fd in potato
TRANSCRIPT

FD in potato Characterising the role of the transcription
factor FD homologue in potato
Mirjam Storcken
LABORATORY OF PLANT BREEDING,
WAGENINGEN UNIVERSITY

FD in potato Characterising the role of the
transcription factor FD homologue in
potato
Mirjam A. Storcken
920321809030
Master Thesis (MSc) in Plant Breeding
PBR-80436
Supervisors:
Dr. Christian Bachem
Dr. Sara Bergonzi
Dr. Guusje Bonnema
November 2018,
Laboratory of Plant Breeding, Wageningen University and Research Institute,
Wageningen, The Netherlands.

ii
Acknowledgements
I would like to thank Sara Bergonzi for supervising in the lab and help with writing
the report. Secondly I would like to thank my other supervisor Christian Bachem
for his help with writing the report. I would also like to thank Marian Oortwijk for
all her help in the lab and Jose Abelenda Vila for helping me with the yeast-two
hybrid. Lastly I would like to thank my family for believing in me that I was able to
do this and all the support they have given me throughout the whole time
especially to my husband William and my father Ton Storcken.

iii
Abstract
Potato (Solanum tuberosum L.) breeding is becoming more interesting the last
years, mainly due to diseases. However, there is a lot unknown about the
molecular mechanisms regulating potato flowering. It is known that StSP3D (FT
homolog of potato) has a role in flower initiation and in other plant species FT
homologues do this together with FD transcription factors. Potato has two StFD
genes, we named these StFD1 and StFD2. This thesis is about these StFD genes and
whether they are involved in flower initiation. By studying the expression in plant
material and the interaction with StSP3D, we found that StFD1 is mainly
transcribed in the leaves and stolon, and can interact with StSP3D. The expression
of StFD2 is primarily restricted to the apex, suggesting that StFD2 is the canonical
FD in potato that sits in the meristem and waits for the StSP3D. StFD2 and StSP3D
can interact and together they are most likely involved in flower initiation.
Key words: potato, StFD, StSPGB, flower initiation, gene expression, yeast-two
hybrid, StSP3D.

iv
Table of contents
1. Introduction .................................................................................................................... 1
2. Aims of the research ....................................................................................................... 5
3. Materials and methods- .................................................................................................. 6
3.1. Sequence comparison ...................................................................................... 6
3.2. Phenotyping and expression analyses ................................................................ 6
3.2.1. Experimental plant material ........................................................................ 6
3.2.2. Experimental set-up .................................................................................. 6
3.2.3. Experimental measurements and sampling.................................................... 7
3.2.4. 24-hour time course .................................................................................. 8
3.2.5. Primer design, development and validation ................................................... 8
3.2.6. Testing of reference genes .......................................................................... 9
3.2.7. RNA isolation, DNase treatment and cDNA synthesis..................................... 10
3.2.8. Quantitative PCR ..................................................................................... 11
3.3. Protein-protein interaction using Yeast-two-Hybrid assay .................................... 12
3.3.1. Amplification of coding sequences .............................................................. 12
3.3.2. Cloning in expression vector for Yeast-two-Hybrid ........................................ 12
3.3.3. Yeast-two hybrid ..................................................................................... 13
3.4. Data analyses .............................................................................................. 15
3.4.1. Phenotypic data ...................................................................................... 15
3.4.2. qPCR data .............................................................................................. 15
3.4.3. Yeast-two hybrid data .............................................................................. 15
4. Results ........................................................................................................................... 16
4.1 StFD genes in potato ...................................................................................... 16
4.2. Sequence comparison .................................................................................... 17
4.3. Phenotyping and expression analyses .............................................................. 18
4.4. 24-hour time course ...................................................................................... 20
4.5. Gene expression during development .............................................................. 21
4.5.1. Leaves ................................................................................................... 21
4.5.2. Apex enriched material ............................................................................ 23
4.5.3. Stolon ................................................................................................... 24
4.6. Yeast-two hybrid .......................................................................................... 25
5. Discussion ...................................................................................................................... 26
5.1. Sequence comparison .................................................................................... 26
5.2. Phenotyping and expression analyses .............................................................. 26
5.3. Gene expression during development .............................................................. 27
5.4. Yeast-two hybrid .......................................................................................... 28
6. Conclusion ..................................................................................................................... 29
7. Recommendation .......................................................................................................... 30

v
References......................................................................................................................... 31
Appendix 1: Primers .......................................................................................................... 36
Appendix 2: Sequences ..................................................................................................... 37

1
1. Introduction
The vast majority of the research on potato development (Solanum tuberosum L.)
focuses on tuber development and little is known about the flowering process. In
this research we focussed on flowering. Potato is a member of the family
Solanaceae or nightshades, along with other agriculturally important species such
as tomato (Solanum lycopersicum), eggplant (Solanum melongena), pepper
(Capsicum annuum) and Tobacco (Nicotiana tabacum). Potato is the third most
important world food crop, after wheat and rice (Ewing & Struik, 1992). It is
cultivated for the tubers, which are storage organs and represent a vegetative
propagation system. Potato tubers are rich in starch, protein and vitamins
(Augustin et al., 1978; Desborough & Weiser, 1974; Negi & Nath, 2002). The ploidy
level of cultivated potato ranges from diploid to hexaploid (Spooner et al., 2005).
Commercial cultivars are autotetraploid out breeders and as a result are highly
heterozygous and show significant inbreeding depression.
The origin of the modern potato cultivar, is from the high altitudes of the
equatorial Andes in South America (Spooner et al., 2005). The potatoes there,
grow under short days (12h of light), with a high light intensity, cool temperatures,
and relatively high humidity (Levy & Veilleux, 2007). In 1567, it was first recorded
that Andean potatoes were cultivated in Europe (Hawkes & Francisco-Ortega,
1993). It is likely that prior to the late blight epidemics in Ireland in 1845, cultivated
potato was crossed with the Chilean potato (Grun, 1979; Grun, 1990). Thus, most
modern potato cultivars contain Chilean germplasm (Huamán & Spooner, 2002).
The Chilean landraces are more adapted to long-day conditions (16h light/8h
darkness) (Ewing & Struik, 1992). The wild potato can still be found in 16 countries,
the main ones being Argentina, Bolivia, Mexico and Peru (Hijmans & Spooner,
2001).
Traditionally, potato propagation is done mainly via tubers, however, sexual
propagation via seeds has many advantages. Seeds can be stored longer, they are
easier to store and transport them in larger amounts (Hirsch et al., 2013). Plus,
numerous diseases such as viruses are not carried over via the seeds (Valkonen,
2007). Seed production is the easiest with diploids. However, since diploids are
usually self-incompatible, generating and maintaining a homozygous line remains
difficult. Introduction of the Sli gene renders diploid potato self-compatible
(Hosaka & Hanneman, 1988 a, b). Selfing also allows the development of
homozygous diploid parental lines (elite lines) that can be used for potato hybrid
breeding and opens the possibility of true-seed potato cultivar (Lindhout et al.,
2011). Hybrid technology makes the breeding process shorter and propagation via
seeds possible.

2
It has been shown that several
components of the tuberisation
pathway in potato are similar to
Arabidopsis thaliana flowering
pathway (figure 1.1). However, the
regulation of flowering in potato
remains to be elucidated in detail. In
flowering plants, floral development
is initiated by the expression of AP1
(APETALA1), LFY (LEAFY) and FUL
(FRUITFULL) (Abe et al., 2005; Turck
et al., 2008). These genes are
positively regulated by SOC1
(Suppressor of Overexpression Constants 1) and the protein-protein interaction
between FT (FLOWERING LOCUS T), FD mediated by 14-3-3 (Taoka et al., 2011).
This protein-protein interaction forms a hexameric florigen activation complex
(FAC) (Tsuji et al., 2013). A mutation in fd in rice has been identified that abolishes
binding with 14-3-3. In this mutant, the complex is unable to be transported into
the nucleus and therefore unable to function (Taoka et al., 2011). The mutation
was located in the interaction motif at the C-terminus of the FD protein, also
known as SAP (SRF [serum response factor] accessory protein) (Tsuji et al., 2013).
SOC1 is likewise positively regulated by the same protein-protein interaction
(Wigge et al., 2005). In Arabidopsis, FT is produced in the leaves and is transported
to the meristem (Chailakhyan, 1936). The expression is dependent on CO
(CONSTANS) (González-Schain et al., 2012). CO is downregulated by CDF (CYCLING
DOF FACTOR) and CDF is post-transcriptionally regulated by FKF (FLAVIN-BINDING,
KELCH REPEAT, F-BOXPROTEIN1) and GI (GIGANTEA) (Imaizumi et al., 2005). FKF
and GI are both regulated by the circadian clock (Sawa et al., 2007). In Arabidopsis,
FD is expressed primarily in the meristem, as this is the site of FD protein activity.
However, FD expression is also present in the root meristem and at lower levels
throughout the plant (Abe et al., 2005; Wigge et al., 2005). FD is also expressed
before the transition to flowering. This indicates that FD protein is not the limiting
factor for floral transition (Wigge et al., 2005).
FD consist of
three exons
(figure 1.2). The
first exon varies
between species and even between different FD alleles within species. The second
exon encodes a bZIP motif. This motif is present in many gene regulatory proteins
in eukaryotic cells (Tsuji et al., 2013). The bZIP motif contains an array of at least
Figure 1.2 The exons of FD. The first exon is not will conserved and is different between species. The second exon contains the bZIP motif, which is thought to be bind to DNA. Lastly, the third exon contains the SAP, which can bind to 14-3-3.
Figure 1.1 Flowering pathway in Arabidopsis thaliana. The different interaction between the different flowering genes (Andrés & Coupland, 2012).

3
five leucines, located at every seventh position facilitating the formation of eight
helical turns. This structure is thought to bind to DNA (Hakoshima, 2005). The third
exon comprises the SAP 14-3-3 binding domain (Tsuji et al., 2013).
It is thought that FD genes originate from the period in which seed plants emerged
and evolved after the emergence of angiosperms. This is assumed because the FD
gene has not been found in moss (bryophyte; Phycomitrella patens) or in the spike
moss (lycophyte, basal vascular plant; Selaginella moellendorffii) (Banks et al.,
2011; Rensing et al., 2008). If the FD gene did emerge from seed plants, then there
should be some evidence in gymnosperms, however this gene has not yet been
found (Tsuji et al., 2013). The function of FD has diversified during evolution and
FD homologs have acquired different roles that are independent from the
interaction with FT (Tylewicz et al., 2015).
In the potato
reference genome
two StFD
homologues were
annotated on
chromosome 2. We
named these StFD1
and StFD2. The
StFDs are also
known as StSPGBs
(Solanum
tuberosum Self-
pruning G-box)
after its tomato
homologue SPGB.
The protein
partners of StFDs
are FT
homologues. In
potato these
include StSP3D (Solanum tuberosum SELF PRUNING 3D) and StSP6A (Solanum
tuberosum SELF PRUNING 6A). StSP3D is involved in flowering, while StSP6A is
active in tuberigen (Navarro et al., 2011). The photoperiodic pathway has been
characterized with respect to expression of StSP6A and tuberisation. During this
thesis, other research showed that StFD1 interacts with StSP6A and form a FAC
together with 14-3-3. This interaction is thought to regulate the tuber formation
(Teo et al., 2017). The photoperiodic pathway for regulating flowering is quite
conserved between a wide variety of flowering plants (Andrés & Coupland, 2012).
Figure 1.3 Flowering pathway and tuberisation pathway in potato. The meristem part is based on tomato and the other parts on potato. StSP3D and StSP6A are produced in the leaves. StSP6A is involved in tuberisation, while in tomato SP3D is involved in flowering together with SPGB and 14-3-3 (Abelenda et al., 2014).

