expression of catenins and e-cadherin during epithelial restitution in inflammatory bowel disease
TRANSCRIPT

J. Pathol. 185: 413–418 (1998)
EXPRESSION OF CATENINS AND E-CADHERINDURING EPITHELIAL RESTITUTION IN
INFLAMMATORY BOWEL DISEASE
. 1, . 2, 2, 2, 2, . 3, 1 2*
1Department of Surgery, Imperial College of Science, Technology and Medicine, Hammersmith Campus, London, U.K.2Department of Histopathology, Imperial College of Science, Technology and Medicine, Hammersmith Campus, London, U.K.
3Department of Gastroenterology, University of Leicester, Leicester, U.K.
SUMMARY
Catenins are cytoplasmic proteins associated with E-cadherin, the prime mediator of cell–cell adhesion. Perturbation in any of thesemolecules results in altered intercellular adhesion, cell differentiation, and increased migration. In this study, the expression and cellularlocalization of catenins and E-cadherin in inflammatory bowel disease were examined. The expression of E-cadherin; á-, â-, andã-catenin; and p120 was evaluated immunohistochemically in 31 paraffin-embedded colonic specimens from 21 patients with ulcerativecolitis and Crohn’s disease. Loss of normal membranous E-cadherin and á-catenin staining was detected at the mucosal edges aroundepithelial ulcerations in all cases of active ulcerative colitis and in 50 per cent of cases with active Crohn’s disease. Reduced expressionof p120 protein was also found at the margins of ulcerated mucosa in all cases of active ulcerative colitis and in 75 per cent of those withactive Crohn’s disease. There was a statistically significant correlation between the expression of E-cadherin, á-catenin and p120 anddisease activity. There were no changes in â- and ã-catenin expression in either ulcerative colitis on Crohn’s disease. These findingsindicate that altered expression of E-cadherin, á-catenin, and p120 occurs during mucosal ulceration in inflammatory bowel disease.These changes may be involved in promoting cell migration during epithelial restitution of the gastrointestinal mucosa. ? 1998 JohnWiley & Sons, Ltd.
KEY WORDS—E-cadherin; catenins; cell migration; epithelial restitution; ulceration; inflammatory bowel disease
INTRODUCTION
E-cadherin is the prime mediator of intercellular ad-hesion in epithelial cells. This transmembrane glycopro-tein, localized mainly in the zonula adherens junctions,mediates by its extracellular domain cell–cell adhesionthrough calcium-dependent homotypic interactions.1 Itscytoplasmic domain associates with a group of under-coat proteins termed catenins (á-, â-, and ã-catenin).2Both â- and ã-catenin bind directly to the cytoplasmicdomain of E-cadherin, while á-catenin links the boundâ- or ã-catenin to the actin microfilament network of thecellular cytoskeleton.3 This binding is essential for theformation of stable cell–cell adhesion and rendersá-catenin a regulatory molecule of the adhesive functionof E-cadherin.4,5 The p120 protein, which was originallyidentified as a substrate of src and several other receptortyrosine kinases, is another member of the cateninfamily.6,7 p120, like its structural homologues â- andã-catenin, binds directly to the cytoplasmic domain ofE-cadherin.7,8
*Correspondence to: Massimo Pignatelli, MD, PhD, MRCPath,Imperial College of Science, Technology and Medicine, Division ofInvestigative Science, Department of Histopathology, HammersmithCampus, Du Cane Road, London W12 0NN, U.K. E-mail:[email protected]
Contract grant sponsors: Crohn’s In Childhood Research Associ-ation (CICRA); Medical Research Council.
CCC 0022–3417/98/040413–06 $17.50? 1998 John Wiley & Sons, Ltd.
