exotic invasive plants in kentucky
TRANSCRIPT

University of Kentucky University of Kentucky
UKnowledge UKnowledge
University of Kentucky Master's Theses Graduate School
2010
EXOTIC INVASIVE PLANTS IN KENTUCKY EXOTIC INVASIVE PLANTS IN KENTUCKY
Yu Liang University of Kentucky, [email protected]
Right click to open a feedback form in a new tab to let us know how this document benefits you. Right click to open a feedback form in a new tab to let us know how this document benefits you.
Recommended Citation Recommended Citation Liang, Yu, "EXOTIC INVASIVE PLANTS IN KENTUCKY" (2010). University of Kentucky Master's Theses. 23. https://uknowledge.uky.edu/gradschool_theses/23
This Thesis is brought to you for free and open access by the Graduate School at UKnowledge. It has been accepted for inclusion in University of Kentucky Master's Theses by an authorized administrator of UKnowledge. For more information, please contact [email protected].

ABSTRACT OF THESIS
EXOTIC INVASIVE PLANTS IN KENTUCKY
Invasion of exotic species is a significant problem in natural ecosystems, reaching
epidemic proportions and resulting in significant economic losses. However, insufficient
knowledge of explicit spatial distribution of invasive species hinders our ability to
prevent and/or mitigate future invasion. In this study, we demonstrate the use of existing
voluntary data to survey invasive plant species in Kentucky. We also reconstructed the
historical distribution of 16 exotic invasive plants typical to Kentucky using herbarium
records. We found that Kentucky is facing a large threat from exotic invasive plants as
they are reported throughout most counties. The distribution maps for four of the top 10
most reported invasive species revealed that Kentucky is presently or was previously a
front of invasion. The majority of the 16 targeted invasive species were scattered
throughout Kentucky with no concentrations within particular regions. Cumulative curves
of occupied counties over time fit a “J” shape expansion curve, which indicates the
potential for further future invasion. This study demonstrates the usefulness of voluntary
data and herbarium data to reconstruct the historical and current distribution of invasive
species. Further studies on other invasive species can take advantage of information
associated with herbarium specimens to achieve more fruitful results.
KEYWORDS: Exotic invasive plant, historical distribution, herbarium records,
expansion rate, predominant land use type.
Yu Liang
June 18, 2010

EXOTIC INVASIVE PLANTS IN KENTUCKY
By
Yu Liang
Dr. Songlin Fei
_______________________________
Director of Thesis
Dr. David B. Wagner
_______________________________
Director of Graduate Studies
June 16, 2010
_______________________________

RULES FOR THE USE OF THESES
Unpublished theses submitted for the Master’s degree and deposited in the University of
Kentucky Library are as a rule open for inspection, but are to be used only with due
regard to the rights of the authors. Bibliographical reference may be noted, but quotations
or summaries of parts may be published only with the permission of the author, and with
the usual scholarly acknowledgements.
Extensive copying or publication of the thesis in whole or in part also requires the
consent of the Dean of Graduate School of the University of Kentucky.
A library that borrows this thesis for use by its patrons is expected to secure the signature
of each user.
Name Date
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________
________________________________________________________________________

THESIS
Yu Liang
The Graduate School
University of Kentucky
2010

EXOTIC INVASIVE PLANTS IN KENTUCKY
________________________________________________________________________
THESIS
________________________________________________________________________
A thesis submitted in partial fulfillment of the
requirements for the degree of Master of Science in the
College of Agriculture at the University of Kentucky
By
Yu Liang
Lexington, Kentucky
Director: Dr. Songlin Fei, Professor of Forestry
Lexington, Kentucky
2010
Copyright © Yu Liang 2010

iii
ACKNOWLEDGEMENTS
The following thesis benefited from the help of many people. I wish to express
sincere appreciation to my advisor, Dr. Songlin Fei, who recruited me from China and
changed my life for the better. I thank him for his understanding and support throughout
my Master’s research. Thanks also go to my committee members: Dr. Jonathan D. Green,
Mr. Robert Paratley, and Dr. John Lhotka. I thank Dr. Green for providing the weed
identification data that served as the foundation of my research. His advising and support
was also very important for my thesis. Rob, I appreciate your help and your kind advice
every time I burst into your office or intercepted you on your way home. Dr. Lhotka, you
made my graduation possible and always gave me strength and courage to face the
difficulties I encountered.
Thanks to all the herbarium curators who welcomed me, sent me data, or allowed me
to download data from their databases. Your persistence and hard work is critical for
advancing science. Many thanks go to my colleagues: John Hast, Ben Augustine, Kevin
Devine, David Parrott, and Dr. Liang Liang for their encouragement and peer reviews.
My deepest thank goes to my family, whose support have helped me during the days I
study abroad. Last but not least, my deepest love goes to my fiancée Shan Liu for her
love, support, sacrifice, and understanding during the whole period.
This thesis is based upon work supported by the Cooperative State Research,
Education and Extension Service, U.S. Department of Agriculture, under Agreement No.
2008-34628-19532. Any opinions, findings, conclusions, or recommendations expressed
in this publication are those of the author(s) and do not necessarily reflect the view of the
U.S. Department of Agriculture.

iv
TABLE OF CONTENTS
ACKNOWLEDGEMENTS ............................................................................................................ iii
TABLE OF CONTENTS ................................................................................................................ iv
LIST OF TABLES .......................................................................................................................... vi
LIST OF FIGURES ....................................................................................................................... vii
CHAPTER ONE: Introduction ........................................................................................................ 1
THE THREAT OF EXOTIC INVASIVE SPECIES ................................................................... 1
EXOTIC INVASIVE PLANTS IN KENTUCKY ....................................................................... 2
Amur honeysuckle ................................................................................................................... 3
Annual bluegrass ...................................................................................................................... 4
Autumn olive ........................................................................................................................... 4
Barnyardgrass .......................................................................................................................... 5
Canada thistle ........................................................................................................................... 5
Climbing euonymus ................................................................................................................. 6
Common chickweed ................................................................................................................. 6
Common lespedeza .................................................................................................................. 7
Common reed ........................................................................................................................... 7
Ground ivy ............................................................................................................................... 8
Japanese honeysuckle .............................................................................................................. 8
Japanese knotweed ................................................................................................................... 9
Japanese stiltgrass .................................................................................................................. 10
Kudzu ..................................................................................................................................... 10
Multiflora rose ....................................................................................................................... 11
Musk thistle ............................................................................................................................ 12
Poison-hemlock...................................................................................................................... 13
Princesstree ............................................................................................................................ 13
Sericea lespedeza ................................................................................................................... 14
Start-of-Bethlehem ................................................................................................................. 14
Tree-of-heaven ....................................................................................................................... 15
White mulberry ...................................................................................................................... 16
DISTRIBUTION OF INVASIVE PLANTS.............................................................................. 16
CHAPTER TWO: A Survey of Invasive Exotic Plants Using a Weed Identification Service
Database ......................................................................................................................................... 20

v
SUMMARY ............................................................................................................................... 20
INTRODUCTION ..................................................................................................................... 21
MATERIALS AND METHODS ............................................................................................... 23
RESULTS .................................................................................................................................. 25
Overview of Invasive Weeds in Kentucky ............................................................................ 25
Spatial and Temporal Distribution of the Top 10 Invasive Weeds ........................................ 27
Predominant Land Use Type for the Top 10 Invasive Weeds ............................................... 28
DISCUSSION ............................................................................................................................ 29
CHAPTER THREE: The Historical Distribution of Sixteen Exotic Invasive Species in Kentucky
based on Herbarium Records ......................................................................................................... 41
SUMMARY ............................................................................................................................... 41
INTRODUCTION ..................................................................................................................... 42
MATERIALS AND METHODS ............................................................................................... 43
RESULTS .................................................................................................................................. 44
Herbaria specimens ................................................................................................................ 44
Historical distribution ............................................................................................................ 45
Distribution rate ..................................................................................................................... 46
DISCUSSION ............................................................................................................................ 47
CONCLUSION .......................................................................................................................... 53
APPENDICES ............................................................................................................................... 65
REFERENCE ................................................................................................................................. 78
VITA .............................................................................................................................................. 92

vi
LIST OF TABLES
Table 2.1 The top 10 most frequently reported invasive exotic weeds in University of
Kentucky Weed Science Identification Program (UKWSIP) since 1986. ........................ 34 Table 2.2 Number of species within each family reported within the Kentucky Weed
Science Identification Program. ........................................................................................ 35
Table 2.3 Percentage of major land use type associated with the top 10 invasive exotic
weeds reported in UKWSIP. The number with an asterisk (*) represents this land use
type has more than twice of percentage than average. ...................................................... 36 Table 2.4 Percentage of predominant Cropland type associated with the top 10 invasive
exotic weeds reported in UKWSIP. The number with an asterisk (*) represents this land
use type has more than twice of the number of case than average. .................................. 37
Table 3. 1 Invasive species first reported year, county, total occupied counties .............. 54

vii
LIST OF FIGURES
Figure 2.1 County level spatial distribution of invasive exotic weeds by (a) year first
reported, (b) total number of invasive species reported based on KY-EPPC list, (c) report
frequency based on UKWSIP database, and (d) the Kentucky population density (2000
Census data). ..................................................................................................................... 38
Figure 2.2 Spatial distribution of (a) Canada thistle (Cirsium arvense), (b) Japanese
stiltgrass (Microstegium vimineum), (c) Japanese knotweed (Polygonum cuspidatum) and
(d) sericea lespedeza (Lespedeza cuneata) at the county level. The strip shadow
represents weed distribution based on EDDMapS data and PLANTS database. Different
solid shadings indicate weed spread in different periods based on weed identification
data. ................................................................................................................................... 39 Figure 2.3 Spatial distribution of (a) common lespedeza (Kummerowia striata), (b)
ground ivy (Glechoma hederacea), (c) star-of-Bethlehem (Ornithogalum umbellatum),
(d) common chickweed (Stellaria media), (e) annual bluegrass (Poa annua), and (f)
barnyardgrass (Echinochloa crus-galli) at the county level. The strip shadow represents
weed distribution based on EDDMapS and PLANTS database. Different solid shadings
indicate weed spread in different periods based on weed identification data. .................. 40
Figure 3.1 Historical distribution of Amur honeysuckle (Lonicera maackii (Rupr.)
Herder) in Kentucky based on herbaria data from 1960 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records. ............................. 55 Figure 3.2 Historical distribution of kudzu (Pueraria montana var. lobata (Willd.)
Maesen & S.M. Almeida) in Kentucky based on herbaria data from 1908 to 2008.
Occupied counties are shaded based on the time period during which the first specimen of
that species was recorded. Gray counties are the locations of the earlier plant records. .. 56
Figure 3.3 Historical distribution of common reed (Phragmites australis (Cav.) Trin. ex
Steud.) in Kentucky based on herbaria data from 1973 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records. ............................. 57 Figure 3.4 Historical distribution of tree-of-heaven (Ailanthus altissima (P. Mill.)
Swingle) in Kentucky based on herbaria data from 1892 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records. ............................. 58 Figure 3.5 Historical distribution of Japanese stiltgrass (Microstegium vimineum (Trin.)
A. Camus) in Kentucky based on herbaria data from 1931 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records. ............................. 59
Figure 3.6 Cumulative number of occupied counties for invasive species introduced from
(a) 1878 to 1909, (b) 1910 to 1941 and (c) 1942 to 1973. ................................................ 60 Figure 3.7 Historical distribution of princesstree (Paulownia tomentosa (Thunb.) Sieb. &
Zucc. ex Steud.) in Kentucky based on herbaria data from 1878 to 2008. Occupied
counties are shaded based on the time period during which the first specimen of that
species was recorded. Gray counties are the locations of the earlier plant records. ......... 61 Figure 3.8 Historical distribution of autumn olive (Elaeagnus umbellata Thunb.) in
Kentucky based on herbaria data from 1939 to 2008. Occupied counties are shaded based
on the time period during which the first specimen of that species was recorded. Gray

viii
counties are the locations of the earlier plant records. ...................................................... 62 Figure 3.9 Historical distribution of Canada thistle (Cirsium arvense (L.) Scop.) in
Kentucky based on herbaria data from 1879 to 2008. Occupied counties are shaded based
on the time period during which the first specimen of that species was recorded. Gray
counties are the locations of the earlier plant records. ...................................................... 63 Figure 3.10 Historical distribution of musk thistle (Carduus nutans L.) in Kentucky based
on herbaria data from 1945 to 2008. Occupied counties are shaded based on the time
period during which the first specimen of that species was recorded. Gray counties are
the locations of the earlier plant records. .......................................................................... 64

1
CHAPTER ONE: Introduction
THE THREAT OF EXOTIC INVASIVE SPECIES
Exotic species are those that have been introduced outside their native ranges
purposely or accidentally (Williams and Grosholz 2008). Not all exotic species become
invasive after they were introduced. Some exotic species, such as rice, corn, and other
food crops, now provide more than 98% of food in the U.S. worth approximately $800
billion per year (Pimentel et al. 2005). Some exotic species, however, cause a decrease in
native species diversity, and alter the function of ecosystems (Reid et al. 2009; Ward and
Jasieniuk 2009; White et al. 2008). In this thesis, an invasive species is defined as an
exotic species whose introduction does or is likely to cause economic or environmental
harm, or harm to human health (Rossman 2001).
Although species establish and spread on their own, increased mobility of humans
have brought about an unparalleled movements of exotic species (Botham et al. 2009;
Chauvel et al. 2006; Flory and Clay 2009; Reichard and White 2001; Rejmanek 1996).
Approximately 50,000 non-native species have been introduced into the United States
alone (Pimentel et al. 2005), compared with a total of about 17,000 native species (Morse
1995). These invasive alien species are considered a threat to native species, and are
widely considered responsible for the decline of species diversity and ecosystem stability.
Exotic invasive species eliminate native species by utilizing various mechanisms, such as
resource competition (Fasola et al. 2009; McNatty et al. 2009; Snyder et al. 2009;
Strubbe and Matthysen 2009) and predation (Bellingham et al. 2010; Kwong et al. 2009;
Pichlova-Ptacnikova and Vanderploeg 2009; Piggott et al. 2008; Strecker and Arnott

