
University Animal Ethics Committee (approval R2175/08), Western Australian
Department of Environment and Conservation (SW012746 and SF0073330) and the
Australian Bird and Bat Banding Scheme (2699).

78
4.3.3 Diet sources
Collections of >100 ripe mistletoe fruit from at least 20 mistletoe plants were collected
at each study site, placed on ice and stored at -20°C. Samples were prepared for stable
isotopes analysis by manually removing and discarding the exocarp from frozen fruits,
which were then freeze dried to facilitate removal of the fruit pulp from the seed.
Individual fruits, minus the seed, were then dried at 60°C for 72 h and stored at 4°C
until stable isotopes analysis. Stable isotope data were analysed for a sub sample of 27
fruit samples from Araluen, 29 from York 1 and 36 from York 2.
Diurnal arthropods were collected from the ground, mistletoe branches and air
during the same sampling period. After capture, arthropods were initially stored in 70%
EtOH prior to drying at 60°C for 72 h, then ground to a fine powder, homogenised and
stored at 4°C until stable isotopes analysis. Arthropods represented 9 orders, with the
number of morphospecies within each order in brackets. Predatory arthropods (York 1
and York 2 only) included: Arachnida (1), Odonata (2), Diptera (5), and Hymenoptra
(7); while non-predatory arthropods (all sites) included: Coleoptera (4), Diptera (15),
Hemiptera (3), Hymenoptra (9), Lepidoptera (1), Mantodea (1), and Orthoptera (10),
with 11 samples from Araluen, 24 from York 1 and 23 from York 2.
4.3.4 Stable isotopes analysis
Prepared samples were transported to the University of Wyoming Stable Isotope
Facility, USA. Total carbon and nitrogen content (%) and δ13
C and δ15
N compositions
(‰) were determined using a Finnigan Delta Plus XP continuous flow inlet Stable
Isotope Ratio Mass Spectrometer (IRMS) online with a Costech EA 1108 Elemental

79
analyser or CE Elantech 2500 CHN Elemental analyser. δ13
C values are expressed with
respect to V-PDB and δ15
N values with respect to atmospheric molecular nitrogen.
Breath samples were analysed using a Thermo Finnigan Gasbench II online gas
preparation and introduction system online with an IRMS as described above.
4.3.5 Isotope mixing model
Isotope mixing models were carried out on whole blood values only, as while we were
able to apply average discrimination values for whole blood, we were unable to find
similar published discrimination values for breath. Due to uncertainty of the location of
mistletoebirds when they moulted, the contribution of various diet sources to the stable
isotope values of feathers also could not be estimated using mixing models.
Isotope values (δ15
N, δ13
C) were compared by 2-way ANOVA with diet source
(all-arthropods or fruit) and location (site) as factors and Tukey HSD post hoc tests for
unequal sample sizes as required. Significant differences in δ13
C and δ15
N were
detected between sites for fruit but not for arthropods (see results). Each site was
therefore analysed by separate isotope mixing models. We classified arthropods into
two groups based on their trophic level (i.e. predatory and non-predatory arthropods, see
Fig. 4.1a, b, c) for York 1 and York 2. This resulted in three diet sources (fruit,
predatory and non-predatory arthropods) for the two York sites while Araluen only
contained two diet sources (fruit and non-predatory arthropods) as no predatory
arthropods were sampled at this site. Contributions of non-predatory and predatory
arthropods were combined for statistical analysis to allow comparison across the three
sites.

80
As it is critical to consider the concentration of elements when attempting to
reconstruct the diet of animals through the use of isotopic signatures (Phillips and Koch
2002), a Bayesian isotopic mixing model, using both carbon and nitrogen isotopes, was
used to determine the contribution of fruit and arthropods to the diet of mistletoebirds in
the package Stable Isotope Analysis in R (SIAR) (Parnell et al. 2010). SIAR uses a
Markov Chain Monte Carlo simulation to estimate probability distributions of different
dietary sources to a mixture (i.e. whole bird blood) while accounting for the observed
variability in isotopic signatures, dietary isotopic fractionation and elemental
concentration. The solo command in SIAR was used with individual mistletoebirds as
single data points, with the resulting values for each individual compared between sites
via MANOVA. As we did not measure the tissue specific discrimination factors for
mistletoebirds, we followed the approach of Herrera et al (2006); the diet source values
of arthropods and fruit were corrected using the average whole blood-diet carbon and
nitrogen discrimination factors for two insectarivores (that also eat fruit) studied to date
(yellow-rumped warblers and garden warblers) which were fed under controlled
conditions on predominately fruit (δ13
C: 1.41±3.68‰; δ15
N: 1.98±0.39‰) or insect
diets (δ13
C: 2.35±0.21‰; δ15
N: 2.6±0.14‰) (Hobson and Bairlein 2003; Pearson et al.
2003).
4.3.6 General statistics:
Stable isotope values of mistletoebird tissues were analysed via mixed-model (MM)
ANOVAs with individual ID as a random factor and tissue and site as fixed factors,
followed by one-way post hoc ANOVAs with tissue or site as fixed factors. Stable
isotope values of breath and blood were compared in separate MM-ANOVAs with

81
species and site as fixed factors, followed by one-way post hoc ANOVAs with species
or site as fixed factors. Data are presented as mean ± 1 SD throughout, with
significance accepted for α<0.05.

82
0
2
4
6
8
10
12
14
0
2
4
6
8
10
12
14b) Araluen c) York 2
Hym
eno
pte
ra (
1)
Dip
tera
(1
0)
a) York 1
15N
(‰
)
0
2
4
6
8
10
12
14
Dip
tera
pre
d (
1)
Hym
eno
pte
ra p
red (
6)
He
mip
tera
(3
)
Co
leop
tera
(2
)
Hym
eno
pte
ra (
5)
Ma
nto
de
a (
1)
Ort
hop
tera
(4
)
Dip
tera
(1
)
Ort
hop
tera
(6
)
Hym
eno
pte
ra (
3)
Dip
tera
(4
)
Co
leop
tera
(2
)
Lep
idop
tera
(1
)
Hym
eno
pte
ra p
red (
1)
Od
on
ata
pre
d (
2)
Ara
chn
ida p
red (
1)
Dip
tera
pre
d (
4)
Figure 4.1 Median, 25th
and 75th
quartile values of δ15
N (‰) signatures of arthropods sampled at three sites in south-west Western Australia.
a) York 1, b) Araluen and c) York 2 with number of samples in parentheses. Pred indicates predatory arthropods.

83
4.4 RESULTS
δ15
N and δ13
C values and C and N concentrations for mistletoebird tissues and diet
sources are summarised in Table 4.2.
4.4.1 Diet sources
Significant differences in both δ13
C and δ15
N were detected between sites for fruit and
all-arthropods (two-way ANOVA; site: δ13
C: F2,131=7.16, P=0.001; δ15
N: F2,131=10.93,
P<0.001; Table 4.2). Mistletoe fruit were significantly depleted in δ13
C and δ15
N
compared to all-arthropods; mistletoe fruit was significantly depleted in δ13
C compared
to the arthropod samples (all-arthropods) (2-way ANOVA; diet source: F1,131=354.9,
P<0.001; Table 4.2). Fruit δ15
N values averaged from 0.53 to 4.08‰ and were also
significantly depleted compared to the arthropod samples (all-arthropod), which ranged
from 6.62 to 7.41‰ (2-way ANOVA: diet source; F1,131=195.7, P<0.001). Mistletoe
fruit from York 1 was significantly enriched in both δ13
C and δ15
N compared to fruit
from Araluen and York 2 (two-way ANOVA; diet source x site: δ13
C: F2,131=22.16,
p<0.001; δ15
N: F2,131=9.78, P<0.001).
Arthropods (across all orders) contained a significantly higher proportion of
nitrogen (averaging 11.63±1.56%) compared to mistletoe fruit (1.37±1.87%) (2-way
ANOVA; diet source: F1,129=1088, P<0.001). Fruit contained a significantly higher
proportion of carbon (50.83±11.32) compared to arthropods (48.84±3.40%) (2-way
ANOVA; diet source: F1,129=6.78, P=0.010), with a significant interaction term (2-way
ANOVA; diet source x site: F2,129=19.62, P<0.001) demonstrating that fruit from York

84
1 had a significantly higher proportion of carbon compared to all other arthropod and
fruit values (Table 4.2).
4.4.2 Mistletoebird tissues
Feathers were significantly enriched in δ13
C compared to blood, with breath showing
the most depleted δ13
C values (2-way MM-ANOVA; tissue: F2,15=25.80, P<0.001).
Significant site (F2,8=5.03, P=0.038) and ID (F9,12=3.16, P=0.033) effects were
recorded. For York 1, the δ13
C values of the three mistletoebird tissues did not vary
significantly (one-way post hoc ANOVA; tissue: F2,12=0.1.23, P=0.326), but for both
Araluen (F2,3=60.19, P=0.004) and York 2 (F2,9=16.26, P=0.001), feathers were
significantly enriched compared to breath (Fig. 4.2a). There was a significant site
difference in breath δ13
C (one-way post hoc ANOVA; site: F2,8=23.10, P<0.001) and
blood δ13
C (F2,9=11.63, P=0.003) values, with samples from York 1 significantly
enriched in comparison to York 2 and Araluen (Fig. 4.2a). There were no site
differences for feather δ13
C values (F2,7=0.05, P=0.950).
Feathers and blood did not differ in their δ15
N values (2-way MM-ANOVA;
tissue: F1,10=0.24, P=0.636, see Fig. 4.2b). There were no ID (F9,5=4.22, P=0.064) or
site (F2,8=0.33, P=0.730) differences.

85
Table 4.2 Stable isotope values δ13
C and δ15
N (‰) and carbon and nitrogen concentrations (%) for mistletoebird tissues (breath, blood and feathers) and diet sources
(arthropods and mistletoe fruit).
Mistletoe fruit: Amyema preissii (Araluen and York 1) and A. miquelii (York 2). Data are presented as mean ± 1 SD with number of samples in parentheses.
York 1 Araluen York 2
δ13C (‰) δ15N (‰) C% N% δ13C (‰) δ15N (‰) C% N% δ13C (‰) δ15N (‰) C% N%
Breath -23.29±0.65 (5) -27.31±0.40 (2) -25.82±1.00 (4)
Whole blood -22.56±0.84 (6) 6.88±0.56 (6) 44.89±1.73 (6) 13.65±0.57 (6) -24.68±0.71 (2) 4.78±0.35 (2) 45.72±0.73 (2) 13.31±0.88 (2) -24.58±0.54 (4) 5.21±1.67 (4) 45.39±0.35 (4) 13.56±0.32 (4)
Feathers -22.14±5.49 (4) 5.49±1.29 (4) 43.89±0.66 (4) 13.79±0.29 (4) -22.07±7.18 (2) 7.18±0.57 (2) 41.56±4.70 (2) 13.09(2)1.52 (2) -22.39±6.18 (4) 6.18±1.10 (4) 43.84±1.18 (4) 13.57±0.31 (4)
All-arthropods
-Non-predatory
-Predatory
-24.01±1.85 (24)
-23.81±2.13 (16)
-24.42±1.09 (8)
7.05±2.60 (24)
5.80±2.14 (16)
9.54±1.31 (8)
48.46±3.61 (24)
48.73±4.35 (16)
48.94±1.34 (8)
11.62±1.71 (24)
11.88±1.90 (16)
11.09±1.17 (8)
-22.95±0.40 (11)
7.41±0.19 (11)
48.77±1.02 (11)
11.28±0.33 (11)
-23.33±1.32 (23)
-23.64±1.30 (16)
-22.64±1.21 (7)
6.62±2.55 (23)
5.24±1.51 (16)
9.75±1.31 (7)
48.51±2.03 (23)
49.45±2.37 (16)
49.62±1.04 (7)
11.81±1.77 (23)
11.25±1.85 (16)
13.10±0.44 (7)
Mistletoe fruit -26.44±1.47 (29) 4.08±1.83 (29) 49.70±11.79 (29) 1.12±0.27 (29) -28.11±1.02 (27) 0.82±1.81 (27) 47.20±7.13 (27) 2.17±3.00 (27) -28.62±1.03 (36) 0.53±1.30 (36) 54.47±12.63 (36) 0.89±0.25 (36)

86
13C
(‰
)
-30
-28
-26
-24
-22
-20
-18
15N
(‰
)
0
1
2
3
4
5
6
7
8
Araluen
York 1
York 2
fruit (A)
fruit (Y 2)
fruit (Y 1)
all-arthropods (A)
all-arthropods (Y 1)
all-arthropods (Y 2)
breath blood feather
a*
a*a*a*
c**
b**
a*
b**
a*
blood feather
a) b)
all-arthropods (A)
all-arthropods (Y 2)
all-arthropods (Y 1)
fruit (A)
fruit (Y 2)
fruit (Y 1)b**
a*
a*
a*
a*
a*
Figure 4.2 δ13
C (a) and δ15
N (b) values (‰) of breath, whole blood and feathers for mistletoebirds at three sites in south-west Western Australia.
Sites are represented by: York 1 (Y 1, white triangles), Araluen (A, black circles) and York 2 (Y 2, black squares). Data are presented as mean ± 1 SD. Horizontal lines
represent δ13
C and δ15
N values for diet sources: mistletoe fruit (solid lines) and arthropods (dashed lines). Diet sources have not been corrected by diet-tissue discrimination
factors. Letters indicate significant differences between sites while asterisks denote differences between tissues obtained from one-way ANOVAs with Tukey HSD post hoc
tests for unequal sample sizes.

87
4.4.3 Isotope mixing model
δ13
C and δ15
N values for mistletoebird whole blood and diet sources are presented in
Fig. 4.3a. There were significant differences in the dietary contribution of arthropods
and fruit between sites (MANOVA: site; F2,8=64.37, P<0.001, Fig. 4.4a). Post hoc tests
revealed that fruit made a significantly greater dietary contribution to mistletoebird diets
at Araluen compared with the other two sites (Araluen: 0.55±0.03; York 1: 0.33±0.01;
York 2: 0.38±0.04) while arthropods made a significantly greater dietary contribution at
York 1 and York 2 (Araluen: 0.45±0.03; York 1: 0.67±0.01; York 2: 0.62±0.04).
At the York sites, we collected non-predatory as well as predatory arthropods,
with their respective dietary contributions as follows; York 1: 0.51±0.03 and 0.16±0.04
and York 2: 0.56±0.01 and 0.07±0.04 Variation amongst individuals (i.e. adults vs
juveniles, males vs females) could not be assessed via statistical methods due to small
sample sizes, but are presented graphically (Fig. 4.4b).
4.4.4 Comparison with other bird species
δ13
C values for breath and δ13
C and δ15
N values for whole blood and feathers from
brown honeyeaters, silvereyes and yellow-rumped thornbills are presented in Table 4.3,
with average δ13
C and δ15
N values for whole blood and diet sources presented in Fig.
4.3b. Breath δ13
C values were depleted at Araluen compared to the two semi-arid sites
at York (2-way MM-ANOVA; site: F2,52=12.58, P<0.001). A significant species effect
was also recorded (F3,52=5.18, P=0.003), with the insectivorous yellow-rumped
thornbill with the most enriched breath δ13
C values while the nectarivorous brown
honeyeater had the most depleted. At Araluen, the breath δ13
C values of the yellow-

88
rumped thornbill were significantly enriched compared to the frugivorous silvereye
(one-way post hoc ANOVA: F3,26=4.50, P<0.011), but species differences were not
evident at either York site (York 1: F3,17=1.79, P=0.190; York 2: F2,9=2.79, P=0.114).
Whole blood δ13
C values were depleted at Araluen, in comparison to both York
1 and 2, with the most enriched δ13
C values from York 1 (2-way MM-ANOVA; site:
F3,53=11.62, P<0.001). A significant effect of species (F3,53=11.62, P<0.001) and an
interaction between site and species were also recorded (F4,53=12.39, P<0.001). At
Araluen and York 2, the whole blood δ13
C values of the yellow-rumped thornbill were
significantly enriched in comparison to silvereyes (Araluen only) and mistletoebirds
(one-way post hoc ANOVA; Araluen: F3,23=18.16, P<0.001; York 2: F1,10=43.96,
P<0.001), while species differences were not evident at York 1 (F3,20=2.47, P=0.091).
Whole blood δ15
N values were more depleted at Araluen in comparison to the
two York sites (2-way MM-ANOVA; site: F2,53=9.75, P<0.001). A significant effect of
species was also evident (F3,53=12.03, P<0.001), with yellow-rumped thornbills having
the most enriched δ15
N values and brown honeyeaters the most depleted. A significant
interaction between site and species were also recorded (F4,53=4.50, P=0.003). At
Araluen and York 2, the δ15
N values of yellow-rumped thornbills were significantly
enriched compared to silvereyes (Araluen only) and mistletoebirds (one-way post hoc
ANOVA; Araluen: F3,23=18.16, P<0.001; York 2: F1,10=42.56, P<0.001). Species
differences were not evident at York 1 (F3,20=3.87, P=0.025) when post hoc analyses
were applied.

89
13C (‰)
-30 -28 -26 -24 -22 -20 -18
15N
(‰
)
0
2
4
6
8
10
12
14
Blood- York 2
Blood- Araluen
Blood- York 1
Fruit- York 2
Fruit- Araluen
Fruit- York 1
All-arthropods- York 1
All-arthropods- Araluen
All-arthropods- York 2
Feathers- York 1
Feathers- Araluen
Feathers- York 2
13C (‰)
-30 -28 -26 -24 -22 -20 -18
15N
(‰
)
0
2
4
6
8
10
12
14
Brown honeyeater- York 1
Brown honeyeater- Araluen
Mistletoebird- York 2
Mistletoebird- Araluen
Mistletoebird- York 1
Silvereye- York 1
Silvereye- Araluen
Yellow-rumped thornbill- York 2
Yellow-rumped thornbil- Araluen
Yellow-rumped thornbill- York 1
Fruit- York 1
Fruit- Araluen
Fruit- York 2
All-arthropods- York 2
All-arthropods- Araluen
All-arthropods- York 1
a)
b)
arthropod signature
Figure 4.3 δ13
C and δ15
N values (‰) of a) whole blood and feathers from individual mistletoebirds
and b) average whole blood values for mistletoebirds, silvereyes, brown honeyeaters and yellow-
rumped thornbills at three sites in south-west Western Australia.
a) Blood samples are represented by circles and feathers by diamonds at the sites York 1 (black), Araluen
(grey) and York 2 (white). δ13
C and δ15
N values for diet sources are represented by inverted triangles
(mistletoe fruit) and squares (arthropods). b) Mistletoebirds are represented by inverted triangles,
silvereyes by squares, brown honeyeaters by circles, and yellow-rumped thornbills by diamonds at the
sites York 1 (black), Araluen (grey) and York 2 (white). δ13
C and δ15
N values for diet sources are
represented by triangles (mistletoe fruit) and hexagons (arthropods). Diet sources have not been corrected
by tissue-diet discrimination factors. Data are presented as mean ± 1 SD.

90
York 1
Pro
po
rtio
na
l co
ntr
ibu
tio
n o
f fr
uit
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Araluen York 2
A,F A,M A,M J,U J,U A,F A,M J,MA,M A,MA,M
b)
Jan Feb Mar April May June Jul Aug Sept Oct Nov Dec
Pro
po
rtio
na
l co
ntr
ibu
tio
n
0.0
0.2
0.4
0.6
0.8
Mistletoe fruit
Non-predatory arthropods
aa
**
*
b
moult
(eggs)
York 1 (n=5) Araluen (n=2) York 2 (n=4)
moult
breeding
Predatory arthropods
**
a)
Figure 4.4 Proportional contribution of fruit and insects to the whole blood of mistletoebirds a)
and proportional contribution of mistletoe fruit to individual mistletoe birds b) obtained from a
dual-isotope (δ13
C, δ15
N) Bayesian isotope mixing model (SIAR).
n= number of mistletoebirds sampled for whole blood at each site. Letters and asterisks indicate
significant differences between sites for all-insects and fruit respectively, obtained from MANOVA with
Tukey HSD post hoc tests for unequal sample sizes. Breeding and moulting information for Western
Australia obtained from Higgins et al. (2006). A indicates adult, J juvenile, F female, M male, U
unknown sex. Arthropod contribution = 1- mistletoe fruit contribution.

91
Table 4.3: Stable isotope values for breath and whole blood for four avian species. Data are presented as mean ± 1 SD with number of samples in parentheses.
York 1 Araluen York 2 δ13C (‰) δ15N (‰) δ13C (‰) δ15N (‰) δ13C (‰) δ15N (‰)
Brown honeyeater Breath -24.27±1.46 (2) -27.50±1.00 (7) -25.82 (1)
Whole blood -22.25± 0.91(2) 3.92±1.97 (2) -23.84±0.33 (6) 6.01±1.13 (6)
Feathers -21.76±1.04 (2) 6.20±0.36 (2) -21.68±0.57 (7) 6.48±0.61 (7) -20.64 (1) 7.71 (1)
Silvereye Breath -24.22± 0.99 (9) -27.36±1.24 (10)
Whole blood -22.13±0.43 (11) 8.11±2.42 (11) -24.61±0.15 (11) 5.86±0.83 (11)
Feathers -21.48±0.51 (9) 10.03±2.60 (9) -22.69±1.52 (15) 5.37±0.49 (15)
Yellow-rumped thornbill Breath -24.53± 0.76 (5) -24.52±3.04 (11) -24.26±1.18 (7)
Whole blood -22.92±0.23 (5) 8.56±0.41 (5) -23.19±0.67 (8) 7.11±0.16 (8) -22.97-±0.32 (8) 9.06±0.37 (8)
Feathers -21.44± 0.60 5) 9.57±0.58 (5) -18.28±1.52 (12) 8.47±0.49 (12) -22.44±1.72 (8) 10.02±1.15 (8)

92
4.5 DISCUSSION
We found that mistletoebirds appear to change their diet over time, as indicated by the
significant differences in δ13
C values of breath and feathers. We also found that the
contribution of arthropods to the diet of mistletoebirds varies depending on the time of
year or between sites, which could be associated with increased protein requirements
during breeding and moulting or differences in availability of food sources between the
sites occupied at these times. For example, our sampling of arthropods at Araluen
during winter was likely to be hampered by low ambient temperatures (less than 20°C
on average), which may also influence arthropod availability for the birds.
4.5.1 Detecting diet switching of mistletoebirds
We measured carbon and nitrogen stable isotopes in three tissues with different isotopic
turn-over rates to determine if we could detect changes in the diet of mistletoebirds over
time. For small birds, diet-tissue discrimination values for carbon and nitrogen isotopes
are generally enriched by 1-3.5‰ relative to diet, with tissue isotope values influenced
by the signature of the diet fed to the animal, the signature of macronutrients within the
diet, as well as the nutrient composition of the tissue (Hobson and Bairlein 2003;
Pearson et al. 2003; Podlesak et al. 2005). Following the approach of Podlesak et al.
(2005), we interpret the significantly different δ13
C values of the various tissues (>3‰)
as indication that mistletoebirds had switched between diets that were isotopically
distinct. For example, the significant differences between feathers, which provide an
estimate of the past diet of mistletoebirds, and breath, which has a very short half-life of

93
~4.5 hours, indicates a recent change in diet at two out of the three sites (Podlesak et al.
2005).
The δ13
C and δ15
N values of mistletoebirds captured at York 1 (sampled in
autumn) were <1.5‰ different across tissues, suggesting that these birds had been
ingesting food with relatively consistent C and N signatures for an extended period of
time. The δ15
N values of mistletoebirds sampled at York 2 (sampled in summer) and
Araluen (sampled in winter) were <1.5‰ different across tissues, suggesting that these
birds had also been ingesting food with relatively consistent N signatures for an
extended period of time, however C signatures varied.
The δ13
C values of breath, blood and feathers from mistletoebirds captured at
York 2 and Araluen were >2‰ different, with the greatest differences evident between
breath and feathers (>3‰), suggesting that these birds had recently changed their diet to
a carbon signature that closely resembles mistletoe fruit (Fig. 4.2a). In birds and bats,
δ13
C values from breath generally reflect immediate oxidation of ingested food, rather
than endogenous energy stores (Welch et al. 2006; Voigt et al. 2008). We mist-netted
birds in front of fruiting mistletoe plants, and noted that 58% of birds excreted mistletoe
seeds while held in calico bags, with no visible evidence of arthropods present in
excreta. This, combined with the δ13
C values obtained from breath samples, strongly
indicates that mistletoebirds were ingesting a high proportion of mistletoe fruit in the
hours preceding capture. Conversely, the δ13
C values of mistletoebird feathers strongly
resembles that of arthropods (Fig. 4.2a), indicating that these birds were supplementing
their fruit diet with arthropods during their moult, a period when dietary protein
requirements are higher (Klasing 1998) .

