fetal rat sympathetic neurons maintained in a serum-free medium retain induced cholinergic...
TRANSCRIPT

DevelopmentalBrain Research, 14 (1984) 71-82 71 Elsevier
BRD 50045
Fetal Rat Sympathetic Neurons Maintained in a Serum-free Medium
Retain Induced Cholinergic Characteristics
DENNIS HIGGINS 1, LORRAINE IACOVITTI 2 and HAROLD BURTON 1
I Department of Anatomy and Neurobiology, Washington University School of Medicine, St. Louis, MO 63110 and 2Laboratory of Neurobiology, Department of Neurology, Cornell University Medical College, New York, NY IO021 (U.S.A.)
(Accepted January 17th, 1984)
Key words: adrenergic neurons - - cholinergic sympathetic neurons - - electrical coupling - - tissue culture - - dual function neurons --neurotransmitters - - embryonic sympathetic neurons
When maintained in some serum-containing media, fetal rat sympathetic neurons acquire substantial choline acetyltransferase ac- tivity and form cholinergic synapses in vitro. However, when they are maintained in a serum-free, defined culture medium, choline
acetyltransferase activity is not detected and cholinergic synapses are not observed. In this study, we have examined the effects of vari- ous times of exposure to a medium inducing cholinergic function on the properties of neurons subsequently maintained in defined me- dium. We report that 2-day, but not 2-h, exposure to this inducing medium causes a long-lasting (> 6 weeks) increase (7-10-fold) in the activity of choline acetyltransferase and that, under these conditions, sympathetic neurons in vitro form cholinergic, electrical and mixed function cholinergic and electrical synapses. We conclude that a relatively brief exposure to media inducing cholinergic function can cause long-lasting changes in the functional properties of sympathetic neurons in vitro.
INTRODUCTION
In tissue culture, post-mitotic sympathetic neurons
of perinatal rats can form several different types of non-adrenergic synapses2,7,10,22, 25. Because the type
or types of synapses formed by these neurons is sub-
ject to experimental manipulation and can be altered by changes in tissue culture media2,22,23, in popula-
tions of non-neuronal cells 23 or in the nature of the
substratum8, sympathetic neurons in vitro have been
used as a model system to study the mechanisms that determine which types of synapses will be formed by
developing autonomic neurons2,22.
We have previously reported that, when main-
tained in a serum-free, chemically-defined medium, sympathetic neurons of embryonic rats form electro- tonic synapses 10. In this medium, no cholinergic syn-
apses have been observed in recordings from more than 200 pairs of cells and the activity of choline ace-
tyltransferase, the enzyme which catalyzes the synthe- sis of acetylcholine, is below the limits of our assay 13.
Correspondence: D. Higgins, Department of Pharmacology, School U.S.A,
In contrast, when sympathetic neurons are main-
tained in a medium supplemented with human pla-
cental serum and chick embryo extract, the neurons acquire substantial amounts of choline acetyltrans-
ferase activityt2. ~7 and form cholinergic chemical syn-
apsesll,16, is. In this study sympathetic neurons have
been exposed to both defined medium and to supple-
mented medium for various lengths of time in order to
determine: (1) how long an exposure to the supple-
mented medium is necessary to allow at least a sub-
population of the neurons to acquire cholinergic
function; (2) whether, once acquired, cholinergic properties will be retained in defined medium; (3)
whether some neurons will respond to the 2 different
media by forming mixed function synapses. Electro- physiological, neurochemical, and immunocytochem-
ical techniques have been used to study the types of
synapses formed by such sympathetic neurons. Our results indicate that a relatively short exposure (48 h)
to a soluble inducing agent or agents can cause a long-lasting (> 1 month) change in the properties
of Medicine, State University of New York, Buffalo, NY 14214,
0165-3806/84/$03.00 © 1984 Elsevier Science Publishers B.V.

72
of sympathetic neurons in vitro and that these neu- rons have the capacity to form cholinergic, electrical or mixed cholinergic and electrical synapses while re- taining at least some of the properties characteristic of an adrenergic neuron.
METHODS
Tissue culture In these experiments, fetal, sympathetic neurons
in dissociated cell cultures were exposed for various lengths of time to a supplemented medium that was previously been shown to induce cholinergic function in these cells 2,11A2,16-18. Subsequently the neurons
were maintained for several weeks in the absence of non-neuronal cells in a serum-free, defined medium that by itself does not cause the expression of cholin- ergic characteristicsl0,x3. 2s, Supplemented medium contains: 65% (v/v) Eagle's minimum essential medi- um, 25% human placental serum, 10% chick embryo (9 days) extract, glutamine (200/~g/ml), glucose (6 mg/ml) and 2.5S nerve growth factor (100 ng/ml). The defined medium used in this study is a modifica- tion of the medium designated N 2 by Bottenstein and Satol; it contains 50% (v/v) Dulbecco's modified Ea- gle's medium and 50% Ham's F-12 medium with added glutamine (200 #g/ml), insulin (5 ktg/ml), hu- man transferrin (100/zg/ml), progesterone (6 ng/ml), putrescine (16 ]~g/ml), selenium (5 ng/ml) and pu- rified 2.5S nerve growth factor (100 ng/ml).
Superior cervical sympathetic ganglia were remov- ed from rat fetuses (20-21 days) and previously de- scribed procedures were used to obtain a suspension of dissociated ganglion cells15,18. Neurons receiving no exposure to supplemented medium were plated in defined medium with 0.25% soybean trypsin inhibi- tor or with 0.5% bovine serum albumin (to promote initial attachment). Two days later this medium was removed and subsequently the cells were maintained on defined medium. All other neurons were plated in supplemented medium and were allowed to remain in this medium for various lengths of time (2 h, 2 days, constant exposure). After this initial exposure to supplemented medium, the cultures were rinsed 4 times with medium F-12 and once with defined medi- um; thereafter they were maintained only on defined medium. These 2 plating methods yield similar num- bers of cells 10. In all the various types of cultures,
non-neuronal cells were eliminated by adding an an- timitotic agent (fluorodeoxyurdine, 10 ktM with uri- dine, 10/~M) to the medium during the 3rd-5th and 14th-16th days in vitro.
