expression of adhesion molecules by lp (a): a potential novel
TRANSCRIPT

17650892-6638/98/0012-1765/$02.25 Q FASEB
/ 383c de21 Mp 1765 Wednesday Nov 04 09:25 AM LP–FASEB de21
Expression of adhesion molecules by Lp(a): a potentialnovel mechanism for its atherogenicity
SEAN ALLEN, SHABEENA KHAN, SHUI-PANG TAM,* MARLYS KOSCHINSKY,*PATRICIA TAYLOR, AND MAGDI YACOUB1
Department of Cardiothoracic Surgery, Imperial College of Science, Technology and Medicine, RoyalBrompton and Harefield NHS Trust Hospital, Harefield Hospital, Harefield, Middlesex UB9 6JH,United Kingdom; and *Department of Biochemistry, Queens University, Kingston, Ontario, Canada
ABSTRACT Lp(a) is a major inherited risk factorfor premature atherosclerosis. The mechanism ofLp(a) atherogenicity has not been elucidated, butlikely involves both its ability to interfere with plas-minogen activation and its atherogenic potential as alipoprotein particle after receptor-mediated uptake.We demonstrate that Lp(a) stimulates production ofvascular cell adhesion molecule 1 (VCAM-1) and E-selectin in cultured human coronary artery endothe-lial cells (HCAEC). This effect resulted from a risein intracellular free calcium induced by Lp(a) andcould be inhibited by the intracellular calcium che-lator, BAPTA/AM. The involvement of the LDL andVLDL receptors in Lp(a) activation of HCAEC wereruled out since Lp(a) induction of adhesion mole-cules was not prevented by an antibody (IgGC7) tothe LDL receptor or by receptor-activating protein,an antagonist of ligand binding to the VLDL recep-tor. Addition of a2-macroglobulin as well as treat-ment with heparinase, chondroitinase ABC, andsodium chlorate did not decrease levels of VCAM-1and E-selectin stimulated by Lp(a), suggesting thatneither the low density lipoprotein receptor-relatedprotein nor cell-surface proteoglycans are involved inLp(a)-induced adhesion molecule production. Nei-ther does the binding site on HCAEC responsible foradhesion molecule production by Lp(a) appear to in-volve plasminogen receptors, as levels of VCAM-1and E-selectin were not significantly decreased by theaddition of glu-plasminogen, the lysine analog e-ami-nocaproic acid, or by trans-4-(aminomethyl)-cyclo-hexanecarboxymethylic acid (tranexamic acid), whichacts by binding to the lysine binding sites carried onthe kringle structures in plasminogen. In contrast, re-combinant apolipoprotein (a) [r-apo(a)] competedwith Lp(a) and attenuated the expression of VCAM-1 and E-selectin. In summary, we have identified acalcium-dependent interaction of Lp(a) with HCAECcapable of inducing potent surface expression ofVCAM-1 and E-selectin that does not appear to in-volve any of the known potential Lp(a) binding sites.Because leukocyte recruitment to the vessel wall ap-pears to represent one of the important early eventsin atherogenesis, this newly described endothelial
cell-activating effect of Lp(a) places it at a crucialjuncture in the initiation of atherogenic disease andmay lead to a better understanding of the role ofLp(a) in the vascular biology of atherosclerosis.—Al-len, S., Khan, S., Tam, S.-P., Koschinsky, M., Taylor,P., Yacoub, M. Expression of adhesion molecules byLp(a): a potential novel mechanism for its atherogen-icity. FASEB J. 12, 1765–1776 (1998)
Key Words: calcium signaling · VCAM-1 · E-selectin · en-dothelial cells
LIPOPROTEIN (a) [Lp(a)]2 is a major inherited riskfactor associated with coronary atherosclerosis, earlysaphenous vein bypass occlusion, and the acceleratedcoronary atherosclerosis of cardiac transplants (1, 2)The mechanism of Lp(a) atherogenicity has not beenelucidated, but likely involves both its ability to inter-fere with plasminogen activation as well as its athero-genic potential as a lipoprotein particle after recep-tor-mediated uptake.
The Lp(a) particle closely resembles low densitylipoprotein (LDL) but is distinguishable by the pres-ence of apolipoprotein (a) apo(a), which is cova-lently linked to the apolipoprotein B100 (apoB100)of LDL by a single disulfide bridge (3). Apolipopro-tein (a) [apo(a)] has extensive structural homologyto the fibrinolytic pre-enzyme plasminogen (4). Bymolecular mimicry, Lp(a) can compete with glu-plas-minogen for the annexin II plasminogen binding do-main on the cell surface (5) and therefore directlyinfluence endothelial cell plasmin generation. By in-
1 Correspondence: FRCS, Department of CardiothoracicSurgery, Imperial College of Science, Technology and Medi-cine, Heart Science Centre, Harefield Hospital, Hill EndRoad, Harefield, Middlesex UB9 6JH, U.K.
2 Abbreviations: ANOVA, analysis of variance; e-APA, e-ami-nocaproic acid; apo(a), apolipoprotein (a); BHT, butylatedhydroxytoluene; HCAEC, human coronary artery endothelialcells; LDL, low density lipoprotein; Lp(a), lipoprotein (a);LRP, LDL receptor-related protein; a2MR, a2-macroglobulinreceptor; RAP, receptor-associated protein; TNF, tumor ne-crosis factor; VCAM-1, vascular cell adhesion molecule 1;VLDL, very low density lipoprotein.

1766 Vol. 12 December 1998 The FASEB Journal ALLEN ET AL.
/ 383c de21 Mp 1766 Wednesday Nov 04 09:25 AM LP–FASEB de21
terfering with the fibrinolytic pathway via direct com-petition with plasminogen binding, Lp(a) down-reg-ulates endothelial cell plasmin generation and shiftsthe vessel surface to a more thrombogenic pheno-type. In addition, Lp(a) may act as a competitive in-hibitor of tissue plasminogen activator in the pres-ence of fibrinogen and also interferes with thebinding of plasminogen to fibrin clots (6). This in-teraction also inhibits TGFb activation (7), resultingin increased smooth muscle cell proliferation and mi-gration. Both inhibition of clot lysis and enhance-ment of cell migration could contribute to the pro-cess of atherogenesis.
By virtue of its similarity with LDL, it has been pro-posed that Lp(a) can effectively compete with LDLfor binding and/or uptake by the LDL receptor. Inaddition to the LDL receptor, several other membersof the LDL receptor family have been identified andmay function in Lp(a) catabolism. The LDL receptor-related protein/a2-macroglobulin receptor (LRP/a2MR) is a recently described multifunctional recep-tor that binds to apoE-enriched lipoproteins (8, 9),as well as to nonlipoprotein ligands such as activateda2-macroglobulin (10) and type-1 plasminogen acti-vator inhibitor/urokinase plasminogen activator(PAI-1/uPA) complexes (11). Owing to its structuralsimilarity with the LDL receptor, a role for the LRPin Lp(a) binding has been postulated; recent evi-dence suggests that LRP may bind to a high molec-ular weight isoform of Lp(a) (12). Recent data havealso suggested that the very low density lipoprotein(VLDL) receptor can bind and internalize Lp(a)(13). All LDL receptor family members bind a 39 kDareceptor-associated protein (RAP) that antagonizesligand binding (14–17), although binding of RAP tothe LDL receptor occurs with low affinity.
