“free” copper: a new endogenous chemical mediator of inflammation in birds
TRANSCRIPT

“Free” Copper: A New Endogenous Chemical Mediatorof Inflammation in Birds
Kaushal Kumar Singh & Mukesh Kumar & Pradeep Kumar &
Madhurendu Kumar Gupta & Dilip Kumar Jha & Suruchi Kumari &Birendra Kumar Roy & Sanjit Kumar
Received: 23 February 2011 /Accepted: 1 July 2011 /Published online: 22 September 2011# Springer Science+Business Media, LLC 2011
Abstract For acceptance of any chemical agent as anendogenous chemical mediator of inflammation, the agent inquestion must fulfill some biological requirements which are(a) it should be ubiquitously present in tissues in inactiveform, (b) it should be activated during process of inflamma-tion whose increase should be identifiable, (c) it should induceor amplify some events of inflammation, (d) there must besome natural inhibitor of such active form in tissues, (e) itshould be able to induce inflammatory reaction after exoge-nous injection, (f) such reaction should be inhibited byexogenous use of their antagonists, and (g) it should beamplified by use of agonists. Copper in its protein free orprotein bound form are reported to act as pathogenic factor ininflammatory processes due to oxidative stress. But their roleas endogenous chemical mediator of inflammation does notappear to be investigated thoroughly in light of above-mentioned biological criterion of mediator. Present studyaims at thorough exploration on role of free copper as
endogenous chemical mediator of inflammation in light ofabove facts. It was done by estimation of total copper, protein-bound copper, and free copper along with estimation of freeradical generation, increase in vascular permeability, andcellular infiltration during acute inflammatory reaction in-duced by carrageenan and concanavalin using chicken skin astest model. It was further evaluated by use of exogenous freecopper in experimental model and their subsequent inhibitionand amplification by chemical chelators of copper. Presentstudy confirms that free copper fulfilled all the biologicalrequirements for accepting it as an endogenous chemicalmediator of inflammation.
Keywords Free copper . Endogenous Chemical Mediator .
Evaluation . Oxygen-derived free radicals . Chemicalchelators
Introduction
Inflammation is the most effective event of both specific aswell as nonspecific immune reaction of the body, theultimate aim of which is to destroy and neutralize injuriousagents and repair the injury caused by them through ahighly complex cellular and vascular changes mediated bya number of endogenous factors accumulated at the site ofinjury. A thorough exploration of these factors hold crucialimportance because it can be exploited for enhancing itskilling effect for the organism and minimizing the risk oftissue damage caused by injurious agents or during theinflammatory reaction itself.
Copper is an essential trace element and acts as a criticalcofactor incorporated into specific cuproenzymes that catalyzeelectron transfer reactions required for cellular respiration,iron oxidation, pigment formation, neurotransmitter biosyn-
K. K. Singh (*) :M. Kumar : P. Kumar :M. K. Gupta :S. KumarDepartment of Veterinary Pathology,Faculty of Veterinary Science & AH,Birsa Agricultural University,Ranchi 834006 Jharkhand, Indiae-mail: [email protected]
D. K. JhaSRF, AICRP Poultry Breeding,Faculty of Veterinary Science & AH,Birsa Agricultural University,Ranchi 834006 Jharkhand, India
S. Kumari : B. K. RoyDepartment of Pharmacology & Toxicology,Faculty of Veterinary Science & AH,Birsa Agricultural University,Ranchi 834006 Jharkhand, India
Biol Trace Elem Res (2012) 145:338–348DOI 10.1007/s12011-011-9198-3

thesis, antioxidant, defensins, peptide amylation, and connec-tive tissue formation [16]. Copper has been found to showalteration in concentration due to a number of inflammatorydiseases. A number of acute and chronic inflammatoryconditions can result in hypercupremia [4, 7] while in manyothers, it has been associated with hypocupremia [3, 17].
The effects of hyper- and hypocupremia with regard toinflammation has not been defined and is still a matter ofdebate. This warrants the exploration of pathophysiologicalrole of copper at the site of inflammation. Protein free ionicform of copper can catalyze generation of superoxide byone electron reduction of molecular oxygen and furtherconvert them into .OH radicals through Fenton reaction [6],which are highly reactive chemical species capable of tissuedamage and thus may have disastrous consequences forbiological system. Consequently, copper in body is boundto or incorporated into various proteins to reduce its toxicityand are desequestrated wherever they are needed.
Free copper can be released from protein moieties due toaltered pH and ionic composition. During inflammation,there is altered pH and ionic concentration [6]. Hence,microenvironment of inflammation may cause release offree copper at that site. Superoxide generated due tooxidative stress during inflammation can also release freecopper from their protein moieties, leading to a viciouscycle of free radical and free copper generation. Thus, freecopper can be one of the important chemical speciesplaying a central role in generation and biotransformationof free radicals at the site of inflammation. One of the mostimportant roles of endogenous mediator of inflammation isgeneration or inhibition of other mediators. Copper in vivohas shown pro-oxidant [2, 10, 11] and antioxidant effects[9, 18]. So increase or decrease in level of copper duringdifferent types of inflammation finger towards its role inaccelerating and inhibiting the generation of free radicalsand may act as endogenous mediator of inflammation. Freeradicals have been recently included in mediator forum ofinflammation [6]. Despite recent induction of free radicalsin mediator forum of inflammation [6], the role of freecopper as endogenous chemical mediator of inflammationdoes not appear to be investigated thoroughly.
In this respect, it is to be noted that for acceptance of achemical as mediator of inflammation, the agent in questionmust fulfill some biological criterion. The criteria support-ing plausibility of a mediator are distribution and availabil-ity of agent in various tissues in inactive form [25], abilityto be activated during inflammation in keeping with itsalleged action [25], and presence of some endogenousnatural inhibitors, which keeps check on its harmful effects[1]. Moreover, the criterion which approves mediation isthat the agent should be able to induce inflammatory eventsafter exogenous injection [1] which must be inhibited byexogenous injection of antagonists [1].
Exploration of role of free copper as a new endogenouschemical mediator in light of these criteria will be helpful inunderstanding the mechanism underlying the debated role ofcopper in inflammation and oxidative stresses (Figs. 1, 2, 3, 4).
Hence, this study was made to evaluate the role of freecopper as endogenous mediator of inflammation capable ofmediating generation and potentiation of oxygen-derivedfree radicals (ODFRs), causing tissue damage and increasedvascular permeability during inflammation in light of abovecriterion using chicken skin as test model. Its role wasfurther confirmed by use of exogenous free copper inexperimental model and their subsequent inhibition andamplification by chemical chelators.
Methods
Chemicals
All chemicals used in this experiment were of analyticalgrade procured from Merck (if not mentioned specifically).
Experimental Design
The experiment was conducted in two phases.
Phase I
During this phase, an attempt was made to explore andassess the role of free copper in induced inflammation andthe chemical interaction between free copper and freeradicals with their biological effect at the site of inflamma-tion, induced by intradermal injection of concanavalin andcarrageenan using chicken skin as test model. The inflamedskin was subjected to estimation of (a) total copper, (b)protein-free copper, (c) protein-bound copper, (d) increasedvascular permeability, (e) generation of free radicals, and (f)microscopic tissue reaction.