2
2008). They crowd out native species from gene pool, which reduces the diversity of
species (Adams and Engelhardt 2009; Anderson and Rosemond 2007; Davis 2003). The
loss of biodiversity, along with the effect of climate change, may decrease the persistence
of ecosystems to stochastic events such as natural disasters and insect pest (Bradley 2010;
Callaway and Maron 2006; Manchester and Bullock 2000). Another concern about
invasive exotic species relates to increasing economic loss in agriculture, forestry and
other segments of the U.S. economy. In agriculture, invasive plants crowd out food crops
for sunlight and water resources (Allaie et al. 2006), reduce crop and forage quality
(Stewart et al. 2009; Suckling and Brockerhoff 2010), and in some case poison livestock
species (Legere 2009). The financial impacts and management costs due to loss of
productivity, costs of herbicides, and other control measures for invasive species is
estimated to exceed $120 billion annually (Pimentel et al. 2005). Invasive disease may
also directly affect human health (Molocznik 2004; Tuiten et al. 2009). For example,
West Nile virus arrived in the United States as recently as 1999 (Edman 2004). In 2003,
4200 people were infected, resulting in 284 deaths (Wonham et al. 2004).
EXOTIC INVASIVE PLANTS IN KENTUCKY
Kentucky faces a huge challenge from exotic species invasion, making this a problem
pressing for a solution. Invasive species can invade into Kentucky from seven
surrounding states. Also, Kentucky is located around 40 degree north latitude, where is
the species transition zone for cool season plants and warm season plants.
Longitudinally, KY has many ecoregions, such as mountains in the east, plain in the
central, and wetland in the west. Its unique geographic location increases the diversity of

3
invasive species in Kentucky. Additionally, the moderate, relatively humid climate in
Kentucky is similar with that in Europe and eastern Asia where most of North American
invasive species came from. It has been reported that, among all the vascular plants, a
quarter of them were introduced (Jones 2005). Ninety-two plant species have been
declared as exotic invasive species by Kentucky Exotic Pest Plant Council (KY-EPPC
2008). The following are the major exotic invasive plants included in this study. General
introductions and biological traits of these invasive plants are described below.
Amur honeysuckle
Amur honeysuckle (Lonicera maackii (Rupr.) Herder) is a deciduous shrub native to eastern
Asia (Luken and Thieret 1996). It was introduced into North America in 1896 as an ornamental
shrub (Luken and Mattimiro 1991). Amur honeysuckle occurs throughout the eastern United
States (Goodell et al. 2010). With the seeds being dispersed by birds and mammals (Bartuszevige
and Gorchov 2006; Luken and Goessling 1995) , this plant forms a dense understory which
restricts native plant growth in forests, and can adversely affect populations of native species in
disturbed open areas and forest edge (Castellano and Boyce 2007; Hartman and McCarthy 2008;
Miller and Gorchov 2004; Swab et al. 2008). Because of its floral and fruit display, the species is
still widely available in nurseries and botanical gardens across the country (Luken and Thieret
1996). The distribution pattern of Amur honeysuckle has been studied in southwestern Ohio
(Medley 1997). The study found honeysuckle abundances were highest in the forest edge area
and small discontinuous patches. Colonization in understory of forest and subsequent
development for Amur honeysuckle was also studied using sample populations in Ohio (Deering
and Vankat 1999). Scattered distributions in multiple locations likely contribute to the
colonization of this invasive shrub. Restoration of natural understory ecosystem was also

4
successfully carried out after the removal of this invasive shrub using two different herbicides
(Hartman and McCarthy 2004).
Annual bluegrass
Annual bluegrass (Poa annua L.) is a cool-season annual, biennial, or perennial grass.
It is a native to Europe and now wildly distributed around the world. It thrives in lawns,
gardens, pastures, roadsides, and disturbed areas throughout the U.S. (USDA 2010).
Annual bluegrass can reduce nutrient availability in the upper soil horizons by forming
dense mats (Johnson 1979). The seeds of this weed can be spread by birds (Koshy 1969).
Vegetative portions are probably eaten by large mammals which also contributes the
spread of this weed. A wide range of invertebrates also feed on annual bluegrass (Beard
et al. 1978).
Autumn olive
Autumn olive (Elaeagnus umbellata Thunb.) was introduced into the United States in 1830
from eastern Asia as an ornamental plant (Catling et al. 1997). This shrub species is found in the
east and northwestern United States (Goldstein et al. 2009). Unlike many other introduced species
which prefer colonizing in forest edge areas (Tlig-Zouari et al. 2009), autumn olive also succeeds
in the adjacent interiors of forest (Yates et al. 2004). It often crowds out native plant species by
creating dense thickets and interfering with natural plant succession and nutrient cycling with
Frankia, a nitrogen-fixing endosymbiont (Brantley and Young 2010; Goldstein et al. 2009; Yates
et al. 2004). Autumn olive produces a great amount of small fleshy drupes, which are consumed
by birds or mammals, and may rapidly expand over large distances (Ahmad et al. 2006; Knapp et
al. 2008; McCay et al. 2009).

5
Barnyardgrass
Barnyardgrass (Echinochloa crus-galli (L.) Beauv.) is an annual of Eurasian origin
that occurs throughout the continental United States (Kaufman and Kaufman 2007).
Barnyardgrass invades cultivated fields, waste places, floodplains and other disturbed
area (Brod 1968). Although it is invasive, it is not strongly competitive with native plants
except in low, moist, disturbed areas (Maun 1977). Barnyardgrass is tolerant to long wet
periods and withstands considerable salt and alkali (Assemat et al. 1981). It is self-
pollinating and a prolific seed producer that can produce as many as one million seeds.
The seed viability drops after one year, but can last up to 13 years (Kennedy et al. 1980).
Canada thistle
Canada thistle (Cirsium arvense (L.) Scop.), a cool season perennial, is native to
Europe and probably arrived in North America in the early 1600s (Kaufman and
Kaufman 2007). It grows in the northern United States and southern Canada (USDA
2010). Canada thistle is particularly problematic in farms and pastures because it causes
economic loss by reducing crop yields and pasture productivity (Demers et al. 2005;
Reece and Wilson 1983; Zimdahl and Foster 1993). This species produces large
quantities of “bristly-plumed” seeds which are dispersed by wind or water (Hayden
1934). Its seeds can remain viable for over 20 years in the soil (Lalonde and Roitberg
1994). Canada thistle can also regenerate vegetatively by roots or shoot sprouts (Laubhan
and Shaffer 2006). Its invasiveness was soon recognized after it was introduced into the
U.S.. Since then, a lot of research work has been carried out about the control strategy for
this weed (Foote et al. 1970; Peschken and Wilkinson 1981; Wedekind 1991). Spatial

6
distribution of this weed has been studied in Germany and a spatially explicit simulation
model for the dispersal of Canada thistle was created to predict further invasion of this
weed (Belde and Mayer 2002).
Climbing euonymus
Climbing euonymus (Euonymus fortunei (Turcz.) Hand.-Maz.) is native to eastern
Asia and was introduced in1907 from China (Kaufman and Kaufman 2007). This
evergreen vine grows in the eastern United States, from New York to Mississippi (USDA
2010). Climbing euonymus is an ever-green clinging vine widely used in residential
landscaping (Harris et al. 2009). It outcompetes the natural vegetation by forming dense
thickets which restricts native species growth (Swearingen 2009). Its vines can climb
overtop native plants and block sunlight (Kaufman and Kaufman 2007). Climbing
euonymus is sold in commercial nurseries and spreads by horticultural trade (Boyer et al.
2008). It escapes the gardens through seed dispersal by birds and small mammals, and
then invades into neighborhoods vegetatively (Rehder 1993).
Common chickweed
Common chickweed (Stellaria media (L.) Vill.), an annual weed introduced from
Eurasia, was first recorded in New England in 1672 (Turkington et al. 1980). It was
probably brought as an herbal remedy by European explorers and emigrants (Shotwell
1993). Common chickweed is now found throughout the United States (USDA 2010). Its
seed was probably spread by emigrants who cultivated the plant for medicinal and leafy
vegetable use and by seeds contaminating grain shipments (Miura and Kusanagi 2001).

7
The weed is particularly well adapted to disturbed agricultural habitats by its ability to
germinate throughout the year in temperate climates, the short after-ripening of the seed,
and a short growth period from germination to flowering and seed set (Inderjit and
Dakshini 1998). Common chickweed seeds can remain viable in the soil for at least 30
years (Storkey and Cussans 2000).
Common lespedeza
Common lespedeza (Kummerowia striata (Thunb.) Schindl.), a Japanese annual herb,
was first reported growing in Georgia in 1846 (Kaufman and Kaufman 2007). Common
lespedeza is widely planted and naturalized in the southeastern U.S. (USDA 2010). This
plant is used extensively on crop lands and pastures for hay, grazing and soil
improvement. Common lespedeza occurs in roadsides, open area, and other disturbed
sites. It may become invasive in some regions or habitats and may displace desirable
vegetation if not properly managed (Nakatsubo 1995). This weed has a rapid ability to
spread through seed production and the seedlings have high vigor (Nakatsubo 1995).
Common reed
Common reed is a large, perennial grass that grows to over 15 feet in height. Both
native and introduced forms of (Phragmites australis (Cav.) Trin. ex Steud.) exist in the
United States (Catling and Carbyn 2006; Meadows and Saltonstall 2007). The introduced
(European) genotype of common reed is held to be responsible for the widespread
invasion of this plant (T'ulbure et al. 2007). Introduced common reed occurs within all of
the lower 48 states (USDA 2010). Their dense stands outcompete native aquatic or marsh

8
plants and eventually reduce flora diversity (Able et al. 2003; Minchinton 2006). This
species was reported to reproduce either by seed or by rhizomes (Brisson et al. 2008).
Historical spread and dispersal pathways of this weed have been analyzed using
herbarium records in Quebec, Canada (Lelong et al. 2007). The study suggested that the
development of the highway network in the 1960s and 1970s strongly contributed to the
inland expansion of the exotic genotype.
Ground ivy
Ground ivy (Glechoma hederacea L.), a perennial herb, was introduced as an ornamental
or medicinal plant from Eurasia as early as the 1800s (Kaufman and Kaufman 2007).
This invasive weed is common throughout the U.S. except in the Southwest. Trailing
over the ground, ground ivy forms a thick ground cover that prevents the establishment
and growth of other native plants (Fernande 1971). It is problematic in lawn and also can
be found in natural area. Ground ivy prefers the moist ground, shade of floodplains and
disturbed area (Kohler et al. 2004a). Stems root at the nodes and stem fragments are the
major source of spread (Mitich 1994).
Japanese honeysuckle
A perennial vine species from eastern Asian, Japanese honeysuckle (Lonicera
japonica Thunb.) was introduced into the U.S. as a horticultural plant by a gardener in
1806 (Schierenbeck 2004). It currently occurs across the southern United States and New
England (USDA 2010). Japanese honeysuckle grows best in high-light forest edges and
open fields (Robertson et al. 1994). It forms a dense ground cover in the understory,

9
outcompeting native plants (Belote and Weltzin 2006; Merriam 2003). Japanese
honeysuckle is still widely available in the nurseries and spreads through the horticultural
trade as an ornamental plant (Kowarik 2003). Further invasion of this species is
facilitated via birds, mammals, and vegetative spread (Schierenbeck 2004). In Oklahoma,
the historical spread of Japanese honeysuckle over the past 100 years has been
reconstructed using herbarium data (Crawford and Hoagland 2009). Maps of the
collection records of this invasive vine illustrated no discernible spatial invasion or
expansion pattern.
Japanese knotweed
Japanese knotweed (Polygonum cuspidatum Sieb. & Zucc.), a herbaceous perennial
plant, was introduced from Japan to the United States in the late 1800s (Kaufman and
Kaufman 2007). This species occurs across 40 states in the U.S., from Maine to
Louisiana and in several midwest and western states (USDA 2010). Japanese knotweed
reproduces primarily by vegetative means via rhizomes, yet prolific seed reproduction is
also observed (Zika and Jacobson 2003). Japanese knotweed spreads quickly, forming
dense stands that exclude indigenous species (Weston et al. 2005). Its underground root
system and viable seed make it extremely difficult to eradicate (Forman and Kesseli
2003). The historical spread of this weed have been recreated using Index Herbarium data
in 2003-2004 in North America (Barney 2006). The study indicated that Japanese
knotweed frequently distributed along waterway and road suggesting large-scale human
related spread across North America. Another study in Canada using Invasive Alien Plant
Program data suggested over half of the suitable habitat for Japanese knotweed in British

10
Columbia have already been occupied, indicating there are still significant areas to be
invaded (Bourchier and Van Hezewijk 2010).
Japanese stiltgrass
Japanese stiltgrass (Microstegium vimineum (Trin.) A. Camus), an annual herbaceous
plant, was first introduced into the United States in Tennessee around 1919 (Fairbrot.De
and Gray 1972a). Japanese stiltgrass is primarily established in 25 states, from Maine to
Florida, west to Texas (USDA 2010). This species spreads mainly by wind or water due
to its extremely light seeds (Cheplick 2005; Mehrhoff 2000). Dense patches of Japanese
stiltgrass established after habitat disturbance can outcompete native communities by
robbing them of sunlight (Rauschert et al. 2010). It also alters soil structures and
properties, such as pH, further interfering with native plants regeneration (Ehrenfeld et al.
2001; Hunt and Zaremba 1992; Kourtev et al. 1998). The spread and distribution of this
weed have been reported along the Hudson River in New York and reported from three
sites in Connecticut (Hunt and Zaremba 1992). A landscape-level survey of Japanese
stiltgrass at east Tennessee was conducted. The study suggested different factors may
interact to control the distribution and performance of this species locally (Cole and
Weltzin 2004).
Kudzu
Kudzu (Pueraria montana var. lobata (Willd.) Maesen & S.M. Almeida) was a
perennial vine introduced into the United States from Japan in 1876 at the Philadelphia
Centennial Exposition (Miller 1983). It has been widely used for soil erosion control