Lp(a) also appears to modulate vascular function.Lp(a) has been demonstrated in several acute inflam-matory and vasculopathic states, including Crohn’sdisease, granulomatous lymph nodes, pericarditis,and thrombotic and thrombocytopenic purpura (5).The deposition of Lp(a) can further modify the stateof vascular activation by decreasing production of en-dothelium-derived growth factor (18) and release ofplasminogen activator inhibitor 1 (19).
One of the earliest signs of endothelial cell activa-tion is a rise in intracellular calcium. We have re-cently shown that LDL binding to the LDL receptorcan induce a rise in intracellular calcium in humanendothelial cells, which triggers the induction of lowlevels of vascular cell adhesion molecule 1 (VCAM-1)and E-selectin (20). These adhesion molecules canincrease the binding of leukocytes to the vascular en-dothelium, which is thought to be an important fac-tor in the early development of atherosclerosis. It ispossible that Lp(a) could also induce adhesion mol-ecules and influence atherogenesis because it sharesglobal homology with LDL. We initiated studies to
investigate the role of the LDL receptor family mem-bers in the activation of cultured human coronaryartery endothelial cells (HCAEC) by Lp(a) and a re-combinant form of apo(a) [r-apo(a)]. Our resultshave identified a calcium-dependent interaction ofLp(a) with HCAEC that does not appear to involveany of the known potential Lp(a) binding sites thatinduce potent surface expression of VCAM-1 and E-selectin.
MATERIALS AND METHODS
Reagents
Endothelial serum-free growth medium and medium 199were purchased from Gibco-BRL (Paisley, U.K.). The fura-2/AM and 1,-2-Bis(o-aminophenoxy)ethane-N,N,N*,N*-tetra-acetic acetomethyl ester (BAPTA/AM) were purchased fromMolecular Probes (Eugene, Oreg.). Plasminogen, trans-4-(aminomethyl)-cyclohexanecarboxymethylic acid (tranex-amic acid), e-aminocaproic acid (e-APA), heparinase III(H8891), chondroitinase ABC (C2905), sodium chlorate, anda2-macroglobulin were purchased from Sigma Chemicals(Poole, U.K.). The LDL receptor antibody IgGC7 was pur-chased from Amersham International (Amersham, U.K.). Hu-man VCAM-1 [clone BBA 6] and E-selectin [clone BBA 2]antibodies were purchased from R&D systems (Oxford, U.K.).Simvastatin, the Sandoz compound 58–035, and r-apo (a)were kindly provided by Dr. Tam and Dr. Koschinsky, QueensUniversity, Canada. RAP was kindly provided by Dr. Gonias,University of Charlottesville,Virginia, and by Dr. Owen, Uni-versity Department of Medicine, The Royal Free HospitalSchool of Medicine, Rowland Hill St., London. All other rea-gents were analar grade and purchased from BDH Chemicals,Poole, U.K.
Purification of proteins
Lipoprotein (a) and LDL (Sigma Chemicals) were isolatedsequentially from plasma under sterile conditions to avoid en-dotoxin contamination as previously described (20). Briefly,Lp(a) was in a solution of 10 mM Tris-HCl containing 1 mg/ml Na2 EDTA, pH 7.2, and LDL was extensively dialyzedagainst 0.15 M NaCl, 0.01% EDTA, pH 7.4–7.5. Both lipopro-teins were checked for their characteristic electrophoretic mo-bility and chemical composition. The lipoproteins were essen-tially free from contamination by other lipoproteins asdetermined by cellulose acetate electrophoresis using fat red7B staining for lipid. Our own sodium dodecyl sulfate-poly-acrylamide gel electrophoresis analysis of Lp(a) confirmed itspurity. Lipoproteins were stored at 47C and used within 2 wkafter isolation. A recombinant form of apolipoprotein (a)[r-apo(a)] containing 17 kringle IV domains, as well as thekringle V and protease domains, was purified from condi-tioned media harvested from stably transfected human em-bryonic kidney cells by lysine-Sepharose affinity chromatog-raphy, as previously described (21).
Cell culture
The HCAEC were cultured in medium 199 containing 10%human AB serum, 10% fetal calf serum, 2 mM L-glutamine,penicillin (100 units/ml), and streptomycin (100 mg/ml) aspreviously described (20). Cells were grown to confluency in24-well tissue culture plates in serum-supplemented media; be-

Lp(a) INDUCES ADHESION MOLECULES IN ENDOTHELIAL CELLS 1767
/ 383c de21 Mp 1767 Wednesday Nov 04 09:25 AM LP–FASEB de21
fore each experiment, cells were preconditioned by an 24 hincubation in serum-free media plus various supplements, asindicated. To down-regulate the LDL receptor, some experi-ments were performed in HCAEC that were cultured in se-rum-containing medium. Cell viability was routinely moni-tored by trypan blue exclusion. In all experiments, thenumber of nonviable cells never exceeded 5% of the total cellnumber and there was no difference in viability between cellsgrown in the presence or absence of serum.
Calcium measurements
The HCAEC were plated on glass coverslips at 105 cells/ml for18–24 h and then loaded with fura-2/AM (1 mM for 30 minat room temperature) in Krebs-HEPES medium (pH 7.4) con-taining 120 mM NaCl, 1.3 mM CaCl2, 1.2 mM MgSO4, 4.8 mMKCl, 1.2 mM KH2PO4, 25 mM HEPES, 25 mM glucose, and0.1% bovine serum albumin. The cells were then washed twicewith Krebs-HEPES and left for 15 min to allow complete hy-drolysis of the fura-2 ester. In experiments involving phar-macological agents, endothelial cells were preincubated for agiven time and temperature (as described) with the agent be-fore the endothelial cells were loaded with fura-2/AM. Thecoverslips were then secured between two plates of a 3 ml-volume coverslip holder, mounted in a temperature-con-trolled incubation holder (347C), and placed onto a micro-scope stage of a Zeiss Axiovert 35 inverted epifluorescencemicroscope. Intracellular calcium measurements were per-formed on individual cells using an ionVision dual excitationsystem (ImproVision, Coventry, U.K.). Absolute calcium levelswere calculated as previously described (20).
Flow cytometry analysis
Changes in VCAM-1 and E-selectin protein in control (TNFa10 ng/ml, positive regulator of these adhesion molecules) andLp(a)-treated endothelial cell cultures were determined usinga Flow Cytometer. Endothelial cells were passaged into 24-wellplates, allowed to grow to confluence, transferred to serum-free medium for 24 h and preincubated with Lp(a) (10 to 400mg/ml) or normal cell culture media for 5 min, 30 min, 5 h,and 12 h. In experiments to investigate whether Lp(a)-in-duced changes in intracellular calcium correlate with adhe-sion molecule expression, endothelial cells were preincubatedwith the intracellular calcium chelator BAPTA/AM (5 mM; 30min) before the addition of Lp(a). At the end of each Lp(a)incubation time, cells were left for 5 h, at which time controlwells [no Lp(a)] were treated with tumor necrosis factor a(TNFa) for 5 h. At the end of this period the cells werewashed, trypsinized, and assayed for adhesion molecule ex-pression. Expression of cell-surface molecules was measuredas fluorescence intensity by use of an EPICS XL-MCL FlowCytometer (Coulter Electronics). Each sample counted 5 1103 cells. Controls for each assay included the absence of theprimary antibody and, in some cases, incubation of cells withan isotype-matched, irrelevant antibody.