Sympathetic neurons were grown in both monolay- er and microisland cultures 6. Because enzyme assays required several thousand cells, monolayer cultures grown on an air-dried collagen substratum15 were used in these experiments. In our initial electrophys- iological experiments with such monolayers, we ob- served heterogeneous synaptic interactions in some cultures. To facilitate the characterization of such synapses, we found it useful to reduce both the num- ber of neurons and the area of their synaptic contacts by using microisland cultures 6. Microislands (9/dish) were prepared by putting small drops ( - 1 mm diam- eter) of collagen from a 25-gauge needle on an Aclar culture dish 15. They were exposed to NH3 vapors, washed with distilled water and air-dried. Then a sec- ond droplet of collagen was placed on top of the first and this was air-dried.
Enzyme assays The activities of the enzymes dopamine-fl-hydrox-
ylase and choline acetyltransferase were determined in the same tissue homogenate ~2. Choline acetyl- transferase activity was determined by a published modification 12 of the method of Schrier and Shus- ter26. To ensure that we were measuring choline ace- tyltransferase, and not the related enzyme, carnitine acetyltransferase, samples were assayed in duplicate in the presence and absence of 300 ktM naphthylvinyl- pyridine, a specific inhibitor of choline acetyltrans- ferase. Dopamine-fl-hydroxylase activityl2 was measured by the method of Henry et ai. 9 in the pres- ence of 8 ~M copper sulfate.
Histological methods A published modification of the peroxidase-anti-
peroxidase method was used for the immunocytoche- mical localization of tyrosine hydroxylase12; the spec- ificity of the antiserum has been described 14. For auto-
radiography, cultures were incubated in Hepes buf- fered balanced salt solution (below) containing L-[7- 3H]norepinephrine (0.2/~M, 14,8 Ci/mM, New En- gland Nuclear) and 10 ktg/ml ascorbate at 35 °C for 30 min. They were then rinsed extensively (5 x) , in- cubated in non-radioactive norepinephrine (10 ~M)

for 2 periods of 10 min at 4 °C, and fixed in 2% glutar- aldehyde in 0.1 M sodium phosphate buffer
(pH 7.4, 4 °C, 16 h). The culture was then coated with Kodak NTB2 emulsion and exposed for 3 days at 4 °C.
Electrophysiological methods Standard electrophysiological techniques, which
have been described 10,11, were used to obtain simulta- neous intracellular recordings from pairs of neurons on islands; a bridge circuit was used to pass stimulat- ing current through the recording electrode. During recording sessions, the cultures were perfused with a balanced salt solution (32-34 °C, pH 7.3) that con- tained: NaCI (125 mM), HEPES (4-(2-hydroxyme- thyl)- 1-piperazinethanesulfonic acid, 30 mM), NaOH (17 mM), KC1 (5 mM), MgC12 (0.2 mM), CaCI 2 (2 raM), and glucose (11 raM).
RESULTS
Enzyme assays Sympathetic neurons that had been both plated
and maintained in defined medium in the absence of non-neuronal cells had no detectable choline acetyl- transferase activity (<0.05 pmol acetylcholine syn- thesized/neuron/h) at either 3 or 6 weeks in vitro (Table I). In contrast, neurons that had been main- tained in supplemented medium had substantial cho- line acetyltransferase activity at both 3 and 6 weeks in vitro and the amount of this activity was at least 44- fold (3 weeks) to 180-fold (6 weeks) greater than in defined medium (Table I). To determine how long of an exposure to supplemented medium was necessary to allow at least some of the cells to express choliner- gic characteristics, some cultures were exposed to
73
supplemented medium for 2 h or 2 days and subse- quently maintained in defined medium; previous work in this laboratory (below) and in other laborato- ries 24 had established that the required exposure was <10 days. In cultures exposed to supplemented medium for 2 h, no choline acetyltransferase activity was observed at any time in vitro (Table I). However, after a 48-h exposure, choline acetyltransferase ac- tivity was detectable after both 3 and 6 weeks and the amount of this activity was at least 7-fold (3 weeks) to 10-fold (6 weeks) greater than in defined medium. Thus, a relatively brief (48 h) exposure allows at least a subpopulation of neurons to express detect- able cholinergic characteristics and is capable of
causing a very long lasting (>40 days) change in the properties of neurons maintained in defined medium. The activity of choline acetyltransferase in cultures
exposed to supplemented medium for 48 h was much less (6-16%) than the activity in cultures constantly maintained in supplemented medium. However, the relatively smaller amount of choline acetyltransfer- ase in the former type of culture appeared to have functional significance since it was associated with a change in the nature of the observed synaptic interac- tions (below).
In both defined medium and supplemented medi- um, sympathetic neurons retain many of the charac- teristics of adrenergic neurons12,13. To determine whether a 48-h exposure to supplemented medium had a general effect on all the enzymes involved in neurotransmitter biosynthesis, or a relatively selec- tive effect on acetylcholine biosynthesis, we meas- ured the activity of dopamine-fl-hydroxylase, the en- zyme which catalyzes the conversion of dopamine to
norepinephrine, in the various types of cultures. More dopamine-fl-hydroxylase activity was observed
TABLE I
The activities of choline acetyltransferase and dopamine-fl-hydroxylase in cultures exposed for different lengths of time to supplemented medium
Exposure to supplemented medium
Choline acetyltransferase activity (pmol acetylcholine/neuron/h)
Dopamine-fl-hydroxylase activity (pmol octopamine/neuron/ h )
3 weeks in vitro 6 weeks in vitro 3 weeks in vitro 6 weeks in vitro
constant 2.20 ± 0.41 (12) 9.0 ± 2.2 (12) 1.58 ± 0.14 (8) 2.90 ± 0,55 (8) 48 h 0.36 ± 0.02 (5) 0.50 ± 0.07 (4) 0.38 ± 0.03 (5) 0.69 ± 0,09 (4)
2 h <0.05 (8) <0.05 (7) 0.43 ± 0.03 (6) 0.67 ± 0.05 (7) 0 h <0.05 (3) <0.05 (3) - - _

74
2 9
3
~ ~ i ~ i~i ~ i ~ i~ii~i!
flail 4 . . . . . iili~i ̧ ?~ ~i!i~ ~?~i,!ii~i ¸¸~
Fig. 1. A photomicrograph of a microisland culture. The sympathetic neurons on this collagen island were exposed to supplemented medium for 2 days and then were maintained in defined medium. On the 17th day in vitro, intracellular recordings were obtained from the numbered cells; these records are shown in Figs. 2 through 6. Subsequently the culture was processed for the immunocytochemical localization of tyrosine hydroxylase; all neurons on this island were judged to be tyrosine hydroxylase-positive. The calibration bar represents 200 ~m.