11
during the 1930s (Forseth and Innis 2004). Kudzu is common throughout most of the
southeastern United States and recently has also been discovered in northern states
(Forseth and Innis 2004; Lamont and Young 2002). This invasive vine thrives in forest
edge areas, abandoned fields, and disturbed areas, where sunlight is abundant (Newton et
al. 2008). Kudzu’s vigorous growth kills trees by girdling, while its dense of leaves
smother native plants (Forseth and Innis 2004). This plant spreads vegetatively in most
cases (Merriam 2003). Rapid stem elongation and frequent rooting at nodes makes this
invasive vine effective in its domination of disturbed areas and habitat gaps (Forseth and
Innis 2004; Merriam 2003). A series of control methods have been developed to control
this invasive vine, such as herbicide (Dickens and Buchanan 1971) and biocontrol using
soybean rust (Phakopsora pachyrhizi) (Hershman et al. 2006; Zidack and Backman
1996). A systematic survey for kudzu biocontrol agents in China was also underway and
several of kudzu foliage, seed, stems, and roots associated insects have be reported (Sun
et al. 2006).
Multiflora rose
Multiflora rose (Rosa multiflora Thunb. ex Murr.) was imported into the eastern
United States in 1866 from eastern Asia as an ornamental and farm hedge (Steavenson
1946). It occurs throughout most of the United States except in the Rocky Mountains and
deserts (USDA 2010). It tolerates a wide range of soil, moisture, and light conditions,
which allows it to grow aggressively into riparian areas, thickets, and woodlands
(Merriam 2003). In herbaceous communities, impenetrable thickets of multiflora rose
exclude native shrubs and herbs from establishing (Yurkonis et al. 2005). Multiflora rose

12
increases native species seed removal and affects forest regeneration by providing cover
for small seed predators (Meiners and LoGiudice 2003). It is a prolific producer and its
seeds can remain viable in the soil for up to 20 years (Banasiak and Meiners 2009). Birds
are a primary dispersal method for the multiflora rose (Borgmann and Rodewald 2004).
However, this invasive shrub can also reproduce vegetatively (Szafoni 1991). Control
methods of multiflora rose have been accessed in many ways (Derr 1989; Hindal and
Wong 1988). For example, rose rosette disease had been used for biocontrol of this
invasive shrub (Epstein et al. 1997).
Musk thistle
Musk thistle (Carduus nutans L.), a biennial weed, was first recorded in central
Pennsylvania in 1852 from its native range of Europe and western Asia (Kaufman and
Kaufman 2007). This species occurs throughout the United States except for Florida
(USDA 2010). Musk thistle forms dense stands, reducing yield in pastures considerably
by crowding out forage plants (Moore et al. 1989; Popay et al. 1989; Wardle et al. 1995).
Musk thistle reproduces and spreads only by seed. One single plant can produce
thousands of seeds which are easily dispersed by wind or water (Jongejans et al. 2008).
Its seeds may remain viable for up to ten years, making it difficult to eradicate (Jongejans
et al. 2006; Metcalf et al. 2009). Effective methods available for control of musk thistle
have also been developed in several countries beside the U.S. such as New Zealand and
Australia (Moore et al. 1989; Shea et al. 2006).

13
Poison-hemlock
Poison-hemlock (Conium maculatum L.), a European biennial weed, was first noticed
in the United States in the 1800s (Kaufman and Kaufman 2007). It grows throughout
southern Canada and the United States (USDA 2010). Poison-hemlock can grow quickly
to occupy disturbed areas and displace native vegetations in moist areas (Schmida 1974).
This weed produces coniine, an alkaloid which is poisonous to both livestock and humans
(Bowman and Sanghvi 1963; Fairbairn and Challen 1959; Lopez et al. 1999). Poison-
hemlock reproduces only from seeds which can adhere to farm equipments, clothes, and
mud (Goeden and Ricker 1982). Its seeds can also be carried by water and wind (Knight
1987).
Princesstree
Princesstree (Paulownia tomentosa (Thunb.) Sieb. & Zucc. ex Steud.) was imported to
Europe by the Dutch East India Company in the 1830s as an ornamental and brought to North
America soon after (Langdon and Johnson 1994). Princesstree occurs in eastern U.S., from Maine
to Texas (USDA 2010). By tolerating drought conditions and acidic soils, it is able to adapt to a
wide variety of habitats (Moore and Lacey 2009). One single princesstree can produce millions of
seeds, which are easily dispersed by wind or water (Grubisic et al. 1985). These seed can
germinate rapidly in many soil types (Hyatt and Casper 2000; Moore and Lacey 2009). This
invasive tree can also reproduce from its extensive root system (Mueller et al. 2001).
Princesstress was reported to displace native plant species in disturbed areas, burns, and forest
edges (Ede et al. 1997; Essl 2007).

14
Sericea lespedeza
Sericea lespedeza (Lespedeza cuneata (Dumont) G. Don), a perennial legume native
to eastern Asia, was introduced in Virginia in 1899 as a forage species (Kaufman and
Kaufman 2007). It occurs throughout the eastern United States from New York to Texas
(USDA 2010). Once established, this species displaces native plants and hinders their
habitat by spreading dynamically (Allred et al. 2010). This plant is a prolific producer as
it can produce as much as 670 kg of seed/hectare annually which remain viable for over
30 years (Logan et al. 1969). Recently, hyperspectral imagery has been used to detect the
sericea lespedeza in pastureland in mid-Missouri and the quantitative distribution of
sericea volume was then mapped using an empirical regression model (Wang et al. 2008).
Start-of-Bethlehem
Start-of-Bethlehem (Ornithogalum umbellatum L.) is a European herbaceous plant that
grows from bulbs with annual renewal (Demars 1994). It was most likely introduced into
the U.S. as a horticultural plant. The exact date of introduction is not known, but there
were reports of large stands of it in 1940 in the forests of Indiana (Demars 1994).
Currently, this plant is found throughout the states except New Mexico, Nevada, Arizona,
Colorado, Wyoming, Montana, and North Dakota (USDA 2010). It was most likely
dispersed via additional plantings as well as the bulbs being washed downstream (Moret
et al. 1991). Star-of-Bethlehem causes potential threats to native vegetation by competing
for the nutrient (Gadella 1972). Star-of Bethlehem is non-responsive to several
herbicides. Research studies found that paraquat provided 70 to 78% control (Main et al.
2004).

15
Tree-of-heaven
Tree-of-heaven (Ailanthus altissima (P. Mill.) Swingle) is a small to medium-sized deciduous tree
native to China (Ding et al. 2006). It was introduced from China into Europe in 1751 and then
introduced via Europe to North America in 1784 (Swingle 1916). It is now widely distributed
across the United States, occurring in 42 states, from Maine to Florida and west to California
(USDA 2010). Once established, tree-of-heaven sends out many root sprouts, rapidly forming
dense thickets that prevent native species growth (Call and Nilsen 2005). Its root system can
cause damage to roadways, sidewalks, and sewer structures. This tree species is also a prolific
seed producer; a mature tree-of-heaven can produce up to 350,000 seeds a year (Kaproth and
McGraw 2008). Seeds maintain a high germination rate even after 5 months (Kaproth and
McGraw 2008). This invasive species is able to disperse long distances into fields and fragmented
landscapes by wind or water (Kaproth and McGraw 2008; Kowarik and Saumel 2008;
Landenberger et al. 2007). In addition to its prolific reproduction and extensive root system, tree-
of-heaven also has alleopathic effects on other tree species growing in its neighborhood by
producing phytotoxic compounds from roots and leaves (Ding et al. 2006; Heisey 1990).
Recently, studies about the invasion mechanism and control method of tree-of-heaven have been
conducted. Call (2005) suggested the positive association between tree-of-heaven and native
black locust (Robinia pseudoacacia) along the highly disturbed skid trails. Ding (2006) reviewed
literature about enemies of the plant in order to access the potential biological control of this
plant. However, the dispersal pattern of tree-of-heaven was only reported in southwest Ohio,
where tree-of-heaven frequently dominate on the edge of forest instead of the interior
(Espenschied-Reilly and Runkle 2008).

16
White mulberry
White mulberry (Morus alba L.), a deciduous tree native of eastern Asia, was most
likely introduced to the United States as an ornamental tree in the mid-1800s (Kaufman
and Kaufman 2007). It grows across the country except for Nevada (USDA 2010).
During colonial times, white mulberry was widely planted to establish a silk industry in
the United States (Noe 2009). Its seeds are spread by birds and mammals that feed on the
fruits. It can also reproduce vegetatively from its roots (Ghersa et al. 2002). White
mulberry forms dense thickets that exclude other native plant species (Burgess and
Husband 2006). The hybridization of white mulberry and native red mulberry (Morus
rubra L.) resulted in threats to the survival of red mulberry by displacing this native
species (Burgess and Husband 2006).
Overall, invasive plants appear to have specific traits or combinations of specific
traits that allow them to outcompete native species. The common traits of these sixteen
invasive plants include: fast growth, tolerance of a wide range of environmental
conditions, the ability to reproduce both asexually as well as sexually, high dispersal
ability, and association with human activities (Rejmanek and Richardson 1996).
DISTRIBUTION OF INVASIVE PLANTS
To stop or mitigate the invasion of existing and new emerging species, effective Early
Detection and Rapid Response (EDRR) systems are needed to detect the occurrence of
invasions (Chornesky et al. 2005; Wittenberg and Cock 2001). These EDRR systems
include functions for identifying the potential high priority species lists, providing

17
adequate and timely information of species distribution patterns to decision-makers and
the public, and determining the at-risk sites by utilizing predictive models (Chornesky et
al. 2005). In EDRR systems, identifying explicit spatial distribution pattern of exotic
invasive species in regions outside their native range is a fundamental component (Hulme
2006). The spatial distribution pattern of alien plants may either predict the species
distribution for a site, or further our understanding of factors that influence these
distributions (Higgins et al. 1999). This information is essential for planning eradication,
containment, and control strategies and treatments. Unfortunately, a common defect of
most EDRR systems is that they have insufficient power to detect the significant change
in the distribution of invasive species due to its limited sampling sites (Van Strien et al.
1997). However, sampling more sites to increase the power of surveys is impractical
because of the enormous amounts of manpower and material resources needed. The
initiatives of citizen volunteers may provide a less costly approach and strengthen the
ability of accurate predicting the invasion tender (Engler et al. 2004). Several regional
systems, Early Detection and Distribution Mapping System (EDDMapS,
http://www.eddmaps.org) created by Southeast Exotic Pest Plant Council (SE-EPPC),
Invasive Plant Atlas of New England (IPANE, http://www.ipane.org) and Invasive Plant
Atlas of MidSouth (IPAMS, http://www.gri.msstate.edu/ipams/), have taken advantage of
such citizen scientist efforts. The development and utilization of voluntary data systems
can add a new catalyst for understanding natural phenomenon, yet its application in
natural resource systems is limited (Orr et al. 2007).
Recognizing the need to identify an invasion of exotic species, many researchers have
begun to reconstruct the historical distribution of invasive species. Recently, progress in

18
this respect has been made by consulting real or virtual herbarium collections (Crawford
and Hoagland 2009; Delisle et al. 2003; Lavoie et al. 2007; Miller et al. 2009; Stadler et
al. 1998). For example, Chauvel (2006), using herbarium specimens in France and in
bordering countries, identified how annual ragweed (Ambrosia artemisiifolia L.) initially
arrived in France and subsequently spread out over time. However, we must be cautious
when the reconstruction of historical spread of invasive species is based on herbarium
specimens. Few studies have taken into consideration the biased nature of natural history
collections such as: unequal sampling effort over time, non-random geographical
representation, and disproportionately represented taxa (Delisle et al. 2003). For instance,
the increase in the number of specimens of an invasive species may indicate an increase
in abundance, or simply may mean an increase in the overall collecting effort that year or
decade (Crawford and Hoagland 2009). To remove such biases, Delisle et al. (2003)
developed a method by selecting indigenous species to provide a collection trend in the
study region. By comparing the collection trend, the increasing collections of invasive
species suggests the increase of the invasive species in range or abundance. Although the
data from herbarium specimens suffer from several biases, they constitute a valuable
source of information to document the early stages of the invasion process (Chauvel et al.
2006; Lelong et al. 2007; Stadler et al. 1998; Weber 1998; Wu et al. 2005). Plant
specimens stored in herbaria are usually well preserved, so the identification of species
can be validated or corrected (Lelong et al. 2007). Furthermore, each specimen includes a
label, usually with information about sampling location and date, which makes the
reconstructing of historical spread feasible (Delisle et al. 2003; Ponder et al. 2001).
Little knowledge is available about the spatial and temporal distribution patterns of

19
exotic invasive plants in Kentucky. This research is designed to gain an overall picture of
exotic invasive plants in the state of Kentucky and identify the historical distributions of
typical exotic invasive species in Kentucky. 24-year weed identification forms from
University of Kentucky Weed Science Identification Program (UKWSIP) were used to
survey exotic invasive plants in Kentucky. Spatial and temporal distribution maps for the
top 10 most reported invasive plants were created and the predominant land use type for
each of these top 10 species was determined according to collection records. To estimate
the long-time historical distribution, we also collected records from 15 herbaria in
Kentucky. Together with the UKWSIP data, herbaria collections dating back to 1870s
revealed the historical distributions of 16 typical exotic invasive plants in Kentucky. The
cumulative number of total occupied counties was used to determine the distribution rate
for each species. Knowledge gained from this project may help to prevent invasions and
mitigate the impact of exotic invasive weeds, producing long-term social, ecological, and
economic benefits.