Role of the LDL receptor
At 24 h prior to the experiment, the medium of the HCAECcultures was replaced with serum-free, lipoprotein-deficientmedium. In some experiments, the cells were preincubatedovernight in serum free medium containing one of the follow-ing supplements—Simvastatin (0.1 to 10 mM), the Sandozcompound 58–035 (5mg/ml), or serum-containingmedium—before the addition of Lp(a). To block the LDL receptor, cellswere preincubated for 30 min at 377C with either the mono-clonal anti-LDL receptor antibody IgGC7 (22) at a concentra-
tion of 50 mg/ml or an irrelevant IgG antibody as a control,before Lp(a) was added. Other cells were treated with a com-bination of Lp(a){ LDL or LP(a)/ r-apo(a) for competitionstudies, or treated with different concentrations of r-apo(a)(10 to 100 mg/ml) alone.
LRP/a2MR and VLDL receptor interactions with Lp(a)
To investigate whether Lp(a) was interacting with other LDLreceptor family members such as the LRP/a2MR and VLDLreceptor, we incubated HCAEC with RAP (100 nM), a mole-cule that binds to these receptors and antagonizes ligand bind-ing (14–17) before the addition of the Lp(a). Coincubationof cells with activated a2-macroglobulin (10 mM) and 100 mg/ml Lp(a) was also performed in order to determine whetheror not competitive binding to the LRPa2MR receptor couldblock Lp(a) activation of HCAEC.
Plasminogen and Lp(a) interactions
The role of plasminogen binding sites was also determined.Before the addition of Lp(a), HCAEC were incubated in se-rum-free media containing either 50 mM e-ACA or 10 mMtranexamic acid for 30 min. Competition experiments wereperformed by the simultaneous addition of 100 mg/ml Lp(a)and glu-plasminogen (0.1 to 1 mM) to each multiwell.
Role of proteoglycan binding sites in Lp(a) function
Experiments were also performed to evaluate proteoglycanbinding sites on HCAEC. The HCAEC were cultured as de-scribed above. Conditions for the use of heparinase III (SigmaH8891) and chondroitinase ABC (Sigma C 2905) were basedon the procedures described by Ji et al. (23) and Oike et al.(24), respectively. Briefly, the cells were washed four times at377C in phosphate-buffered saline and then incubated for 2 hat 377C with either heparinase (2 units/ml) or chondroitinase(0.25 units/ml) in serum-free medium. The cells were washedan additional four times with phosphate-buffered saline at377C and Lp(a) was added as described above. In some ex-periments, the cells were preincubated for 48 h with 30 mMsodium chlorate prior to cell activation with Lp(a).
Immunofluoresence
Lipoprotein (a) (100 mg/ml) or TNFa (10 ng/ml was addedto HCAEC grown on glass coverslips 12–18 h previously. Somecoverslips were preincubated with BAPTA/AM (5 M for 30min) before cells were treated with Lp(a). Coverslips werewashed after 5 h and fixed in 1% (v/v) para-formaldehyde. E-selectin and VCAM-1 were visualized by indirect immunoflu-orescence using either a monoclonal anti E-selectin (cloneBBA2) or anti-VCAM-1 antibody (clone BBA 6), followed bybiotinylated rabbit antimouse IgG and then Streptavidin-Texas red. Cells were visualized with a Laser-Scanning Con-focal Microscope (Noran Instruments, U.K.).
Statistical analysis
Statistical analysis were carried out using the commercial pro-gram Unistat version 4 for Windows (Unistat Ltd, U.K.). Theresults are expressed as mean { SEM. Calcium measurementswere analyzed by an unpaired Student’s t test when appropri-ate. Adhesion molecule data was analyzed using repeated mea-sures ANOVA (analysis of variance) coupled with a post hocBonferroni correction analysis. Differences were consideredto be significant when the probability value was õ0.05.

1768 Vol. 12 December 1998 The FASEB Journal ALLEN ET AL.
/ 383c de21 Mp 1768 Wednesday Nov 04 09:25 AM LP–FASEB de21
Figure 1. a) [Ca2/]i changes after addition of Lp(a) to HCAECbathed in Krebs-HEPES. Lp(a) (100 mg/ml) added at 4 s (A)and paired images were acquired at 4 s intervals (B–I). Imageswere processed as indicated in Material and Methods; calciumchanges are color coded (color bar) such that warm colorsindicate high calcium. A representative experiment of at least10 cell preparations obtained from three donor coronary ar-teries. b) Concentration-dependent effect of Lp(a) added toHCAEC on [Ca2/]i. The values represent mean { SEM of 50individual HCAEC obtained from three donor coronary ar-teries. c) Time course showing continuous recording of[Ca2/]i after addition of 100 mg/ml Lp(a) to HCAEC cultures.The values represent the change in [Ca2/]i of four individualcells. Note the oscillations in [Ca2/]i. (Figure 1C is on next page.)

Lp(a) INDUCES ADHESION MOLECULES IN ENDOTHELIAL CELLS 1769
/ 383c de21 Mp 1769 Wednesday Nov 04 09:25 AM LP–FASEB de21
Figure 1. (Continued)
RESULTS
Lp(a) induces calcium transients in HCAEC
To investigate the effects of Lp(a) on endothelial cellcalcium, we subjected fura-2/AM loaded, culturedHCAEC to Lp(a). Addition of Lp(a) (100 m/ml) toHCAEC adherent on glass coverslips and bathed inKrebs-HEPES buffer caused a transient calcium risefrom a resting level of 78 { 12.4 nM to 524 { 127nM (mean{SEM, nÅ19) (Fig. 1a). The calcium tran-sients in HCAEC were dependent on the concentra-tion of Lp(a) and were saturable (Fig. 1b). Further-more, the responses of the cells to Lp(a) wereheterogeneous and asynchronous, with some endo-thelial cells not responding and some exhibiting os-cillations in calcium transients (Fig. 1c). Lp(a)-in-duced calcium signaling in HCAEC cultured in thepresence of serum (500{80 nM; mean{SEM, nÅ12)was not significantly different from that triggered inthe absence of serum. Elevation of intracellular cal-cium by Lp(a) was not a sole characteristic ofHCAEC, as both human aortic and human heart mi-crovascular endothelial cells also responded to Lp(a)with a rise in intracellular calcium (data not shown).The average calcium response induced by the cal-cium ionophore A23187 in HCAEC was 7.0 { 3 mM,which is 13-fold greater than that triggered by Lp(a).
