field crops research crop physiology and productivity
TRANSCRIPT

E L S E V I E R Field Crops Research 53 (1997) 3-16
Field Crops Research
Crop physiology and productivity
Marie-Hrlbne Jeuffroy *, Bertrand Ney Laboratoire d'Agronomie, INRA-INA PG, 78 850 Thiverval-Grignon, France
Abstract
This paper reviews aspects of crop physiology and productivity for selected grain legumes. Vegetative development, including phases of leaf area increase and branching are described, then, the main reproductive stages and their progression along the stem are discussed. The effects of water and nitrogen shortage on reproductive development are briefly described. A model for reproductive development along a stem is proposed and applied to several grain legumes, and effects of genetic variability are discussed. Growth, and its analysis in terms of intercepted radiation and radiation use efficiency are then reviewed. The variability of these two components is analysed according to differences due to species, genotypes (mainly characterized by different foliage structures), environmental conditions and methods of measurement. Yield is then analysed as a direct consequence of crop growth. Finally, a pattern of assimilate partitioning is described, and its consequences for reproductive structure formation, i.e. the grain number on each node of the stem, are discussed. © 1997 Elsevier Science B.V.
Keywords: Grain legume; Development; Growth; Assimilate partitioning; Yield determination
1. Introduct ion
Yields of grain legumes are smaller and generally more variable than those of many other crop species. In developed countries, grain yields of legumes have not increased as rapidly as those of cereal crops. Between 1940 and 1981, for instance, winter wheat yields in the UK increased at approximately twice the rate of those for pea and faba bean (82, 44 and 31 kg h a - l year-1, respectively) (Heath and Heb- blethwaite, 1985a). In France, between 1981 and 1996, yield increases were 75 kg ha -1 year -1 for pea and 120 for wheat (Carroure, 1996). There is thus a need to increase the performance of pulse
* Corresponding author. Tel.: (+33)1-30-81-55-36; fax: ( + 33)1-30-81-55-64; e-mail: jeuffroy @bcgn.grignon.inra.fr.
crops, particularly in developing countries, where most grain legume production is for human con- sumption and demand is increasing due to population increase. Oram and Agcaoili (1992) estimated that yields in developing countries were only 45% of those of developed countries for pea, and 75% for faba bean and chickpea.
The agronomic problems associated with pulse crops differ between geographical areas. In the As ia-Afr ica-Oceania regions (Johansen et al., 1992), North America (Slinkard et al., 1992), and Europe (Monti et al., 1992), drought and biotic stresses appear to be the major limiting factors, while other stresses (such as extreme temperatures and nutrient deficiencies) have less frequent impact. Drought and biotic stresses can be alleviated in various ways, for example by breeding or crop management. Although resistance cultivars can be bred to combat diseases,
0378-4290/97/$17.00 © 1997 Elsevier Science B.V. All rights reserved. PII S0378-4290(97)00019- 1

4 M.-H. Jeuffroy, B. Ney/Field Crops Research 53 (1997) 3-16
often with time pathogens evolve to overcome the resistance. Irrigation and fungicidal sprays are not always available in developing countries to prevent drought or control foliar diseases.
Another approach is to escape stress. Damage due to drought can be reduced if the period of crop sensitivity does not fall in the period of water deficit (Ney et al., 1994). This type of approach requires an improved understanding of crop physiology and of plant responses to environment. This chapter reviews current knowledge of phenology, growth and of plant structure determination, and their consequences on yield formation.
2. Development
2.1. Vegetative development
2.1.1. Plastochron For pea and many other species, successive nodes
are initiated in the apex at a constant rate throughout the crop cycle, the plastochron, calculated in cumula- tive degree-days (Lyndon, 1977; Dracup and Kirby, 1993; Jeuffroy, 1991; Lecoeur et al., 1995), with 0°C as a base temperature (Etrv6 and Derieux, 1982). Thermal time units have been widely used to de- scribe the progress of crop development in legumes and other crops (Wilhelm and McMaster, 1995). Although other factors, such as daylength, soil com- paction and nitrogen nutrition also affect the rate of leaf emergence, their influence is minor (Kirby, 1995). The end of nodal initiation varies according to the situation, but the final number of initiated nodes is, at least in the situations observed, greater than the final number of visible nodes (Lecoeur, 1994; Jeuf- froy and Sebillotte, 1997).
2.1.2. Phyllochron Development of successive nodes along a stem is
also constant throughout the vegetative stage in cu- mulative degree-days, determining the phyllochron (Leong and Ong, 1983, for peanut; Truong and Duthion, 1993, for pea). This rate is enhanced fol- lowing floral initiation (Lecoeur, 1994). Variation of pea leaf appearance rate in the field is due to differ- ences in mean temperature and plant growth rate (Truong and Duthion, 1993).
2.1.3. Leaf growth and leaf area Individual leaf area depends on cell number and
cell size. The changes with time of cell number and leaf area for individual leaves of pea (Pisum sativum L.) show two distinct and successive phases: phase 1, during 2 / 3 of leaf development, involves cell division, while phase 2, during the last 1 /3 of leaf development, involves cell expansion (Lecoeur et al., 1995). The effects of water deficit on leaf area have been studied at different periods in the leaf life. Early deficit during phase 1 reduces leaf area through an effect on cell number, while deficit during phase 2 reduces leaf area by reducing cell area (Lecoeur et al., 1995). These results suggest that long-term tem- poral analysis may be necessary to study dicot leaf expansion, whereas spatial analysis is more effective for monocot leaves. A scale was proposed by Maurer et al. (1966) to describe the macroscopic develop- ment of a pea leaf.
2.1.4. Branching Branch number in leguminous plants is highly
variable, and is an important determinant of grain yield. Low plant densities can be compensated by substantial branching (Hedley and Ambrose, 1981). Plant structure varies among cultivated grain legumes. Pea and soybean branches appear mainly on the first vegetative nodes, whereas white lupin produces branches at the top of the main stem. For lupins, several orders of branches develop sequen- tially as described by Farrington and Greenwood (1975), but branches of the same order develop synchronously. Branching capacities vary among genotypes. Genotypes with greater branching ability generally have more determinate habits, probably because of the competition between branches in the canopy.
Branching is also strongly influenced by environ- mental conditions such as soil physical conditions or soil water status (Dawkins et al., 1984). Environmen- tal conditions can modify the contributions of branches to final yield (Duthion et al., 1994, for white lupin). The potential number of branches of a given order on white lupin depends on the number of axillary buds on the stems of the previous order. The number of branches produced is linked to the plant growth rate at the beginning of branch elongation (Munier-Jolain et al., 1996a).

M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16 5
The branching pattern of peanut has been de- scribed by Gupton et al. (1968) and Bunting et al. (1985). The first lateral branches appear at about the time that the second true node appears on the main stem, and the rate of appearance of the other primary branches is about half that of node development on the main stem (Leong and Ong, 1983). Very high mean temperatures inhibit branching of peanut (Leong and Ong, 1983).
