factors affecting the growth of chinese hamster cells in halt selection media
TRANSCRIPT

445
Mutation Research, 35 ( 1 9 7 6 ) 4 4 5 - - 4 6 4 © Elsevier Scient i f ic Pub l i sh ing C o m p a n y , A m s t e r d a m - - P r i n t e d in T h e N e t h e r l a n d s
FACTORS AFFECTING THE GROWTH OF CHINESE HAMSTER CELLS IN HAT SELECTION MEDIA
M A R G A R E T F O X and J.M. B O Y L E
Paterson Laboratories Christie Hospital and Holt Radium Institute Manchester M20 9BX (England)
(Rece ived O c t o b e r 13 th , 1 9 7 5 ) (Rev i s ion rece ived J a n u a r y 26 th , 1 9 7 6 ) ( A c c e p t e d J a n u a r y 27 th , 1 9 7 6 )
Summary
Factors affecting the efficiency of selection of "revertants" of salvage path- way mutants in media containing amethopterin have been examined. Our V79 Chinese hamster cell line was found to require a significantly higher level of thymidine for optimal growth in such media than has been reported for other cell lines. Hypoxanthine (but not glycine) was also required for reversal of amethopterin toxicity, bu t levels did not differ significantly from those re- ported elsewhere. Growth in HAT was also dependent on plating density and serum batch. Our modification (VHAT) was compared with published HAT re- cipies in back selection reconstruction experiments. A sharp fall in EOR (effi- ciency of recovery) of wild type cells from mixtures with mutants at plating densities greater than 3500 cells/cm: (10 s cells/6 cm dish) was observed for VHAT. EOR with other HAT recipes was lower still, and was affected also by the particular mutant used in the mixture.
EMS induced "revertants" were isolated from three 8AZ ~ mutants by plating in VHAT. All revertants were however amethopterin resistant, they were also 8AZ resistant and the mobili ty of residual HGPRT (as measured by polyacryl- amide gel electrophoresis) was similar to that of their 8AZ r parents i.e. dissimi- lar from that in wild type. The modal chromosome number of V79 wild type cells was 21. No significant deviation from this mode was detected in any of
Abbreviations: 10MEM, Eagle's Minimal Essential medium with 10% foetal calf serum; IODMEM, Dulbec- co's modification of MEM with 10% foetal calf serum; HGPRT, hypoxanthine guanine phosphoribosyl transferase; EMS, ethyl methanesulphonate; 8AZ, S-azaguanine; 8AZ r, 8-azaguanine resistant clone; HAT, medium, MEM or DMEM supplemented with hypoxanthine, thymidine and amethopterin, HAT +, clones which are able to grow in HAT medium; LHAT, HAT medium as described in ref. [16]; VHAT, HAT media as used in present study, containing hypoxanthine 1 X 10 -4 M, amethopterin 4 X 10 -~ M; and thymidine 5 × 10 -4 M; f.e.s., foetal calf serum; EOR, efficiency of recovery of wild type V79 cells from mixtures with mutants.

446
the mutant lines examined. The data indicate that the recovery of colonies in HAT from 8AZ r mutants does not necessarily indicate that a back mutat ion in the structural gene for HGPRT has occurred. Thus, the frequency of HAT + col- onies cannot be taken as a direct indication of reversion frequencies.
Introduction
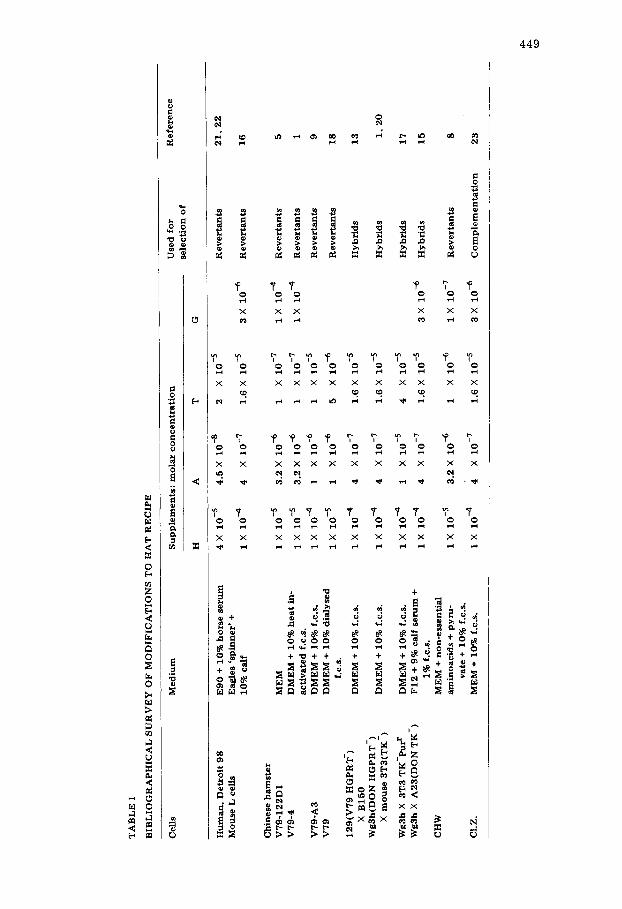
Inhibition of de novo purine and pyrimidine synthesis (by competitive inhi- bition of dihydrofolate reductase) with aminopterin or amethopterin forces cells to utilise exogenous supplies of purines and pyrimidines via the respective salvage pathways. This is the rationale for the use of medium supplemeted with hypoxanthine, aminopterin, and thymidine (plus glycine by some workers (see Table I)) for the selection both of revertants of mutations in salvage pathways [5,8,9,21,22] and hybrids between salvage pathway mutants [1,13,15--17].
Some of the factors that may affect the efficiency of HAT selection, particu- larly dialysable components of serum, have been discussed recently by Peterson et al. [18] . Since the rapid cell doubling time and the relatively stable karyo- type of V79 Chinese hamster cells have already prompted their extensive use in the s tudy of forward and back mutat ion at the HGPRT locus [2--5,14,19] it can be anticipated that they will continue to be used for such studies. Hence we thought it timely to define more carefully the factors affecting the use of HAT medium with high density populations, particularly in relation to the se- lection of revertants and measurement of reversion frequencies from 8AZ r to 8AZ s. In this paper we report the results of back selection reconstruction ex- periments which indicate that the yield of "revertants" from high density pop- ulations may be critically affected by the degree of growth inhibition produced by the particular HAT recipe used. Our data also suggest that V79 lines held by different laboratories may differ with respect to their resistance to aminop- terin. This, together with the variation in HAT response seen with the use of different serum batches [7,18] makes it obvious that it is not sufficient to rely on previously published data for suitable HAT recipes (Table I). Continuous monitoring of HAT media is therefore advisable.
These considerations lead us to recommend that future papers reporting re- version frequencies obtained with HAT selection, should contain a statement of the controls performed to check the efficiency of HAT selection under the spe- cific conditions used. This s tatement could be in the form of reconstruction ex- periments, and data indicating the amethopterin and 8-azaguanine sensitivity of the "revertants" scored.
Materials and methods
Cells The V79 cells used in this s tudy were derived from those of Dr. E.H.Y. Chu
via Dr. Colin Arlett. Mutants defective in hypoxanthine-phosphoribosyl trans- ferase (HGPRT-) were derived from V79 following EMS or UV mutagenesis

447
and their characteristics have been described [7 ]. DON cells were kindly pro- vided by Dr. Peter Goldfarb and routinely maintained in DMEM supplemented with 13% foetal calf serum. A mutant derivative of DON, wg3h (HGPRT-) was obtained from Professor G. Pontecorvo.
CHO cells were provided by Dr. C.H. Ockey and maintained in McCoy's me- dium supplemented with 10% foetal calf serum. All stocks were shown to be free from mycoplasma by the agar plating method.
Media MEM and McCoy's media were obtained from Flow Laboratories; DMEM
was prepared using the recipe of Dulbecco and Vogt [6] . Various batches of foetal calf serum from Flow Laboratories are referred to by their control num- bers. Experiments with V79 were incubated in either 5 or 10% CO2 in air mix- tures without appreciably altering the results. DON and CHO cells were incu- bated with 10% CO2 in air.
Cells were washed prior to trypsinisation with Hanks' balanced salt solution without calcium and magnesium [12] which will be referred to as HSS.
Dialysed serum was obtained from GIBCO batch no. A643119 -- Cat. no. BCL-078d.
Chemicals Hypoxanthine and thymidine were purchased from Sigma. Aminopterin was
bought from Nutritional Biochemicals Corporation; amethopterin was pur- chased as Methotrexate Sodium Parenteral or provided as USP 81.5% pure as a gift from Lederle Laboratories. Stock solutions of these chemicals were made in double distilled water and stored frozen at --20 ° C.
Folinic acid supplied as Calcium Leucovorin (injection) was obtained from Lederle. [14C] hypoxanthine (62 .0 mCi/mM) and [ 14C] formaldehyde (5.0 MCi/ mM) were obtained from Radiochemical Centre, Amersham.