Lp(a) induces VCAM-1 and E-selectin expression onHCAEC
Further studies were performed to elucidate the na-ture of the rise in intracellular calcium by Lp(a). In-cubation of endothelial cells with Lp(a) (10–400 mg/ml) resulted in a dose-dependent increase in VCAM-1and E-selectin (Fig. 2A, B) expression, which stabi-lized after 100 mg/ml Lp(a). In all experiments Lp(a)at a concentration of 100 mg/ml induced significantly
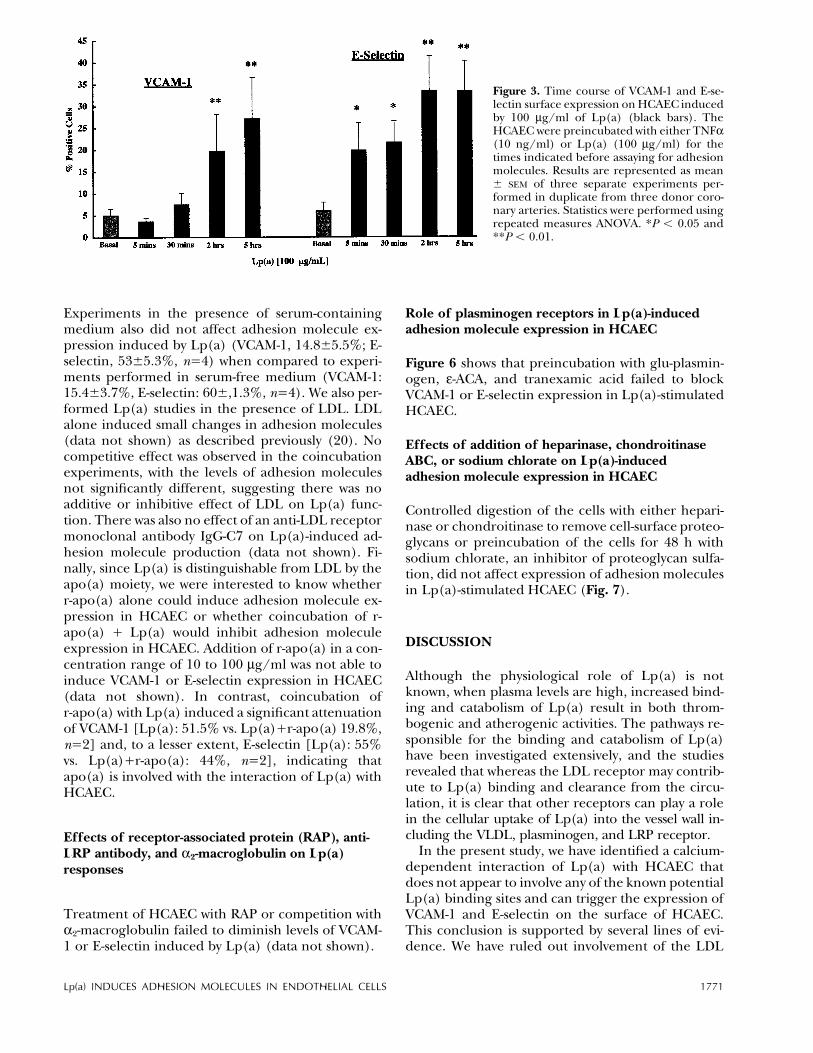
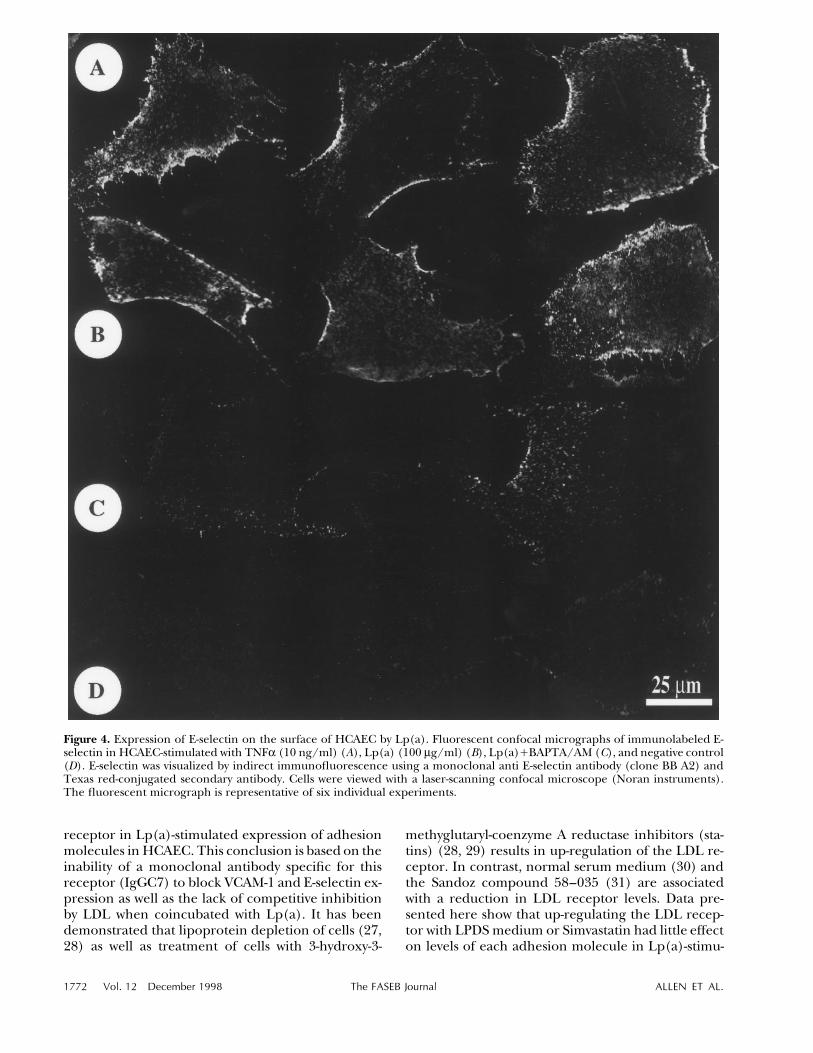
more E-selectin than VCAM-1 expression: 61 { 8%vs. 35 { 10%, P õ 0.01, respectively. The expressionof VCAM-1 and E-selectin was inhibited in theHCAEC that were preloaded with the intracellularcalcium chelator BAPTA/AM prior to Lp(a) treat-ment (Fig. 2A, B), suggesting that calcium transientsare involved in Lp(a)-induced adhesion moleculeproduction. However, as BAPTA also has IP3 antag-onist properties, we cannot rule out that IP3 antago-nism with BAPTA, which would result in the inhibi-tion of release of calcium, could also contribute tothe effect observed. Figure 3 shows the effects of ex-posure to Lp(a) (100 mg/ml) over time. Both VCAM-1 and E-selectin showed similar kinetics, with signifi-cant levels of each protein occurring after 2 hincubation with Lp(a) and peaking at 5 h. Indirectimmunofluorescence confirmed the flow cytometrydata, showing VCAM-1 (data not shown) and E-selec-tin (Fig. 4) surface staining induced by TNFa andLp(a), which was inhibited by treating the HCAECwith BAPTA/AM.
It seems unlikely that the effects of Lp(a) are dueto the mild oxidation of the lipoprotein since exper-iments performed in the presence of 20 mmol/l bu-tylated hydroxytoluene (BHT), which is known toeliminate nonspecific oxidation of lipoprotein parti-cles during endothelial cell culture experiments (25,26), had no significant effect on the levels of adhe-sion molecules expressed in the HCAEC comparedto non-BHT treated cells (data not shown).
Effects of up- and down-regulation of the LDLreceptor as well as an anti-LDL receptor antibodyon Lp(a)-induced adhesion molecule expression
Figure 5 demonstrates that Lp(a)-induced adhesionmolecule expression of both VCAM-1 and E-selectindo not alter significantly with up- and down-regula-tion of the LDL receptor activity induced by Simvas-tatin or the Sandoz compound 58–035, respectively.