Normal E-cadherin expression and function are essen-tial for the induction and maintenance of polarized anddifferentiated epithelial phenotypes during embryonicdevelopment and morphogenesis.1 There is evidence thatthe E-cadherin/catenin complex is also involved in thegrowth control of epithelial cells.9 The importance ofcadherin-mediated cell adhesion in intestinal epitheliumis evident from studies using chimeric-transgenic mice.10
In this experimental model, perturbation of E-cadherinfunction in crypt enterocytes was associated withincreased cell migration along the crypt–villus axis,increased cell proliferation and apoptosis, disruption ofmucosal barrier function, and development of progres-sive inflammatory changes, with features of Crohn’sdisease.10 Conversely, forced expression of E-cadherinrestored the altered cell cycle.11 Similarly, disruption ofE-cadherin-mediated cell adhesion in villous enterocytesresulted in loss of their differentiated polarized pheno-type and alteration of their cell cycle.12 These changeswere associated with cellular redistribution of E-cadherin from the cell membrane to the cytoplasm.11
Altered E-cadherin expression in inflammatory boweldisease (IBD) has also been demonstrated in humantissues,13,14 but the expression of the catenins in IBDand their relationship to E-cadherin have not beenstudied before. In this study, we evaluated immuno-histochemically the expression of E-cadherin con-comitantly with á-, â-, and ã-catenin, and p120 in IBD.
Received 13 March 1997Accepted 20 March 1998

414 A. J. KARAYIANNAKIS ET AL.
The relationship between their expression and clinico-pathological data was also evaluated.
MATERIALS AND METHODS
Tissue specimens
Thirty-one formalin-fixed, paraffin-embedded colonictissue specimens from 21 patients undergoing colectomyfor Crohn’s disease (n=9) or ulcerative colitis (n=12)were retrieved from the archival material of the Histo-pathology Department of Hammersmith Hospital,London, U.K. Representative blocks containing areas ofulceration were selected and 5 ìm sections were cut. Incases of active disase, a second section from an area withmild activity was also taken. All sections werere-evaluated and graded by the same histopathologist(MP) before immunostaining.
Antibodies
A mouse monoclonal anti-human E-cadherin anti-body (HECD-1) was kindly provided by Professor M.Takeichi (Kyoto University, Kyoto, Japan) and wasused undiluted as culture supernatant.15 Commerciallyavailable mouse monoclonal antibodies against á-, â-,and ã-catenin, and p120 (Transduction Laboratories,Lexington, KY, U.S.A.) were used at the followingdilutions: anti-á-catenin: 1:500; anti-â-catenin; and anti-ã-catenin and anti-p120: 1:1000.
Immunohistochemistry
Streptavidin–biotin indirect immunostaining was usedafter application of the microwave antigen retrievaltechnique.16 Sections were immersed in 0·1 citratebuffer at pH 6·0 and heated in a microwave oven at750 W for four 5-min cycles. After rinsing in 0·01 phosphate buffered saline (PBS), normal rabbit serumdiluted at 1:20 was applied for 20 min to block non-specific antibody binding. Subsequently, sections wereincubated overnight at 4)C with the primary antibodiesagainst E-cadherin; á-, â-, and ã-catenin; and p120respectively. After further rinsing in PBS, sections wereincubated with biotinylated rabbit anti-mouse IgG(Dako Ltd, High Wycombe, U.K.) and streptavidin–peroxidase conjugate (Amersham International,Buckinghamshire, U.K.). The peroxidase reaction wasdeveloped with 0·01 per cent hydrogen peroxide in 0·05per cent diaminobenzidine tetrahydrochloride (SigmaChemical Co., Poole, U.K.) solution in PBS. For p120staining, sections were incubated with streptavidin–alkaline phosphatase complex (Dako). The alkalinephosphatase activity was developed with naphtholAS-MX phosphate as substrate and fast red as achromogen (Sigma).
Normal colonic tissues of homogeneous immuno-phenotype for the studied antigens were included aspositive controls. Negative controls consisted of dupli-cate sections stained simultaneously, in which the pri-mary antibody was omitted and replaced by PBS.
? 1998 John Wiley & Sons, Ltd.
Evaluation of immunostaining
Sections were examined under light microscopy bythree independent observers (AJK, KS, MP). Whenmore than 50 per cent of cells exhibited uniformly thesame intensity and pattern of immunostaining as theadjacent normal mucosa, the antigen expression wasconsidered normal. Antigen expression was consideredabnormal when less than 50 per cent of cells stained fora particular antigen, when cells showed a heterogene-ously weak or altered distribution (i.e., cytoplasmic ornuclear) of immunostaining, or, finally, when completeabsence of staining was observed.