8 i i
k__..d w i ^
I I
b T
III I
!
- - - ~ _ I A
13 ; I ! 0 i 0 I
iJ i I
_ - i,,____
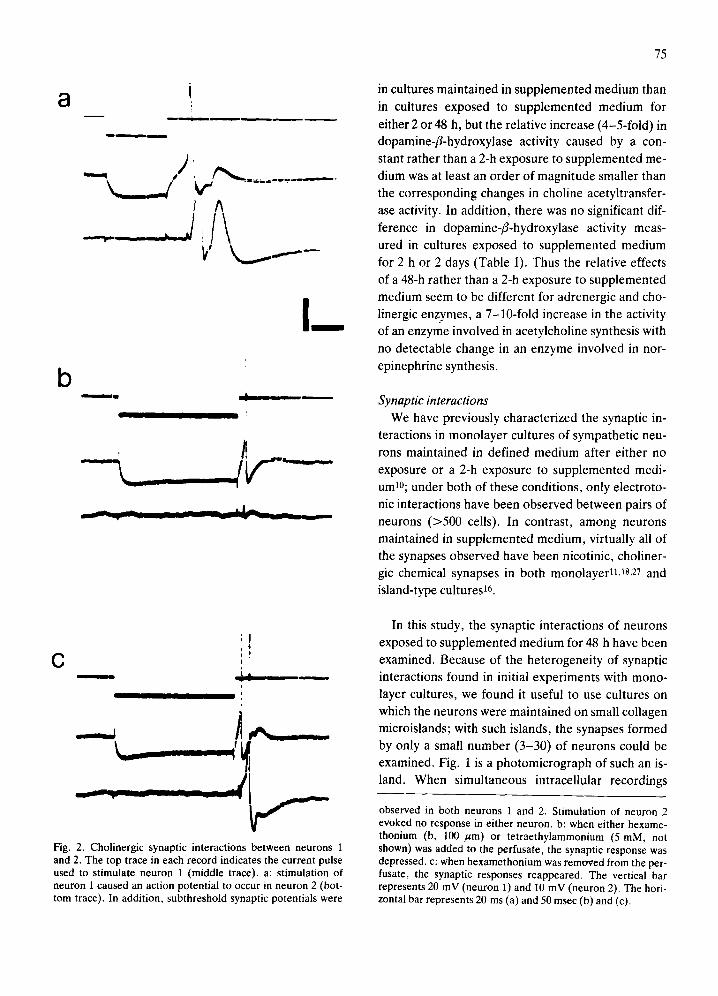
Fig. 2. Cholinergic synaptic interactions between neurons 1 and 2. The top trace in each record indicates the current pulse used to stimulate neuron 1 (middle trace), a: stimulation of neuron 1 caused an action potential to occur in neuron 2 (bot- tom trace). In addition, subthreshold synaptic potentials were
75
in cultures mainta ined in supplemented medium than
in cultures exposed to supplemented medium for
either 2 or 48 h, but the relat ive increase (4-5-fold) in
dopamine-f l -hydroxylase activity caused by a con-
stant ra ther than a 2-h exposure to supplemented me-
dium was at least an order of magni tude smaller than
the corresponding changes in choline acetyltransfer-
ase activity. In addit ion, there was no significant dif-
ference in dopamine-f l -hydroxylase activity meas-
ured in cultures exposed to supplemented medium
for 2 h or 2 days (Table I). Thus the relative effects
of a 48-h rather than a 2-h exposure to supplemented
medium seem to be different for adrenergic and cho-
linergic enzymes, a 7-10-fold increase in the activity
of an enzyme involved in acetylcholine synthesis with
no detectable change in an enzyme involved in nor-
epinephrine synthesis.
Synaptic interactions We have previously character ized the synaptic in-
teractions in monolayer cultures of sympathet ic neu-
rons mainta ined in defined medium after ei ther no
exposure or a 2-h exposure to supplemented medi-
um10; under both of these condit ions, only electroto-
nic interactions have been observed between pairs of
neurons (>500 cells). In contrast , among neurons
maintained in supplemented medium, virtually all of
the synapses observed have been nicotinic, choliner-
gic chemical synapses in both monolayerl~,ls,27 and
island-type cultures 16.
In this study, the synaptic interactions of neurons
exposed to supplemented medium for 48 h have been
examined. Because of the heterogenei ty of synaptic
interactions found in initial exper iments with mono-
layer cultures, we found it useful to use cultures on
which the neurons were mainta ined on small collagen
microislands; with such islands, the synapses formed
by only a small number (3-30) of neurons could be
examined. Fig. 1 is a photomicrograph of such an is-
land. When simultaneous intracel lular recordings
observed in both neurons 1 and 2. Stimulation of neuron 2 evoked no response in either neuron, b: when either hexame- thonium (b, 100 /~m) or tetraethylammonium (5 mM, not shown) was added to the perfusate, the synaptic response was depressed, c: when hexamethonium was removed from the per- fusate, the synaptic responses reappeared. The vertical bar represents 20 mV (neuron 1) and 10 mV (neuron 2). The hori- zontal bar represents 20 ms (a) and 50 msec (b) and (c).
76
I 8 , , ¸
Im
b
t
!