1770 Vol. 12 December 1998 The FASEB Journal ALLEN ET AL.
/ 383c de21 Mp 1770 Wednesday Nov 04 09:25 AM LP–FASEB de21
Figure 2. A) VCAM-1 surface expression on HCAEC induced by increasing concentrations of Lp(a) (black bars). The HCAECwere preincubated with either TNFa (10 ng/ml) or increasing concentrations of Lp(a) (10 to 400 mg/ml) for 5 h before assayingfor VCAM-1. Some cells were pretreated with BAPTA/AM (5 mM for 30 min) before Lp(a) was added (stippled bars). Resultsare represented as mean { SEM of three separate experiments performed in duplicate from three donor coronary arteries.Statistics were performed using repeated-measures ANOVA. *P õ 0.05, **P õ 0.01, and ***P õ 0.001. B) Bar graph shows E-selectin surface expression on HCAEC induced by increasing concentrations of Lp(a) (black bars). The HCAEC were prein-cubated with either TNFa (10 ng/ml) or increasing concentrations of Lp(a) (10 to 400 mg/ml) for 5 h before assaying forE-selectin. Some cells were pretreated with BAPTA/AM (5 mM for 30 min) before Lp(a) was added (stippled bars). Results arerepresented as mean { SEM of three separate experiments performed in duplicate from three donor coronary arteries. Statisticswere performed using repeated measures ANOVA. *P õ 0.05, **P õ 0.01, and ***P õ 0.001.
Lp(a) INDUCES ADHESION MOLECULES IN ENDOTHELIAL CELLS 1771
/ 383c de21 Mp 1771 Wednesday Nov 04 09:25 AM LP–FASEB de21
Figure 3. Time course of VCAM-1 and E-se-lectin surface expression on HCAEC inducedby 100 mg/ml of Lp(a) (black bars). TheHCAEC were preincubated with either TNFa(10 ng/ml) or Lp(a) (100 mg/ml) for thetimes indicated before assaying for adhesionmolecules. Results are represented as mean{ SEM of three separate experiments per-formed in duplicate from three donor coro-nary arteries. Statistics were performed usingrepeated measures ANOVA. *P õ 0.05 and**P õ 0.01.
Experiments in the presence of serum-containingmedium also did not affect adhesion molecule ex-pression induced by Lp(a) (VCAM-1, 14.8{5.5%; E-selectin, 53{5.3%, nÅ4) when compared to experi-ments performed in serum-free medium (VCAM-1:15.4{3.7%, E-selectin: 60{,1.3%, nÅ4). We also per-formed Lp(a) studies in the presence of LDL. LDLalone induced small changes in adhesion molecules(data not shown) as described previously (20). Nocompetitive effect was observed in the coincubationexperiments, with the levels of adhesion moleculesnot significantly different, suggesting there was noadditive or inhibitive effect of LDL on Lp(a) func-tion. There was also no effect of an anti-LDL receptormonoclonal antibody IgG-C7 on Lp(a)-induced ad-hesion molecule production (data not shown). Fi-nally, since Lp(a) is distinguishable from LDL by theapo(a) moiety, we were interested to know whetherr-apo(a) alone could induce adhesion molecule ex-pression in HCAEC or whether coincubation of r-apo(a) / Lp(a) would inhibit adhesion moleculeexpression in HCAEC. Addition of r-apo(a) in a con-centration range of 10 to 100 mg/ml was not able toinduce VCAM-1 or E-selectin expression in HCAEC(data not shown). In contrast, coincubation ofr-apo(a) with Lp(a) induced a significant attenuationof VCAM-1 [Lp(a): 51.5% vs. Lp(a)/r-apo(a) 19.8%,nÅ2] and, to a lesser extent, E-selectin [Lp(a): 55%vs. Lp(a)/r-apo(a): 44%, nÅ2], indicating thatapo(a) is involved with the interaction of Lp(a) withHCAEC.
Effects of receptor-associated protein (RAP), anti-LRP antibody, and a2-macroglobulin on Lp(a)responses
Treatment of HCAEC with RAP or competition witha2-macroglobulin failed to diminish levels of VCAM-1 or E-selectin induced by Lp(a) (data not shown).
Role of plasminogen receptors in Lp(a)-inducedadhesion molecule expression in HCAEC
Figure 6 shows that preincubation with glu-plasmin-ogen, e-ACA, and tranexamic acid failed to blockVCAM-1 or E-selectin expression in Lp(a)-stimulatedHCAEC.
Effects of addition of heparinase, chondroitinaseABC, or sodium chlorate on Lp(a)-inducedadhesion molecule expression in HCAEC
Controlled digestion of the cells with either hepari-nase or chondroitinase to remove cell-surface proteo-glycans or preincubation of the cells for 48 h withsodium chlorate, an inhibitor of proteoglycan sulfa-tion, did not affect expression of adhesion moleculesin Lp(a)-stimulated HCAEC (Fig. 7).
DISCUSSION
Although the physiological role of Lp(a) is notknown, when plasma levels are high, increased bind-ing and catabolism of Lp(a) result in both throm-bogenic and atherogenic activities. The pathways re-sponsible for the binding and catabolism of Lp(a)have been investigated extensively, and the studiesrevealed that whereas the LDL receptor may contrib-ute to Lp(a) binding and clearance from the circu-lation, it is clear that other receptors can play a rolein the cellular uptake of Lp(a) into the vessel wall in-cluding the VLDL, plasminogen, and LRP receptor.
In the present study, we have identified a calcium-dependent interaction of Lp(a) with HCAEC thatdoes not appear to involve any of the known potentialLp(a) binding sites and can trigger the expression ofVCAM-1 and E-selectin on the surface of HCAEC.This conclusion is supported by several lines of evi-dence. We have ruled out involvement of the LDL
1772 Vol. 12 December 1998 The FASEB Journal ALLEN ET AL.
/ 383c de21 Mp 1772 Wednesday Nov 04 09:25 AM LP–FASEB de21
Figure 4. Expression of E-selectin on the surface of HCAEC by Lp(a). Fluorescent confocal micrographs of immunolabeled E-selectin in HCAEC-stimulated with TNFa (10 ng/ml) (A), Lp(a) (100 mg/ml) (B), Lp(a)/BAPTA/AM (C), and negative control(D). E-selectin was visualized by indirect immunofluorescence using a monoclonal anti E-selectin antibody (clone BB A2) andTexas red-conjugated secondary antibody. Cells were viewed with a laser-scanning confocal microscope (Noran instruments).The fluorescent micrograph is representative of six individual experiments.