Cell growth measurements Cells were generally plated at 1--5 × 104 per plastic petri dish, 6 cm in dia-
meter except where specified. At intervals, medium was removed, the cells washed with HSS and trypsinised using 0.1% trypsin (lyophilised trypsin 188 U/mg, Worthington Biochemical Company) in HSS. After cells had detached the action of trypsin was nullified by addition of growth medium containing 10% foetal calf serum. Cell suspensions were counted microscopically using a haemocytometer .
Kinetics of inactivation by HAT Cells were plated 5 X 102 per dish in normal growth medium and allowed to
attach for 4 h in the absence of HAT. Subsequently normal medium was aspi- rated from each plate and replaced with HAT medium. At intervals thereafter HAT medium was aspirated and replaced with either growth medium or growth medium containing hypoxanthine and thymidine. The plates were then further incubated for 7 days or until colonies of approximately normal size had devel- oped. The numbers of colonies per plate obtained after varying lengths of HAT

448
t reatment were compared with those obtained on control plates that had not been treated.
Changes in total cell numbers per plate during incubation in the presence of HAT were determined as described under cell growth measurements. Approx. 5 × 104 mutant cells per dish were incubated (4 h) in the absence of HAT to permit attachment. Subsequently the medium was replaced by HAT and total cell counts determined at intervals.
Reconstruction of revertant selection The efficiency of recovery of wild type cells (V79 or DON) from high densi-
ty populations of their HGPRT deficient mutants was determined as a function of both the composit ion of HAT medium and the density of mutant cells. 10-- 25 wild type cells were plated in growth medium either alone (control), or with increasing numbers of mutant cells from 1 × 104 to 1 X 106. After 4 h incuba- tion to allow attachment, growth medium was removed and replaced with HAT medium of the required composition. Plates were incubated for 6--7 days then colonies were fixed, stained and counted.
Isolation of revertants and characterisation The techniques used for isolation of "revertant" HAT + clones from 8AZ r
mutants and their characterisation i.e. assay of survival, measurement of HGPRT activity, hypoxanthine uptake and polyacrylamide gel electrophoresis of HGPRT have been described in detail elsewhere [7].
Chromosome preparations Chromosome preparations were made from V79 cells and various mutants
after exposure of exponentially growing cultures to Vinblastine sulphate (Velbe : Lilley) 0.1 /~g/ml for 1 h followed by hypotonic t reatment in KC1 0.07 M (8 min). Preparations were fixed in alcohol : acetic acid 3 : 1 and after air drying were stained in acetic orcein. At least 25 cells were analysed from each clone.
Results
Growth of V79 in "Littlefield" HAT A survey of the literature (Table I) revealed much variation in HAT recipes.
Some of these have been used extensively and are apparently suitable for V79 cell lines. As a starting point we determined the growth kinetics for V79 in 10MEM and in 10MEM supplemented with 1 X 10 -4 M hypoxanthine, 4 X 10 -7 M amethopterin, 1.6 × 10 -s M thymidine and 3 X 10 -6 M glycine [16] . The presence of glycine was not required to rescue V79 cells from amethopterin toxicity (Figs. 1 and 2) and was subsequently not added to MEM LHAT and VHAT, (see below). Control cultures in 10MEM double approximately every 8--9 h and reach a saturation density of about 5 X 106 cells per 6 cm dish. The initial doubling time of cells in LHAT was similar to that of controls (Fig. 1), but the saturation density was only about 2 × 10 s cells per dish. At low plating densities ( 5 × 102 per plate) similar saturation densities were achieved which allowed clones to be produced after 4 days incubation, at plating efficiencies similar to that of controls although by 8 days LHAT clones generally had degen-
TA
BL
E I
BIB
LIO
GR
AP
HIC
AL
SU
RV
EY
O
F M
OD
IFIC
AT
ION
S
TO
HA
T R
EC
IPE
Cel
ls
Me
diu
m
Su
pp
lem
en
ts:
mo
lar
co
nc
en
tra
tio
n
H
A
T
G
Use
d f
or
sele
ctio
n o
f
Re
fere
nc
e
Hu
ma
n,
De
tro
it 9
8
Mo
use
L c
ells
Ch
ines
e h
am
ste
r V
79
-12
2D
I V
79
-4
V7
9-A
3
V7
9
12
9(V
79
HG
PR
T-)
X
B1
50
W
g3
h(D
ON
HG
PR
T-)
X
mo
use
3T
3(T
K
)
Wg
3h
X 3
T3
TK
-Pu
r r
Wg
3h
X A
23
(DO
N T
K-)
CH
W
CL
Z.
E9
0 +
10
% h
ors
e s
eru
m
Eag
les
'sp
inn
er'
+
10
% c
alf
ME
M
DM
EM
+ 1
0%
he
at
in-
ac
tiv
ate
d f
.c.s
. D
ME
M +
10
% f
.c.s
. D
ME
M +
10
% d
ialy
sed
I,
C.S
.
DM
EM
+ 1
0%
f.c
.s.
DM
EM
+ 1
0%
f.c
.s.
DM
EM
+ 1
0%
f.c
.s.
F1
2 +
9%
cal
f se
rum
+
1%
f.c
.s.
ME
M +
no
n-e
ssen
tial
a
min
oa
cid
s +
py
ru-
vat
e +
10
% f
.c.s
. M
EM
+ 1
0%
f.c
.s.
4 X
1
0 -5
4
.5 X
1
0 -
8
2 X
1
0 -
5
1 X
10
-4
4 X
1
0 -
7
1.6
X
10
-5
1 ×
1
0-5
3
.2 ×
1
0-5
1
X
10
-7
1 ×
1
0 -5
3
.2 X
1
0-5
1
X
10
-7
1 X
10
-4
1 ×
10 -6
1
X 1
0-5
1 X
10
-5
1 X
1
0-5
5
X
10
-5
1 X
1
0 .-
4 4
X
10
-7
1
.6 X
1
0 -5
1 X
1
0-4
4
X
10 -7
1,
6 X
1
0-5
1 X
10
-4
1 X
10
-5
4 X
1
0-5
1X
1
0-4
4
X 1
0 -
7
1.6
X
10
-5
1 X
1
0 -
s 3
.2 X
1
0 -
6
1 X
1
0-5
1 X
10
-4
4
X
10
-7
1
.6 X
10
-5
3 ×
10
-5
1 X
1
0 -4
i X
1
0 -
4
3 X
1
0-5
1 X
1
0 -
7
3 X
1
0-5
Re
ve
rta
nts
Re
ve
rta
nts
Re
ve
rta
nts
Re
ve
rta
nts
Re
ve
rta
nts
Re
ve
rta
nts
Hy
bri
ds
Hy
bri
ds
Hy
bri
ds
Hy
bri
ds
Re
ve
rta
nts
Co
mp
lem
en
tati
on
21
, 2
2
16
5 1 9
18
13
1,
20
17
15
8
23
¢.O
450
lo\
10 6
1 0 5 _
10 4
71.(:.1 / •
10 7
10 {
o y ' 13.C. •
/
DaVy s 3 ~ 0 i Days2 3 ~"
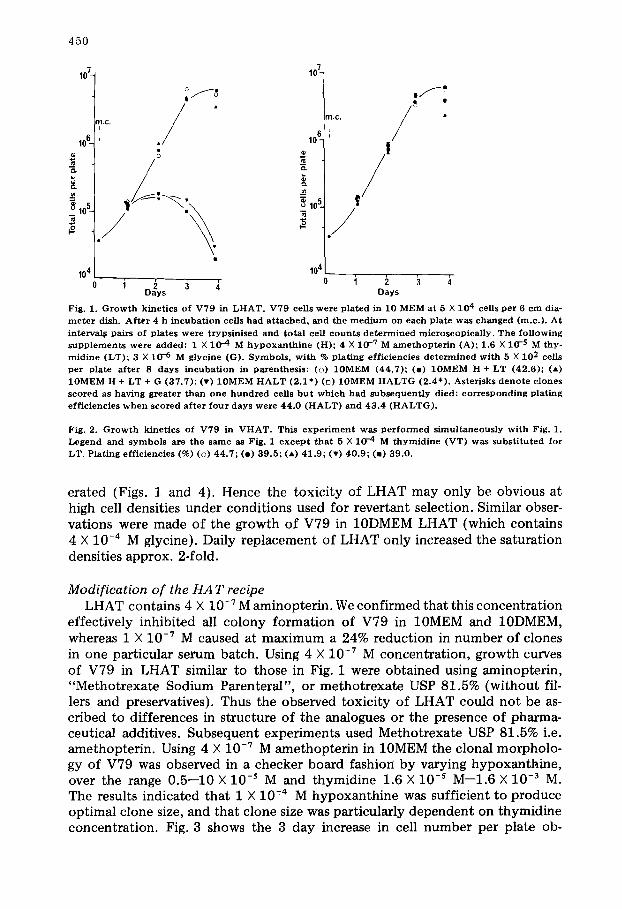
Fig. 1. G r o w t h k i n e t i c s o f V 7 9 in L H A T . V 7 9 cells we re p l a t e d in 10 MEM a t 5 X 104 cells p e r 6 c m dia- m e t e r dish. A f t e r 4 h i n c u b a t i o n cel ls h a d a t t a c h e d , a n d the m e d i u m o n e a c h p l a t e w a s c h a n g e d (m.c . ) . A t in te rva l$ pa i r s of p l a t e s we re t r y p s i n i s e d a n d t o t a l cel l c o u n t s d e t e r m i n e d m i c r o s c o p i c a l l y . T h e f o l l o w i n g s u p p l e m e n t s we re a d d e d : 1 X 1 0 --4 M h y p o x a n t h i n e (H); 4 X 1 0 -7 M a m e t h o p t e r i n (A); 1 .6 X 10- 5 M t h y - m i d i n e (LT) ; 3 X 1 0 -6 M g lyc ine (G) . S y m b o l s , w i t h % p l a t i n g e f f ic ienc ies d e t e r m i n e d w i t h 5 X 102 cells p e r p l a t e a f t e r 8 d a y s i n c u b a t i o n in p a r e n t h e s i s : (o) 1 0 M E M (44 .7 ) ; (e ) 1 0 M E M H + L T ( 4 2 . 6 ) ; (A) 1 0 M E M H + LT + G ( 3 7 . 7 ) ; (v) 1 0 M E M H A L T ( 2 . 1 " ) (D) 1 0 M E M H A L T G (2 .4* ) . As te r i sks d e n o t e c lones s c o r e d as h a v i n g g r e a t e r t h a n o n e h u n d r e d cells b u t w h i c h h a d s u b s e q u e n t l y d ied : c o r r e s p o n d i n g p l a t i n g e f f i c i enc ies w h e n s c o r e d a f t e r f o u r d a y s we re 4 4 . 0 ( H A L T ) a n d 4 3 . 4 ( H A L T G ) .