Jl )t f "
_ - - J
G I
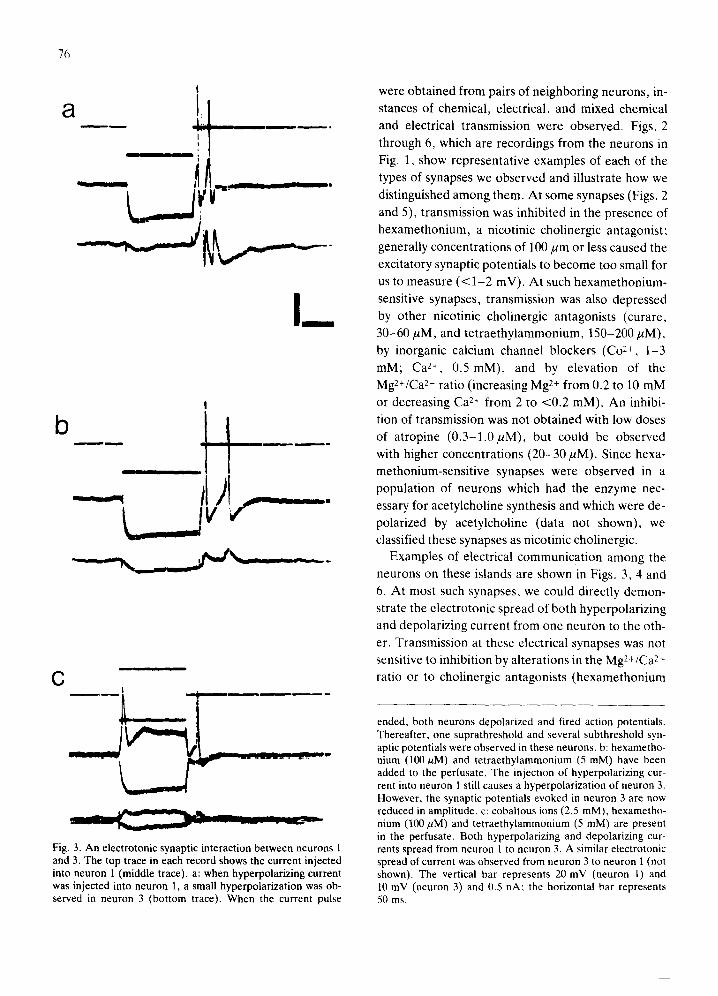
Fig. 3. An electrotonic synaptic interaction between neurons 1 and 3. The top trace in each record shows the current injected into neuron 1 (middle trace), a: when hyperpolarizing current was injected into neuron 1, a small hyperpolarization was ob- served in neuron 3 (bottom trace). When the current pulse
were obta ined from pairs of neighboring neurons, in-
stances of chemical , electrical, and mixed chemical
and electrical t ransmission were observed. Figs, 2
through 6, which are recordings from the neurons in
Fig. 1, show representat ive examples of each of the
types of synapses we observed and il lustrate how we
distinguished among them. At some synapses (Figs. 2
and 5), transmission was inhibited in the presence of
hexamethonium, a nicotinic cholinergic antagonist ;
generally concentrat ions of 100 ~tm or less caused the
excitatory synaptic potent ials to become too small for
us to measure ( < 1 - 2 mV). At such hexamethonium-
sensitive synapses, t ransmission was also depressed
by other nicotinic cholinergic antagonists (curare,
30-60 /aM, and te t rae thy lammonium, 150-200/aM),
by inorganic calcium channel blockers (Co 2÷, 1 -3
mM; Ca 2÷, 0.5 mM), and by elevat ion of the
Mg2÷/Ca 2+ ratio (increasing Mg 2+ from 0.2 to 10 mM
or decreasing Ca 2÷ from 2 to <0 .2 mM). An inhibi-
tion of transmission was not obta ined with low doses
of a tropine (0 .3 -1 .0 /aM) , but could be observed
with higher concentrat ions (20-30/aM) , Since hexa-
methonium-sensi t ive synapses were observed in a
populat ion of neurons which had the enzyme nec-
essary for acetylcholine synthesis and which were de-
polarized by acetylcholine (data not shown), we
classified these synapses as nicotinic cholinergic.
Examples of electrical communicat ion among the
neurons on these islands are shown in Figs. 3, 4 and
6. At most such synapses, we could directly demon-
strate the electrotonic spread of both hyperpolar iz ing
and depolar izing current from one neuron to the oth-
er. Transmission at these electrical synapses was not
sensitive to inhibition by al terat ions in the Mg2+/Ca 2+
ratio or to cholinergic antagonists (hexamethonium
ended, both neurons depolarized and fired action potentials. Thereafter, one suprathreshold and several subthreshold syn- aptic potentials were observed in these neurons, b: hexametho- nium (100/~M) and tetraethylammonium (5 mM) have been added to the perfusate. The injection of hyperpolarizing cur- rent into neuron 1 still causes a hyperpolarization of neuron 3. However, the synaptic potentials evoked in neuron 3 are now reduced in amplitude, c: cobaltous ions (2.5 mM), hexametho- nium (100 ~M) and tetraethylammonium (5 mM) are present in the perfusate. Both hyperpolarizing and depolarizing cur- rents spread from neuron 1 to neuron 3. A similar electrotonic spread of current was observed from neuron 3 to neuron 1 (not shown). The vertical bar represents 20 mV (neuron 1) and 10 mV (neuron 3) and 0.5 nA; the horizontal bar represents 50 ms.

(200/~m); curare (60/~M)). In addition, the spread
of current was not reduced, and usually was facili-
tated (Fig. 4), in the presence of tetraethylammoni-
um (5 mM), which at miUimolar concentrat ions is a
K + channel blocker, and/or Co 2+ (3 mM); this in-
crease was probably due to a decreased membrane
conductance caused by these agents 20. At a few syn-
apses that we classified as electrical, we could not di-
rectly demonstrate the passage of current from soma
to soma. At such synapses, transmission was facili-
tated in the presence of millimolar concentrations of
te traethylammonium and, therefore, was not nic-
a
• ,qll n
I m
b
~ O
, . I
Fig. 4. An electrotonic interaction between neurons 4 and 5. The top trace indicates the current injected into neuron 4 (mid- dle trace), a: when hyperpolarizing current was injected into neuron 4, there was either no change or a very small change in the membrane potential of neuron 5 (bottom trace). However, a small synaptic potential occurred in neuron 5 when neuron 4 fired an action potential, b: in the presence of Co 2÷ (3 mM) and tetroethylammonium (5 mM), the electrotonic spread of cur- rent from neuron 4 to neuron 5 was observed. The vertical bar represents 20 mV (neuron 4), 10 mV (neuron 5) and 1.0 nA. The horizontal bar represents 50 ms.