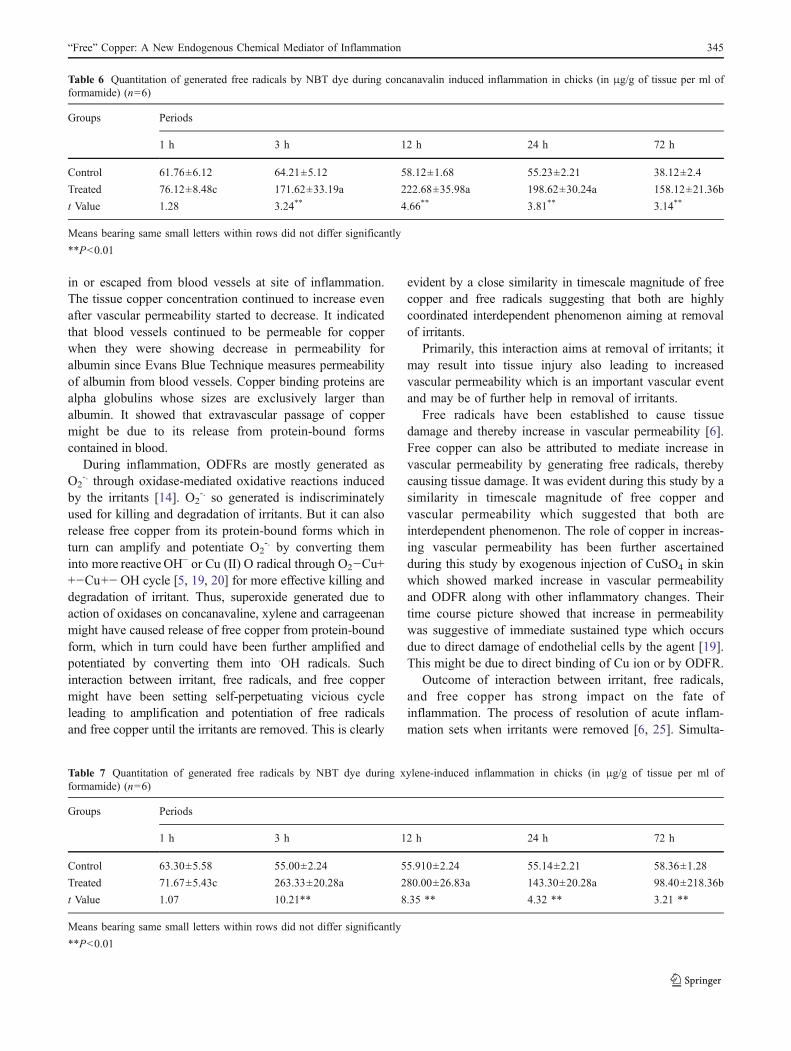
Fig. 1 Microscopic section of chicken skin after 72 h of intradermalinjection of copper sulfate showing heavy infiltration of mononuclearcells in perivascular as well as interstitial spaces of dermis. H&E ×400
“Free” Copper: A New Endogenous Chemical Mediator of Inflammation 339
In this phase, all estimations were done at 1, 3, 12, 24,and 72 h post-incubation at inflammatory site along withtheir respective control sites.
Phase II
The experiment of this phase aimed at confirming role of freecopper as a mediator of inflammation by assessment of theirability to induce inflammatory reaction by use of exogenousfree copper in experimental model and their subsequentinhibition and activation by copper binding/reacting agents.
Inflammation was induced by intradermal injection ofcopper sulfate alone and pretreated with copper-chelatingagent, ascorbic acid, and ethylenediaminetetraacetic acid(EDTA) using chicken skin as a test model, during which
1. Increase in vascular permeability2. Generation of free radicals3. Microscopic tissue reaction in inflammation was evaluated
at 15 min, 1, 3, 6, 24 h, days 3 and 6 after induction ofinflammation along with their respective control sites.
Experimental Procedures
Induction of Inflammation
One day before the inductions of inflammation, feathersfrom the lateral thoracic regions were plucked; the skincleared by cotton moistened with sterile distilled water andallowed to dry. On the following day, cleaning of the skinwas repeated. Local inflammatory reaction was elicited inseveral ways:
1. By intradermal injection of 0.1 ml of saline containing200 μg of carrageenan (Sigma)
2. By intradermal injection of 0.1 ml of saline containing200 μg of concanavalin (Merck)
3. By tropical application of Xylene4. By intradermal injection of 0.05 ml 2% copper sulfate
solution5. Prior to the intradermal injection of copper sulfate, 1 ml
of 4% ascorbic acid and 1 ml of 4% EDTA solutionwere inoculated separately subcutaneously.
6. Normal saline solution (0.1 ml) was inoculated ascontrol for the treated sites.
Estimation of vascular permeability
It was done as per the method of [12] using Evans Blue dyein place of Trypan Blue 36 during each phase. The methodin brief is as follows:
Cutaneous inflammatory lesions were produced byagents as per methods mentioned above; 0.5% Evans bluesolution was injected intravenously at 25 mg/kg bodyweight. Thirty minutes after dye injection, the dyed skinwas excised, weighed, and immersed in a tube containing4 ml formamide. The amount of dye extracted after 24 hof incubation at 37°C in formamide was determined byabsorption evaluation at 618 nm wavelength using