Fig. 2. G r o w t h k i n e t i c s o f V 7 9 in V H A T . This e x p e r i m e n t was p e r f o r m e d s i m u l t a n e o u s l y w i t h Fig. 1. L e g e n d a n d s y m b o l s are t he s ame as Fig. 1 e x c e p t t h a t 5 X 1 0 -4 M t h y m i d i n e (VT) w a s s u b s t i t u t e d fo r LT. P l a t ing e f f i c i enc ies (%) (o) 4 4 . 7 ; (e) 3 9 . 5 ; (A) 4 1 . 9 ; (v) 4 0 . 9 ; (m) 3 9 . 0 .
erated (Figs. 1 and 4). Hence the toxicity of LHAT may only be obvious at high cell densities under conditions used for revertant selection. Similar obser- vations were made of the growth of V79 in 10DMEM LHAT (which contains 4 X 10 -4 M glycine). Daily replacement of LHAT only increased the saturation densities approx. 2-fold.
Modification of the HA T recipe LHAT contains 4 X 10 -7 M aminopterin. We confirmed that this concentration
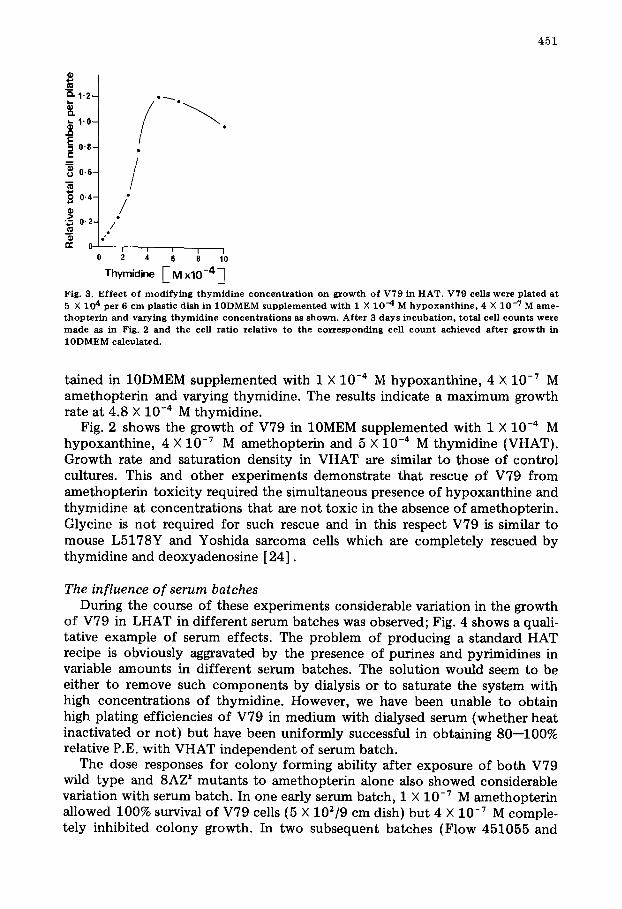
effectively inhibited all colony formation of V79 in 10MEM and 10DMEM, whereas 1 X 10 -7 M caused at maximum a 24% reduction in number of clones in one particular serum batch. Using 4 X 10 -7 M concentration, growth curves of V79 in LHAT similar to those in Fig. 1 were obtained using aminopterin, "Methotrexate Sodium Parenteral", or methotrexate USP 81.5% (without fil- lers and preservatives). Thus the observed toxicity of LHAT could not be as- cribed to differences in structure of the analogues or the presence of pharma- ceutical additives. Subsequent experiments used Methotrexate USP 81.5% i.e. amethopterin. Using 4 X 10 -7 M amethopterin in 10MEM the clonal morpholo- gy of V79 was observed in a checker board fashion by varying hypoxanthine, over the range 0.5--10 X 10 -s M and thymidine 1.6 X 10 -s M--1.6 X 10 -3 M. The results indicated that 1 X 10 =4 M hypoxanthine was sufficient to produce optimal clone size, and that clone size was particularly dependent on thymidine concentration. Fig. 3 shows the 3 day increase in cell number per plate ob-
451
D. 1 . 2 -
k. 1 .0 -
~ 0 . 8 - e-
~ 0-6-
~ 0.4-
• ,,~ 11.2-
~ 0
• ---.~ e
/ /
/°
Thymidine E M x10-4"~ Fig. 3. E f f ec t of m o d i f y i n g t h y m i d i n e concen txa t ion on g r o w t h of V79 in H A T . V79 cel l s w e r e p lated at 5 X 104 per 6 c m plast ic dish in 10DMEM s u p p l e m e n t e d wi th 1 X 10 -4 M h y p o x a n t h i n e , 4 X 10 -7 M a m e - thopter i n and varying t h y m i d i n e c o n c e n t r a t i o n s as shown . Af t e r 3 days i ncuba t ion , to ta l cell c o u n t s w e r e m a d e as in Fig. 2 and the cell ra t io re la t ive to the corresponding ce l l c o u n t achieved af ter g r o w t h in 10DMEM calcula ted .
tained in 10DMEM supplemented with 1 X 10 -4 M hypoxanthine, 4 X 10 -7 M amethopterin and varying thymidine. The results indicate a maximum growth rate at 4.8 X 10 -4 M thymidine.
Fig. 2 shows the growth of V79 in 10MEM supplemented with 1 X 10 -4 M hypoxanthine, 4 X 10 -7 M amethopterin and 5 X 10 -4 M thymidine (VHAT). Growth rate and saturation density in VHAT are similar to those of control cultures. This and other experiments demonstrate that rescue of V79 from amethopterin toxicity required the simultaneous presence of hypoxanthine and thymidine at concentrations that are not toxic in the absence of amethopterin. Glycine is not required for such rescue and in this respect V79 is similar to mouse L5178Y and Yoshida sarcoma cells which are completely rescued by thymidine and deoxyadenosine [24].
The influence o f serum batches During the course of these experiments considerable variation in the growth
of V79 in LHAT in different serum batches was observed; Fig. 4 shows a quali- tative example of serum effects. The problem of producing a standard HAT recipe is obviously aggravated by the presence of purines and pyrimidines in variable amounts in different serum batches. The solution would seem to be either to remove such components by dialysis or to saturate the system with high concentrations of thymidine. However, we have been unable to obtain high plating efficiencies of V79 in medium with dialysed serum (whether heat inactivated or not) but have been uniformly successful in obtaining 80--100% relative P.E. with VHAT independent of serum batch.
The dose responses for colony forming ability after exposure of both V79 wild type and 8AZ r mutants to amethopterin alone also showed considerable variation with serum batch. In one early serum batch, 1 X 10 -7 M amethopterin allowed 100% survival of V79 cells (5 × 102/9 cm dish) but 4 X 10 -T M comple- tely inhibited colony growth. In two subsequent batches (Flow 451055 and

452
42208) 100% survival was observed at 1 X 10 -s M but inhibition of colony for- mation was complete at 4 X 10 -s M. At higher plating densities (5 X 104/9 cm dish) inhibition apparently occurred at lower amethopterin concentration (1 X 10 -7 M a n d 4 X 10 -9 M in different sera). This latter inhibition was apparently due to growth of cultures to saturation density and subsequent detachment of cells. Amethopterin toxicity toward wild type cells has been reversed by folinic acid [10,11]. Addition of 1 X 10 -s M folinic acid to VHAT also reversed the growth inhibition of 8AZ r mutants. Hence variations in the concentrations of exogenous folate in serum may well explain the variation we have seen in both amethopterin and HAT toxicity. The precise mechanism whereby folate alters amethopterin toxicity is not clear, since total folate pools have been found to correlate not with amethopterin sensitivity per se, but with the ability of thy- midine to reverse amethopterin toxicity [24].