94
4.5.2 Contribution of fruit vs arthropods
We were able to estimate the proportional contribution of mistletoe fruit and arthropods
to the diet of mistletoebirds by sampling whole bird blood, arthropods and mistletoe
fruit from the area where birds were mist-netted. Bird and plant surveys indicated that
mistletoebirds were sighted in the study sites during months when ripe mistletoe fruit
was available (Napier et al. 2013), so we were confident that we managed to obtain an
accurate representation of the dietary sources available to mistletoebirds at each
location.
Mistletoebirds varied the contribution of arthropods to their diet, ranging from
45% to 67% (Fig. 4.4a), depending on the time of year. Mistletoebirds are known to
ingest insects, primarily during the breeding season, or when fruit is scare (Higgins et
al. 2006). As mistletoe fruit was abundant at each of the sites during the sampling
period (with proportions of ripe mistletoe fruit available ranging from 0.13 to 0.38 per
30 cm of branch, Table 4.1), it is not likely that a lack of fruit availability was the cause
of this variation. Frugivorous birds are also known to supplement their fruit diet with
insects during moulting periods (Klasing 1998). As mistletoebirds moult from summer
to autumn (Higgins et al. 2006), we may expect arthropods to contribute a higher
proportion to the diet of mistletoebirds at these times. Indeed, we see that
mistletoebirds obtained significantly greater contributions of arthropods to their diet at
York 1 and York 2, which were sampled in April (autumn) and December/January
(summer) and presumably during their moulting period which is associated with an
increased demand for dietary nitrogen. Araluen was sampled during June (winter),
presumably post moulting. The breeding period of mistletoebirds in WA is poorly

95
described, with unspecified breeding records from the months of July to January (with
eggs recorded in September and October) and from March to May. We did mist-net
juvenile mistletoebirds at two sites, York 1 (2 out of 5 birds) and York 2 (1 out of 4
birds), with juveniles of undetermined sex sampled at York 1 and a single first
immature male (as defined by Higgins et al. 2006) mist-netted at York 2 (Fig. 4.4b).
Therefore, it is likely that a combination of breeding and moulting contributed to the
increase in arthropods in the diet of mistletoebirds at York 1 and York 2, compared to
Araluen.
We did not see the level of intrapopulation variation described by Barea and
Herrera (2009), rather, differences were evident between populations of mistletoebirds
(Figs. 4a, b). We could not test for sex- (or age-) based dietary differences although
none were evident in our data, as also noted by Barea and Herrera (2009).
4.5.3 Comparison with other bird species
As we did not determine the tissue specific discrimination factors for mistletoebirds for
use in the isotope mixing model, these estimates were obtained through the application
of average discrimination values from two studies that fed two species of warblers fruit
and insect based diets (Hobson and Bairlein 2003; Pearson et al. 2003), following the
approach of Herrera et al. (2005; 2006; 2009) and Barea and Herrera (2009). Yellow-
rumped and garden warblers belong to two diverse families of insectivorous birds
(Parulidae and Sylvidae respectively), while mistletoebirds are specialised frugivores
belonging to the family Dicaeidae. To date, there have been no studies on tissue to diet
discrimination factors in specialist frugivorous birds (see Hahn et al. 2012). As

96
discrimination factors have been shown to vary between taxonomic groupings (Caut et
al. 2009), the extrapolation of these average discrimination factors is problematic, and
our conclusions based upon the diet mixing model must be treated with some caution.
We therefore also compared the average values of δ13
C and δ15
N with birds
from other taxonomic and feeding guilds, captured and sampled at the same time as
mistletoebirds. As insectivores feed at higher trophic levels than frugivores, we expect
that the tissues of yellow-rumped thornbills would have higher δ15
N values compared to
mistletoebirds (Gagnon and Hobson 2009). The average increase of 3.4 in δ15
N for
each increase in trophic level (Kelly 2000; Post 2002) allows the detection of diet shifts,
such as insectivory vs. frugivory (Herrera et al. 2001b; Herrera et al. 2003).
For birds sampled at Araluen and York 2, yellow-rumped thornbills had more
enriched δ15
N and δ13
C values for whole blood compared to mistletoebirds also sampled
at these sites (Fig 4.3b). The δ15
N and δ13
C values for mistletoebirds and yellow-
rumped thornbills at York 1 were similar, lending support to the findings of the isotope
mixing model that mistletoebirds were supplementing their diets with a higher
proportion of arthropods at this site.
4.5.4 Arthropods and mistletoe fruit are both important sources of dietary
nitrogen
Mistletoebirds did not depend on mistletoe fruit as a sole food source; they incorporate
arthropods in their diet to varying degrees. However, despite its low nitrogen content
(Table 4.2), mistletoe fruit makes a significant contribution to the assimilated nitrogen
of mistletoebirds. The high reliance on mistletoe fruit as a source of protein by these

97
specialised frugivores is assisted by their high intake rates of mistletoe fruit, as well as
specialised intestinal morphology (Richardson and Wooller 1988) that results in
extremely rapid transit times of mistletoe fruit compared to other non-specialised
frugivores (Murphy et al. 1993; Barea 2008; Napier et al. 2013).
Stable isotope techniques are an extremely useful tool in the reconstruction of
animal diets. This study is the first to investigate the changes in diets of mistletoebirds
over time, and demonstrates the increased contribution of arthropods to the diet of
mistletoebirds during breeding or moulting.

98
5 SUGAR PREFERENCES OF AVIAN NECTARIVORES ARE
CORRELATED WITH INTESTINAL SUCRASE ACTIVITY
Kathryn R. Napier1, Todd J. McWhorter
1,2, Susan W. Nicolson
3 and Patricia A.
Fleming1
1 School of Veterinary and Life Sciences, Murdoch University, Murdoch, WA 6150,
AUSTRALIA
2 School of Animal and Veterinary Sciences, University of Adelaide, Roseworthy Campus, SA
5371, AUSTRALIA
3 Department of Zoology and Entomology, University of Pretoria, Pretoria 0002, SOUTH
AFRICA
This chapter has been published in the journal Physiological and Biochemical Zoology
(86(5):499–514, DOI: 10.1086/672013).

99
STATEMENT OF AUTHOR CONTRIBUTION
K.R. Napier: Initiated the compilation of the research, mist-netted and kept
birds in captivity, performed all laboratory experiments on mistletoebirds, silvereyes
and singing honeyeaters and collected, entered and analysed the data. Also performed
sugar preference experiments on the New Holland honeyeaters detailed in Fleming et al.
(2008). Collected intestinal tissue and performed enzyme assays on five species (as
detailed in Table 5.2, years 2010–2011), and wrote the manuscript.
T.J. McWhorter: Collected intestinal tissue and performed enzyme assays on four
species (as detailed in Table 5.2, years 2006–2007) and provided access to unpublished
data as cited throughout. Assisted with the design and initiation of the research, advised
and assisted K.R. Napier with enzyme assay and apparent assimilation efficiency
procedures and statistical analyses, and provided editorial comments to versions of the
manuscript from draft to publication.
S.W. Nicolson: Provided data on nectar composition as detailed in Figure 5.6 and
Table 5.4 and provided editorial comments to versions of the manuscript from draft to
publication.
P.A. Fleming: Assisted with the design and initiation of the research, assisted
K.R. Napier with mist-netting, advised and assisted K.R. Napier with statistical
analyses, and provided editorial comments to versions of the manuscript from draft to
publication.

100
5.1 ABSTRACT
Nectar-feeding birds generally demonstrate preference for hexose solutions at low sugar
concentrations, switching to sucrose/no preference at higher concentrations. Species
vary in the concentration at which the switch from hexose preference occurs; this could
reflect physiological constraints that would also influence nectar selection when
foraging. We recorded concentration-dependent sugar type preferences in three
opportunistic/generalist Australian nectarivorous species: the mistletoebird (Dicaeum
hirundinaceum), silvereye (Zosterops lateralis) and singing honeyeater (Lichenostomus
virescens). All three preferred hexoses up to sugar concentrations of 0.25 mol·L-1
and
switched to sucrose/no preference for higher concentrations. Using these and literature
records, we investigated physiological mechanisms that may explain the concentration-
dependence of sugar type preferences and compared diet preference data with foraging
records. We measured sucrase activity in silvereyes and singing honeyeaters as well as
three specialised nectarivorous species; red wattlebirds (Anthochaera carunculata),
New Holland honeyeaters (Phylidonyris novaehollandiae) and rainbow lorikeets
(Trichoglossus haematodus) for comparison with published concentration-dependent
sugar preference data. Sucrase activity varied between these species (P=0.003). The
minimum diet concentration at which birds show no sugar preference was significantly
correlated with sucrase activity for the eleven species analysed (P=0.005). Birds with
the lowest sucrase activity showed hexose preference at higher diet concentrations and
birds with the greatest sucrase activity either showed no hexose preference or hexose
preference on only the most dilute diets. Foraging data compiled from the literature
also support the laboratory analyses, e.g. rainbow lorikeets (preference for hexose over
a wide range of diet concentrations, low sucrase activity) also feed primarily on hexose

101
nectars in the wild. Intestinal sucrase activity is likely to contribute to diet selectivity in
nectarivorous bird species.

102
5.2 INTRODUCTION
Nectar and fruit are an important carbohydrate-rich food source for many bird species.
The disaccharide sucrose and its monosaccharide components glucose and fructose (i.e.
hexoses, which are similar in chemical structure and in energy content per unit mass)
are among the most common carbohydrates in nectar and fruit (Levey and Martinez Del
Rio 2001). The composition and concentration of sugars in nectar and fruit pulp varies
widely amongst plant species (Whiting 1970; Pyke and Waser 1981; Baker and Baker
1982b; Baker and Baker 1983; Baker et al. 1998; Nicolson and Van Wyk 1998;
Nicolson 2002; Wilson and Downs 2012). Fruit pulp tends to be hexose-dominant, with
sucrose content averaging only 8% of total sugars in fruits consumed by passerines
(Martínez del Rio et al. 1992; Baker et al. 1998). Nectar may be sucrose-dominant,
hexose-dominant, or contain a mixture of both sucrose and hexoses (Nicolson and
Fleming 2003b; Johnson and Nicolson 2008). Many nectarivorous and frugivorous bird
species exhibit distinct preferences for these sugars (see review by Lotz and Schondube
2006), although past studies were commonly conducted using a single sugar
concentration and so the role that energy density may play in sugar selection is not
clear. These past studies also used a wide variety of experimental methodologies,
which can make comparing results among different studies difficult (Brown et al.
2008).
The potential physiological mechanisms underlying the sugar preferences of
birds and the extent to which the sugar composition of natural nectars reflects selection
by birds have long been debated. Dramatic differences in the composition of sugars in
nectar were first reported by Baker and Baker (1982b; 1983). While these differences

103
in plant nectar sugar composition were first thought to reflect selective pressures from
their hummingbird (sucrose-dominant nectars) or passerine (hexose-dominant nectars)
pollinators (Martínez del Rio 1990; Martínez del Rio et al. 1992), subsequent studies on
the digestive enzymes of various avian lineages have shown that both hummingbirds
and nectar-specialist passerines are capable of efficient digestion and assimilation of
sucrose (see review by Lotz and Schondube 2006). The intestinal enzyme sucrase-
isomaltase is responsible for the hydrolysis of sucrose into its monosaccharide hexose
components. Most specialist and occasional nectarivores and frugivores are able to
efficiently assimilate both sucrose and hexoses (Lotz and Schondube 2006; Fleming et
al. 2008; Napier et al. 2008a), with the exception of frugivores in the Sturnidae-
Muscicapoidea lineage that lack sucrase (Martínez del Rio and Stevens 1989; Brugger
et al. 1993; Sabat and Gonzalez 2003; Gatica et al. 2006; Brown et al. 2012). Some
occasional nectarivores, however, exhibit lower apparent assimilation efficiencies for
both sucrose and hexoses (Brown et al. 2010b; Brown et al. 2010a) and some
occasionally nectarivorous and frugivorous passerines exhibit sucrose assimilation
efficiency that is significantly lower than that for hexoses (Lane 1991; Odendaal et al.
2010). These patterns are consistent with findings presented by Johnson and Nicolson
(2008), who demonstrated that nectars of plants pollinated by specialist nectarivorous
passerines are strongly convergent with those of plants pollinated by hummingbirds.
Specifically, plants pollinated by specialist avian nectarivores tend to have small
volumes of concentrated, sucrose-dominant nectars, while those pollinated by
generalists tend to have large volumes of dilute, hexose-dominant nectars.
One important finding of recent studies is that sugar type preference varies with
sugar concentration. Nectarivorous birds tested using a range of concentrations of

104
‘equicaloric’ (Fleming et al. 2004) sucrose or hexose diets generally demonstrate
preference for hexose solutions at low sugar concentrations (i.e. energy densities), with
a switch to sucrose or no preference at higher concentrations. This has been
demonstrated in specialist nectarivores including sunbirds, hummingbirds, honeyeaters
and lorikeets (Schondube and Martinez del Rio 2003; Fleming et al. 2004; Lotz and
Schondube 2006; Fleming et al. 2008; Brown et al. 2010c), and occasional nectarivores
such as the speckled mousebird (Colius striatus) and village weaver (Ploceus
cucullatus) (Brown et al. 2010a; Odendaal et al. 2010, see Table 5.1). Although these
species demonstrate a similar pattern in sugar preferences, they differ in the
concentration at which the switch from hexose preference to no preference occurs.
Most specialist nectarivores prefer hexoses at extremely dilute diets only, e.g. the red
wattlebird (Anthochaera carunculata), New Holland honeyeater (Phylidonyris
novaehollandiae), white-bellied sunbird (Cinnyris talatala), Malachite sunbird
(Nectarina famosa), magnificent hummingbird (Eugenes fulgens) and cinnamon-bellied
flowerpiercer (Diglossa baritula) (Table 5.1). Some opportunistic nectar feeders (the
speckled mousebird and village weaver) prefer hexoses up to slightly higher
concentrations than these specialised nectarivores, yet the dark-capped bulbul
(Pycnonotus tricolor, a nectar generalist), and rainbow lorikeet (Trichoglossus
haematodus, a nectar specialist) prefer hexoses at much higher sugar concentrations
(Table 5.1). Brown and colleagues suggested that these findings help to explain the
dichotomy reported by Johnson and Nicolson (2008); however, aside from the work by
Brown et al. (2010a; 2010b) and Odendaal et al. (2010), little comparative data on sugar
preferences in generalist nectar feeders has been available to date. Compared with

105
nectarivores, we know far less about the concentration-dependence of sugar preferences
of opportunistic or generalist avian frugivores.
Compensatory feeding, where birds increase volumetric intake rate as food
energy density decreases, allows birds to deal with variations in nectar concentration
(Martínez del Rio et al. 2001; Nicolson and Fleming 2003a). Lotz and Schondube
(2006) and Fleming et al. (2008) have hypothesised that the concentration-dependence
of sugar preferences in nectarivorous birds may be attributed to varying levels of
sucrase activity and the need for constant energy assimilation (i.e. compensatory
feeding). Birds that exhibit a lower capacity to hydrolyse sucrose are more likely to
show preference for hexoses over sucrose solutions on dilute diets in this scenario,
because digesta transit rates will be faster and substrate concentration for the sucrase
enzyme will be lower, limiting the hydrolysis rate (McWhorter and Martinez del Rio
2000). In this study, we have tested this prediction with new and available published
data. We investigated sugar preferences and apparent assimilation efficiency in three
opportunistic/generalist Australian nectarivorous species: the mistletoebird (Dicaeum
hirundinaceum), silvereye (Zosterops lateralis) and singing honeyeater (Lichenostomus
virescens). We also analysed the activity of the intestinal enzymes sucrase-isomaltase
(EC 3.2.1.48, hereafter ‘sucrase’) and maltase-glucoamylase (EC 3.2.1.20, hereafter
‘maltase’) in silvereyes and singing honeyeaters, as well as three specialised
nectarivorous species (New Holland honeyeaters, rainbow lorikeets and red wattlebirds)
for comparison with published sugar preference data for these species (Fleming et al.
2008). Finally, we compiled foraging data for these species and Australian nectar sugar
compositions, where available, from the literature. We predicted that:

106
1) specialised nectarivorous species would exhibit greater apparent assimilation
efficiencies for both hexoses and sucrose than generalist nectarivores;
2) the degree of preference for hexose over sucrose solutions would be
correlated with variation in the capacity to hydrolyse sucrose; and
3) specialist nectarivores should preferentially forage on sucrose-rich nectars
compared with generalist species.

107
Table 5.1 Summary of sugar type preferences, apparent assimilation efficiency (AE*) and digestive capacity.
- indicates not measured or not available. Diet: Specialist nectarivore (SN, bold), generalist nectarivore (GN), frugivore (Fr). References: 1this study;
2Fleming et al. (2008);
3Napier et al. (2008a);
4Fleming et al. (2004);
5Schondube and Martínez del Rio (2003);
6Schondube and Martínez del Rio (2004);
7McWhorter et al. (unpublished),
8Köhler et al.
(2010), 9Brown et al. (2010a),
10Brown et al. (2010b),
11Brown et al. (2010c),
12Odendaal et al. (2010),
13Wilson and Downs (2011),
14Downs (1997)
15Brown et al. (2012)
16Martínez del Rio (1990)
17Martínez del Rio (1990),
18McWhorter and Martínez del Rio (2000),
19Bizaare et al.(2012). Sugar preferences: dark grey: hexose (H) or glucose (G),
light grey: sucrose (S), ns: no significant preference. Total activity: nd: not detectable. Vmax: *we were advised by the authors that this published value is incorrect, and should be
9.16±1.1. This correct value was used in the analyses detailed in Figure 5.4. Km and pH optima: Kinetic parameters obtained using at least n=1 tissue homogenate (proximal
intestinal section): † two data sets for birds caught in 2010–2011 (n=1) or 2006–2007 (n=1).
Sugar type preferences
Apparent assimilation efficiency (AE*)
Digestive capacity (sucrase)
Body
mass (g)
Diet Concentration (Sucrose
Equivalents, mol·L-1)
Gut nominal surface area
(cm2) Total activity (µmol·min-1)
Vmax (μmol∙min-1) Km (mmol)
pH optima Bird species Diet Ref 0.075 0.1 0.25 0.5 0.75 1 2
Sucrose Glucose Fructose
n
Rainbow lorikeet (Trichoglossus haematodus) SN 137±14 1,2,3 H H H H H ns S
>98 99.7±0.1 -
7 31.6±7.1 25.8±12.1 42.8±21.7 21.7, 16.7† 5, 5† New Holland honeyeater (Phylidonyris novaehollandiae) SN 20.5±3.4 1,2 H H ns ns ns ns ns
>99 - -
9 6.0±1.7 12.6±7.9 25.3±15.3 39.0, 25.0† 6, 6†
Red wattlebird (Anthochaera carunculata) SN 105±3 1,2 H ns ns ns ns S S
>99 99.8±0.1 -
8 15.3±4.2 41.7±23.6 77.9±37.0 42.9, 20.2† 5.5, 6† White-bellied sunbird (Cinnyris talatala) SN 9.0±1.4 4,7,8 - H ns ns S ns -
99.8±0.05 99.7±0.2 99.7±0.1
4 3.3±0.5 8.3±2.2 12.7±3.1 15.4±4.5 5.5
Broad-tailed hummingbird (Selasphorus platycercus) SN 3.3±0.1 4,6,7,18 - - ns ns ns ns -
95.0±0.02 - -
3 2.1±0.4 4.8±1.6 Silvereye (Zosterops lateralis) Fr, GN 9.0±0.4 1 H H H ns S S S
98.7±0.3 99.9±0.1 97.7±0.5
4 6.2±1.1 4.1±1.5 7.5±2.8 22.9 5
Singing honeyeater (Lichenostomus virescens) GN 28.9±4.1 1 H H H ns ns ns ns
99.6±0.2 99.9±0.1 99.3±0.3
7 8.3±1.7 13.6±8.6 23.7±14.5 24.9, 20.0† 5.5, 6† Mistletoebird (Dicaeum hirundinaceum) Fr, GN ~8 1 H H H ns ns ns ns
98.4±1.4 99.8±0.2 99.3±0.4
- - - - -
0.146 0.584 1.168
Magnificent hummingbird (Eugenes fulgens) SN 7.1±0.2 5,6 H ns S
99 99 99
3 3.5±0.5 21.4±4.2* - - - Cinnamon-bellied flowerpiercer (Diglossa baritula) SN 8.1±0.2 5,6 H ns S
99 99 99
4 3.7±0.2 3.3±0.6 10.2±1.9 59.5 6
0.146 0.73 1.022
Broad-billed hummingbird (Cynanthus latirostris) SN 2.9±0.2 16,17 ns ns ns
99±2.4 97±4.9 98±2.4
3 1.7 5.6±0.9 - - -
0.146 0.29 0.438 0.584 0.73
'Hexoses'
Malachite sunbird (Nectarina famosa ) SN ~16 11,14 H ns ns ns S
>99 >99 Village weaver (Ploceus cucullatus) GN 36.7±2.8 12 H H ns ns ns
~90-94 ~96–98
Speckled mousebird (Colius striatus) GN ~47 9 H H ns ns S
~84-87 ~89–93 Dark-capped bulbul (Pycnonotus tricolor) GN ~37 10 H H H H H
~65-85 ~75–95
Red-winged starling (Onychognathus morio) GN ~126 15,19 H H H H ns
0 ~64–73
nd
0.193 0.643
Knysna tauraco (Tauraco corythaix) Fr ~260 13 S(vs G) ns Purple-crested tauraco (Tauraco porphyreolophus) Fr ~250 13 ns ns

108
5.3 MATERIALS AND METHODS
5.3.1 Birds and their maintenance
The mistletoebird is a specialised frugivore that feeds primarily on mistletoe fruit
(Richardson and Wooller 1988), but also includes nectar and insects in its diet (Reid
1990). The silvereye is a generalist, feeding on fruit, nectar and insects (Wilkinson
1931; Thomas 1980; Richardson and Wooller 1986). The singing honeyeater is a
nectarivore that also ingests a relatively high proportion of insects (Collins and
Morellini 1979; Richardson and Wooller 1986); both singing honeyeaters and silvereyes
have more muscular gizzards than specialised nectarivores due to their ingestion of
insects (Richardson and Wooller 1986), therefore we have classified these species as
generalist nectarivores (Table 5.1).
Singing honeyeaters (n=8) and silvereyes (n=8) were captured on the grounds of
Murdoch University, Perth, Western Australia (WA; 32°04′S, 115°50′E) by mist-netting
in May 2009 and January 2010, respectively. There is no measure for sexual
dimorphism in plumage for either species, so their sex could not be determined.
Mistletoebirds (four male and two female) were captured on private property at York,
WA (31°50′S, 116°44′E), in December 2010, January and March 2011. All birds were
acclimated to captive conditions for at least two weeks before the commencement of
experimental trials.
Birds were housed in individual outdoor aviaries (116 x 160 x 210 cm), but were
confined to smaller cages (47 x 54 x 41 cm) placed within each aviary for the
experiments. During the period of captivity, all three species were fed a maintenance

109
diet of Wombaroo® nectarivore mix (Wombaroo Food Products, South Australia),
which contains sucrose as the main sugar type, supplemented with additional sucrose or
equal parts of glucose and fructose for a total sugar content of c. 25% w/w dry matter.
Birds fed through a small hole (c. 1-1.5 mm diameter) from plastic, stoppered syringes
hung on the sides of the cage. The frugivorous silvereyes and mistletoebirds were also
fed a variety of fleshy fruits (e.g. mistletoe fruit, watermelon, grapes, apricots) daily.
Martínez del Rio (1990) reported that measured sugar preferences in hummingbirds
were not correlated with the sugar type of their maintenance diet. All animal care
procedures and experimental protocols adhered to Murdoch University Animal Ethics
Committee regulations (R1137/05 and R2175/08). Birds were collected under permits
issued by the Western Australian Department of Environment and Conservation (DEC).
5.3.2 Apparent assimilation efficiency (AE*)
Singing honeyeaters (n=8), silvereyes (n=8) and mistletoebirds (3 male, 2 female) fed
ad libitum from sucrose and hexose solutions at three concentrations (0.25, 0.5, 1 mol·L-
1) for 24 h. Each bird fed from each sugar solution at each diet concentration, with
sugar type and concentration randomised. Trials commenced within 30 min after
sunrise (0500 to 0716 WST). Maintenance diet was removed one hour before sunrise to
ensure all previously ingested food (i.e. from the previous day) was voided before trials
commenced. Trays were placed under experimental cages to collect excreta, and liquid
paraffin was placed in containers directly beneath feeders to collect any diet spilt. Food
intake was recorded over 24 h by weighing feeders (0.01 g). Excreta produced over 24
h were allowed to evaporate and were then reconstituted and collected with a known
volume of distilled H2O and stored at -20 °C until analysis.