2.2. Reproductive development
2.2.1. The reproductive stages The development of legume seeds has been de-
scribed in several studies (Dure, 1975; Pate, 1975; Pate and Flinn, 1977). An initial phase of cell divi- sions is followed by a period of cell expansion, and then by an accumulation of reserve compounds in the cells. Four main stages can be defined in the life of a reproductive organ: (1) organ initiation, (2) anthesis, (3) final stage in seed abortion and (4) physiological maturity. Floral initiation is very early in the plant life (about 4-6-leaf stage). Anthesis of a flower is the time when 'petals are opened', accord- ing to the scale of Maurer et al. (1966) and is later than ovule fertilization by about 24 hours (Cooper, 1938; Linck, 1961). For an ovule, Pigeaire et al. (1986) demonstrated the existence of a stage after
which the probability of seed abortion is lower than 5%, called the final stage in seed abortion (FSSA). This stage corresponds to the transition between a period of active cell division and a phase of rapid dry matter accumulation in the seed (Ney et al., 1993). The first period, during which the seed en- larges rapidly, lasts about 300 degree-days after an- thesis (base temperature of 0°C). At the end of this phase, the final cell number is reached in the ovule. At the pod scale, the final stage in seed abortion can be defined as the time when all the ovules of the pod have reached this stage. At FSSA, the seed number in the pod is determined. Thereafter, dry matter accumulation in the seed correlates with cumulative degree-days. Two criteria can be used to determine the time of transition between these main phases, seed number formation and seed filling. First, Duthion and Pigeaire (1991) have shown, on a great number of seeds, that there is a maximum length for aborted seeds, 8.8 mm for cultivar Solara for exam- ple. Ney et al. (1993) confirmed that, at this length, the final cell number of the embryo was reached. Second, seed water content may also be used. Ney et al. (1993) have shown that seed water content re- mained stable between 0.80 and 0.86 gg-1 during the period of seed abortion, and decreased from 0.80 to 0.55 gg- 1 during the period of linear seed growth. The water content of 0.55 gg-1 determines seed
Table 1 Variation of the values determining the developmental stages and the durations of the successive phases among species
Species Seed Seed water Duration Seed water Duration Reference length content FLO-FSSA content at FSSA-PM FSSA FLO-FSSA (degree-days) PM (g. g - l ) (degree-days) (mm) (g . g - i )
Pea 6-8.5 80-86 300 55 About 300
cv Solara 181 333-471 cv Finale 195 386-427 cv Frisson 234
55
Soybean 10-12 85 250-390 60 500-620
White lupin 11.5-13 85-86 600-650 65 300-400
60-65
Chickpea 7.6-8.7 85 60
Duthion and Pigeaire, 1991 Ney et al., 1993 Ney and Turc, 1993
Le Deunff and Rachidian, 1988
Duthion and Pigeaire, 1991 Munier-Jolain et al., 1993
Duthion and Pigeaire, 1991 Munier-Jolain, 1996 Noffsinger et al., 1994
Turc et al., 1994
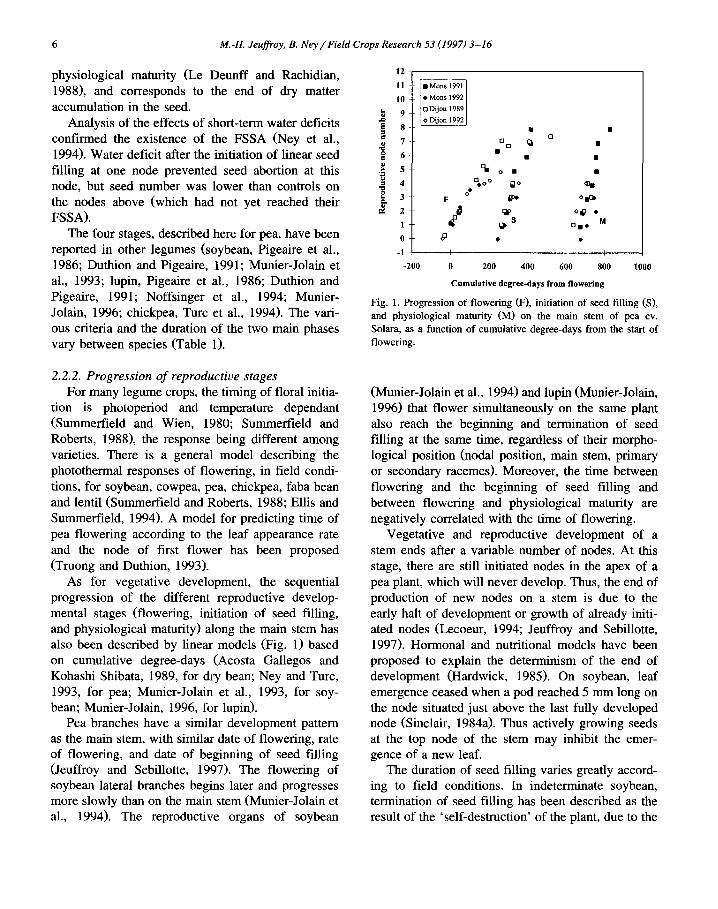
6 M.-H. Jeuffroy, B. Ney/Field Crops Research 53 (1997) 3-16
physiological maturity (Le Deunff and Rachidian, 1988), and corresponds to the end of dry matter accumulation in the seed.
Analysis of the effects of short-term water deficits confirmed the existence of the FSSA (Ney et al., 1994). Water deficit after the initiation of linear seed filling at one node prevented seed abortion at this node, but seed number was lower than controls on the nodes above (which had not yet reached their FSSA).
The four stages, described here for pea, have been reported in other legumes (soybean, Pigeaire et al., 1986; Duthion and Pigeaire, 1991; Munier-Jolain et al., 1993; lupin, Pigeaire et al., 1986; Duthion and Pigeaire, 1991; Noffsinger et al., 1994; Munier- Jolain, 1996; chickpea, Turc et al., 1994). The vari- ous criteria and the duration of the two main phases vary between species (Table 1).
12
=.
*d
10
9
8
7
6
5
4
3 F 2 8 1 £ 0 ~
-1 I
-200 0
I I • O
° o ~ •
01 0 • •
~o o Oo a~ o*
t~p o@ . s M
I I I :
200 400 600 800
Cumulative degree-days from flowering
1000
Fig. 1. Progression of f lowering (F), initiation of seed fil l ing (S), and physiological maturity (M) on the main stem of pea cv. Solara, as a function of cumulat ive degree-days from the start of flowering.
2.2.2. Progression of reproductive stages For many legume crops, the timing of floral initia-
tion is photoperiod and temperature dependant (Summerfield and Wien, 1980; Summerfield and Roberts, 1988), the response being different among varieties. There is a general model describing the photothermal responses of flowering, in field condi- tions, for soybean, cowpea, pea, chickpea, faba bean and lentil (Summerfield and Roberts, 1988; Ellis and Summerfield, 1994). A model for predicting time of pea flowering according to the leaf appearance rate and the node of first flower has been proposed (Truong and Duthion, 1993).
As for vegetative development, the sequential progression of the different reproductive develop- mental stages (flowering, initiation of seed filling, and physiological maturity) along the main stem has also been described by linear models (Fig. 1) based on cumulative degree-days (Acosta Gallegos and Kohashi Shibata, 1989, for dry bean; Ney and Turc, 1993, for pea; Munier-Jolain et al., 1993, for soy- bean; Munier-Jolain, 1996, for lupin).
Pea branches have a similar development pattern as the main stem, with similar date of flowering, rate of flowering, and date of beginning of seed filling (Jeuffroy and Sebillotte, 1997). The flowering of soybean lateral branches begins later and progresses more slowly than on the main stem (Munier-Jolain et al., 1994). The reproductive organs of soybean
(Munier-Jolain et al., 1994) and lupin (Munier-Jolain, 1996) that flower simultaneously on the same plant also reach the beginning and termination of seed filling at the same time, regardless of their morpho- logical position (nodal position, main stem, primary or secondary racemes). Moreover, the time between flowering and the beginning of seed filling and between flowering and physiological maturity are negatively correlated with the time of flowering.