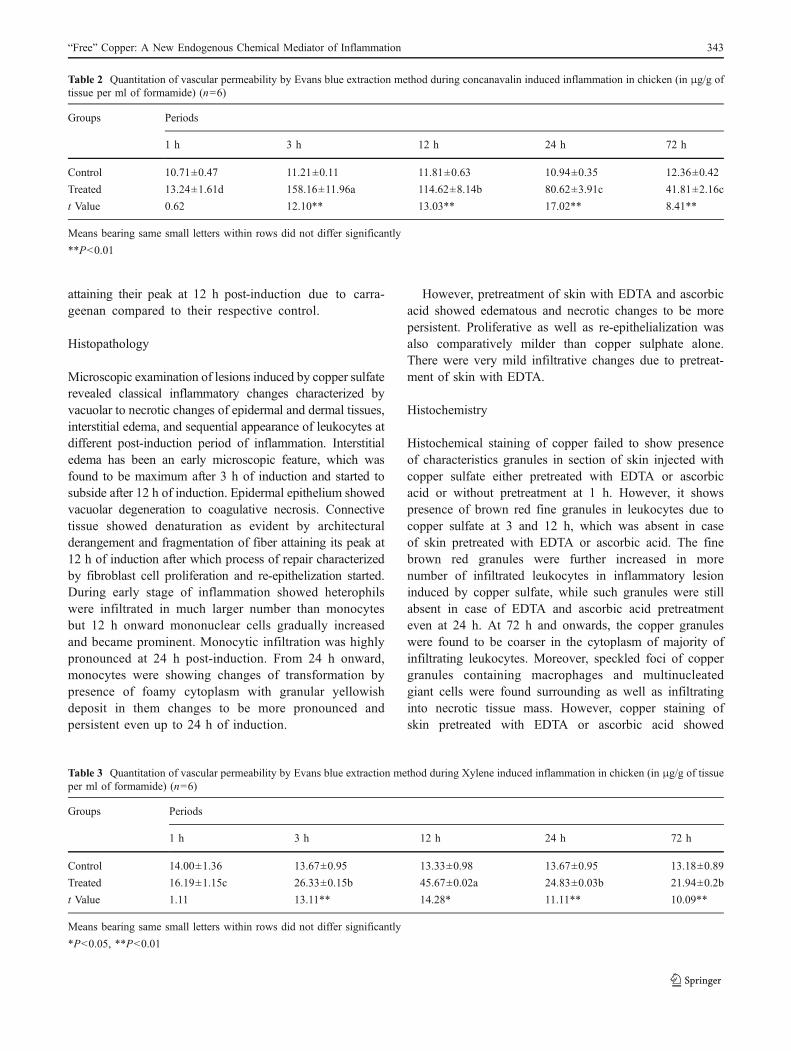
Fig. 4 Microscopic section of chicken skin after 24 h of intradermalinjection of copper sulphate showing dark-colored deposition incytoplasm of infiltrated mononuclear cells. Mallory and Parker’sHematoxylin stain ×400
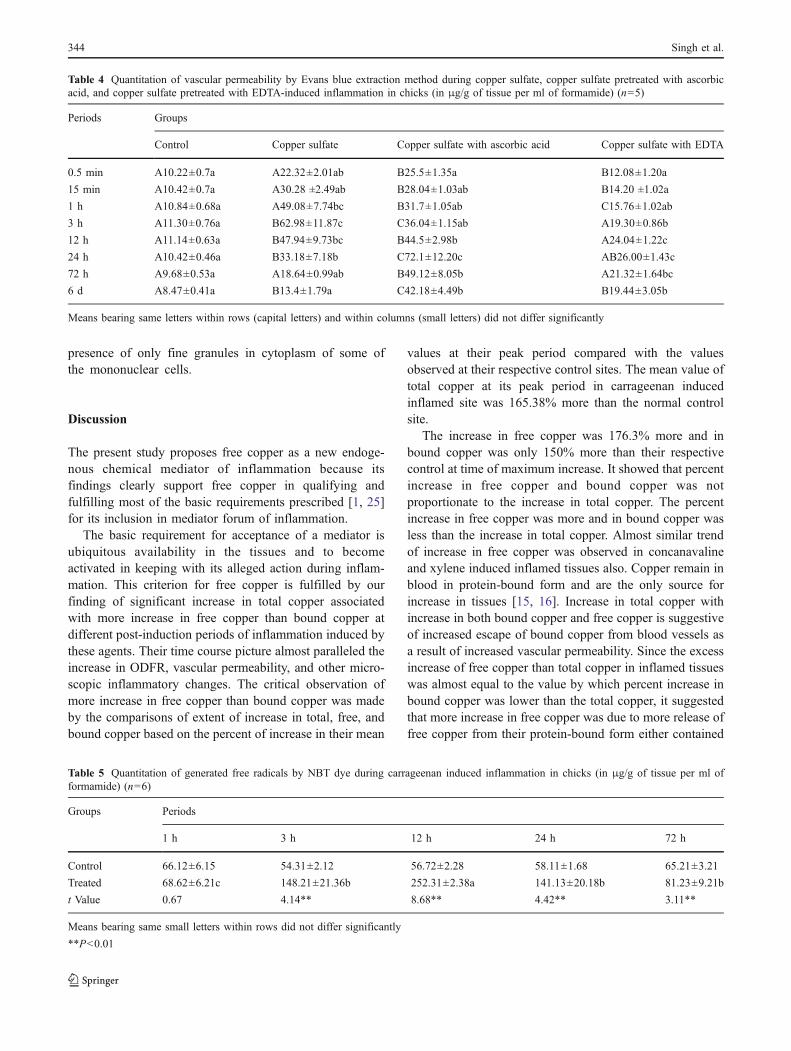
Fig. 3 Microscopic section of chicken skin after 24 h of intradermalinjection of copper sulphate showing marked infiltration of mononu-clear cells in superficial as well as in deep dermis. H&E ×400
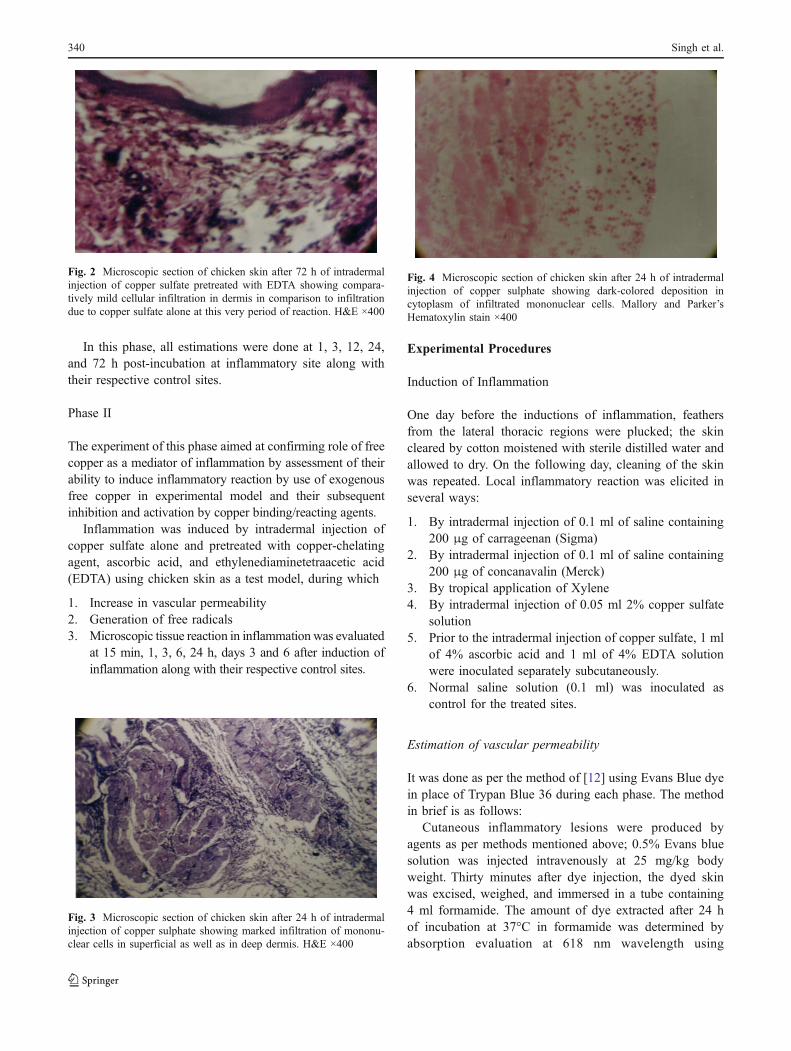
Fig. 2 Microscopic section of chicken skin after 72 h of intradermalinjection of copper sulfate pretreated with EDTA showing compara-tively mild cellular infiltration in dermis in comparison to infiltrationdue to copper sulfate alone at this very period of reaction. H&E ×400
340 Singh et al.

spectrophotometer. The concentration of the dye informamide was determined with the help of linear graphmade with known concentration of Evans blue informamide. Thereafter, the concentration of the dye intissues was calculated.
Estimation of Free Radicals
Quantitative estimation of free radicals was done by estimatingthe reduction of nitroblue tetrazolium (NBT) salt [23].
The method in brief is described as below:
1. Preparation of tissue homogenate—tissues collectedfrom the site were immediately kept in Hank’s balancesalt solution (H.B.S.S) containing 0.01 M EDTA permilliliter. After a few minutes, the tissue was pressedand rubbed in a wire mesh (200 G), and cellsuspensions were collected in H.B.S.S. Cells werewashed twice with H.B.S.S. A working suspension ofcell was made in a known volume of H.B.S.S.containing 2% bovine albumin.
2. Prepration of NBT Solution - It was prepared at aconcentration of 2 mg/ml in 0.13 M phosphate buffer(pH 7.2). The solution was shaken well for 5 min to getthe NBT salts dissolved.
3. Preparation of standard curve of NBT—0.5 ml of NBTsolution was added to 2 ml of 1 mM ascorbic acid in0.2 N sodium hydroxide. Insoluble residue wascollected on a 0.3-milipore filter. The filtrate wasweighed, dissolved in 2 ml formamide. With solution,a serial dilution of known and decreasing concentrationwas made. Their optical density was determined at580 nm, and a curve was drawn to determine theconcentration of unknown solution.