4
Figure 1.3 shows a possible flowering pathway for potato, which is based on the
pathway that is found in tomato. However, it was found that StSP3D promotes
flowering in potato, this based on delayed flowering in StSP3D knock-down plants.
Although the function of StFD is not known, it is likely to act in a similar way as was
shown in Arabidopsis and other plant species.
This research focused on StFD and its role in potato. We tried to recover this role
of both StFDs using six different genotypes. First, by comparing sequences we
looked for similarities between the potato StFDs and other FDs from different
plant species. It is found that StFD2 from potato is more similar to a SPGB from
tomato, than to StFD1. Due to this, it is expected that the function of the former
two will be similar. It is known that the tomato SPGB interacts with SINGLE
FLOWER TRUSS (homologue of SP3D) to trigger flowering (Pneuli et al., 2001).
StFD2 is located in a QTL for the unset of flowering (Hurtado-Lopez, 2012). For
these reasons, it is expected that StFD2 has a role in flowering. StFD1 interacts
with StSP6A via 14-3-3 and causes initiation of the tubers (Teo et al., 2017).
Therefore it is expected that StFD1 is higher expressed in the stolons, just before
tuber initiation.
In this report the aim of the research will be told first, followed by the methods.
The results will come next, tailed by the discussion, conclusion and
recommendation.

5
2. Aims of the research
To investigate the role of StFD1 and StFD2 in potato flowering, the research is split
into two different sections: 1) phenotyping plant growth and correlating the
phenotypes with gene expression analysis, 2) testing the interaction between the
different StFDs and StSP3D.
Phenotyping plant growth together with expression analysis of the genes
Genotype C, E, CE3027, and CE3130 (see Materials and Methods) were
phenotyped for developmental traits, in order to investigate whether differences
in growth correlate with the StFDs expression. With a particular focus on
flowering, plastochron length (i.e. time of appearance of consecutive leaves) and
flowering abortion. If a correlation is found between the observation of the start
of flowering and StFD expression, it is likely that the StFD is then involved in
initiating flowering.
Genes can be dependent on the circadian clock, which can reveal in which way the
gene functions. FD in A. thaliana is independent of the circadian clock (Wigge et
al., 2005). Therefore, it is expected that StFDs are also independent of the
circadian clock. To verify this a 24-hour time course was analysed, the samples
were coming from S. andigena.
Looking for interaction between different StFDs with StSP3D
If StFD is involved in flowering together with StSP3D it is likely that they are
interacting with each other, as found in different plant species (Abe et al., 2005; Li
& Dubcovsky, 2008; Sussmilch et al., 2015). The protein-protein interaction was
examined with a yeast-two hybrid approach, to find out whether the two StFD
proteins of S. andigena are able to form a FAC with StSP3D.

6
3. Materials and methods-
There are six different genotypes used for this research: (1) S. tuberosum group
Phureja DM1-3 516 R44 (hereafter referred to as DM), (2) S. andigena, (3) C, (4)
E, (5) CE3027 and (6) CE3130. DM is a double monoploid whose genome was
sequenced. In this research it is used as a base line to determine the sequence of
the genes. S. andigena is a tetraploid wild type which is used as a model
genotype for potato and is used as a reference for certain parts of this research.
Genotype C and E are heterozygous diploid potato developed for research
purposes. This development was based on the different genetic background of S.
phureja and S. tuberosum, and their phenotypic contrast (Jacobs et al., 1995).
In a previous experiment conducted in the glasshouse E flowered two weeks
earlier than C. Also, E formed tubers a bit quicker than C. The two were crossed
for QTL mapping and CE3027 and CE3130 were created here. CE3027 forms tubers
relatively late and CE3130 forms tubers relatively early, compared to genotype C
and E. However, they flower around the same time, between the flowering time
of genotype E and C.
3.1. Sequence comparison
The amino acid sequence of the StFD conceptually translated proteins and their
allelic variants were aligned with other similar bZIP transcription factors, retrieved
from the public repository NCBI. The alignment program used is Clustal Omega
Multiple Sequence Alignment (http://www.ebi.ac.uk/Tools/msa/clustalo/).
Additional information was used from PGSC and ITAG annotations.
3.2. Phenotyping and expression analyses
3.2.1. Experimental plant material
Plant material was propagated from in vitro plantlets maintained in the
Wageningen Plant Breeding collection. Plants were grown on standard media for
plant growth (Murashige and Skoog MS20 L-1: 4.4g MS+Vitamine, 20g sucrose, pH
5.8 [with KOH] and 8g agar, and final step autoclaving to sterilize). Cuttings were
made consisting out of at least two leaves and transferred onto new standard
media. A second round of only apical propagation was taken a week later to
equalise the developmental stage of the plantlets. All in vitro plant material was
kept in a climate chamber at 24o C and under long day conditions (16h light/8h
darkness; LD). This holds for genotypes: C, E, CE3027 and CE3130 in this research
but also for the plant material from which RNA/cDNA has been used in this
experiment.
3.2.2. Experimental set-up
After four weeks in vitro, the plants were potted in 14cm pots using Potground #4
(supplied by Horticoop). At this time each plant had exactly five leaves. The plants

7
were grown in a climate chamber under LD, with a light intensity of 200μ mol m-2
s-1, 80% relative humidity and a day/night temperature of 20o C/18o C. The
moisture content of the pots was monitored and pots were watered according to
requirements. For the first three days the plants had a transparent plastic cup
covering them to raise the humidity and to prevent dehydration.
For phenotyping 15 individual plants of each genotype were used. For expression
17 plants from genotype E and 24 plants from genotype C were used. The two
groups were (for genotype and expression) kept separate from each other and
they were manually randomized.
3.2.3. Experimental measurements and sampling
Phenotyping
Leaf, flower, stolon and tuber development have been examined over seven
weeks. Plastochron interval was recorded by keeping track of the number of new
leaves developed per week. This was achieved by piercing a hole in the youngest
leaf at every observation. Where the youngest leaf is the leaf that has been formed
latest on the main stem being at least 0.5cm wide.
Flower development was only recorded at the apical meristem on the main stem.
It was examined on a daily basis when the plants had flowering buds, had
undergone anthesis, and flower abortion occurred. Formation of flower buds was
considered as entry into the generative phase. Anthesis on the other hand, is when
the flower opens. Comparison of bud formation and anthesis was done to verify
transition time. This comparison also highlights possible differences between the
genotypes. Flower abortion is when the plants terminates flowering and therefore
flowering stops and the flowers will wilt. This can occur at different stages before
and after anthesis.
Tuber development was phenotyped weekly, only for genotypes E and C. This to
assess a correlation between flowering and tuberisation or flower abortion and
tuberisation. In figure 3.1 shows the different developmental stages of
tuberisation. It was scored when the plants had formed stolon (stage one) and
when tubers were starting to develop (stage three). Plants used for tuber
phenotyping were not used for any further phenotyping.

8
Sampling of a developmental Time course
Samples were taken weekly, from genotypes C and E to examine the gene
expression in different tissues and over time. For each sampled three plants were
pooled together. Except for the second and third sampled where four plants were
pooled together. Each sampling was done in duplicate. The sampling started when
the plants had been in the soil for one week. For genotype E samples were taken
for five weeks after planting, while genotype C was sampled for seven weeks. The
difference of sampling time is based on previous research where C appeared to
flower later then E. Likewise, gene expression from C was carried out longer.
Samples were taken from the apex enriched meristem (in a vegetative o
generative state depending on the time point), the fifth leaf counted from the top
of the plant and the stolon/tubers if present. After harvesting the samples were
immediately frozen in liquid nitrogen and stored at -80o C and the sampled plants
were destroyed.
3.2.4. 24-hour time course
In previous research, a 24-hour time course was sampled using S. andigena,
where, whole above ground plants material was sampled every three hours. The
plants were grown in the same conditions as for the earlier explained time course
(3.2.2. Experimental set-up), except only half of the plants were under LD. The
other half were exposed to short day conditions (8h light/16hrdarkness; SD).
Plants were approximately three weeks on soil. After collection, the samples were
immediately frozen in liquid nitrogen and stored at -80o C.
3.2.5. Primer design, development and validation
Primers were used for qPCR and for cloning of the whole coding sequence (CDS)
for the protein-protein interaction. Some of the primers were already developed
and validated. The other primers were designed based on the information from
previous research. This by comparing the sequences with the transcript sequence
from DM of the PGSC Genome Browser, to determine any differences. The primers
Figure 3.1 Potato tuber development stages, over a time period of 25 days after a switch from a 16 hours light per day to an 8 hours light period (Kloosterman et al., 2005).

9
were designed without any known mismatches to the reference sequence (DM for
S. andigena, for C and E low coverage whole genome sequence is available) and
that the forward and reverse primer do not align to each other’s (primer dimer).
Primer3 software was used to design primer pair when possible. The melting
temperature was calculated with Oligonucleotide Properties Calculator software.
Self-complementarity was checked with the same program. In case there were no
problems the sequence was compared with the PGSC DM Transcripts (v.3.4) using
the BLASTN nucleotide search (Altschul et al., 1997).
Primers were tested on cDNA, using the Thermo Scientific Dream Taq DNA
Polymerase and GeneAmp® PCR System 2700 for amplification of nucleic acids.
Each PCR reaction (25µL) consisted out of 1µL cDNA (10ng µL-1 concentration),
2.5µL DreamTaq buffer (10x), 1µL Forward Primer (10µM), 1µL Reverse Primer
(10µM), 2.5µL dNTP Mix (2mM each), 0.75µL DreamTaq polymerase, and 16.25µL
Milli-Q. Water-controls were included by replacing cDNA with Milli-Q. The initial
denaturation step was at 95o C for 1:00, the denaturation (95o C for 0:30),
annealing (60o C for 0:30) and extension (for this specific gene 72o C for 1:00) steps
were 35 times repeated. The final extension was at 72o C for seven minutes
followed by a hold at 4o C.
Per 5µL PCR product 1µL of loading dye was added to the PCR product, for running
a gel on a 1-2% agarose gel using 0.5 X TBE buffer for ± 45 minutes at 100V with a
1kb+ ladder for reference. After running the gel was photographed and the image
was examined to find out whether the primer pairs gave a single clear band. A full
list of the primers is given in Appendix 1.
Primer amplification efficiency was checked using a twofold dilution series of
pooled cDNA samples. A linear regression was calculated with the Bio-RAD CFX
Manager 3.0 software and the reaction efficiency (E) was calculated. It shows the
amount of target is produced with each cycle, an efficiency of 100% means that
the target amplicon is doubling with each cycle. The coefficient of determination
(R2) measures how correct the regression line describes the data, the closer to one
the better it fits. Efficiencies in the rage of 90-110% are generally deemed
acceptable (Pfaffl, 2001).
3.2.6. Testing of reference genes
Three reference genes were tested: NAC, EIF3e and Actin. They are sourced from
a previous research where they were tested on samples from potato (Bourke,
2014; Kloosterman et al., 2013). For selecting the most suitable reference gene,
they were tested on available cDNA from leaf and apex enriched tissues of S.
andigena. The plants were grown under similar conditions as explained earlier.
The only difference was that half of the plants were transferred after one week in

10
LD conditions to SD conditions and sampling happened over a period of four
weeks.
Figure 3.2 depicts the non-normalized expression of the three reference genes.
Actin variates in expression over different time points, between LD, SD, and
especially different tissue with about two Cts difference between leaves and
apices. EIF3e and NAC do not fluctuate much over time and different light regimes
lead to similar expressions. However, there is a difference in expression between
different tissues, this difference is bigger for NAC than for EIF3e. Due to this EIF3e
was chosen for the reference gene.
3.2.7. RNA isolation, DNase treatment and cDNA synthesis
For RNA isolation the frozen samples were grinded using liquid nitrogen, in order
to prevent RNA degradation. RNA was extracted from the frozen powder using a
Qiagen RNeasy Mini Kit (Cat. No. 74104 and 74106). The RLT buffer was used,
which is a lysis buffer for lysing cells and tissues prior to RNA isolation. This by
placing the RNeasy spin column in a new 2mL collection tube and discard the old
collection tube with the flow-through. The samples are then centrifuged on full
speed for one minute. Purified on-column RNA was collected in 30µL of RNase-
free water in a sterile Eppendorf tube. This was done twice resulting in a final
volume of 60µL. For the stolon tissue, which are rich in starch, another buffer was
used instead of RLT buffer, due to that this tissue contains more starch. To make
this buffer, 600µL Z-6 buffer (8M guanidinium-HCl, 20mM MES, 20mM EDTA [pH
7.0] and 0.7% [v/v] 2-mercaptoethanol) was thoroughly mixed with 500µL PCI
solution (phenol: chloroform: isoamylalcohol= 25:24:1) and subjected to
centrifugation (20,000xg, 10 minutes, 4o C) for phase separation. Only the upper
layer is used for buffer.
15,00
17,00
19,00
21,00
23,00
25,00
27,00
29,00
LDw1
LDw2
SDw2
LDw3
SDw3
LDw4
SDw4
LDw1
LDw2
SDw2
LDw3
SDw3
LDw4
SDw4
Leaves Apical meristem
Ct
EIF3e
Actin
NAC
Figure 3.2 Expression in different leaves and apex enriched material tissue over time, of the reference genes: EIF3e, Actin and NAC. LD stands for long day and SD for short day.