110
Glucose assays: Two replicates of each excreta sample (100 µl) were incubated
at room temperature (~21 °C) for 15 min with 500 µl of hexokinase-glucose-6-
phosphate dehydrogenase enzymatic assay reagent (G3293, Sigma Aldrich).
Absorbance was then measured at 340 nm relative to distilled water by
spectrophotometry (UV mini 1240, Shimadzu Scientific Instruments, Balcatta, WA,
Australia). Standard ‘reagent blanks’ were included for each excreta sample.
Fructose assays: Two replicates of each excreta sample (45 µl) were incubated
at room temperature (~21 °C) for 15 min with 650 µl of hexokinase-glucose-6-
phosphate dehydrogenase enzymatic assay reagent (G3293, Sigma Aldrich) and 5 µl
phosphoglucose isomerase from baker’s yeast (F2668, Sigma Aldrich). Absorbance
was then measured at 340 nm relative to distilled water by spectrophotometry. Standard
‘reagent blanks’ were included for each excreta sample.
Sucrose assays: Two replicates of each excreta sample (25 µl) were incubated
at room temperature (~21 °C) for 10 min with 25 µl invertase from baker’s yeast
sucrose assay reagent (S1299, Sigma Aldrich). 650 µl of hexokinase-glucose-6-
phosphate dehydrogenase enzymatic assay reagent (G3293, Sigma Aldrich) was then
added, and samples incubated for a further 15 min. Absorbance was then measured at
340 nm relative to distilled water by spectrophotometry. Standard ‘reagent blanks’
were included for each excreta sample.
Apparent assimilation efficiency (AE*) was estimated separately for sucrose,
glucose and fructose as:
AE* = (sugarin-sugarout) / (sugarin) (eq. 5.1)

111
where sugarin (g) is calculated as the concentration (g·L-1
) of sugar in the ingested diet
multiplied by the volume of solution ingested (L), and sugarout (g) is the sugar
concentration (g·L-1
) in the total volume of excreta plus rinse water (L).
AE* data were arcsine square root transformed (Zar 1999) before analysis.
Differences in AE between sugar type, sugar concentration, species and total sugar
intake were assessed by ANCOVA with total sugar intake as a covariate and Tukey-
Kramer post hoc tests for unequal sample sizes as required. Additional data for sucrose
AE* (excluding species from the sub-family Muscicapoidea) were obtained from
Fleming et al. (2008) and differences between specialist (n=21 species) and generalist
(n=13 species) nectarivores assessed by Mann-Whitney U test.
5.3.3 Sugar preference trials
Singing honeyeaters (n=8), silvereyes (n=8) and mistletoebirds (four males) participated
in sugar preference trials which, following the methodology of Fleming et al (2008) for
consistency, lasted for 6 h, commencing within 30 min of sunrise (0535 to 0705 WST).
Sugar preferences were examined by comparing the intake of seven paired
concentrations of sucrose and energetically equivalent hexose (1:1 glucose:fructose)
solutions: 0.075, 0.1, 0.25, 0.5, 0.75, 1 and 2 mol·L-1
Sucrose Equivalents (SE). Hexose
diets were equicaloric with, but had approximately twice the osmolality of sucrose
solutions (Fleming et al. 2008). Birds were simultaneously presented with pairs of
feeders containing sucrose and hexose concentrations of the same SE molarity in
random order. To account for potential sources of side bias (Jackson et al. 1998a;
Jackson et al. 1998b), the start position of each feeder was random, with the positions of

112
the feeders switched half way though each trial. Each concentration was also tested on
each bird twice, with the starting position of the feeders reversed on the second trial.
Liquid paraffin was placed in containers directly below feeders to collect any diet spilt.
Sugar intake was determined by weighing the feeders before and after trials (0.01 g) and
calculating the mass of sugar ingested by taking into account the density of each diet.
Trials were conducted approximately every second day, with at least one day of rest and
maintenance diet between trials. Trials were repeated for a third time in the instance of
low diet intake (a few individuals did not drink when first offered the lowest
concentration of 0·075 mol·L-1
SE, but increased intake during subsequent trials). The
average intake over all trials for each diet was used to calculate a sugar preference
index, with hexose intake expressed as a proportion of total sugar intake (H/(H+S),
where a value of 0.5 indicates no preference whilst a value close to 1 indicates a strong
hexose preference).
Average food intake (g sugar in 6 h of each trial) was analysed via one-way
ANOVA for each species, with diet sugar concentration as the independent factor and
Tukey’s Honest Significant Differences (HSD) post-hoc tests as required. Preference
data were arcsine square root transformed (Zar 1999) before analysis by one-way
ANOVA for each species, with diet sugar concentration as the independent factor and
Tukey’s HSD tests as required. Differences in preferences between species and diet
concentrations were assessed via two-way ANOVA with Tukey-Kramer post hoc tests
for unequal sample sizes as required. For each species, sugar preference at each
concentration was analysed by one-sample t-tests (Sokal and Rohlf 1995) comparing the
arcsine-transformed square root of preference indices against 0.5 (no preference).

113
5.3.4 Intestinal enzymes
Study species and dissection: Red wattlebirds (n=3), singing honeyeaters (n=7),
silvereyes(n=4), New Holland honeyeaters (n=9) and rainbow lorikeets (n=7) were
captured by mist- or cannon-netting near Perth, Western Australia, between 2007 and
2011 (see Table 5.2 for details). We did not have access to mistletoebirds for this part
of the study. Birds were not fasted prior to euthanasia. Birds were euthanised via
Isoflurane overdose or a 1:1 sodium pentobarbital:distilled H2O solution injected into
the heart. Sex was determined by examination of reproductive organs upon dissection.
The intestines were removed from stomach to cloaca within 10 min of euthanasia,
dissected length-wise, cut into three sections (proximal, medial and distal) and
measured (length and width to calculate nominal surface area, cm2). The intestinal
sections were then rinsed in 0.75 mol·L-1
NaCl, blotted, and weighed (0.001 g). Each
section was then frozen in liquid nitrogen and stored at -80 °C until enzyme activity
analysis (<12 months after euthanasia). All animal care procedures and experimental
protocols adhered to Murdoch University Animal Ethics Committee regulations
(R1137/05). Birds were collected under permits issued by DEC. Some tissues were
kindly provided by Joao Coimbra (The University of Western Australia Animal Ethics
Committee RA/3/100/927 and DEC permit SF007556).
Disaccharidase assays: Intestinal samples were thawed at room temperature
(21±2 °C) and homogenised (Heidolph ‘DIAX 600’, Heidolph Instruments, Schwabach,
Germany) in 0.3 mol·L-1
mannitol in 0.001 mol·L-1
HEPES/KOH pH 7.5 buffer (99 to
128 mg intestine·mL-1
of homogenate). Aliquots of homogenates were immediately
diluted in 0.3 mol·L-1
mannitol in 1.0 mmol·L-1
HEPES/KOH pH 7.5 buffer (1:40 for

114
sucrase, 1:300 for maltase) and frozen in liquid nitrogen and stored at -80 °C until
disaccharidase (sucrase and maltase) assays were performed.
Disaccharidase activity was measured according to Dahlqvist (1984) as modified
by Martínez del Rio et al. (1995). Diluted intestinal homogenates (30 µL) were
incubated with 30 µL of 0.056 mol·L-1
sugar substrate (maltose or sucrose) solutions in
0.1 mol·L-1
maleate NaOH pH 6.5 buffer at 40 °C for 20 min. 400 µL of a stop/develop
reagent was then added, and samples were vortexed and incubated at 40 °C for a further
30 min. Stop/develop reagent was made by dissolving one bottle of Glucose
oxidase/peroxidase reagent (G3660, Sigma Aldrich) in 19 mL 0.5 mol·L-1
phosphate
buffer (NaH2PO4/Na2HPO4) pH 7.0 plus 19 mL 1 mol·L-1
Tris/HCl pH 7.0, plus 2 mL
O-dianisidine solution (2.5 mg O-dianisidine dihydrochloride [D3252, Sigma Aldrich]
per mL dH2O). Lastly, 400 µL 12NH2SO4 was added and the absorbance read at 540
nm. Maltase and sucrase activity (µmol·min-1
) was measured for each section of
intestine and summed together to calculate ‘total activity’ for each individual. Total
enzyme activity for each individual bird was then adjusted to optimal pH, and then
standardised for nominal gut surface area (µmol·min-1
·cm2).
Differences in standardised sucrase activity between the five species were
assessed by one-way ANOVA followed by Tukey-Kramer post hoc tests for unequal
sample sizes. Least squares linear regression was also used, with data averaged for
species to assess relationships between log body mass (mb) and log gut nominal surface
area and log total sucrase and maltase activity.
As maltose may be hydrolysed by both sucrase and maltase (Alpers 1987;
Martinez del Rio 1990), the activity of both disaccharidases were measured. The slope

115
of the relationship between sucrase and maltase indicates the amount of maltase activity
relative to sucrase activity and the y-intercept provides an estimate of maltase activity
that occurs in the absence of sucrase (Martinez del Rio 1990). The relationship between
standardised sucrase and maltase activities was therefore examined using least squares
linear regression.
Least squares multiple linear regression was also used to assess the relationship
between hexose preference (scored as the minimum diet concentration at which birds
show no sugar preference) and standardised sucrase activity with data averaged for all
individuals for eleven species (Table 5.1). Studies that have used only a few diet
concentrations may not yield accurate information in this regard, but the use of the
minimum no-preference concentration is a conservative estimate of sugar type
preference. Minimum no-preference concentration values also allowed inclusion of
species that do not exhibit hexose preference, e.g. broad-billed (Cynanthus latirostris)
and broad-tailed hummingbirds (Selasphorus platycercus). The red-winged starling
(Onychognathus morio), like other starlings, lacks the intestinal enzyme sucrase and
therefore has non-detectable levels of sucrase activity (Bizaare et al. 2012). We then
included the red-winged starling in the analyses with a sucrase activity value of 0.
Phylogenetic analyses: As phylogenetic relationships may confound the
inferences of allometric analyses (Garland et al. 1992; Garland and Adolph 1994;
Rezende and Diniz-Filho 2012), these conclusions were corroborated using
phylogenetically-independent contrasts. Felsenstein’s (1985) independent contrasts
method was used in the computer program PDAP (Garland et al. 1992; Garland et al.
1993; Garland et al. 1999; Garland and Ives 2000) running through Mesquite (Version

116
2.75) (Midford et al. 2009). Phylogenetically-independent contrasts (PIC) of dependent
and independent variables were calculated and standardised utilising the branch length
transformation (Garland et al. 1992). Evolutionary relationships (Fig. 5.1) were
determined using the phylogenetic tree of Ericson et al. (2006) as a ‘backbone’ with sets
of pseudo-prosterior samples of the dated phenologies built by Jetz et al. (2012)
subsampled and then pruned for our full set of species. Regressions were fitted to
standardised PIC values, forcing the data through the origin (Garland et al. 1992).
Cynanthus latirostris
Eugenes fulgens
Selasphorus platycercus
Trichoglossus haematodus
Lichenostomus virescens
Anthochaera carunculata
Phylidonyris novaehollandiae
Zosterops lateralis
Onychognathus morio
Cinnyris talatala
Diglossa baritula
Figure 5.1 Evolutionary relationships.
Phylogenetically independent contrast values were calculated using the evolutionary phylogenetic tree of
Ericson et al. (2006) as a backbone, with sets of pseudo-posterior samples of the dated phenologies built
by Jetz et al. (2012) subsampled and then pruned for our full set of species, through the website
birdtree.org.

117
5.3.5 Foraging data and Australian nectar composition
Foraging data for silvereyes, singing and New Holland honeyeaters, red wattlebirds and
rainbow lorikeets were compiled from the Western Australian Pollination Database
(Brown et al. 1997). As the foraging records for rainbow lorikeets in Western Australia
(Brown et al. 1997) were rather limited (due to their recent introduction in the 1960s
and subsequent establishment as a pest species in Perth, WA), detailed foraging records
for rainbow lorikeets were also compiled from the Queensland-New South Wales
border region (Cannon 1984). Nectar compositions of Australian plants were compiled
from published and unpublished sources (Baker and Baker 1982b; Paton 1982;
Gottsberger et al. 1984; McFarland 1985; Nicolson 1994; Davis 1997; Baker et al.
1998; Nicolson and Van Wyk 1998; Hölscher et al. 2008; Morrant et al. 2010; S.W.
Nicolson and B.-E. Van Wyk, pers. comm.). The ratio of hexoses to sucrose was
calculated as H/(H+S), and nectars classed as hexose-dominant (>0.8), hexose-rich
(0.6–0.8), mixed (0.4–0.6), sucrose-rich (0.2–0.4) or sucrose-dominant (<0.2). Ratios
were adapted from Baker and Baker (1982b), with new classifications developed for
this study. To examine the relationship between nectar type and foraging preference of
the five species, a contingency table was constructed for foraging data for each species
and the five nectar classifications (excluding plants for which we lack information on
the nectar composition – classified as ‘unknown’) and analysed for significance by
Pearson’s χ2 square analysis (with Bonferroni correction applied).

118
5.3.6 General statistical analysis
Data are reported as mean ± 1 SD throughout, with n referring to the number of animals.
Statistical analyses were performed with Statistica (StatSoft Inc 2007) and SPSS (SPSS
Inc, Chicago, IL, USA). Statistical significance was accepted for α<0.05.

119
Table 5.2 Details of birds euthanased for digestive enzymes.
*Nedlands – grounds of The University of Western Australia (UWA); Murdoch – grounds of Murdoch
University; Shenton Park – grounds of the UWA Shenton Park Field Station; Roleystone – grounds of
the Araluen Country Club; Bentley – grounds of Curtin University (bird flew into window and died ~1 h
later); Wattle Grove – bird obtained from Wattle Grove Veterinary Clinic after an unknown period in
captivity; Perth Airport – grounds of Perth Domestic Airport as part of a Department of Conservation
(DEC) culling program. +Birds were euthanased via 1:1 sodium pentobarbital:distilled H2O solution
injected into the heart or by Isoflurane overdose.
Species Location of capture*
Year of capture
Method of capture
Sex (M, F)
Period of
captivity Method of euthanasia+
Red wattlebird
(Anthochaera carunculata)
Nedlands, WA (31°58′S, 115°49′E)
Murdoch, WA (32°04′S, 115°50′E)
2010
2007
Mist-netting
Mist-netting
(3, 0)
(4, 1)
< 72 h
5 mo
Sodium pentobarbital
Isoflurane
Singing honeyeater
(Lichenostomus virescens)
Shenton Park, WA (31°57′S, 115°47′E)
Murdoch, WA (32°04′S, 115°50′E)
2010
2007
Mist-netting
Mist-netting
(2, 1)
(4, 0)
< 7 h
5 mo
Sodium pentobarbital
Isoflurane
New Holland honeyeater
(Phylidonyris novaehollandiae)
Roleystone, WA (32°08′S, 116°05′E)
Murdoch, WA (32°04′S, 115°50′E)
2011
2007
Mist-netting
Mist-netting
(3, 1)
(5, 0)
< 7 h
14 mo
Sodium pentobarbital
Isoflurane
Silvereye
(Zosterops lateralis)
Roleystone, WA (32°08′S, 116°05′E) 2011 Mist-netting (3, 1) < 7 h Sodium pentobarbital
Rainbow lorikeet
(Trichoglossus haematodus)
Bentley, WA (32°0′S, 115°53′E)
Wattle Grove, (32°0′S, 115°59′E)
Perth Airport, WA (31°55′S, 115°57′E)
2011
2011
2006
Unknown
Unknown
Cannon-netting
(1, 0)
(1, 0)
(5, 0)
< 1 h
Unknown
>12 mo
Natural death
Sodium pentobarbital
Isoflurane

120
5.4 RESULTS
5.4.1 Apparent assimilation efficiency (AE*)
Silvereyes, singing honeyeaters and mistletoebirds displayed high assimilation
efficiencies for all three sugar types (>97.5%, Table 5.1). AE* was not different
between sugar concentrations (F1,191 =2.56, P=0.111) but varied with sugar intake (F1,191
=10.20, P=0.002), where AE* increased as sugar intake decreased. AE* varied between
species (F2,191=7.19, P=0.001), being greatest for singing honeyeaters and least for
silvereyes overall. AE* also varied between sugars (F2,191 =65.54, P<0.001), being
greatest for glucose and least for fructose. The significant sugar type by species
interaction (F4,191=11.54, P<0.001) demonstrated that silvereyes assimilated less
fructose than mistletoebirds and singing honeyeaters, and singing honeyeaters
assimilated more sucrose than mistletoebirds and silvereyes.
AE* for sucrose differed significantly between generalist (86.24±16.21%, n=13)
and specialist nectarivores (98.09±1.25%, n=21) (U=64, Z=2.62, P=0.0093).
Comparable data for hexoses were not available.
5.4.2 Sugar preferences
Silvereyes, singing honeyeaters and mistletoebirds all failed to consume sufficient
volumes to maintain energy balance on the most dilute diets, with significantly lower
intakes of sugar compared with the more concentrated diets. Compensatory feeding
(identified here as diet concentrations where sugar intake was not significantly different
from the most concentrated diets) was observed for singing honeyeaters for diets ≥0.25

121
mol·L-1
, but only for diets ≥0.5 mol·L-1
in mistletoebirds and silvereyes (Fig. 5.2).
Sugar preferences were influenced by sugar concentration (F6,279=36.17, P<0.001), with
all three species showing significant preferences for hexose solutions at low sugar
concentrations (Fig. 5.3). Sugar preferences differed significantly between the species
(F2,279=4.460, P=0.012), with silvereyes displaying significant preferences for sucrose
diets at the higher concentrations (i.e. ≥0.75 mol·L-1
; Fig. 5.3).
Diet concentration (mol·L-1
)
0.0 0.5 1.0 1.5 2.0 2.5
Die
t su
ga
r in
take
(g
)
0
1
2
3
4
(n=8)
(n=8)
(n=4)
Figure 5.2 Concentration-dependent total sugar intake.
Mistletoebirds (circle), silvereyes (triangle) and singing honeyeaters (square) were offered paired sucrose
and hexose (fructose + glucose) solutions of varying concentrations: 0·075, 0·1, 0·25, 0·5, 0·75, 1 and 2
mol·L-1
sucrose equivalents (SE). Diets where birds did not achieve energy balance (statistically lower
intake than the maximal sugar intake) are indicated with increasingly lighter shaded symbols (one-way
ANOVA and Tukey’s HSD test). n refers to number of individuals.

122
0.0 0.5 1.0 1.5 2.0
Sug
ar
pre
fere
nce
0.0
0.2
0.4
0.6
0.8
1.0
Diet concentration mol·L-1
0.0 0.5 1.0 1.5 2.0
0.0
0.2
0.4
0.6
0.8
1.0
0.0 0.5 1.0 1.5 2.0
0.0
0.2
0.4
0.6
0.8
1.0
a) Mistletoebirds (n=4) b) Silvereyes (n=8) c) Singing honeyeaters (n=8)
*a
*a
*a
b
b
b
b
*a
*a
*a
b *b *b
*b
*a,b
*a
*b,c
c,dc,d
c,dd
He
xo
se
Sucro
se
Figure 5.3 Concentration-dependent sugar preferences.
a) mistletoebirds (circle), b) silvereyes (triangle) and c) singing honeyeaters (square) were offered paired sucrose and hexose (fructose + glucose) solutions of varying concentrations:
0·075, 0·1, 0·25, 0·5, 0·75, 1 and 2 mol·L-1
sucrose equivalents (SE). Diets where birds did not achieve energy balance (statistically lower intake than the maximal sugar intake) are
indicated with increasingly lighter shaded symbols. Letters indicate diets that are statistically different from each other (one-way ANOVA with Tukey’s HSD test). Asterisks
indicates concentrations where there was a significant preference for either hexose or sucrose diets (one-sample t-test). n refers to number of individuals.

123
5.4.3 Intestinal enzymes
Body mass, gut nominal surface area, total sucrase activity and kinetic parameters for
rainbow lorikeets, silvereyes, singing and New Holland honeyeaters and red wattlebirds
are summarised in Table 5.1, with data for additional species reported from the
literature. Total maltase activity and associated kinetic parameters are detailed in Table
5.3. Sucrase and maltase activity, as a function of substrate concentration, followed
Michaelis-Menten kinetics. Sucrase and maltase activities were highest in proximal
sections of the intestine and decreased distally (data not shown). There were species
differences in standardised sucrase activity (one-way ANOVA: F5,26=4.87, P=0.003);
rainbow lorikeets and silvereyes had significantly lower sucrase activity than red
wattlebirds (post hoc: P=0.012, P=0.032). When comparing data averaged for each
species, gut nominal surface area increased with body mass (F1,9=120.88, P<0.001,
R2=0.94; Fig. 5.4a). This result was confirmed by PIC analysis of log10body mass
PIC
and log10gut nominal surface areaPIC
(F1,8=62.53, P<0.001, R2=0.887). Total sucrase
activity was also significantly correlated with body mass (F1,9=12.46, P=0.008,
R2=0.61, Fig. 5.4b; confirmed by PIC analysis: F1,8=15.08, P=0.006, R
2=0.653). Total
maltase activity showed a borderline correlation with body mass, which was not upheld
by PIC analysis (F1,8=5.97, P=0.04, R2=0.43, Fig. 5.4c; PIC: F1,8=0.432, P=0.532,
R2=0.051). Standardised maltase activity was not significantly correlated with
standardised sucrase activity (F1,9=0.18, P=0.686, R2=0.02; Fig. 5.4d), which was
confirmed by PIC analysis (F1,8=4.113, P=0.082, R2=0.340).
For eleven species tested, hexose preference (the minimum no-preference
concentration) was significantly correlated with standardised sucrase activity

124
(F1,10=13.44, P=0.005, R2=0.60; Fig. 5.4e). Phylogenetically-corrected analysis
confirmed this result (F1,9=18.0, P=0.003, R2=0.667). Birds with the lowest
standardised sucrase activity showed hexose preference at more concentrated diets (i.e.
greater minimum no-preference concentration), and birds with the greatest standardised
sucrase activity either showed no hexose preference (e.g. hummingbirds) or hexose
preference on only the most dilute diets.

125
sucrase activity (µmol·min-1
·cm2
)
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0
Min
imu
m n
o-p
refe
ren
ce c
once
ntr
ation
(m
ol)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Broad-tailed hummingbird
Broad-billed hummingbird
Cinnamon-bellied flowerpiercer
Magnificent hummingbird
New Holland honeyeater
Rainbow lorikeet
Red wattlebird
Red-winged starling
Silvereye
Singing honeyeater
White-bellied sunbird
0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.2
log
[su
cra
se
activity, µ
mo
l·m
in-1
]
0.4
0.6
0.8
1.0
1.2
1.4
1.6
log [body mass, g]
0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.2
log
[g
ut n
om
ina
l su
rfa
ce a
rea
, cm
2]
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Cinnamon-bellied flowerpiercer
Magnificent hummingbird
New Holland honeyeater
Rainbow lorikeet
Red wattlebird
Silvereye
Singing honeyeater
White-bellied sunbird
Broad-billed hummingbird
Broad-tailed hummingbird
0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.2
log
[m
alta
se a
ctivity, µ
mo
l·m
in-1
]
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
sucrase activity (µmol·min-1
·cm2)
0 1 2 3 4 5
ma
lta
se a
ctivity (
µm
ol·m
in-1
·cm
2)
2
4
6
8
10
12
14
16
18
log [body mass, g]
log [body mass, g]
a) b)
c) d)
Rainbow lorikeet
Silvereye
Cinnamon-bellied flowerpiercer
Singing honeyeater
New Holland honeyeater
Magnificent hummingbird
Red wattlebird
White-bellied sunbird
Broad-tailed hummingbird
Broad-billed hummingbird
e)
Figure 5.4 Gut nominal surface area and intestinal enzymes.
Relationships between body mass and a) gut nominal surface area; b) total sucrase activity and c) total
maltase activity. d) Relationship between maltase and sucrase activity (both standardised by gut nominal
surface area). e) Relationship between degree of hexose preference (i.e. minimum no-preference
concentration) and standardised sucrase activity. Data are averaged for each species. White symbols
denote generalist nectarivores, grey symbols denote specialised nectarivores. See Table 5.1 for details of
references, numbers of individuals and diet categories.

126
5.4.4 Foraging data and Australian nectar composition
Foraging data (Cannon 1984; Brown et al. 1997) are summarised in Fig. 5.5. Foraging
records indicate that all of the focal species have a diverse diet, including multiple plant
taxa in their diets (Fig. 5.5a). Nectar composition was available for 16 Australian
genera (Fig. 5.6, Table 5.4). There was a significant association between bird species
and nectar type (χ2
16=532.77, P<0.001; Fig. 5.5b), with rainbow lorikeets avoiding
sucrose and mixed nectars in favour of hexose-rich nectars and the three honeyeater
species avoiding hexose nectars in favour of sucrose-dominant, -rich and mixed nectars.
The foraging preferences of silvereyes were not very clear, which may reflect few
foraging records (n=44) for this species.

127
Pro
port
ion o
f fo
ragin
g o
bserv
ations
0.0
0.2
0.4
0.6
0.8
1.0
unknown
sucrose dominant
sucrose rich
mixed
hexose rich
hexose dominant
Pro
port
ion o
f fo
ragin
g o
bserv
ations
0.0
0.2
0.4
0.6
0.8
1.0Other
Tristania
Schefflera
Macaranga
Paraserianthes
Beaufortia
Acacia
Lambertia
Calothamnus
Grevillea
Banksia
Melaleuca
Eucalyptus
Adenanthos
Callistemon
Amyema
Anigozanthos
Corymbia
Hakea
b)
a)
Rainbow lorik
eet (1174, 3
0)
New Holla
nd honeyeater (
433, 31)
Red wattle
bird (2
64, 19)
Singing honeyeater (
198, 25)
Silvereye (5
9, 23)
***A
***P
***P
***P
***P
***A***A
***P
***A *A
***P
***P
*P
*A
Figure 5.5 Feeding observations for New Holland honeyeaters, red wattlebirds, singing
honeyeaters and silvereyes in Western Australia (Brown et al. 1997) and rainbow lorikeets in
Western Australia and the Queensland-New South Wales border region (Cannon 1984; Brown et
al. 1997).

128
a) Feeding observations grouped by plant genus. White lines indicate hexose dominant and rich nectars,
solid light grey indicates mixed sugars, dark grey lines indicates sucrose dominant and rich nectars. Dots
indicate unknown sugar composition, solid white indicates other genera comprising <2% of feeding
observations (including Agonis, Adansonia, Astroloma, Billardiera, Blancoa, Bombax, Bossiaea,
Braxychiton, Brachysema, Chasmanthe, Chorilaena, Cosmelia, Crotalaria, Darwinia, Diplolaena,
Eremophilia, Erythina, Gastrolobium, Hardenbergia, Hybanthus, Jacksonia, Jansonia, Kunzea,
Leptosema, Leptospermum, Loranthus, Lysiana, Macropidia, Microcorys, Muiriantha, Nematolepis,
Nicotiana, Nutysia, Pimelea, Pittosporum, Psoralea, Regelia, Temletonia, and Xanthorrea). In
parentheses: total number of foraging observations, number of plant genera. b) Feeding observations
grouped by nectar composition; see text for definitions of nectar categories. Asterisks denote significant
preference (P) or avoidance (A) of nectar categories as determined by χ2 analysis with Bonferroni
correction applied (P<0.05*, P<0.01**, P<0.001***).