4. NBT reduction test—to two siliconized glass centrifugetubes were added 0.03 ml of 0.1 M freshly preparedpotassium azide in H.B.S.S. medium and 0.8 ml of thecell suspension; 0.8 ml of H.B.S.S. medium was added tothe first tube, and 0.8 ml of NBTsolution was added to thesecond tube. Both the tubes were shaken in a water bath at37°C. After 5 min of shaking at 37°C, 1 ml of ice-coldformal saline solution was added and shaken well. Twomilliliters of formamide was added, and the dye wasextracted by heating the extract at 85°C for 15 min. Theoptical densities of the solutes cleared by centrifugation at1,000×g for 15 min were determined at 580 nm.
Estimation of Total Copper, Protein-Free Copper,and Protein-Bound Copper
Total Copper - For estimation of total copper, weighedquantity of tissues were digested by ternary acid mixture asper method used by [21]. In brief, the samples were
digested at 50°C in 4 ml of ternary acid mixture of nitric,sulfuric, and perchloric acid in a proportion of 10:4:1. Thedigested materials were doubly diluted with distilled waterand filtered. The filtrates were used for determination ofcopper by GBC902 Double Beam Atomic AbsorptionSpectro Photometer at wavelength 248.3 nm using tripledistilled water as a blank. Copper concentration (microgramper gram) was calculated.
Free Copper - Quantitation of free copper was done byestimating the trichloroacetic acid (TCA) soluble copper inskin. It was done as per method described by [21]. In brief,a piece of about 500 mg of washed skin was taken andweighted accurately. It was thoroughly trichurated andhomogenized in 5 ml of distilled water. An equal volumeof 15% TCA solution was added to this 5 ml of trichuratedsolution of this skin. The precipitates were filtered, and thefiltered sample was utilized for the determination of TCAsoluble copper. The TCA soluble copper of the skin(microgram per gram) was calculated by multiplying thedilution factor with the concentration of copper recorded infiltered sample.
Bound form of Copper - It was determined by taking thedifference between total copper and free copper.
Histopathological Study
The skin sites bearing the lesions or control were collectedand fixed in 10% formal saline solution and were processedroutinely for paraffin embedding. The embedded tissueswere cut at 5-μm thickness. The slides were then stainedwith routine H&E stain. Leukocyte emigrations, cellular,vascular, and exudative responses along with other inflam-matory reactions were assessed by microscopic examinationof the section.
Histochemical Studies
Copper was stained by Mallory and Parker’s Hematoxylinmethod [8]. The method is briefed below:
The paraffin section was brought to distilled water andstained in fresh hematoxylin solution prepared bydissolving10mg hematoxylin in few drops of absolute alcoholfollowed by adding 10 ml of a filtered 2% potassiumdihydrogen phosphate. Then, it was washed in running waterfor 10 min, dehydrated, cleared, and mounted in DPX.
Statistical Analysis
Student’s t test was done to compare the value of treatedsite of different time intervals with the values in control
“Free” Copper: A New Endogenous Chemical Mediator of Inflammation 341

sites as per method of [22]. Analysis of variance and criticaldifference test were carried out to test the variation withinthe same group between periods as well as betweentreatments as per method [22].
Results
Quantitation of Vascular Permeability
Increase in vascular permeability due to injury induced bycarrageenan, concanavalin, xylene, copper sulfate, coppersulfate pretreated with ascorbic acid, and copper sulfatepretreated with EDTA along with their correspondingcontrol is shown in Tables 1, 2, 3, and 4. Table 1 showeda significant time-dependent increase in vascular perme-ability attaining peak at 3 h post-induction which wasabsent in corresponding control sites.
Table 2 showed a significant time-dependent increase,attaining peak at 3 h post-injury due to concanavalin.However, Table 3 showed a significant time-dependentincrease, attaining peak at 12 h post-injury due to xylene.Likewise, Table 4 indicated the peak value at 3 h post-induction of copper sulfate injury. The increase in vascularpermeability after copper sulfate with ascorbic acid treat-ment was comparatively more than their correspondingvalue of vascular permeability due to copper sulfate alone,while no such increase could be recorded from control sites.However copper sulfate with EDTA caused increase invascular permeability comparatively less than that founddue to copper sulfate alone.
Quantitation of Generated Free Radicals
The mean values of free radicals generated at different stags ofinflammation induced by carageenan, concanavalin, xylene,copper sulfate alone, and copper sulfate pretreated withascorbic acid and EDTA are shown in Tables 5, 6, 7, and8 along with their corresponding control. Table 5 showed asignificant time-dependent increase in ODFR generation due
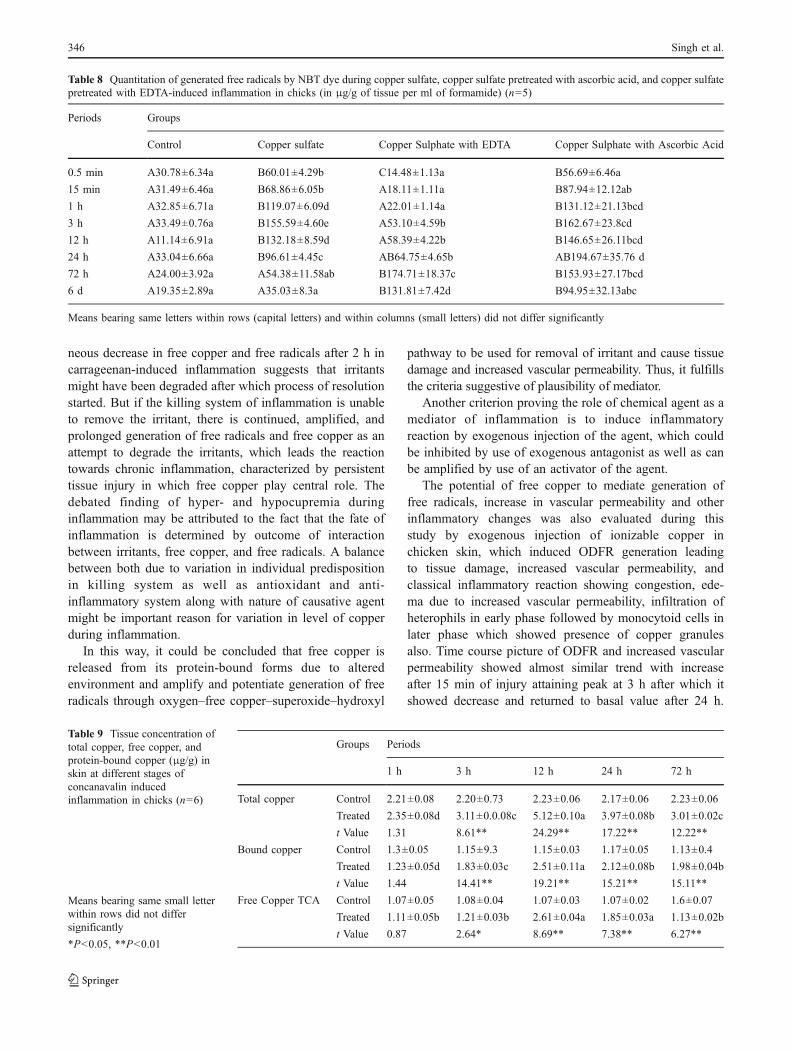
to carrageenan attaining peak at 12 h post-injury incomparison to their control. Table 6 showed a significanttime-dependent marked increase in ODFR generation attain-ing peak at 12 h post-injury in respect to their control.Table 7 showed a significant time-dependent increase inODFR generation due to xylene attaining peak at 12 h post-injury in comparison to their control. Table 8 showed thepeak value at 3 h post-injury. Such time-dependent increasewas not observed from their corresponding control sites. Themean value of generated free radical due to inflammatoryreaction induced by copper sulfate differed significantly atdifferent periods of observations as presented in Table 8.Their mean values were much higher than their correspondingcontrol sites. The peak value was observed at 3 h of inducedinjury. The peak value of free radicals generated due to coppersulfate pretreated with ascorbic acid as well as EDTA wasobserved at 24 and 72 h of injury, respectively. Moreover, theincrease in values of free radicals after copper sulfate withascorbic acid treatment was comparatively more than theircorresponding value of free radicals due to copper sulfatealone, while no such increase could be recorded from controlsites. However, copper sulfate with EDTA caused increasein values of free radicals comparatively less than thatfound due to copper sulfate alone.