Vegetative and reproductive development of a stem ends after a variable number of nodes. At this stage, there are still initiated nodes in the apex of a pea plant, which will never develop. Thus, the end of production of new nodes on a stem is due to the early halt of development or growth of already initi- ated nodes (Lecoeur, 1994; Jeuffroy and Sebillotte, 1997). Hormonal and nutritional models have been proposed to explain the determinism of the end of development (Hardwick, 1985). On soybean, leaf emergence ceased when a pod reached 5 mm long on the node situated just above the last fully developed node (Sinclair, 1984a). Thus actively growing seeds at the top node of the stem may inhibit the emer- gence of a new leaf.
The duration of seed filling varies greatly accord- ing to field conditions. In indeterminate soybean, termination of seed filling has been described as the result of the 'self-destruction' of the plant, due to the

M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16 7
remobilization of large amounts of nitrogen (Sinclair and de Wit, 1976). In different environmental condi- tions, seed filling has been shown to end when nitrogen available in the plant for remobilization was exhausted (Munier-Jolain et al., 1996b), except in the situations where seeds had reached their potential size, which depends on seed cell number.
2.2.3. Effects of water stress and nitrogen deficiency In glasshouse and field experiments, short-term
water stresses did not change the rate of progression of flowering, or the beginning of seed filling and physiological maturity in pea (Ney et al., 1994). They did not change the length of either the period between flowering and beginning of seed filling or the period between the beginning of seed filling and physiological maturity. The main effect was an ear- lier cessation of flowering, and thus a smaller num- ber of flowering nodes, which potentially bear pods
and seeds. These results are consistent with the effect of water stress on other indeterminate species, such as dry bean (Acosta Gallegos and Kohashi Shibata, 1989). The beginning and end of seed filling are earlier under long-term water stress compared with control conditions (Turc, 1995), from 30 to 50 de- gree-days and from 50 to 100 degree-days respec- tively for the first node, resulting in a shorter period of seed filling. The duration of soybean seed filling is affected by water deficit (Sionit and Kramer, 1977; Korte et al., 1983).
Nitrogen deficiency induces an early end of pea flowering (Sagan et al., 1993; Jeuffroy and Sebil- lotte, 1997). The rate of progression of flowering along the stem is not modified. Nitrogen deficiency for various periods in the cycle have been studied. Cessation of flowering is only affected if the defi- ciency starts before the onset of flowering on the stem and if it has a long duration (Jeuffroy and
1 6 -
uj¢ 14- rn
12- z
10-
uJ 8 -
O o= 4 -
¢ 2 -
0-
1 6 -
~ 14-
12- z
10- o z w 8 -
~ 6- o ~ 4 - n
U/
O-
Colmo • 1311991 ,0 D 2 1991 • D I 1992 zx D2 1992
A
0~. 0 • 0/~ O /"
200 400 600 800 1000
FinaJe * 1311991 0 D 2 199"1 • D1 1992 z~ D2 1992
/x # &'" o
~'~F °~S o o • •z~ PM
0 200 400 600 800 1000 C U M U L A ' R V E D E G R E E DAYS F R O M B E G I N N I N G O F F L O W E R I N G
14
12
10
Bingo ~ D1 1aS1 D2 1991
• D1 1992 z~ D2 1992
8q
o ~ o • 6d ~• •q~ OO•&
0-4, 0 200 400 600 800 1000
1214161 Countess 0
10 Z~,• •
0 • 8-1 • /x
6-t • • '~ '~ o#,A
q4 # * s 0 ~ ,
0 200 400
[311991 0 D2 199"1 • . D1 1992 ,~ D 2 1992
e~ 0 L~
O •Z /x,& O •
o • PM
600 800 1000 C U M U L A T I V E D E G R E E DAYS F R O M B E G I N N I N G O F F L O W E R I N G
Fig. 2. Genotypic variability of progression of flowering (F), beginning of seed filling (S), and physiological maturity (PM) on the main stem of pea, as a function of cumulative degree-days from the start of flowering, for four genotypes in different environments.

8 M.-H. Jeuffroy, B. Ney/Field Crops Research 53 (1997) 3-16
Sebillotte, 1997). Truong and Duthion (1993), exam- ining the deviations from their model for predicting the time of flowering, observed that deficient nitro- gen nutrition (characterized by a low nitrogen con- centration of aerial parts at flowering) promoted flowering. Although grain legumes can fix N 2 in symbiosis with rhizobia, nitrogen deficiencies have often been observed in pea fields (Dorr, 1992). In these situations, there are always few branches per plant.
2.2.4. Genetic variability In ten genotypes of pea, varying in their seed size,
foliage type and number of branches (Dumoulin et al., 1994), the progression of flowering, beginning of seed filling, and physiological maturity were linearly correlated to cumulative degree-days since the begin- ning of flowering. Significant genotypic differences were observed in the rates of progression of flower- ing and of beginning of seed filling (Fig. 2). The time between flowering and beginning of seed filling at the first node and the duration of seed filling were very similar in all genotypes, despite differences in seed size.
2.2.5. A model for reproductive development Ney and Turc (1993) proposed a general descrip-
tion of reproductive development on the main stem of a pea plant. Reproductive development can be entirely defined by the start, the end and the rate of progression of each stage along the stem: flowering, beginning of seed filling, and physiological maturity (Fig. 3). This general pattern of development has been successfully applied to soybean (Munier-Jolain et al., 1993) and lupin (Munier-Jolain, 1996). As some of the parameters in this model are constant or easily calculated, the characterization of the whole reproductive development requires only a few obser- vations. Thus, this model can be used to determine precisely the developmental stages of each reproduc- tive organ on a plant, all along the cycle. It also allows the period of seed formation to be deter- mined, during which time abortions can occur: it begins with the flowering of the first reproductive node and ends when the last reproductive node reaches the beginning of the seed filling stage. The period of seed filling, for an individual plant, lasts between the beginning of seed filling of the first
,+
E Z
Q3
0 Z
N F T
NST . . . . L
/ , - - 1 / R F II 1
i I , I
, s~, FT, ~s V O [ 11 I
I VG YP III
s. I
i
I
I
IV I
SF I
Cumula t ive Degree Days From Sta r t of Flowering
Fig. 3. A general description of pea reproductive development. S F, S s and S], are the start of flowering, seed filling, and physiological maturity; R F, R s and Rp are the rates of progres- sion of the three stages along the stem; E F, E s and Ep are the ends of the stages. NFT and NST are the highest flowering and fruiting nodes.
reproductive node and physiological maturity of the last. Thus, the periods of seed formation and seed filling overlap between the beginning of seed filling of the first and of the last reproductive nodes. This description allows analysis of the effects of limiting factors: for example a drought period can have an effect on seed number or on mean weight per seed, depending on which period is affected (Ney et al., 1994).
3. Growth
3.1. Radiation interception
As for other species, dry matter accumulation in legume crops can be analysed according to radiation interception and photosynthesis of the canopy. There has been much effort to model radiation interception by canopies of grain legumes. Generally, the fraction of incident radiation which is intercepted by the canopy, I / I o, is calculated as a function of the leaf area index (LAI) as:
I / I o = 1 - e x p ( - k × LAI) (1)
where k is the extinction coefficient of the canopy. In some studies, LAI is directly correlated with time

M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16 9
expressed in cumulative degree days (for instance for alfalfa (Medicago sativa): Lemaire and Allirand, 1993). Some other authors calculate LAI from the product of leaf area per plant and plant density. Leaf area per plant was related by Sinclair (1984b), Sin- clair (1986) to the plastochron index, hypothezing that both variables are dependent on temperature if there is no drought. This model was used by Sinclair et al. (1987) to simulate LAI of cowpea (Vigna unguiculata) and black gram (Vigna mungo) and more recently by Hammer et al. (1995) for peanut ( Arachis hypogea L.).