Relevance to other Chinese hamster lines To det/ermine how anomalous is the requirement of V79 cells for high thy-
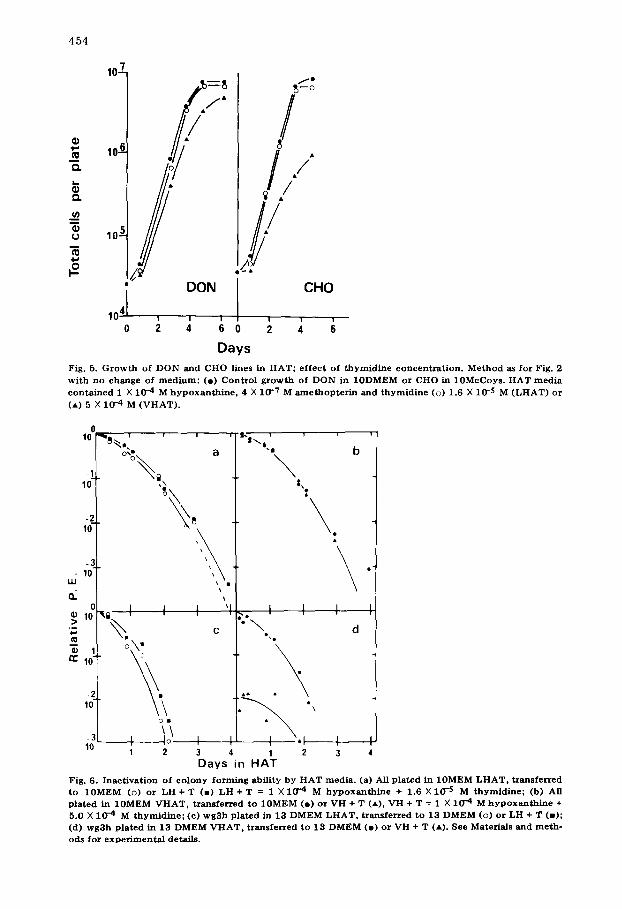
midine levels in HAT, we compared the growth of DON and CHO lines, grown in 13DMEM and 10 McCoy's respectively, with their growth in LHAT and mo- dified HAT. The results {Fig. 5) confirm that both cell lines are able to grow equally well in LHAT and unsupplemented media. Furthermore, increasing the level of thymidine in the HAT recipe proved toxic. Thus it is probable that V79 is atypical of Chinese hamster lines in a requirement for high thymidine con- centration.
Kinetics o f cellular inactivation by HA T media The rate of inactivation of HGPRT- mutants wg3h and A l l in both VHAT
and LHAT and its relationship to plating density was determined in two ways. Firstly, cells were plated in the presence of HAT which was replaced by nonse- lective media at various intervals to assay the proportion of cells able to pro- duce clones as a function of the length of HAT treatment. The results (Fig. 6) show that inactivation occurs at an increasing rate with time of t reatment for both strains. Further as would be expected, since the amethopterin concentra- tion is constant, the rates of inactivation are virtually the same in VHAT and LHAT. There was marginal restoration of residual toxicity of amethopterin by post incubation of treated cells in H + T as shown by the similarity of control curves to H + T curves. Both cell lines required 2--3 days exposure to HAT to inactivate 99% of the population. Chu et al. [3] have also reported that the ma- jority of mutant cells degenerated after 2--3 days incubation in THAG.
Secondly, changes in total cell numbers per plate were measured to deter- mine whether any cell division occurred during HAT treatment. Fig. 7 shows that the population may increase 3--4 fold before HAT inhibits growth. Inhibi- tion of growth is however density dependent and a 5-fold increase in plating density resulted in an immediate cessation of cell division and detachment of
50% of the cells by 48 h.
Effects o f H A T composit ion and plating density on back selection In order to assess the relevance of modifications to the HAT recipe in the
selection of either revertants of salvage pathway mutants, or somatic cell hy-
453
b
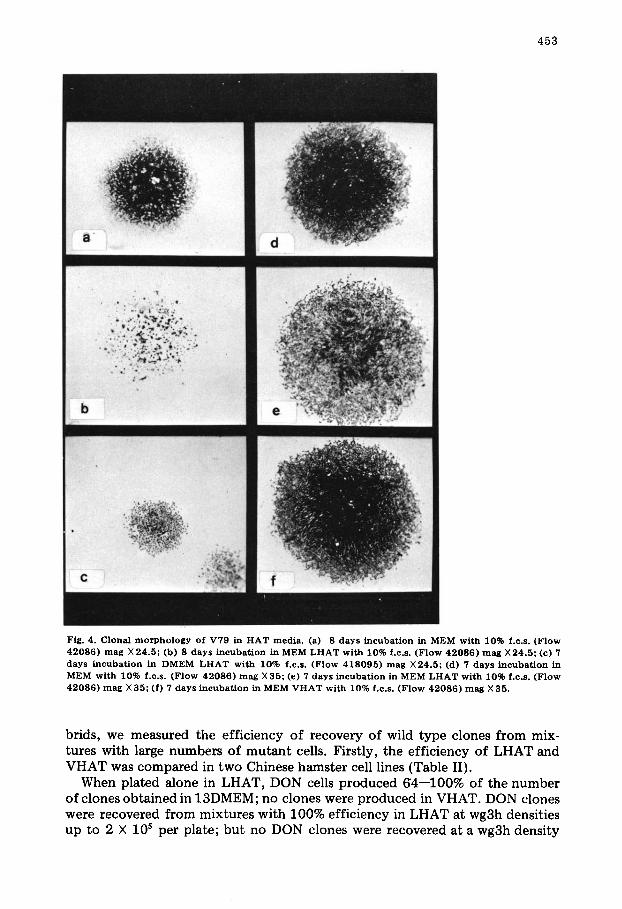
Fig. 4. Clonal morphology of V79 in HAT media. (a) 8 days incubation in MEM with 10% f.c.s. (Flow 42086) mag X 24.5; (b) 8 days incubation in MEM LHAT with 10% f.c.s. (Flow 42086) mag X24.5: (c) 7 days incubation in DMEM LHAT with 10% f.c.s. (Flow 418095) mag X24.5; (d) 7 days incubation in MEM with 10% f.c.s. (Flow 42086) mag X 35; (e) 7 days incubation in MEM LHAT with 10% f.c.s. (Flow 42086) mag X35; (f) 7 days incubation in MEM VHAT with 10% f.c.s. (Flow 42086) mag X 35.
brids, we measured the efficiency of recovery of wild type clones from mix- tures with large numbers of mutant cells. Firstly, the efficiency of LHAT and VHAT was compared in two Chinese hamster cell lines (Table II).
When plated alone in LHAT, DON cells produced 64-100% of the number of clones obtained in 13DMEM; no clones were produced in VHAT. DON clones were recovered from mixtures with 100% efficiency in LHAT at wg3h densities up to 2 X lo5 per plate; but no DON clones were recovered at a wg3h density
454
m o.
o.
¢J
1 0 :
10 ~1 "1 ! i
0 2 4 6
/
Y CHO
i i i
0 2 4 6
Days Fig. 5. G r o w t h of D O N and CHO lines in H A T ; e f fec t of t h y m i d i n e c o n c e n t r a t i o n . M e t h o d as for Fig. 2 w i th no change of m e d i u m ; (e) Con t ro l g r o w t h of DON in 10DMEM or CHO in 10McCoys . H A T m e d i a c o n t a i n e d I X 10 -4 M h y p o x a n t h i n e , 4 X 10 -7 M a m e t h o p t e r i n and t h y m i d i n e (o) 1.6 X 10- S M ( L H A T ) or (A) 5 X 10 -4 M ( V H A T ) .
-t lO
LU
- 2
10 ~
3 10
0 ~ , . a
\
~= 1 I I "1 c
o m
1 2
l . | ~ 6 J i ,
\ 1,
!
\.
i 1 . ' \ e~ e
I I .I 3 4 1 2 Days in HAT
\
Fig. 6. Inac t iva t ion of co lony f o r m i n g abi l i ty by H A T media . (a) All p l a t ed in 10MEM L H A T , t r ans fe r red to 10MEM (o) or L H + T (m) L H + T = 1 × 1 0 -4 M h y p o x a n t h i n e + 1.6 X 1 0 -5 M t h y m i d i n e ; (b) All p la ted in 10MEM V H A T , t r ans fe r red to 10MEM (e ) or VH + T (A), VH + T = 1 X 10 -4 M h y p o x a n t h i n e + 5.0 X 10 -4 M t h y m i d i n e ; (c) w g 3 h Plated in 13 DMEM LHAT0 t rans fe r red to 13 DMEM (o) or L H + T ( . ) ; (d) w g 3 h p la t ed in 13 DMEM V H A T , t r ans fe r red to 13 DMEM (o) or VH + T (A). See Materials and me th - ods for e x p e r i m e n t a l details.