Estimation of Total Copper, Protein-Free Copper,and Protein-Bound Copper
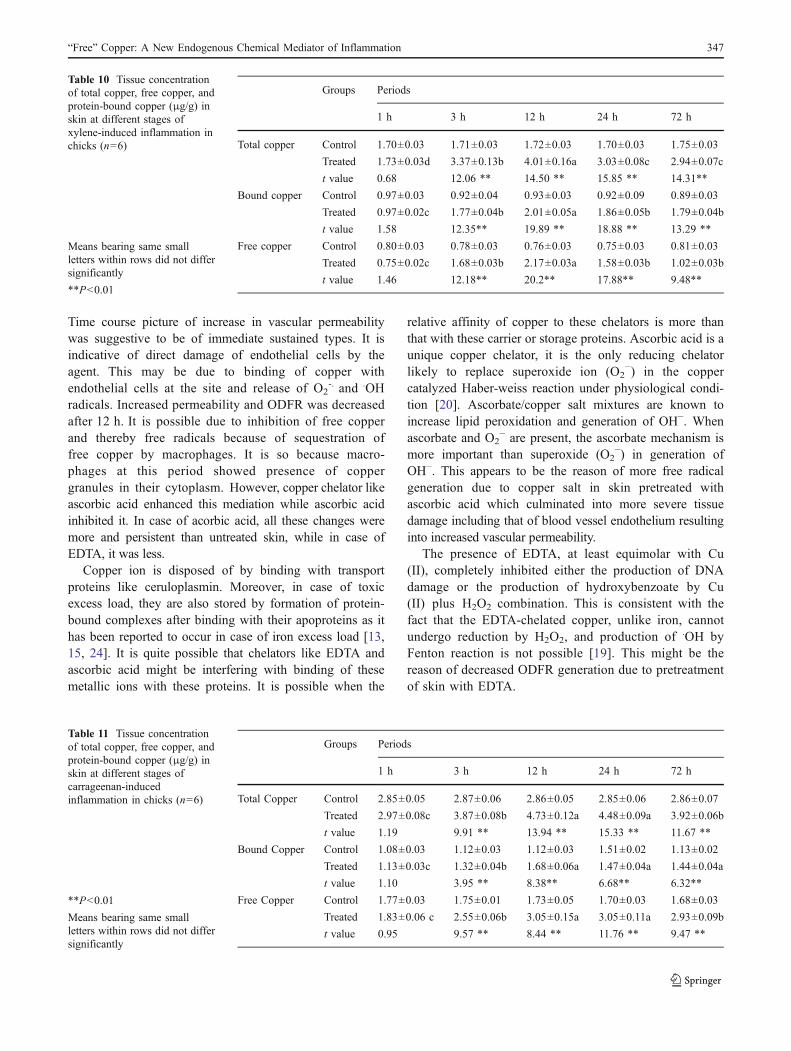
The mean concentration of total copper, protein-free copper,and protein-bound copper in tissue during concanavalin,xylene, and carrageenan induced inflammation at differentpost-induction periods along with their control is shown inTables 9, 10, and 11, respectively. Table 9 showed asignificant increase during early phase of inflammationattaining their peak at 12 h post-induction due to conca-navalin as compare to their respective control. Likewise,Table 10 showed a significant time dependent increase inthese values due to xylene as compare to control, attainingtheir peak at 12 h post-induction. Table 11 showed asignificant increase during early phase of inflammation
Table 1 Quantitation of vascular permeability by Evans blue extraction method during carrageenan induced inflammation in chicken (in μg/g oftissue per ml of formamide) (n=6)
Groups Periods
1 h 3 h 12 h 24 h 72 h
Control 11.23±0.12 10.94±0.92 10.72±0.48 11.94±0.36 10.82±0.36
Treated 12.5±0.79c 46.12±6.81a 42.96±0.89a 24.33±1.10b 18.42±1.11b
t Value (between control and treated) 1.46 35.21** 34.95** 10.31** 8.25**
Means bearing same small letters within rows did not differ significantly
**P<0.01
342 Singh et al.
attaining their peak at 12 h post-induction due to carra-geenan compared to their respective control.
Histopathology
Microscopic examination of lesions induced by copper sulfaterevealed classical inflammatory changes characterized byvacuolar to necrotic changes of epidermal and dermal tissues,interstitial edema, and sequential appearance of leukocytes atdifferent post-induction period of inflammation. Interstitialedema has been an early microscopic feature, which wasfound to be maximum after 3 h of induction and started tosubside after 12 h of induction. Epidermal epithelium showedvacuolar degeneration to coagulative necrosis. Connectivetissue showed denaturation as evident by architecturalderangement and fragmentation of fiber attaining its peak at12 h of induction after which process of repair characterizedby fibroblast cell proliferation and re-epithelization started.During early stage of inflammation showed heterophilswere infiltrated in much larger number than monocytesbut 12 h onward mononuclear cells gradually increasedand became prominent. Monocytic infiltration was highlypronounced at 24 h post-induction. From 24 h onward,monocytes were showing changes of transformation bypresence of foamy cytoplasm with granular yellowishdeposit in them changes to be more pronounced andpersistent even up to 24 h of induction.
However, pretreatment of skin with EDTA and ascorbicacid showed edematous and necrotic changes to be morepersistent. Proliferative as well as re-epithelialization wasalso comparatively milder than copper sulphate alone.There were very mild infiltrative changes due to pretreat-ment of skin with EDTA.
Histochemistry
Histochemical staining of copper failed to show presenceof characteristics granules in section of skin injected withcopper sulfate either pretreated with EDTA or ascorbicacid or without pretreatment at 1 h. However, it showspresence of brown red fine granules in leukocytes due tocopper sulfate at 3 and 12 h, which was absent in caseof skin pretreated with EDTA or ascorbic acid. The finebrown red granules were further increased in morenumber of infiltrated leukocytes in inflammatory lesioninduced by copper sulfate, while such granules were stillabsent in case of EDTA and ascorbic acid pretreatmenteven at 24 h. At 72 h and onwards, the copper granuleswere found to be coarser in the cytoplasm of majority ofinfiltrating leukocytes. Moreover, speckled foci of coppergranules containing macrophages and multinucleatedgiant cells were found surrounding as well as infiltratinginto necrotic tissue mass. However, copper staining ofskin pretreated with EDTA or ascorbic acid showed
Table 2 Quantitation of vascular permeability by Evans blue extraction method during concanavalin induced inflammation in chicken (in μg/g oftissue per ml of formamide) (n=6)
Groups Periods
1 h 3 h 12 h 24 h 72 h
Control 10.71±0.47 11.21±0.11 11.81±0.63 10.94±0.35 12.36±0.42
Treated 13.24±1.61d 158.16±11.96a 114.62±8.14b 80.62±3.91c 41.81±2.16c
t Value 0.62 12.10** 13.03** 17.02** 8.41**
Means bearing same small letters within rows did not differ significantly
**P<0.01
Table 3 Quantitation of vascular permeability by Evans blue extraction method during Xylene induced inflammation in chicken (in μg/g of tissueper ml of formamide) (n=6)
Groups Periods
1 h 3 h 12 h 24 h 72 h
Control 14.00±1.36 13.67±0.95 13.33±0.98 13.67±0.95 13.18±0.89
Treated 16.19±1.15c 26.33±0.15b 45.67±0.02a 24.83±0.03b 21.94±0.2b
t Value 1.11 13.11** 14.28* 11.11** 10.09**
Means bearing same small letters within rows did not differ significantly
*P<0.05, **P<0.01
“Free” Copper: A New Endogenous Chemical Mediator of Inflammation 343
presence of only fine granules in cytoplasm of some ofthe mononuclear cells.