Very different extinction coefficients have been reported for a given species as reviewed by Varlet-
Grancher et al. (1989). These variations are due to the differences in methods of measurements and to the environmental conditions. Solar radiation balance is calculated as the difference between the radiation received (incident and soil-reflected radiation) and lost (crop-reflected and transmitted radiation) by the canopy. Methods differ by the type of radiation measured (total radiation or photosynthetically active radiation, PAR, measured as energy flux or photon flux) and also by the calculation of the balance. Some authors calculate absorbed radiation and others intercepted radiation (in this case, reflected radiation by soil and canopy are neglected). Variation in re- ported extinction coefficients also results from ef-
Table 2 Extinction coefficient k reported by various authors, and methods of calculation of radiation balance (absorbed and intercepted) and type of radiation measured
Species Absorbed Intercepted
Tot. R. EPAR QPAR TotalRad. EPAR QPAR Other a
Reference
Glycine max 0.88 0.60
Phaseolus vulgaris 0.4
Cajanus cajan 0.38
Medicago sativa 0.88
Vigna unguiculata 0.93
Vigna mungo
Lupinus albus
Cicer arietinum
Arachis hypogea
Pisum sativum leafed
Pisum sativum semi-leafless
Pisum sativum leafless
0.83
0.33/0.49
0.49
0.33-0.49 cor. at/2
0.46
0.55/0.75 cot. zr/2
0.62
0.95
0.90 0.68
0.97
0.47-0.61
0.50
0.33
0.33 cor. ~r/2
Shibles and Weber (1965) Sinclair (1986)
Gardiner et al. (1979)
Natarajan and Willey (1985) Sivakumar and Virmani (1984) Hugues et al. (1981)
Gosse et al. (1982) Fuess and Tesar (1968)
Varlet-Grancher and Bonhomme (1974) Littleton et al. (1979)
Muchow and Charles-Edwards (1982)
Varlet-Grancher et al. (1989)
Hugues et al. (1987)
Hammer et al. (1995)
Heath and Hebblethwaite (1985b) Martin et al. (1994) Ney (unpublished)
Heath and Hebblethwaite (1985b)
Martin et al. (1994)
Ney (unpublished)
Heath and Hebblethwaite (1985b)
a Other methods (for example photometric units) or not specified.
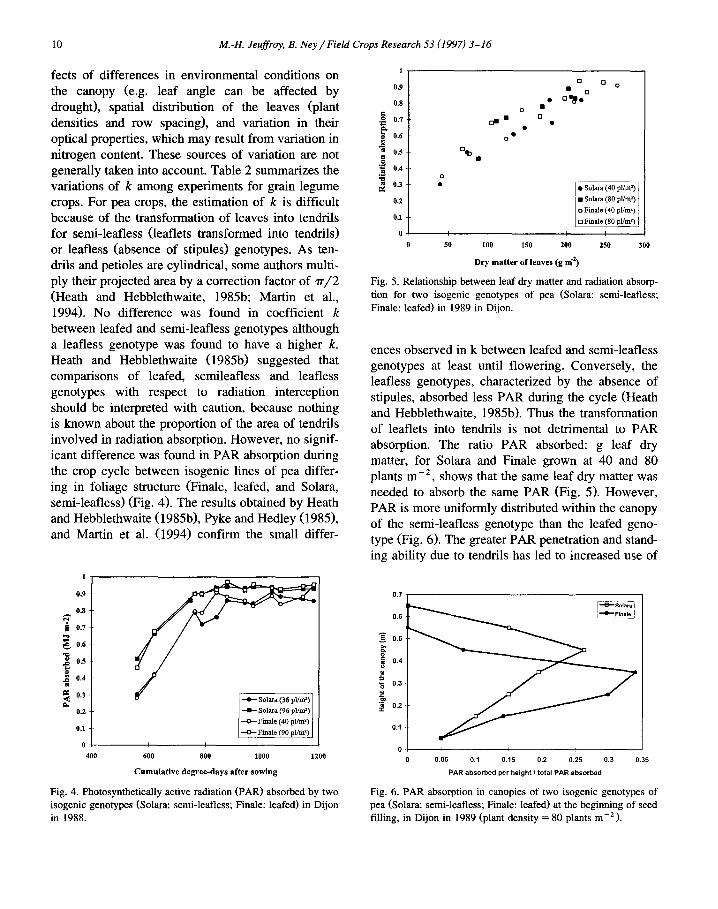
10 M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16
fects of differences in environmental conditions on the canopy (e.g. leaf angle can be affected by drought), spatial distribution of the leaves (plant densities and row spacing), and variation in their optical properties, which may result from variation in nitrogen content. These sources of variation are not generally taken into account. Table 2 summarizes the variations of k among experiments for grain legume crops. For pea crops, the estimation of k is difficult because of the transformation of leaves into tendrils for semi-leafless (leaflets transformed into tendrils) or leafless (absence of stipules) genotypes. As ten- drils and petioles are cylindrical, some authors multi- ply their projected area by a correction factor of 7r/2 (Heath and Hebblethwaite, 1985b; Martin et al., 1994). No difference was found in coefficient k between leafed and semi-leafless genotypes although a leafless genotype was found to have a higher k. Heath and Hebblethwaite (1985b) suggested that comparisons of leafed, semileafless and leafless genotypes with respect to radiation interception should be interpreted with caution, because nothing is known about the proportion of the area of tendrils involved in radiation absorption. However, no signif- icant difference was found in PAR absorption during the crop cycle between isogenic lines of pea differ- ing in foliage structure (Finale, leafed, and Solara, semi-leafless) (Fig. 4). The results obtained by Heath and Hebblethwaite (1985b), Pyke and Hedley (1985), and Martin et al. (1994) confirm the small differ-
0.9
0.8 ~7 ~a 0.7
5 0 . 6
~ 0~ ~ 0.4
~ 0J 0.2
0 3
• -4m-- Selma (96 pl/m 2) • -"0-- Finale (40 pl/m 2) • ...42-- Finale (90 pl]m 2)
¢- + ,¢
400 600 800 1000 1200
C u m u l a t i v e d e g r e e - d a y s a f t e r s o w i n g
Fig. 4. Photosynthetically active radiation (PAR) absorbed by two isogenic genotypes (Solara: semi-leafless; Finale: leafed) in Dijon in 1988.
0.9
0.8
0.7
~ 0.6
~ 0.5
~ 0.4
~ 0J
0.2
0.I
o~
om •
o
o
• o • DI~@
O • Q
U
Q o
• Solara (40 pUm2)~
• Soinra (80 pl/m2) 1 o Finale (40 pl/mz) [ 0 Finale (80 pl/m=) I
I b I I
I00 150 200 250 300
Dry m a t t e r of leaves (g m "2)
Fig. 5. Relationship between leaf dry matter and radiation absorp- tion for two isogenic genotypes of pea (Solara: semi-leafless; Finale: leafed) in 1989 in Dijon.
ences observed in k between leafed and semi-leafless genotypes at least until flowering. Conversely, the leafless genotypes, characterized by the absence of stipules, absorbed less PAR during the cycle (Heath and Hebblethwaite, 1985b). Thus the transformation of leaflets into tendrils is not detrimental to PAR absorption. The ratio PAR absorbed: g leaf dry matter, for Solara and Finale grown at 40 and 80 plants m -2, shows that the same leaf dry matter was needed to absorb the same PAR (Fig. 5). However, PAR is more uniformly distributed within the canopy of the semi-leafless genotype than the leafed geno- type (Fig. 6). The greater PAR penetration and stand- ing ability due to tendrils has led to increased use of
0.7 i r
0.6
0 . 5
0.4
0.3.