20
CHAPTER TWO: A Survey of Invasive Exotic Plants Using a Weed Identification Service
Database
SUMMARY
Invasive species is a significant problem in many ecosystems, reaching epidemic
proportions and resulting in significant ecologic and economic losses. Our ability to
prevent and/or mitigate invasive species is in part hindered by insufficient knowledge of
spatial distribution and spread of invasive species in ecosystems that have been
challenged. In this study, we demonstrate the use of existing voluntary data to survey
invasive plant species in Kentucky. Data used in this study was collected through the
University of Kentucky Weed Science Identification Program (UKWSIP) which provides
a weed identification service for the public through the local county Cooperative
Extension Offices across the state. Distribution maps for the top 10 most reported
invasive plants were reconstructed to understand their distribution patterns. Predominant
land use type for each of these ten invasive species was determined by comparing
proportion of associated land use type for each species with proportion of each land use
type for the overall records. We found that Kentucky is facing a large threat from exotic
invasive weeds. We believe the invasive weeds, both in number of species, as well as
frequency, were underestimated because our data is on a voluntary base. The distribution
maps for the top 10 invasive species recorded indicated Kentucky was likely to be the
southern invasion front for Canada thistle and Japanese knotweed, and the northern
invasion front for Japanese stiltgrass and sericea lespedeza. Knowledge gained from this
project will help to prevent and mitigate the impact of exotic invasive weeds in Kentucky
and beyond.

21
INTRODUCTION
Exotic plant invasions have gained increasing attention globally due to their
substantial ecological impacts and dramatic economic cost. The loss of biodiversity that
invasive exotic species cause, along with other factors such as climate change, may
decrease the resilience of an ecosystem to natural disturbance and disease (Callaway and
Maron 2006; Manchester and Bullock 2000). To stop or mitigate the invasion of existing
and new emerging species, an effective Early Detection and Rapid Response (EDRR)
system is needed to detect the occurrence of invasions (Chornesky et al. 2005;
Wittenberg and Cock 2001). EDRR systems help to identify the potential priority species
list, provide adequate and timely information of species distribution patterns to decision-
makers and the general public, and determine at-risk sites by utilizing predictive models
(Chornesky et al. 2005; Lodge et al. 2006; Ries et al. 2004). In EDRR systems,
identifying spatial explicit distribution patterns of invasive species in regions outside
their native range is a fundamental component of their functions (Hulme 2006). The
spatial distribution pattern of invasive plants may help to either predict the species
distribution for a site, or further our understanding of factors that influence these
distributions (Higgins et al. 1999). This information is also essential for planning
eradication, containment, and control strategies of invasive species.
A common limitation of EDRR systems is that they have insufficient power to detect
the significant change in distribution of invasive species due to limited sites sampled
(Van Strien et al. 1997). However, sampling more sites to increase the power of surveys
is often impractical because it needs enormous manpower and resources. Recently,
researchers have begun to explore applications of remotely sensed imagery (e.g., satellite
imagery and aerial photography) for identifying the distribution or impacts of invasive

22
exotic species (Cuneo et al. 2009; Huang and Asner 2009; Kimothi et al. 2010; Wilfong
et al. 2009). These geospatial technologies can reduce costs and increase the efficiency
and effectiveness of weed management programs for certain weeds (Wilson et al. 2008).
However, many invasive plant species are not good candidates for remote sensing
because they are indistinguishable from other native plants, particularly during vegetative
growth (D'Iorio et al. 2007; Kimothi et al. 2010). On the other hand, some studies try to
infer temporal dynamics using historical records, such as herbarium or museum
collections (Chauvel et al. 2006; Graham et al. 2004; Lavoie et al. 2007). Success of
these studies varied depending on the availability of the data. Utilizing existing voluntary
data or initiating citizen scientist may provide a less costly approach and strengthen the
likelihood of predicting trends of exotic invasions by providing more observed data with
minimal cost (Engler et al. 2004). Several regional systems, such as Early Detection and
Distribution Mapping System (EDDMapS, http://www.eddmaps.org) created by
Southeast Exotic Pest Plant Council (SE-EPPC), Invasive Plant Atlas of New England
(IPANE, http://www.ipane.org) and Invasive Plant Atlas of MidSouth (IPAMS,
http://www.gri.msstate.edu/ipams/), have taken the advantage of the citizen scientist
approach. The development and utilization of the voluntary data system can add a new
catalyst for understanding natural phenomenon by offering more sample data, yet its
application in a natural resource system is limited (Orr et al. 2007).
In this study, we demonstrate the use of existing voluntary data from the University
of Kentucky Weed Science Identification Program (UKWSIP) to survey invasive species
in Kentucky. We also reconstructed the spatial and temporal distribution maps of the top
10 most reported invasive species and determined their predominant habitat types.

23
Knowledge gained from this project will help to prevent and mitigate the impact of exotic
invasive weeds, which will have long-term social, ecological, and economic benefits.
MATERIALS AND METHODS
Data collected through the UKWSIP from 1986 to 2009 were used in this study. The
UKWSIP identifies weedy plants for the public and provides weed control
recommendations in support of the county offices of the Cooperative Extension Service
throughout Kentucky. Since 1986, weed identification forms have been used to generate
written records pertaining to information on each plant specimen submitted for
identification. Slight modifications have been made during the last 24 years, but, in
general, identification forms have the following associated information: submitter’s
contact information, date, county, habitat associated with the collected specimen, and
species name and control recommendation. A total of 8,289 weed identification forms
collected across Kentucky were used in this study. All records were digitized into a
database including the following key fields: collection date, collection county, associated
land use type, scientific name, and common name of the submitted specimen. Records
with imprecise or incomplete information were excluded from further analyses.
In this study, we only focused on plants considered to be invasive exotic weeds.
There are many invasive exotic species lists available both at the state and national level.
To ensure local relevance, we consider a weed as an invasive exotic weed if it is listed as
an invasive exotic species by the Kentucky Exotic Pest Plant Council (KY- EPPC;
http://www.se-eppc.org/ky/list.htm). Nomenclature was unified through the database
following the Kentucky Exotic Pest Plant Council list. Statistical summaries were

24
generated to infer the status of invasive exotic weeds in Kentucky, including total number
of invasive exotic weeds, the most frequently reported invasive species and families, the
most wide spread species, and counties with the most invasive species reported. We also
conducted regression analysis to determine if an association existed between human
population and the total number of invasive species in each county.
Second, information from the exotic invasive weed database was used to create
distribution maps. Year and county of each collection were used to define the time and
location of the presence of a given invasive species. To illustrate spatial pattern, tabular
records of invasive species were first attributed to different counties in ArcGIS 9.3
(ESRI, Inc. Redlands, CA) based on the location information of samples. The earliest
record for a given invasive species in each county was used to represent its spatial
dynamic. This means counties once occupied by a given invasive species were defined as
permanently “infested” by this species. To ensure sufficient data to understand the
invasion pattern, only the top 10 most frequently reported exotic invasive weeds were
mapped (Table 2.1). Distribution maps generated from our database were then
overlapped with maps from USDA PLANTS database (USDA NRCS 2010) and
EDDMapS (2010) in Kentucky and adjacent states. By combining these geographic
distributions, we inferred whether Kentucky was located on the edge of the current
overall range of a given weed. If it was, then Kentucky was likely to be the invasion front
of this specific invasive weed.
Associated land use types in the database were used to determine the predominant
land use type for each of the top 10 invasive species. Because land use type categories on
the weed identification forms were slightly different in various time periods, we

25
recategorized land use type into eight groups: Cropland, Pasture and Hayfields,
Lawn/Turf, Non-Cropland, Ornamental, Trees/Woodlot, Vegetable/Garden, and Others
(e.g., aquatic and roadside). The Cropland group was further divided into sub-categories
for additional analysis. To detect the invasive species predominant habitat, a total of 6483
records with explicit land use type were used to estimate the average proportion of each
land use type group. Proportion of associated land use type for each species was then
compared with the overall average proportion of each land use type in order to eliminate
some biases inherent in herbarium data by reducing the effect of collecting effort. For
example, although 35% records of Japanese knotweed (Polygonum cuspidatum Siebold
& Zucc.) were collected in Lawn/Turf land use type, we can’t confirm this invasive weed
is highly associated with this habitat because 32% of the UKWSIP records were collected
in Lawn/Turf (Table 2.3). If a given land use type for a specific invasive exotic species
has more than twice the percentage than average proportion, this land use type was
considered as the predominant habitat for this given invasive species. Based on the 928
records with Cropland land use type, a similar process was also implemented for the sub-
categories under the Cropland group to find the predominant crop type for the top 10
invasive species.
RESULTS
Overview of Invasive Weeds in Kentucky
Based on our survey, Kentucky is facing a large threat from exotic invasive weeds. A
total of 79 different exotic invasive weed species were reported 1,488 times during the 24
year time period (1986-2009) in Kentucky, covering 86% of the invasive plant species on

26
the KY-EPPC list. The top ten most frequently reported invasive exotic weeds were
ground ivy (Glechoma hederacea L.), Japanese knotweed (Polygonum cuspidatum Sieb.
& Zucc.), common lespedeza (Kummerowia striata (Thunb.) Schindl.), annual bluegrass
(Poa annua L.), common chickweed (Stellaria media (L.) Vill.), sericea lespedeza
(Lespedeza cuneata (Dumont) G. Don), Star-of-Bethlehem (Ornithogalum umbellatum
L.), barnyardgrass (Echinochloa crus-galli (L.) Beauv.), Canada thistle (Cirsium arvense
(L.) Scop.), and Japanese stiltgrass (Microstegium vimineum (Trin.) A. Camus) (Table
2.1). The majority of the invasive plants were herbaceous (68 spp.), followed by shrubs
and trees (7 and 4 spp., respectively). All 79 invasive species reported through the
UKWSIP belong to 31 families. Twenty-three percent of these invasive exotic species are
from the Poaceae family (18 species), the highest among all families, followed by the
Fabaceae, Asteraceae, and Lamiaceae families (9, 5, and 5 species, respectively) (Table
2.2). The rest of the families have four species or less, 18 families each has only one
invasive exotic weed.
The number of invasive exotic species reported was the highest right after written
records were kept through the weed science identification program in 1986, and
decreased as time went by. In the first four years (1986-89), 49 invasive species (62%)
were reported in 74 counties (Figure 2.1a). In the next 10 years (1990-99), 23 new
species were reported. After 2000, only seven species have been added to the database.
Exotic invasive weeds have been widely reported across Kentucky. All counties had
at least one exotic invasive species reported except for four counties (Figure 2.1b). Fayette
County, where Lexington, the second largest city in the Commonwealth is located, had
the most number of invasive exotic weed species (33) and contributed the most weed

27
reports (74) (Figure 2.1c). However, counties with a larger human population not
necessarily submitted more samples (Figure 2.1c, 2.1d). For example, Jefferson County
which had the largest population only had eight invasive weeds reported during 24 years
study period. We also noticed that counties with more invasive weed records often had
more total number of species reported.
Spatial and Temporal Distribution of the Top 10 Invasive Weeds
Spatial and temporal distribution maps created in our study revealed a clear pattern of
the top 10 exotic invasive weeds distribution. By comparing our database with
EDDMapS and PLANTS database, we found that Kentucky was likely to be the invasion
front for Canada thistle, Japanese stiltgrass, Japanese knotweed, and sericea lespedeza
(Figure 2.2). Canada thistle had invaded many states north of Kentucky (Figure 2.2a).
Our distribution map showed it was concentrated in the triangle area among Louisville,
Lexington, and Covington in northern Kentucky. Most recent reports of this weed were
mainly from the southern part of the state. The overall distribution indicates that southern
Kentucky is likely to be the current invasion front for Canada thistle. Japanese stiltgrass
was first introduced into Tennessee around 1919 (Fairbrot.De and Gray 1972b). It
currently exists across Tennessee and Virginia and also occupies the eastern and western
portion of Kentucky according to EDDMapS and PLANTS database (Figure 2.2b). Our
data filled the gap between the east and west while records from northern Kentucky were
absent, which indicates northern Kentucky is likely to be the invasion front for Japanese
stiltgrass. The distribution of Japanese knotweed was quite patchy, with most populations
being concentrated in the central and east of Kentucky (Figure 2.2c). As indicated by our

28
data, this species was widely distributed across Kentucky with a possible invasion front at
the southwestern portion of the state. Sericea lespedeza was prevalent across Kentucky
with a potential invasion front in northern Kentucky (Figure 2.2d).
The other six species are distributed widely in Kentucky and adjacent states. New
localities were reported simultaneously in different regions of the state (Figure 2.3). As
time passed by, these exotic weeds were then reported in surrounding counties from an
early location in a concentric candlewick pattern. Ground ivy was only reported in central
and northern Kentucky in EDDMapS and PLANTS database. Our data gave evidence
that this weed was also reported in other areas, which indicated ground ivy is prevalent
throughout Kentucky (Figure 2.3b). Star-of-Bethlehem and common chickweed were not
limited to a particular region of the state (Figure 2.3c, 2.3d). EDDMapS and PLANTS
database reported that annual bluegrass and barnyardgrass were scattered across the states
around Kentucky except for Ohio (Figure 2.3e, 2.3f). Our data further suggested that
these two weeds were currently prevalent across Kentucky. None of the above invasive
weeds was reported frequently in any particular region of the state.
Predominant Land Use Type for the Top 10 Invasive Weeds
The majority of the weeds reported in this database were from areas with high levels
of human impact. Three quarters of the invasive weeds reported were from Lawn/Turf
areas (32%), Pasture and Hayfields (32%), and Cropland (14%) habitats. Trees/Woodlot
habitat contributed less than one percent of the total case, which made it the rarest land
use type in our study. Each of the top 10 invasive weeds was associated with a dominant
land use type. Compared with the land use proportion from all records, ground ivy,