455
5 10.[ . _ ,
'~ "1 • "° ~e
1 IP
I
p_ 111
0 1 2 3 Days
Fig. 7. Changes in t o t a l cel l c o u n t s o f H G P R T - m u t a n t s in H A T m e d i u m . A p p r o x . 3 X 10 4 cel ls per 6-cm dish w e r e p lated in H A T m e d i u m . A t intervals plates w e r e tryps inised and ce l l c o u n t s m a d e microscop ica l - ly as descr ibed in Materials and m e t h o d s . Closed circles, w g 3 h in 13DMEM L H A T ; o p e n circles , A l l in 10MEM V H A T .
of 1 X 1 0 6 per plate. (Table IIb). Thus LHAT provides an adequate medium for revertant or hybrid selection from this cell line and has been used for this pur- pose (Table I). VHAT on the other hand is inadequate probably due to the cytotoxic i ty of excess thymidine.
The results for the selection of V79 from high density populations of mutant A l l are presented in Table IIa. The number of clones scored in both VHAT and LHAT were not significantly different from those obtained in 10MEM. However, we failed to recover any V79 clones from mixtures with A l l selected in LHAT, and achieved 100% recovery only from 1 X 104 A l l cells/plate se- lected in VHAT.
Secondly, we compared, using mixtures of A l l and V79, the' effect of varia- tion in concentration of other HAT constituents (Table III). Modifications 4 (VHAT) and 6 gave 100% recovery for V79 at plating densities up to 1 X 10s/ plate. A comparison of data for 4 and 6 shows that recovery of V79 clones is
T A B L E II
E F F E C T OF H A T M E D I A C O M P O S I T I O N ON R E C O V E R Y OF H G P R T ÷ C L O N E S F R O M M I X T U R E S W I T H H G P R T - C E L LS
N u m b e r s o f cel ls p lated per dish
H G P R T + 25 0 25 25 25 H G P R T - 0 1 X 10 6 1 X 10 4 2 X 10 S 1 X I 0 6
N u m b e r s o f c l o n e s r e c ove r e d p e r dish
(a) V79 ( H G P R T +) + All ( H G P R T - )
MEM 25.7 + 2.09 V H A T 22.9 + 1.42 0 29 .5 + 1.01 2 .80 ± 1 .00 0 L H A T 27.5 + 1.71 0 0 0 0
(b) DON ( H G P R T +) + w g 3 h ( H G P R T - )
DMEM 23.8 + 1 .00 V H A T 0 0 0 0 0 L H A T 15.3 ± 0 .92 0 16 .0 ± 1.96 19 .8 ± 0 .89 0
456
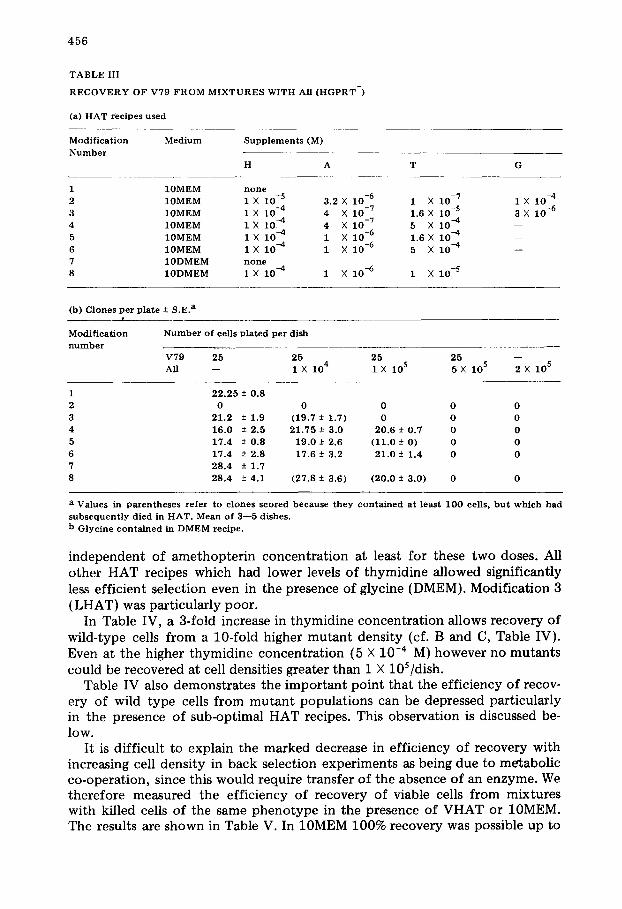
T A B L E I I I
R E C O V E R Y OF V79 F R O M M I X T U R E S W I T H All ( H G P R T - )
(a) H A T rec ipes used
M o d i f i c a t i o n M e d i u m S u p p l e m e n t s (M)
N u m b e r
H A T G
1 10MEM n o n e 2 1 0 M E M 1 X 10 -5 3.2 X 10 -6 1 X 10 -7 1 X 10 -4
3 10MEM 1 X 10 -4 4 X 10 -7 1.6 × 10 -S 3 X 10 -6 4 10MEM 1 X 10-4 4 X 10 -7 5 X 10 --4 - -
5 10MEM 1 X 10-4 1 X 10 -6 1.6 X 10 -4 - -
6 10MEM 1 X 10 -4 1 X 10 -6 5 X 10-4 - -
7 1 0 D M E M n o n e 8 1 0 D M E M 1 × 10-4 1 X 10 -6 1 X 10 -5
(b) Clones per p la te -+ S.E. a
M o d i f i c a t i o n N u m b e r o f cells p la ted pe r d i sh
n u m b e r V79 25 25 25 25 - - All - - 1 X 104 1 X 105 5 X 105 2 X 105
1 22 .25 + 0.8
2 0 0 0 0 0 3 21 .2 ± 1.9 (19.7-+ 1.7) 0 0 0
4 16.0 ± 2.5 21 .75 + 3.0 20.6 ± 0.7 0 0 5 17.4 ± 0.8 19.0 + 2.6 (11 .0 ± 0) 0 0
6 17.4 ± 2.8 17.6 + 3.2 21 .0 -+ 1.4 0 0 7 28 .4 + 1.7
8 28 .4 -+ 4.1 (27 .8 ± 3.6) (20 .0 ± 3 .0) 0 0
a Values in paxen theses r e f e r to c l o n e s scored because t h e y c o n t a i n e d a t least 100 cells, b u t w h i c h had
s u b s e q u e n t l y d ied in H A T . Mean of 3- -5 dishes .
b G lyc ine c o n t a i n e d in D M E M recipe .
independent of amethopterin concentration at least for these two doses. All other HAT recipes which had lower levels of thymidine allowed significantly less efficient selection even in the presence of glycine (DMEM). Modification 3 (LHAT) was particularly poor.
In Table IV, a 3-fold increase in thymidine concentration allows recovery of wild-type cells from a 10-fold higher mutant density (cf. B and C, Table IV). Even at the higher thymidine concentration (5 X 10 -4 M) however no mutants could be recovered at cell densities greater than 1 X 10S/dish.
Table IV also demonstrates the important point that the efficiency of recov- ery of wild type cells from mutant populations can be depressed particularly in the presence of sub-optimal HAT recipes. This observation is discussed be- low.
It is difficult to explain the marked decrease in efficiency of recovery with increasing cell density in back selection experiments as being due to metabolic co-operation, since this would require transfer of the absence of an enzyme. We therefore measured the efficiency of recovery of viable cells from mixtures with killed cells of the same phenotype in the presence of VHAT or 10MEM. The results are shown in Table V. In 10MEM 100% recovery was possible up to

457
T A B L E IV
R E C O V E R Y OF V79 FROM M I X T U R E S OF D I F F E R E N T H G P R T - M U T A N T S
Basic m e d i u m was MEM con ta in ing 10 f.c.s. (F low b a t c h No. 4 2 0 8 6 ) . Fo r H A T select ion, s u p p l e m e n t s w e r e m a d e of 1 X 10 -4 M h y p o x a n t h i n e , 4 X 10 -7 M a m e t h o p t e r i n and t h y m i d i n e a t (A) 1.6 X 10 -5 M (B) 1.6 X 10 ̀ 4 M and (C) 5 X 10 -4 M. Values in pa ren theses indica te c lones of g rea t e r t h a n 100 cells wh ich had subsequen t ly d ied in H A T . Resul t s axe m e a n ± S.E. of co lony on 3 - -5 dishes.
M u t a n t M e d i u m N u m b e r of cells p l a t ed pe r dish used
V79 10 10 10 10 - - M u t a n t - - 1 × 104 1 X 105 2 X 105 2 × 105
None 10MEM 8 . 2 ± 0 . 9 A 6 . 4 ± 1 . 8 0 0 0 0
All B 7 . 8 ± 1 . 8 7.0 ± 1 . 6 0 0 0 All C 6 . 0 ± 0 . 3 4.6 ± 1 . 5 5 . 0 ± 1 . 5 0 0
A6 B 3.5 ±1.2 6.7±2.3 0 0
AI2 B 7.0 ±0.6 (3.5±1.7) 0 0
A2 B 3.75±0.3 2.0±1.0 0 0
A13 B 7 4.6±0.9 0 0
densities of 4.0 X 10 s but was less efficient in VHAT. These data suggest that failure to recover "revertants" from high density populations is probably due to killing of potential "revertants" by toxic products released by cellular break- down. Such products could contribute significantly to the already near toxic levels of hypoxanthine and thymidine present in VHAT. Thus, for both DON and V79, the HAT medium that showed least toxicity gave the most efficient recovery.