~ 0.2
0.1
0 0 0.05 0.1 0.15 0.2 0.25 0.3 0.35
PAR absorbed per height / total PAR absorbed
Fig. 6. PAR absorption in canopies of two isogenic genotypes of pea (Solara: semi-leafless; Finale: leafed) at the beginning of seed filling, in Dijon in 1989 (plant density = 80 plants m-2).

M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997)3-16 11
this trait in new varieties of pea during the last ten years.
3.2. Radiation use efficiency
Numerous studies have addressed photosynthesis in legume crops. Radiation use efficiency (RUE), i.e. the ratio aerial dry matter produced to PAR ab- sorbed, was found to be widely variable for a given species. In pea for instance, Heath and Hebbleth- waite (1985b) found 0.96 to 1.46 g aerial dry matter per MJ of total radiation intercepted depending on the experimental location; Pyke and Hedley (1985) 1.25 g DM MJ-1 total radiation intercepted; Martin et al. (1994) 1.43 g DM MJ -1 PAR intercepted under drought conditions, and Wilson et al. (1985) 2.36 g DM MJ- l PAR intercepted. One reason for this wide variability is the large effects of environ- mental conditions. Reductions in RUE under drought have been recorded in many studies for grain legumes including faba bean (Green et al., 1985), pigeon pea (Cajanus cajan L.) (Hughes and Keatinge, 1983) or pea (Keatinge et al., 1985). RUE is sensitive to minimum temperature (Bell et al., 1992, for peanut (Arachis hypogea L.)) and nitrogen nutrition (Sinclair and Horie, 1989).
RUE also varies along the crop cycle. As RUE depends on the relationship between sources and sinks (Shibles et al., 1987), it increases with pod growth and then declines when seeds are filling, as shown for pea (Fig. 7).
Finally, numerous studies (Boerma and Ashley, 1988; Ashley and Boerma, 1989; see Shibles et al., 1987 for a review) have reported genetic variability in canopy apparent photosynthesis for soybean. In our experiments for three years in Dijon (France) with pea, RUE ranged from 1.55 to 2.05 g aerial dry matter M J-1 PAR absorbed for 6 genotypes of pea before flowering. On the other hand, no significant difference was found between genotypes differing only in their leaf structure (Heath and Hebblethwaite, 1985b; Pyke and Hedley, 1985).
3.3. Growth and yield determination
A positive relationship has been found between canopy apparent photosynthesis (CAP) during the reproductive period and yield in soybean (review by
4
3.5
3
2.5.
~1 2
0.5
0
200
D RUE • seed DM
+ pod DM . . . . . . Flowering
P 0
600
500
400 -""
300
200
100
400 600 800 1000 1200 1400
Cumulative degree-days from emergence
Fig. 7. Var ia t ions o f r ad ia t ion use e f f ic iency ( R U E ) wi th t ime in
relation to growth of pods and seeds for pea (Solara, Dijon, 1989).
Ashley and Boerma, 1989). For some of these stud- ies, such as those involving source-sink manipula- tions, the results must be interpreted with caution: if CAP during reproductive development is influenced by sinks, it is obvious that CAP will correlate with yield.
Other authors have related yield or yield compo- nents directly to the production of assimilates by the canopy. Jiang and Egli (1995) found a linear rela- tionship for soybean between seed number, the most predictive component of yield, and crop growth rate during flowering and pod set.
4. Assimilate partitioning and structure determi- nation
4.1. Assimilate partitioning
There is little knowledge of processes involved in carbon partitioning to the various organs of a plant (Whisler et al., 1986). Assimilate partitioning is thus the main weak point of current crop simulation models. Usually, carbon is translocated from a ma- ture leaf to the nearest actively growing region, but this general pattern can be changed if leaves are removed or shaded or depending on sink require- ments (Shibles et al., 1987).
In pea, the presence of a sink and a source on the same reproductive node led some authors to study

12 M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16
the possible preferential relationship between them. At a node scale, the assimilates allocated to the pods come mainly from the leaf of the node, and to a lesser extent from the hull of the node (Flinn and Pate, 1970). However, translocation of assimilates between nodes can occur (Harvey, 1973; Szynkier, 1974; Jeuffroy and Warembourg, 1991). In pea and soybean, during the reproductive phase, carbon is preferentially exported to pods on the same node and, secondarily, to pods two nodes below or above (Shibles et al., 1987), according to the vascularisa- tion of the plant (see review of Pate, 1975).
In a 14C-labelling experiment, with plants grow- ing under two different radiation intensities, Jeuffroy and Warembourg (1991) showed that there was no priority among the sinks in the plant during the period of seed formation. When the amount of assim- ilates produced by the whole plant was severely reduced, the amount of ~4C allocated to each individ- ual organ was proportionally reduced. This led to a model for assimilate partitioning between the organs on a stem, at different dates during this period. In the model, the assimilates produced by the stem during a time interval are allocated to each pod in proportion to its demand. This demand is linked to the initial amount of dry matter of the pod before its final stage in seed abortion, and to the seed number of the pod
1 1 \ ~ -II---- D3S 1 observed
10 & ~ ' ~ / --£3-- D3SI simulated 9 ~ 4 --o-- S i 1 observed
--o-- SI 1 simulated 8
~ 6
~ 5
z e 4 '
3
2
0 I I L I h I 1
0 1 2 3 4 5 6 7
S E E D N U M B E R P E R N O D E P E R S T E M
Fig. 8. C o m p a r i s o n o f o b s e r v e d seed n u m b e r s pe r node o b s e r v e d
and va lues s imula ted wi th the m o d e l o f J eu f f roy (1994b) , f r o m
two f ie ld loca t ions ( D 3 S 1 = Char t res , s o w n 1 4 / 0 3 / 8 9 , 257 s t ems
m - 2 ; S l l = Gr ignon , s o w n 2 4 / 0 2 / 9 2 , 97 s t ems m - 2 ) .
after this stage (Jeuffroy and Devienne, 1995). Parti- tioning between reproductive and vegetative organs is dependent on the plant growth during the two preceding phyllochrons (Jeuffroy, 1994a). This model was tested in various field conditions (Jeuffroy and Devienne, 1995), with good accordance between ob- served and simulated growth of the different organs, i.e. pods of each node and vegetative parts. How- ever, in situations with water stress, the growth rate of seeds which had begun to fill before the beginning of the stress remained stable, while abortions oc- curred on pods which had not yet reached this stage (Ney et al., 1994). This suggests that filling pods have priority over young pods in assimilate partition- ing. In soybean, defoliation reducing the supply of assimilates did not also modify seed growth after the beginning of the linear growth phase of the seed (Munier-Jolain, 1994). This observation could be used in a general model of assimilate partitioning.
4.2. Relationship between assimilate partitioning and structure formation
At the pod scale, seed number can be predicted from pod growth dynamics during the first 120 degree-days after anthesis (Jeuffroy and Chabanet, 1994). Thus, the pattern of assimilate partitioning (see above) and the relationship between early pod growth rate and seed number per pod have been used to construct a model for seed number production on each node of a pea stem (Jeuffroy, 1992; Jeuffroy, 1994b). The inputs of the model are the development parameters (as presented above), the growth rate of the whole stem during the period of seed formation, and the climatic data (mean daily temperature). When tested in various field conditions (Jeuffroy, 1994b), this model gave a good account of the variability in seed number partitioning among the nodes of the stems (Fig. 8). This model requires measurements of the assimilate demand of the filling seeds. In soy- bean (Munier-Jolain, 1994) and pea (Munier-Jolain and Ney, 1995), the potential seed growth rate is determined by seed cell number, which is fixed at the beginning of seed filling. Thus, if the cell num- ber is low at this stage, the accumulation of reserves in the seed until physiological maturity can be lim- ited even if assimilate sources are large. The rela-

M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16 13
t ionship be tween seed g rowth rate and cell number
does not s eem to di f fer be tween genotypes , even
with substantial d i f ferences in individual seed weight .