11
RNA concentration was measured with Thermo Scientific’s NanoDropTM 1000
Spectrophotometer. The concentrations of a 1µL sample of the purified RNA was
measured in ng µL-1 and the sample purity was assessed by examining the
absorbance ratio at 280/260nm and 260/230nm. The quality of the RNA was then
checked with a gel electrophoresis.
Estimated from the values obtained using the Nanodrop spectrohotomerter, 1µg
of RNA was treated with Deoxyribonuclease I (Amplidication grade) to eliminate
any residual DNA in the samples. This was done by adding 4µL RNA (with a
concentration of 250ng µL-1), 1µL 10X DNase I Reaction Buffer, 1µL DNAse I (Amp
Grade 1U µL-1) and 4µL Milli-Q water. The tubes were incubated at room
temperature (22o C) for 15 minutes. 1µL of 25mM EDTA solution was added to the
mixture to inactivate DNase I. The samples were heated for ten minutes at 65o C.
To synthesize cDNA (iScript cDNA Synthesis kit, Biorad), the following was added
to the RNA: 4µL 5x iScript reaction, 1µL iScript Reverse Transcriptase and sufficient
RNase-free water to make up a total reaction volume of 20µL. The samples were
kept for five minutes at 25o C, for 30 minutes at 42o C, for five minutes at 85o C, for
five minutes at 4o C, and finally for five minutes at 85o C. Thereafter they were
cooled down to 4o C. Synthesised cDNA was diluted with 130µL deionised water
(MQ), making a final concentration of 6.67ng µL-1. The samples prepared for the
dilution curve (in qPCR) were kept at higher concentration by diluting them with
100µL MQ, making a final concentration of 8.33ng µL-1. All samples were stored at
-4o C and placed at 4o C before using it for PCR.
3.2.8. Quantitative PCR
Quantitative (Real-Time) PCR (qPCR) was performed on a 96-well Bio Rad C1000
Thermal Cycler using iQ-SYBR-green super mix. All reactions were performed in
triplicate (technical replicates) and water-controls (containing MQ instead of
cDNA) were included for all primer pairs. Each well contained 10µL consisting out
of: 5µL SYBR-green, 0.25µL forward primer (10µM), 0.25µL reverse primer (10µM),
0.5µL MQ, and 4µL diluted cDNA.
The qPCR programme was in four stages: 1) 95o C for 30 minutes; 2) 40 cycles of
95o C for 15 seconds and 60o C for one minute followed by plate read; 3) 95o C for
10 seconds; 4) melting-curve analysis from 65o C-95o C in 0.5o C increments.
This was all done for genotypes C, E and S. andigena. With sample tissues coming
from apex enriched material and leaves for all three genotypes. Further for
genotype C and E stolon samples were taken. The RNA expression was measured
for the genes EIF3e, StFD1, StFD2, StSP3D and StAP1 in the eight different sample
tissues, with an exemption for StAP1 in the stolon sample tissues. All together
forming 38 different tests.

12
3.3. Protein-protein interaction using Yeast-two-Hybrid assay
3.3.1. Amplification of coding sequences
The coding sequences of the gene of interest were amplified from the cDNA of S.
andigena, C and E. This was achieved using a Thermo Scientific Phusion High-
Fidelity DNA Polymerase and GeneAmp® PCR System 2700 for amplification of
nucleic acids. Each PCR reaction (50µL) consisted out of: 3µL cDNA (10ng µL-1
concentration), 10µL 5X Phusion HF Buffer, 2µL Forward Primer (10µM), 2µL
Reverse Primer (10µM), 1µL dNTPs mix (10mM), 0.5µL Phusion DNA Polymerase,
31.5µL Milli-Q. Water-controls were included by replacing cDNA with Milli-Q.
The initial denaturation step was at 98o C for 30 seconds, the denaturation (98o C
for 10 seconds), annealing (61o C for 30 seconds) and extension (72o C for 15
seconds) steps were repeated 35 times in the cycle. The final extension was at 72o
C for seven minutes followed by a hold at 4o C. Per 5µL PCR product 1µL of loading
dye was added to the PCR product, for running a gel on a 1-2% agarose gel using
0.5 X TBE buffer for ±45 minutes at 100V with a 1kb+ ladder for reference. After
running the gels were photographed and the image was examined and the
amplified DNA was extracted from the gel.
The amplified fragment was purified from the gel by using a ZymocleanTM Gel DNA
Recovery Kit. Using the Nanodrop spectrophotometer the concentration of DNA
was measured and validated. In the gel, 1µL of the DNA was runned again to make
sure that only one band was purified. The gel was made the same way and run in
the same conditions as previously described. When only one single band was
visible the DNA was ready to be transferred into Escherichia coli. This was done for
one of the alleles of StFD1, StFD2, StSP3D and StTFL1 coming from S. andigena.
3.3.2. Cloning in expression vector for Yeast-two-Hybrid
The first step for the cloning is a ligation in the gateway donor vector using TOPO®
Cloning Reaction. The ligation mix consisted of: 1µL Salt Solution, 1µL TOPO®
vector, 0.5-4µL fresh PCR product, and MQ water was added till a final volume of
6µL. The molar ratio PCR to product:TOPO® vector should be between 0.5:1 and
2:1. The reaction was gently mixed and incubated at room temperature (22o C) for
30 minutes.
To chemically transform competent E. coli cells 3µL of the earlier mentioned
reaction was put together with 15-20µL of E. coli TOP10 cells and put on ice for 30
minutes, heat-shocked at 42o C for 30 seconds and immediately transferred back
to ice. Per reaction 250µL S.O.C. Medium was added and was shaken horizontally
(200 rpm) at 37o C for one to three hours. Of the reaction was 100µL on a LB agar
plates with 50µg mL-1 kanamycin (for pENTR/TOPO) and left overnight at 37o C.
Growing colonies were picked and grown, overnight, in LB liquid media containing
antibiotics.

13
The plasmids were extracted using plasmid miniprep (QIAprep Spin Miniprep Kit)
and the presence of the wanted gene tested using M13 primers (designed on the
plasmids) in a PCR reaction. The plasmids of the positive amplified clones were
sent for sequencing to check whether the gene is complete and whether it was
correctly situated in the plasmid. The correct plasmids were used for the gateway
LR reaction, to transfer the genes in the destination vectors (pGADT7 [150ng µL-1]
and pGBKT7 [300ng µL-1]). This by making two different mixtures with both 1µL
PCR product (20ng µL-1), and either 2µL destination vector pGADT7 or 1µL of
pGBKT7 and 1µL 1X TE buffer (pH 8.0) for pGADT7 or 2µL for pGBKT7, making an
end volume of 4µL for both mixtures. LR ClonaseTM II Plus enzyme mix was kept on
ice for 2 minutes. Afterwards 1µL was added to every mix and incubate the
reaction for one hour at 25o C. After this 1µL of proteinase K solution was added,
mixed briefly and incubated for ten minutes at 37o C. Transformation into E.coli
cells was performed by heat-shock at 42o C for 30 seconds and immediately
transferred back to ice. Per reaction 250µL S.O.C. Medium was added and was
shaken horizontally (200 rpm) at 37o C for one to three hours. Of the reaction
100µL was plated on LB agar plates with 50µg mL-1 kanamycin (pGBKT7) or
ampicillin (pGADT7) and left overnight at 37o C. Growing colonies were picked and
grown, overnight, in LB liquid media containing antibiotics. The plasmids were
extracted using small scale plasmid preparation (QIAprep Spin Miniprep Kit) and
the presence of the wanted gene tested using the T7 forward primer (on the
plasmid) and the specific reverse primer in a PCR reaction. The plasmids of the
positive amplified clones were sent for sequencing, to determine whether the
gene is complete and whether it was correctly situated in the plasmid. The
confirmed plasmids were used for the yeast-two hybrid (Y2H).
3.3.3. Yeast-two hybrid
The Y2H system was used to determine whether different proteins can interact
with each other within a cell. In our case for example, it was used to find this out
for FD proteins and StSP3D protein. The target protein was fused to a DNA binding
domain that directs the fusion protein to the regulatory region of a reporter gene
as “bait” (pGBKT7). In case this target protein binds to another specially designed
protein (“prey”, pGADT7) in the cell nucleus of the yeast, their interaction binds
two halves of a transcriptional activator. This then switches on the expression of
the reporter gene. The pGADT7 (prey) contains TRP1 nutritional marker and
pGBKT7 (bait) contains LEU2 nutritional marker. Therefor the plates used to check
yeast grow, showing that it was carrying both plasmids, contained histidine and
adenine. If there was growth it was grown on plates without histidine and adenine,
to get a better population.
Preparation of fresh yeast competent cells

14
For yeast transformation the lithium acetate method was followed. The Gold yeast
strain was used from a stock plate with separate colonies (with a diameter of 2-
3mm) and grown in YPD (this is liquid media that contains per 250mL: 12.5g Difco
YPD Broth, 7.5g purified agar and 245MQ). The sample was vortexed vigorously
for five minutes to disperse any clumps. Then, the sample was then put in a flask
containing 50mL of YPD and thereafter was incubated at 30o C for 16-18 hours
while being shaken at 250rpm to a stationary phase (OD600>1.5). In a new flask
containing 100mL YPD, the sample was added to reach an OD600 concentration
between 0.2-0.3 and incubated at 30o C for three to five hours while being shaken
(230rpm). The incubation time was depending on the OD600 value. This value
should get to an OD600 of 0.4-0.6. The cells were placed in a 50mL tube and
centrifuged at 3000xg for five minutes at room temperature (22o C). The
supernatant was discarded and the cell pellet was re-suspended with TE and after
that transferred to a new tube. It was centrifuged again at 3000xg for five minutes
at room temperature (washing step). The supernatant was discarded and the cell
pellet re-suspended with 1.5mL of freshly prepared 1X TE/1X lithium acetate (LiAc)
(competent cells ready for transformation).
Yeast transformation and selection
For transformation 0.1µg plasmid DNA (10µL per plasmid: pGADT7 [100ng] or
pGBKT7 [100ng]) and 0.1µg Fish (before using needs to be desaturated by boiling
for two to five minutes at 95o C) were added. This was done separately for all the
genes. Thereafter this was put in a 2mL tube and mixed. 0.1mL of yeast competent
cells was added and vortexed before adding 0.6mL of TE/LiAc/PEG (1mL TE 20X
1mL-1 LiAc10x 8mL-1 PEG 50%) solution and vortexed for ten seconds. The tubes
were then incubated at 30o C for 30 minutes (where being shaken at 200rpm).
From DMSO was 70µL added and mixed gently by inversion. The samples received
a heat shock for 30 minutes in a 42o C water bath, followed by a cool down on ice
for one to two minutes. The supernatant was removed after centrifuging the cells
for five seconds at 14,000rpm at room temperature. The cells were re-suspended
with 0.25mL 1X TE buffer and then 100µL plated on SD agar plates (-2aa [3.35g
nitrogen base, 10g glucose, 0.3 ODD supplement, 0.22g adenine, 0.22g histidine,
and MQ added to 500µL] and -4aa [3.35g nitrogen base, 10g glucose, 0.3 ODD
supplement, and MQ added to 500µL]). This was to select for the desired
transformation (-2aa) and for the interaction (-4aa). The plates were incubated for
two to four days at 30o C. Colonies were picked and diluted 10, 100 and 1000 times.
They were drop plated in triple on -2aa liquid media and grown overnight. If
growth is observed it indicates that an interaction is found, while if there is no
growth it indicates an absence of interaction between the two proteins.