129
Nectar compostion (% fructose, glucose, sucrose)
0 20 40 60 80 100
Lambertia (3)
Macadamia (1)
Calothamnus (1)
Grevillea (25)
Banksia (23)
Melaleuca (6)
Eucalyptus (18)
Adenanthos (3)
Erythrina (1)
Callistemon (1)
Telopea (1)
Amyema (1)
Anigozanthos (1)
Corymbia (1)
Hakea (4)
Stenocarpus (1)% fructose
% glucose
% sucrose
Figure 5.6 Average nectar composition from 16 Australian plant genera (mean fructose, glucose, sucrose).
In parentheses: number of species sampled for each genus. Fructose is represented by white bars, glucose by grey and sucrose by black. Hexose-dominant nectars include:
Sternocarpus, Hakea, Corymbia, Anigozanthos, Amyema, Telopea, Callistemon, Erythrina and Adenanthos; hexose-rich nectars: Eucalyptus and Melaleuca; mixed nectars: Banksia
(including former Dryandra species); sucrose-rich nectars: Grevillea and Calothamnus; sucrose-dominant nectars: Lambertia and Macadamia.

130
Table 5.3 Summary of maltase activity.
Data are presented as mean ± 1 SD. - indicates not tested or not available. References: 1Data was obtained from this study;
6Schondube and Martínez del Rio (2004);
7McWhorter
et al. (unpublished), 17
Martínez del Rio (1990). Km and pH optima: Kinetic parameters obtained using at least n=1 tissue homogenate (proximal intestinal section): † two data sets
for birds caught in 2010–2011 (n=1) or 2006–2007 (n=1).
Digestive capacity (maltase)
ref Total activity (µmol·min-1) Vmax
(μmol∙min-1) Km (mM) pH optima
Rainbow lorikeet, Trichoglossus haematodus (n=7) 1 174±89.7 207.4±103.2 4.5, 5.8 5, 6 Silvereye, Zosterops lateralis (n=4) 1 50.7±20.3 60.5±24.2 5.4 6.5 Singing honeyeater, Lichenostomus virescens (n=7) 1 91.3±47.9 100.6±51.3 3.9, 2.6 5.5, 4.5 New Holland honeyeater, Phylidonyris novaehollandiae (n=9) 1 40.6+19.0 50.5±22.0 12.5, 4.3 4.5, 5 Red wattlebird, Anthochaera carunculata (n=8) 1 213.9±119 258.7±130.2 12.6, 4.3 5, 5.5 Broad-tailed hummingbird, Selaphorus platycercus (n=2) 6 7.7±1.4 - - - Magnificent hummingbird, Eugenes fulgens (n=3) 6 17.0±3.3 - - - Cinnamon-bellied flowerpiercer, Diglossa baritula (n=4) 6 30.1±4.0 33.2±4.4 2.8 5.5 White-bellied sunbird, Cinnyris talatala (n=4) 7 41.0±7.9 44.3±8.5 2.2 5 Broad-billed hummingbird, Cynanthus latirostris (n=3) 17 14.0±2.3 - - -

131
Table 5.4 Nectar composition of 16 Australian plant genera.
Data are presented as mean ± 1 SD, n=total number of species.
Family Genus Fructose (%) Glucose (%) Sucrose (%) n Reference
Fabaceae Erythrina 46 51 3 1 (Baker and Baker 1982a)
Haemodoraceae Anigozanthos 45 55 0 1 (Hölscher et al. 2008)
Loranthaceae Amyema 59 40 1 1 (Paton 1982)
Myrtaceae Callistemon 48 50 2 1 S.W. Nicolson and B.-E Van Wyk, unpublished Calothamnus 12 8 80 1 S.W. Nicolson and B.-E Van Wyk, unpublished Corymbia 52 48 0 1 (Nicolson 1994) Eucalyptus 41.6±15.3 31.40±12.15 26.91±21.68 18 (Nicolson 1994; Davis 1997; Baker et al. 1998; Morrant et al.
2010; S.W. Nicolson and B.-E Van Wyk, unpublished) Melaleuca 41.5±7.5 31.76±12.95 26.70±19.56 6 (Morrant et al. 2010; S.W. Nicolson and B.-E Van Wyk,
unpublished)
Proteaceae Adenanthos 46.0±3.0 46.7±5.1 7.3±8.1 3 (Nicolson and Van Wyk 1998) Banksia 23.6±21.0 24.1±21.0 52.2±41.7 23 (McFarland 1985; Nicolson and Van Wyk 1998) Grevillea 8.5±16.9 12.5±24.6 82.7±33.7 25 (Gottsberger et al. 1984; Nicolson and Van Wyk 1998) Hakea 49.4±1.3 50.6±1.3 0 4 (Nicolson and Van Wyk 1998) Lambertia 1.3±0.6 1.0±1.0 97.7±1.5 3 (Nicolson and Van Wyk 1998) Macadamia 4 4 92 1 (Nicolson and Van Wyk 1998) Stenocarpus 51.7±3.1 48.3±3.1 0 1 (Nicolson and Van Wyk 1998) Telopea 49.0±0.0 49.3±1.5 1.7±1.5 1 (Nicolson and Van Wyk 1998)

132
5.5 DISCUSSION
We investigated physiological mechanisms that may explain the concentration-
dependence of sugar type preferences using data obtained from laboratory trials and
literature records, and compared diet preference data with foraging records.
Supporting our first prediction, we found that specialised nectarivorous species
exhibited greater apparent assimilation efficiencies for sucrose than generalist
nectarivores when comparing broadly, using data available from this study (Table 5.1)
and the literature (Fleming et al. 2008). Not enough information for glucose and
fructose assimilation was available for generalist nectarivores (n=3) so we were unable
to make this broader comparison for hexoses. However, the Australian generalist
nectarivore species studied exhibited high apparent assimilation efficiencies (AE*) for
sucrose, glucose and fructose (all >97.5%) comparable with specialist nectarivores.
These results suggest that these Australian generalist nectarivores should be as capable
of feeding on both sucrose- and hexose-rich nectars as specialist nectarivores.
In terms of our second prediction, both specialist and generalist nectarivores
demonstrated concentration-dependent sugar preferences. The degree of preference for
hexose over sucrose solutions on dilute diets (assessed as the minimum no-preference
concentration) was negatively correlated with the capacity to hydrolyse sucrose. For
example, rainbow lorikeets, a specialist nectarivore, had one of the lowest sucrase
activity levels and correspondingly preferred hexose diets over a broad range of diet
concentrations. Hummingbirds, with the greatest sucrase activity levels, showed no
preference for hexose over sucrose.

133
Our third prediction was that diet preferences would match foraging records.
While some specialist nectarivores (e.g. New Holland honeyeaters) preferentially
foraged on sucrose nectars over hexose nectars in the wild, others (e.g. rainbow
lorikeets) preferred hexose-rich nectars to mixed and sucrose-rich and sucrose-dominant
nectars. These data therefore do not support a simplistic differentiation in diet
preferences between specialist and generalist nectarivores and indicate that the digestive
physiology of each species is more closely correlated with its diet preferences
(measured in the laboratory or foraging records in the field) than broad classifications
have led us to expect.
5.5.1 Are there differences in apparent assimilation efficiency between sugar
types?
Although all three Australian generalist nectarivore species assessed show high apparent
assimilation efficiencies of sucrose and hexoses, there were some differences between
these sugar types. AE* was greatest for glucose and least for fructose, and varied by
species. Greater AE* for glucose over sucrose has been noted in studies of other
species (Table 5.1) and may reflect the direct assimilation of glucose, but the need for
hydrolysis of sucrose before its constituent monosaccharides can be assimilated.
Fructose absorption (by GLUT5 transporters) appears to be more concentration-
dependent than the absorption of D-glucose (Holdsworth and Dawson 1964; Rand et al.
1993), therefore the lower AE* for fructose may reflect the availability of GLUT
transporters and reliance on facilitated diffusion (rather than secondary active transport
via SGLT1 transporter proteins as for glucose).

134
Our data revealed a clear distinction between specialist and generalist
nectarivores in terms of their AE*for sucrose. Specialist nectarivores uniformly have
high AE* for sucrose, while many generalist nectarivores have lower AE* which could
reflect lower sucrase activity. We could only compare AE* between generalist and
specialist nectarivores for sucrose, due to lack of available data for the other sugars.
However, because AE* values for Australian generalist and specialist nectarivores
feeding on all three sugars are >97.5%, these differences are not likely to be
functionally significant or impact the sugar preferences or foraging choices of these
birds.
5.5.2 Can we explain hexose preferences on dilute diets?
We examined whether hexose preference on dilute diets could be influenced by the
amount of intestinal sucrase activity. Across eleven bird species, hexose preference
(minimum no-preference concentration) was significantly negatively correlated with
sucrase activity (Fig. 5.4e). Birds with lesser capacities to digest sucrose show a
significant preference for hexose solutions at higher sugar concentrations. For example,
the rainbow lorikeet assessed in this study does not have the same sucrose digestive
capacity shown by other specialist nectarivores, with only one third the sucrase activity
of the red wattlebird, a similar-sized honeyeater (Fig. 5.4d, e). Rainbow lorikeets
prefered hexose solutions up to 0.75 mol·L-1
. By contrast, birds with greater capacities
to digest sucrose showed hexose preference on only the most dilute diets or no
preference over the range of concentrations tested. We included data for two
hummingbird species which show no sugar type preference at room temperature for the
minimum diet concentrations they have been tested on (0.146 and 0.25 mol·L-1
diets,

135
respectively). When tested at lower diet concentrations (0.1 mol·L-1
), the broad-tailed
hummingbird resorted to torpor rather than feeding on the dilute solutions (Fleming et
al. 2004). Challenging them with colder ambient temperatures (i.e. increasing their
metabolic demands; Fleming et al. 2004) may be the only way to test for evidence of a
hexose preference in hummingbirds. These data, together, demonstrates that preference
for hexose at low diet concentrations reflects the digestive capacity of bird species.
Most specialist nectarivores prefer hexoses at only the most dilute diet
concentrations tested, while many species of generalist nectarivores (e.g. the dark-
capped bulbul and red-winged starling) show hexose preference across a greater range
of diet concentrations (Table 5.1). However, the rainbow lorikeet (a specialist
nectarivore) shows significant hexose preference for more concentrated diets than other
specialist nectarivores. Furthermore, the simplistic categorisation of honeyeater species
as specialist or generalist, in itself, may also be problematic. These data therefore do
not support a simplistic distinction between specialist and generalist nectarivores across
all avian lineages. Compared to specialist avian nectarivores, we know far less about
the concentration-dependence of sugar preferences of avian frugivores (see Table 5.1).
5.5.3 Can we explain sucrose preference on concentrated diets?
A number of species have now been shown to switch to preference for sucrose solutions
at high diet concentrations (Table 5.1). Significant sucrose preference has been
somewhat puzzling, since these solutions have similar energetic value compared with
the hexose equivalents, and sucrose solutions require sucrose hydrolysis before
assimilation. As sucrose-dominant nectars tend to be more concentrated than

136
predominantly hexose nectars (Nicolson 1998), birds may prefer sucrose at high
concentrations and hexose at low diet concentrations as this reflects the pattern found in
natural floral nectars (Lotz and Schondube 2006).
It has also been suggested that the preference for sucrose on high sugar
concentrations could reflect taste preferences. By human tastes, fructose is 1.3x sweeter
than sucrose while glucose has only 0.7x the sweetness of sucrose (Harborne 1993).
Birds may also show discrimination in sugar tastes. A recent study demonstrated that
the broad-billed hummingbird perceives glucose, fructose and sucrose differently and is
able to detect fructose at ~30% lower concentrations than sucrose or ~20% lower than
glucose, indicating that fructose has a more intense flavour for this hummingbird
(Medina-Tapia et al. 2012). These authors suggested that hummingbirds were selecting
sugar solutions in relation to their relative sweetness, and that gustatory thresholds may
play an important role in determining sugar selection at least for more dilute diets
(Medina-Tapia et al. 2012). The role of taste in sugar type preference for concentrated
diets remains to be tested.
5.5.4 Do laboratory results reflect foraging preferences in the wild?
The three honeyeater species examined (New Holland and singing honeyeaters and red
wattlebirds) feed preferentially on sucrose nectars, avoiding hexose nectars; these
foraging data reflect their preferences for hexoses only on very dilute sugar
concentrations when tested in the laboratory. By contrast, rainbow lorikeets feed
predominantly on hexose nectars, avoiding sucrose nectars; again, these data reflect
preferences of these birds for hexoses at much higher sugar concentrations under

137
laboratory conditions. We have few foraging data for silvereyes to date, therefore it is
difficult to make any conclusions about their foraging preferences.
We have been limited by several constraints in our comparison between
laboratory sugar type preferences and foraging choices in the wild. Firstly, there are
very few data available on nectar sugars of Australian plants. While foraging
observations are identified to plant species, the nectar composition data for these same
plant species are often unavailable. We therefore present nectar composition data for
plant genera rather than species (even so, we still lack data on nectar composition data
for plant genera accounting for an average of 15% of foraging records for the five bird
species examined for this measure). Secondly, these bird species also forage widely at
plant species outside of Western Australia (we have not found comparative data of
foraging observations for the rest of the country). Finally, where nectar data are
available for multiple species of a plant genus, averaging values for sugar composition
obscures the fact that some genera (notably Banksia and Grevillea), show a dichotomy
in nectar composition, with some species having sucrose nectars and other species
hexose nectars (Nicolson and Van Wyk 1998). Many species included in this data set
(e.g. Grevillea spp.; Table 5.4) may be not be primarily bird-pollinated, although birds
may visit their flowers on an opportunistic basis.
5.5.5 Conclusions
In the Americas and Africa, nectar-feeding birds are relatively easily categorised as
specialised (hummingbirds and sunbirds, respectively) or generalist (all other bird taxa)
due to distinctions between bird lineages. However, there are ~180 species of

138
Australasian honeyeaters (Family Meliphagidae) which exhibit a range of diets, from
predominantly nectar through to predominantly insect diets. This makes a simplistic
dichotomy between specialised and generalist/opportunistic nectarivores difficult for
Australian honeyeaters.
We have identified that sucrase activity is likely to be a key digestive constraint
directly influencing the concentration-dependence of sugar type preferences shown in
birds. To our knowledge, this is the first study to compare sugar preferences assessed in
the laboratory with both aspects of digestive physiology and wild foraging observations.
We suggest that further comparative work on generalist and specialist nectarivores,
particularly in larger birds such as lorikeets, takes a similarly multi-faceted approach by
incorporating avian ecology and behaviour with digestive physiology.

139
6 MISTLETOEBIRDS AND XYLOSE: AUSTRALIAN FRUGIVORES
DIFFER IN THEIR HANDLING OF DIETARY SUGARS
Kathryn R. Napier1, Patricia A. Fleming
1 and Todd J. McWhorter
2
1 School of Veterinary and Life Sciences, Murdoch University, Murdoch, WA 6150,
AUSTRALIA
2 School of Animal and Veterinary Sciences, University of Adelaide, Roseworthy Campus, SA 5371,
AUSTRALIA
This chapter has been submitted to the journal Physiological and Biochemical Zoology
for consideration of publication.

140
STATEMENT OF AUTHOR CONTRIBUTION
K.R. Napier: Initiated the research, mist-netted and kept birds in captivity,
performed all laboratory experiments on mistletoebirds, silvereyes and singing
honeyeaters, collected, entered and analysed the data and wrote the manuscript.
P.A. Fleming: Assisted with the design and initiation of the research, assisted
K.R. Napier with mist-netting and paracellular absorption experiments, advised and
assisted K.R. Napier with statistical analyses, and provided editorial comments to
versions of the manuscript from draft to publication.
T.J. McWhorter: Assisted with the design and initiation of the research, advised
K.R. Napier with apparent assimilation efficiency, paracellular absorption, and HPLC
experimental methodologies, procedures and statistical analyses, and provided editorial
comments to versions of the manuscript from draft to publication.

141
6.1 ABSTRACT
Carbohydrate-rich mistletoe fruits are consumed by a wide range of avian species.
Small birds absorb a large portion of water-soluble nutrients, such as glucose, via the
paracellular pathway. D-xylose, a pentose monosaccharide, is abundant in some nectars
and mistletoe fruits consumed by birds, and it has been suggested that it is most likely
absorbed via the paracellular pathway in birds. We measured apparent assimilation
efficiency (AE*) and bioavailability (f) for D-xylose and D- and L-glucose in three
frugivorous Australian bird species. Mistletoebirds, silvereyes and singing honeyeaters
showed significantly lower AE* for D-xylose than for D-glucose. Across two diet
sugar concentrations, silvereyes and singing honeyeaters significantly increased f of
both L-glucose (a metabolically inert isomer of D-glucose commonly used to quantify
paracellular uptake) and D-xylose on the more concentrated diet, probably due to
increased gut processing time. By contrast, mistletoebirds (mistletoe fruit specialists)
did not vary f of either sugar with diet concentration. Mistletoebirds also showed higher
f for D-xylose than L-glucose and eliminated D-xylose more slowly than silvereyes and
singing honeyeaters, demonstrating differences in the handling of dietary xylose
between these species. Our results suggest that D-xylose may be absorbed by both
mediated and non-mediated mechanisms in mistletoebirds.

142
6.2 INTRODUCTION
Mistletoe fruit is used by a wide range of bird species, with at least 30 bird species
recorded feeding on Australian mistletoe fruits (Reid 1986; Reid 1989; Barker and
Vestjens 1990; Higgins et al. 2001). The composition of the fleshy fruit pulp varies
among species, but most mistletoe fruits are high in carbohydrates, lipids and protein
(Walsberg 1975; Herrera 1987; McPherson 1987; Witmer 1996; López de Buen and
Ornelas 2001; Barea 2008), with 24–74% of the dry mass comprising of soluble
carbohydrates (Godschalk 1983a; Lamont 1983b; Restrepo 1987; Snow and Snow
1988; Gedalovich-Shedletzky et al. 1989). The sugar composition varies between
species, with fructose, rhamnose, mannose, glucose, arabinose, galactose and xylose
present in varying amounts in four Santalaceae and Loranthaceae species studied to date
(Table 6.1, Gedalovich-Shedletzky et al. 1989; Azuma et al. 2000).
Xylose is a pentose monosaccharide that has been reported as one of the most
abundant sugars in the fruit of several species of the North American mistletoe families
Santalaceae and Loranthaceae and is present in one species of Australian Loranthaceae
mistletoe, Amyema preissii (see Table 6.1). Xylose is also abundant in nectar sugar of
two genera of Proteaceae, comprising up to 39% of the total nectar sugar by weight in
Protea and Faurea nectars examined (Nicolson and Van Wyk 1998). The high
concentration of xylose in these mistletoe fruits and floral nectars suggests that this
sugar could be an important dietary component for nectarivores and frugivores.
However, few vertebrates are able to tolerate high concentrations of xylose in food, and
it is unclear whether many vertebrates are able to directly metabolise it (Lotz and
Nicolson 1996; Franke et al. 1998; Jackson et al. 1998a; Jackson et al. 1998b). Xylose

143
appears to be absorbed to some degree in nectar feeding birds, indicated by reasonably
high apparent assimilation efficiencies (AE*) in two southern African species: the Cape
sugarbird (Promerops cafer, AE* ~53%) and the Cape white-eye (Zosterops pallidus,
AE* ~61%) (Franke et al. 1998; Jackson et al. 1998a; Jackson et al. 1998b), although
assimilation of sucrose and hexoses including glucose and fructose are still much higher
with AE* close to 100% in most nectarivores and frugivores tested to date (Lotz and
Schondube 2006; Fleming et al. 2008; Napier et al. 2008a). Some birds have been
shown to exhibit osmotic diarrhoea (resulting from unabsorbed xylose) after ingesting
pure xylose solutions (Lotz and Nicolson 1996; Jackson et al. 1998b; Jackson and
Nicolson 2002). While nectar feeding bird species tested to date have shown avoidance
of pure xylose solutions (Lotz and Nicolson 1996), a southern African mammal
pollinator, the Namaqua rock mouse, Aethomys namaquensis, freely consumed xylose
solutions with an AE* of 97% (Johnson et al. 1999). Xylose only has nutritional value,
however, if it is able to be absorbed and metabolised. The mechanisms of absorption of
this sugar are therefore of direct interest in interpreting how animals deal with the
presence of xylose in nectar and fruit.
There are two routes for sugar absorption at the intestine, via carrier-mediated
transport through the intestinal epithelial cells themselves (the transcellular route), or
diffusion between these cells (the paracellular route). Small birds and bats with reduced
intestinal absorptive surface area, as an adaptation to flight (Caviedes-Vidal et al. 2007),
absorb a large portion of their water-soluble nutrients (e.g. glucose) via the paracellular
pathway (reviewed by McWhorter 2005). Paracellular absorption provides a non-
saturable absorptive mechanism that matches absorption capacity to acute changes in
dietary nutrient concentration (Afik et al. 1997; Ferraris 2001). The relative

144
contribution of paracellular to total glucose absorption in nectarivorous birds increases
with an increase in diet sugar concentration, an effect which may be largely due to
increased digesta retention time of more energy dense nectars in the intestine
(McWhorter et al. 2006; Napier et al. 2008b). Frugivorous birds and bats also rely
extensively on the paracellular pathway for the absorption of glucose (Tracy et al. 2007;
McWhorter et al. 2010), but the effects of diet energy density and thus digesta transit or
residence times have not been assessed in frugivorous species to date.
The mechanism of absorption of xylose appears to vary amongst species, and
probably includes both paracellular and carrier-mediated components. There is
evidence that intestinal absorption of xylose in chickens, hamsters, rats, frogs, rabbits
and cows occurs at least in part by active carrier-mediated transport; it requires energy
expenditure and it is Na+-dependent and inhibited by phlorizin (Salem et al. 1965;
Alvarado 1966; Lassen and Csaky 1966; Heyman et al. 1980; Scharrer and Grenacher
2000). However in humans, xylose appears to be absorbed only via passive (i.e.
paracellular) diffusion (Ohkohchi et al. 1986; Fine et al. 1993). It has been suggested
that xylose is likely to be absorbed via the paracellular pathway in nectarivorous birds
(Jackson and Nicolson 2002), although no direct uptake measurements have been done.
We investigated the AE* of D-xylose (assumed to be absorbed by paracellular
mechanisms) and D-glucose (absorbed by both paracellular and transcellular pathways)
in three bird species: a mistletoe fruit specialist, a generalist frugivore and a generalist
nectarivore that also takes mistletoe fruit. We also explored the mechanism(s) of
absorption by assessing the bioavailability of radiolabelled L-glucose (an isomer of D-
glucose that is not metabolised and is absorbed only by the paracellular pathway) and

145
D-xylose. We predicted, firstly, that all three study species would exhibit high AE* for
D-glucose but lower AE* for D-xylose (as shown in previous studies, e.g. Lotz and
Nicolson 1996; Franke et al. 1998; Jackson et al. 1998b). Because L-glucose is only
absorbed via the paracellular pathway (Chang and Karasov 2004), a high bioavailability
of L-glucose is indicative of significant paracellular uptake of D-glucose. Therefore, we
further predicted that if D-xylose is also absorbed solely by the paracellular pathway,
both L-glucose and D-xylose bioavailability would increase with diet sugar
concentration.
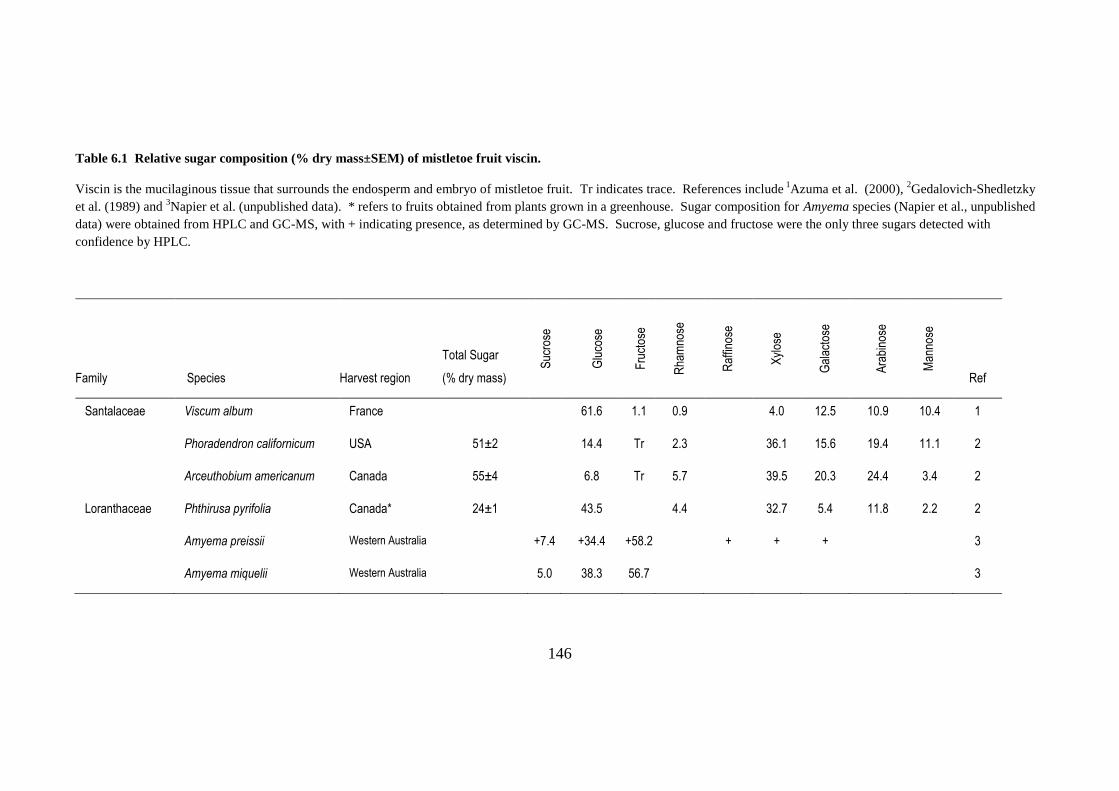
146
Table 6.1 Relative sugar composition (% dry mass±SEM) of mistletoe fruit viscin.
Viscin is the mucilaginous tissue that surrounds the endosperm and embryo of mistletoe fruit. Tr indicates trace. References include 1Azuma et al. (2000),
2Gedalovich-Shedletzky
et al. (1989) and 3Napier et al. (unpublished data). * refers to fruits obtained from plants grown in a greenhouse. Sugar composition for Amyema species (Napier et al., unpublished
data) were obtained from HPLC and GC-MS, with + indicating presence, as determined by GC-MS. Sucrose, glucose and fructose were the only three sugars detected with
confidence by HPLC.
Family Species Harvest region
Total Sugar
(% dry mass)
Suc
rose
Glu
cose
Fru
ctos
e
Rha
mno
se
Raf
finos
e
Xyl
ose
Gal
acto
se
Ara
bino
se
Man
nose
Ref
Santalaceae Viscum album France 61.6 1.1 0.9 4.0 12.5 10.9 10.4 1
Phoradendron californicum USA 51±2 14.4 Tr 2.3 36.1 15.6 19.4 11.1 2
Arceuthobium americanum Canada 55±4 6.8 Tr 5.7 39.5 20.3 24.4 3.4 2
Loranthaceae Phthirusa pyrifolia Canada* 24±1 43.5 4.4 32.7 5.4 11.8 2.2 2
Amyema preissii Western Australia +7.4 +34.4 +58.2 + + + 3
Amyema miquelii Western Australia 5.0 38.3 56.7 3

147
6.3 MATERIALS AND METHODS
6.3.1 Study species
Our study species were three Australian bird species that are known to ingest mistletoe
fruit (Chaffer 1966; Paton and Ford 1977; Bernhardt 1984; Forde 1986; Reid 1986).
The mistletoebird (Dicaeum hirundinaceum) is a specialised frugivore that feeds
primarily on mistletoe fruit (Richardson and Wooller 1988). The short, specialised
alimentary tract of the mistletoebird is typical of specialised frugivores; mistletoebirds
possess smaller and less muscular gizzards than insectivorous birds of similar sizes, and
the gizzard, proventriculus and duodenum are all in the same plane, which allows the
more direct and rapid processing of the large number of mistletoe fruit that are ingested
(Richardson and Wooller 1988).
The silvereye (Zosterops lateralis) is a generalist frugivore (Wilkinson 1931;
Thomas 1980; Richardson and Wooller 1986) and the singing honeyeater
(Lichenostomus virescens) is considered a generalist nectarivore (Collins and Morellini
1979; Richardson and Wooller 1986). The digestive tract of honeyeaters is less
specialised then that of the mistletoebird (Richardson and Wooller 1988). Silvereyes
and singing honeyeaters show no distinct morphological adaptations for frugivory, with
intestine lengths expected for nectarivorous birds of their size, and more muscular
gizzards that fall between those of specialist frugivores and specialist insectivores
(Richardson and Wooller 1986; Stanley and Lill 2002). The junction of the gizzard and
duodenum is also on a separate plane from that of the proventriculus and gizzard in
these species (Richardson and Wooller 1988).