Discussion
The present study proposes free copper as a new endoge-nous chemical mediator of inflammation because itsfindings clearly support free copper in qualifying andfulfilling most of the basic requirements prescribed [1, 25]for its inclusion in mediator forum of inflammation.
The basic requirement for acceptance of a mediator isubiquitous availability in the tissues and to becomeactivated in keeping with its alleged action during inflam-mation. This criterion for free copper is fulfilled by ourfinding of significant increase in total copper associatedwith more increase in free copper than bound copper atdifferent post-induction periods of inflammation induced bythese agents. Their time course picture almost paralleled theincrease in ODFR, vascular permeability, and other micro-scopic inflammatory changes. The critical observation ofmore increase in free copper than bound copper was madeby the comparisons of extent of increase in total, free, andbound copper based on the percent of increase in their mean
values at their peak period compared with the valuesobserved at their respective control sites. The mean value oftotal copper at its peak period in carrageenan inducedinflamed site was 165.38% more than the normal controlsite.
The increase in free copper was 176.3% more and inbound copper was only 150% more than their respectivecontrol at time of maximum increase. It showed that percentincrease in free copper and bound copper was notproportionate to the increase in total copper. The percentincrease in free copper was more and in bound copper wasless than the increase in total copper. Almost similar trendof increase in free copper was observed in concanavalineand xylene induced inflamed tissues also. Copper remain inblood in protein-bound form and are the only source forincrease in tissues [15, 16]. Increase in total copper withincrease in both bound copper and free copper is suggestiveof increased escape of bound copper from blood vessels asa result of increased vascular permeability. Since the excessincrease of free copper than total copper in inflamed tissueswas almost equal to the value by which percent increase inbound copper was lower than the total copper, it suggestedthat more increase in free copper was due to more release offree copper from their protein-bound form either contained
Table 4 Quantitation of vascular permeability by Evans blue extraction method during copper sulfate, copper sulfate pretreated with ascorbicacid, and copper sulfate pretreated with EDTA-induced inflammation in chicks (in μg/g of tissue per ml of formamide) (n=5)
Periods Groups
Control Copper sulfate Copper sulfate with ascorbic acid Copper sulfate with EDTA
0.5 min A10.22±0.7a A22.32±2.01ab B25.5±1.35a B12.08±1.20a
15 min A10.42±0.7a A30.28 ±2.49ab B28.04±1.03ab B14.20 ±1.02a
1 h A10.84±0.68a A49.08±7.74bc B31.7±1.05ab C15.76±1.02ab
3 h A11.30±0.76a B62.98±11.87c C36.04±1.15ab A19.30±0.86b
12 h A11.14±0.63a B47.94±9.73bc B44.5±2.98b A24.04±1.22c
24 h A10.42±0.46a B33.18±7.18b C72.1±12.20c AB26.00±1.43c
72 h A9.68±0.53a A18.64±0.99ab B49.12±8.05b A21.32±1.64bc
6 d A8.47±0.41a B13.4±1.79a C42.18±4.49b B19.44±3.05b
Means bearing same letters within rows (capital letters) and within columns (small letters) did not differ significantly
Table 5 Quantitation of generated free radicals by NBT dye during carrageenan induced inflammation in chicks (in μg/g of tissue per ml offormamide) (n=6)
Groups Periods
1 h 3 h 12 h 24 h 72 h
Control 66.12±6.15 54.31±2.12 56.72±2.28 58.11±1.68 65.21±3.21
Treated 68.62±6.21c 148.21±21.36b 252.31±2.38a 141.13±20.18b 81.23±9.21b
t Value 0.67 4.14** 8.68** 4.42** 3.11**
Means bearing same small letters within rows did not differ significantly
**P<0.01
344 Singh et al.
in or escaped from blood vessels at site of inflammation.The tissue copper concentration continued to increase evenafter vascular permeability started to decrease. It indicatedthat blood vessels continued to be permeable for copperwhen they were showing decrease in permeability foralbumin since Evans Blue Technique measures permeabilityof albumin from blood vessels. Copper binding proteins arealpha globulins whose sizes are exclusively larger thanalbumin. It showed that extravascular passage of coppermight be due to its release from protein-bound formscontained in blood.
During inflammation, ODFRs are mostly generated asO2
-. through oxidase-mediated oxidative reactions inducedby the irritants [14]. O2
-. so generated is indiscriminatelyused for killing and degradation of irritants. But it can alsorelease free copper from its protein-bound forms which inturn can amplify and potentiate O2
-. by converting theminto more reactive OH− or Cu (II) O radical through O2−Cu++−Cu+− OH cycle [5, 19, 20] for more effective killing anddegradation of irritant. Thus, superoxide generated due toaction of oxidases on concanavaline, xylene and carrageenanmight have caused release of free copper from protein-boundform, which in turn could have been further amplified andpotentiated by converting them into .OH radicals. Suchinteraction between irritant, free radicals, and free coppermight have been setting self-perpetuating vicious cycleleading to amplification and potentiation of free radicalsand free copper until the irritants are removed. This is clearly
evident by a close similarity in timescale magnitude of freecopper and free radicals suggesting that both are highlycoordinated interdependent phenomenon aiming at removalof irritants.
Primarily, this interaction aims at removal of irritants; itmay result into tissue injury also leading to increasedvascular permeability which is an important vascular eventand may be of further help in removal of irritants.
Free radicals have been established to cause tissuedamage and thereby increase in vascular permeability [6].Free copper can also be attributed to mediate increase invascular permeability by generating free radicals, therebycausing tissue damage. It was evident during this study by asimilarity in timescale magnitude of free copper andvascular permeability which suggested that both areinterdependent phenomenon. The role of copper in increas-ing vascular permeability has been further ascertainedduring this study by exogenous injection of CuSO4 in skinwhich showed marked increase in vascular permeabilityand ODFR along with other inflammatory changes. Theirtime course picture showed that increase in permeabilitywas suggestive of immediate sustained type which occursdue to direct damage of endothelial cells by the agent [19].This might be due to direct binding of Cu ion or by ODFR.