5. Conclusion
Recen t w o r k has substant ial ly improved under-
s tanding of the func t ion ing o f l e g u m e crops. This
a l lows bet ter def ini t ion o f the var ious improvemen t s
that are possible in product ion, y ie ld stability, and
seed qual i ty o f these crops. The more precise charac-
ter isat ion o f the deve lopmen ta l stages a long the cy-
cle, and the de te rmina t ion o f sensi t ive per iods may
make poss ible the avo idance o f stresses. For example
in pea, water stresses (Ney et al., 1994) or high
tempera tures (Jeuff roy et al., 1990) affect y ie ld re-
duc ing seed number when they occur before the final
s tage in seed abortion. The later the stress, the
smal ler the decrease in yield. Conce rn ing growth,
there is genet ic var iabi l i ty for radiat ion in tercept ion
and radiat ion use ef f ic iency. Lit t le increase is to be
expec ted in potent ia l R U E , but genotypes wi th low
R U E must be e l iminated. The part i t ioning o f growth
into harves ted yie ld also cou ld be improved . Harves t
index (HI) remains wide ly var iable on some species.
De te rmina te types have, in s o m e condit ions, more
stable HI, but these types are not avai lable for all
grain l e g u m e species. For some species, it is neces-
sary to unders tand the reproduc t ive structure forma-
t ion in relat ion to env i ronmenta l condit ions. Recen t
studies have shown, as descr ibed in this chapter, that
plant structure can be s imula ted f rom plant sequen-
tial d e v e l o p m e n t and growth.
References
Acosta Gallegos, J. and Kohashi Shibata, J., 1989. Effect of water stress on growth and yield of indeterminate dry-bean (Phaseolus vulgaris) cultivars. Field Crops Res., 20: 81-93.
Ashley, D.A. and Boerma, H.R., 1989. Soybean canopy photo- synthesis and seed yield. In: Proc. IV World Soybean Confer- ence 1989, Buenos Aires, Argentina, pp. 169-176.
Bell, M.J., Wright, G.C. and Hammer, G.L., 1992. Night tempera- ture affects radiation use efficiency in peanut. Crop Sci., 32: 1329-1335.
Boerma, H.R. and Ashley, D.A., 1988. Canopy photosynthesis and seed fill duration in recently developed soybean cultivars and selected plant introduction. Crop Sci., 28: 137-140.
Bunting, A.H., Gibbons, R.W. and Wynne, J.C., 1985. Groundnut (Arachis hypogaea L.). In: eds. R.J. Summerfield and E.H. Roberts, Grain Legume Crops. Collins Technical Books, Lon- don, pp. 747-800.
CarrouEe, B., 1996. Personal communication. Cooper, D.C., 1938. Embryology of Pisum sativum. Bot. Gaz.
(Chicago), 100: 123-132. Dawkins, T.C.K., Hebblethwaite, P.D. and McGowan, M., 1984.
Soil compaction and the growth of vining peas. Ann. Appl. Biol., 105: 329-343.
Dort, T., 1992. Analyse, par voie d'enqu~te, de la variabilitE des rendements et des effets prEc&lents du pois protEagineux de printemps (Pisum sativum L.). ThOse de Doctorat, INA-PG, Paris, 214 pp.
Dracup, M. and Kirby, E.J.M., 1993. Patterns of growth and development of leaves and intemodes of narrow-leafed hipin. Field Crops Res., 34: 209-225.
Dumoulin, V., Ney, B. and EttvE, G., 1994. Variability of seed and plant development in pea. Crop Sci., 34: 992-998.
Dure, L.S., 1975. Seed formation. Ann. Rev. Plant Physiol., 26: 259-278.
Duthion, C. and Pigeaire, A., 1991. Seed lengths corresponding to the final stage in seed abortion of three grain legumes. Crop Sci., 31: 1579-1583.
Duthion, C., Ney, B. and Munier-Jolain, N.M., 1994. Develop- ment and growth of white lupin: implication for crop manage- ment. Agron. J., 86: 1039-1045.
Ellis, R.H. and Summerfield, R.J., 1994. A phenological model and its potential use in legume crop breeding. In: eds. J.D.H. Keatinge and I. Kiismenoglu, Autumn Sowing of Lentil in the Highlands of West Asia and North Africa. Central Research Institute for Field Crops, Ankara, pp. 107-120.
EtEvE, G. and Derieux, M., 1982. Variabilit6 de la durEe de la phase vEgttative chez le pois (Pisum sativum L.). Application h la selection de types rtsistants h l'hiver et h la determination de la date de semis. Agronomie, 2: 813-817.
Farrington, P. and Greenwood, E.A.N., 1975. Description and specification of the branching structure of lupins. Aust. J. Agric. Res., 26: 507-510.
Flinn, A.M. and Pate, J.S., 1970. A quantitative study of carbon transfer from pod and subtending leaf to the ripening seeds of the field pea (Pisum arvense L.). J. Exp. Bot., 21: 71-82.
Fuess, F.W. and Tesar, M., 1968. Photosynthetic efficiency, yields and leaf loss in alfalfa. Crop Sci., 8: 159-163.
Gardiner, T.R., Vietor, D.M. and Craker, L.E., 1979. Growth habit and row width effects on leaf area development and light interception of field beans. Can. J. Plant Sci., 59: 191-199.
Gosse, G., Chattier, M., Varlet-Grancher, C., Bonhomme, R., 1982. Interception du rayonnement utile h la photosynth~se chez la luzeme: variations et modElisation. Agronomie, 6: 583-588.
Green, C.F., Hebblethwaite, P.D. and Ison, D.A., 1985. A quanti- tative analysis of varietal and moisture status effects on the growth of Vicia faba in relation to radiation absorption. Ann. appl. Biol., 106: 143-155.
Gupton, C.L., Emery, D.A. and Benson, J.A., 1968. Reproductive efficiency of virginia type peanuts: III. Relating the time of

14 M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16
peg placement to the branching pattern of the plant. Oleagineux, 23: 247-250.
Hammer, G.L., Sinclair, T.R., Boote, K.J., Wright, G.C., Meinke, H. and Bell, M.J. 1995. A peanut simulation model: I: Model development and testing. Agron. J., 87: 1085-1093.
Hardwick, R.C., 1985. Yield components and processes of yield production in vining peas. In: eds. P.D. Hebblethwaite, M.C. Heath and T.C.K. Dawkins, The Pea Crop. Butterworths, London, pp. 317-326.
Harvey, D.M., 1973. The translocation of 14C photosynthate in Pisum sativum L. Ann. Bot., 37: 787-794.
Heath, M.C. and Hebblethwaite, P.D., 1985a. Agronomic prob- lems associated with the pea crop. In: eds. P.D. Hebbleth- walte, M.C. Heath and T.C.K. Dawkins, The Pea Crop. But- terworths, London, pp. 19-29.
Heath, M.C. and Hebblethwaite, P.D., 1985b. Solar radiation interception by leafless, semileafless and leafed peas (Pisum sativum) under contrasting field conditions. Ann. appl. Biol., 107: 309-318.
Hedley, C.L. and Ambrose, M.J., 1981. Designing 'leafless' plants for improving yields of the dried pea crop. Adv. Agron., 34: 225-277.
Hughes, G. and Keatinge, J.D.H., 1983. Solar radiation intercep- tion, dry matter production and yield in pigeon pea (Cajanus cajan (L.) Millspaugh). Field Crops Res., 6: 171-178.