receptor in Lp(a)-stimulated expression of adhesionmolecules in HCAEC. This conclusion is based on theinability of a monoclonal antibody specific for thisreceptor (IgGC7) to block VCAM-1 and E-selectin ex-pression as well as the lack of competitive inhibitionby LDL when coincubated with Lp(a). It has beendemonstrated that lipoprotein depletion of cells (27,28) as well as treatment of cells with 3-hydroxy-3-
methyglutaryl-coenzyme A reductase inhibitors (sta-tins) (28, 29) results in up-regulation of the LDL re-ceptor. In contrast, normal serum medium (30) andthe Sandoz compound 58–035 (31) are associatedwith a reduction in LDL receptor levels. Data pre-sented here show that up-regulating the LDL recep-tor with LPDS medium or Simvastatin had little effecton levels of each adhesion molecule in Lp(a)-stimu-

Lp(a) INDUCES ADHESION MOLECULES IN ENDOTHELIAL CELLS 1773
/ 383c de21 Mp 1773 Wednesday Nov 04 09:25 AM LP–FASEB de21
Figure 5. The effect of regulating the expression of the LDLreceptor on VCAM-1 and E-selectin surface expression onHCAEC induced by 100 mg/ml of Lp(a). The HCAEC werepreincubated with either Lp(a) (black bars), Lp(a) plus in-creasing concentrations of Simvastatin from left to right (di-agonal bars) or Lp(a) plus Sandoz compound (vertical lines)for the times indicated in the methods before assaying foradhesion molecules. Results are represented as mean{SEM ofthree separate experiments performed in duplicate fromthree donor coronary arteries. Statistics were performed usingrepeated measures ANOVA. **P õ 0.01 and ***P õ 0.001.
Figure 6. The effect of regulating plasminogen binding siteson VCAM-1 and E-selectin surface expression on HCAEC in-duced by 100 mg/ml of Lp(a). The HCAEC were preincubatedwith either Lp(a) (black bars), Lp(a) / glu-plasminogen (di-agonal bars), Lp(a) / e-APA (vertical lines) or Lp(a) / tra-nexamic acid (horizontal bars) for the times indicated inMaterials and Methods before assaying for adhesion mole-cules. Results are represented as mean{ SEM of three separateexperiments performed in duplicate from three donor coro-nary arteries. Statistics were performed using repeated mea-sures ANOVA. **P õ 0.01 and ***P õ 0.001.
Figure 7. Bar-graph shows the effect of removal of proteogly-can binding sites from HCAEC on VCAM-1 and E-selectin ex-pression induced by 100 mg/ml of Lp(a). The HCAEC werepreincubated with either Lp(a) (black bars), Lp(a) / Hepar-inase (diagonal bars), Lp(a) / Chondroinase ABC (verticalbars), or Lp(a) / sodium chlorate (horizontal bars) for thetimes indicated in the methods before assaying for adhesionmolecules. Results are represented as mean { SEM of threeseparate experiments performed in duplicate from three do-nor coronary arteries. Statistics were performed using re-peated-measures ANOVA. **P õ 0.01 and ***P õ 0.001.
lated HCAEC. Similarly, down-regulation of the LDLreceptor with the Sandoz compound 58–035 or cul-turing cells in normal serum-containing medium hadno significant effects on the levels of VCAM-1 and E-selectin induced by LDL or Lp(a).
The presence of serum in the medium of these ex-periments reinforces the point that Lp(a) effects arespecific and not due to artifacts such as oxidation ortoxicity. While it is possible that in the absence ofserum, fatty acids can be hydrolyzed from LDL andachieve concentrations that are toxic to the endothe-lial cells, it is unlikely that in serum-free conditionsLDL will be hydrolyzed by lipoprotein lipase. Eventhough the endothelial cells can produce this en-zyme, LDL is not a substrate for this enzyme. Theactual substrates are VLDL or chylomicrons, whichwould not be present in the absence of serum. Fur-ther confirmation that serum-free media does not al-ter the function of Lp(a) is indicated by the fact thatLp(a)-induced calcium transients are not signifi-cantly different in the presence or absence of serum-containing medium. Finally, trypan blue staining ofviability of HCAEC was not different between endo-thelial cells grown in serum-containing medium, se-rum-free medium, or incubated with BHT, clearly in-dicating that in our model, no detectable oxidationor toxicity is occurring that would affect the ability ofLp(a) to induce adhesion molecule expression.
A recent study has shown that the interaction ofLp(a) with HepG2 cells can be mediated through theLDL receptor by a ‘hitchhiking’ process. This path-
way appears to involve binding of apo(a) to apoB-containing particles prior to their uptake by the LDLreceptor (32). In the present study, we directly ex-amined the ability of a recombinant form of apo(a)to activate cultured HCAEC. Incubation of HCAECwith increasing concentrations of r-apo(a) alone

1774 Vol. 12 December 1998 The FASEB Journal ALLEN ET AL.
/ 383c de21 Mp 1774 Wednesday Nov 04 09:25 AM LP–FASEB de21
failed to stimulate VCAM-1 or E-selectin expression.In contrast, coincubation of Lp(a)/r-apo(a) pro-duced a significant reduction in levels of VCAM-1 andE-selectin. These results suggest that apo(a) is re-sponsible for mediating the interaction of Lp(a) withthe HCAECs, and also confirms that nonspecific ef-fects of Lp(a) such as oxidation or endotoxin con-tamination are unlikely to be responsible for Lp(a)-induced adhesion molecule expression.
Compared to our previous findings with LDL onadhesion molecule expression in HCAEC (20), Lp(a)is approximately fivefold more potent in inducingVCAM-1 and E-selectin. This suggests that linkage ofapo(a) to apoB could be important for Lp(a) to in-duce greater expression of adhesion molecules com-pared to LDL. Also, unlike LDL, which only inducedtransient calcium rises, Lp(a) frequently induced cal-cium oscillations in the HCAEC. Whether differentcalcium signals produced by LDL and Lp(a) relate tothe different abilities of both lipoproteins to induceVCAM-1 and E-selectin requires further investigation.
We have also provided data to exclude the LRP/a2MR and VLDL receptor as the binding site forLp(a)-induced adhesion molecule production. Weperformed Lp(a) experiments in the presence of ac-tivated a2-macroglobulin. No competition effect wasobserved with the levels of each adhesion moleculeunchanged. In addition, incubation of HCAEC withRAP, a molecule known to inhibit ligand binding toLDL receptor family members such as LRPa2MR andthe VLDL receptor, failed to significantly alter levelsof VCAM-1 or E-selectin in Lp(a)-stimulated HCAEC.These data suggest that adhesion molecule produc-tion after the interaction of Lp(a) with HCAEC is notmediated by the LRP or VLDL receptor.
It is also unlikely that the Lp(a) binding site onHCAEC responsible for VCAM-1 and E-selectin ex-pression represents the plasminogen receptor. Li-poprotein (a) has been previously shown to competewith plasminogen for cellular binding sites, mediatedby lysine affinity sites present in both apo(a) and plas-minogen. We have demonstrated here that plasmin-ogen did not compete with Lp(a) for induction ofVCAM-1 and E-selectin since the levels of adhesionmolecules in HCAEC were unaffected by its presence.Further confirmation that plasminogen binding siteswere not involved in Lp(a) activation of HCAECcame from experiments with the lysine analog e-ACA(33) and tranexamic acid, which acts by binding tothe lysine binding sites carried on the kringle struc-tures in plasminogen (34). Neither inhibitor had aneffect on the levels of each adhesion molecule afterLp(a) treatment.