Outcome of interaction between irritant, free radicals,and free copper has strong impact on the fate ofinflammation. The process of resolution of acute inflam-mation sets when irritants were removed [6, 25]. Simulta-
Table 6 Quantitation of generated free radicals by NBT dye during concanavalin induced inflammation in chicks (in μg/g of tissue per ml offormamide) (n=6)
Groups Periods
1 h 3 h 12 h 24 h 72 h
Control 61.76±6.12 64.21±5.12 58.12±1.68 55.23±2.21 38.12±2.4
Treated 76.12±8.48c 171.62±33.19a 222.68±35.98a 198.62±30.24a 158.12±21.36b
t Value 1.28 3.24** 4.66** 3.81** 3.14**
Means bearing same small letters within rows did not differ significantly
**P<0.01
Table 7 Quantitation of generated free radicals by NBT dye during xylene-induced inflammation in chicks (in μg/g of tissue per ml offormamide) (n=6)
Groups Periods
1 h 3 h 12 h 24 h 72 h
Control 63.30±5.58 55.00±2.24 55.910±2.24 55.14±2.21 58.36±1.28
Treated 71.67±5.43c 263.33±20.28a 280.00±26.83a 143.30±20.28a 98.40±218.36b
t Value 1.07 10.21** 8.35 ** 4.32 ** 3.21 **
Means bearing same small letters within rows did not differ significantly
**P<0.01
“Free” Copper: A New Endogenous Chemical Mediator of Inflammation 345
neous decrease in free copper and free radicals after 2 h incarrageenan-induced inflammation suggests that irritantsmight have been degraded after which process of resolutionstarted. But if the killing system of inflammation is unableto remove the irritant, there is continued, amplified, andprolonged generation of free radicals and free copper as anattempt to degrade the irritants, which leads the reactiontowards chronic inflammation, characterized by persistenttissue injury in which free copper play central role. Thedebated finding of hyper- and hypocupremia duringinflammation may be attributed to the fact that the fate ofinflammation is determined by outcome of interactionbetween irritants, free copper, and free radicals. A balancebetween both due to variation in individual predispositionin killing system as well as antioxidant and anti-inflammatory system along with nature of causative agentmight be important reason for variation in level of copperduring inflammation.
In this way, it could be concluded that free copper isreleased from its protein-bound forms due to alteredenvironment and amplify and potentiate generation of freeradicals through oxygen–free copper–superoxide–hydroxyl
pathway to be used for removal of irritant and cause tissuedamage and increased vascular permeability. Thus, it fulfillsthe criteria suggestive of plausibility of mediator.
Another criterion proving the role of chemical agent as amediator of inflammation is to induce inflammatoryreaction by exogenous injection of the agent, which couldbe inhibited by use of exogenous antagonist as well as canbe amplified by use of an activator of the agent.
The potential of free copper to mediate generation offree radicals, increase in vascular permeability and otherinflammatory changes was also evaluated during thisstudy by exogenous injection of ionizable copper inchicken skin, which induced ODFR generation leadingto tissue damage, increased vascular permeability, andclassical inflammatory reaction showing congestion, ede-ma due to increased vascular permeability, infiltration ofheterophils in early phase followed by monocytoid cells inlater phase which showed presence of copper granulesalso. Time course picture of ODFR and increased vascularpermeability showed almost similar trend with increaseafter 15 min of injury attaining peak at 3 h after which itshowed decrease and returned to basal value after 24 h.
Table 8 Quantitation of generated free radicals by NBT dye during copper sulfate, copper sulfate pretreated with ascorbic acid, and copper sulfatepretreated with EDTA-induced inflammation in chicks (in μg/g of tissue per ml of formamide) (n=5)
Periods Groups
Control Copper sulfate Copper Sulphate with EDTA Copper Sulphate with Ascorbic Acid
0.5 min A30.78±6.34a B60.01±4.29b C14.48±1.13a B56.69±6.46a
15 min A31.49±6.46a B68.86±6.05b A18.11±1.11a B87.94±12.12ab
1 h A32.85±6.71a B119.07±6.09d A22.01±1.14a B131.12±21.13bcd
3 h A33.49±0.76a B155.59±4.60e A53.10±4.59b B162.67±23.8cd
12 h A11.14±6.91a B132.18±8.59d A58.39±4.22b B146.65±26.11bcd
24 h A33.04±6.66a B96.61±4.45c AB64.75±4.65b AB194.67±35.76 d
72 h A24.00±3.92a A54.38±11.58ab B174.71±18.37c B153.93±27.17bcd
6 d A19.35±2.89a A35.03±8.3a B131.81±7.42d B94.95±32.13abc
Means bearing same letters within rows (capital letters) and within columns (small letters) did not differ significantly
Table 9 Tissue concentration oftotal copper, free copper, andprotein-bound copper (μg/g) inskin at different stages ofconcanavalin inducedinflammation in chicks (n=6)
Means bearing same small letterwithin rows did not differsignificantly
*P<0.05, **P<0.01
Groups Periods
1 h 3 h 12 h 24 h 72 h
Total copper Control 2.21±0.08 2.20±0.73 2.23±0.06 2.17±0.06 2.23±0.06
Treated 2.35±0.08d 3.11±0.0.08c 5.12±0.10a 3.97±0.08b 3.01±0.02c
t Value 1.31 8.61** 24.29** 17.22** 12.22**
Bound copper Control 1.3±0.05 1.15±9.3 1.15±0.03 1.17±0.05 1.13±0.4
Treated 1.23±0.05d 1.83±0.03c 2.51±0.11a 2.12±0.08b 1.98±0.04b
t Value 1.44 14.41** 19.21** 15.21** 15.11**
Free Copper TCA Control 1.07±0.05 1.08±0.04 1.07±0.03 1.07±0.02 1.6±0.07
Treated 1.11±0.05b 1.21±0.03b 2.61±0.04a 1.85±0.03a 1.13±0.02b
t Value 0.87 2.64* 8.69** 7.38** 6.27**
346 Singh et al.
Time course picture of increase in vascular permeabilitywas suggestive to be of immediate sustained types. It isindicative of direct damage of endothelial cells by theagent. This may be due to binding of copper withendothelial cells at the site and release of O2
-. and .OHradicals. Increased permeability and ODFR was decreasedafter 12 h. It is possible due to inhibition of free copperand thereby free radicals because of sequestration offree copper by macrophages. It is so because macro-phages at this period showed presence of coppergranules in their cytoplasm. However, copper chelator likeascorbic acid enhanced this mediation while ascorbic acidinhibited it. In case of acorbic acid, all these changes weremore and persistent than untreated skin, while in case ofEDTA, it was less.
Copper ion is disposed of by binding with transportproteins like ceruloplasmin. Moreover, in case of toxicexcess load, they are also stored by formation of protein-bound complexes after binding with their apoproteins as ithas been reported to occur in case of iron excess load [13,15, 24]. It is quite possible that chelators like EDTA andascorbic acid might be interfering with binding of thesemetallic ions with these proteins. It is possible when the
relative affinity of copper to these chelators is more thanthat with these carrier or storage proteins. Ascorbic acid is aunique copper chelator, it is the only reducing chelatorlikely to replace superoxide ion (O2
−) in the coppercatalyzed Haber-weiss reaction under physiological condi-tion [20]. Ascorbate/copper salt mixtures are known toincrease lipid peroxidation and generation of OH−. Whenascorbate and O2
− are present, the ascorbate mechanism ismore important than superoxide (O2
−) in generation ofOH−. This appears to be the reason of more free radicalgeneration due to copper salt in skin pretreated withascorbic acid which culminated into more severe tissuedamage including that of blood vessel endothelium resultinginto increased vascular permeability.
The presence of EDTA, at least equimolar with Cu(II), completely inhibited either the production of DNAdamage or the production of hydroxybenzoate by Cu(II) plus H2O2 combination. This is consistent with thefact that the EDTA-chelated copper, unlike iron, cannotundergo reduction by H2O2, and production of .OH byFenton reaction is not possible [19]. This might be thereason of decreased ODFR generation due to pretreatmentof skin with EDTA.