Statistical analysis
For statistical analysis, antigen expression was consid-ered either normal or abnormal. Correlations betweenantigen expression and clinicopathological variableswere evaluated by Fisher’s exact test. P<0·05 was con-sidered statistically significant.
RESULTS
Expression of E-cadherin
Uniform, membranous E-cadherin staining localizedat the intercellular borders was observed in all caseswithin normal colonic mucosa present in the sections, aswell as in all cases with inactive disease. Reducedexpression of E-cadherin was observed in 38 per cent ofcases (Table I). A significantly higher proportion ofcases with active disease (80 per cent) exhibited reducedE-cadherin expression compared with inactive disease (0per cent) (P=0·0002). In these cases, heterogeneousstaining with loss of normal membranous E-cadherinimmunoreactivity and cytoplasmic staining wasobserved in the cuboidal non-polarized cells forming aregenerating monolayer adjacent to the ulceratedmucosa (Figs 1A and 2A). In contrast, epithelial cellswith a columnar shape and polarized nuclei distant fromthe ulcer margin expressed normal membranousE-cadherin immunoreactivity similar to that seen in theadjacent normal mucosa. This abnormal pattern ofE-cadherin expression was observed in all cases of activeulcerative colitis and in 50 per cent of cases with activeCrohn’s disease.
Expression of á-catenin
In normal colonic epithelium, á-catenin was presentuniformly at the lateral cell borders. Staining in theapical or basal cell borders was not detected. All speci-mens of inactive disease expressed á-catenin homogene-ously with the same intensity and pattern of distributionas normal colonic mucosa. Abnormal á-catenin expres-sion was observed in ten cases (80 per cent) of activedisease. Loss of membranous immunoreactivity withcytoplasmic accumulation was evident in epithelial cellsat the mucosal end adjacent to the ulceration (Figs 1Band 2B). The non-ulcerated mucosa showed normalá-catenin expression. Abnormal á-catenin expression