15
3.4. Data analyses
3.4.1. Phenotypic data
Data was exported in Excel and analysed with IBM SPSS Statistics version 22. The
means were analysed for significant differences between genotypes using ANOVA
and LSD with an α=0.05.
3.4.2. qPCR data
qPCR data was extracted with CFX Manager (Bio-Rad) version 3.0 and imported
into Excel for further analysis. The mean of the estimated starting quantity (SQ)
and the associated standard deviation (SD) are used for expression analysis. The
expression of the gene of interest is the SQ of this gene divided by the SQ of the
reference gene. With the associate SD calculated by:
√𝑆𝑄 𝑔𝑒𝑛𝑒 𝑜𝑓 𝑖𝑛𝑡𝑒𝑟𝑒𝑠𝑡
𝑆𝑄 𝑟𝑒𝑓𝑒𝑟𝑒𝑛𝑐𝑒 𝑔𝑒𝑛𝑒𝑋 (
𝑆𝐷 𝑔𝑒𝑛𝑒 𝑜𝑓 𝑖𝑛𝑡𝑒𝑟𝑒𝑠𝑡
𝑆𝑄 𝑔𝑒𝑛𝑒 𝑜𝑓 𝑖𝑛𝑡𝑒𝑟𝑒𝑠𝑡)
2
+ (𝑆𝐷 𝑟𝑒𝑓𝑒𝑟𝑒𝑛𝑐𝑒 𝑔𝑒𝑛𝑒
𝑆𝑄 𝑟𝑒𝑓𝑒𝑟𝑒𝑛𝑐𝑒 𝑔𝑒𝑛𝑒)
2
T-test and ANOVA, both with LSD, were used to statistically analyse the data with
α=0.05. This to find out whether there was a significant difference between the
genotypes and between the different samples within a genotype. The SD is based
on two biological replicates and for each biological replicate has 3 technical
replicates. IMB SPSS Statistics version 22 was used for this.
3.4.3. Yeast-two hybrid data
To calculate the cotransformation of one of the plasmid types, the amount of
growing colonies have to be counted (cfu). This from the dilution drop plates. The
formula to calculate the cotransformation (cfu ng-1 DNA) is:
𝑉𝑜𝑙. 𝑝𝑙𝑎𝑡𝑒𝑑 (𝜇𝐿)𝑋 𝑑𝑖𝑙𝑢𝑡𝑖𝑜𝑛 𝑓𝑎𝑐𝑡𝑜𝑟 𝑋 𝑎𝑚𝑜𝑢𝑛𝑡 𝑜𝑓 𝐷𝑁𝐴 𝑢𝑠𝑒𝑑 (𝜇𝑔)

16
4. Results
4.1 StFD genes in potato
During genome sequencing two StFD homologues genes were annotated, both are
located on chromosome 2. We named these StFD1 (PGSC0003DMT00061403; also
known as StFDL1) and StFD2 (PGSC0003DMT00009393; also known as StFD; Teo
et al., 2017) (figure 4.1). StFD2 is located in a QTL interval found by Hurtado-Lopez
(2012) for the onset of flowering and is therefore a potential candidate gene for
being involved in flowering. Looking at the publicly available gene expression data,
from the potato genome browser (solanaceae.plantbiology.msu.edu), StFD1
appears to be more expressed throughout the plant, while StFD2 is mainly
expressed in the meristem. While the role of the two StFD genes in potato
flowering has not yet been established, it was found that StFD1 is involved in the
regulation of tuber formation together with StSP6A (Teo et al., 2017).
Previous flowering phenotyping showed that genotype E flowers earlier than
genotype C, while both genotypes form tubers around the same time. StFD1 and
StFD2 loci have different allelic variants in C and E genotypes (Willemsen,
unpublished). The sequences of the different alleles are shown in Appendix 2.
These differences in flowering phenotype and gene sequences of E and C make
them ideal to determine possible expression differences and to find out whether
there is a correlation between the StFDs and flower initiation. Due to limited time,
however, no distinction was made in allelic expression and was only looked at the
difference of gene expression from StFD1 and StFD2.
Figure 4.1 Chromosome 2 with the two StFD genes. StFD1 is the first one (positioned at around 20 Mb) and StFD2 is the second (positioned at about 39 Mb; solanaceae.plantbiology.msu.edu).

17
4.2. Sequence comparison
Next to sequences of StFD from genotype S. andigena and DM, 50 other sequences
were found of different plant species, which all contain a bZIP motif at the c-
terminus part of the encoded transcription factor. Figure 4.2 shows a tree where
all the available sequences are compared with each other. It is clear that the
sequences from Solanaceae and Brassicaceae form individual clades. StFD2 is
more similar to the SPGB gene from tomato (S. lycopersicum), than to StFD1. This
might indicate that the function of StFD2 is the same as the function of the SPGB
gene in tomato.
Figure 4.2 The of the StFD conceptually translated proteins and their genetic variants compared with other similar bZIP transcription factors gathered from the public repository NCBI.

18
4.3. Phenotyping and expression analyses
In order to find out if the StFD genes were involved in flowering, the growth and
development of the plants were measured and observed. It was found that the
flowering times of the four genotypes (C, E, CE3027, and CE3130) differed
maximally two days, where E and CE3027 were the earliest and C was the latest
(table 4.1). The same difference is observed for anthesis. For genotype CE3130
only two plants had opened one flower before both plants aborted flowering, the
rest of the plants had flower abortion before anthesis could happen. The E and
CE3027 genotypes formed more side shoots than CE3130 and C. Though the C
genotypes were forming side shoots at the end of the experiment. Next to this C
finished with flowering at the earliest time after 52 DAP (days after planting).
When examining the tuberisation, genotype E had formed stolons after 18 DAP,
while only 50% of the C plants had formed stolons. After 25 DAP, all plants from
both these genotypes had formed stolons. At 32 DAP these stolons developed into
tubers, while the C genotypes had fewer but larger tubers than the E genotypes.
Table 4.1 Observations of apical flower bud, anthesis and flower abortion for genotype E, CE3027, CE3130 and C.
As described in section 3.2.2. Experimental set-up each plant had five leaves at
the time of transplanting from in vitro media to soil. Over time, leaf development
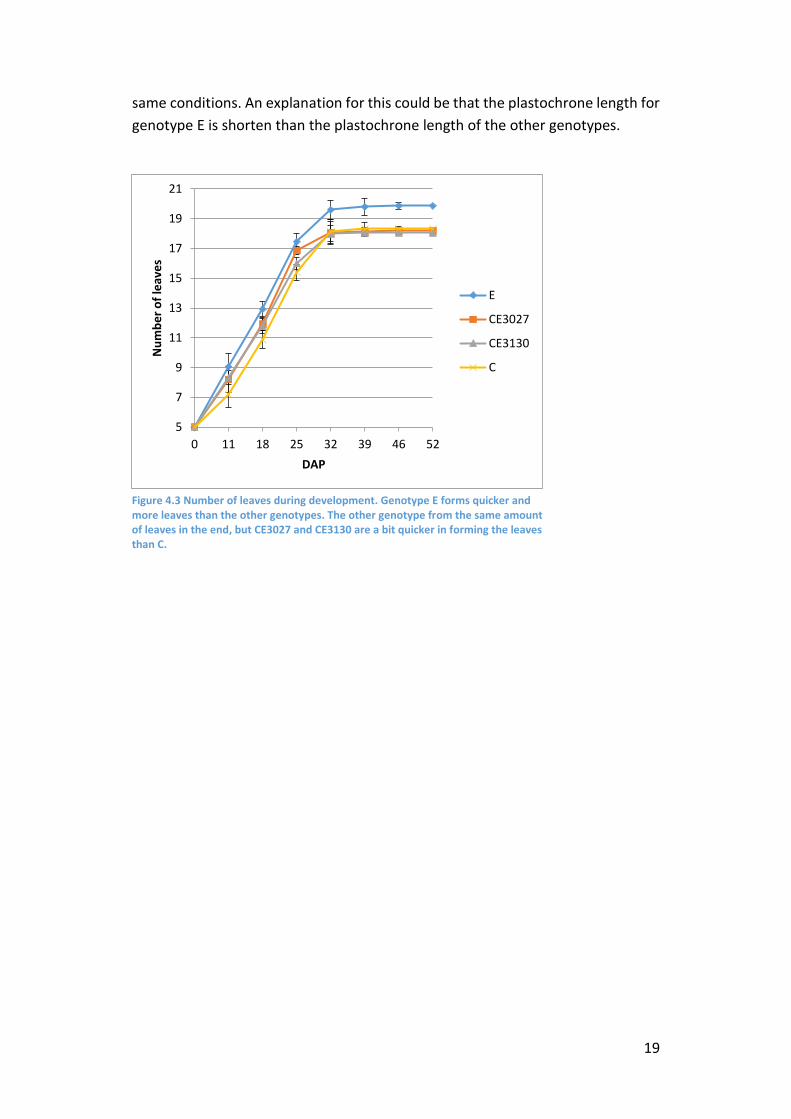
shows that genotype E formed significantly (p<0.002) more leaves than the other
genotypes (figure 4.3). After 23-25 DAP the plants started flowering. It is well
known that leaf development stops if on the same shoot the plant starts flowering.
However, here the plants still formed four to five leaves. This was because these
leaves were undersized before they were counted.
We can conclude that there is a small difference in flowering time between
genotype E and C. Further, genotype CE3130 was the only genotype with flower
abortion. Also was found, that genotype E forms more leaves than the other
genotypes, which all formed a similar amount of leaves and were grown under the
Apical flower bud (days) Anthesis (days) Aborts flowering (days)
E 23 ± 0 37 ± 0.41
CE3027 23 ± 0.26 37 ± 1.68
CE3130 24 ± 0.88 40 ± 2.83 43 ± 0.26
C 25 ± 1.13 40 ± 1.96
19
same conditions. An explanation for this could be that the plastochrone length for
genotype E is shorten than the plastochrone length of the other genotypes.
5
7
9
11
13
15
17
19
21
0 11 18 25 32 39 46 52
Nu
mb
er
of
leav
es
DAP
E
CE3027
CE3130
C
Figure 4.3 Number of leaves during development. Genotype E forms quicker and more leaves than the other genotypes. The other genotype from the same amount of leaves in the end, but CE3027 and CE3130 are a bit quicker in forming the leaves than C.

20
4.4. 24-hour time course
Genes can be dependent on the circadian clock, which might reveal their function.
In order to find out if StFD1 and StFD2 are regulated by the circadian clock a 24-
hour time course was performed. We found that the relative expression of StFD1
was higher than the relative expression of StFD2 this for both LD and SD (figure
4.4). The StFD1 relative expression goes down throughout the day and goes up
during the night. Indicating that it is regulated by the circadian clock, this for both
LD and SD conditions. However, we can’t conclude whether StFD2 is regulated by
the circadian clock, as the relative expression is too low.
Figure 4.4 The relative expression of StFD1 and StFD2 in S. andigena, over a period of 24 hours. Zt means zeitgeber time. The dark grey part is the dark period for the long day (LD). The light and dark grey parts form together the dark period for the short day (SD).

21
4.5. Gene expression during development
To find out whether the StFD genes were involved in flowering, the mRNA
production in different plant tissue was measured. In previous research it was
found that E flowered approximately two weeks earlier than C. Based on these
results we chosen to grow E for only five weeks and C for seven weeks. In order to
find out where StFD1, StFD2, StSP3D, and StAP1 are expressed, we took samples
from different plant material: leaf, apex enriched material, and stolon.
4.5.1. Leaves
In the leaf samples was found that StFD1 is significantly higher expressed than
StFD2, this is especially the case in genotype C (figure 4.5). StFD1 mRNA expression
increased in the leaves during development and reached a maximum after five
weeks, which is just one week after the floral buds were first observed and is
significant from the rest of the weeks (P<0.01). For StFD2 there was no difference
between the two genotypes and it was very low expressed in the leaves. StSP3D
mRNA was also low expressed but there were two significant peaks for E (P<0.02).
These peaks occurred just after the plants started flowering. As StFD2 also StAP1,
which is a marker for the floral transition in the meristem, is least expressed in the
leaves. Although in genotype E increased in expression and had a peak in week
four which was significant to all of the other time point except for week five
(P<0.03). The expression of StSP3D is low, except in genotype E which has an
increase in expression in week 4 and 5.