77
I m
19
_ . I w ~ . ~ l ~ l g ~ l ~ 8 ~ . ~ n ~ . llg~.~,~ ~ . ~ ~ ~ ~ q
Fig. 5. A cholinergic synaptic interaction between neurons 6 and 7. The top trace in (a) indicates the current used to stimu- late neuron 7 (bottom trace), a: stimulation of neuron 7 caused subthreshold and suprathreshold synaptic potentials to occur in both neurons 6 and 7. Stimulation of neuron 6 evoked no re- sponse in either neuron, b: the synaptic interaction was de- pressed when hexamethonium (100/~M) was added to the per- fusate. The vertical bar represents 10 mV (neuron 6), 20 mV (neuron 7) and 1 nA. The horizontal bar represents 20 ms.
otinic cholinergic. It was also unaffected by elevation
of the Mg2+/Ca 2+ ratio. When Co 2÷ or Cd2+ were
added to the medium, the postsynaptic potential of-
ten became somewhat smaller as the presynaptic ac-
tion potential was reduced in amplitude, but failures
of transmission were not observed.
Some neurons in these cultures communicated
with other neurons by means of both chemical and
electrical synapses. An example of such a neuron is
shown in Figs. 2 and 3. Fig. 2 illustrates that stimula-
tion of neuron 1 caused a complex series of excitatory
synaptic potentials and action potentials to appear in
neuron 2 (and in itself); in the presence of either hex-
amethonium or te t rae thylammonium transmission
was reversibly depressed. Neuron 1 also interacted
with neuron 3 (Fig. 3); thus, when hyperpolarizing
current was injected into either neuron, it spread elec-
trotonically to the other neuron of the pair. In addi-
tion a component of the synaptic response of neuron
3 to an action potential in neuron 1 was sensitive to
hexamethonium or hexamethonium with tetraethyl-
78
El !
l / !
- - _ ! k
I / b
/ I
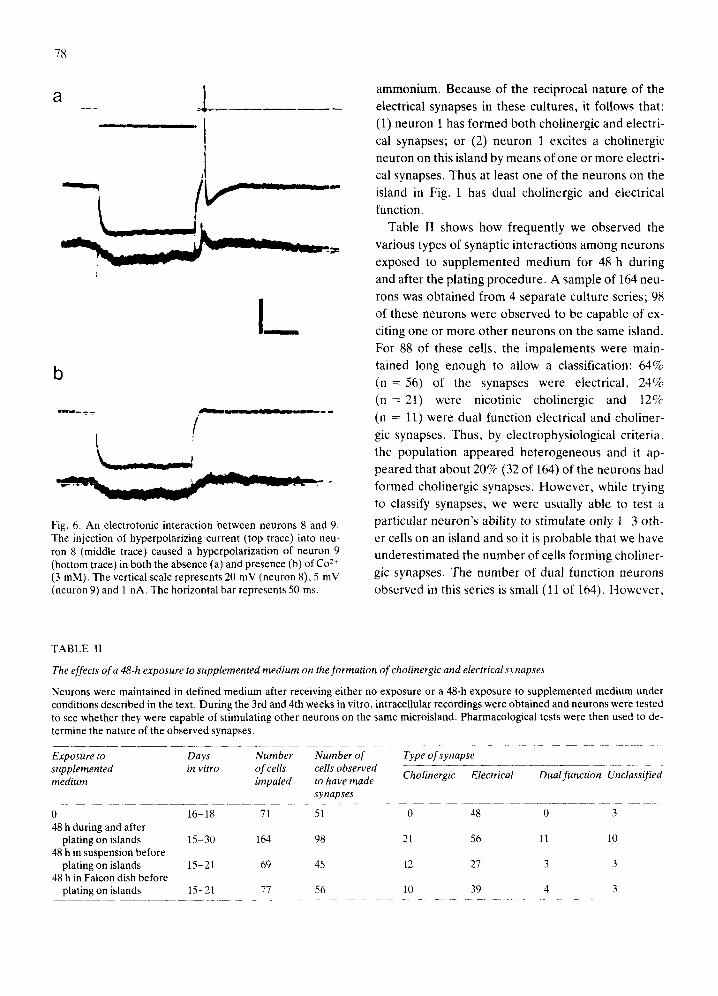
Fig. 6. An electrotonic interaction between neurons 8 and 9. The injection of hyperpolarizing current (top trace) into neu- ron 8 (middle trace) caused a hyperpolarization of neuron 9 (bottom trace) in both the absence (a) and presence (b) of Co 2+ (3 mM). The vertical scale represents 20 mV (neuron 8), 5 mV (neuron 9) and 1 nA. The horizontal bar represents 50 ms.
ammonium. Because of the reciprocal nature of the
electrical synapses in these cultures, it follows that: (1) neuron 1 has formed both cholinergic and electri-
cal synapses; or (2) neuron 1 excites a cholinergic
neuron on this island by means of one or more electri-
cal synapses. Thus at least one of the neurons on the
island in Fig. 1 has dual cholinergic and electrical
function.
Table II shows how frequently we observed the
various types of synaptic interactions among neurons
exposed to supplemented medium for 48 h during
and after the plating procedure. A sample of 164 neu-
rons was obtained from 4 separate culture series; 98 of these neurons were observed to be capable of ex-
citing one or more other neurons on the same island.
For 88 of these cells, the impalements were main-
tained long enough to allow a classification: 64%
(n-- -56) of the synapses were electrical, 24%
(n = 21) were nicotinic cholinergic and t2%
(n : 11) were dual function electrical and choliner-
gic synapses. Thus, by electrophysiological criteria,
the population appeared heterogeneous and it ap-
peared that about 20% (32 of 164) of the neurons had
formed cholinergic synapses. However, while trying to classify synapses, we were usually able to test a
particular neuron's ability to stimulate only 1-3 oth-
er cells on an island and so it is probable that we have
underestimated the number of ceils forming choliner-
gic synapses. The number of dual function neurons
observed in this series is small (11 of 164). However,
TABLE II
The effects of a 48-h exposure to supplemented medium on the formation of cholinergic and electrical synapses
Neurons were maintained in defined medium after receiving either no exposure or a 48-h exposure to supplemented medium under conditions described in the text. During the 3rd and 4th weeks in vitro, intracellular recordings were obtained and neurons were tested to see whether they were capable of stimulating other neurons on the same microisland. Pharmacological tests were then used to de- termine the nature of the observed synapses.