29
common lespedeza, and annual bluegrass were more frequently found in Lawn/Turf land
use areas (Table 2.3). Japanese knotweed, common chickweed, sericea lespedeza, and
star-of-Bethlehem were primarily found on Non-cropland, Vegetable/Garden, Pasture and
Hayfields, and Ornamental land use types, respectively. Whereas, barnyardgrass and
Canada thistle had high proportions in Cropland land use types. Although Japanese
stiltgrass had a high proportion in Trees/Woodlot, we can’t confirm it had a strong habitat
type preference because it had only two samples. In the subcategory of Cropland, Annual
bluegrass and barnyardgrass showed preference for wheat and soybean habitat,
respectively (Table 2.4).
DISCUSSION
Kentucky is facing a large threat from exotic invasive weeds. Seventy-nine declared
invasive plants from Kentucky Exotic Pest Plant Council’s list have been reported
through the UKWISP from 116 counties across Kentucky during the last 24 years. We
believe the invasive weeds, both in number of species, as well as frequency, were
underestimated because the weed identification service is on a voluntary base. Samples
were submitted by individuals through the local county Cooperative Extension Service
and forwarded to the UK Weed Science program. One example to support this point is
that kudzu (Pueraria montana var. lobata (Willd.) Maesen & S.M. Almeida) was only
reported once in our entire dataset. The likely explanation for the low reports number of
kudzu is that it is already well known as an exotic invasive species whereby either the
individual or the local county agent was able to identify.
Generally, after the initial introduction of an invasive species, the pattern of invasion

30
begins with a lag period of few collections followed by a period of rapid, exponential
expansion. Alien invasive species recently studied in France (Chauvel et al. 2006),
Quebec (Lavoie et al. 2007), and North America (Barney 2006) follow the same temporal
invasion pattern. Sixty-three percent of the invasive species were reported in the first four
years since the earliest records, which indicates these invasive plants have been in
Kentucky for more than 20 years. Further research about how long these invasive species
have been in Kentucky can be studied base on older records from herbaria or museums.
There is no significant correlation between population and occurrence of invasive
species based on our records. But, we still need to take into account the effect of local
county extension agents on the diversity and frequency of weed reports (Crawford and
Hoagland 2009). We suspect that we had more samples submitted for weed identification
from some counties than others partly because either the county extension agents in those
counties are less familiar with identifying and control of unwanted plants and forward
these samples for identification, or there are more people in those counties submitting
samples. It may depend on the specific plant species as to whether or not it is reported
frequently. Such submissions should be decreased as specific weeds become familiar for
the extension agents and the public. Other institutions offering similar plant identification
service such as botanical gardens might also have received weed reports which was not
included in our database. This may result in the lack of weed reports in the two east
counties, Pike and Martin because they may submit weed to the institutions in Virginia
for identification. In addition, we referenced the KY-EPPC list to define the invasive
species in our study, which does not include all invasive species. More species would be
recognized as invasive species if we referenced other regional or national invasive

31
species databases. Additional studies by referencing other invasive species databases
would bear their own significances.
Maps of plant distributions based on records from the UKWSIP, along with the other
two voluntary data, EDDMapS and PLANTS database, gave us a reasonably accurate
picture of the current distribution of a given invasive species within Kentucky. Baker
(1974) described the typical North American invasion pattern to be scattered populations
expanding to fill in absences between populations. The distributions of the top 10
invasive alien weeds in our study appear to follow this pattern. Plant distribution maps
will be more accurate as the number of plant collections increases. Given the short
history of the weed records embodied in the current weed identification system and the
limitations on how samples are submitted, analysis of the change in species distribution
over time can be somewhat unreliable. However, weed identification data still
contributed to sketching a big picture of current distribution of alien weeds. A good
illustration is that four of the top 10 species distribution maps indicate Kentucky was
likely to be the southern invasion front for Canada thistle and Japanese knotweed, and the
northern front for Japanese stiltgrass and sericea lespedeza (Figure 2.2). Plus, our
voluntary data provided more details for the exotic invasive weed distribution by filling
the gaps of other weed detection systems such as EDDMapS.
It’s not surprising that most of the weed reports were submitted from human related
land use types since this weed identification program was designed to detect weeds in
urban and agriculture areas where invasive species more likely gain attention from
people. Ground ivy, common lespedeza, and annual bluegrass have been reported as
common weeds in lawn turf (Busey 2003; Kohler et al. 2004b), and sericea lespedeza is

32
considered an usual invasive weed in the tall grass prairies of the Southern Great Plains
(Koger et al. 2002). Common chickweed is often found in gardens and arable areas
(Briggs et al. 1991). In additional, previous studies have reported that annual bluegrass
and barnyardgrass prefer wheat and soybean field, respectively (Borovickova 1992; Vail
and Oliver 1993; Wilson 1981). The associated habitats for the top 10 species revealed in
our study can assist in identifying the predominant land use/habitat type(s) of a given
invasive species, which helps natural resource managers to concentrate limited resources
to habitats that are highly susceptible to certain invasive species. It can also facilitate the
prediction of the spread of exotic invasive weeds by identifying the potential invasion
area.
The data resource we used has both strengths and weaknesses. The advantages
involve the nature of the source of weed identification database. It allows reports to be
submitted from more extensive areas in a shorter time, which is more economical and
efficient than herbarium collections and extensive field surveys. The disadvantages
include the bias for all such plant records, the nonrandom sampling bias, which is caused
by factors such as: unequal sampling effort over time and non-random geographical
representation. Poor location information in the specimens and incorrect identification
may reduce the accuracy of result. Since Kentucky’s forests cover 47 percent of the State
(Forest inventory 2004), the concentration on a particular urban and agriculture related
group of plants in this study may produced underrepresentation of forest related invasive
plants. Overall, these biases may lead to inadequate reports, resulting in both
underestimation of invasive species occurrence and disproportionately representation.
Our study suggests that weed identification system can be a valuable data resource to

33
reconstruct the historic distribution patterns of invasive species. It can help to fill gaps of
other invasive database such as EDDMapS and improve the accuracy of exotic invasive
species distribution maps. Similar weed identification services already exist in other
states. The wide application of these systems in invasive species EDRR systems will
greatly enhance our ability to combat exotic invasion.
The knowledge gained from this study would provide timely information to policy
makers in Kentucky and throughout the Southeast region in support of effective policy
development. The invasive species report frequency generated from our database could
be used to identify the priority invasive plant list. With predictive models, distribution
maps and habitat associations of given invasive species could help to determine at–risk
sites and to concentrate limited resource on the potential vulnerable areas to prevent
future invasion of exotic species. The UKWSIP forms could also reveal new exotic
invasive species which are not included in the KY-EPPC list so as to increase policy
makers and public awareness of new exotic species invasion.

34
Table 2.1 The top 10 most frequently reported invasive exotic weeds in University of
Kentucky Weed Science Identification Program (UKWSIP) since 1986.
Weed Species Common Name Total Reports
Glechoma hederacea L. ground ivy 99
Polygonum cuspidatum Sieb. & Zucc. Japanese knotweed 91
Kummerowia striata (Thunb.) Schindl. common lespedeza 86
Poa annua L. annual bluegrass 83
Stellaria media (L.) Vill. common chickweed 77
Lespedeza cuneata (Dumont) G. Don sericea lespedeza 62
Ornithogalum umbellatum L. star-of-Bethlehem 57
Echinochloa crus-galli (L.) Beauv. barnyardgrass 57
Cirsium arvense (L.) Scop. Canada thistle 53
Microstegium vimineum (Trin.) A. Camus Japanese stiltgrass 51

35
Table 2.2 Number of species within each family reported within the Kentucky Weed
Science Identification Program.
Number of species Family name
18 Poaceae
9 Fabaceae
5 Asteraceae, Lamiaceae
4 Polygonaceae
3 Brassicaceae, Celastraceae, Convolvulaceae, Rosaceae
2 Apiaceae, Caprifoliaceae, Chenopodiaceae, Liliaceae 1
Apocynaceae, Araliaceae, Boraginaceae, Caryophyllaceae, Clusiaceae, Commelinaceae, Dioscoreaceae, Dipsacaceae, Elaeagnaceae, Lardizabalaceae, Moraceae, Oleaceae, Oxalidaceae, Ranunculaceae, Salicaceae, Scrophulariaceae, Simaroubaceae, Solanaceae

36
Table 2.3 Percentage of major land use type associated with the top 10 invasive exotic weeds reported in UKWSIP. The
number with an asterisk (*) represents this land use type has more than twice of percentage than average.
Weed Species
Records (%)
Lawn/Turf
Pasture and
Hayfields Cropland Non-
Cropland Ornamental Vegetable/
Garden Other Trees/
Woodlot Total
Glechoma hederacea 65 (81*) 3 (4) 2 (3) 1 (1) 9 (11) 0 (0) 0 (0) 0 (0) 80
Polygonum cuspidatum 22 (35) 10 (16) 6 (10) 18 (29*) 2 (3) 3 (5) 1 (2) 0 (0) 62
Lespedeza striata 59 (82*) 10 (14) 1 (1) 0 (0) 2 (3) 0 (0) 0 (0) 0 (0) 72
Poa annua 48 (72*) 4 (6) 10 (15) 3 (4) 1 (1) 1 (1) 0 (0) 0 (0) 67
Stellaria media 38 (57) 11 (16) 0 (0) 2 (3) 3 (4) 10 (15*) 3 (4) 0 (0) 67
Lespedeza cuneata 1 (2) 38 (86*) 1 (2) 4 (9) 0 (0) 0 (0) 0 (0) 0 (0) 44
Ornithogalum umbellatum 30 (58) 8 (15) 6 (12) 2 (4) 6 (12*) 0 (0) 0 (0) 0 (0) 52
Echinochloa crus-galli 3 (7) 25 (58) 14 (33*) 0 (0) 0 (0) 0 (0) 1 (2) 0 (0) 43
Cirsium arvense 1 (2) 19 (42) 16 (36*) 1 (2) 5 (11) 1 (2) 2 (4) 0 (0) 45
Microstegium vimineum 19 (49) 12 (31) 0 (0) 4 (10) 1 (3) 1 (3) 0 (0) 2 (5*) 39
Total UKWSIP records 2079 (32) 2064 (32) 928 (14) 377 (6) 382 (6) 233 (4) 384 (6) 36 (0) 6483
36

37
Table 2.4 Percentage of predominant Cropland type associated with the top 10 invasive exotic weeds reported in UKWSIP.
The number with an asterisk (*) represents this land use type has more than twice of the number of case than average.
Area Collected (%)
Weed species Corn Soybean Tobacco Wheat Other Total case
Glechoma hederacea 0 (0) 2 (100) 0 (0) 0 (0) 0 (0) 2
Polygonum cuspidatum 4 (67) 1 (17) 1 (17) 0 (0) 0 (0) 6
Kummerowia striata 0 (0) 1 (100) 0 (0) 0 (0) 0 (0) 1
Poa annua 3 (30) 1 (10) 1 (10) 5 (50*) 0 (0) 10
Stellaria media 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0
Lespedeza cuneata 0 (0) 1 (100) 0 (0) 0 (0) 0 (0) 1
Ornithogalum umbellatum 3 (50) 1 (17) 1 (17) 1 (17) 0 (0) 6
Echinochloa crus-galli 2 (14) 9 (64*) 1 (7) 0 (0) 2 (14) 14
Cirsium arvense 9 (56) 2 (13) 4 (25) 1 (6) 0 (0) 16
Microstegium vimineum 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0
Total UKWSIP records 334 (36) 272 (29) 160 (17) 120 (13) 42 (5) 928
37

38
Figure 2.1 County level spatial distribution of invasive exotic weeds by (a) year first
reported, (b) total number of invasive species reported based on KY-EPPC list, (c) report
frequency based on UKWSIP database, and (d) the Kentucky population density (2000
Census data).

39
Figure 2.2 Spatial distribution of (a) Canada thistle (Cirsium arvense), (b) Japanese
stiltgrass (Microstegium vimineum), (c) Japanese knotweed (Polygonum cuspidatum) and
(d) sericea lespedeza (Lespedeza cuneata) at the county level. The strip shadow
represents weed distribution based on EDDMapS data and PLANTS database. Different
solid shadings indicate weed spread in different periods based on weed identification
data.

40
Figure 2.3 Spatial distribution of (a) common lespedeza (Kummerowia striata), (b)
ground ivy (Glechoma hederacea), (c) star-of-Bethlehem (Ornithogalum umbellatum),
(d) common chickweed (Stellaria media), (e) annual bluegrass (Poa annua), and (f)
barnyardgrass (Echinochloa crus-galli) at the county level. The strip shadow represents
weed distribution based on EDDMapS and PLANTS database. Different solid shadings
indicate weed spread in different periods based on weed identification data.

41
CHAPTER THREE: The Historical Distribution of Sixteen Exotic Invasive Species in Kentucky
based on Herbarium Records
SUMMARY
Regional distribution and abundance data for invasive plant species are urgently needed
for planning the management of invasive species, modeling of invasion risks and impacts,
and communicating the scope of the problem. Yet, detailed regional distribution data are
rare. In this project, historical distributions of 16 exotic invasive species in Kentucky
were reconstructed based on records from 15 Kentucky herbaria. Sampling locations and
dates were used to reveal the historical spatial distributions. The date of the first recorded
collection in each county was identified and the expansion pattern for each species was
examined. The majority of the invasive species in this study were first reported in
multiple locations over 50 years ago. The present distribution shows that the majority of
these 16 invasive species are scattered throughout Kentucky, indicating no concentrations
in particular regions. The cumulative curves for all species fitted a “J” shape expansion
curve, indicating the potential for further invasion in the future. This study demonstrates
the usefulness of herbarium data to estimate the distribution of invasive species and to
test hypothesis about their historical dynamics. Spatially explicit knowledge gained from
this study can aid natural resource managers to prevent and/or mitigate biological
invasion of exotic species.