J. Pathol. 185: 413–418 (1998)
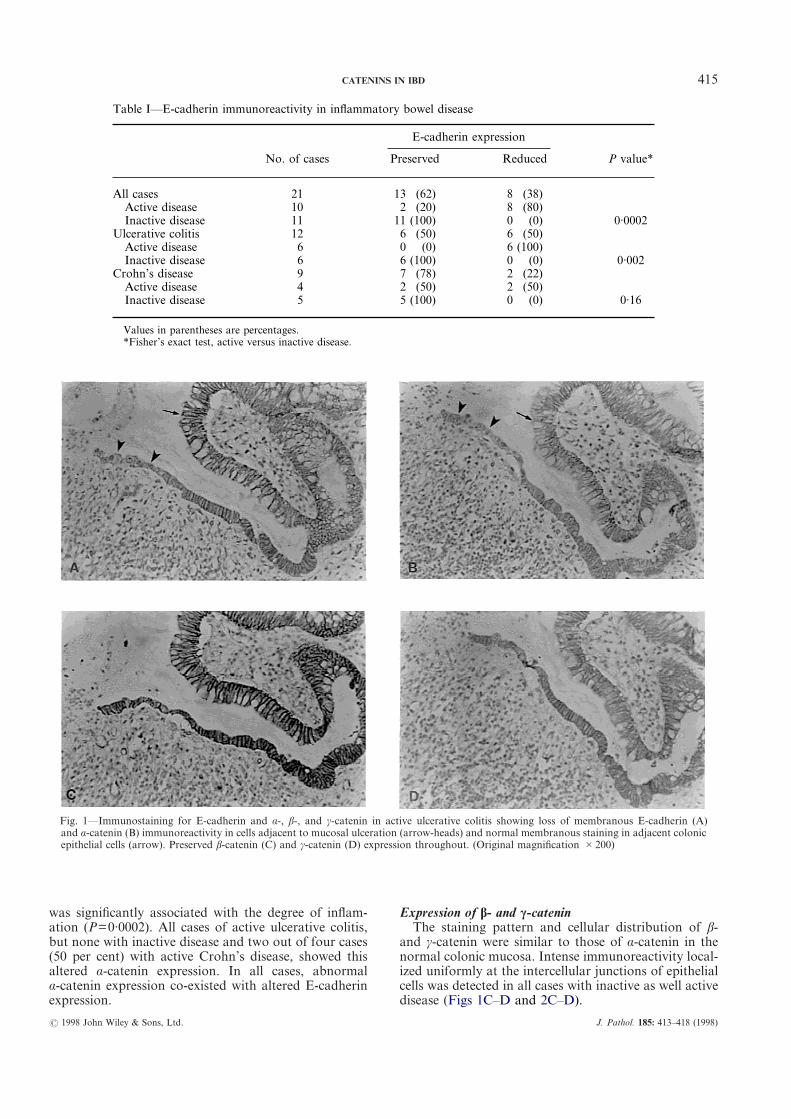
415CATENINS IN IBD
was significantly associated with the degree of inflam-ation (P=0·0002). All cases of active ulcerative colitis,but none with inactive disease and two out of four cases(50 per cent) with active Crohn’s disease, showed thisaltered á-catenin expression. In all cases, abnormalá-catenin expression co-existed with altered E-cadherinexpression.
? 1998 John Wiley & Sons, Ltd.
Expression of â- and ã-cateninThe staining pattern and cellular distribution of â-
and ã-catenin were similar to those of á-catenin in thenormal colonic mucosa. Intense immunoreactivity local-ized uniformly at the intercellular junctions of epithelialcells was detected in all cases with inactive as well activedisease (Figs 1C–D and 2C–D).
Fig. 1—Immunostaining for E-cadherin and á-, â-, and ã-catenin in active ulcerative colitis showing loss of membranous E-cadherin (A)and á-catenin (B) immunoreactivity in cells adjacent to mucosal ulceration (arrow-heads) and normal membranous staining in adjacent colonicepithelial cells (arrow). Preserved â-catenin (C) and ã-catenin (D) expression throughout. (Original magnification #200)
Table I—E-cadherin immunoreactivity in inflammatory bowel disease
No. of cases
E-cadherin expression
P value*Preserved Reduced
All cases 21 13 (62) 8 (38)Active disease 10 2 (20) 8 (80)Inactive disease 11 11 (100) 0 (0) 0·0002
Ulcerative colitis 12 6 (50) 6 (50)Active disease 6 0 (0) 6 (100)Inactive disease 6 6 (100) 0 (0) 0·002
Crohn’s disease 9 7 (78) 2 (22)Active disease 4 2 (50) 2 (50)Inactive disease 5 5 (100) 0 (0) 0·16
Values in parentheses are percentages.*Fisher’s exact test, active versus inactive disease.