Hugues, G., Keating, J.D.H. and Scott, S.P., 1981. Pigeon pea as a dry season crop in Trinidad, West Indies. H: Interception and utilization of solar radiation. Trop. Agric., 191-199.
Hugues, G., Keating, J.D.H., Cooper, P.J.M. and Dee, N.F., 1987. Solar radiation interception and utilization by chick pea (Cicer arietinum L.) crops in northern Syria. J. Agric. Sci., Camb., 108: 419-424.
Jeuffroy, M.-H. and Chabanet, C., 1994. A model to predict seed number per pod from early pod growth rate in pea (Pisum sativum L.). J. Exp. Bot., 45: 709-715.
Jeuffroy, M.-H. and Devienne, F., 1995. A simulation model for assimilate partitioning between pods in pea (Pisum sativum L.) during the period of seed set; validation in field conditions. Field Crops Res., 41: 79-89.
Jeuffroy, M.-H. and Sebillotte, M., 1997. The end of flowering in pea: influence of plant nitrogen nutrition. Eur. J. Agron., in press.
Jeuffroy, M.-H. and Warembourg, F.R., 1991. Carbon transfer and partitioning between vegetative and reproductive organs in Pisum sativum L. Plant Physiol., 97: 440-448.
Jeuffroy, M.-H., 1991. RSle de la vitesse de croissance, de la rrpartition des assimilats et de la nutrition azotre dans l'61aboration du nombre de graines du pois protragineux de printemps (Pisum sativum L.). Thbse de Doctorat, Universit6 Paris XI, Orsay, 208 pp.
Jeuffroy, M.-H., 1992. P'tipois: a simulation model for pea seed number. In: ed. A. Scaife, Proceedings of 2nd Congress of European Society for Agronomy, Warwick, pp. 90-91.
Jeuffroy, M.-H., 1994a. Les rbgles de partage des assimilats. In: eds. INRA, ITCF and UNIP, Agrophysiologie du Pois Protragineux, Applications h la Production Agricole, pp. 49- 73.
Jeuffroy, M.-H., 1994b. Le nombre de graines par tige. In: eds. INRA, ITCF and UNIP, Agrophysiologie du Pois Protragineux, Applications h la Production Agricole, pp. 93-110.
Jeuffroy, M.-H., Duthion, C., Meynard, J.-M. and Pigeaire, A., 1990. Effect of a short period of high day temperatures during flowering on the seed number per pod of pea (Pisum sativum L.). Agronomie, 2: 139-145.
Jiang, H.-F. and Egli, D.B., 1995. Soybean seed number and crop growth rate during flowering. Agron. J., 87: 264-267.
Johansen, C., Baldev, B., Brouwer, J.B., Erskine, W., Jermyn, W.A., Li-Juan, L., Malik, B.A., Ahad Miah, A. and Silim, S.N., 1992. Biotic and abiotic stresses constraining productiv- ity of cool season food legumes in Asia, Africa and Oceania. In: eds. F.J. Muehlbauer and W.J. Kaiser, Expanding the Production and Use of Cool Season Food Legumes. Kluwer, Dordrecht, pp. 175-194.
Keatinge, J.D.H., Cooper, P.J.M. and Hughes, G., 1985. The potential of peas as a forage in the dryland cropping rotations of Western Asia. In: eds. P.D. Hebblethwalte, M.C. Heath and T.C.K. Dawkins, The Pea Crop. Butterworths, London, pp. 185-192.
Kirby, E.J.M., 1995. Factors affecting rate of leaf emergence in barley and wheat. Crop Sci., 35: 11-19.
Korte, L.L., Specht, J.E., Williams, J.H. and Sorensen, R.C., 1983. Irrigation of soybean genotypes during reproductive ontogeny. II: Yield component responses. Crop Sci., 23: 528- 533.
Le Deunff, Y. and Rachidian, Z., 1988. Interruption of water delivery at physiological maturity is essential for seed devel- opment, germination and seedling growth in pea (Pisum sativum L.). J. Exp. Bot., 39: 1221-1230.
Lecoeur, J., 1994. Rrponses phrnologiques et morphologiques du pois (Pisum sativum L.) h la contrainte hydrique. Consr- quences sur le nombre de phytom~res reproducteurs. ThOse de Doctorat, ENSAM, Montpellier, 99 pp.
Lecoeur, J., Wery, J., Turc, O. and Tardieu, F., 1995. Expansion of pea leaves subjected to short water deficit: cell number and cell size are sensitive to stress at different periods of leaf development. J. Exp. Bot., 46: 1093-1101.
Lemaire, G. and Allirand, J.M., 1993. Relation entre croissance et qualit6 de la luzerne: interaction grnotype-mode d'exploita- tion. Fourrages, 134: 183-198.
Leong, S.K. and Ong, C.K., 1983. The influence of temperature and soil water deficit on the development and morphology of groundnut (Arachis hypogaea L.). J. Exp. Bot, 34: 1551-1561.
Linck, A.J., 1961. The morphological development of the fruit of Pisum sativum, var. Alaska. Phytomorpholoy, 11: 79-84.
Littleton, E.J., Dennet, M.D., Monteith, J.L. and Elston, E.L., 1979. The growth and development of cowpeas (Vigna un- guiculata) under tropical field conditions. 2: Accumulation and partition of dry weight. J. Agric. Sci., Camb., 93: 309-320.
Lyndon, R.F., 1977. The shoot apical meristem. In: eds. I.F. Sutcliffe and J.S. Pate, The Physiology of the Garden Pea. Academic Press, New York, pp. 183-212.
Martin, I., Tenorio, J.L. and Ayerbe, L., 1994. Yield, growth and water use of conventional and semi-leafless peas in semiarid environments. Crop Sci., 34: 1576-1583.

M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16 15
Maurer, A.R., Jaffray, D.E. and Fletcher, H.F., 1966. Response of peas to environment. III: Assessment of the morphological development of peas. Can. J. Plant Sci., 46: 285-290.
Monti, L., Biddle, A.J., Moreno, M.T. and Plancquaert, P., 1992. Biotic and abiotic stresses of pulse crops in Europe. In: eds. F.J. Muehlbaner and W.J. Kaiser, Expanding the Production and Use of Cool Season Food Legumes. Kluwer, Dordrecht, pp. 204-218.
Muchow, R.C. and Charles-Edwards, D.A., 1982. A physiological analysis of growth of mungbeans at a range of plant densities in tropical Australia. I: Dry matter production. Aust. J. Agric. Res., 33: 41-51.
Munier-Jolain, N.G. and Ney, B., 1995. Variability of seed growth rate for various pea cultivars (Pisum sativum L.) in relation to their seed cell number. Proceedings of 2nd European Confer- ence on Grain Legumes, Copenhagen, pp. 32-33.
Munier-Jolain, N.G., 1994. Etude de la variabilit6 du poids indi- viduel des graines du soja de type ind&ermin6 (Glycine max. L. Merrill, cv. Maple Arrow). Influence de l'appadtion srquentielle des organes reproducteurs. Th~se de Doctorat, INA-PG, Paris, 85 pp.
Munier-Jolain, N.G., Ney, B. and Duthion, C., 1993. Sequential development of flowers and seeds on the mainstem of an indeterminate soybean. Crop Sci., 33: 768-771.
Munier-Jolain, N.G., Ney, B. and Duthion, C., 1994. Reproduc- tive development of an indeterminate soybean as affected by morphological position. Crop Sci., 34: 1009-1013.
Munier-Jolain, N.G., Ney, B. and Duthion, C., 1996b. Termina- tion of seed growth in relation to nitrogen content of vegeta- tive parts in soybean plants. Eur. J. Agron., 5: 219-225.