42
INTRODUCTION
The introduction and spread of exotic invasive species into human-influenced and
natural environments has garnered increasing attention globally. The biology of invasive
species has attracted the interest of ecologists and conservation biologists (Ruiz and
Carlton 2003; Vila and Weiner 2004; Westbrooks et al. 1998; Wu et al. 2005). In spite of
the many studies that have been conducted, little is known regarding regional historical
dynamics of successful exotic species invasion (Callaway and Aschehoug 2000). Indeed,
increased human mobility over the past 100 years and the modification of the biosphere
has exposed disturbed habitats to invasive species and altered community structure and
ecosystem processes (Callaway and Maron 2006; Manchester and Bullock 2000).
Recently, the spatial distribution of exotic invasive species has become a hot topic
(Christen and Matlack 2009; Kelly et al. 2009; Liu et al. 2005; Merriam 2003). As
reported by Strayer et al.(2006), however, most invasive species studies have lacked a
temporal context, which can lead to unreliable or even controversial results. Long-term
data is needed to understand the attributes of the invasion process. To understand the
historical distribution of non-indigenous species, many researchers are turning to physical
or online herbarium collections (Lavoie et al. 2003; Lindgren 2003; Miller et al. 2009;
Wu et al. 2005; Zangerl and Berenbaum 2005). Although the data from herbarium
specimens suffer from several biases, they constitute a valuable source of information to
document the early stages of the invasion process (Crawford and Hoagland 2009; Lavoie
et al. 2007). Plant specimens stored in herbaria are numerous and usually well preserved.
Furthermore, most herbarium specimens have informative labels which indicate sampling
location, date, and the habitat type in which the plant was collected.
Currently, the Commonwealth of Kentucky is facing a significant threat from exotic

43
species invasion to its natural ecosystems. According to Jones (2005), nearly 25 percent
of vascular plants in Kentucky are non-indigenous. Kentucky Exotic Pest Plant Council
(KY- EPPC; http://www.se-eppc.org/ky/list.htm) also lists 92 plant species as threats to
local communities. Although research and extension programs to determine management
strategies exist in this region, no major effort exists for identifying temporal and spatial
distribution patterns of exotic invasive species. This paper describes the historical
distribution of 16 representative exotic invasive species in tree, shrub, vine, and herb
groups (Table 3.1) from the time of first recorded observation to the present status in
Kentucky. The historical distribution of each species was reconstructed using plant
specimens found in 15 herbaria in or around Kentucky dating back to 1870s (Appendix
Table 3.A). The cumulative number of total occupied counties was used to determine the
distribution rate for each species. The focus of this study is the utility of herbarium data
for the reconstruction of spatial and temporal distribution patterns of exotic invasive
species and the identification of factors contributing to the invasion of these exotic
species.
MATERIALS AND METHODS
This study is based exclusively on herbarium specimens from 15 sources (herbaria,
botanical gardens or databases of online herbaria; see Appendix Table 3.A) in and around
Kentucky. Since these herbaria have numerous exotic invasive plant specimens, only 16
typical species representing various growth forms and regions were included in this
study. The sampling location and year of collection of each invasive species specimen
was identified as precisely as possible. Due to the variability of spatial information

44
provided on herbarium specimen labels, all sampling locations were specified to county
level. Specimens of the same year and location were treated as one record. Nomenclature
in this study follows the Kentucky Exotic Pest Plant Council list. Specimens having
incomplete or imprecise information about the sampling location or collection date were
excluded from further analysis.
To reconstruct the historical distribution of exotic invasive species in Kentucky,
specimens in the resulting dataset were georeferenced and mapped using ArcGIS 9.3
(ESRI, Inc. Redlands, CA) with the time attributes attached. Because of the long
duration of herbarium records, a temporal resolution of one year was used. For each
invasive species, the earliest collection in each county was used to represent its temporal
dynamic. Historical distribution maps for each species were reconstructed at six time
periods, starting with the first record, to depict the spread of the species through time. In
order to compare the distribution rate of invasive exotic species introduced over time, we
categorized all invasive species based on the year they were first reported into three
groups: 1878-1909, 1910-1941, and 1942-1973. Distribution rates for invasive species
were estimated using cumulative numbers of total occupied counties through time based
on herbaria specimens and compared among three groups.
RESULTS
Herbaria specimens
There were a total of 2,233 specimens of the 16 exotic invasive plants in this study
kept in the 15 Kentucky herbaria. Several of these invasive species dated back 100 years
(Table 3.1). The earliest collection was a specimen of princesstree (Paulownia tomentosa

45
(Thunb.) Sieb. & Zucc. ex Steud. ) found in Fayette county in 1878. Common Reed
(Phragmites australis (Cav.) Trin. ex Steud.) was one of the more recent invasive species
within the herbarium collections (1973) in Kentucky. Based on herbaria specimens,
Japanese honeysuckle (Lonicera japonica Thunb.) was found in 100 counties across
Kentucky, which made it the most widespread invasive species in this study, whereas, the
presence of common reed was only recorded in 11 counties. The majority of the 16
invasive species were currently found throughout Kentucky (Figure 3.4, 3.5, 3.7-10,
Appendix Figure 3.C-I). The exceptions were Amur honeysuckle (Lonicera maackii
(Rupr.) Herder), which was restricted to northern and central Kentucky, kudzu (Pueraria
montana var. lobata (Willd.) Maesen & S.M. Almeida), which was limited to southern
Kentucky, and common reed, which was limited to western Kentucky (Figure 3.1-3). The
Herbarium at Eastern Kentucky University contributed 489 specimens, the biggest
collection among 15 institutions. The specimens from each herbarium were mainly
collected in the regions where herbarium located except for University of Kentucky
Weed Science Identification program whose collections cover most of the state (App.
Figure 3A, 3B). The detailed information on herbaria specimens was shown in Appendix
Table 3B.
Historical distribution
The maps generated from herbaria specimens indicated that the majority of the 16
invasive species illustrated no discernible spatial invasion or expansion pattern. However,
the historical distribution maps of Amur honeysuckle and tree-of-heaven (Ailanthus
altissima (P. Mill.) Swingle) suggested that each of these two species may have been

46
introduced in different regions of Kentucky simultaneously (Figure 3.1, 3.4). Based on
the historical distribution maps, tree-of-heaven was first found in western, central and
eastern Kentucky separately at an early time period (Figure 3.4). During the following
years, specimens of this species were generally collected in neighboring counties near the
first colonies. It suggested that the original invasion in western Kentucky was responsible
for the subsequent invasion in western Kentucky, while the central colonies accounted for
the invasion in central Kentucky. The first few collections of Amur honeysuckle were
found in central and northern Kentucky (Figure 3.1). Subsequent specimens were
collected in the surrounding counties of original colonies over the next 20 years.
Thereafter, these two separate colonies connected around 1997 and afterward spread
further. We also found that common reed and Japanese stiltgrass (Microstegium
vimineum (Trin.) A. Camus) were introduced in one particular region and then spread
from that area (Figure 3.3, 3.5). Specimens of common reed were first collected in 1973
in western Kentucky (Figure 3.3). After that, subsequent colonies of this species were
only found nearby, except for Kenton county in northern Kentucky. Somewhat
differently, Japanese stiltgrass was first introduced into Knoxville, TN in 1919 and was
first reported in Kentucky near the southeast border in 1931 (Figure 3.5). Referring to the
historical distribution map generated, the spread of this species was scattered throughout
Kentucky, however, it showed a remarkable invasion trend from southeast to northwest.
Distribution rate
Our data suggest the majority of invasive plants have few collections at their early
invasion period, and experienced a rapid expansion after that. The cumulative distribution

47
of occupied counties for all invasive species fitted a “J” shape curve. We found invasive
species first reported between 1910 and 1941 generally invaded more quickly than
species introduced in earlier or later time periods (Figure 3.6). Four species first reported
from 1929 to 1936, Japanese knotweed (Polygonum cuspidatum Sieb. & Zucc.), Japanese
stiltgrass, sericea lespedeza (Lespedeza cuneata (Dumont) G. Don), and multiflora rose
(Rosa multiflora Thunb. ex Murr.), expanded to a wider range than most alien species
found in Kentucky before 1910. The other two alien species in this group, climbing
euonymus (Euonymus fortunei (Turcz.) Hand.-Maz.) and autumn olive (Elaeagnus
umbellata Thunb.), occupied similar ranges to invasive plants that arrived in Kentucky at
an earlier time, indicating a faster rate of spread. All invasive species introduced before
1910 showed a constant increase over time except for Japanese honeysuckle. In addition,
the cumulative curves of occupied counties suggested no similarity among invasive
species first collected after 1942.
DISCUSSION
Herbarium specimens are an important source of information for reconstructing the
introduction and colonization of a species (Barney et al. 2008; Chauvel et al. 2006;
Lavoie et al. 2007). For example, Dessaint et al. (2005) used herbarium collections to
study the historical distribution of ragweed (Ambrosia artemisiifolia L.) in France.
Information from herbarium collections now became available on the internet, and can be
used to answer a wide range of ecological questions (Graham et al. 2004). Temporal and
geographical biases can exist in herbarium collections (Delisle et al. 2003). A temporal
bias may be present because of irregular collecting intensity. Collection intensity can

48
decline once the species is considered well represented in herbaria collections. But we
can still assume herbarium observations approximate the distribution of an invasive
species since the first specimens of most species investigated in this study can be traced
back to the beginning of last century, close to the time at which these species were first
recorded in North America. A geographical bias might be present because of non-random
geographical representation. The species sampled in herbaria usually reflect the flora of a
specific geographical area. Analysis of herbarium collections can potentially
underestimate the current range of invasive species and thus, the magnitude of their
ecological impact.
Using information provided by historical distribution maps of invasive species, we
can examine the invasion pattern and suggest explanations for the spread of these
invasive species. All exotic invasive plants in this study were introduced more than half a
century ago except for common reed, which was first reported in 1973 (Table 3.1). Initial
specimens of 7 invasive species were first collected more than 100 years ago. For most
species, the spatial and temporal distribution in Kentucky showed no clear front: new
locations separated by large distances were colonized simultaneously (Figure 3.2-4, 3.7-
10, Appendix Figure 3.C-I). Each species was introduced in various locations at different
periods of time, and spread in different directions.
We suggest a clear northwestward invasion pattern based on the historical distribution
maps for Japanese stiltgrass (Figure 3.5). This invasive species was introduced in
Knoxville, TN in 1919, as a packing material for porcelain (Kaufman and Kaufman
2007). According to the Early Detection and Distribution Mapping System (EDDMapS,
2010), the current distribution of this species was concentrated in Tennessee, Virginia,

49
and the western Carolinas. The first specimen of Japanese stiltgrass was collected in 1931
near the southeast boundary of Kentucky, which is adjacent to the original invasion site
and only 12 years later than its first appearance in the United States (Figure 3.5). It was
believed that these earlier colonies were responsible for the following northwestward
invasion although new locations, separated from the earlier colonies, were detected.
Recently, Flory (2010) reported the appearance of Japanese stiltgrass in southern Indiana,
which further suggests this species is expanding in a northeastward direction. A previous
study suggests that human disturbance, such as forest road management, is responsible
for the rapid spread of this invasive species (Rauschert et al. 2010). Future studies to
continue monitoring this species are needed to further confirm its spread north of
Kentucky.
Historical invasion maps generated from herbarium specimens of Amur honeysuckle
indicated a concentric circle candlewick pattern during the last 50 years (Figure 3.1).
Amur honeysuckle was first introduced in central and northern Kentucky. During the
following years, this exotic invasive species was generally found in areas surrounding the
original occupied counties. Based on this invasion pattern, areas surrounded by the
current range of Amur honeysuckle are potential vulnerable areas for invasion. This
species has been used as an ornamental (Harris et al. 2009), so it is not surprising that
Amur honeysuckle was mainly found near urban areas in central and northern Kentucky.
Amur honeysuckle produces abundant fruit which are attractive to bird and small
animals. Its appearance in rural areas may have resulted from seed dispersal by birds. The
limited, distant distribution of this species in western and southern Kentucky can also be
explained either by horticultural trade or the migration of birds. Overall, the distribution

50
pattern of Amur honeysuckle suggests human activity is responsible for the colonization
of this invasive species and dispersal by fruit-eating birds and horticultural activity is the
main cause of spread in this species.
The historical distributions of other species show no clear dispersal pattern during
their spread. For example, princesstree was first reported in central Kentucky before 1910. A
small number of specimens were collected in different regions of the state during the
following 60 years (Figure 3.7). Till 1970, the distribution of princesstree had been
scattered throughout the state. As time passed by, this invasive tree was generally
reported in surrounding counties from an early location. The current distribution of
princesstree were scattered throughout Kentucky, indicating no concentrations in
particular regions.
The invasion patterns of kudzu (Pueraria montana var. lobata (Willd.) Maesen &
S.M. Almeida), autumn olive, and common reed were primarily related to the fact that
they had been used to prevent soil erosion, reclaim mine spoil, or help with water
treatment (Batty and Younger 2004; Forseth and Innis 2004; Shields et al. 1995; Zipper
et al. 2007). Only one specimen of kudzu was collected before 1930s in Kentucky (Figure
3.2). In the 1930s and 1940s, the Soil Conservation Service promoted kudzu for erosion
control. After that, kudzu was commonly found scattered in distant regions of southern
Kentucky. The U.S. government stopped advocating the use of kudzu in 1953 after they
found this vine can also cause damage to the forests by preventing trees from obtaining
sunlight. The USDA later declared kudzu to be a weed in 1972 (Forseth and Innis 2004).
From then on, the established areas of kudzu in Kentucky expanded into surrounding
counties at a relative slow rate because it is limited to vegetative expansion by rhizomes