J. Pathol. 185: 413–418 (1998)
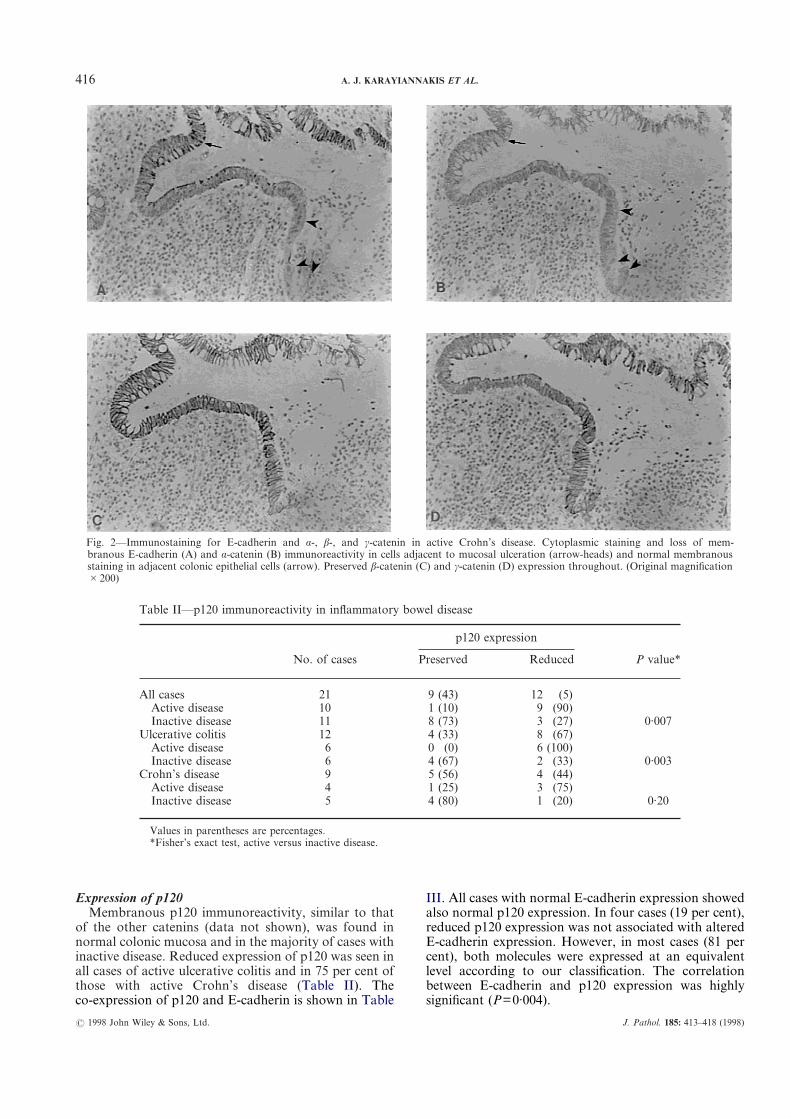
416 A. J. KARAYIANNAKIS ET AL.
Expression of p120Membranous p120 immunoreactivity, similar to that
of the other catenins (data not shown), was found innormal colonic mucosa and in the majority of cases withinactive disease. Reduced expression of p120 was seen inall cases of active ulcerative colitis and in 75 per cent ofthose with active Crohn’s disease (Table II). Theco-expression of p120 and E-cadherin is shown in Table
? 1998 John Wiley & Sons, Ltd.
III. All cases with normal E-cadherin expression showedalso normal p120 expression. In four cases (19 per cent),reduced p120 expression was not associated with alteredE-cadherin expression. However, in most cases (81 percent), both molecules were expressed at an equivalentlevel according to our classification. The correlationbetween E-cadherin and p120 expression was highlysignificant (P=0·004).
Fig. 2—Immunostaining for E-cadherin and á-, â-, and ã-catenin in active Crohn’s disease. Cytoplasmic staining and loss of mem-branous E-cadherin (A) and á-catenin (B) immunoreactivity in cells adjacent to mucosal ulceration (arrow-heads) and normal membranousstaining in adjacent colonic epithelial cells (arrow). Preserved â-catenin (C) and ã-catenin (D) expression throughout. (Original magnification#200)
Table II—p120 immunoreactivity in inflammatory bowel disease
No. of cases
p120 expression
P value*Preserved Reduced
All cases 21 9 (43) 12 (5)Active disease 10 1 (10) 9 (90)Inactive disease 11 8 (73) 3 (27) 0·007
Ulcerative colitis 12 4 (33) 8 (67)Active disease 6 0 (0) 6 (100)Inactive disease 6 4 (67) 2 (33) 0·003
Crohn’s disease 9 5 (56) 4 (44)Active disease 4 1 (25) 3 (75)Inactive disease 5 4 (80) 1 (20) 0·20
Values in parentheses are percentages.*Fisher’s exact test, active versus inactive disease.