Munier-Jolain, N.M., 1996. Analyse et modrlisation de la struc- ture du peuplement et de la production de graines chez le lupin blanc (Lupinus albus L.) de printemps. Effets de la date et de la densit6 de semis. ThOse de Doctorat, INA-PG, Pads, 144
Pp. Mnnier-Jolaln, N.M., Ney, B. and Duthion, C., 1996a. Analysis of
branching in spring-sown white lupins (Lupinus albus L.): the significance of the number of axillary buds. Ann. Bot., 77: 123-131.
Natarajan, M. and Willey, R.W., 1985. Effects of row arrange- ment on light interception and yield in sorghum-pigeon pea intercropping. J. Agric. Sci., Camb., 104: 263-270.
Ney, B. and Turc, O., 1993. Heat-unit-based description of the reproductive development of pea. Crop Sci., 33: 510-514.
Ney, B., Duthion C., and Turc, O., 1994. Phenological response of pea to water stress during reproductive development. Crop Sci., 34: 141-146.
Ney, B., Duthion, C. and Fontaine, E., 1993. Timing of reproduc- tive abortions in relation to cell division, water content, and growth of pea seeds. Crop Sci., 33: 267-270.
Noffsinger, S.L., Kraepiel, A., Huyghe, C. and van Santen, E., 1994. Mean seed mass in relation to moisture content, and pod length and width in antumn-sown white lupin (Lupinus albus L). 1st Australian Lupin Technical Symposium.
Oram, P.A. and Agcaoili, M., 1992. Title? In: eds. F.J. Muehlbauer and W.J. Kaiser, Expanding the Production and Use of Cool Season Food Legumes. Kluwer, Dordrecht, pp. 3-52.
Pate, J.S. and Flinn, A.M., 1977. Fruit and seed development. In: eds. I.F. Sutcliffe and J.S. Pate, The Physiology of the Garden Pea. Academic Press, New York, pp. 431-468.
Pate, J.S., 1975. Pea. In: ed. L.T. Evans, Crop Physiology: Some Case Histories. Camb. Univ. Press, Cambridge, pp. 191-224.
Pigeaire, A., Duthion, C. and Turc, O., 1986. Characterization of the final stage in seed abortion in indeterminate soybean, white lupin and pea. Agronomie, 6: 371-378.
Pyke, K.A. and Hedley, C.L., 1985. Growth and photosynthesis of different pea phenotypes. In: eds. P.D. Hebblethwaite, M.C. Heath and T.C.K. Dawkins, The Pea Crop. Butterworths, London, pp. 297-305.
Sagan, M., Ney, B. and Duc, G., 1993. Plant symbiotic mutants as a tool to analyse nitrogen nutrition and yield relationship in field-grown peas (Pisum sativum L.). Plant and Soil, 153: 33-45.
Shibles, R., Secor, J. and Ford, D.M., 1987. Carbon assimilation and metabolism. In: Soybeans: Improvement, Production, and Uses. ASA-CSSA-SSSA, pp. 535-588.
Shibles, R.M. and Weber, C.R., 1965. Leaf area, solar radiation interception and dry matter production by soybeans. Crop Sci., 5: 575-577.
Sinclair, T.R. and de Wit, C.T., 1976. Analysis of the carbon and nitrogen limitations to soybean yield. Agron. J., 68: 319-324.
Sinclair, T.R. and Horie, T., 1989. Leaf nitrogen, photosynthesis, and crop radiation use efficiency: a review. Crop Sci., 29: 90-98.
Sinclair, T.R., 1984a. Cessation of leaf emergence in indetermi- nate soybeans. Crop Sci., 24: 483-486.
Sinclair, T.R., 1984b. Leaf area development in field-grown soy- beans. Agron. J., 76: 141-146,
Sinclair, T.R., 1986. Water and nitrogen limitations in soybean grain production. I. Model development. Field Crops Res., 15: 125-141.
Sinclair, T.R., Muchow, R.C., Ludlow, M.M., Leach, G.J., Lawn, R.J. and Foale, M.A., 1987. Field and model analysis of the effect of water deficits on carbon and nitrogen accumulation by soybean, cowpea and black gram. Field Crops Res., 17: 121-140.
Sionit, B.K. and Kramer, P.J., 1977. Effect of water stress during different stages of growth of soybean. Agron. J., 69: 274-278.
Sivakumar, M.V.K. and Virmani, S.M., 1984. Crop productivity in relation to intercception of photosynthetically active radia- tion. Agric. For. Meteorol., 31: 131-141.
Slinkard, A.E., Bascur, G. and Heruandez-Bravo, G., 1992. In: eds. F.J. Muehlbauer and W.J. Kaiser, Expanding the Produc- tion and Use of Cool Season Food Legumes. Kluwer, Dor- drecht, pp. 195-203.
Summerfield, R.J. and Roberts, E.H., 1988. Photothermal regula- tion of flowering in pea, lentil, faba bean and chickpea. In: ed. R.J. Summerfield, World Crops: Cool Season Food Legumes. Kluwer, Dordrecht, pp. 911-922.
Summerfield, R.J. and Wien, H.C., 1980. Effects of photoperiod and air temperature on growth and yield of economic legumes. In: eds. R.J. Summerfield and A.H. Bunting, Advances in Legume Science. Royal Botanic Gardens, Kew, pp. 17-36.
Szynkier, K., 1974. The effect of removing of supply or sink

16 M.-H. Jeuffroy, B. Ney / Field Crops Research 53 (1997) 3-16
organs on the distribution of assimilates in two varieties of garden pea, Pisum sativum L. Acta Agric. Scand., 4: 7-12.
Truong, H.H. and Duthion, C., 1993. Time of flowering of pea (Pisum sativum L.) as a function of leaf appearance rate and node of first flower. Ann. Bot., 72: 133-142.
Turc, O., 1995. Drought stress accelerates seed growth and devel- opment in pea. In: Proceedings of 2nd European Conference on Grain Legumes, Copenhagen, pp. 106-107.
Turc, O., Farinha, N.C., Sousa, C.J.F. and Ney, B., 1994. A characterization of seed growth and water content to describe the development of chickpeas seeds. In: eds. M. Borin and M. Sattin, Proceedings of 3rd European Society of Agronomy Congress, Abano-Padova, pp. 258-259.
Varlet-Grancher, C. and Bonhomme, R., 1974. Utilisation de l'6nergie solaire par une culture de Vigna sinensis. III: Evolu- tion du rendement 6nerg6tique pendant la phase v6g6tative. Ann. Agron., 25: 811-819.
Varlet-Grancher, C., Gosse, G., Chattier, M., Sinoquet, H., Bon- homme, R. and Allirand, J.M., 1989. Mise au point: rayon- nement solaire absorb6 ou intercept6 par un couvert v6g~tal. Agronomie, 9: 419-439.
Whisler, F.D., Acock, B., Baker, D.N., Fye, R.E., Hodges, H.F., Lambert, J.R., Lemmon, H.E., McKinion, J.M. and Reddy, V.R., 1986. Crop simulation models in agronomic systems. Adv. Agron., 40: 141-208.
Wilhelm, W.W. and McMaster, G.S., 1995. Importance of the phyllochron in studying development and growth in grasses. Crop Sci., 35: 1-3.
Wilson, D.R, Jamieson, P.D., Jermyn, W.A. and Hanson, R., 1985. Models of growth and water use of field peas (Pisum sativum L.). In: eds. P.D. Hebblethwaite, M.C. Heath and T.C.K. Dawkins, The Pea Crop. Butterworths, London, pp. 139-151.