51
(Merriam 2003). Autumn olive was introduced in Kentucky in 1939. Only one specimen
of this species was recorded in the following 45 years (Figure 3.8). After the Surface
Mining Control and Reclamation Act of 1977, it has been widely used for strip mine
reclamation (Zipper et al. 2007). Based on herbaria specimens, its current distribution is
spread across western and eastern Kentucky coal fields. Another invasive species,
common reed, was mainly found in wetland habitat. It has been used for water treatment
(Batty and Younger 2004). This trait of common reed can explain why the collections of
common reed were generally restricted to wetlands in western Kentucky (Figure 3.3).
The spatial and temporal maps generated also suggest the present distribution of some
species is limited by their suitable habitat. Kudzu grows best where summer temperatures
are above 80º F, and winters are mild. This species was introduced in various points of
southern Kentucky over 100 years ago. Currently, kudzu is still concentrated in the south
which indicates the importance of latitude in limiting the spatial distribution of this
species (Cheng et al. 2007, Figure 3.2). Both Canada thistle (Cirsium arvense (L.) Scop.)
and musk thistle (Carduus nutans L.) usually grow in open area and along roadsides.
They compete with desirable forage and grain crops, and are problematic weeds on farms
and pastures (Reece and Wilson 1983). The present distributions of these two thistles
indicate the specimens were mostly collected in pasture, and are rare in eastern Kentucky,
which is dominated by forest (Figure 3.9, 3.10).
Generally, after the initial introduction of an invasive species, the pattern of invasion
begins with a lag period of few collections, followed by a period of rapid, exponential
expansion. The distribution rate held constant after invasive species completely occupied
in a region (Pysek and Richardson 2006; Stadler et al. 1998; Strayer et al. 2006). Exotic

52
invasive species studied in Quebec (Delisle et al. 2003) , France (Chauvel et al. 2006),
and North America (Barney 2006) followed this invasion pattern. Our data suggests a lag
period with a short, flat portion of the curve at the beginning of the time period for
invasive species (Figure 3.6). It appeared that most invasive species in this study were
still in their expansion period. The cumulative curves for all species were fitted the “J”
shape expansion curve, indicating a potential further invasion of these invasive plants. By
combining the invasion curve of species introduced in different time periods, we suggest
the early introduction doesn’t necessary contribute to its latter expansion (Figure 3.6).
The overall distribution trend for sixteen invasive species revealed a correspondence
between the rapid increase of invasive species and big events in U.S. history. First, during
the Great Depression in 1930s, the Agricultural Adjustment Act (AAA) of the New Deal
era restricted agricultural production by letting all farms lie fallow. These unmanaged
fields provided space for exotic species to invade, which might cause the first explosion
around 1930. At the end of World War II in 1945, the United States economy entered a
time of unprecedented growth. As a result, urbanization caused an increase in human-
disturbed habitat, which might lead to the second explosion of invasion (Addo-Fordjour
et al. 2009; Botham et al. 2009). Later, major highways such as I-75 and I-64, completed
in 1963 and 1976, respectively, perhaps facilitated the third expansion of existing
invasive species and might play an important role during the introduction of some exotic
species. Recently, the increasing volume and efficiency of local and global transportation
not only continue to introduce new exotic species from abroad, but also serve to spread
invasive species already present in a region (Colunga-Garcia et al. 2009).

53
CONCLUSION
This study on the historical distribution of 16 exotic invasive plants species in
Kentucky illustrates the usefulness of herbarium specimens in not only reconstructing the
distribution of invasive species but also by identifying potential expansion pathway. It
highlights the increasing importance of anthropological activities for non-indigenous
species invasion (King et al. 2009; Pejchar and Mooney 2009; Second and Rouhan 2008).
Herbaria are invaluable data sources for a wide range of ecological topics. We hope
studies such as ours will encourage others to take advantage of the information gathered
by botanists and to design novel research utilizing herbarium records.
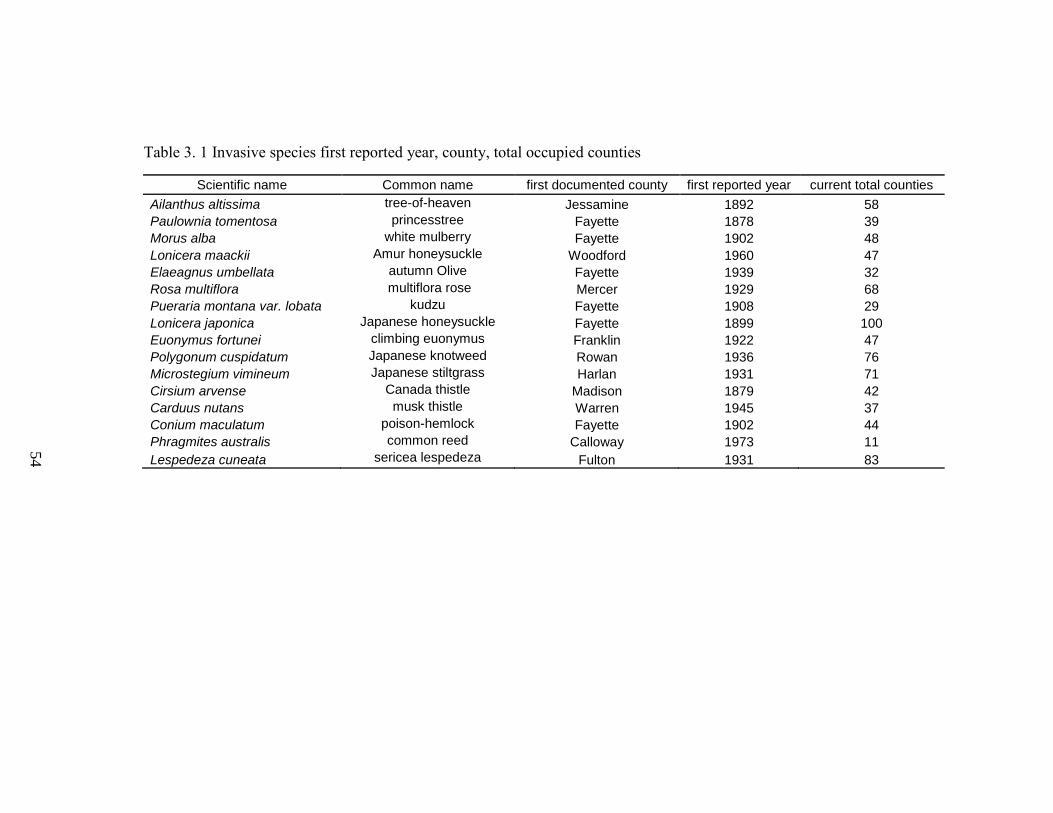
54
Table 3. 1 Invasive species first reported year, county, total occupied counties
Scientific name Common name first documented county first reported year current total counties
Ailanthus altissima tree-of-heaven Jessamine 1892 58
Paulownia tomentosa princesstree Fayette 1878 39
Morus alba white mulberry Fayette 1902 48
Lonicera maackii Amur honeysuckle Woodford 1960 47
Elaeagnus umbellata autumn Olive Fayette 1939 32
Rosa multiflora multiflora rose Mercer 1929 68
Pueraria montana var. lobata kudzu Fayette 1908 29
Lonicera japonica Japanese honeysuckle Fayette 1899 100
Euonymus fortunei climbing euonymus Franklin 1922 47
Polygonum cuspidatum Japanese knotweed Rowan 1936 76
Microstegium vimineum Japanese stiltgrass Harlan 1931 71
Cirsium arvense Canada thistle Madison 1879 42
Carduus nutans musk thistle Warren 1945 37
Conium maculatum poison-hemlock Fayette 1902 44
Phragmites australis common reed Calloway 1973 11
Lespedeza cuneata sericea lespedeza Fulton 1931 83
54

55
Figure 3.1 Historical distribution of Amur honeysuckle (Lonicera maackii (Rupr.)
Herder) in Kentucky based on herbaria data from 1960 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records.

56
Figure 3.2 Historical distribution of kudzu (Pueraria montana var. lobata (Willd.)
Maesen & S.M. Almeida) in Kentucky based on herbaria data from 1908 to 2008.
Occupied counties are shaded based on the time period during which the first specimen of
that species was recorded. Gray counties are the locations of the earlier plant records.

57
Figure 3.3 Historical distribution of common reed (Phragmites australis (Cav.) Trin. ex
Steud.) in Kentucky based on herbaria data from 1973 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records.

58
Figure 3.4 Historical distribution of tree-of-heaven (Ailanthus altissima (P. Mill.)
Swingle) in Kentucky based on herbaria data from 1892 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records.

59
Figure 3.5 Historical distribution of Japanese stiltgrass (Microstegium vimineum (Trin.)
A. Camus) in Kentucky based on herbaria data from 1931 to 2008. Occupied counties are
shaded based on the time period during which the first specimen of that species was
recorded. Gray counties are the locations of the earlier plant records.

60
Figure 3.6 Cumulative number of occupied counties for invasive species introduced from
(a) 1878 to 1909, (b) 1910 to 1941 and (c) 1942 to 1973.
0
10
20
30
40
50
60
70
80
90
100
1878 1898 1918 1938 1958 1978 1998
tree-of-heaven
pricesstree
white mulberry
Canada thistle
poison-hemlock
kudzu
Japanese honeysuckle
0
10
20
30
40
50
60
70
80
90
100
1878 1898 1918 1938 1958 1978 1998
climbing euonymus
Japanese knotweed
Nepalese browntop
sericea lespedeza
Multiflora rose
Autumn olive
0
10
20
30
40
50
60
70
80
90
100
1878 1898 1918 1938 1958 1978 1998
Amur honeysuckle
musk thistle
common reed
(a)
(b)
(c)

61
Figure 3.7 Historical distribution of princesstree (Paulownia tomentosa (Thunb.) Sieb. &
Zucc. ex Steud.) in Kentucky based on herbaria data from 1878 to 2008. Occupied
counties are shaded based on the time period during which the first specimen of that
species was recorded. Gray counties are the locations of the earlier plant records.

62
Figure 3.8 Historical distribution of autumn olive (Elaeagnus umbellata Thunb.) in
Kentucky based on herbaria data from 1939 to 2008. Occupied counties are shaded based
on the time period during which the first specimen of that species was recorded. Gray
counties are the locations of the earlier plant records.

63
Figure 3.9 Historical distribution of Canada thistle (Cirsium arvense (L.) Scop.) in
Kentucky based on herbaria data from 1879 to 2008. Occupied counties are shaded based
on the time period during which the first specimen of that species was recorded. Gray
counties are the locations of the earlier plant records.