J. Pathol. 185: 413–418 (1998)

417CATENINS IN IBD
As shown in Tables I–III, we were able to demon-strate that abnormal expression of E-cadherin,á-catenin, and p120 correlated significantly with diseaseactivity (P<0·001). In fact, abnormal cytoplasmic stain-ing was observed in the acute IBD cases, with exacer-bation and ulceration, but not in the inactive cases.Finally, with regard to the abnormal E-cadherin/cateninexpression, a statistically significant difference wasobserved between ulcerative colitis and Crohn’s disease(P<0·01) (Tables I–III). These disease-specific altera-tions correlated with the presence of active chronicinflammation and ulceration.
DISCUSSION
This study demonstrated alterations in E-cadherin/catenin expression in both ulcerative colitis and Crohn’sdisease. Loss of normal membranous immunoreactivityof E-cadherin and á-catenin, as well as p120, wasobserved in cells around areas of ulceration. Non-ulcerated mucosa and mucosa in inactive disease dem-onstrated the normal distribution of the cadherin/catenin complex. Preserved E-cadherin and cateninexpression within inflamed epithelium suggests thestability of this complex; its alteration is consistent withan acquired feature during mucosal ulceration.
It is of note that abnormal á-catenin expressionalways co-existed with altered E-cadherin expression.Results from in vitro studies may serve as an explanationfor this finding. L-cells lacking endogenous cadherinalso have a minimal amount of á-catenin, although theyexpress á-catenin mRNA.17 Their transfection withE-cadherin cDNA resulted not only in cadherin, but alsoin á-catenin upregulation, without concomitant changesin á-catenin mRNA. When carboxy-tail truncatedE-cadherin, which cannot bind to á-catenin, was usedfor transfection, á-catenin expression did not increase.17
These results suggest that á-catenin binding toE-cadherin is essential for stable á-catenin proteinexpression. Therefore, the downregulation of E-cadherinobserved in this study could have resulted in reducedá-catenin expression.
Diffuse E-cadherin and á-catenin cytoplasmic im-munoreactivity has been observed in several adeno-carcinomas and has been correlated with cell–celladhesion-dependent histological features such as differ-
? 1998 John Wiley & Sons, Ltd.
entiation, invasiveness, and metastasis.18–23 Interest-ingly, the most aggressive tumours were those withsimultaneously reduced E-cadherin and á-cateninexpression.23 Presumably, cytoplasmic localization ofE-cadherin and á-catenin represents structural or func-tional alterations of these molecules, which render themnon-functional with subsequent loss of cell–cell adhe-sion. Cellular redistribution of E-cadherin from the cellmembrane to the cytoplasm and increased cell migrationwere observed after disruption of E-cadherin-mediatedcell adhesion in the murine small intestine.11 We haverecently shown that perturbation of E-cadherin-mediated cell adhesion is associated with cell migrationand epithelial restitution in an in vitro model of epithelialinjury.15 It is likely that downregulation of E-cadherinexpression in cells at the margins of mucosal ulcerationmay represent an important step for epithelial restitutionin vivo. Reduced cell adhesion would permit an increasedmotility of marginal epithelial cells and allow theirmigration over the ulcerated area in order to restoremucosal integrity. Since E-cadherin and á-cateninexpression in non-ulcerated mucosa was normally pre-served, this disruption of intercellular adhesion must betransient, occurring only during mucosal injury. Aftercell migration and before re-establishment, cells mustre-express E-cadherin and the catenins on the cell sur-face, in order to avoid cell shedding and to form anintact and stable mucosal layer.
The expression of p120 and E-cadherin were alsoclosely correlated. The binding site of p120 onE-cadherin is in close proximity to, but distinct fromthat of catenins.24 p120, like â- and ã-catenin, is phos-phorylated in response to stimulation by several growthfactors.8 Its tyrosine phosphorylation by the src tyrosinekinase is associated with cell transformation.6 Thesedata suggest that p120 may participate in the modu-lation of E-cadherin-mediated cell adhesion, perhapsindependently from the catenins.