148
The transit time of mistletoe seeds is shorter in specialist mistletoe feeders such
as the mistletoebird and painted honeyeater (Grantiella picta) than in more generalist
frugivores and honeyeaters. For example, the mean gut passage time of Amyema
quandong mistletoe seeds by the spiny-cheeked honeyeater (Acanthagenys rufogularis),
an opportunistic mistletoe fruit feeder, in Australia, is 40.57±0.60 (SE) min, nearly three
times longer than for the mistletoebird at 13.67±0.48 (SE) min (Murphy et al. 1993) and
nearly twice as long as painted honeyeaters at 24.43±1.27 (SE) min (Barea 2008).
Keast (1958) also reported longer voiding times in silvereyes feeding on A. miquelii and
A. qaudichaudii fruits (30 to 80 min) compared to mistletoebirds (25 to 60 min). Seed
transit times for silvereyes feeding on Coprosma quadrifida and Rhagodia parabolica
fruits (slightly smaller than A. preissii fruit) average 18 and 31.5 min, respectively
(French 1996; Stanley and Lill 2002).
Silvereyes (body mass; mb 9.0±0.4 g) and singing honeyeaters (mb 28.9±4.1 g)
were captured on the grounds of Murdoch University, Perth, Western Australia (WA;
32°04′S, 115°50′E) by mist-netting in May 2009 and January 2010, respectively. There
is no measure of sexual dimorphism by plumage in either species and we did not
conduct genetic sexing, so gender is not known for these species. Mistletoebirds (mb
8.7±0.6 g) were captured on a private property at York, WA (31°50′S, 116°44′E), in
December 2010 and January and March 2011. All birds were acclimatised to outdoor
captive conditions (see below) for at least two weeks before the commencement of
experimental trials. All three species were fed a maintenance diet of Wombaroo ®
nectarivore mix (Wombaroo Food Products, Glen Osmond, South Australia), which
contains sucrose as the main sugar type, supplemented with additional sucrose or equal
parts of glucose and fructose for a total sugar content of ~25% w/w dry matter.

149
Silvereyes and mistletoebirds were also fed a variety of fleshy fruits daily (e.g. mistletoe
fruit, watermelon, grapes, apricots).
Birds were housed in individual outdoor aviaries (116 x 160 x 210 cm), and
confined to individual cages (47 x 54 x 41 cm) within the aviary for apparent
assimilation efficiency (AE*) and gut processing time (GPT) experiments. During
pharmacokinetic experiments, birds were housed individually in opaque plastic cages
(42 x 54 x 50 cm) with an automatic lighting regime (12:12 h), and a one way mirror to
minimise disturbance during sample collection. Excreta were collected from waxed
paper which was rolled through slots in the bottom of the cage, allowing samples to be
collected immediately upon defecation.
All animal care and experimental procedures were approved by the Murdoch
University Animal Ethics Committee (approval R2175/08).
6.3.2 Apparent assimilation efficiency (AE*):
Seven silvereyes, 8 singing honeyeaters and 5 mistletoebirds (3 male, 2 female) fed ad
libitum from D-glucose:D-xylose (4:1) solutions at three concentrations (0.25, 0.5, 1
mol·L-1
total sugar) for 24 h, through inverted, stoppered syringes. Each bird fed from
each solution with sugar concentration randomised and trials commencing within 30
min after sunrise (0500 to 0716 WST). Maintenance diet was removed 1 h before
sunrise to ensure all ingested food was voided before trials commenced. Trays were
placed under experimental cages to collect excreta and small containers of liquid
paraffin were placed directly below feeders to collect any diet spilt. Food intake was
recorded over 24 h by weighing feeders (0.01 g). Dried excreta were reconstituted with

150
a recorded amount of distilled rinse water. AE* was estimated separately for glucose
and xylose as:
AE* = (sugarin-sugarout) / (sugarin) (eq. 6.1)
where sugarin (mg) is the concentration (mg·ml-1
) of sugar in the ingested diet multiplied
by the volume of solution ingested (ml), and sugarout (mg) is the sugar concentration
(mg·ml-1
) in the total volume of excreta plus rinse water (ml).
Glucose assays: Two replicates of each excreta sample (100 µl) were incubated
at room temperature (~21 °C) for 15 min with 500 µl of hexokinase-glucose-6-
phosphate dehydrogenase enzymatic assay reagent (G3293, Sigma Aldrich, Castle Hill,
NSW, Australia). Standard ‘reagent blanks’ were included for each excreta sample.
Absorbance was then measured at 340 nm relative to distilled water by
spectrophotometry (UV mini 1240, Shimadzu Scientific Instruments, Balcatta, WA,
Australia).
Xylose assays: Two replicates of each excreta sample (50 µl) and two standard
‘reagent blank’ samples were incubated at room temperature (~21 °C) for 10 min with
200 µl of TEA/MgCl2 buffer solution, 200 µl of NAD+/ATP solution and 10 µl of
hexokinase suspension (D-xylose assay kit, Megazyme International, Wicklow, Ireland)
in order to remove any D-glucose present in the sample. Absorbance (A1) was then
measured at 340 nm relative to distilled water by spectrophotometry. The xylose
detection reaction was then initiated by the addition of 10 µl of β-xylose
dehydrogenase/xylose mutarotase solution and incubated for 10 min. Absorbance (A2)
was then measured at 340 nm relative to distilled water by spectrophotometry, with the
absorbance difference calculated (A2-A1).

151
6.3.3 Pharmacokinetic experiments
Bioavailability of L-glucose and D-xylose was measured using [14
C] and [3H]
radiolabelled L-glucose (180.16 g mol-1
) and D-xylose (150.13 g mol-1
), administered
orally and by intramuscular (IM) injection to 8 silvereyes, 8 singing honeyeaters and 4
mistletoebirds (2 male, 2 female) as described below. Oral and IM trials were
performed in separate experiments. To vary food intake rate, birds received two diet
sugar concentrations (0.25 and 1 mol·L-1
hexose solutions) in separate feeding
experiments. Both the order of trials and treatments given were randomly assigned, and
followed published protocol (McWhorter et al. 2006; Napier et al. 2008b).
Bioavailability (f) was calculated as:
F = (P·S·Kel) / I (eq. 6.2)
where P is the steady-state feeding concentration of radiolabelled sugars in plasma
(d.p.m.·mg-1
of plasma); S is the probe distribution space of radiolabelled sugars in
plasma (mg of plasma); Kel is the elimination rate constant for the removal of
radiolabelled sugars from plasma and its excretion in urine (min-1
); and I is the ingestion
rate of radiolabelled sugars (d.p.m.·min-1
) (Karasov and Cork 1994; McWhorter et al.
2006; Napier et al. 2008b).
For IM administration, each bird was injected into the pectoralis muscle with
~40 μl of a solution of 175 mmol·L-1
NaCl containing 500 KBq [14
C]-L-glucose per mg
mb, or 2,220 KBq [3H]-D-xylose per mg mb. The osmolality of the IM injection solution
was controlled at approximately 350mmol·kg-1
, so that the solution was isosmotic with
avian blood (Goldstein and Skadhauge 2000). After IM administration, excreta were

152
collected continuously for ~2 h, followed by collection of a small blood sample from
the brachial vein. The parameters for the mono- and bi-exponential models were
derived for each individual by non-linear curve fitting of the concentration of
radiolabelled sugars in excreta after IM administration versus time, by use of the
Marquardt-Levenberg algorithm (Marquardt 1963). For oral administration, birds fed in
separate trials from a hexose solution containing radiolabelled sugars ad libitum for ~3
h (3H-D-xylose: 120 KBq·ml
-1 and 200 KBq·ml
-1;
14C-L-glucose: 60 KBq·ml
-1 and 90
KBq·ml-1
for 0.25 and 1 mol·L-1
hexose diets respectively). One small blood sample
was collected 3 h after introduction of the radiolabelled diet; 4 silvereyes, 8 singing
honeyeaters and 4 mistletoebirds (3 male, 1 female) reached steady-state with regard to
radiolabel ingestion and excretion by 90 min, after returning to steady-state feeding
within 30 min after introduction of the radiolabelled diet (data not shown). The final
analyses were conducted on these individuals only, as four silvereyes and one
mistletoebird did not return to steady-state feeding despite repeated attempts at oral
trials, violating this assumption (Napier et al. 2012).
6.3.4 Gut passage times (GPT)
Eight silvereyes, 8 singing honeyeaters and 5 mistletoebirds (3 male, 2 female) were
offered fresh, ripe mistletoe fruits on branches, either A. miquelii (singing honeyeaters)
or A. preissii (mistletoebirds and silvereyes), depending on time of year and seasonal
availability (the bird species were not held simultaneously due to space and logistic
constraints). Birds were then observed via video camera, and the GPT of mistletoe
fruits was determined by recording the time of ingestion and defecation of mistletoe
fruits and calculating the difference; it was assumed that fruits were defecated in the

153
order they were ingested (Karasov and Levey 1990; Murphy et al. 1993; Witmer 1994).
Trials commenced within 1 h after sunrise (0500 to 0716 WST) and lasted for up to 3 h,
until all ingested seeds had been defecated. Maintenance diet was removed 1 h before
sunrise to increase appetite for the mistletoe fruit (Murphy et al. 1993). GPT was
calculated for each fruit defecated during the feeding trial by viewing of the video
footage, with up to two trials per bird conducted on separate days.
6.3.5 Statistical analysis
Proportional AE* data were arcsine square root transformed (Zar 1999) before analysis.
Differences in AE*, total sugar intake and pharmacokinetic parameters between sugar
type, sugar concentration and species were assessed by 3-way RM-ANOVA and 2-way
RM-ANOVA with sugar concentration and sugar type as repeated measures with
Tukey-Kramer post hoc tests for unequal sample sizes as required. Data are reported as
mean ± 1 SD throughout. Statistical analyses were performed with Statistica (StatSoft
Inc 2007) and SPSS (SPSS, Inc, Chicago, IL, USA). Statistical significance was
accepted for α<0.05.
6.4 RESULTS
6.4.1 Apparent assimilation efficiency (AE*)
All three species modulated their sugar intake with changes in diet concentration (i.e.
exhibited compensatory feeding), ingesting the same mass of sugar irrespective of diet
concentration (3-way RM-ANOVA; sugar concentration: F4,68=0.605, P=0.660). A
separate analysis examining AE* revealed a significant three-way interaction term

154
(sugar concentration x sugar type x species; F4,34=3.1, P=0.029) which indicated that the
three bird species were handling the sugars differently. Glucose AE* was higher than
xylose AE* (sugar type: F1,17=733.8, P<0.001). Glucose AE* was extremely high
(averaged over all concentrations: mistletoebirds: 99.77±0.39%; silvereye:
99.85±0.14%; singing honeyeater: 99.85±0.14%) and did not differ with species or diet
concentration (Fig. 6.1; post hoc analyses shown). Xylose AE* ranged from
56.08±8.10 to 78.3±4.07% (Fig. 6.1). Diet concentration did not have a significant
effect on xylose AE* for silvereyes, but singing honeyeaters assimilated significantly
less xylose on the 1 mol·L-1
diet compared to the 0.25 mol·L-1
diet, and mistletoebirds
assimilated significantly less xylose on the 0.25 mol·L-1
diet compared to the 0.5 mol·L-1
diet (Fig. 6.1).

155
Mistletoebird Silvereye Singing honeyeater
Ap
pa
ren
t a
ssim
ilatio
n e
ffic
ien
cy (
AE
*, %
)
0
20
40
60
80
100
0.25 mol·L-1
D-xylose
0.5 mol·L-1
D-xylose
1 mol·L-1
D-xylose
0.25 mol·L-1
D-glucose
0.5 mol·L-1
D-glucose
1 mol·L-1
D-glucose
d
b,c
b,c,d
b,c
b,c,db,c,d
b
b,c,d
c,d
a a a a a aaaa
Figure 6.1 Apparent assimilation efficiency (AE*) of D-xylose (solid) and D-glucose (lines).
Birds were fed solutions containing 80% D-glucose and 20% D-xylose at three total sugar concentrations
(0.25 (white), 0.5 (light grey) and 1 mol·L-1
(grey)) for mistletoebirds, silvereyes and singing honeyeaters.
Values are presented as mean ± 1SD; mistletoebirds (n=5), silvereyes (n=7), singing honeyeaters (n=8).
Superscripts refer to results of Tukey-Kramer post hoc tests for unequal sample sizes (3-way RM-
ANOVA).

156
6.4.2 Pharmacokinetic experiments
Pharmacokinetic data were available for all singing honeyeaters (n=8) tested, but only
for four out of the original eight silvereyes and four of the original five mistletoebirds,
due to silvereyes and one mistletoebird failing to recommence steady-state feeding
within a sufficient timeframe. These individuals were therefore excluded from
pharmacokinetic analyses due to the violation of steady-state feeding conditions (Napier
et al. 2012). Over the three hour trial period, birds drank approximately three times the
volume of the dilute diet (0.25 mol·L-1
) compared to the more concentrated diet (Table
6.2). The mean steady-state concentration (P) of L-glucose and D-xylose in plasma was
relatively high in all species, indicating significant absorption of both compounds
(Table 6.2). The elimination of both L-glucose and D-xylose after IM injection did not
fit a bi-exponential model significantly better than a mono-exponential model for all
individual birds of all species (glucose: F<3.29, P>0.1; xylose: F<4.49, P>0.06),
indicating single compartment kinetics. Xylose was eliminated faster than glucose in
singing honeyeaters, but the opposite was the case for mistletoebird (2-way RM-
ANOVA: Kel; sugar type x species; F2,13=240, P<0.001, Table 6.2). Probe distribution
space (S) did not differ between glucose and xylose for mistletoebirds and silvereyes but
did for singing honeyeaters (Table 6.2).
In terms of bioavailability (f) of the two sugars, there was a significant species
by sugar type interaction (3-way RM-ANOVA; sugar type x species: F2,13=7.65,
P=0.006) indicating the three species handled glucose and xylose differently; we
therefore analysed the data for each species separately by two-way RM-ANOVA (sugar
and diet concentration as the repeated-measures factors). Mistletoebirds did not vary

157
either glucose or xylose f with diet concentration (concentration: F1,3=0.006, P=0.941),
while the other two species did (silvereyes concentration: F1,3=97.69, P=0.002; singing
honeyeaters concentration: F1,7=33.38, P=0.001; see Fig. 6.2). Furthermore,
mistletoebirds showed higher f for xylose compared with glucose, while the opposite
pattern was observed for singing honeyeaters (no significant effect for silvereyes, see
Fig. 6.2).

158
Table 6.2 Parameters used to determine bioavailability (f) of radiolabelled L-glucose and D-xylose in mistletoebirds, silvereyes and singing honeyeaters at two diet
concentrations (0.25 and 1 mol·L-1
hexose solutions).
Values are presented as mean ± 1 SD. Feeding rate: mistletoebirds and singing honeyeaters were presented both L-glucose and D-xylose in a single solution with double-labelled
isotopes while silvereyes were fed L-glucose and D-xylose in separate feeding trials. Superscripts refer to significant differences obtained from 2-way RM-ANOVA (between
species: Kel and S; within species: f) and Tukey post hoc tests for unequal sample sizes. †calculated only once for each bird while feeding on 0.25 mol·L-1 hexose solution.
Mistletoebirds (n=4) Silvereyes (n=4) Singing honeyeaters (n=8)
L-glucose D-xylose L-glucose D-xylose L-glucose D-xylose
Feeding rate (mg·min-1) 0.25: 48.44±8.14
1: 16.08±6.93
0.25: 48.44±8.14
1: 16.08±6.93
0.25: 26.99±4.95
1: 5.72±2.66
0.25: 26.42±9.55
1: 9.611±3.61
0.25: 65.16±32.97
1: 22.28±5.78
0.25: 65.16±32.97
1: 22.28±5.78
Intake rate, I (d.p.m.·min-1) 0.25: 76215±12811
1: 43132±40343
0.25: 86764±14583
1: 63568±27376
0.25: 67002±16493
1: 16289±6503
0.25: 99876±52152
1: 25860±8544
0.25: 597098±30308
1: 218258±59577
0.25: 61976±30509
1: 31294±9709
Steady-state plasma, P
(d.p.m.·mg-1 plasma)
0.25: 282±156
1: 101.89±56
0.25: 832±156
1: 658±256
0.25: 707±160
1: 500±193
0.25: 1310±465
1: 653±291
0.25: 2335±667
1: 1668±456
0.25: 157±56
1: 142±63
Elimination constant, Kel (min-1) † 0.044±0.005a
0.014±0.003b,c
0.021±0.009 b,c
0.025±0.009 b,c
0.014±0.009 c
0.022±0.005 b
Probe distribution space, S
(mg plasma) †
2100±1062b
4015±1772a,b
1456±590 b
1631±1117 b
6268±2243a
3631±2033 b
Bioavailability, f (%) 0.25: 32.88±19.72 b
1: 32.92±27.84 b
0.25: 51.98±23.46 a
1: 54.15±14.75 a
0.25: 29.70±12.69 d
1: 81.81±17.23 a,b
0.25: 53.31±16.24 b,d
1: 89.44±12.07 a
0.25: 36.53±13.35 b
1: 65.22±21.93 a
0.25: 25.06±17.13 d
1: 36.86±14.89 b

159
Mistletoebirds Silvereyes Singing honeyeaters
Bio
availa
bili
ty (
f, %
)
0
20
40
60
80
100
0.25 mol·L-1
L-glucose
1 mol·L-1
L-glucose
0.25 mol·L-1
D-xylose
1 mol·L-1
D-xylose
b
b
a
a
a
d
b,d
a,b
a
b
d
b
Figure 6.2 Bioavailability (f, %) of radiolabelled L-glucose (white) and D-xylose (grey) at two diet
concentrations (0.25 and 1 mol·L-1
, solid and lines respectively).
Values are presented as mean ± 1 SD; mistletoebirds (n=4), silvereyes (n=4), singing honeyeaters (n=8).
Superscripts refer to significant differences between diets within species (2-way RM-ANOVA with
Tukey-Kramer post hoc tests for unequal sample sizes).

160
6.4.3 Gut passage times (GPT)
Mistletoebirds were the only species to ingest mistletoe fruits whole, after removing the
exocarp. None of the 8 singing honeyeaters ingested mistletoe fruit over their two 3-
hour trials (i.e. 6 h observations for each individual bird). One of 8 silvereyes ingested
small amounts of A. preissii fruit pulp (2 out of 14 available fruits; in one trial) by
prising open the exocarp of the fruit using its beak in a pliers-like motion to open a
small hole into a wider split that it could feed from, but did not consume any seeds.
When the seed was removed from the exocarp, the bird stuck it to the mistletoe branch
and continued to eat the fruit pulp.
The average GPT of A. preissii ingested by mistletoebirds was 16.45±7.21 min,
and ranged from 2.0 to 40.0 min (n=177, total number of trials n=8). The number of
fruits presented to individual birds ranged from 22 to 32, with 84–100% ingested by
birds and 0–16% of fruits manipulated, but dropped before consumption. The average
feeding period (71.99±16.73 min) ranged from 49.0 to 98.0 min, while the total trial
period (last defecation-first feeding) averaged 87.95±18.89 min (ranging from 54.0 to
109.0 min). The average number of fruits in the birds intestine over the total trial period
was 5.49±2.49 (range from 1–12). The average number of seeds defecated over the
total trial period was 3.08±1.03 (range from 1–5).
6.5 DISCUSSION
Xylose was only first reported as a nectar sugar in southern African Proteaceae by Van
Wyk and Nicolson in 1995, but we know very little about its prevalence and presence in
other plant nectars across the globe. Xylose is also evident in the fruits of some

161
mistletoe species (Table 6.1 and references therein). Xylose may therefore be more
prevalent in nectar and fruit diets for birds than we currently are aware of, and birds that
have evolved with a high-xylose diet are likely to have developed mechanisms of
handling this sugar. Mistletoe birds clearly process xylose differently from the other
two frugivores tested in this study. While there was no clear pattern between the
species in terms of the AE* of xylose, the three species handled the absorption of
glucose and xylose differently: silvereyes and singing honeyeaters showed significantly
greater bioavailability for both sugars on the more concentrated diet, but mistletoebirds
did not vary f with diet sugar concentration. Furthermore, mistletoebirds showed higher
f for D-xylose compared with L-glucose; while the opposite pattern was observed for
the singing honeyeaters (no significant effect for silvereyes). We discuss these findings
firstly in terms of the assimilation of these sugars, followed by their mechanisms of
absorption.
6.5.1 Apparent assimilation efficiency (AE*)
All three species assimilated significantly less D-xylose than D-glucose. As
mistletoebirds consume mistletoe fruit (previous studies reveal several species of
mistletoe contain large quantities of xylose sugar, Table 6.1 and references therein),
these birds should have been better able to deal with xylose than the other two bird
species. However we did not find this, as mistletoebirds did not demonstrate higher D-
xylose AE* values then the other two species.
All three species exhibited compensatory feeding, that is, they were able to
adjust their intake rate so they ingested a similar total mass of sugar over the range of

162
sugar concentrations. Sugar concentration did not affect D-xylose AE* in silvereyes,
but varying effects of concentration were demonstrated in mistletoebirds and singing
honeyeaters, with no clear discernable pattern. The AE* experiments integrate
digestive function over a longer time period in comparison to the pharmacokinetic
measures of bioavailability, and so may be more sensitive to variation in feeding rate
and frequency than the latter. It is also possible that varying amounts of microbial
degradation of xylose in the excreta collection pans may contribute to this variation, but
the absolute causes of the varying effects of diet concentration between bird species on
the D-xylose AE* values are unclear.
We also found slightly higher AE* of D-xylose in our study species (56–78%)
compared to other bird species studied to date (53–61%) (Franke et al. 1998; Jackson et
al. 1998b), but much lower than the Namaqua rock mouse (~97%) (Johnson et al.
1999), suggesting that, like other bird species studied, xylose is not absorbed efficiently
by these Australian birds.
6.5.2 Mechanism of sugar absorption
Silvereyes and singing honeyeaters showed greater bioavailability of both L-glucose
and D-xylose on the more concentrated diet, where slower gut passage time would
promote increased absorption via the paracellular route (likely due to increased contact
time of digesta with absorptive surfaces), as has been previously described in three
specialised nectarivores to date (McWhorter et al. 2006; Napier et al. 2008b). On the
other hand, mistletoebirds did not vary f with diet concentration for either sugar. This
could suggest that absorption of these compounds known to be absorbed by non-

163
mediated (i.e. paracellular) mechanisms is somehow decoupled from digesta retention
time in this species. Although we were unable to directly compare the transit rates of
mistletoe fruit between our study species, mistletoebirds have previously demonstrated
much faster transit rates of mistletoe fruit through their highly specialised intestinal
tracts compared to silvereyes and honeyeaters (Keast 1958; Murphy et al. 1993; French
1996; Stanley and Lill 2002; Barea 2008). We are not aware of any measurements of
digesta retention time in mistletoebirds feeding on liquid diets, but it is highly likely that
mistletoebirds, like many other species (Lopez-Calleja et al. 1997; Levey and Martinez
del Rio 1999; McWhorter and Lopez-Calleja 2000; McWhorter et al. 2006; Wilson and
Downs 2011), do vary retention time with energy density. A high capacity for mediated
glucose uptake (not quantified in this study) might mean that mistletoebirds can meet
their energy needs without an increased reliance on paracellular uptake. Indeed, the
significantly lower f of L-glucose for mistletoebirds in comparison to silvereyes and
singing honeyeaters suggests a decreased reliance on the paracellular pathway in
mistletoebirds. One other specialised frugivore, the cedar waxwing, has three times the
active transport of glucose compared to the American robin (Karasov and Levey 1990).
This allows cedar waxwings to maintain a high and efficient absorption of sugars, even
at relatively high intake and processing rates of fruit (Witmer and van Soest 1998). If
mistletoebirds have relatively high capacity for mediated carbohydrate uptake, digesta
retention time may vary less with energy density than in other species studied,
providing a potential mechanism to explain the apparent decoupling of f and diet sugar
concentration in this species.
The paracellular space discriminates according to molecular size, similar to a
sieve (Friedman 1987; Chediack et al. 2003). Therefore, if D-xylose and L-glucose

164
were both absorbed via the paracellular pathway, we would expect higher f values for
D-xylose based on its lower molecular mass (Chediack et al. 2003). Due to these
differences in molecular mass between D-xylose and L-glucose and the fact that
diffusion in water declines with molecular weight (MW0.5
), the bioavailability values of
D-xylose can be corrected by a decrease of 8.7% [(180.160.5
-150.130.5
)/180.160.5
)*100]
to assess the effects of molecule size on absorption. Mistletoebirds were the only
species to demonstrate a significantly higher f for D-xylose than L-glucose, and this
difference is not solely accounted for by differences in molecular mass (i.e. average D-
xylose f values are >8.7% higher than L-glucose, Table 6.2). This suggests that xylose
could possibly be absorbed by both mediated and paracellular routes in these birds-
possibly in the same manner as D-glucose (i.e. both paracellular and carrier-mediated
active transport). Further work is required to determine if xylose is actively transported
by membrane proteins in these birds.
Evidence also suggests that in some other animals, D-xylose is actively
transported along with D-glucose, although the affinity of the transporter for D-xylose is
much lower than that of D-glucose (Salomon et al. 1961; Bihler et al. 1962; Csaky and
Lassen 1964; Alvarado 1966; Ohkohchi and Himukai 1984). In chickens, Savory
(1992) reported D-xylose was absorbed slower than D-glucose and D-galactose, but
faster than D-arabinose and D-mannose, which concurs with results previously reported
in chicks (Wagh and Waibel 1967), rats (Kohn et al. 1965) and humans (Wood and
Cahill 1963); suggesting that the absorption rates of these sugars may depend on their
relative contributions of active and passive (transcellular and paracellular) transfer
mechanisms (Savory 1992).