22
0
2
4
6
8
10
1 2 3 4 5 6 7
StFD
1/E
IF3
e
Time (weeks)
StFD1E
C
0,00
0,05
0,10
0,15
0,20
1 2 3 4 5 6 7
StFD
2/E
IF3
e
Time (weeks)
StFD2E
C
0,00
0,10
0,20
0,30
0,40
0,50
1 2 3 4 5 6 7
StSP
3D
/EIF
3e
Time (weeks)
StSP3DE
C
0,000
0,005
0,010
0,015
0,020
0,025
1 2 3 4 5 6 7
StA
P1
/EIF
3e
Time (weeks)
StAP1E
C
Figure 4.5 Expression of different genes in leave tissue of genotype E and C. Genotype E started flowering just after three weeks DAP and C just before four weeks DAP. The expression of the genes in genotype E was only measured for the first five weeks and for C the expression was measured for seven weeks.
23
4.5.2. Apex enriched material
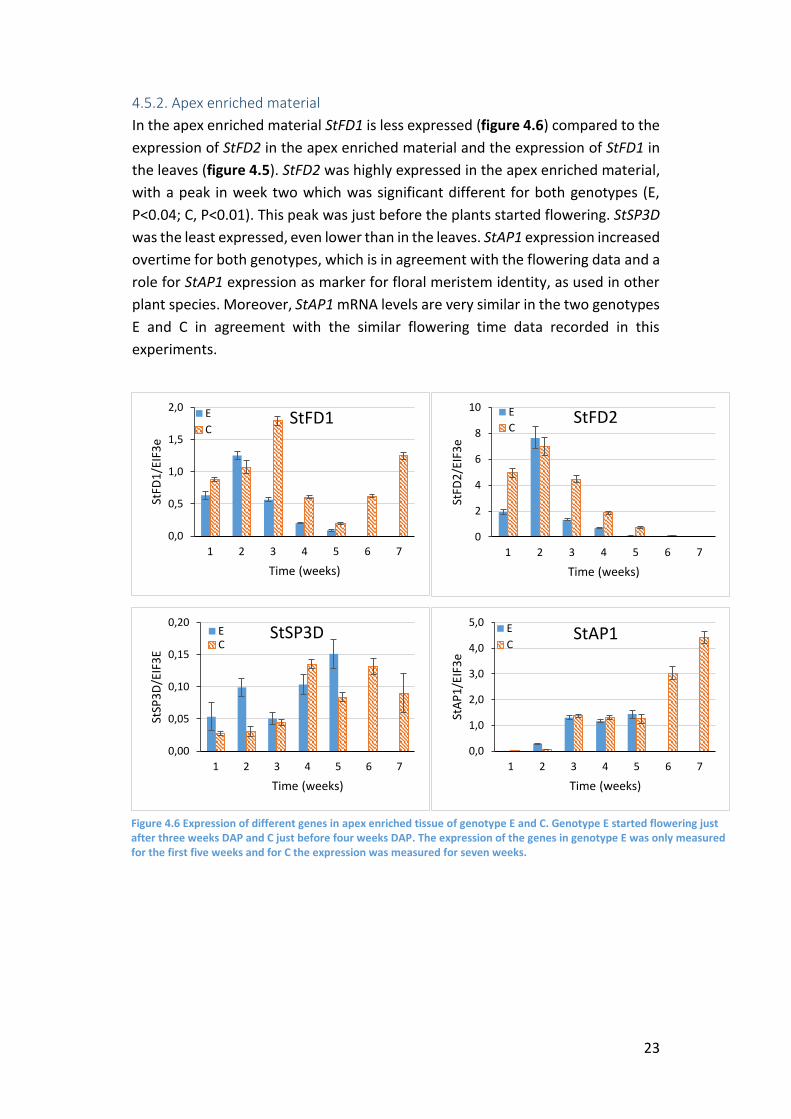
In the apex enriched material StFD1 is less expressed (figure 4.6) compared to the
expression of StFD2 in the apex enriched material and the expression of StFD1 in
the leaves (figure 4.5). StFD2 was highly expressed in the apex enriched material,
with a peak in week two which was significant different for both genotypes (E,
P<0.04; C, P<0.01). This peak was just before the plants started flowering. StSP3D
was the least expressed, even lower than in the leaves. StAP1 expression increased
overtime for both genotypes, which is in agreement with the flowering data and a
role for StAP1 expression as marker for floral meristem identity, as used in other
plant species. Moreover, StAP1 mRNA levels are very similar in the two genotypes
E and C in agreement with the similar flowering time data recorded in this
experiments.
0,0
0,5
1,0
1,5
2,0
1 2 3 4 5 6 7
StFD
1/E
IF3
e
Time (weeks)
StFD1E
C
0
2
4
6
8
10
1 2 3 4 5 6 7
StFD
2/E
IF3
e
Time (weeks)
StFD2E
C
0,0
1,0
2,0
3,0
4,0
5,0
1 2 3 4 5 6 7
StA
P1
/EIF
3e
Time (weeks)
StAP1E
C
0,00
0,05
0,10
0,15
0,20
1 2 3 4 5 6 7
StSP
3D
/EIF
3E
Time (weeks)
StSP3DEC
Figure 4.6 Expression of different genes in apex enriched tissue of genotype E and C. Genotype E started flowering just after three weeks DAP and C just before four weeks DAP. The expression of the genes in genotype E was only measured for the first five weeks and for C the expression was measured for seven weeks.

24
4.5.3. Stolon
StFD1, StFD2 and StSP3D are all less expressed in the stolon (figure 4.7). StFD1
expression decreased over time, while the expression of StFD2 fluctuated over
time. StSP3D had a peak in week two for genotype C and for the rest of the weeks
the expression was low.
We conclude that StFD1 is highest expressed in leaves and stolon. The expression
of StSP3D is low in the leaves. Which was expected as it is known that that LD
conditions have a negative effect on this expression (Navarro et al., 2011). StFD2
and StAP1 are mainly expressed in the apex enriched material and the expression
of StAP1 increases after the peak of StFD2.
0,0
0,5
1,0
1,5
2,0
2,5
2 3 4 5 6 7
StFD
1/E
IF3
e
Time (weeks)
StFD1EC
0,0
1,0
2,0
3,0
4,0
5,0
2 3 4 5 6 7
StFD
2/E
IF3
e
Time (weeks)
StFD2EC
0,00
0,05
0,10
0,15
0,20
0,25
0,30
2 3 4 5 6 7
StSP
3D
/EIF
3e
Time (weeks)
StSP3DEC
Figure 4.7 Expression of different genes in stolon tissue of genotype E and C. The expression of the genes in genotype E was only measured for the first five weeks and for C the expression was measured for seven weeks. The vertical blue line represents the time that genotype E got stolons, the vertical red line represents when genotype C got stolons and the vertical green line represents when the stolons from both genotypes were starting to become tubers.

25
4.6. Yeast-two hybrid
To gather information about the function of the StFDs several yeast-two hybrids
were performed to find an interaction with StSP3D. The yeast-two hybrids were
two times performed. The first time only an interaction was found between StFD1
and StSP3D from S. andigena. However, the colonies took a week to grow. The
second time was done after the research thesis by Bergonzi & Gol (unpublished)
and an interaction was found between StFD1 and StSP3D, but also between StFD2
and StSP3D (figure 4.8). We concluded that StSP3D can interact with both of the
StFDs, which indicates that they might be involved in the initiation of flowering.
Figure 4.8The culture growth from the yeast-two hybrid (Bergonzi & Gol, unpublished). Growth on SD -3aa, means that an interaction between the two proteins is possible. Therefore, StFD1 can interact with StSP3D and StFD2 can also interact with StSP3D. All the potato genes are coming from S. andigena.
SD -2aa (–H-A)
SD -3aa (-H) 4 mM 3AT
102
102
10 102
102
10

26
5. Discussion
5.1. Sequence comparison
Comparing the different FD genes, the exon with the bZIP motif was generally well
conserved. This was expected as this part can bind specifically to the short
palindromic or pseudo palindromic target DNA sequence and as a result is
required for the functioning of the FD as transcription factor (Hakoshima, 2005).
The same is true for the SAP exon, which is required for the binding to 14-3-3, as
it was shown in different plant species including rice (Taoka et al., 2011). The first
exon of FD is not that well conserved as it varies between different families. There
are two families for which this difference stands out Solanaceae and Brassicaceae.
The reason why these families stood out is because more sequences of the
different FD genes from these families are available. Nevertheless, the gene in
general is well conserved, which indicates that it has an important function in the
angiosperms.
StFD2 is more similar to a SPGB gene from tomato than to StFD1, which might
indicate that the function of the protein is similar. In tomato SPGB interacts with
SP3D and triggers flowering (Cao et al., 2016; Pneuli et al., 2001). So, it is likely
that StFD2 is involved initiating flowering in potato.
5.2. Phenotyping and expression analyses
In order to find out whether the StFD genes were involved in flowering, the growth
and development of the plants were measured and observed. A difference was
observed in flowering times between genotype C and E, but the difference was
less than what was to be expected based on previous researches. This could be
due to the growing condition used in the earlier and described experiments,
glasshouse and climatic chamber respectively. Additionally, Celis-Gamboa et al.,
(2003) found that genotype C and E flowered and formed stolons within a
narrower than anticipated time period, relative to each other also when
genotypes were grown in the open field. Indicating that the observed flowering
time difference between genotype C and E in this research is possible.
Genotype CE3130 was the only genotype that showed flower bud abortion and
flower abortion. Kooman (1995) found that the rate at which the complete shift
of assimilate partitioning from shoot to tuber occurs, varies between cultivars and
depends on temperature and photoperiod. Meaning that the assimilates were
redirected towards tubers rather than the flowers. CE3130 is strongly induced to
tuberisation due to the condormation of the StCDF1 alleles and promotion of
StSP6A expression (kloosterman et al 2013). Flowering and tuberisation are in
competition for assimilates and the signals for initiating both are similar
(Almekinders & Struik, 1996).

27
5.3. Gene expression during development
In genotype C and E StFD2 was highest expressed in the apex enriched material,
with a peak expression one week before flowering was observed. This indicated
that it is likely that StFD2 is involved in flowering, which is in agreement with the
found QTL for the unset of flowering where StFD2 is in located (Hurtado-Lopez,
2012). After the peak expression, the amount expressed degrades over time.
There is a difference in the degradation of StFD2 between the genotypes. In E the
increase of StFD2 occurs quickly and it degrades even quicker. In genotype C the
change in the expressions from the week before and after the peak was less than
in genotype E. A possible explanation for this is that there is a slight difference in
flowering time between the plants from genotype C of one day, while in genotype
E there was no gap. The rapid reduction of FD has also been noticed in Arabidopsis
(Seedat et al., 2013). It could be that FD is only needed to start up flowering and
thereafter it no longer needed and expression reduces.
From our results we can’t conclude if StFD2 is regulated by the circadian clock, due
to that the relative expression levels are low. The results from the later
experiments show that the samples we took are inappropriate.
The highest expression of StSP3D was found in the leaves. This was expected as
StSP3D is produced in these (Navarro et al., 2011). It is similar to tomato where all
four FT-like genes (including SFT which is the homologue of StSP3D) are expressed
exclusively in leaf tissue (Cao et al., 2016). However, the expression levels were
low, likely because the plants were growing under LD conditions and StSP3D is
mainly produced during SD conditions (Navarro et al., 2011). Despite the low
levels, genotype E showed higher expression of StSP3D than genotype C. This
could be due to differences in StCDF1 alleles. StCDF1 indirectly downregulates the
production of StSP3D. Genotype E has alleles StCDF1.1 and StCDF1.2 and genotype
C has alleles StCDF1.1 and StCDF1.3. StCDF1.2 has an insertion resulting in a frame-
shift introducing a premature stop codon, which causes earlier flowering and
tuberisation (Kloosterman et al., 2013).
The expression of StAP1 mRNA was tested as a possible marker for the floral
transition. In Arabidopsis is AP1 produced in the apex, where it specifically marks
the presence of flower meristems (Wigge et al., 2005). In different plant species it
is shown that FD and FT initiate this expression (Abe et al., 2005; Taoka et al., 2011;
Wigge et al., 2005). We found that StAP1 is mainly expressed in the apex enriched
material and the expression starts after StFD2 is produced. The expression of
StAP1 kept increasing over the measuring time. This suggests that the StAP1
expression is conserved in potato, just like in other plant species (Abe et al., 2005;
Parmentier-Line & Coleman, 2015; Wigge et al., 2005).