Exposure to Days Number Number of Type of synapse supplemented in vitro of cells cells observed medium impaled to have made Cholinergic Electrical Dual function Unclassified
synapses
0 16-18 71 51 0 48 0 3 48 h during and after
plating on islands 15-30 164 98 21 56 l I 10 48 h in suspension before
plating on islands 15-21 69 45 12 27 3 3 48 h in Falcon dish before
plating on islands 15-21 77 56 10 39 4 3

the presence of such a subpopulation has been a con- sistent finding. We have observed 10 other instances
of dual function in culture series where we did not classify all of the synapses and they have been ob- served in all 7 of the culture series used in this study.
In a previous study, we reported that sympathetic neurons which had received no exposure or a 2-h ex-
posure to supplemented medium formed electrotonic but not cholinergic synapses 10. However, in that
study, only monolayer cultures were used. To rule out the possibility that the detection of cholinergic synapses among neurons exposed to supplemented
medium was due to a change in the culture system (monolayer vs island), we obtained recordings from 71 neurons on islands which had been both plated and maintained in defined medium. Fifty-one cells sam-
pled were found to have formed synapses; all 48 that we were able to classify were electrotonic.
The data in Table II concerning the frequency of cholinergic interactions among neurons exposed to supplemented medium for 48 h were obtained be-
tween the 16th and 30th days in vitro. We were unable to obtain satisfactory impalements at longer times in vitro because the islands began to detach during the 4th or 5th week in vitro from the plastic culture dishes that we use. However, we were able to use the mono- layer culture system to confirm what the enzyme as- says of choline acetyltransferase indicated, i.e. that, once it has been induced, cholinergic function can be retained for at least 6 weeks in defined medium. In these experiments, sympathetic neurons were ex- posed to supplemented medium for 10 days and then maintained in defined medium for 35-42 days. When
intracellular recordings were obtained, 41 of 67 neu- rons were capable of exciting at least one other neu- ron on the dish; all the synapses tested (n = 23) were inhibited by hexamethonium or tetraethylammoni- a m .
Immunocytochemistry and autoradiography Four cultures containing neurons which had re-
ceived a 48-h exposure to supplemented medium were incubated with a specific antibody to tyrosine hydroxylase and were subsequently stained by the peroxidase-anti-peroxidase method. More than 97% of the neurons (n = 613) on the 36 islands in these cultures had specific dark brown deposits with- in their perikaryon (Fig. 1). Three of the stained cul-
79
tures contained islands that had a total of 4 dual func-
tion electrical/cholinergic neurons; all of the neurons on the islands containing mixed function neurons were tyrosine hydroxylase-positive.
Four cultures were incubated in tritiated norepi-
nephrine and subsequently processed for autoradio- graphy. Again more than 95% of the somata of the neurons were labelled. The number of grains over
the neurons was reduced to background levels in cul- tures incubated with tritiated norepinephrine in the presence of desipramine (10 pM); thus, the labelling represented specific catecholamine uptake. On two islands containing dual function neurons, all of the
neurons were labelled by tritiated norepinephrine. These data indicate that virtually all of the neurons exposed to supplemented medium for 48 h, includ-
ing those forming dual cholinergic/electrical syn- apses, retain at least 2 of the properties characteristic of an adrenergic neuron.
Mechanisms We have tried to determine whether the long-last-
ing change in cholinergic function caused by a 2-day exposure to supplemented medium is caused by: (1) molecules which bind to the substratum 8 and contin-
ue to exert their effects after supplemented medium has been removed; or (2) soluble molecules which act upon the neurons only during the 48-h exposure. In one series of experiments, we tested the effect of ex- posing the collagen microislands to supplemented medium for 48 h before adding cells. A 2-day expo- sure to either supplemented medium or a serum-free medium (L-15) caused a change in the properties of
the air-dried collagen such that no neurons adhered to the islands. We were thus unable to determine whether some of the cholinergic induction was due to
material bound to the substratum. However, we have been able to obtain data that indicate that at least some of the changes in cholinergic function do not re- quire a substrate-bound material. In the first experi- ment, dissociated neurons were exposed to supple- mented medium for 2 days in suspension. They were then pelleted by centrifugation (75-100 g for 7 min); resuspended in defined medium with 0.25% trypsin inhibitor, plated onto islands, and maintained in de- fined medium for 17 days. In a second experiment neurons were maintained in supplemented medium for 2 days in a 60 mm Falcon dish without a collagen

8(t
substratum. They were then detached from the sur- face with trypsin (0.05% in Ca2+,Mg2+-free Hanks
Balanced Salt Solution), pelleted, and then treated
like the neurons in the first experiment. In both ex-
periments a subpopulation of cholinergic neurons
was observed among neurons maintained in defined
medium (Table 1I). In the first experiment, 36% (15
of 42) of the classified synapses were either choliner-
gic or dual function; in the second experiment, 26%
(14 of 53) of the synapses had a cholinergic compo- nent.