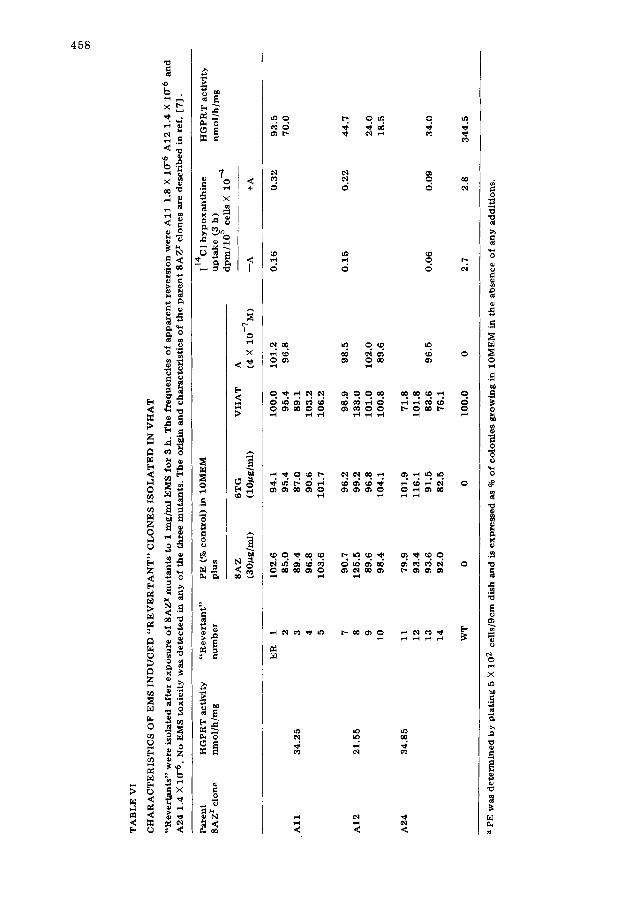
Nature of "revertant" V79 colonies recovered in VHA T We have previously presented evidence that 8AZ r clones isolated from our
V79 Chinese hamster cell line probably represent structural gene mutations, but we have been unable to isolate spontaneous revertants [7]. "Revertants" were obtained after exposure of a number of 8AZ r mutants to EMS [7], at fre- quencies similar to those reported by others [4,9,19]. Thirteen such induced "revertant" clones have been isolated. Their characteristics are summarised in Table VI. All were phenotypically stable and were 8AZ r, 6TG r and HAT + even
T A B L E V
R E C O V E R Y OF V79 F R O M M I X T U R E S W I T H I R R A D I A T I O N K I L L E D CELLS
Number of cells plated/dish
V79 20 20 20 20 20 0 V79 non-viable a 4 X 103 4 X 104 4 X 105 4 X 106 0 4 X 104
VHAT 11.4 + 0.8 15.8 + 0.5 0 0 21.2 + 0.8 0 10MEM 16.0 ± 2.6 14.2 ± 0.8 30.9 ± 1.1 0 25.1 ± 1.9 0
a V79 cells were a l lowed to a t t a c h 4 h t h e n irradiated wi th 2000 e r g s / m m 2 254 n m UV in HSS. A p p r o x . 20 viable V79 cells we re a d d e d to each dish t o g e t h e r w i th 10 ml of 10MEM or V H A T . Resul ts are m e a n -+ S.E. of c o l o n y c o u n t s on three rep l i ca te dishes.
¢91
TA
BL
E V
I
CH
AR
AC
TE
RIS
TIC
S O
F E
MS
IN
DU
CE
D "
RE
VE
RT
AN
T"
CL
ON
ES
IS
OL
AT
ED
IN
VH
AT
"Re
ve
rt~
nts
" w
ere
iso
late
d a
fter
ex
po
sure
of
8A
Z r
mu
tan
ts t
o 1
mg
/ml
EM
S f
or
3 h
. T
he
fre
qu
en
cie
s o
f a
pp
are
nt
rev
ers
ion
we
re A
ll
1.8
X 1
0 -6
A1
2 1
.4 X
10
- 6
an
d
A2
4 1
.4 X
10
-6.
No
EM
S t
ox
icit
y w
as d
ete
cte
d i
n a
ny
of
the
thre
e m
uta
nts
. T
he
ori
gin
an
d c
ha
rac
teri
stic
s o
f th
e p
are
nt
8A
Z r
clo
nes
are
de
scri
be
d i
n r
ef.
[7].
Par
ent
HG
PR
T a
ctiv
ity
"R
ev
ert
an
t"
PE
(%
co
ntr
ol)
in
10
ME
M
[14
C]
hy
po
xa
nth
ine
H
GP
RT
ac
tiv
ity
8
AZ
r cl
on
e n
mo
lfh
/mg
n
um
be
r p
lus
up
tak
e ~
3 h
) n
mo
l/h
/mg
d
pm
/10
v c
ells
X
10
-4
8
AZ
6
TG
V
HA
T
A
(30
/ag
/ml)
(1
0/z
g/m
l)
(4 X
10
-TM
) --
A
+A
• All
3
4.2
5
A1
2
21
.55
A2
4
34
.85
ER
1
10
2.6
9
4.1
1
00
.0
10
1.2
0
.16
0
.32
9
3.5
2
85
.0
95
.4
95
.4
96
.8
70
.0
3 8
9.4
8
7.0
8
9.1
4
96
.8
90
.6
10
3.2
5
10
3.6
1
01
.7
10
6.2
7 9
0.7
9
6.2
9
8.9
9
8.5
0
.15
0
.22
4
4.7
8
12
5.5
9
9.2
1
33
.0
9 8
9.6
9
6.8
1
01
.0
10
2.0
2
4.0
1
0
98
.4
10
4.1
1
00
.8
89
.6
18
.5
11
7
9.9
1
01
.9
71
.8
12
9
3.4
1
16
.1
10
1.8
1
3
93
.6
91
.5
83
.6
14
9
2.0
8
2.5
7
6.1
9
6.5
0
.06
0
.09
3
4.0
WT
0
0 1
00
.0
0 2
.7
2.8
3
44
.5
a P
E w
as d
ete
rmin
ed
by
pla
tin
g 5
X 1
02
c
eU
s/9
cm
dis
h a
nd
is
ex
pre
sse
d a
s %
of
co
lon
ies
g~
ow
ing
in
10
ME
M i
n t
he
ab
sen
ce
of
an
y a
dd
itio
ns.
459
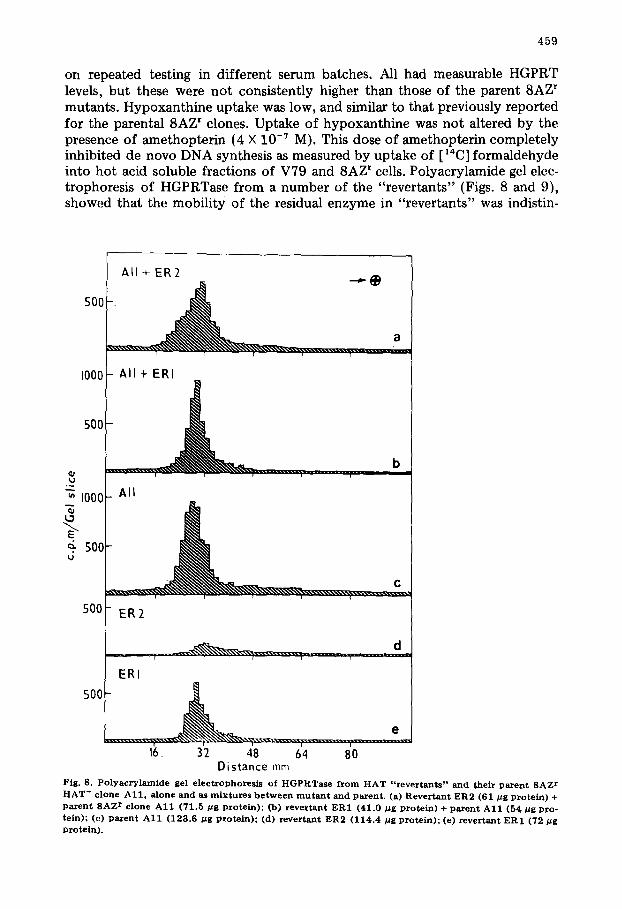
on repeated testing in different serum batches. All had measurable HGPRT levels, but these were n o t consistently higher than those of the parent 8AZ r mutants. Hypoxanthine uptake was low, and similar to that previously reported for the parental 8AZ r clones. Uptake of hypoxanthine was not altered by the presence of amethopterin (4 X 10 -7 M). This dose of amethopterin completely inhibited de novo DNA synthesis as measured by uptake of [ ~4C] formaldehyde into hot acid soluble fractions of V79 and 8AZ ~ cells. Polyacrylamide gel elec- trophoresis of HGPRTase from a number of the "revertants" (Figs. 8 and 9), showed that the mobil i ty of the residual enzyme in "revertants" was indistin-
500
I000
500
~j
I000
~- 500
500
Al l + ER2
.A - A l l + E R I
-..-@
All
ER2
ERI
500
16 32 48 64 80 Distance mm
Fig. 8. P o l y a c r y l a m i d e gel e l e c t ropbo re$ i s o f H O P R T a s e f r o m H A T " r e v e r t a n t s " and their p a r e n t 8 A Z r H A T - c l o n e A l l , a lone and as m i x t u r e s b e t w e e n m u t a n t and p a r e n t , (a) R e v e r t a n t E R 2 (61 /zg p r o t e i n ) + p a r e n t 8 A Z r c l o n e A l l ( 7 1 . 5 # g P r o t e i n ) ; (b) r e v e r t a n t E R 1 ( 4 1 . 0 ~g p r o t e i n ) + p a r e n t A l l (54 /~g p r o - t e in ) ; (c) p a r e n t A l l ( 1 2 3 . 6 /~g p r o t e i n ) ; (d) r e v e r t a n t E R 2 ( 1 1 4 . 4 ~g p r o t e i n ) ; (e) r e v c r t a n t E R 1 (72 ~tg Prote in) .