28
We found that StFD1 is controlled by the circadian clock, where it decreases during
the day time and increases during the night time. Also was found that StFD1 was
highest expressed in the leaves and stolon. In the leaves of genotype C was a peak
expression a week after floral buds were observed. In tomato there is a FD
homolog found (SPGBi) which is mainly expressed in the leaves. It is thought that
the high FD expression in the leaves makes it unnecessary for SFT RNA to travel
towards its interacting partners as implied for Arabidopsis (Lifschitz et al., 2006).
This could also be the function of StFD1 in potato, accounting for the higher
expression in the leaves for genotype C.
The transition of stolon to tuber is a highly regulated process, involving
components from the photoperiod-dependent transition which are also shared
with the flowering regulatory pathway (Rodríguez-Falcón et al., 2006). However,
at the time of this research it had not been shown that FD is involved in
tuberisation. Later, Teo et al. (2017) published that StFD1 is involved in the
regulation of tuber formation together with StSP6A. Which could explain the high
expression in the stolon during the first sampling time. Also, a week after the high
expression the first stolons were observed.
5.4. Yeast-two hybrid
In this research an interaction was found between StFD1 and StSP3D, and StFD2
and StSP3D (Bergonzi & Gol, unpublished). The FT-FD complex interaction has
already been reported in other plant species including: rice, tomato, Arabidopsis,
wheat, pea and kiwi (Li & Dubcovsky, 2008; Lifschitz et al., 2006; Pnueli et al., 2001;
Sussmilch et al., 2015; Tsuji et al., 2013; Varkonyi-Grasic et al., 2013; Wigge et al.,
2005). This would suggest that it is a general mechanism for FT-mediated gene
activation in the shoot apical meristem in all higher plants.

29
6. Conclusion
The aim of this research was to investigate the role of StFD1 and StFD2 in potato
flowering, this was done by splitting the research into two different sections: 1)
phenotyping plant growth and correlating the phenotypes with gene expression
analysis, 2) testing the interaction between the different StFDs and StSP3D with a
yeast-two hybrid. We found that the expression of StFD2 is primarily restricted to
the apex, with a peak expression one week before flowering. After this peak, the
expression levels degraded over time. We also found that StFD2 can interact with
StSP3D and form a FAC. Overall, it is most likely that StFD2 is the canonical FD in
potato that sits in the meristem and interacts with StSP3D. Together they are most
likely involved in flower initiation.
We found that StFD1 is regulated by the circadian clock. StFD1 is mainly
transcribed in the leaves and can interact with StSP3D. It is unclear why StFD1 is
expressed in the leaves in a diurnally regulated manner. However, since FD
proteins are unknown to be transported, it is unlikely that this expression is linked
with flower transition. A peak expression was found in the stolon just a few weeks
before tuberisation was observed.
However, the functional characterisation, through generation of mutants and
transgenic lines is needed to elucidate the role of StFD1 and StFD2 in potato.

30
7. Recommendation
More research is needed to find the cause of flower abortion. It can be due to
genetic differences, to abiotic, to biotic influences, or to a combination of these.
In this research, we observed that the genotype with high tuberisation aborted all
the flowers. Indicating that it could be a problem in resource allocation and that
the plant allots more energy to tuberisation than to flowering, resulting in flower
abortion.
It is unclear whether StFD2 is regulated by the circadian clock, as samples that
were used were coming from whole vegetative plants. StFD2 is mainly produced
just before flowering and is in low levels expression in the leaves. Therefore, a new
24-hour time course of apex enriched material with plants that are almost
flowering could give more insight in whether StFD2 is dependent of the circadian
clock.
Only one S. andigena allele of StFD1 and one allele from StFD2 were used in the
yeast-two hybrid, however, there are more alleles from StFD1 and StFD2.
Particularly C and E present interesting allelic variation in StFD2 which is
hypothesis to underline the QTL for onset of flowering mapped in the CxE
population (Hurtado et al., 2012). To give a better view on the protein-protein
interaction, these different alleles have to be tested whether they can interact in
a yeast-two hybrid with StSP3D.
It is unknown whether FD can interact with TFL1, as different researches proven
opposite results (Abe et al., 2005; Wigge et al., 2005) but generally TFL1
homologues act as floral repressors. The interaction between StFD and StTFL1
might be interesting with regard to flowering in potato, as it could play a role in
repression of flowering and flower abortion.
In this research two protein-protein interactions were found between StFD1 and
StSP3D, and StFD2 and StSP3D. To prove that this is correct and possible it should
also be proven with the other systems pull down and BiFC. To finalise the
functional role of the two StFDs involves a transgenic approach. It is the only way
to understand the biological function of a given gene. All the previous results are
only correlation suggesting relations. Plants have to be down regulated in either
StFD1, StFD2 and both to understand existence of redundant roles, in order to find
the different effects between different StFD expressions. It is expected that the
results will show that StFD2 is involved in flowering, because it expresses highest
in the apex enriched material and it can form a FAC with StSP3D.

31
References
Abe M., Kobayashe Y., Yamamoto S., Daimon Y., Yamaguchi A., Ikeda Y., Ichinoki
H., Notaguchi M., Goto K. & Araki T. (2005) FD, a bZIP Protein Mediating Signals
form the Floral Pathway Integrator FT at the Shoot Apex. Science Vol. 309 No. 5737
Pp. 1052-1056.
Abelenda J.A., Navarro C. & Prat S. (2014) Flowring and tuberization: a tale of two
nightshades. Trends in Plant Science Vol. 19, No. 2, Pp. 115-122.
Almekinders C.J.M. & Struik P.C. (1996) Shoot development and flowering in
potato (Solanum tubersorum L.). Potato Research Vol. 39 No. 4 Pp. 581-607.
Altschul S.F., Madden T.L., Schäffer A.A., Zhang J., Zhang Z., Miller W. & Lipman
D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database
search progreams. Nucleic acids research Vol. 25 No. 17 Pp. 3389-3402.
Andrés F. & Coupland G. (2013) The genetic basis of flowering responses to
seasonal cues. Nature revieuws Genetics Vol. 13 Pp. 627-639.
Augustin J., Johnson S.R., Teitzel C., True R.H., Hogan J.M., Toma R.B., Shaw R.L. &
Deutsch R.M. (1978) Changes in nutrient composition of potatoes during home
preparation: II. Vitamins. American Potato Journal Vol. 55 Pp. 653-662.
Banks J.A., Nishiyama T., Hasebe M., Bowman J.L., Gribskov M., dePamphilis C.,
Albert V.A., Aono N., Aoyama T., Ambrose B.A., Ashton N.W., Axtell M.J., Barker
E., Barker M.S., Bennetzen J.L., Bonawithz N.D., Chapple C., Cheng C., Correa
L.G.G., Dacre M., DeBarry J., Dreyer I., Elias M. Engstrom E.M., Estelle M., Feng L.,
Finet C., Floyd S.K., Frommer W.B., Fujita T., Gramzow L., Gutensohn M., Harholt
J., Hattori M., Heyl A., Hirai T., Hiwatashi Y., Ishikawa M., Iwata M., Karol K.G.,
Koehler B., Kolukisaoglu U., Kubo M., Kurata T., Lalonde S., Li K., Li K., Li Y., Litt A.,
Lyons E., Manning G., Maruyama T., Michael T.P., Mikami K., Miyazaki S., Morinaga
S., Murat T., Mueller-Roeber B., Nelson D.R., Obara M., Oguri Y., Olmstead R.G.,
Onodera N., Petersen B.L., Pils B., Prigee M., Rensing S.A., Riaño-Pachón D.M.,
Robers A.W., Sato Y., Scheller H.V., Schulz B., Schulz C., Shakirov E., Shibagaki N.,
Shinohara N., Shippen D.E., Sørensen I., Sotooka R., Sugimoto N., Sugita M.,
Sumikawa N., Tanurdzic M., Theißen G., Ulvskov P., Wakazuki S., Weng J.K., Willats
W.W.G.T., Wipf D., Wolf P.G., Yang L., Zimmer A.D., Zhu Q., Mitros T., Hellsten U.,
Loqué D., Otillar R., Salamov A., Schmutz J., Shapiro H., Lindquist E., Lucas S.,
Rokhsar D. &Grigoriev I. (2011) The compact Selaginella genome identifies
changes in gene content associated with the evolution of vascular plants. Science
Vol. 332 No. 6032 Pp. 960-963.
Bergonzi S. & Gol D. (unpublished) Yeast-two hybrid results.
Bourke P. (2014) Tuber inhibition under abiotic stress. Master Thesis University
Wageningen.
Cao K., Cui L., Zhou X., Ye L., Zou Z. & Deng S. (2016) Four Tomato FLOWERING
LOCUS T-Like Proteins Axt Antagonistically to Regulate Floral Initiation. Frontiers
in Plant Science Pp. 6.

32
Celis-Gamboa C., Struik P.C., Jacobsen E. & Visser R.G.F. (2003) Temporal dynamics
of tubers formation and related processes in a crossing population of potato
(Solanum tuberosum). Ann. appl. Biol. Vol. 143 Pp. 175-186.
Chailakhyan M.K. (1936) New facts in support of the hormonal theory of plant
development. C. R. Acad. Sci. URSS No. 13 Pp. 79-83.
Desborough S.L. & Weiser C.J. (1974) Improving potato protein. I. Evaluation of
selection techniques. American Potato Journal Vol. 51 Pp. 185-196.
Ewing E.E. & Struik P.C. (1992) Tuber formation in potato: induction, initiation and
growth. Horicultural Reviews Vol. 14 Pp. 89-197.
González-Schain N.D., Díaz-Mendoza M., Zurczak M. & Suárez-López P. (2012)
Potato CONSTANS is involved in photoperiodic tuberization in a graft-transmissible
manner. Plant Journal Vol. 70 PP. 678-690.
Grun P. (1979) Evolution of the cultivated potato: a cutoplasmic analysis. Hawkes
J.G., Lester R.N., Skelding A.D. ed(s). The biology and taxonomy of the Solanaceae.
Pp. 655-665.
Grun P. (1990) The evolution of the cultivated potatoes. Economic Botany Vol. 44
No. 3 Pp.39–55.
Hakoshima, T. (2005). Leucine Zippers. Encyclopedia of Life Science Pp. 1-5.
Hawkes J.G. & Francisco-Ortega J. (1993) The early history of the potato in Europe.
Euphytica Vol. 70 Pp. 1-7.
Hijmans R.J. & Spooner D.M. (2001) Geographic distribution of wild potato
species. American Journal of Botany Vol. 88 No. 11 Pp. 2101-2112.
Hirsch C.N., Hirsch C.D., Felcher K., Coombs J., Zarka D., Deynze van A., Jong De
W., Veilleux R.E., Jansky S., Bethke P., Douches D.S. & Buell C.R. (2013)
Retrospective View of North American Potato (Solanum tuberosum L.) Breeding in
the 20th and 21st Centuries. G3: Genes| Genomes| Genetics Vol. 3 No. 6 Pp. 1003-
1013.
Hosaka K. & Hanneman R.E. (1998a) Genetics of self-compatibility in a self-
incompatible wild diploid potato species Solanum chacoense. 1. Detection of an S
locus inhibitor (Sli) gene. Euphytica Vol. 99 Pp.191–197.
Hosaka K. & Hanneman R.E. (1998b) Genetics of self-compatibility in a self-
incompatible wild diploid potato species Solanum chacoense. 2. Localization of an
S locus inhibitor (Sli) gene on the potato genome using DNA markers. Eutphytica
Vol. 103 Pp. 265–271.
Huamán Z. & Spooner D.M. (2002) Reclassification of landrace populations of
cultivated potatoes (Solanum Sect. Petota). American Journal of Botany Vol. 89 Pp.
947-965.