165
6.5.3 Metabolism of xylose
Xylose is only beneficial as a source of energy to these birds if it is able to be
metabolised, either by gastrointestinal microbes or directly by bird tissues (discussed in
depth by Jackson and Nicolson 2002). Animals such as the Namaqua rock mouse,
through its caecal fermentation chamber in its gut, are able to ferment xylose to convert
it into a source of energy (Johnson et al. 2006). The caecal enlargement demonstrated
in chicks fed xylose suggests a similar process of bacterial fermentation (Schutte 1990;
Schutte et al. 1992). While frugivorous and nectarivorous birds like mistletoebirds and
honeyeaters tend to have rudimentary or small caeca (Richardson and Wooller 1986;
Richardson and Wooller 1988), the presence of intestinal microbes (that may be able to
metabolise xylose), has not been studied in these birds. Xylose is also able to be
catabolised by certain mammalian tissues, with mammalian cells demonstrated to be
able to survive with xylose or xylitol as their sole energy source (Demetrakopoulos and
Amos 1978), but to date, this has not been investigated in birds.
The presence of xylose in nectar and fruit remains puzzling due to the aversion
shown by birds and rodent pollinators (Jackson and Nicolson 2002). Although the
Namaqua rock mouse is able to efficiently absorb and metabolise xylose, it is the least
preferred sugar in preference tests (Johnson et al. 1999). A co-evolutionary explanation
for xylose as an attractant for pollinators and dispersers therefore remains contentious
(Jackson and Nicolson 2002), and the study of the potential catabolism of xylose by
intestinal microbes or systemic catabolism in birds certainly warrants further
investigation.

166
6.5.4 Conclusions
Mistletoebirds showed higher bioavailability for xylose compared with glucose; exactly
the opposite pattern that was observed for singing honeyeaters (data for silvereyes were
not statistically significant). This implies that mistletoebirds may be absorbing D-
xylose by both mediated and non-mediated mechanisms. Mistletoebirds also eliminated
xylose more slowly than silvereyes and singing honeyeaters, suggesting that xylose may
have been incorporated into cells or used in biochemical pathways in mistletoebirds; it
might also reveal a delay due to xylose passing through the gut enterocytes on its way to
the circulation.
The observation that mistletoebirds did not vary f with diet sugar concentration,
unlike silvereyes and singing honeyeaters, is also very intriguing – while possible
explanations may include the decoupling of retention time and absorption of compounds
absorbed by non-mediated mechanisms, a decreased reliance on the paracellular
pathway, relatively less modulation of digesta retention time with diet energy density,
or the presence of intestinal microbes in this species, we did not measure digesta mean
retention time, mediated carbohydrate uptake or quantify the intestinal microbiome of
birds in the present study.
These data build upon our understanding of the handling of sugars in
frugivorous Australian birds, with new insight in particular to a specialised frugivore.
While mistletoebirds do not assimilate more xylose than the more generalist frugivores
assessed in this study, they may absorb xylose differently. These three species therefore
reveal differences in how they handle dietary sugars.

167
7 GENERAL CONCLUSIONS
Australian mistletoes are a fascinating example of biological diversity, and are
considered a keystone species due to the key role they play in ecosystems. There is an
extremely diverse community of birds, ants, butterflies and mammals that live around
mistletoes and their hosts, with mistletoe associations ranging from the highly specific
relationships between mistletoes and their hosts and the specialised dispersers of
mistletoe fruit, to the more general relationships between opportunistic feeders of
mistletoe fruit, nectar and leaves.
The main aim of this thesis was to use a multi-faceted approach in the
investigation of the interactions between two mistletoe species, Amyema miquelii and A.
preissii, their hosts, and their avian consumers in the south-west of Western Australia
(WA). This was achieved through intensive field surveys and sampling, followed by a
series of laboratory studies assessing various aspects of avian digestive physiology. By
using a variety of experimental and analytical techniques, including field surveys, stable
isotopes analysis, multivariate analysis and pharmacokinetic modelling, this thesis has
provided new insight into the relationships between mistletoes and their hosts, and the
food resources mistletoes provide to a mistletoe specialist, the mistletoebird.
7.1 MISTLETOE-HOST INTERACTIONS
The mistletoe story presented in this thesis began with a study of the parasitic and host
relationship that determines how Australian mistletoes obtain their water and nutrients
(Chapter 2). Unlike other mistletoe species studied to date, A. miquelii obtained little

168
nitrogen from its nitrogen-poor eucalypt hosts, while A. preissii obtained more nitrogen
from its nitrogen-fixing (and therefore nitrogen-rich) acacia hosts. This study also
demonstrated that a greater differential in water use efficiency was required for
mistletoes on nitrogen-poor eucalypt hosts to extract nitrogen, resulting in more
negative differences in δ13
C in comparison to mistletoes on acacia hosts. The
investigation of the degree of heterotrophy in Australian mistletoes would also be a
valuable contribution to this area of research. The future investigation of proportional
heterotrophy requires a back-to-basics approach, however; greenhouse-based pot-
culture experimental methodologies that measure gas exchanges and biomass and
control for water availability, as well as investigations into the assumptions such as
estimated carboxylation factors and confounding effects of identical carbon isotope
discrimination, are recommended (Bannister and Strong 2001; Tennakoon et al. 2011).
7.2 MISTLETOE-ANIMAL INTERACTIONS
This thesis then transitioned into the investigation of the importance of mistletoe as a
food resource to bird communities in south-west WA, with particular focus on the
mistletoebird. The presence of fruiting mistletoe was associated with significant
changes in bird community structure, and mistletoebirds were more likely to be
recorded during months when ripe mistletoe fruit was present (Chapter 3). The use of
stable isotopes allowed the estimation of the proportional contribution of mistletoe fruit
to the diet of mistletoebirds, which ranged from 33% to 55% across the three
sites/sampling times tested (Chapter 4). The low nitrogen content of mistletoe fruit
(0.89–2.17%) is significantly less then that of arthropods (11.09–13.10%).

169
Mistletoebirds therefore supplement their diet with arthropods, and appear to increase
the dietary contribution of arthropods during breeding and moulting periods.
One limitation of the study of the dietary contributions of arthropods and
mistletoe fruit to mistletoebirds detailed in Chapter 4 is the limited sample sizes of both
dietary sources and mistletoebird tissues, particularly at the Araluen site. Mistletoebirds
proved difficult to mist-net in large numbers, with only two individuals mist-netted and
sampled at Araluen. I was unable to attempt mist-netting and sampling at two of the
five original study sites due to the extreme height of mistletoe in the canopy (Paruna),
and the overlapping availability of ripe fruit between Paruna and York 2 meant that field
work could only be conducted at one site (York 2). Cold weather conditions also
contributed to the decreased diversity of arthropods sampled at Araluen. Nevertheless,
this study is the first attempt to estimate the diets of mistletoebirds at more than one site
and at different times throughout the year through the use of dual isotopes (δ13
C and
δ15
N) and multiple avian tissues, and also takes into account the variation in dietary
sources and differences in the elemental concentration of isotopically distinct dietary
sources.
The nitrogen content of mistletoe fruit sampled in this study from York 1 (A.
preissii, 1.12±0.27%) and York 2 (A. miquelii, 0.89±0.25%) is similar to other bird
dispersed fruits (0.90±0.57%, Levey et al. 2000), and also to another Australian
Amyema species (A. quandang, 0.7%, Barea 2008). The higher nitrogen content of A.
preissii fruits sampled from Araluen (2.17±3.00%) is intriguing; while A. preissii,
parasitising nitrogen-rich acacia hosts, exhibit higher foliar nitrogen contents compared
to A. miquelii (parasitising nitrogen-poor eucalypt hosts, Chapter 2), there is no clear

170
pattern in fruit nitrogen content. Considerable variation in the nutritional content of
fruit, due to environmental and genetic factors, has previously been demonstrated within
species (Herrera 1987; Obeso and Herrera 1994; Izhaki et al. 2002), but further research
is required to determine if the differences demonstrated within A. preissii in this study
are due to local abiotic factors.
The specialisation to a diet consisting of high carbohydrate, low protein fruits
and nectar is associated with relatively low nitrogen requirements (Witmer 1998;
Schondube and Martinez del Rio 2004; Tsahar et al. 2006). This, combined with high
intake rates of mistletoe fruit and specialised intestinal morphology, has been
hypothesised to allow mistletoebirds to rely on mistletoe fruit as a dietary source of
protein (Barea and Herrera 2009). It would have been particularly valuable to have
quantified the nitrogen requirements of mistletoebirds (i.e. minimal nitrogen
requirements, MNR), which would be beneficial for future research (see ‘future
directions’). If mistletoebirds maintain the general pattern observed in other frugivores
(and specialised nectarivores) of having lower than expected nitrogen requirements
compared with generalist-feeding bird species, the physiological mechanisms which
they use to be able to survive with such low nitrogen requirements would also be of
great interest.
7.3 PHYSIOLOGICAL ADAPTATIONS TO FRUGIVORY
This mistletoe story concluded with the study of various digestive adaptations
associated with consuming a fruit diet in three focal avian species that consume fruit to
various degrees in their diet. Like the majority of bird-consumed fruits, fruit pulp from
mistletoe species studied to date contains low amounts of, or no, sucrose (Chapter 6).

171
For the mistletoe species analysed to date, the sugar composition of mistletoe fruit pulp
varies, with glucose and xylose the most prevalent sugars. The two Australian mistletoe
species studied in this thesis appear to differ from published studies of other mistletoe
species due to the high levels of fructose present in mistletoe fruit pulp (with the caveat
that the HPLC analysis performed was not sensitive enough to detect prevalence or
concentrations of sugars other than sucrose and hexoses). Mistletoebirds demonstrated
comparable values for the assimilation of sucrose and hexoses to both the more
generalist feeders in this study (silvereyes and singing honeyeaters) and other
specialised nectarivores studied to date, with apparent assimilation efficiency (AE*)
values >98% for sucrose, glucose and fructose. While intestinal enzyme data were not
available for mistletoebirds due to ethical reasons, the study detailed in Chapter 5 is the
first to demonstrate that intestinal sucrase activity is likely to contribute to diet
selectivity in frugivorous and nectarivorous bird species. Foraging data available for
nectarivorous species supported the laboratory analyses, with the rainbow lorikeet,
which demonstrated preferences for hexose over a wide range of diet concentrations in
laboratory preference trials, was also demonstrated to feed primarily on hexose nectars
in the wild. Mistletoebirds, unlike silvereyes, did not demonstrate a significant
preference for sucrose solutions at high diet concentrations, which may be influenced by
the low sucrose concentrations in mistletoe fruit.
The mistletoebird is currently the only specialised frugivore that has been
examined for concentration-dependent sugar preferences over a wide range of dietary
concentrations. More conclusive research into the sugar composition of Australian
mistletoe fruit is required to confirm that mistletoe fruits in general contain low amounts
of sucrose, as well as the assessment of sucrase activity.

172
Mistletoebirds demonstrated similar D-xylose AE* values to silvereyes and
singing honeyeaters (Chapter 6). Both silvereyes and singing honeyeaters showed
significantly increased bioavailability (f) of L-glucose and D-xylose on a more
concentrated diet, where slower gut passage time would promote increased absorption
via the paracellular route. By contrast, mistletoebirds did not vary f with diet
concentration and mistletoebirds appear to absorb xylose through both mediated and
paracellular mechanisms. This lower dependence on paracellular sugar absorption
(compared with the other two species tested in this thesis as well as various
nectarivorous bird species tested to date) is likely due to the short intestinal tract of
mistletoebirds, specialised for processing fruit. Mistletoebirds may rely more on the
mediated transport of glucose in comparison to silvereyes and singing honeyeaters
(whose relatively slower gut passage times would promote increased absorption via the
paracellular route).
7.4 FUTURE DIRECTIONS
How frugivorous and specialist nectarivores are able to survive with such low nitrogen
requirements remains unknown. While it has been suggested that these birds may have
lower rates of endogenous protein turnover and high nitrogen recycling capacity
(Witmer 1998; Pryor et al. 2001), yet this has not been confirmed to date. Birds excrete
urate in their urine in spherical concretions, which also contain protein and inorganic
ions (Casotti and Braun 1997; Casotti and Braun 2004). Tsahar et al. (2005), upon
reporting that protein concentration was lower in excreta than in ureteral urine in two
frugivorous bird species, hypothesised that some of the protein associated with urate-
containing spheres is digested in the lower intestine and recovered. This corresponds

173
with the findings of high activities of aminopeptidase-N (which is often used as a proxy
measure for the ability to digest protein, Maroux et al. 1973; Kania et al. 1977;
Martínez del Rio et al. 1995) in the terminal portion of the intestine of cedar waxwings
(Witmer and Martínez del Rio 2001). McWhorter et al. (unpublished data) also
demonstrated that aminopeptidase activity was highest for medial and distal portions of
the intestine for honeyeaters, sunbirds and a lorikeet. McWhorter et al. (unpublished
data) also reported the curious finding that nectarivorous and non-nectarivorous
passerines have similar total aminopeptidase activity, despite the lower than expected
nitrogen requirements of small nectarivores. Future research into the nitrogen
requirements of frugivores such as mistletoebirds and silvereyes is therefore of great
interest.
While singing honeyeaters and silvereyes have been recorded feeding on the
fruits of the two focal mistletoe species in this thesis in the literature, neither species
ingested mistletoe fruit whilst in captivity, nor were recorded feeding on mistletoe fruit
in the wild during surveys. Stable isotopes analysis has the advantage of allowing the
evaluation of the origin of assimilated nutrients – in this thesis, this approach
demonstrated that mistletoebirds supplement their diets with arthropods, despite the lack
of arthropod remains evident in the faeces of captured birds. During the field sampling
of mistletoebirds, I also sampled other bird species, including silvereyes and singing
honeyeaters (captured in mist-nets in front of fruiting mistletoe), with the aim to
compare differences in dietary contributions between species; however several
limitations resulted in the exclusion of this data from this thesis. Limitations included
the low capture numbers of some species, and the unsuccessful stable isotopes analysis
of other available dietary sources, such as nectar and sap.

174
Stable isotopes analysis has also been used in the study of other keystone species
in ecosystems around the world. For example, Symes et al. (2011) used carbon stable
isotopes (δ13
C) to investigate the importance of Aloe marlothii nectar to opportunistic
avian nectarivores in South Africa. Wolf and Martínez del Rio (2003) also used δ13
C
and δD to demonstrate how the nectar and fruit resources provided by saguaro
(Carnegiea gigantea) provide for a large proportion of the diets of many insectivorous,
granivorous and frugivorous avian species in the Sonoran Desert. This thesis has
demonstrated that stable isotopes analysis can be extremely useful in the investigation
of the nutritional ecology of mistletoebirds, and with a more complete isotopic sample
of available dietary sources, this approach can be utilised to investigate the nutritional
importance of mistletoe fruit and nectars to a wide range of bird species.
While this thesis makes some progress, the mechanisms of xylose absorption in
birds are still unclear. It appears that mistletoebirds may absorb D-xylose by both
mediated and paracellular mechanisms of absorption; further research to confirm the
mechanisms of absorption and potential catabolism of xylose by intestinal microbes or
systemic catabolism in birds would be advantageous.
7.5 CONCLUSIONS
This thesis presents new insight into the parasitic relationship between mistletoes and
hosts, and the relationship between mistletoes and its avian consumers. This thesis
presents the first data on mistletoe fruiting and flowering phenology and the influence
of mistletoe on bird communities in south-west WA, and demonstrates differences in
the mechanisms of sugar absorption by a mistletoe specialist, the mistletoebird. This

175
thesis also provides conclusive evidence that intestinal sucrase activity is likely to
contribute to diet selectivity in frugivorous and nectarivorous bird species.

176
8 REFERENCES
Afik, D., and Karasov, W. H. (1995). The trade-offs between digestion rate and efficiency in
warblers and their ecological implications. Ecology 76, 2247–2257.
Afik, D., McWilliams, S. R., and Karasov, W. H. (1997). A test for passive absorption of
glucose in yellow rumped warblers and its ecological implications. Physiological Zoology 70,
370–377.
Alpers, D. H. (1987). Digestion and absorption of carbohydrates and proteins. In 'Physiology of
the gastrointestinal tract.' (Ed. L. R. Johnson.) pp. 1469–1487. (Raven Press: New York.)
Alvarado, F. (1966). D-Xylose active transport in the hamster small intestine. Biochimica et
Biophysica Acta 112, 292–306.
Azuma, J.-i., Kim, N., Heux, L., Vuong, R., and Chanzy, H. (2000). The cellulose system in
viscin from mistletoe berries. Cellulose 7, 3–19.
Azuma, J.-i., and Sakamoto, M. (2003). Cellulosic hydrocolloid system present in seed of
plants. Trends in Glycoscience and Glycotechnology 15, 1–14.
Baker, H. G., and Baker, I. (1982a). Chemical constituents of nectar in relation to pollination
mechanisms and phylogeny. In 'Biochemical Aspects of Evolutionary Biology.' (Ed. M. H.
Nitecki.) pp. 131–171. (University of Chicago Press: Chicago.)
Baker, H. G., and Baker, I. (1983). Floral nectar sugar constituents in relation to pollinator type.
In 'Handbook of experimental pollination biology.' (Eds. C. E. Jones and R. J. Little.) pp. 117–
141. (Van Nostrand Reinhold: New York.)
Baker, H. G., Baker, I., and Hodges, S. A. (1998). Sugar compositions of nectars and fruits
consumed by birds and bats in the tropics and subtropics. Biotropica 30, 559–586.
Baker, I., and Baker, H. G. (1982b). Some chemical constituents of floral nectars of Erythrina
in relation to pollinators and systematics. Allertonia 3, 25–37.
Bannister, P. (1989). Nitrogen concentration and mimicry in some New Zealand mistletoes.
Oecologia 79, 128–132.
Bannister, P., and Strong, G. L. (2001). Carbon and nitrogen isotope ratios, nitrogen content and
heterotrophy in New Zealand mistletoes. Oecologia 126, 10–20.
Barea, L. P., and Watson, D. (2007). Temporal variation in food resources determines onset of
breeding in an Australian mistletoe specialist. Emu 107, 203–209.
Barea, L. P. (2008). Interactions between frugivores and their resources: case studies with the
painted honeyeater Grantiella picta. PhD Thesis, Charles Sturt University, Albury.
Barea, L. P., and Herrera, M. L. G. (2009). Sources of protein in two semi-arid zone mistletoe
specialists: insights from stable isotopes. Austral Ecology 34, 821–828.
Barker, R. D., and Vestjens, W. J. M. (1990). 'The food of Australian birds. II. passerines.'
(CSIRO: Lyneham.)

177
Barlow, B. A. (1984). Loranthaceae. In 'Flora of Australia. Vol. 22.' (Ed. A. S. George.) pp. 68–
131. (Australian Government Publishing Service: Canberra.)
Barlow, B. A. (1992). Conspectus of the genus Amyema Tieghem (Loranthaceae). Blumea 36,
293–381.
Bennetts, R. E., White, G. C., Hawksworth, F. G., and Severs, S. E. (1996). The influence of
dwarf mistletoe on bird communities in Colorado ponderosa pine forests. Ecological
Applications 6, 899–909.
Bernhardt, P. (1984). Mistletoes on mistletoes: the floral ecology of Amyema miraculosum and
its host, Amyema miquelii (Loranthaceae). Australian Journal of Botany 32, 73–86.
Beuchat, C. A., Calder, W. A., and Braun, E. J. (1990). The integration of osmoregulation and
energy-balance in hummingbirds. Physiological Zoology 63, 1059–1081.
Bihler, I., Hawkins, K. A., and Crane, R. K. (1962). Studies on the mechanism of intestinal
absorption of sugars. VI. The specificity and other properties of Na+-dependent entrance of
sugars into intestinal tissue under anaerobic conditions, in vitro. Biochimica et Biophysica Acta
59, 94–102.
Bizaare, L., Downs, C. T., and Coetzer, T. (2012). Disaccharidase presence and activities in a
range of southern African frugivores. Ostrich 83, 165–168.
Blakers, M., Davies, S., and Reilly, P. (1984). 'The atlas of Australian birds.' (Melbourne
University Press: Melbourne.)
Brown, E. M., Burbidge, A. H., Dell, J., Edinger, D., Hopper, S. D., and Wills, R. T. (1997).
'Pollination in Western Australia: a database of animals visiting flowers.' (Western Australian
Naturalists Club: Perth.)
Brown, M., Downs, C. T., and Johnson, S. D. (2008). Sugar preferences of nectar feeding birds
- a comparison of experimental techniques. Journal of Avian Biology, 279–483.
Brown, M., Downs, C. T., and Johnson, S. D. (2010a). Sugar preferences of a generalist non
passerine flower visitor, the African speckled mousebird (Colius striatus). The Auk 127, 781–
786.
Brown, M., Downs, C. T., and Johnson, S. D. (2010b). Sugar preferences and digestive
efficiency in an opportunistic avian nectarivore, the Dark-capped Bulbul Pycnonotus tricolor.
Journal of Ornithology 151, 637–643.
Brown, M., Downs, C. T., and Johnson, S. D. (2010c). Concentration-dependent sugar
preferences of the Malachite sunbird (Nectarinia famosa). The Auk 127, 151–155.
Brown, M., Downs, C. T., and Johnson, S. D. (2012). African Red-winged Starlings prefer
hexose sugar solutions, but do not like them too sweet. Journal of Ornithology 153, 265–272.
Brugger, K. E., Nol, P., and Phillips, C. I. (1993). Sucrose repellency to European starlings: will
high-sucrose cultivars deter bird damage to fruit? Ecological Applications 3, 256–261.
Bureau of Meteorology (2011). 'Climate statistics for Australian locations.' (Australian
Government) Available at http://www.bom.gov.au/climate/data/index.shtml

178
Burns, A. E., Cunningham, S. A., and Watson, D. M. (2011). Arthropod assemblages in tree
canopies: a comparison of orders on box mistletoe (Amyema miquelii) and its host eucalypts.
Australian Journal of Entomology 50, 221–230.
Calder, D. M. (1983). Mistletoes in focus: an introduction. In 'The biology of mistletoes.' (Eds.
D. M. Calder and P. Bernhardt.) pp. 1–18. (Academic Press: Sydney.)
Cannon, C. E. (1984). The diet of lorikeets Trichoglossus spp in the Queensland-New South
Wales border region. Emu 84, 16–22.
Carleton, S. A., Bakken, B. H., and Martinez del Rio, C. (2006). Metabolic substrate use and the
turnover of endogenous energy reserves in broad-tailed hummingbirds (Selasphorus
platycercus). Journal of Experimental Biology 209, 2622–2627.
Casotti, G., and Braun, E. J. (1997). Ionic composition of urate-containing spheres in the urine
of domestic fowl. Comparative Biochemistry and Physiology Part A: Physiology 118, 585–588.
Casotti, G., and Braun, E. J. (2004). Protein location and elemental composition of urine spheres
in different avian species. Journal of Experimental Zoology Part A: Comparative Experimental
Biology 301A, 579–587.
Caut, S., Angulo, E., and Courchamp, F. (2009). Variation in discrimination factors (Δ15N and
Δ13C): the effect of diet isotopic values and applications for diet reconstruction. Journal of
Applied Ecology 46, 443–453.
Caviedes-Vidal, E., McWhorter, T. J., Lavin, S. R., Chediack, J. G., Tracy, C. R., and Karasov,
W. H. (2007). The digestive adaptation of flying vertebrates: high intestinal paracellular
absorption compensates for smaller guts. Proceedings of the National Academy of Sciences of
the USA 104, 19132–19137.
Chaffer, N. (1966). Birds of the mistletoe. Wildlife Australia 3, 140–141.
Chamberlain, C. P., Blum, J. D., Holmes, R. T., Feng, X., Sherry, T. W., and Graves, G. R.
(1997). The use of isotope tracers for identifying populations of migratory birds. Oecologia 109,
132–141.
Chan, K. (2001). Partial migration in Australian landbirds: a review. Emu 101, 281–292.
Chang, M. H., and Karasov, W. H. (2004). Absorption and paracellular visualization of
fluorescein, a hydrosoluble probe, in intact house sparrows (Passer domesticus). Zoology 107,
121–133.
Chediack, J. G., Caviedes-Vidal, E., Fasulo, V., Yamin, L. J., and Karasov, W. H. (2003).
Intestinal passive absorption of water-soluble compounds by sparrows: effect of molecular size
and luminal nutrients. Journal of Comparative Physiology 173, 187–197.
Christidis, L., and Boles, W. E. (2008). 'Systematics and taxonomy of Australian birds.' (CSIRO
Publishing: Melbourne.)
Clarke, K. R. (1993). Non-parametric multivariate analysis of changes in community structure.
Australian Journal of Ecology 18, 117–143.
Collins, B. G., and Morellini, P. C. (1979). The influence of nectar concentration and time of
day upon energy intake and expenditure by the singing honeyeater, Meliphaga virescens.
Physiological Zoology 52, 165–175.