Table 10 Tissue concentrationof total copper, free copper, andprotein-bound copper (μg/g) inskin at different stages ofxylene-induced inflammation inchicks (n=6)
Means bearing same smallletters within rows did not differsignificantly
**P<0.01
Groups Periods
1 h 3 h 12 h 24 h 72 h
Total copper Control 1.70±0.03 1.71±0.03 1.72±0.03 1.70±0.03 1.75±0.03
Treated 1.73±0.03d 3.37±0.13b 4.01±0.16a 3.03±0.08c 2.94±0.07c
t value 0.68 12.06 ** 14.50 ** 15.85 ** 14.31**
Bound copper Control 0.97±0.03 0.92±0.04 0.93±0.03 0.92±0.09 0.89±0.03
Treated 0.97±0.02c 1.77±0.04b 2.01±0.05a 1.86±0.05b 1.79±0.04b
t value 1.58 12.35** 19.89 ** 18.88 ** 13.29 **
Free copper Control 0.80±0.03 0.78±0.03 0.76±0.03 0.75±0.03 0.81±0.03
Treated 0.75±0.02c 1.68±0.03b 2.17±0.03a 1.58±0.03b 1.02±0.03b
t value 1.46 12.18** 20.2** 17.88** 9.48**
Table 11 Tissue concentrationof total copper, free copper, andprotein-bound copper (μg/g) inskin at different stages ofcarrageenan-inducedinflammation in chicks (n=6)
**P<0.01
Means bearing same smallletters within rows did not differsignificantly
Groups Periods
1 h 3 h 12 h 24 h 72 h
Total Copper Control 2.85±0.05 2.87±0.06 2.86±0.05 2.85±0.06 2.86±0.07
Treated 2.97±0.08c 3.87±0.08b 4.73±0.12a 4.48±0.09a 3.92±0.06b
t value 1.19 9.91 ** 13.94 ** 15.33 ** 11.67 **
Bound Copper Control 1.08±0.03 1.12±0.03 1.12±0.03 1.51±0.02 1.13±0.02
Treated 1.13±0.03c 1.32±0.04b 1.68±0.06a 1.47±0.04a 1.44±0.04a
t value 1.10 3.95 ** 8.38** 6.68** 6.32**
Free Copper Control 1.77±0.03 1.75±0.01 1.73±0.05 1.70±0.03 1.68±0.03
Treated 1.83±0.06 c 2.55±0.06b 3.05±0.15a 3.05±0.11a 2.93±0.09b
t value 0.95 9.57 ** 8.44 ** 11.76 ** 9.47 **
“Free” Copper: A New Endogenous Chemical Mediator of Inflammation 347

In this way, it could be concluded that increase in boundcopper in extravascular space as a result of initial increase invascular permeability due to initial injury might be releasingfree copper under the influence of altered pH, ionic conc.,and generation of O2
-.. This in turn caused amplification andpotentiation of ODFR and free copper generation to be usedfor removal of irritant as well as tissue damage and therebyprolongation and amplification of increased vascular perme-ability. These can be inhibited by copper-binding proteins ortheir sequestration in phagocytic cells. Ability of free copperwas further confirmed by exogenous injection of ionizablecopper which increased ODFR generation, increased vascu-lar permeability, and other classical inflammatory signs.
In this way, it fulfills almost all the requirements foracceptability as endogenous inflammatory mediator.
Acknowledgements Authors acknowledge the financial support pro-vided by Dr N. N. Singh, Vice Chancellor, Birsa Agricultural University,and Dr S. K. Singh, Dean, Ranchi Veterinary College, Kanke, Ranchi,Jharkhand, India, for conducting the present experiment.
References
1. Anderson JR (1987) Muir’s Textbook of pathology, vol 12. ELBS,London, pp 41–438
2. Andrewartha KA, Caple IW (1980) Effects of changes innutritional copper on erythrocyte superoxide dismutase activityin sheep. Res Vet Sci 28:101–104
3. Cartwright GE, Gubler CJ, Wintrobe MM (1954) Studies oncopper metabolism. J Clin Invest 33(4):685–698
4. Clegg MS, Ferrel F, Keen CL (1987) Hypertension inducedalterations in copper and zinc metabolism in Dahl rats. Hypertension9:624–628
5. Cohen G (1985) In: Greenwald RA (ed) In Handbook of methodsof oxygen radicals research. CRC, Boca Raton, pp 55–64
6. Cotran RS, Majno G (1964) The delayed and prolonged vascularleakage in inflammation, topography of the leaking vessels afterthermal injury. Am J Pathol 45:261–281
7. Cousins RJ (1985) Absorption, transport, and hepatic metabolismof copper and zinc: special reference to metallothionein andceruloplasmin. Physiol Rev 65:238–309
8. Culling CFA, Allison RT, Barr WT (1985) Cellular pathology, 9thedn. Butterworth, Maple Ridge
9. DiSilvestro RA, Elizabeth JMS, Yanga FL (1992) Copperdeficiency impairs survival in endotoxin-treated rats. Nutr Res12:501–508
10. Dougherty JJ, Hoekstra WG (1982) Effects of vitamin E andselenium on copper induced lipid peroxidation in vivo and onacute copper toxicity. Proc Soc Exp Biol Med 169:201–208
11. Hochstein P, Kumar KS, Forman SJ (1980) Lipid peroxidation andthe cytotoxicity of copper. Ann NY Acad Sci 355:240–248
12. Ito NMK, Bohm GM (1986) Turpentine induced acute inflamma-tory response in gallus gallus, oedema, vascular permeability andeffects of non-steroidal anti-inflammatory drugs. Res Vet Sci41:231–236
13. Johnson GF, Morell AG, Stockert RJ (1981) Hepatic lysosomalcopper-protein in dogs with an inherited copper toxicosis.Hepaology 1:1243–1248
14. Kafy ML, Haigh CG, Lewis DA (1986) In vitro interactionsbetween dangerous endogenous polyamines and superoxide anion.Agents Actions 18(516):555–559
15. Kaneko JJ (1979) Clinical Biochemistry of Domestic Animals, vol4. Academic, London, pp 257–258
16. Keen CL, Graham TW (1979) Trace elements. In: Kaneko JJ (ed)Clinical Biochemistry of Domestic Animals, 4th edn. Academic,New York, pp 753–793
17. Lea CM, Luttrell VAS (1965) Copper content of hair inKwashiorkor. Nature 206:413
18. Marklund SL (1984) Extracellular superoxide dismutase and othersuperoxide dismutase isoenzymes in tissues from nine mammalianspecies. Biochem J 222:649–655
19. Mukherjee U, Chaterjee SN (1994) Oxidative damage of DNAand benzoate by chelated and non-chelated copper in presence ofhydrogen peroxide. I J Biochem Biophy 32:32–36
20. Prabhu HR, Krishnamurthy S (1993) Ascorbate dependentformation of hydroxyl radicals in the presence of iron chelates.Indian J Biochem Biophys 30:289–292
21. Singh KK, Gupta MK, Ram M, Singh V, Roy BK (2010)Effect of Chronic Fenvalerate Intoxication on Tissue Concen-tration of Copper in Goats and further Exploration on ItsMechanism. Biol Trace Elem Res. doi:10.1007/s1211-010-8629-x
22. Snedecor GW, Cochran WG (1994) Statistical Methods, vol 6.Iowa State University Press, Ames, pp 258–298
23. Stossel TP (1973) Evaluation of opsonic and leukocyte functionwith a spectrophotometric test in patients with infection andphagocytic disorders. Blood 42:121–130
24. Theil EC (1987) Ferritin structure gene regulation and cellularfunction in animals, plants and microorganisms. Ann RevBiochem 56:289–315
25. Walter JB, Israel MS (1987) General pathology, vol 6. ChurchillLivingston, New York, pp 81–96
348 Singh et al.