DISCUSSION
Embryonic rat sympathetic neurons form choliner-
gic synapses when they are maintained in a tissue cul-
ture medium supplemented with human placental se-
rum and chick embryo extract 2,1 ~,12,16-18. In this study we have examined the effects of various times of ex-
posure to supplemented medium on the properties of
neurons maintained subsequently in a serum-free de-
fined medium. Our results indicate that: (1) a short
exposure to supplemented medium is sufficient to
cause a long-lasting change in the functional prop-
erties of neurons maintained in defined medium; and
(2) some sympathetic neurons in vitro are capable of expressing simultaneously parts of 3 different synap-
tic transmission systems. Patterson and Chun 24 reported that sympathetic
neurons which received an initial 10-day exposure to
heart conditioned medium produced more acetylcho-
line on the 30th day in vitro than neurons which had
never been exposed to conditioned medium. In our
initial experiments we were able to confirm this ob-
servation. Sixty percent of the sympathetic neurons
which had been exposed to supplemented medium for 10 days and then had been maintained in defined
medium for 5 -6 weeks were found to have made cho- linergic synapses; this percentage is comparable to
the percentage ( - 8 5 % ) of cholinergic neurons ob- served in cultures of sympathetic neurons maintained in the constant presence of supplemented medi- um 1~,16, In contrast, cholinergic synapses have not
been observed among neurons maintained in defined medium without an exposure to supplemented medi- um 10. Thus, once they have been exposed to a medi- um inducing cholinergic properties, sympathetic neu- rons can retain cholinergic function for long times in
both serum-free and serum-containing media. After
obtaining these data, we examined the effects of shorter exposures to supplemented medium to see
whether we could find an effective exposure duration
that was short enough to allow eventual characteriza-
tion of the mechanism of induction by the use of me-
tabolic inhibitors. A 2-day, but not a 2-h, exposure to
supplemented medium caused a measurable amount
of choline acetyltransferase activity to be present at
both 3 and 6 weeks in vitro; however, there was no
significant difference in dopamine-fi-hydroxylase ac-
tivity measured in cultures exposed to supplemented medium for 2 or 48 h. Others j3,2s have reported that
tyrosine hydroxylase activity is similar in sympathetic neurons maintained in serum-free and in serum-con-
taining media. Thus, the long-lasting effects of a 48-h
exposure to supplemented medium appeared to be at
least somewhat selective; they did not represent an
increase in the activity of all enzymes involved in
neurotransmitter biosynthesis. The magnitude of the
increase in choline acetyltransferase activity (7-10-
fold more than in defined medium) caused by a 48-h
exposure was modest when compared to the amount
induced by a constant exposure to supplemented me-
dium (44-180-fold), but it was readily detectable. Furthermore, it was associated with changes in syn-
aptic function. We found that 36% of the synaptic in-
teractions (20% of all cells tested) were either cholin-
ergic or had a cholinergic component . Freschi ~ has also observed some cholinergic synapses among sym-
pathetic neurons maintained in defined medium', in
his study, the neurons were initially maintained in a medium (18 h) containing fetal calf serum, a serum
known to induce cholinergic function 4. Thus, a 2-day
exposure to supplemented medium or to some other cholinergic inducing media is sufficient to cause at
least a subpopulation of sympathetic neurons to ex-
press cholinergic function in defined medium.
Since the hormones present in defined medium do not prevent the acquisition of cholinergic function 5
and since the addition of adult rat serum to an L-15- based defined medium promotes the synthesis of ace- tylcholine 28. it appears that the effects of a 48-h expo- sure to supplemented medium are mediated by a cho-
linergic-inducing factor(s) present in one or both of the biological extracts used in this medium. It has been reported that both soluble and substrate-bound factors are capable of inducing cholinergic function

in sympathetic neurons in vitro 8. Since the factor or factors present in supplemented medium have not
been isolated, it seemed possible that some of the long-lasting effects of supplemented medium on cho- line acetyltransferase activity in monolayer cultures
could be due to material bound to the collagen sub- stratum. However, we have found about the same percentage of cholinergic synapses in cultures in which the neurons were exposed to supplemented
medium before plating on islands as in cultures that were exposed to supplemented medium while the neurons were on the islands. Thus, at least a substan-
tial part of the effect of a 48-h exposure to supple- mented medium is mediated by a molecule(s) that can be removed from the medium after 48 h and that need not bind to the substratum. In unpublished ex- periments (Higgins) we have also observed choliner-
gic synapses among neurons exposed to heart condi- tioned medium for 48 h; with this culture medium cholinergic induction is known to be mediated by a soluble factor 23.
When maintained in defined medium with no ex- posure to supplemented medium, sympathetic neu- rons form electrotonic, but not cholinergic, syn- apses ~0. Among neurons exposed to supplemented medium for 48 h, we observed electrical, cholinergic and dual cholinergic and electrical interactions; often 2 or more of these types of interaction were observed on the same island. Furshpan et al. 7 have also ob-
served a heterogeneity of synaptic phenotype among neurons on a single island. It seems possible that the cholinergic synapses observed after a 48-h exposure to supplemented medium may have been formed by only a subpopulation of the neurons that is somewhat more sensitive to cholinergic induction. However, due to the small size of the cells, we were generally unable to record from all of the cells on an island and so cannot rule out the possibility that a low level of cholinergic function is being expressed by all of the neurons. While apparently heterogeneous in some respects, the population of neurons exposed to sup- plemented medium was homogenous in others; more than 95% of the neurons had the capacity to take up norepinephrine by a desipramine-sensitive mecha- nism 27 and a similar percentage of the cells had mole- cules of tyrosine hydroxylase within their somata. Furthermore, there was substantial dopamine-/3-hy- droxylase activity in homogenates of these cultures.
81
Thus, after a 48-h exposure to supplemented medi-
um, most neurons which have formed either cholin- ergic or electrical synapses have retained at a readily detectable level several of the properties appropriate to their sympathetic and adrenergic lineage.
It has previously been reported that sympathetic neurons can form either cholinergic2,7. 25 or electrical synapses10.21 under a variety of conditions in vitro.
However, this is the first report that sympathetic neu- rons, like some other types of motorneurons 3, can
form dual function electrical and cholinergic syn- apses in vitro; it therefore extends previous obser- vations as to the capacity of these autonomic neurons to use multiple transmitter systems 7. We have found that all of the dual function neurons which we have examined were expressing at least one of properties
of an adrenergic neuron. Thus, sympathetic neurons in vitro have the capacity to express simultaneously parts of 3 different transmission systems, and, under other conditions in vitro, to release 3 different neuro-
transmitters (norepinephrine, acetylcholine and adenosine) 25.
During neurogenesis, many neurons undergo brief interactions which cause long-lasting changes in their properties in vivo. For example, precursors of sym- pathetic neurons come into the vicinity of the neural tube, notochord and somatic mesenchyme for a few hours during their ventral migration and shortly thereafter these neuroblasts begin to synthesize cate- cholamines (reviewed by LeDouarin19). In this
study, we have observed a phenomenon in vitro which, at least in one respect, resembles changes oc- curring in vivo; we have found that a 2-day exposure to supplemented medium causes a long-lasting change in the functional properties of some sympa- thetic neurons that is still evident after 6 weeks in vit- ro. Since the exposure to supplemented medium is brief and since the inducing agent(s) can act without binding to the substrate, it seems possible to use in- hibitors of both nucleotide metabolism and process extension in this system to determine some of the events necessary to allow sympathetic neurons to ex- press cholinergic characteristics.