33
Hurtado P.X., Schnabel S.K., Zaban A., Veteläinen M., Virtanen E., Eilers P.H., van
Eeuwijk F.A., Visser R.G.F. & Maliepaard C. (2012) Dynamics of senescence-related
QTLs in potato. Euphytica Vol. 183 No. 3 Pp. 289-302.
Hurtado-Lopez P.Z. (2012) Investigating genotype by environment and QTL by
environment interactions for developmental traits in potato.Imaizumi T., Schultz
T.F., Harmon F.G., Ho L.A. & Kay S.A. (2005) FKF1 F-box protein mediates cyclic
degradation of a repressor of CONSTANS in Arabidopsis. Science Vol. 309 No. 5732
Pp. 293-297.
Jacobs J.M.E., Van Eck H.J., Arens P., Verkerk-Bakker B., te Lintel Hekkert B.,
Bastiaanssen H.J.M., El-Kharbotly A., Pereira A., Jacobsen E. & Stiekema W.J.
(1995) A genetic map of potato (Solanum tuberosum) integrating molecular
markers, inclurding transposons, and classical markers. Theoretical and Applied
Genetics Vol. 91 Pp. 289-300.
Kloosterman B., Vorst O., Hall R.D., Vissers R.G.F. & Bachem C.W. (2005) Tuber on
a chip: differential gene expression during potato tuber development. Plant
Biotechnology Jounal Vol. 3 Pp. 505-519.
Kloosterman B., Abelenda J.A., Gomez M.M.C., Oortwijn M., de Boer J.M.,
Kowitwanich K., Horvath B.M., van Eck H.J., Smaczniak C., Prat S., Visser G.F. &
Bachem C.W.B. (2013) Naturally occurring allele diversity allows potato cultivation
in northern latitudes. Nature Vol. 495 Pp. 246-252.
Kooman P. (1995) Yielding ability of potato crops as influenced by temperature
and daylength. Proefschrift Wagenigen University.
Levy D. & Veilleux E. (2007) Adaptation to High Temperatures and Salinity – A
Review. Amer J of Potato Res Vol. 84 Pp. 487-506.
Li C. & Dubcovsky J. (2008) Wheat FT protein regulates VRN1 transcription through
interactions with FDL2. The Plant Journal Vol. 55 Pp. 543-554.
Lifschitz E., Eviatar T., Rozman A., Shalit A., Goldshimdt A., Amsellem Z., Alvarez
J.P. & Eshed Y. (2006) The tomato FT ortholog triggers systemic signals that
regulate growth and flowering and substitute for diverse environmental stimuli.
PNAS Vol. 103 No. 16 Pp. 6398-6403.
Liljegren S.J., Gustafson-Brown C., Pinyopich A., Ditta G.S. & Yanofsky M.F. (1999)
Interactions among APETALA1, LEAFY, and TERMINAL FLOWER1 Specify Meristem
Fate. The Plant Cell Vol. 11 Pp. 1007-1018.
Lindhout P., Meijer D., Schotte T., Hutten R.C.B., Visser R.G.F. & Eck van H.J. (2011)
Towards F1 Hybrid seed Potato Breeding. Potato Research Vol. 54 Pp. 301-312.
Navarro C., Abelenda J.A., Cruz-Oró E., Cuéllar C.A., Tamaki S., Silva J., Shimamoto
K. & Prat S. (2011) Control of flowering and storage organ formation in potato by
FLOWEING LOCUS T. Nature Vol. 478, Pp. 119-123.
Negi P. & Nath N. (2002) Effect of partial dehydration on quality of canned
potatoes. European Food Research and Technology Vol. 215 No. 3 Pp. 231-234.

34
Parmentier-Line C.M. & Coleman G.D. (2015) Constitutive expression of the Poplar
FD-like basic leucine zipper transcription factor alters growth and bud
development. Plant Biotechnology Journal Pp. 1-11.
Pfaffl M.W. (2001) A new mathematical model for relative quantification in real-
time RT-PCR. Nucleic acids research Vol. 29 No. 9 Pp. e45-e45.
Pnueli L., Gutfinger T., Hareven D., Ben-Naim O., Ron N., Adir N. & Lifschitz E.
(2001) Tomato SP-Interacting Proteins Define a Conserved Signaling System That
Regulates Shoot Architecture and Flowering. The Plant Cell Vol. 13 Pp. 2687-2702.
Rensing S.A., Lang D., Zimmer A.D., Terry A., Salamov A., Shapiro H., Nishiyama T.,
Perroud P.F., Lindquist E.A., Kamisugi Y, Tanahashi T., Sakakibara K., Fujita T., Oishi
K., Shin-I T., Kuroki Y., Toyoda A., Suzuki Y., Hashimoto S., Yamaguchi K., Sugano
S., Kohara Y., Fujiyama A., Anterola A., Aoki S., Ashton N., Barbazuk B., Barker E.,
Bennetzen J.L., Blankenship R., Cho S.H., Dutcher S.K., Estelle M., Fawcett J.A.,
Gundlach H., Hanada K., Heyl A., Hicks K.A., Hughes J., Lohr M., Mayer K.,
Melkozernov A., Murata T., Nelson D.R., Pils B., Prigge M., Reiss B., Renner T.,
Rombauts S., Rushton P.J., Sanderfoot A., Schween G., Shiu S.H., Stueber K.,
Theodoulou F.L., Tu H., Van de Peer Y., Verrier P.J., Waters E., Wood A., Yang L.,
Cove D., Cuming A.C., Hasebe M., Lucas S., Mishler B.D., Reski R., Grigoriev I.V. &
Quatrano R.S. Boore J.L. (2008) The Physcomitrella Genome Reveals Evolutionary
Insights into the Conquest of Land by Plants. Science Vol. 319 Pp. 64-69.
Rodríguez-Falcón M., Bou J. &Prat S. (2006) Seasonal Control of Tuberizion in
Potato: Covnseved Elements with the Flowering Response. The Annual Review
Plant Biology Vol. 57 Pp. 151-180.
Sawa M., Nusinow D.A., Kay S.A. & Imaizumi T. (2007) FKF1 and GIGANTEA
complex formation in required for day-length measurement in Arabidopsis.
Science Vol. 318 No. 5848 Pp. 261-265.
Seedat N., Dinsdale A., Ong E.K. & Gendall A.R. (2013) Acceleration of flowering in
Arabidopsis thaliana by Cape Verde Islands alleles of FLOWERING H is dependent
on the floral promoter FD. Journal of Experimental Botany Vol. 64 No. 10 Pp. 2767-
2778.
Spooner D.M., McLean K., Ramsay G., Waugh R. & Bryan G.J. (2005) A single
domestication for potato based on multilocus amplified fragment length
polymorphism genotyping. PNAS Vol. 102 No. 41 Pp. 14694-14699.
Sussmilch F.C., Berbel A., Hecht V., Vander Schoor J.K., Ferrándiz C., Madueño F &
Weller J.L. (2015) Pea VEGETATIVE2 Is an FD Homolog That Is Essential for
Flowering and Compound Inflorescence Development. The Plant Cell Vol. 27 Pp.
1046-1060.
Taoka K.I., Ohki I., Tsuji H., Furuita K., Hayashi K., Yanase T., Yamaguchi M.,
Nakashima C., Purwestri Y.A., Tamaki S., Ogaki Y., Shimada C., Nakagawa A., Kojima
C. & Shimamoto K. (2011) 14-3-3 proteins act as intracellular receptors for rice
Hd3a florigen. NATURE Vol. 476 Pp. 332-338.

35
Teo C.J., Takahashi K., Shimizu K., Shimamoto K. & Taoka K. (2017) Potato Tuber
Induction in Regulated by Interactions Between Components of a Tuberigen
Complex. Plant Cell Physiology Vol. 58 No. 2 Pp. 365-374.
Tsuji H., Nakamura H., Taoka K. & Shimamoto K. (2013) Functional Diversification
of FD Transcription Factors in Rice, Components of Florigen Activation Complexes.
Plant cell Physiol. Vol. 54 No. 3 Pp. 385-397.
Turck F., Fornara F. & Coupland G. (2008) Regulation and identity of Florigen:
FLOWERING LOCUS T Moves Center Stage. Annual Review Plant Biology Vol. 59 Pp.
573-594.
Tylewicz S., Tsuji H., Miskolczi P., Petterle A., Azeez A., Jonsson K., Shimamoto K.
& Bhalerao R.P. (2015) Dual role of tree florigen activation complex component
FD in photperiodic growth control and adaptive response pathway. PNAS Vol . 112
No. 10 Pp. 3140-3145.
Valkonen J.P (2007) Viruses: economical losses and biotechnological potential.
Potato biology and biotechnology Pp. 619-641.
Varkonyi-Gasic E., Moss S.M.A., Voogd C., Wang T., Putterill J. & Hellens R.P. (2013)
Homologs of FT, CEN and FD respond to developmental and environmental signals
affecting growth and flowering in the perennial vine kiwifruit. New Phytologist Vol.
198 Pp. 732-746.
Wigge P.A., Kim M.C., Jaeger K.E., Busch W., Schmid M., Lohmann J.U. & Weigel D.
(2005) Integration of Spatial and Temporal Information During Floral Induction in
Arabidopsis. Science Vol. 309 Pp. 1056-1059.

36
Appendix 1: Primers Table App 1.1 Primers that were used, the usage of the primers are all for potato
Primer Sequence
FD1 forward for qPCR and cloning CACCATGTGGTCATCAAGTAATGAAG
FD1 reverse for cloning TCAAAATGGGGCCGTTGATG
FD1 reverse for qPCR TGGGATGATTATAGCTACAAGATGT
FD2 forward DM for cloning CACCATGTGGTCATCAAGCAGGTC
FD2 reverse for cloning TCAAAATGGAGCGGTTGACG
FD2 forward C1 for cloning CACCATCTGGTCATCAAGCAGGTC
FD2 forward C2 for cloning CACCATGGAAGAAGTGTGGAAAG
FD2 forward C3 for sequencing CATTCCCAACTCTTTGTTCTC
FD2 forward E cloning CACCATGTGGTCATCAAGCAGTGAC
FD2 negative control reverse cloning TCAATGAAGAGAGTTCTTTTTTG
FD2 forward for qPCR TTCCCCATTTTCTCCAAGAC
FD2 reverse for qPCR TCCACCAAAATTAGCACCTTG
StSP3D forward for qPCR GGACCCAGATGCTCCAAGTC
stSP3D reverse for qPCR CTTGCCAAAACTTGAACCTG
TFL1 forward for cloning CACCATGGCTTCCAAAATGTGTGAAC
TFL1 reverse for cloning TCAACGCCTTCTAGCGGCAG
AP1 forward for qPCR GGCACCAGCAAACTAATCAAG
AP1 reverse for qPCR CTCCTTGCTTCTGCTGCTTC
ELF3e (349C1) forward for housekeeping gene
GGAGCACAGGAGAAGATGAAGGAG
ELF3e (349C1) reverse for housekeeping gene
CGTTGGTGAATGCGGCAGTAGG
NAC forward for housekeeping gene ATATAGAGCTGGTGATGACT
NAC reverse for housekeeping gene TCCATGATAGCAGAGACTA *Since for S.andigena we have no genome information, primers were designed on the reference genome DM.