It has been suggested that lipoprotein lipase en-hances the binding of Lp(a) or lipoproteins to cellsby acting as a bridge between lipoproteins in the me-dium and proteoglycans on the cell surface (35). Nosignificant effect of Lp(a)-induced expression of ad-
hesion molecules was observed in HCAEC eitherwhen the cells were depleted of heparan sulfate byheparinase treatment or when sulfation was inhibitedby the addition of sodium chlorate treatment. Fur-thermore, removal of chondroitin sulfate by chon-droitinase ABC treatment did not seem to influenceLp(a) effects on adhesion molecule levels. Our datademonstrate a lack of involvement of cell-surface gly-cosaminoglycans or proteoglycans in the expressionof VCAM-1 and E-selectin by Lp(a). This is supportedby a previous report that also failed to show a directinteraction between Lp(a) and cell-surface glycosa-minoglycan chains (36). Together, these experi-ments provide evidence indicating that a novel bind-ing site for Lp(a) may mediate the cellular activationand subsequent expression of VCAM-1 and E-selectinin cultured HCAEC. Bottalico et al. (37) showed thatcholesterol loading of macrophages leads to an en-hancement of Lp(a) internalization and degradationvia induction of a specific receptor that has yet to beidentified.
An increased adherence of leukocytes to the en-dothelium appears to be a crucial event in the devel-opment of atherosclerosis. In this context, there is agrowing body of evidence to suggest that modifiedand unmodified LDL molecules can induce adhesionmolecules in vascular endothelial cells. Recently ithas been shown that chylomicrons can induceVCAM-1 and E-selectin expression on endothelialcells, suggesting that the apoB moiety of these lipo-proteins may be important for mediating these ef-fects. The evidence to date suggesting a link betweenlipoprotein induction of adhesion molecules and thedevelopment of atherosclerosis is very important inlight of the recent findings by Hackman et al. (38),who reported that patients with hypercholesterol-emia had increased levels of soluble VCAM-1 and E-selectin compared to control subjects, which the au-thors propose may be a marker for atherosclerosis.
In summary, we have shown here that the expres-sion of VCAM-1 and E-selectin on the surface of cul-tured HCAEC can be induced by Lp(a) in a dose- andtime-dependent manner. The response is triggeredby a rise in intracellular free calcium, and can beblocked by the calcium chelator BAPTA/AM and viacompetition with r-apo(a). Furthermore, substantialadhesion molecule expression by Lp(a) occurs at lev-els of Lp(a) that are considered to constitute a riskfactor for cardiovascular disease (above 300 mg/ml)(39). Despite this risk, the pathogenic mechanism(s)of Lp(a) remains obscure. We suggest that Lp(a) maycontribute to atherogenesis by inducing adhesionmolecules on endothelial cells and thus promotingthe interaction of leukocytes with the vessel wall. Be-cause leukocyte recruitment to the vessel wall appearsto represent one of the important early events in ath-erogenesis, this newly described endothelial cell-ac-tivating effect of Lp(a) places it at a crucial juncture

Lp(a) INDUCES ADHESION MOLECULES IN ENDOTHELIAL CELLS 1775
/ 383c de21 Mp 1775 Wednesday Nov 04 09:25 AM LP–FASEB de21
in the initiation of atherogenic disease and may leadto a better understanding of the role of Lp(a) in thevascular biology of atherosclerosis.
Professor Sir Magdi Yacoub is supported by the British HeartFoundation.
REFERENCES
1. Budde, T., Fechtrup, C., Bosenberg, E., Vielhauer, C., Enberg,A., Schulte, H., Assman, G., and Breithardt, G. (1994) Plasmaand Lp(a) levels correlate with number, severity, and length-extension of coronary arteriography for lesions in male patientsundergoing coronary arteriography for clinically suspected cor-onary atherosclerosis. Arterioscler. Thromb. 14, 1730–1736
2. Barbir, M., Kushwaha, S., Hart, B., Macken, H., Thompson,G. R., Mitchell, A., Robinson, O., and Yacoub, M. (1992) Lipo-protein (a) and accelerated coronary artery disease in cardiactransplant recipients. Lancet 340, 1500–1501
3. Koschinsky, M. L., Cote, G. P., Gabel, B., and van der Hoek,Y. Y. (1993) Identification of the cysteine residue in apolipopro-tein (a) which mediates extracellular coupling with apolipopro-tein B-100. J. Biol. Chem. 268, 19819–19825
4. McLean, J. W., Tomlinson, J. E., Khang, W. J., Eaton, O., Chen,E., Fless, G. M., and Scorli, A. M. (1987) cDNA sequence of hu-man apolipoprotein (a) is homologous to plasminogen. Nature(London) 330, 132–137
5. Nachman, R. L. (1992) Thrombosis and atherogenesis: molec-ular connections. Blood 79, 1897–1906
6. Loscalzo, J., Weinfeld, M., Fless, G. M. and Scanu, A. M. (1990)Lipoprotein (a), fibrin binding and plasminogen activation. Ar-teriosclerosis 10, 240–245
7. Grainger, D. J., Kirschenlohr, H. L., Metcalfe, J. C., Weissberg,P. L., Wade, D. P., and Lawn, R. M. (1993) Proliferation of hu-man smooth muscle cells promoted by lipoprotein (a). Science260, 1655–1658
8. Kowal, R. C., Herz, J., Goldstein, J. L., Esser, V., and Brown,M. S. (1989) Low density lipoprotein receptor-related proteinmediates uptake of cholesteryl esters derived from apoproteinE-enriched lipoproteins. Proc. Natl. Acad. Sci. USA 86, 5810–5814
9. Kowal, R. C., Herz, J., Goldstein, J. L., Esser, V., and Brown, M.S. (1990) Opposing effects of apolipoprotein E and C on lipo-protein binding to low density lipoprotein receptor-related pro-tein. J. Biol. Chem. 265, 10771–10779
10. van Leuven, F., Marynan, P., Scottrup-Jensen, L., Cassiman, J. J.,and van den Berrghe, H. (1986) The receptor-binding domainof human alpha 2-macroglobulin. Isolation after limited prote-olysis with a bacterial proteinase. J. Biol. Chem. 261, 11369–11373
11. Herz, J., Clouthier, D. E., and Hammer, R. E. (1992) LDL re-ceptor-related protein internalizes and degrades uPA-PAI-1 com-plexes and is essential for embryo implantation. Cell 71, 411–421
12. Marz, W., Beckmann, A., Scharnagl, H., Siekmeier, R., Mondorf,U., Held, I., Schneider, W., Pressner, K. T., and Curtis, L. K.(1993) Heterogenous lipoprotein (a) size isoforms differ bytheir interaction with the low density lipoprotein receptor andthe low density lipoprotein receptor-related protein/alpha 2-macroglobulin receptor. FEBS Lett. 325, 271–275
13. McTigue, K., Kozarsky, K. F., Fallon, J. T., Harpel, P. C., andStrickland D. K. (1997) The atherogenic lipoprotein Lp (a) isinternalized and degraded in a process mediated by the VLDLreceptor. J. Clin. Invest. 100, 2170–2181