Components of the E-cadherin/catenin complex arephosphorylated by a number of growth factor receptorsupon their ligand binding activation. Tyrosine phospho-rylation of the catenins appears to be a possible mech-anism for functional modulation of the E-cadherin/catenin adhesion system. Epidermal growth factor(EGF), as well as hepatocyte growth factor/scatterfactor (HGF/SF), induces tyrosine phosphorylation ofâ- and ã-catenin and p120 with subsequent cellularredistribution of E-cadherin, cell dissociation, andincreased cell motility.8,25,26 EGF, HGF/SF, and mem-bers of the transforming and fibroblast growth factorfamily have been shown to promote cell migration andthereby to enhance epithelial restitution in vitro.27–29 It ispossible that the integrated action of multiple growthfactors during different stages of ulceration and mucosalhealing may contribute to the changes in E-cadherin/catenin expression seen in vivo.
In conclusion, this study demonstrated that changesin the expression of catenins and E-cadherin occurduring ulceration in inflammatory bowel disease. Evalu-ation of the interactions between the E-cadherin/catenincomplex and growth factors may provide some insightinto the reparative process of the gastrointestinal tract.
Table III—Relationship between E-cadherin and p120expression
E-cadherinexpression
p120 expression
Total P value*Preserved Reduced
Preserved 9 (43) 4 (19) 13Reduced 0 (0) 8 (38) 8Total 9 12 21 0·004
Values in parentheses are percentages.*Fisher’s exact test.
J. Pathol. 185: 413–418 (1998)

418 A. J. KARAYIANNAKIS ET AL.
ACKNOWLEDGEMENTS
We thank Professor M. Takeichi for kindly providingthe HECD-1 antibody. This work was supported bythe Crohn’s In Childhood Research Association(CICRA) and the Medical Research Council. Dr A. J.Karayiannakis is a recipient of a Greek State Scholar-ships Foundation scholarship. Mr J. Efstathiou is arecipient of a Wellcome Vacation scholarship.
REFERENCES1. Takeichi M. Cadherin cell adhesion receptors as a morphogenetic regulator.
Science 1991; 251: 1451–1455.2. Gumbiner BM, McCrea PD. Catenins as mediators of the cytoplasmic
functions of cadherins. J Cell Sci 1993; 17 (Suppl): 155–158.3. Hinck L, Näthke IS, Papkoff J, Nelson WJ. Dynamics of cadherin/catenin
complex formation: novel protein interactions and pathways of complexassembly. J Cell Biol 1994; 125: 1327–1340.
4. Hirano S, Kimoto N, Shimoyama Y, Hirohashi S, Takeichi M. Identifica-tion of a neural á-catenin as a key regulator of cadherin function andmulticellular organization. Cell 1992; 70: 293–301.
5. Shimoyama Y, Nagafuchi A, Fujita S, et al. Cadherin dysfunction in ahuman cancer cell line: possible involvement of loss of á-catenin expressionin reduced cell–cell adhesiveness. Cancer Res 1992; 52: 5770–5774.
6. Reynolds AB, Herbert L, Cleveland JL, Berg ST, Gaut JR. p120, a novelsubstrate of protein tyrosine kinase receptors and of p60v-src, is related tocadherin-binding factors â-catenin, plakoglobin and armadillo. Oncogene1992; 7: 2439–2445.
7. Reynolds AB, Daniel J, McCrea PD, Wheelock MJ, Wu J, Zhang Z.Identification of a new catenin: the tyrosine kinase substrate p120casassociates with E-cadherin complexes. Mol Cell Biol 1994; 14: 8333–8342.
8. Shibamoto S, Hayakawa M, Takeuchi K, et al. Association of p120, atyrosine kinase substrate, with E-cadherin/catenin complexes. J Cell Biol1995; 128: 949–957.
9. Watabe M, Nagafuchi A, Tsukita S, Takeichi M. Induction of polarizedcell–cell association and retardation of growth by activation of theE-cadherin–catenin adhesion system in a dispersed carcinoma line. J CellBiol 1994; 127: 247–256.
10. Hermiston ML, Gordon JI. Inflammatory bowel disease and adenomas inmice expressing a dominant negative N-cadherin. Science 1995; 270: 1203–1207.
11. Hermiston ML, Wong MH, Gordon JI. Forced expression of E-cadherin inthe mouse intestinal epithelium slows cell migration and provides evidencefor nonautonomous regulation of cell fate in a self-renewing system. GenesDev 1996; 10: 985–996.