460
! 100[- ER 13
!
2001 ER I0 ' ' ' ' ' 1 (
w i ! v
g 2ooi ER9
-G C
)
d 4 0 0 ' ~
200
1000
Ib 32 48 64 80 Dlstance mm
Fig. 9. Mobility in polyacrylamide gels of HGPRT activity from a number of "revertant" HAT + c l o n e s compared with that of wild type enzyme. (a) Revert~mt I~R13 (57.6 ~g protein); (b) revertant ERIO (82.6 ~g protein); (c) revertant ER9 (94.0 pg protein); (d) revertant ER7 (101 #g protein); (e) V79 Wild type (72.5 jug protein). N.B. different scale for cpm in this ease.
guishable from that of their parent 8AZ r HAT- clones and differed significantly from that of wild type V79 cells.
Other factors may contribute to the ability of clones to survive in HAT me- dium. Firstly the HAT + clones may be tetraploid and survive because of gene dosage effects at the HGPRT locus [25]. The lack of any consistent increase in HGPRT activity (Table VI) would however argue against this. No detailed karyotypic analysis has yet been performed, however, modal chromosome numbers in wild type V79 cells, three 8AZ r clones and three HAT + "revertants" so far examined do not differ significantly. 83.2% of all cells from wild type and mutant clones had a modal chromosome number of 21. The incidence of poly- ploids was on average 6.3% and was similar in all clones. Therefore, our HAT* clones do not survive because of gene dosage effects.
A second possibility is that the HAT+ clones carry new mutations conferring enhanced resistance to amethopterin. This is probably the case as shown in Ta-

461
ble VI. "Rever tants" which had retained resistance to 8AZ show 100% survival at 4 X 10 -7 M amethopterin, whereas in the same serum batch (451055) there was no survivors from wild type or the parental 8AZ r clones at doses greater than 1 X 10 -s M. Other authors [21,22] have suggested the use of higher doses of amethopterin to eliminate amethopterin resistant colonies provided that wild-type cells can be efficiently rescued. Our initially isolated "revertant" col- onies were unable to grow in 4 X 10 -6 M amethopterin, but complete rescue of V79 cells from amethopterin toxici ty at this dose was achieved by addition of 1 X 10 -4 M hypoxanthine and 5 X 10 -4 M thymidine. Using HAT of this com- position we failed again to detect spontaneous revertants. However, HAT + col- onies were recovered from EMS treated cultures at frequencies similar to those obtained at lower amethopterin doses. These HAT + colonies proved to be resis- tant to amethopterin 4 X 10 -6 M.
Discussion
We have shown that the selection of wild type cells from mutant populations under conditions simulating the selection of HGPRT ÷ revertants, is most effi- cient with HAT recipes that support optimal growth of wild type cells. Subop-~ timal HAT recipes result in the recovery of significantly fewer revertants. The practical importance of these observations is that if sensible comparisons of reversion data are to be made then it will be necessary for future publications to provide data, in the form of back selection reconstruction experiments, to demonstrate the efficiency of the selection systems. It is not sufficient to dem- onstrate that wild type cells will clone in HAT since some HAT recipes, al- though capable of supporting clonal growth are demonstrably not optimal for revertant selection.
The influence of some serum factors on HAT selection has already been de- scribed in this s tudy and elsewhere [7,18]. We therefore recommend the reas- sesment of HAT used for revertant selection with each batch of serum used and, if studies involve the use of cells over many passages, the controls should be performed to check that the cellular response to HAT has remained con- stant.
The rationale for using HAT is that reversion or suppression of salvage path- way mutat ions will allow growth in HAT media. Our results imply that other (stable} variations in the cellular metabolism of HAT components can alter their effective concentrations and necessitate modification of the HAT recipe. In this respect V79 cell lines held in different laboratories appear to be particu. laxly variable (Table I). A corollary of this argument is that mutagenic treat- ments may induce not only reversion of the salvage pathway mutat ion or sup- pressor mutations [3] bu t may also modify the ancillary metabolism of HAT. The results of Table IV suggest this idea. The efficiency of recovery of V79 in suboptimal HAT, where stresses on the selection system may show a greater ef- fect than with optimal VHAT, is dependent on the mutant with which V79 is mixed. This result is consistent with the idea that some HGPRT mutants might have additional mutat ions affecting exogenous thymidine utilization. A low- ering of this requirement for thymidine could, for example, result in sufficient residual thymidine to be cyto toxic for V79 in high density populations.

462
Since many regimes of mutagenesis undoubtedly produce multiple mutations it is to be expected that in some systems where no revertants are recovered from control {spontaneous) populations, that induced "revertants" [4,7] might occur as a result of concomitant effects of the mutagen either directly on cellu- lar metabolism during the critical first two days of HAT selection, or via ancil- lary mutations affecting the metabolism of HAT components. We have recov- ered HAT + colonies after mutagenesis which show little or no alteration in re- sidual HGPRT activity, no loss of 8AZ resistance, and no change in electropho- retic mobility of HGPRT when compared with their HAT- parents {Table VI). These clones are resistant to amethopterin, at least up to 4 X 10 -6 M. Thus al- though many back mutat ion changes may occur without alteration in electro- phoretic mobility of the gene product and remain undetected, the absence of any change in 8AZ resistance or HGPRT activity in our HAT + colonies strong- ly suggests that they are not revertants at the HGPRT locus, but carry new mu- tations resulting in altered amethopterin sensitivity.
There is a limited amount of data in the literature relating to the character- isation of other HAT + revertants from 8AZ r cell lines. Chu et al. [3] isolated eight spontaneous HAT + revertants from an 8AZ r V79 cell line which had an HGPRT- phenotype and was unable to utilize hypoxanthine. Of these, only one proved to be 8AZ sensitive and had HGPRT activity higher than wild type. The remainder were 8AZ r and had HGPRT levels 3--23% of wild type; hypo- xanthine uptake was demonstrable only in the presence of amethopterin.
Gillen et al. [9] and Caskey et al. [2] also report the isolation of HAT + clones, both spontaneous and induced from a number of HAT- CRM- HGPRT- mutants. HGPRT activity was shown to be partially restored in some clones but did not reach wild type levels. In others it was barely detectable. Ability.to uti- lise hypoxanthine was also partially restored but again only in the presence of amethopterin to inhibit de novo synthesis.
Peterson et al. [19] also isolated and characterised spontaneous revertants from spontaneous and induced 8AZ r, HGPRT- HAT- V79 cell lines. 72% of these revertants were indistinguishable from wild type with respect to 8AZ sen- sitivity, the remainder showed up to 5% plating efficiency in 8AZ. HGPRT levels in revertants were not reported and no induced revertants were charac- terised although extensive data on specificity of reversion after exposure of dif- ferent clones to various mutagens were given.
In none of the above cases was evidence of a qualitative alteration in HGPRT gene product presented. On the other hand Gee et al. [8] have reported that HGPRT mobility was dissimilar to that of wild type in presumed revertants of an 8AZ-resistant Chinese hamster line. Thus, although a proportion of HAT* clones, particularly those occurring spontaneously, may represent true salvage pathway revertants, many induced revertants apparently do not, and in our case apparently represent new mutations at another locus.
An additional complication in the interpretation of reversion frequencies also exists [25]. The frequency of HAT + clones derived from a BUdR ~ Syrian hamster cell line was not altered by exposure to nitrosoguanidine, and the PE of isolated revertant clones was similar in HAT and BUdR. These HAT + clones did not maintain resistance in a stable way and showed extensive variation of karyotype with a strong tendency towards polyploidy suggesting that gene dos-

463
age effects may be responsible for survival in HAT in this case. Hence the fol- lowing possibilities exist, some of which are supported by examples quoted above, for the formation of HAT’ clones from 8AZ’ or BUdR’ mutants:
(a) True back mutation in the HGPRT or TK structural gene. (b) A suppressor mutation at another site. (c) Spontaneous fusion, polyploidisation and subsequent segregation resul-
ting in cell lines which carry multiple copies of the originally defective gene. (d) Occurrence of a new mutation e.g. increase in resistance to amethopterin,
increased ability to degrade thymidine or altered permeability to HAT compo- nents.
(e) Technical reasons e.g. variation in serum batches and plating density. We have examined the effects of a number of these possibilities and have
been unable to demonstrate the occurrence of back mutation in the HGPRT structural gene; all HAT’ colonies recovered under our selective conditions were amethopterin resistant relative to wild type and 8AZ’ clones. A reapprais- al of reports of induced back mutation, in particular those which occur at high frequencies, would appear to be necessary.