ACKNOWLEDGEMENTS
We are grateful to our colleagues R. P. Bunge and

82
M. I. J ohnson for thei r helpful discussions of this pro-
ject and to S. Mant i a for secre tar ia l aid. This p ro j ec t
was suppor t ed by N I H Gran t s NS 09808, NS 09923
and NS 14416 and by N I H Tra in ing Gran t NS 07071.
REFERENCES
1 Bottenstein, J. E. and Sato, G. H., Growth of a rat neuro- blastoma cell line in serum-free supplemented media, Proc. nat. Acad. Sci. U.S.A., 76 (1979) 514-517.
2 Burton, H, and Bunge, R. P., The expression of cholinergic and adrenergic properties by autonomic neurons in tissue culture. In P. G. Nelson and M. Lieberman (Eds.), Excita- ble Cells in Tissue Culture, Plenum Press, New York, 1981, pp. 1-36.
3 Fischbach, G. D., Synapse formation between dissociated nerve and muscle cells in low density culture, Develop. Biol., 28 (1972) 407-429.
4 Freschi, J. E., Effects of serum-free medium on growth and differentiation of sympathetic neurons in culture, Develop. Brain Res., 4 (1982) 455-464.
5 Fukada, K., Hormonal control of neurotransmitter choice in sympathetic neurone cultures, Nature (Lond.), 287 (1980) 553-555.
6 Furshpan, E. J., MacLeish, P. R., O'Lague, P. H. and Pot- ter, D. D., Chemical transmission between sympathetic neurons and cardiac myocytes developing in microcultures: evidence for cholinergic, adrenergic, and dual-function neurons, Proc. nat. Acad. Sci. U.S.A., 76 (1976) 4225-4229.
7 Furshpan, E. J., Potter, D. D. and Landis, S. C., On the transmitter repertoire of sympathetic neurons in culture, The Harvey Lectures, Series 76 (1982) 149-189.
8 Hawrot, E., Cultured sympathetic neurons: Effects of cell- derived and synthetic substrata on survival and devel- opment, Develop. Biol., 74 (1980) 136-151.
9 Henry, D. P., Johnson, D. G., Starman, B. J. and Wil- liams, R. H., Kinetic characterization of rat serum dopa- mine-beta-hydroxylase using a simplified radioenzymatic assay, Life Sci., 17 (1975) 1179-1186.
10 Higgins, D. and Burton, H., Electrotonic synapses are formed by fetal rat sympathetic neurons maintained in a chemically-defined culture medium, Neuroscience, 7 (1982) 2241-2253.
11 Higgins, D., Iacovitti, L., Burton, H. and Joh, T. H., The immunocytochemical localization of tyrosine hydroxylase within rat sympathetic neurons that release acetylcholine in culture, J. Neurosci., 1 (1981) 126-131.
12 lacovitti, L., Joh, T. H., Park, D. H. and Bunge, R. P., Dual expression of neurotransmission synthesis in cultured autonomic neurons, J. Neurosci., 1 (198l) 685-690.
13 Iacovitti, L., Johnson, M. I., Joh, T. H. and Bunge, R. P., Biochemical and morphological characterization of sympa- thetic neurons grown in chemically defined medium, Neu- roscience, 7 (1982) 2225-2239.
14 Joh, T. H., Geghman, C. and Reis, D. J., Immunochemical demonstration of increased accumulation of tyrosine hy-
droxylase protein in sympathetic ganglia and adrenal me- dulla elicited by reserpine, Proc. nat. Acad. Sci. U.S.A., 70 (1973) 2767-2771.
15 Johnson, M. I. and Argiro, V., Techniques in tissue culture of rat sympathetic neurons, Meth. Enzymol., 103 (1984) 334-347.
16 Johnson, M. I., Paik, K. P. and Higgins, D., Alteration from clear to dense-cored of the vesicle population in cul- tured cholinergic sympathetic neurons, in press.
17 Johnson, M. I., Ross, C. D., Meyers, E., Spitznagel, L. and Bunge, R. P. Morphological and biochemical studies on the development of cholinergic properties in cultured sympa- thetic neurons. I. Correlative changes in choline acetyl- transferase and synaptic vesicle cytochemistry, J. Cell Biol., 84 (1980) 680-691.
18 Ko, C.-P., Burton, H., Johnson, M. I. and Bunge, R. P., Synaptic transmission between rat superior cervical gangli- on neurons in dissociated cell cultures, Brain Res., 117 (1976) 461-485.
19 LeDouarin, N., The Neural Crest, Cambridge University Press, Cambridge, 1982.
20 O'Lague, P. H., Potter, D. D. and Furshpan, E. J., Studies on rat sympathetic neurons developing in cell culture. I. Growth characteristics and electrophysiological properties, Develop. Biol., 67 (1978) 384-403.
21 O'Lague, P. H., Furshpan, E. J. and Potter, D. D.. Studies on rat sympathetic neurons developing in cell culture. I1. Synaptic mechanisms, Develop. Biol., 67 (1978) 404-423.
22 Patterson, P. H., Environmental determination of auto- nomic neurotransmitter functions, Ann. Rev. Neurosci., 1 (1980) 1-17.
23 Patterson, P. H. and Chun, L. L. Y., The induction of ace- tylcholine synthesis in primary cultures of dissociated rat sympathetic neurons. I. Effects of conditioned medium, Develop. Biol., 56 (1977) 263-280.
24 Patterson, P. H. and Chun, L. L. Y., The induction of ace- tylcholine synthesis in primary cultures of dissociated rat sympathetic neurons. II. Developmental aspects, Develop. Biol., 60 (1977) 473-481.
25 Potter, D. D., Furshpan, E. J. and Landis, S. C., Transmit- ter status in cultured rat sympathetic neurons: plasticity and multiple function, Fed. Proc., 42 (1983) 1626-1632.
26 Schrier, B. K. and Shuster, L., A simplified radiochemical assay for choline acetyltransferase, J. Neurochem., 14 (1967) 977-985.
27 Wakshull, E., Johnson, M. I. and Burton, H., Persistence of an amine uptake system in cultured rat sympathetic neu- rons which use acetylcholine as their transmitter, J. Cell Biol., 79 (1975) 121-131.
28 Wolinsky, E., Patterson, P. H., Landis, S. C. and Willard, H., The influence of serum on the development of sympa- thetic neurons in culture, Soc. Neurosci. Abstr., 9 (1983) 4.