12. Hermiston ML, Gordon JI. In vivo analysis of cadherin function in themouse intestinal epithelium: essential roles in adhesion, maintenance ofdifferentiation, and regulation of programmed cell death. J Cell Biol 1995;129: 489–506.
? 1998 John Wiley & Sons, Ltd.
13. Dogan A, Wang ZD, Spencer J. E-cadherin expression in intestinal epi-thelium. J Clin Pathol 1995; 48: 143–146.
14. Hanby AM, Chinery R, Poulsom R, Playford RJ, Pignatelli M. Downregu-lation of E-cadherin in the reparative epithelium of the human gastrointes-tinal tract. Am J Pathol 1996; 148: 723–729.
15. Shimoyama Y, Hirohashi S, Hirano S, et al. Cadherin cell adhesionmolecules in human epithelial tissues and carcinomas. Cancer Res 1989; 49:2128–2133.
16. Cattoretti G, Pileri S, Parravicini C, et al. Antigen unmasking on formalin-fixed, paraffin-embedded tissue sections. J Pathol 1993; 171: 83–97.
17. Nagafuchi A, Takeichi M, Tsukita S. The 102 kd cadherin-associatedprotein: similarity to vinculin and posttranscriptional regulation of expres-sion. Cell 1991; 65: 849–857.
18. Pignatelli M, Ansari TW, Gunter P, et al. Loss of membranous E-cadherinexpression in pancreatic cancer: correlation with lymph node metastasis,high grade, and advanced stage. J Pathol 1994; 174: 243–248.
19. Gagliardi G, Kandemir O, Liu D, et al. Changes in E-cadherin immunore-activity in the adenoma–carcinoma sequence of the large bowel. VirchowsArch 1995; 426: 149–154.
20. Syrigos KN, Krausz T, Waxman J, et al. E-cadherin expression in bladdercancer using formalin-fixed, paraffin-embedded tissues: correlation withhistopathological grade, tumour stage and survival. Int J Cancer 1995; 64:367–370.
21. Shiozaki H, Iihara K, Oka H, et al. Immunohistochemical detection ofá-catenin expression in human cancers. Am J Pathol 1994; 144: 667–674.
22. Kadowaki T, Shiozaki H, Inoue M, et al. E-cadherin and á-cateninexpression in human esophageal cancer. Cancer Res 1994; 54: 291–296.
23. Matsui S, Shiozaki H, Inoue M, et al. Immunohistochemical evaluation ofalpha-catenin expression in human gastric cancer. Virchows Arch 1994; 424:375–381.
24. Daniel JM, Reynolds AB. The tyrosine kinase substrate p120cas bindsdirectly to E-cadherin but not to the adenomatous polyposis coli protein orá-catenin. Mol Cell Biol 1995; 15: 4819–4824.
25. Downing JR, Reynolds AB. PDGF, CSF-1, and EGF induce tyrosinephosphorylation of p120, a pp60src transformation-associated substrate.Oncogene 1991; 6: 607–613.
26. Shibamoto S, Hayakawa M, Takeuchi K, et al. Tyrosine phosphorylation ofâ-catenin and plakoglobin enhanced by hepatocyte growth factor andepidermal growth factor in human carcinoma cells. Cell Adhes Commun1994; 1: 295–305.
27. Dignass AU, Podolsky DK. Cytokine modulation of intestinal epithelial cellrestitution: central role of transforming growth factor â. Gastroenterology1993; 105: 1323–1332.
28. Dignass AU, Lynch-Devaney K, Podolsky DK. Hepatocyte growth factor/scatter factor modulates intestinal epithelial cell proliferation and migra-tion. Biochem Biophys Res Commun 1994; 202: 701–709.
29. Dignass AU, Tsunekawa S, Podolsky DK. Fibroblast growth factorsmodulate intestinal epithelial cell growth and migration. Gastroenterology1994; 106: 1254–1262.
J. Pathol. 185: 413–418 (1998)