Acknowledgements
We wish to thank S. McMillan, P.J. Smith, Josephine Hopkins and Moira Bloomfield for excellent technical assistance at different phases of this study. We are grateful to Peter Davis, Pathology Laboratory, Withington Hospital, Manchester for screening our cells for Mycoplasma contamination.
Supported by block grants from the Medical Research Council and Cancer Research Campaign.
References
1
2
3
4
5
6
I
8
9
10
11
Boyd, Y.L. and H. Harris, Correction of genetic defects in mammalian cells by the input of small amounts of foreign genetic material, J. Cell Sci., 13, (1973) 841-861. Caskey. CT., A.L. Beaudet, D.J. Roufa and F.D. Gillen, Characterisation of 8-azaguanine resistant Chinese hamster fibroblast mutants, in Molecular and Environmental Aspects of Mutagenesis Ch. 12.
C. Thomas Publishers, Springfield, Illinois, 1974. PP. 196-207. Chu. E.H.Y., P. Brimer, K.B. Jacobson and E.V. Merriam, Mammalian cell genetics. I. Selection and chsracterisation of mutations auxotiophic for L-glutsmine or resistant to 8-azaguanine in Chinese hamster cells in vitro, Genetics, 62 (1969) 359-377. Chu. E.H.Y., P.A. Brimer. C.K. Schenley. T. Ho and H.V. Malliig, Reversion studies of chemically in- duced mutants in Chinese hamster cells in culture. in Molecular and Environmental Aspects of Muta- genesis. Ch. 11. C. Thomas Publishers. Springfield, Illinois, 1974. PP. 178-196. Chu, E.H.Y. and H.V. Mailing. Mammalian cell genetics, II. Chemical induction of specific locus muta- tions in Chinese hamster cells in vitro, Proc. Natl. Acad. Sci. U.S.A., 61 (19681 1306-1312. Dulbecco. R. and M. Vogt. Plaque formation end isolation of pure lines of poliomyelitis viruses, J. EXP. Med., 99 (1954) 167-182. Fox, M., J.M. Boyle and B.W. Fox. Biological and biochemical characterisation of purine analogue re- sistant clones of Chinese hamster cells, Mutation Res. (in press). Gee. P.A.. M. Ray, T. Mohandas, G.R. Douglas, H.R. Palser, B.J. Richardson and J.L. Hamerton, Char- acteristics of an HPRT-deficient Chinese hamster line, Cytogenet. Cell Genet., 13 (1974) 437447. Gillen, F.D.. D.J. Roufa, A.L. Beaudet and C.T. Caskey, Azaguanine resistance in mammalian cells. I. Hypoxanthine-guanine phosphoribosyl transferase. Genetics, 72 (1972) 239-252. Goldin, A.. N. Mantel, S.W. Greenhouse, J.M. Venditti and S.R. Humphreys, Estimation of the snti- leukemia potency of antimetabollte aminopterin administered alone or in combination with Citro- vorum factor or Folinic acid, Cancer Res., 13 (1953) 843-850. Goldin. A., J.M. Venditti, I. Kline and N. Mantil. Eradication of leukemia cells (L1210) by metho-

4 6 4
t r e x a t e a n d m e t h o t r e x a t e p lu s C i t r o v o r u m f a c t o r , N a t u r e , 2 1 2 ( 1 9 6 6 ) 1 5 4 8 - - 1 5 5 0 . 12 H a n k s , J .H . a n d R .E . Wal lace , R e l a t i o n o f o x y g e n a n d t e m p e r a t u r e in t he p r e s e r v a t i o n o f t i ssue b y re-
f r i ge r a t i on , P roc . Soc. Exp . Biol. (N.Y. ) 71 ( 1 9 4 9 ) 1 9 6 - - 2 0 1 . 13 Har r i s , M., N o n - m e n d e l i a n s e g r e g a t i o n in h y b r i d s b e t w e e n Chinese h a m s t e r cells, J . Cell. Phys io l . ,
86 ( 1 9 7 5 ) 4 1 3 - - 4 3 1 . 14 H u b e r m a n , E., L. Asp i ras , C. H e i d e l b e r g e r , P .L . G r o v e r a n d P. Sims, M u t a g e n i c i t y to m a m m a l i a n
cells o f e p o x i d e s a n d o t h e r de r iva t ives o f p o l y c y c l i c h y d r o c a r b o n s , P roc . Nat . A c a d . Sci. U.S .A. , 6 8 ( 1 9 7 1 ) 3 1 9 5 - - 3 1 9 9 .
1 5 L i m b o s c h , S., V. H e i l p o r n , A. Lievens , J .L . D e c o e n a n d F. Z a m p e t t i - B o s s e l e r , R a d i a t i o n r e s p o n s e o f a s o m a t i c cell h y b r i d , In t . J . R a d i a t . Biol . , 26 ( 1 9 7 4 ) 1 9 7 - - 2 0 1 .
16 L i t t l e f i e ld , J .W. , The inos in ic ac id p y r o p h o s p h o r y l a s e ac t i v i t y o f m o u s e f i b r o b l a s t s p a r t i a l l y r e s i s t an t t o 8 - a z a g u a n i n e , P roc . Nat . A c a d . Sci. U.S .A. , 50 ( 1 9 6 3 ) 5 6 8 - - 5 7 3 .
17 Mat in , G. a n d L. P u g l i a t t i - C r i p p a , P r e f e r e n t i a l s e g r e g a t i o n o f h o m o s p e c i f i c g r o u p s o f c h r o m o s o m e s in h e t e r o s p e c i f i c s o m a t i c cell h y b r i d s , E x p . Cell Res . , 7 0 ( 1 9 7 2 ) 2 5 3 - - 2 5 6 .
18 P e t e r s o n , A . R . , H. P e t e r s o n a n d C. He ide lbe rge r , The i n f l u e n c e o f s e r u m c o m p o n e n t s o n the g r o w t h a n d m u t a t i o n o f Ch inese h a m s t e r cells in m e d i u m c o n t a i n i n g a m i n o p t e r i n , M u t a t i o n Res . , 2 4 ( 1 9 7 4 ) 2 5 - - 3 3 .
19 P e t e r s o n , A .R . , H. P e t e r s o n a n d C. H e i d e l b e r g e r , R e v e r s i o n o f t he 8 - a z a g u a n i n e r e s i s t an t P h e n o t y p e o f v a r i a n t Ch inese h a m s t e r cel ls t r e a t e d w i t h a l k y l a t i n g agen t s a n d 5 - b r o m o - 2 - d e o x y u r i d i n e , M u t a t i o n Res . , 29 ( 1 9 7 5 ) 1 2 7 - - 1 3 7 .
2 0 P o n t e c o r v o , G. , I n d u c t i o n o f d i r e c t i o n a l c h r o m o s o m e e l i m i n a t i o n in s o m a t i c cell h y b r i d s , N a t u r e , 2 3 0 ( 1 9 7 1 ) 2 6 7 - - 3 6 9 .
21 S z y b a l s k a , E .H. a n d W. Szyba l sk i , G e n e t i c s o f h u m a n cell l ines. IV. D N A - m e d i a t e d he r i t ab l e t r a n s f o r - m a t i o n o f a b i o c h e m i c a l t r a i t , P roc . Nat l . A c a d . Sci. U.S .A. , 4 8 ( 1 9 6 2 ) 2 0 2 6 - - 2 0 3 4 .
22 Szyba l sk i , W., E .H. S z y b a l s k a a n d G. Ragn i , G e n e t i c s t ud i e s w i th h u m a n cell l ines, Na t . C a n c e r Inst . Monog~. , 7 ( 1 9 6 2 ) 7 5 - - 8 8 .
23 Sek iguch i , T. a n d F. S e k i g u c h i , In te ra l le l l c c o m p l e m e n t a t i o n in h y b r i d cells de r ived f r o m Ch inese h a m s t e r d i p l o i d c lones d e f i c i e n t in b y p o x a n t h i n e g u a n i n e p h o s p h o r i b o s y l t r a n s f e r a s e ac t i v i t y , E x p . Cell Res . , 77 ( 1 9 7 3 ) 3 9 1 - - 4 0 4 .
24 Ta t t e r sa l l , M .H .N . , R .C . J a c k s o n , S .T.M. J a c k s o n a n d K . R . H a r r a p , F a c t o r s d e t e r m i n i n g cell sens i t iv i ty t o m e t h o t r e x a t e : s tud ie s o f f o l a t e a n d d e o x y r i b o n u c l e o s i d e t r i p h o s p h a t e p o o l s in five m a m m a l i a n cell l ines, Eur . J . C a n c e r , 10 ( 1 9 7 4 ) 8 1 9 - - 8 2 6 .
2 5 Terzi , M., C h r o m o s o m a l v a r i a t i o n a n d the o r ig in o f d r u g r e s i s t a n t m u t a n t s in m a m m a l i a n cell l ines , P roc . Na t . A c a d . Sci. U .S .A. , 71 ( 1 9 7 4 ) 5 0 2 7 - - 5 0 3 1 .