179
Cooney, S. J. N., Watson, D. M., and Young, J. (2006). Mistletoe nesting in Australian birds: a
review. Emu 106, 1–12.
Cooney, S. J. N., and Watson, D. M. (2008). An experimental approach to understanding the
use of mistletoe as a nest substrate for birds: nest predation. Wildlife Research 35, 65–71.
Csaky, T. Z., and Lassen, U. V. (1964). Active intestinal transport of D-xylose. Biochimica et
Biophysica Acta 82, 215–217.
Dahlqvist, A. (1984). Assay of intestinal disaccharides. Scandinavian Journal of Clinical
Laboratory Investigation 44, 69–72.
Davidar, P. (1983). Similarity between flowers and fruits in some flowerpecker pollinated
mistletoes. Biotropica 15, 32–37.
Davidar, P. (1985). Ecological interactions between mistletoes and their avian pollinators in
south India. Journal of the Bombay Natural History Society 82, 45–60.
Davidson, N., True, K., and Pate, J. S. (1989a). Water relations of the parasite/host relationship
between the mistletoe Amyema linophyllum (Fenzl) Tieghem and Casuarina obesa Miq.
Oecologia 80, 321–330.
Davidson, N. J., True, K. C., and Pate, J. S. (1989b). Water relations of the parasite:host
relationship between the mistletoe Amyema linophyllum (Fenzl) Tieghem and Casuarina obesa
Miq. Oecologia 80, 321–330.
Davidson, N. J., and Pate, J. S. (1992). Water relations of the mistletoe Amyema fitzgeraldii and
its host Acacia acuminata. Journal of Experimental Botany 43, 1549–1555.
Davis, A. R. (1997). Influence of floral visitation on nectar-sugar composition and nectary
surface changes in Eucalyptus. Apidologie 28, 27–42.
Demetrakopoulos, G. E., and Amos, H. (1978). Xylose and xylitol. World Review of Nutrition
and Dietetics 32, 96–122.
DEP (1997). 'State of the Environment Reference Group draft working papers. Section 4, Land.'
(State of the Environment Reporting Unit, Department of Environmental Protection: Perth,
WA.)
Dörr, I. (1997). How Striga parasitizes its host: a TEM and SEM study. Annals of Botany 79,
463–472.
Downs, C. T. (1997). Sugar digestion efficiencies of Gurney's sugarbirds, malachite sunbirds,
and black sunbirds. Physiological Zoology 70, 93–99.
Ehleringer, J. R., Schulze, E.-D., Ziegler, H., Lange, O. L., Farquhar, G. D., and Cowan, I. R.
(1985). Xylem-tapping mistletoes: water or nutrient parasites? Science 227, 1479–1481.
Ehleringer, J. R., Ullmann, I., Lange, O. L., Farquhar, G. D., Cowan, I. R., Schulze, E. D., and
Ziegler, H. (1986). Mistletoes: a hypothesis concerning morphological and chemical avoidance
of herbivory. Oecologia 70, 234–237.
Ehleringer, J. R., and Marshall, J. D. (1995). Water relations. In 'Parasitic plants.' (Eds. M. C.
Press and J. D. Graves.) pp. 125–140. (Chapman & Hall: London.)

180
Ericson, P. G. P., Anderson, C. L., Britton, T., Elzanowski, T. A., Johansson, U. S., Källersjö,
M., Ohlson, J. I., Parsons, T. J., Zuccon, D., and Mayr, G. (2006). Diversification of Neoaves:
integration of molecular sequence data and fossils. Biology Letters 2, 543–547.
Farquhar, G. D., O'Leary, M. H., and Berry, J. A. (1982). On the relationship between carbon
isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian
Journal of Plant Physiology 9, 121–137
Farquhar, G. D., Ehleringer, J. R., and Hubick, K. T. (1989). Carbon isotope discrimination and
photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503–537
Felsenstein, J. (1985). Phylogenies and the comparative method. American Naturalist 125, 1–
15.
Ferraris, R. P. (2001). Dietary and developmental regulation of intestinal sugar transport.
Biochemical Journal 360, 265–276.
Field, S. A., Tyre, A. J., and Possingham, H. P. (2002). Estimating bird species richness: how
should repeat surveys be organized in time? Austral Ecology 27, 624–629.
Fine, K. D., Santa Ana, C. A., Porter, J. L., and Fordtran, J. S. (1993). Effect of D-glucose on
intestinal permeability and its passive absorption in human small intestine in vivo.
Gastroenterology 105, 1117–1125.
Fleming, P. A., Bakken, B. H., Lotz, C. N., and Nicolson, S. W. (2004). Concentration and
temperature effects on sugar intake and preferences in a sunbird and a hummingbird. Functional
Ecology 18, 223–232.
Fleming, P. A., Xie, S., Napier, K., McWhorter, T. J., and Nicolson, S. W. (2008). Nectar
concentration affects sugar preferences in two Australian honeyeaters and a lorikeet. Functional
Ecology 18, 223–232.
Forde, N. (1986). Relationships between birds and fruits in temperate Australia. In 'The
dynamic partnership: birds and plants in southern Australia.' (Eds. H. A. Ford and D. C. Paton.)
pp. 42–58. (Government Printer: Adelaide.)
Franke, E., Jackson, S., and Nicolson, S. (1998). Nectar sugar preferences and absorption in a
generalist African frugivore, the Cape White-eye Zosterops pallidus. Ibis 140, 501–506.
French, K. (1996). The gut passage rate of Silvereyes and its effect on seed viability. Corella
20, 16–19.
Friedman, M. H. (1987). 'Principles and models of biological transport.' (Springer: Berlin.)
Gagnon, C., and Hobson, K. A. (2009). Using stable isotopes to track frugivory in migratory
passerines. Canadian Journal of Zoology 87, 981–992.
Garland, T. J., Harvey, P. J., and Ives, A. R. (1992). Procedures for the analysis of comparative
data using phylogenetically independent contrasts. Systematic Biology 41, 18–32.
Garland, T. J., Dickerman, A. W., Janis, C. M., and Jones, J. A. (1993). Phylogenetic analysis of
covariance by computer simulation. Systematic Biology 42, 265–292.
Garland, T. J., and Adolph, S. C. (1994). Why not do two-species comparative studies:
limitations on inferring adaptation. Physiological Zoology 67, 797–828.

181
Garland, T. J., Midford, P. E., and Ives, A. R. (1999). An introduction to phylogenetically based
statistical methods, with a new method for confidence intervals on ancestral values. American
Zoologist 39, 374–388.
Garland, T. J., and Ives, A. R. (2000). Using the past to predict the present: confidence intervals
for regression equations in phylogenetic comparative methods. American Naturalist 155, 346–
364.
Gatica, C. D. L., Gonzalez, S. P., Vasquez, R. A., and Sabat, P. (2006). On the relationship
between sugar digestion and diet preference in two Chilean avian species belonging to the
Muscicapoidea superfamily. Revista Chilena De Historia Natural 79, 287–294.
Gedalovich-Shedletzky, E., Delmer, D. P., and Kuijt, J. (1989). Chemical composition of viscin
mucilage from three Mistletoe species- a comparison. Annals of Botany 64, 249–252.
Gedalovich, E., Kuijt, J., and Carita, N. C. (1988). Chemical composition of viscin, an adhesive
involved in dispersal of the parasite Phoradendron californicum (Viscaceae). Physiological and
Molecular Plant Pathology 32, 61–76.
Godschalk, S. K. B. (1983a). A biochemical analysis of the fruit of Tapinanthus leendertziae.
South African Journal of Botany 2, 42–54.
Godschalk, S. K. B. (1983b). Mistletoe dispersal by birds in South Africa. In 'The biology of
mistletoes.' (Eds. D. M. Calder and P. Bernhardt.) pp. 117–128. (Academic Press: Sydney.)
Goldstein, D. C., and Skadhauge, E. (2000). Renal and extrarenal regulation of body fluid
composition. In 'Sturkies avian physiology, 5th edition.' (Ed. G. Causey Whittow.) pp. 265–297.
(Academic Press: California.)
Gottsberger, G., Schrauwen, J., and Linskens, H. F. (1984). Amino acids and sugars in nectar,
and their putative evolutionary significance. Plant Systematics and Evolution 145, 55–77.
Govier, R. N., Nelson, M. D., and Pate, J. S. ( 1967). Hemiparasitic nutrition in Angiosperms I.
The transfer of organic compounds from host to Odontites verna (Bell.) Dum.
(Scrophulariaceae) New Phytologist 66, 285–297.
Hahn, S., Hoye, B. J., Korthals, H., and Klaassen, M. (2012). From food to offspring down:
tissue-specific discrimination and turn-over of stable isotopes in herbivorous waterbirds and
other avian foraging guilds. PLoS ONE 7, e30242.
Hammer, Ø., Harper, D. A. T., and Ryamn, P. D. (2001). PAST: paleontological statistics
software package for education and data analysis. Palaeontologia Electronica 4, 9.
Harborne, J. B. (1993). 'Introduction to ecological biochemistry.' 4 edn. (Academic Press)
Hawkeswood, T. J. (1981). Notes on the pollination of Nuytsia floribunda (Labill.) R. Br.
(Loranthaceae) and some literature reviewed. Western Australian Naturalist 15, 17–21.
Hawksworth, F. G., and Geils, B. W. (1996). Biotic associates. In 'Dwarf mistletoes: biology,
pathology, and systematics. Agricultural Handbook 709.' 2 edn. (Eds. F. G. Hawksworth and D.
Wiens.) pp. 73–89. (USDA Forest Service: Washington, DC.)
Hawksworth, F. G., and Wiens, D. (1996). 'Dwarf mistletoes: biology, pathology, and
systematics. Agricultural Handbook 709.' 2 edn. (USDA Forest Service: Washington, DC.)

182
Herrera, C. M. (1987). Vetebrate-dispersed plants of the Iberian Peninsula: a study of fruit
characteristics. Ecological Monographs 57, 305–331.
Herrera, L. G., Hobson, K. A., RamÍrez P, N., Méndez C, G., and Sánchez-Cordero, V. (2001a).
Sources of protein in two species of phytophagous bats in a seasonal dry forest: evidence from
stable-isotope analysis. Journal of Mammalogy 82, 352–361.
Herrera, L. G., Hobson, K., Rodríguez, M., and Hernandez, P. (2003). Trophic partitioning in
tropical rain forest birds: insights from stable isotope analysis. Oecologia 136, 439–444.
Herrera, L. G. M., Hobson, K. A., Hernández, P. C., and Rodríguez, M. G. (2005). Quantifying
differential responses to fruit abundance by two rain forest birds using long-term isotopic
monitoring. The Auk 122, 783–792.
Herrera, M. L. G., Hobson, K. A., Manzo, A., Estrada, B., Sánchez‐Cordero, V., and Méndez,
C. (2001b). The role of fruits and insects in the nutrition of frugivorous bats: evaluating the use
of stable isotope models. Biotropica 33, 520–528.
Herrera, M. L. G., Hobson, K. A., Martínez, J. C., and Méndez C., G. (2006). Tracing the origin
of dietary protein in tropical dry forest birds. Biotropica 38, 735–742.
Herrera, M. L. G., Rodríguez, M. G., and Hernández, P. P. (2009). Sources of assimilated
protein in a specialized tropical frugivorous bird, the yellow-throated Euphonia (Euphonia
hirundinacea). The Auk 126, 175–180.
Heyman, M., Dumontier, A., and Desjeux, J. (1980). Xylose transport pathways in rabbit ileum.
American Journal of Physiology 238, G326–G331.
Higgins, P. J. (Ed.) (1999). 'Handbook of Australian, New Zealand and Antarctic Birds. Volume
4: Parrots to Dollarbird.' (Oxford University Press: Melbourne.)
Higgins, P. J., Peter, J. M., and Steele, W. K. (Eds) (2001). 'Handbook of Australian, New
Zealand and Antarctic birds.Volume 5: Tyrant-flycatchers to Chats.' (Oxford University Press:
Melbourne.)
Higgins, P. J., Peter, J. M., and Cowling, S. J. (Eds) (2006). 'Handbook of Australian, New
Zealand and Antarctic Birds. Volume 7: Boatbill to Starlings.' (Oxford University Press:
Melbourne.)
Hobson, K. A. (1999). Tracing origins and migration of wildlife using stable isotopes: a review.
Oecologia 120, 314–326.
Hobson, K. A., and Bairlein, F. (2003). Isotopic fractionation and turnover in captive garden
warblers (Sylvia borin): implications for delineating dietary and migratory associations in wild
passerines. Canadian Journal of Zoology 81, 1630–1635.
Hobson, K. A. (2007). Isotopic tracking of migrant wildlife. In 'Stable Isotopes in Ecology and
Environmental Science.' 2 edn. (Eds. R. Michener and K. Lajtha.)Blackwell Publishing Ltd:
Malden, MA.)
Holdsworth, D. C., and Dawson, A. M. (1964). The absorption of monosaccharides in man.
Clinical Science 27.
Hölscher, D., Brand, S., Wenzler, M., and Schneider, B. (2008). NMR-based metabolic
profiling of Anigozanthos floral nectar. Journal of Natural Products 71, 251–257.

183
Hull, R. J., and Leonard, O. A. (1964). Physiological aspects of parasitism in mistletoes
(Arceuthobium and Phoradendron) 1. The carbohydrate nutrition of mistletoes. Plant
Physiology 39, 996–1007.
Izhaki, I., Tsahar, E., Paluy, I., and Friedman, J. (2002). Within population variation and
interrelationships between morphology, nutritional content, and secondary compounds of
Rhamnus alaternus fruits. New Phytologist 156, 217–223.
Jackson, S., Nicolson, S. W., and Lotz, C. N. (1998a). Sugar preferences and "side bias" in Cape
Sugarbirds and Lesser Double-collared Sunbirds. Auk 115, 156–165.
Jackson, S., Nicolson, S. W., and van Wyk, B. E. (1998b). Apparent absorption efficiencies of
nectar sugars in the Cape Sugarbird, with a comparison of methods. Physiological Zoology 71,
106–115.
Jackson, S., and Nicolson, S. W. (2002). Xylose as a nectar sugar: from biochemistry to
ecology. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology 131,
613–620.
Jetz, W., Thomas, G. H., Joy, J. B., Hartmann, K., and Mooers, A. O. (2012). The global
diversity of birds in space and time. Nature 491, 444–448.
Johnson, S. A., van Tets, I. G., and Nicolson, S. W. (1999). Sugar preferences and xylose
metabolism of a mammal pollinator, the Namaqua rock mouse (Aethomys namaquensis).
Physiological and Biochemical Zoology 72, 438–444.
Johnson, S. A., Nicolson, S. W., and Jackson, S. (2006). Nectar xylose metabolism in a rodent
pollinator (Aethomys namaquensis): Defining the role of gastrointestinal microflora using 14C-
labeled xylose. Physiological and Biochemical Zoology 79, 159–168.
Johnson, S. D., and Nicolson, S. W. (2008). Evolutionary associations between nectar properties
and specificity in bird pollination systems. Biology Letters 4, 49–52.
Kania, R. K., Santiago, N. A., and Gray, G. M. (1977). Intestinal surface amino-oligopeptidases.
II. Substrate kinetics and topography of the active site. Journal of Biological Chemistry 252,
4929–4934.
Karasov, W. H., and Levey, D. J. (1990). Digestive system tradeoffs and adaptations in
frugivorous passerine birds. Physiological Zoology 63, 1246–1270.
Karasov, W. H., and Cork, S. J. (1994). Glucose absorption by a nectarivorous bird: the passive
pathway is paramount. American Journal of Physiology 267, G18–G26.
Keast, A. (1958). The influence of ecology on variation in the mistletoebird (Dicaeum
hirundinaceum). Emu 58, 195–206.
Kelly, D., Ladley, J. J., Robertson, A. W., and Norton, D. A. (2000). Limited forest
fragmentation improves reproduction in the declining New Zealand mistletoe Peraxilla
tetrapetala (Loranthaceae). In 'Genetics, demography and viability of fragmented populations.'
(Eds. A. Young and G. Clarke.) pp. 241–252. (Cambridge University Press: Cambridge.)
Kelly, J. F. (2000). Stable isotopes of carbon and nitrogen in the study of avian and mammalian
trophic ecology. Canadian Journal of Zoology 78, 1–27.
Klasing, K. C. (1998). 'Comparative avian nutrition.' (C.A.B International: New York.)

184
Knutson, D. M. (1983). Physiology of mistletoe parasitism and disease responses in the host. In
'The biology of mistletoes.' (Eds. D. M. Calder and P. Bernhardt.) pp. 295–316. (Academic
Press: Sydney.)
Köhler, A., Verburgt, L., McWhorter, T. J., and Nicolson, S. (2010). Energy management on a
nectar diet: can sunbirds meet the challenges of low temperature and dilute food? Functional
Ecology 24, 1241–1251.
Kohn, P., Dawes, E. D., and Duke, J. W. (1965). Absorption of carbohydrates from the intestine
of the rat. Biochimica et Biophysica Acta 107, 358–362.
Kotliar, N. B. (2000). Application of the new keystone-species concept to prairie dogs: how
well does it work? Conservation Biology 14, 1715–1721.
Kuijt, J. (1969). 'The biology of parasitic flowering plants.' (University of California Press:
Berkeley, Los Angeles.)
Ladley, J. J., Kelly, D., and Robertson, A. W. (1997). Explosive flowering, nectar production,
breeding systems, and pollinators of New Zealand mistletoes (Loranthaceae). New Zealand
Journal of Botany 35, 345–360.
Lamont, B. (1982). Host range and germination requirements of some South African mistletoes.
South African Journal of Science 78, 41–42.
Lamont, B. (1983a). Germination of mistletoes. In 'The biology of mistletoes.' (Eds. D. M.
Calder and P. Bernhardt.) pp. 129–143. (Academic Press: Sydney.)
Lamont, B. (1983b). Mineral nutrition of mistletoes. In 'The biology of mistletoes.' (Eds. D. M.
Calder and P. Bernhardt.) pp. 185–204. (Academic Press: Sydney.)
Lane, S. (1991). Preferences and apparent digestibilities of sugars by fruit damaging birds in
Japan. Annals of Applied Biology 130, 361–370.
Lassen, U. V., and Csaky, T. Z. (1966). Active transport of d-xylose in the isolated small
intestine of the bullfrog. Journal of General Physiology 49, 1029–1041.
Levey, D. J. (1988). Spatial and temporal variation in Costa Rican fruit and fruit-eating bird
abundance. Ecological Monographs 58, 351–269.
Levey, D. J., and Grajal, A. (1991). Evolutionary implications of fruit-processing limitations in
Cedar Waxwings. The American Naturalist 138, 171–189.
Levey, D. J., and Martinez del Rio, C. (1999). Test, rejection, and reformulation of a chemical
reactor-based model of gut function in a fruit-eating bird. Physiological and Biochemical
Zoology 72, 369–383.
Levey, D. J., Bissell, H. A., and O'Keefe, S. F. (2000). Conversion of nitrogen to protein and
amino acids in wild fruits. Journal of Chemical Ecology 26, 1749–1763.
Levey, D. J., and Martinez Del Rio, C. (2001). It takes guts (and more) to eat fruit: lessons from
avian nutritional ecology. The Auk 118, 819–831.
Liddy, J. (1983). Dispersal of Australian mistletoes: the Cowiebank study. In 'The biology of
mistletoes.' (Eds. M. Calder and P. Bernhardt.) pp. 101–116. (Academic Press: Sydney.)

185
Lopez-Calleja, M. V., Bozinovic, F., and Martinez del Rio, C. (1997). Effects of sugar
concentration on hummingbird feeding and energy use. Comparative Biochemistry and
Physiology a-Molecular & Integrative Physiology 118, 1291–1299.
López de Buen, L., and Ornelas, J. F. (2001). Seed dispersal of the mistletoe Psittacanthus
schiedeanus by birds in central Veracruz, Mexico. Biotropica 33, 487–494.
Lotz, C. N., and Nicolson, S. W. (1996). Sugar preferences of a nectarivorous passerine bird,
the Lesser Double-collared Sunbird (Nectarinia chalybea). Functional Ecology 10, 360–365.
Lotz, C. N., and Martínez del Rio, C. (2004). The ability of rufous hummingbirds Selasphorus
rufus to dilute and concentrate urine. Journal of Avian Biology 35, 54–62.
Lotz, C. N., and Schondube, J. E. (2006). Sugar preferences in nectar- and fruit-eating birds:
behavioral patterns and physiological causes. Biotropica 38, 3–15.
Malizia, L. R. (2001). Seasonal fluctuations of birds, fruits, and flowers in a subtropical forest
of Argentina. The Condor 103, 45–61.
March, W., and Watson, D. (2007). Parasites boost productivity: effects of mistletoe on litterfall
dynamics in a temperate Australian forest. Oecologia 154, 339–347.
March, W. A., and Watson, D. M. (2010). The contribution of mistletoes to nutrient returns:
evidence for a critical role in nutrient cycling. Austral Ecology 35, 713–721.
Maroux, S., Louvard, D., and Baratti, J. (1973). The aminopeptidase from hog intestinal brush-
border. Biochimica et Biophysica Acta 321, 282–295.
Marquardt, D. W. (1963). An algorithm for least squares estimation of parameters. Journal of
the Society of Industrial and Applied Mathematics 11, 431–441.
Marshall, J. D., and Ehleringer, J. R. (1990). Are xylem-tapping mistletoes partially
heterotrophic? Oecologia 84, 244–248.
Marshall, J. D., Ehleringer, J. R., Schulze, E.-D., and Farquhar, G. (1994). Carbon isotope
composition, gas exchange and heterotrophy in Australian mistletoes. Functional Ecology 8,
237–241.
Martinez del Rio, C. (1990). Dietary, phylogenetic, and ecological correlates of intestinal
sucrase and maltase activity in birds. Physiological Zoology 63, 987–1011.
Martínez del Rio, C., and Stevens, B. R. (1989). Physiological constraint on feeding behavior:
intestinal membrane disaccharides of the starling. Science 243, 794–796.
Martínez del Rio, C. (1990). Sugar preferences in hummingbirds – the influence of subtle
chemical differences on food choice. Condor 92, 1022–1030.
Martínez del Rio, C., Baker, H. G., and Baker, I. (1992). Ecological and evolutionary
implications of digestive processes – bird preferences and the sugar constituents of floral nectar
and fruit pulp. Experientia 48, 544–550.
Martínez del Rio, C., Brugger, K. E., Rios, J. L., Vergara, M. E., and Witmer, M. C. (1995). An
experimental and comparative study of dietary modulation of intestinal enzymes in European
starlings (Sturnus vulgaris). Physiological Zoology 68, 490–511.