14. Williams, S. E., Ashcom, J. D., Argraves, W. S., and Strickland,D. K. (1992) A novel mechanism for controlling the activity ofa2-macroglobulin receptor/low density lipoprotein receptor-related protein. Multiple regulatory sites for 39-kDa receptor-associated protein. J. Biol. Chem. 267, 9035–9040
15. Kounnas, M. Z., Argraves, W. S., and Strickland, D. K. (1992)The 39-kDa receptor-associated protein interacts with two mem-bers of the low density lipoprotein-receptor family, a2-macro-
globulin receptor and glycoprotein 330. J. Biol. Chem. 267,21162–21166
16. Battey, F., Gafreis, M. E., Fitzgerald, D. J., Argraves, W. S., Chap-pell, D. A., Strauss, J. F., III, and Strickland, D. K. (1994) The 39kDa receptor associated protein regulates ligand binding by thevery low density lipoprotein receptor. J. Biol. Chem. 269, 23268–23273
17. Medh J. D., Fry, G. L., Bowen, S. L., Pladet, M. W., Strickland,D. K., and Choppell, D. A. (1995) The 39 kDa receptor associ-ated protein modulates lipoprotein catabolism by binding to theLDL receptors. J. Biol. Chem. 270, 536–540
18. Sorenson, K. E., Celermayer, D. S., Georgakaupoulos, D.,Hatcher, G., Betteridge, O. J., and Dearfield, J. (1994) Impair-ment of endothelium-dependent dilation is an early event inchildren with familial hypercholesterolemia and is related to thelipoprotein (a) level. J. Clin. Invest. 93, 50–55
19. Etingin, O., Hajjar, D. P., Hajjar, K. A., Harpel, P. C., and Nach-man, R. L. (1991) Lipoprotein (a) regulates plasminogen acti-vator-1 expression in endothelial cells. J. Biol. Chem. 266, 2459–2465
20. Allen, S. P., Khan, S., Al-Mohanna, F. A., Batten, P. and Yacoub,M. H. (1998) Native low-density induced calcium transients trig-ger VCAM-1 and E-selectin expression in cultured human vas-cular endothelial cells. J. Clin. Invest. In press
21. Sangrar, W., Gabel, B. R., Boffa, M. B., Walker, J. B., Hancock,M. A., Marcovina, S. A., Horrevoets, A. J. G., Nesheim, M. E., andKoschinsky, M. L. (1997) The solution phase interaction be-tween apolipoprotein (a) and plasminogen inhibits the bindingof plasminogen to a plasmin-modified fibrinogen surface. Bio-chemistry 36, 10353–10363
22. Beisiegel, U., Schneider, W. J., Goldstein, J. L., Anderson,R. G. W., and Brown, M. S. (1981) Immunoblot analysis of lowdensity lipoprotein receptors in fibroblasts from subjects withfamilial hypercholesterolemia. J. Biol. Chem. 256, 11923–11931
23. Ji, Z. S., Brecht, W. J., Miranda, R. D., Hussain, M. M., Innerarity,T. L., and Mahley, R. W. (1993) Role of heparan sulphate pro-teoglycans in the binding and uptake of apolipoprotein E-en-riched remnant lipoproteins by cultured cells. J. Biol. Chem.268, 10160–1016724
24. Oike, Y., Kimata, K., Shinomura, T., Nakazawa, K., and Suzuki,S. (1980) Structural analysis of chick-embryo cartilage proteo-glycan by selective degradation with chondroitin lysases (chon-droitinase) and endo-b-D-galactosidase (keratinase). Biochem. J.191, 193–207
25. Pritchard, K. A., Jr., Tota, R. R., Lin, J., Danishefsky, K. J., Kurilla,B. A., Holland, J. A., and Stemerman, B. A. (1991) Native low-density lipoprotein: endothelial cell recruitment of mononu-clear cells. Arterioscler. Thromb. 11, 1175–1181
26. Smalley, D. M., Lin, J. H. C., Curtis, M. L., Kobari, Y., Stemerman,M. B., and Pritchard, C. A., Jr. (1996) Native LDL increases en-dothelial cell adhesiveness by inducing intercellular adhesionmolecule-1. Arterioscler. Thromb. Vasc. Biol. 16, 585–590
27. Goldstein, J. L,. and Brown, M. S. (1990) Regulation of the mev-alonate pathway. Nature (London) 343, 425–430
28. Tam, S. P., Brissette, L., Ramharack, R., and Deeley, R. G. (1991)Differences between the regulation of 3-hydroxy-3-methylglu-taryl-coenzyme A reductase and low density lipoprotein receptorin human hepatoma cells and fibroblasts reside primarily at thetranslational and posttranslational levels. J. Biol. Chem. 266,16764–16773
29. Shepherd, J., Cobbe, S. M., Ford, I., Isles, C. G., Lorimer, A. R.,Macfarlane, P. W., McKillop, J. H., and Packard, C. J. (1995)Prevention of coronary heart disease with pravastatin in menwith hypercholesterolemia. N. Engl. J. Med. 333, 1301–1307
30. Schaefer, H. I. M. P., Hold, K. M., Egas-Kenniphaas, J. M., andvan der Laarse, A. (1993) Intracellular calcium signalling afterbinding of low-density lipoprotein to confluent and noncon-fluent cultures of an endothelial cell line, EA.hy 926. Cell Cal-cium 14, 507–516
31. Havekes, L. M., deWit, E. C. M., and Princen, H. M. G. (1987)Cellular free cholesterol in HepG2 cells is only partially availablefor down-regulation of low-density lipoprotein receptor activity.Biochem. J. 247, 739–746
32. Kostner, G.M. (1993) Interaction of Lp(a) and of apo(a) withliver cells. Arterioscler. Thromb. 13, 1101–1109
33. Hajjar, K. A., Gavish, D., Breslow, J. L. and Nachman, R. L.(1989) Lipoprotein (a) modulation of endothelial cell surface

1776 Vol. 12 December 1998 The FASEB Journal ALLEN ET AL.
/ 383c de21 Mp 1776 Wednesday Nov 04 09:25 AM LP–FASEB de21
fibrinolysis and its potential role in atherosclerosis. Nature (Lon-don) 339, 303–305
34. Saksela, O. (1989) Plasminogen activation and regulation of per-icellular proteolysis. Biochem. Acta 823, 35–65
35. Williams, K. J., Fless, G. M., Petrie, K. A., Snyder, M. L., Brocia,R. W. and Swenson, T. L. (1992) Mechanisms by which lipopro-tein lipase alters cellular metabolism of lipoprotein (a), low den-sity lipoprotein, and nascent lipoproteins. J. Biol. Chem. 267,13284–13292
36. Tam, S. P., Zhang, X., and Koschinsky, M. L. (1996) Interactionof a recombinant form of apolipoprotein (a) with human fibro-blasts and with the human hepatoma cell line HepG2. J. LipidRes. 37, 518–533
37. Bottalico, L. A., Keesler, G. A., Fless, G. M., and Tabas, I. (1993)Cholesterol loading of macrophages leads to marked enhance-ment of native lipoprotein (a) and apoprotein (a) internaliza-tion and degradation. J. Biol. Chem. 268, 8569–8573
38. Hackman, A., Abe, Y., Insull, W., Jr., Pownall, H., Smith, L.,Dunn, K., Gotto, A. M., Jr., and Ballantyne C. M. (1996) Levelsof soluble cell adhesion molecules in patients with dyslipidemia.Circulation 93, 1334–1338
39. Scanu, A. M., and Scandiani, L. (1991) Lipoprotein (a): struc-ture, biology and clinical relevance. Adv. Int. Med. 36, 249–270
Received for publication February 25, 1998.Revised for publication June 23, 1998.