37
Appendix 2: Sequences
The sequences highlighted in yellow are the introns of the gene sequences, the
not highlighted parts are the exons.
>StFD1_Genome.DNA.allele_DM
ATGTGGTCATCAAGTAATGAAGAACATCAAGGGTTAAACAAAAACCTTGCAACTAACTC
TTCAAAATCATCATCATCATCATCATCTACATCTTGTAGCTATAATCATCCCATTAATCCT
ATACTAATAAATAGTAAAACCATGGAAGAAGTATGGAAAGACATAAATCTTTCATGTTC
TAGAGATCCACAAAATACAACTGCTGGTGGTATAATTTTGCAAGATTTATTGGCAAGAC
CATTTGCTAATAACCCTTCAACAGCAGCAGCTTATGGTTCCCCTGTTCCTCCCCTGCCTCC
TCCCCCTGTTACCATGTTGACATTGAACTCTGGCCCTGAATTTCATTTCTTTAGCACCTTA
AACCCAATCAGACAAACTCAACAAAAAAACATTTCTAATGAGGGATTAGCTTCACCAGC
TCCTGGAAGAAAAAGACATCCTGAATCAAATAACAACAATTCTGGTGACCAGAAAAGC
AAGAGGATGATAAAGAACCGTGAGTCCGCTGCTCGATCCCGGGCTCGAAAGCAGGTA
AGTTTTTATTCTCTTCTTATAATAAAATGCATCATTGCCTGATTGAACAGTCACCTGTACA
TCATAATCTTACTCCTTATAAATGGAGTGACATATTATAGTTGACAAGTTCAAACTTGTTT
GAGACTGAAACGTAGTAATCGAGACTCTTTTGGTCAGTTTTGCTGATTTTAGTAGTGAT
ATGTTTTCATATAACTTGCAGGCTTACACGAATGAATTAGAGATGGAAGTAGCCAATTT
GATGGAAGAAAATGCCAGGCTCAAGAGGCAGCAGCAACAGGTTACTTTTTATGGTTTA
GCTAAATCTTTTGGAGTGTAAAAAACTCATCTTTTTTCTCATACATCTGTGTCCTTTTTCAT
GTGGTGGTCAGTTATGCTTAGCTTCAGCTGGTGCTCTACTTCCAAAAATGAAGTCACTC
AATAGAACATCAACGGCCCCATTTTGA
>StFD1_Genome.DNA.Allele_E
ATGTGGTCATCAAGTAATGAAGAACATCAAGGGTTAAACAAAAACCTTGCAACTAACTC
TTCAAAATCATCATCATCATCATCATCTACATCTTGTAGCTATAATCATCCCATTAATCCT
ATGCTAATAAATAGTAAAACCATGGAACAAGTATGGAAAGACATAAATCTTTCATGTTC
TAGAGATCCACAAAATACAACTGCTGGTGGTATAATTTTGCAAGATTTATTGGCAAGAC
CATTTGCTAATAACCCTTCAACAGCAGCAGCAGCAGCTTATGGTTCCCCTGTTCCTCCCT
TGCCTCCTCCCCCTGTTACCATGTTGACATTGAACTCTGGCCCTGAATTTCATTTCTTTAG
CACCACCTTAAACCCAATCAGACAAACTCAACAAAAAAACATTTCTATTGAGGGATTAG
CTTCACCAGCTCCAAGAAGAAAAAGACATCCTGAATCAAATAACAACAATTCTGGTGAC
CAGAAAAGCAAGAGGATGATCAAGAACCGTGAGTCCGCTGCTCGATCCCGGGCTCGAA
AGCAGGTAAGTTTTTATTCTCTTCTTATAATAAAATGCATCATTGCCTGATTGAACAGTC
ACCTGTACATCATAATCTTACTCCTTATAAATAGAGTGACATATTATAGTTGACAAGTTC
AAACTTGTTTGAGACTGAAACGTAGTAATCGAGACTCTTTTGATCAGTTTTGCTGATTTT
AGTAGTGATATGTTTTCATATAACTTGCAGGCTTACACGAATGAATTGGAGATGGAAGT
AGCCAATTTGATGGAAGAAAATGCCAGGCTCAAGAaGCAGCAGCAACAGGTTACTTTT
TATGGTTTAGCTAAATCTGTTGGAGTGTAAAAAACTCATCTTTTTTCTCATTCATCTGTGC
CCTTTTTCATGTGGTGGTCAGTTATGCTTAGCTTCAGCTGGTGCTCAACTTCCAAAAATG
AAGTCATCACTCAATAGAACATCAACGGCCCCATTTTGA

38
>StFD1_Genome.DNA.allele_C
ATGTGGTCATCAAGTAATGAAGAACATCAAGGGTTAAACAAAAACCTTGCAACTAACTC
TTCAAAATCATCATCATCATCATCATCTACATCTTGTAGCTATAATCATCCCATTAATCCT
ATGCTAATAAATAGTAAAACCATGGAAGAAGTATGGAAAGACATAAATCTTTCATGTTC
TAGAGATCCACAAAATACAACTGCTGGTGGTATAATTTTGCAAGATTTATTGGCAAGAC
CATTTGCTAATAACCCTTCAACAGCAGCAGCTTATGGTTCCCCTGTTCCTCCCCTGCCTCC
TCCCCCTGTTACCATGTTGACATTGAACTCTGGCCCTGAATTTCATTTCTTTAGCACCTTA
AACCCAATCAGACAAACTCAACAAAAAAACATTTCTAATGAGGGATTAGCTTCACCAGC
TCCAAGAAGAAAAAGACATCCTGAATCAAATAACAACAATTCTGGTGACCAGAAAAGC
AAGAGGATGATCAAGAACCGTGAGTCCGCTGCTCGATCCCGGGCTCGAAAGCAGGTAA
GTTTTTATTCTCTTCTTATAATAAAATGCATCATTGCCTGATTGAACAGTCACCTGTACAT
CATAATCTTACTCCTTATAAATGGAGTGACATATTATAGTTGACAAGTTCAAACTTGTTT
GAGACTGAAACGTAGTAATCGAGACTCTTTTGGTCAGTTTTGCTGATTTTAGTAGTGAT
ATGTTTTCATATAACTTGCAGGCTTACACGAATGAATTGGAGATGGAAGTAGCCAATTT
GATGGAAGAAAATGCCAGGCTCAAGAAGCAGCAGCAACAGGTTACTTTTTATGGTTTA
GCTAAATCTGTTGGAGTGTAAAAAACTCATCTTTTTTCTCATTCATCTGTGCCCTTTTTCA
TGTGGTGGTCAGTTATGCTTAGCTTCAGCTGGTGCTCAACTTCCAAAAATGAAGTCACT
CAATAGAACATCAACGGCCCCATTTTGA
>FD2.1_Genomic.DNA.Allele_C
>ATC mutation in the start codon
AATAGTTCAACATAAAAAGAGTACATTCTCCTCGTTTTGTTCTCTATATACTGTATTTTTT
TTCAAGAATCCAATTATCTGGTCATCAAGCAGGTCTTCTTCATCTTCATCTCATTCCCCAT
TTTCTCCAAGACTCAAAACAATGGAAGAAGTGTGGAAAGATATTAATCTTTCTTCACTTC
AAGATCACACTACTAATTACTCGAAAGACCATCAACATCATCTTCATGATCATAATCATC
AAGGTGCTAATTTTGGTGGAATGATTTTACAAGATTTTTTGGCTAGGCCTTTTGCTAAG
GAATCTTCAGCAGCAGCAGCAGCCTCCTCTGTTTCAGCTACAACTATGCTGAATTTGAAC
TCTGTTCCAGAGCTTCATTTCTTTGATAACCCTTTAAGGCAAAACTCAATCTTGCACCAAC
CAAATGTTAGTGGAAGAAAAAGGGTTGTCCCTGAAACAGAGGACAATTCTACAGGGG
ACAGAAGAAATCAGAGGATGATCAAGAACAGAGAGTCTGCTTCTAGATCAAGAGCTAG
AAAGCAGGAAAGTGACACTCTCTTTATCTTAATCATCATTTGTTCTTTTCTGTTGTCCTGT
CGATTTTAAATTATGTTTCTTCATAAAAACACAACAGATAAACTATGTTGCTTCAATATGC
TGATTTTCTCTGTTTTCTTTTTTGGCCATTGATGATTTTTGCAGGCCTATTTGAATGAGTT
GGAGACGGAAGTGGCACATTTAGTTGAAGAAAATGCAAGGCTCAAGAAGCAGCAGCA
ACAGGTCCTTTTATCTTTTATTACACCATGTTTTAAATTCTGTTTATAACTAATGACTTTCT
TTAATTAGAACAAACAAATAAATGACTCAGAATTGATTGGTGTTTTTCTTAATTTGGACA
GTTACGCTTAGCTGCAGCTATTCAAGTTCCAAAAAAGAACTCTCTTCATCGGACGTCAAC
CGCTCCATTTTGA
>FD2.2_Genomic.DNA.Allele_E

39
AATAGTTCAACATAAAAAGAGTACATACCCCACTCTTTGTTCTCTATATATTTTTTTTGAA
GAATCCAATTATGTGGTCATCAAGCAGTGACAACAGGTCTTCTTCATCTTCATCTTCATC
TCATTCCCCATTTTCTCCAAGACTCAAAACAATGGAAGAAGTGTGGAAAGATATTAATC
TTTCTTCACTTCAAGATCACACTACTAATTACTCTAAAGACCATCAACATCATCTTCATGA
TCATAATCATCAAGGTGCTAATTTTGGTGGAATGATTTTACAAGATTTTTTGGCTAGGCC
TTTTGCTAATGAATCTTCAGCAGCAGCCTCCTCTGTTTCAGCTACAACTATGCTGAATTT
GAACTCTGTTCCAGAGCTTCATTTCTTTGATAACCCTTCAAGGCAAAACTCAATCTTGCA
CCAACCAAATGTTAGTGGAAGAAAAAGGGTTGTCCCTGAAACAGAGGACAATTCTACA
GGGGACAGAAGAAATCAGAGGATGATCAAGAACAGAGAGTCTGCTTCTAGATCAAGA
GCTAGAAAGCAGGAAAGTGACACTCTCTTTATCTTAATCATCATTTGTTCTAGTATAAAT
CTTTTCTGTTGTCCTGTCGATTTTTAGCAATAAATTATGTTTCTTTATAAAAACACAACAG
ATAAACTGTGTTGCTTCAATATGCTGATTTTCTCTGTTTTCTTTTTTGGCCATTGATGATTT
TTGCAGGCCTATTTGAATGAGTTGGAGACAGAAGTGGCACATTTAGTTGAAGAAAATG
CAAGGCTCAAGAAGCAGCAGCAACAGGTCCTATTATCTTTTATTACACCATGTTTTAAAT
TCTATTTATAACTAATGACTTTCTTTAATTAGAACAAACAAATAAATGACTCAGAATTGA
TTGGTGTTTTTCTTAATTTGGACAGTTACGCTTAGCTGCAGCTATTCAAGTTCCAAAAAA
GAACTCTCTTCATCGGACGTCAACCGCTCCATTTTGAGAAAA
>FD2.3_Genomic.DNA.Allele_C&E
AATAGTTCAACATAAAAAGAGTACATTCCCAACTCTTTGTTCTCTATATATTTTTTTTCAA
GAATCCAATTATGTGGTCATCAAGCAGGTCTTCTTCATCTTCATCTTCATCTCATTCCCCA
TTTTCTCCAAGACTCAAAACAATGGAAGAAGTGTGGAAAGATATTAATCTTTCTTCACTT
CAAGATCACACTACTAATTACTCTAAAGATCATCAACATCATCTTCATGATCATAATCAT
CAAGGTGCTAATTTTGGTGGAATGATTTTACAAGAATTTTTGGCTAGGCCTTTTGCTAAT
GAATCTTCACCAGCAGCAGCAGCCTCCCCTGTTTCAGCTACAACTATGCTGAATTTGAAC
TCTGTTCCAGAGCTTCATTTCTTTGATAACCCTTTAAGGCAAAACTCAATCTTGCACCAAC
CAAATGTTAGTGGAAGAAAAAGGGTTGTACCTGAAACAGAGGACAATTCTACAGGGG
ACAGAAGAAATCAGAGGATGATCAAGAACAGAGAGTCTGCTTCTAGATCAAGAGCTAG
AAAGCAGGAAAGTGACACTCTCTTTATCTTAATCATCATTTGTTCTTTTCTGTTGTCCTGT
CGATTTTTAGCAATAAATTATGTTTTTTTATAAAAACACAACAGATAAACTATGTTGCTTC
AATATGCTGATTTTCTCTGTTTTCTTTTTTGGCCATTGATGATTTTTGCAGGCCTATTTGA
ATGAGTTGGAGACGGAAGTGGCACATTTAGTTGAAGAAAATGCAAGGCTCAAGAAGC
AGCAGCAACAGGTCCTTTTATCTTTTATTACACCATGTTTTAAATTCTGTTTATAACTAAT
GACTTTCTTTAATTAGAACAAACAAATAAATGACTCAGAATTGATTGGTGTTTTTCTTAA
TTTGGACAGTTACGCTTAGCTGCAGTTATTCAAGTTCCAAAAAAGAACTCTCTTCATCGG
ACGTCAACCGCTCCATTTTGAGAAAA