186
Martínez del Rio, C., Schondube, J. E., McWhorter, T. J., and Herrera, L. G. (2001). Intake
responses in nectar feeding birds: digestive and metabolic causes, osmoregulatory
consequences, and coevolutionary effects. American Zoologist 41, 902–915.
Mathiasen, R. L., Daugherty, C. M., Howell, B. E., Melgar, J. C., and Sesnie, S. E. (2007). New
morphological measurements for Psittacanthus angustifolius and Psiitacanthus pinicola
(Lornathaceae). Madroño 36, 156–163.
Mathiasen, R. L., Nickrent, D. L., Shaw, D. C., and Watson, D. M. (2008). Mistletoes:
pathology, systematics, ecology, and management. Plant Disease 92, 988–1006.
McFarland, D. C. (1985). Flowering biology and phenology of Banksia integrifolia and B.
spinulosa (Proteaceae) in New England National Park, N.S.W. Australian Journal of Botany 33,
705–714.
McPherson, J. M. (1987). A field study of winter fruit preferences of Cedar Waxwings. The
Condor 89, 293–306.
McWhorter, T. J., and Martinez del Rio, C. (1999). Food ingestion and water turnover in
hummingbirds: how much dietary water is absorbed? Journal of Experimental Biology 202,
2851–2858.
McWhorter, T. J., and Lopez-Calleja, M. V. (2000). The integration of diet, physiology, and
ecology of nectar-feeding birds. Revista Chilena De Historia Natural 73, 451–460.
McWhorter, T. J., and Martinez del Rio, C. (2000). Does gut function limit hummingbird food
intake? Physiological and Biochemical Zoology 73, 313–324.
McWhorter, T. J., Martinez del Rio, C., and Pinshow, B. (2003). Modulation of ingested water
absorption by Palestine sunbirds: evidence for adaptive regulation. Journal of Experimental
Biology 206, 659–666.
McWhorter, T. J. (2005). Paracellular intestional absorption of carbohydrates in mammals and
birds. In 'Physiological and ecological adaptations to feeding in vertebrates.' (Eds. J. M. Starck
and T. Wang.) pp. 113–140. (Science Publishers: Enfield, New Hampshire.)
McWhorter, T. J., Hartman Bakken, B., Karasov, W. H., and Martinez del Rio, C. (2006).
Hummingbirds rely on both paracellular and carrier-mediated intestinal glucose absorption to
fuel high metabolism. Biology Letters 2, 131–134.
McWhorter, T. J., Green, A. K., and Karasov, W. H. (2010). Assessment of radiolabeled d-
glucose and the nonmetabolizable analog 3-O-methyl-d-glucose as tools for in vivo absorption
studies. Physiological and Biochemical Zoology 83, 376–384.
Medina-Tapia, N., Ayala-Berdon, J., Morales-Pérez, L., Mirón Melo, L., and Schondube, J. E.
(2012). Do hummingbirds have a sweet-tooth? Gustatory sugar thresholds and sugar selection in
the broad-billed hummingbird Cynanthus latirostris. Comparative Biochemistry and
Physiology, Part A 161, 307–314.
Midford, P. E., Garland, J. T., and Maddison, W. P. (2009). 'PDAP:PDTREE package for
Mesquite.' Version 1.15. (National Center for Biotechnology Information: Bethesda, MD.)
Miller, A. C., Watling, J. R., Overton, I. C., and Sinclair, R. (2003). Does water status of
Eucalyptus largiflorens (Myrtaceae) affect infection by the mistletoe Amyema miquelii
(Loranthaceae)? Functional Plant Biology 30, 1239–1247.

187
Montaño-Centellas, F. A. (2013). Effectiveness of mistletoe seed dispersal by Tyrant
Flycatchers in a mixed Andean landscape. Biotropica 45, 209–216.
Morrant, D. S., Petit, S., and Schumann, R. (2010). Floral nectar sugar composition and
flowering phenology of the food plants used by the western pygmy possum, Cercartetus
concinnus, at Innes National Park, South Australia. Ecological Research 25, 579–589.
Morton, E. S. (1973). On the evolutionary advantages and disadvantages of fruit eating in
tropical birds. The American Naturalist 107, 8–22.
Murphy, S. R., Reid, N., Yan, Z., and Venables, W. N. (1993). Differential passage time of
mistletoe fruits through the gut of honeyeaters and flowerpeckers: effects on seedling
establishment. Oecologia 93, 171–176.
Napier, K. R., McWhorter, T. J., and Fleming, P. A. (2008a). Mechanism and rate of glucose
absorption differ between an Australian honeyeater (Meliphagidae) and a lorikeet (Loriidae).
Journal of Experimental Biology 211, 3544–3553.
Napier, K. R., Purchase, C., McWhorter, T. J., Nicolson, S. W., and Fleming, P. A. (2008b).
The sweet life: diet sugar concentration influences paracellular glucose absorption. Biology
letters 4, 530–533.
Napier, K. R., McWhorter, T. J., and Fleming, P. A. (2012). A comparison of pharmacokinetic
methods for in vivo studies of nonmediated glucose absorption. Physiological and Biochemical
Zoology 85, 200–208.
Napier, K. R., Mather, S. H., McWhorter, T. J., and Fleming, P. A. (2013). Do bird species
richness and community structure vary with mistletoe flowering and fruiting in Western
Australia? Emu In press.
Nickrent, D. L. (2002). Mistletoe phylogenetics: current relationships gained from analysis of
DNA sequences. In 'Proceedings of the forty-eighth Western international forest disease work
conference.' (Eds. F. G. Hawksworth and R. F. Scharpf.) pp. 48–57. (USDA Forest Service:
Redding, California, Kona, Hawaii.)
Nicolson, S. W. (1994). Eucalyptus nectar: Production, availability, composition and osmotic
consequences for the larva of the eucalypt nectar fly, Drosophila flavohirta. South African
Journal of Science 90, 75–79.
Nicolson, S. W. (1998). The importance of osmosis in nectar secretion and its consumption by
insects. American Zoologist 38, 418–425.
Nicolson, S. W., and Van Wyk, B. E. (1998). Nectar sugars in Proteaceae: patterns and
processes. Australian Journal of Botany 46, 489–504.
Nicolson, S. W. (2002). Pollination by passerine birds: why are the nectars so dilute?
Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology 131, 645–652.
Nicolson, S. W., and Fleming, P. A. (2003a). Energy balance in the Whitebellied Sunbird
Nectarinia talatala: constraints on compensatory feeding, and consumption of supplementary
water. Functional Ecology 17, 3–9.
Nicolson, S. W., and Fleming, P. A. (2003b). Nectar as food for birds: the physiological
consequences of drinking dilute sugar solutions. Plant Systematics and Evolution 238, 139–153.

188
Norton, D. A., Hobbs, R. J., and Atkins, L. (1995). Fragmentation, disturbance, and plant
distribution: mistletoes in woodland remnants in the Western Australian wheatbelt.
Conservation Biology 9, 426–458.
Obeso, J. R., and Herrera, C. M. (1994). Inter- and intraspecific variation in fruit traits in co-
occurring vertebrate-dispersed plants. International Journal of Plant Sciences 155, 382–387.
Odendaal, T. C., Brown, M., Downs, C. T., and Johnson, S. D. (2010). Sugar preferences and
digestive efficiency of the village weaver: a generalist avian pollinator of African plants.
Journal of Experimental Biology 213, 2531–2535.
Ohkohchi, N., and Himukai, M. (1984). Species difference in mechanisms of D-xylose
absorption by the small intestine. Japanese Journal of Physiology 34, 669–677.
Ohkohchi, N., Himukai, M., Igarashi, Y., and Kasai, M. (1986). Mechanism of D-xylose
transport in human small intestine. Journal of Pediatric Gastroenterology and Nutrition 5, 372–
378.
Panvini, A. D., and Eickmeier, W. G. (1993). Nutrient and water relations of the mistletoe,
Phoradendron leucarpum (Viscaceae): how tightly are they integrated? American Journal of
Botany 80, 872–878.
Parnell, A. C., Inger, R., Bearhop, S., and Jackson, A. L. (2010). Source partitioning using
stable isotopes: coping with too much variation. PLoS ONE 5, e9672.
Paton, D. C., and Ford, H. A. (1977). Pollination by birds of native plants in South Australia.
Emu 77, 73–85.
Paton, D. C. (1982). The diet of the New Holland honeyeater, Phylidonyris novaehollandiae.
Australian Journal of Ecology 7, 279–298.
Pearson, S. F., Levey, D. J., Greenberg, C. H., and Martínez del Rio, C. (2003). Effects of
elemental composition on the incorporation of dietary nitrogen and carbon isotopic signatures in
an omnivorous songbird. Oecologia 135, 516–523.
Peres, C. A. (2000). Identifying keystone plant resources in tropical forests: the case of gums
from Parkia pods. Journal of Tropical Ecology 16, 287–317.
Phillips, D. L., and Koch, P. L. (2002). Incorporating concentration dependence in stable
isotope mixing models. Oecologia 130, 114–125.
Podlesak, D. W., McWilliams, S. R., and Hatch, K. A. (2005). Stable isotopes in breath, blood,
feces and feathers can indicate intra-individual changes in the diet of migratory songbirds.
Oecologia 142, 501–510.
Polhill, R., and Wiens, D. (1998). 'Mistletoes of Africa.' (Royal Botanic Gardens: Kew.)
Post, D. M. (2002). Using stable isotopes to estimate trophic position: models, methods, and
assumptions. Ecology 83, 703–718.
Power, M. E., Tilman, D., Estes, J. A., Menge, B. A., Bond, W. J., Mills, L. C., Daily, G.,
Castilla, J. C., Lubchenco, J., and Paine, R. T. (1996). Challenges in the quest for keystones.
BioScience 46, 609–620.

189
Press, M. C. (1995). Carbon and nitrogen relations. In 'Parasitic flowering plants.' (Eds. M. C.
Press and J. D. Graves.) pp. 103–124. (Chapman and Hall: London.)
Press, M. P. (1998). Dracula or robin hood? A functional role for root hemiparasites in nutrient
poor ecosystems. Oikos 82, 606–611.
Pryor, G. S., Levey, D. J., and Dierenfeld, E. S. (2001). Protein requirements of a specialized
frugivore, Pesquet's Parrot (Psittrichas fulgidus). The Auk 118, 1080–1088.
Pyke, G. H., and Waser, N. M. (1981). The production of dilute nectars by hummingbird and
honeyeater flowers. Biotropica 13, 260–270.
Rand, E. B., Depaoli, A. M., Davidson, N. O., Bell, G. I., and Burant, C. F. (1993). Sequence,
tissue distribution, and functional characterization of the rat fructose transporter GLUT5.
American Journal of Physiology 264, G1169–G1176.
Rawsthorne, J., Watson, D. M., and Roshier, D. A. (2011). Implications of movement patterns
of a dietary generalist for mistletoe seed dispersal. Austral Ecology 36, 650–655.
Rawsthorne, J., Watson, D. M., and Roshier, D. A. (2012). The restricted seed rain of a
mistletoe specialist. Journal of Avian Biology 43, 9–14.
Reid, N. (1986). Pollination and seed dispersal of mistletoes (Loranthaceae) by birds in southern
Australia. In 'The dynamic partnership: birds and plants in southern Australia.' (Eds. H. A. Ford
and D. C. Paton.) pp. 64–84. (Government Printer: Adelaide.)
Reid, N. (1987). The mistletoebird and Australian mistletoes: co-evolution or coincidence? Emu
87, 130–131.
Reid, N., and Lange, R. T. (1988). Host specificity, dispersion and persistence through drought
of two arid zone mistletoes. Australian Journal of Botany 36, 299–313.
Reid, N. (1989). Dispersal of mistletoes by honeyeaters and flowerpeckers: components of seed
dispersal quality. Ecology 70, 137–145.
Reid, N. (1990). Mutualistic interdependence between mistletoes (Amyema quandang), and
honeyeaters and mistletoebirds in an arid woodland. Australian Journal of Ecology 15, 175–
190.
Reid, N. (1991). Coevolution of mistletoes and frugivorous birds? Australian Journal of
Ecology 16, 457–469.
Reid, N., Yan, Z., and Fittler, J. (1994). Impact of mistletoes (Amyema miquelii) on host
(Eucalyptus blakelyi and Eucalyptus melliodora) survival and growth in temperate Australia.
Forest Ecology and Management 70, 55–65.
Reid, N., Stafford Smith, M., and Yan, Z. (1995). Ecology and population biology of mistletoes.
In 'Forest canopies.' (Eds. M. D. Lowman and N. M. Nadkarn.) pp. 285–310. (Academic Press:
San Diego, CA.)
Restrepo, C. (1987). Aspectos ecol´ogicos de la diseminaci´on de cinco especies de mu´erdagos
por aves. Humboldtia 1, 65–116.
Rezende, E. L., and Diniz-Filho, J. A. F. (2012). Phylogenetic analyses: comparing species to
infer adaptations and physiological mechanisms. Comprehensive Physiology 2, 639–674.

190
Richardson, K. C., and Wooller, R. D. (1986). The structures of the gastrointestinal tracts of
honeyeaters and other small birds in relation to their diets. Australian Journal of Zoology 34,
119–124.
Richardson, K. C., and Wooller, R. D. (1988). The alimentary tract of a specialist frugivore, the
Mistletoebird, Dicaeum hirundinaceum, in relation to its diet. Australian Journal of Zoology 36,
373–382.
Robertson, A. W., Kelly, D., Ladley, J. J., and Sparrow, A. D. (1999). Effects of pollinator loss
on endemic New Zealand mistletoes (Loranthaceae). Conservation Biology 13, 499–508.
Rowe, J. S. (1983). Concepts of fire effects on plant individuals and species. In 'The role of fire
effects in northern circumpolar ecosystems.' (Ed. D. A. Maclean.) pp. 135–154. (Wiley:
Chichester, UK.)
Sabat, P., and Gonzalez, S. P. (2003). Digestive enzymes in two species of marine Cinclodes
(Passeriformes: Furnariidae). The Condor 106, 830–833.
Salem, A. A., Coco, A. E., and Hendrix, T. R. (1965). Inhibition of absorption of D-xylose by
phlorizin in the anesthetized rat. American Journal of Physiology 209, 165–168.
Salle, G. (1983). Germination and establishment of Viscum album L. In 'The biology of
mistletoes.' (Eds. M. Calder and P. Bernhardt.) pp. 145–159. (Academic Press: Sydney.)
Salomon, L. L., Allums, J. A., and Smith, D. E. (1961). Possible carrier mechanism for the
intestinal transport of D-xylose. Biochemical and Biophysical Research Communications 4,
123–126.
Sargent, S. (1995). Seed fate in a tropical mistletoe: the importance of host twig size. Functional
Ecology 9, 197–204.
Saunders, D. A. (1993). Comunity-based observer scheme to assess avian responses to habitat
reduction and fragmentation in south-western Australia. Biological Conservation 64, 203–218.
Savory, C. J. (1992). Metabolic fates of U-14
C-labelled monosaccharides and an enzyme-treated
cell-wall substrate in the fowl. British Journal of Nutrition 67, 103–114.
Scharrer, E., and Grenacher, B. (2000). Na+-dependent transport of d-xylose by bovine intestinal
brush border membrane vesicles (BBMV) is inhibited by various pentoses and hexoses. Journal
of Veterinary Medicine Series A 47, 617–626.
Schondube, J. E., and Martinez del Rio, C. (2003). Concentration-dependent sugar preferences
in nectar-feeding birds: mechanisms and consequences. Functional Ecology 17, 445–453.
Schondube, J. E., and Martinez del Rio, C. (2004). Sugar and protein digestion in flowerpiercers
and hummingbirds: a comparative test of adaptive convergence. Journal of Comparative
Physiology B: Biochemical, Systemic, and Environmental Physiology 174, 263–273.
Schulze, E.-D., Turner, N. C., and Glatzel, G. (1984). Carbon, water and nutrient relations of
mistletoes and their hosts: a hypothesis. Plant, Cell and Environment 7, 293–299.
Schulze, E. D., Lange, O. L., Ziegler, H., and Gebauer, G. (1991). Carbon and nitrogen isotope
ratios of mistletoes growing on nitrogen and non-nitrogen-fixing hosts and on CAM plants in
the Namib desert confirm partial heterotrophy. Oecologia 88, 457–462.

191
Schutte, J. B. (1990). Nutritional implications and metabolizable energy value of D-xylose and
L-arabinose for broiler chicks. Poultry Science 69, 1724–1730.
Schutte, J. B., de Jong, J., van Weerden, E. J., and van Baak, M. J. (1992). Nutritional value of
D-xylose and L-arabinose for broiler chicks. British Poultry Science 33, 89–100.
Serventy, D. L. (1973). The Mistletoebird. Australian Wildlife Heritage 5, 744–747.
Shepherd, D. P., Beeston, G. R., and Hopkins, A. J. M. (2001). 'Native vegetation in Western
Australia, technical report 249.' (Department of Agriculture, Western Australia: South Perth.)
Smith, R. B., and Wass, E. F. (1979). Infection trials with three dwarf mistletoe species within
and beyond their known ranges in British Columbia. Canadian Journal of Plant Pathology 1,
47–57.
Snow, B. K., and Snow, D. W. (1988). 'Birds and berries: a study of an ecological interaction.'
(T. and A.D. Poyser: Carolton, UK.)
Sokal, R. R., and Rohlf, F. J. (1995). 'Biometry.' 3rd edn. ( W.H. Freeman & Co: New York.)
Stanley, M. C., and Lill, A. (2002). Does seed packaging influence fruit consumption and seed
passage in an avian frugivore. The Condor 104, 136–145.
Start, A. N. (2011). Fire responses and survival strategies of mistletoes (Loranthaceae) in an
arid environment in Western Australia. Australian Journal of Botany 59, 533–542.
StatSoft Inc (2007). 'STATISTICA (data analysis software system), version 8.0.' (Tulsa, OK.)
Available at www.statsoft.com
Stewart, G. R., and Press, M. C. (1990). The physiology and biochemistry of parasitic
angiosperms. Annual Review of Plant Physiology and Plant Molecular Biology 41, 127–151.
Stewart, G. R., Turnbullm, M. H., Schmidt, S., and Erskine, P. D. (1995). 13
C natural abundance
in plant communities along a rainfall gradient: a biological integrator of water availability.
Australian Journal of Plant Physiology 22, 51–55.
Stiles, F. G., and Freeman, C. E. (1993). Patterns in floral nectar characteristics of some bird-
visited plant species from Costa Rica. Biotropica 25, 191–205.
Sutherland, W. J., Freckleton, R. P., Godfray, H. C. J., Beissinger, S. R., Benton, T., Cameron,
D. D., Carmel, Y., Coomes, D. A., Coulson, T., Emmerson, M. C., Hails, R. S., Hays, G. C.,
Hodgson, D. J., Hutchings, M. J., Johnson, D., Jones, J. P. G., Keeling, M. J., Kokko, H., Kunin,
W. E., Lambin, X., Lewis, O. T., Malhi, Y., Mieszkowska, N., Milner-Gulland, E. J., Norris, K.,
Phillimore, A. B., Purves, D. W., Reid, J. M., Reuman, D. C., Thompson, K., Travis, J. M. J.,
Turnbull, L. A., Wardle, D. A., and Wiegand, T. (2013). Identification of 100 fundamental
ecological questions. Journal of Ecology 101, 58–67.
Symes, C. T., McKechnie, A. E., Nicolson, S. W., and Woodborne, S. M. (2011). The
nutritional significance of a winter‐flowering succulent for opportunistic avian nectarivores. Ibis
153, 110–121.
Tennakoon, K. U., and Pate, J. S. (1996). Effects of parasitism by a mistletoe on the structure
and functioning of branches of its host. Plant, Cell and Environment 19, 517–528.

192
Tennakoon, K. U., Chak, W. H., and Bolin, J. (2011). Nutritional and isotopic relationships of
selected Bornean tropical mistletoe- host associations in Brunei Darussalam. Journal of
Functional Plant Biology 38, 505–513.
Těšitel, J., Plavcová, L., and Cameron, D. D. (2010). Interactions between hemiparasitic plants
and their hosts. Plant Signaling & Behavior 5, 1072–1076.
Thomas, D. G. (1980). The bird community of Tasmanian temperates rainforest. Ibis 122, 298–
306.
Toft, N., Anderson, J., and Nowak, R. (1989). Water use efficiency and carbon isotope
composition of plants in a cold desert environment. Oecologia 80, 11–18.
Tracy, C. R., McWhorter, T. J., Korine, C., Wojciechowski, M. S., Pinshow, B., and Karasov,
W. H. (2007). Absorption of sugars in the Egyptian fruit bat (Rousettus aegyptiacus): A paradox
explained. Journal of Experimental Biology 210, 1726–1734.
Tsahar, E., Martínez del Rio, C., Izhaki, I., and Arad, Z. (2005). Can birds be ammonotelic?
Nitrogen balance and excretion in two frugivores. Journal of Experimental Biology 208, 1025–
1034.
Tsahar, E., Arad, Z., Izhaki, I., and Martínez del Rio, C. (2006). Do nectar- and fruit-eating
birds have lower nitrogen requirements than omnivores? An allometric test. The Auk 123, 1004–
1012.
Turner, R. J. (1991). Mistletoe in eucalypt forest-a resource for birds. Australian Forestry 54,
226–235.
van Ommeren, R. J., and Whitham, T. G. (2002). Changes in interactions between juniper and
mistletoe mediated by shared avian frugivores: parasitism to potential mutualism. Oecologia
130, 281–288.
Van Wyk, B.-E., and Nicolson, S. W. (1995). Xylose is a major nectar sugar in Protea and
Faurea. S Afr J Sci 91, 151–153.
Voigt, C. C., Baier, L., Speakman, J. R., and Siemers, B. M. (2008). Stable carbon isotopes in
exhaled breath as tracers for dietary information in birds and mammals. Journal of
Experimental Biology 211, 2233–2238.
Wagh, P. V., and Waibel, P. E. (1967). Alimentary absorption of L-arabinose and D-xylose in
chicks. Proceedings of the Society for Experimental Biology and Medicine 124, 421–424.
Walsberg, G. E. (1975). Digestive adaptations of Phainopepla nitens associated with the eating
of mistletoe berries. The Condor 77, 169–174.
Wang, L., Kgope, B., D’Odorico, P., and Macko, S. A. (2007). Carbon and nitrogen parasitism
by a xylem-tapping mistletoe (Tapinanthus oleifolius) along the Kalahari Transect: a stable
isotope study. African Journal of Ecology 46, 540–546.
Ward, M. J., and Paton, D. C. (2007). Predicting mistletoe seed shadow and patterns of seed
rain from movements of the mistletoebird, Dicaeum hirundinaceum. Austral Ecology 32, 113–
121.
Watson, D. M. (2001). Mistletoe – a keystone resource in forests and woodlands worldwide
Annual Review of Ecology and Systematics 32, 219–249.

193
Watson, D. M. (2002). Effects of mistletoe on diversity: a case-study from southern New South
Wales. Emu 102, 275–281.
Watson, D. M. (2003). The 'standardized search': an improved way to conduct bird surveys.
Austral Ecology 28, 515–525.
Watson, D. M. (2004). Comparative evaluation of new approaches to bird surveys. Wildlife
Research 31, 1–11.
Watson, D. M. (2011). 'Mistletoes of southern Australia.' (CSIRO Publishing: Collingwood,
VIC.)
Watson, D. M., McGregor, H. W., and Spooner, P. G. (2011). Hemiparasitic shrubs increase
resource availability and multi-trophic diversity of eucalypt forest birds. Functional Ecology 25,
889–899.
Watson, D. M., and Herring, M. (2012). Mistletoe as a keystone resource: an experimental test.
Proceedings of the Royal Society B 279, 3853–3860.
Welch, K. C., Bakken, B. H., Martínez de Rio, C., and Suarez, R. K. (2006). Hummingbirds
fuel hovering flight with newly ingested sugar. Physiological and Biochemical Zoology 79,
1082–1087.
Western Australian Herbarium (1998–). 'FloraBase- the Western Australian Flora.' (Department
of Environment and Conservation) Available at http://florabase.dec.wa.gov.au/
Whiting, G. C. (1970). Sugars. In ' The biochemistry of fruits and their products.' (Ed. H. A.C.)
pp. 1–31. (Academic Press: London and New York.)
Wilkinson, A. E. (1931). Habits of the silvereye. Emu XXXI, 157–159.
Wilson, A.-L., and Downs, C. T. (2011). Food preferences of Knysna and purple-crested turacos
fed varying concentrations of equicaloric and equimolar artificial fruit. Journal of Experimental
Biology 214, 613–618.
Wilson, A.-L., and Downs, C. T. (2012). Fruit nutritional composition and non-nutritive traits of
indigenous South African tree species. South African Journal of Botany 78, 30–36.
Witmer, M. C. (1994). Contrasting digestive strategies of frugivorous birds. Cornell University,
Ithaca, N.Y.
Witmer, M. C. (1996). The annual diet of the cedar waxwing described from U.S. Biological
Survey records (1885–1950): dietary patterns, digestive function, and natural history. The Auk
113, 414–430.
Witmer, M. C. (1998). Ecological and evolutionary implications of energy and protein
requirements of avian frugivores eating sugary diets. Physiological Zoology 71, 599–610.
Witmer, M. C., and van Soest, P. J. (1998). Contrasting digestive strageties of fruit-eating birds.
Functional Ecolgy 12, 728–741.
Witmer, M. C. (1999). Do avian frugivores absorb fruit sugars inefficiently? How dietary
nutrient concentration can affect coefficients of digestive efficiency. Journal of Avian Biology
30, 159–164.

194
Witmer, M. C., and Martínez del Rio, C. (2001). The membrane‐bound intestinal enzymes of
waxwings and thrushes: adaptive and functional implications of patterns of enzyme activity.
Physiological and Biochemical Zoology 74, 584–593.
Wolf, B., and Martínez del Rio, C. (2003). How important are columnar cacti as sources of
water and nutrients for desert consumers? A review. Isotopes in Environmental and Health
Studies 39, 53–67.
Wood, F. C., and Cahill, G. F. (1963). Mannose utilization in man. Journal of Clinical
Investigation 42, 1300–1312.
Wright, D. D. (2005). Diet, keystone resources and altitudinal movement of dwarf cassowaries
in relation to fruiting phenology in a Papua New Guinean rainforest. In 'Tropical fruits and
frugivores: the search for strong interactions.' (Eds. J. L. Dew and J. P. Boubli.) pp. 205–236.
(Springer: Dordrecht, The Netherlands.)
Yan, Z. (1993). Seed dispersal of Amyema preissii and Lysiana exocarpi by Mistletoebirds and
Spiny-cheeked Honeyeaters. Emu 93, 214–219.
Yan, Z., and Reid, N. (1995). Mistletoe (Amyema miquelii and A. pendulum) seedling
establishment on eucalypt hosts in eastern Australia. Journal of Applied Ecology 32, 778–784.
Yokota, S. D., Benyajati, S., and Dantzler, W. H. (1985). Comparative aspects of glomerular
filtration in vertebrates. Renal Physiology Basel 8, 193–221.
Zar, J. H. (1999). 'Biostatistical Analysis.' 4th edn. (Prentice-Hall: Upper Saddle River.)