64
Figure 3.10 Historical distribution of musk thistle (Carduus nutans L.) in Kentucky based
on herbaria data from 1945 to 2008. Occupied counties are shaded based on the time
period during which the first specimen of that species was recorded. Gray counties are
the locations of the earlier plant records.
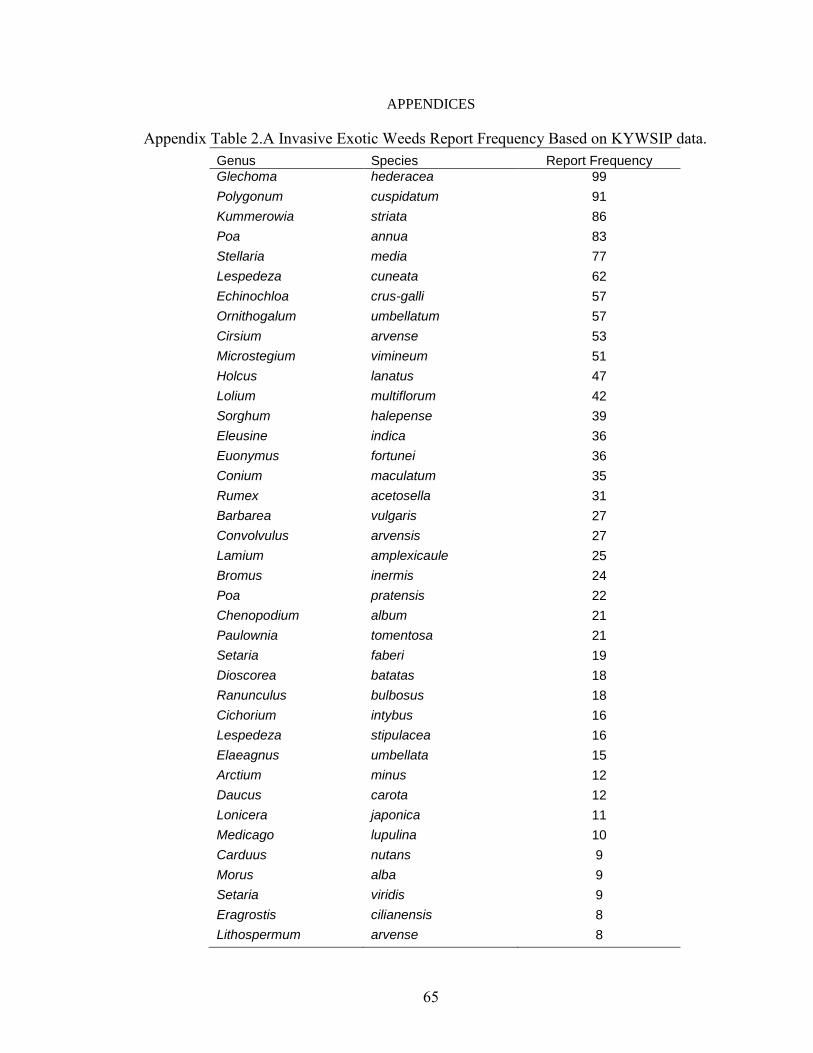
65
APPENDICES
Appendix Table 2.A Invasive Exotic Weeds Report Frequency Based on KYWSIP data.
Genus Species Report Frequency
Glechoma hederacea 99
Polygonum cuspidatum 91
Kummerowia striata 86
Poa annua 83
Stellaria media 77
Lespedeza cuneata 62
Echinochloa crus-galli 57
Ornithogalum umbellatum 57
Cirsium arvense 53
Microstegium vimineum 51
Holcus lanatus 47
Lolium multiflorum 42
Sorghum halepense 39
Eleusine indica 36
Euonymus fortunei 36
Conium maculatum 35
Rumex acetosella 31
Barbarea vulgaris 27
Convolvulus arvensis 27
Lamium amplexicaule 25
Bromus inermis 24
Poa pratensis 22
Chenopodium album 21
Paulownia tomentosa 21
Setaria faberi 19
Dioscorea batatas 18
Ranunculus bulbosus 18
Cichorium intybus 16
Lespedeza stipulacea 16
Elaeagnus umbellata 15
Arctium minus 12
Daucus carota 12
Lonicera japonica 11
Medicago lupulina 10
Carduus nutans 9
Morus alba 9
Setaria viridis 9
Eragrostis cilianensis 8
Lithospermum arvense 8
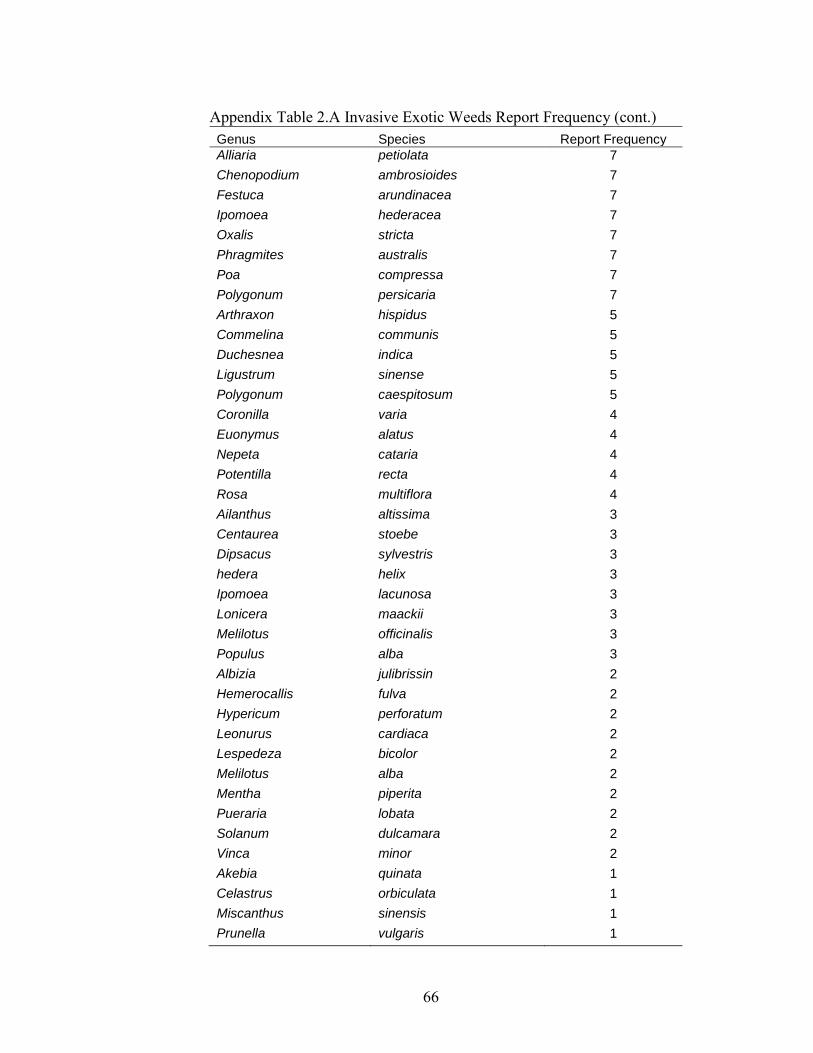
66
Appendix Table 2.A Invasive Exotic Weeds Report Frequency (cont.)
Genus Species Report Frequency
Alliaria petiolata 7
Chenopodium ambrosioides 7
Festuca arundinacea 7
Ipomoea hederacea 7
Oxalis stricta 7
Phragmites australis 7
Poa compressa 7
Polygonum persicaria 7
Arthraxon hispidus 5
Commelina communis 5
Duchesnea indica 5
Ligustrum sinense 5
Polygonum caespitosum 5
Coronilla varia 4
Euonymus alatus 4
Nepeta cataria 4
Potentilla recta 4
Rosa multiflora 4
Ailanthus altissima 3
Centaurea stoebe 3
Dipsacus sylvestris 3
hedera helix 3
Ipomoea lacunosa 3
Lonicera maackii 3
Melilotus officinalis 3
Populus alba 3
Albizia julibrissin 2
Hemerocallis fulva 2
Hypericum perforatum 2
Leonurus cardiaca 2
Lespedeza bicolor 2
Melilotus alba 2
Mentha piperita 2
Pueraria lobata 2
Solanum dulcamara 2
Vinca minor 2
Akebia quinata 1
Celastrus orbiculata 1
Miscanthus sinensis 1
Prunella vulgaris 1

67
Appendix Table 3.A Real and virtual Herbaria in and around Kentucky
Herbarium location Abbreviation
Berea College BEREA
Eastern Kentucky University EKY
Georgetown College GC
Mammoth Cave National Park Herbarium MCNPH
Missouri Botanical Garden MBG
Morehead State University MDKY
Murray State University MUR
Northern Kentucky University KNK
University of Kentucky KY
University of Louisville DHL
University of the Cumberlands UC
Weed Science Identification Program WSIP
Weed Science Herbarium at University of Kentucky WSH
Western Kentucky University WKY

68
Appendix Table 3.B Numbers of specimens for sixteen exotic invasive species in herbaria
Species name BEREA EKY GC MCNPH MBG MDKY MUR KNK KY DHL UC WSIP WSH WKU Total
Ailanthus altissima 32 39 1 0 2 15 4 14 27 4 0 3 6 2 149
Paulownia tomentosa 20 0 0 1 0 8 19 10 15 8 1 19 5 5 111
Morus alba 17 46 0 0 0 9 13 0 28 13 1 8 8 12 155
Lonicera maackii 9 64 0 0 5 0 0 31 2 0 0 3 0 0 114
Elaeagnus umbellata 30 0 0 0 1 10 0 8 5 1 3 14 1 0 73
Rosa multiflora 58 82 1 0 5 35 14 54 11 9 4 4 2 4 283 Pueraria montana var. lobata 4 20 0 1 0 12 6 0 0 1 2 1 2 3 52
Lonicera japonica 25 99 1 1 8 22 65 28 2 41 6 11 4 0 313
Euonymus fortunei 26 20 2 0 0 0 0 0 2 1 1 35 2 1 90
Polygonum cuspidatum 10 27 0 0 0 21 5 0 3 0 0 87 1 2 156
Microstegium vimineum 50 40 0 0 22 15 12 26 10 2 0 44 6 7 234
Cirsium arvense 8 9 0 0 2 5 0 5 1 0 0 51 16 0 97
Carduus nutans 8 0 0 0 4 4 7 6 6 2 0 8 4 9 58
Conium maculatum 5 0 1 0 2 8 2 9 12 1 1 30 20 5 96
Phragmites australis 0 3 0 0 1 0 2 2 0 0 0 7 0 0 15
Lespedeza cuneata 23 40 0 0 6 28 14 28 14 8 1 61 5 9 237
Total 325 489 6 3 58 192 163 221 138 91 20 386 82 59 2233
68

69
Appendix Figure 3.A Locations of herbaria in Kentucky.

70
Appendix Figure 3.B Sixteen Invasive plants records distribution by the major herbaria in
Kentucky.

71
Appendix Figure 3.C Historical distribution of sericea lespedeza in Kentucky based on
herbaria data from 1931 to 2008. Occupied counties are shaded based on the time period
during which the first specimen of that species was recorded. Gray counties are the
locations of the earlier plant records.

72
Appendix Figure 3.D Historical distribution of white mulberry in Kentucky based on
herbaria data from 1902 to 2008. Occupied counties are shaded based on the time period
during which the first specimen of that species was recorded. Gray counties are the
locations of the earlier plant records.

73
Appendix Figure 3.E Historical distribution of poison-hemlock in Kentucky based on
herbaria data from 1902 to 2008. Occupied counties are shaded based on the time period
during which the first specimen of that species was recorded. Gray counties are the
locations of the earlier plant records.

74
Appendix Figure 3.F Historical distribution of climbing euonymus in Kentucky based on
herbaria data from 1922 to 2008. Occupied counties are shaded based on the time period
during which the first specimen of that species was recorded. Gray counties are the
locations of the earlier plant records.

75
Appendix Figure 3.G Historical distribution of Japanese honeysuckle in Kentucky based
on herbaria data from 1899 to 2008. Occupied counties are shaded based on the time
period during which the first specimen of that species was recorded. Gray counties are
the locations of the earlier plant records.

76
Appendix Figure 3.H Historical distribution of Japanese knotweed in Kentucky based on
herbaria data from 1936 to 2008. Occupied counties are shaded based on the time period
during which the first specimen of that species was recorded. Gray counties are the
locations of the earlier plant records.

77
Appendix Figure 3.I Historical distribution of multiflora rose in Kentucky based on
herbaria data from 1929 to 2008. Occupied counties are shaded based on the time period
during which the first specimen of that species was recorded. Gray counties are the
locations of the earlier plant records.

78
REFERENCE
Able, K. W., Hagan, S. M., and Brown, S. A. 2003. Mechanisms of marsh habitat
alteration due to phragmites: Response of young-of-the-year Mummichog
(Fundulus heteroclitus) to treatment for Phragmites removal. Estuaries 26: 484-
494.
Adams, S. N., and Engelhardt, K. A. M. 2009. Diversity declines in Microstegium
vimineum (Japanese stiltgrass) patches. Biological Conservation 142: 1003-1010.
Addo-Fordjour, P., Obeng, S., Addo, M. G., and Akyeampong, S. 2009. Effects of human
disturbances and plant invasion on liana community structure and relationship
with trees in the Tinte Bepo forest reserve, Ghana. Forest Ecology and
Management 258: 728-734.
Ahmad, S. D., Sabir, S. M., and Zubair, A. 2006. Ecotypes diversity in autumn olive
(Elaeagnus umbellata Thunb): A single plant with multiple micronutrient genes.
Chem Ecol 22: 509-521.
Allaie, R. R., Reshi, Z., Rashid, I., and Wafai, B. A. 2006. Effect of aqueous leaf leachate
of Anthemis cotula - An alien invasive species on germination behaviour of some
field crops. J Agron Crop Sci 192: 186-191.
Allred, B. W., Fuhlendorf, S. D., Monaco, T. A., and Will, R. E. 2010. Morphological
and physiological traits in the success of the invasive plant Lespedeza cuneata.
Biological Invasions 12: 739-749.
Anderson, C. B., and Rosemond, A. D. 2007. Ecosystem engineering by invasive exotic
beavers reduces in-stream diversity and enhances ecosystem function in Cape
Horn, Chile. Oecologia 154: 141-153.
Assemat, L., Morishima, H., and Oka, H. I. 1981. Neighbor Effects between Rice (Oryza-
Sativa-L) and Barnyard Grass (Echinochloa-Crus-Galli Beauv) Acta Oecol-Oec
Plant 2: 63-78.
Baker, H. G. 1974. The Evolution of Weeds. Annual Review of Ecology and Systematics
5: 1-24.
Banasiak, S. E., and Meiners, S. J. 2009. Long term dynamics of Rosa multiflora in a
successional system. Biological Invasions 11: 215-224.
Barney, J. N. 2006. North American history of two invasive plant species:
phytogeographic distribution, dispersal vectors, and multiple introductions.
Biological Invasions 8: 703-717.
Barney, J. N., Whitlow, T. H., and Lembo, A. J. 2008. Revealing Historic Invasion
Patterns and Potential Invasion Sites for Two Non-Native Plant Species. PLoS
One 3(2): e1635.
Bartuszevige, A. M., and Gorchov, D. L. 2006. Avian seed dispersal of an invasive shrub.
Biological Invasions 8: 1013-1022.
Batty, L. C., and Younger, P. L. 2004. Growth of Phragmites australis (Cav.) Trin ex.
Steudel in mine water treatment wetlands: effects of metal and nutrient uptake.
Environmental Pollution 132: 85-93.
Beard, J. B., Rieke, P. E., Turgeon, A. J., and Vargas, J. M. 1978. Annual Bluegrass
(Poa-Annua L) Description, Adaptation, Culture and Control. Mich Aes Res Rep:
3-32.

79
Belde, M., and Mayer, F. 2002. Analysis of spatial distribution of Cirsium arvense (L).
Scop.: a simulation study. Z Pflanzenk Pflanzen: 367-374.
Bellingham, P. J., Towns, D. R., Cameron, E. K., Davis, J. J., Wardle, D. A., Wilmshurst,
J. M., and Mulder, C. P. H. 2010. New Zealand island restoration: seabirds,
predators, and the importance of history. New Zeal J Ecol 34: 115-136.
Belote, R. T., and Weltzin, J. F. 2006. Interactions between two co-dominant, invasive
plants in the understory of a temperate deciduous forest. Biological Invasions 8:
1629-1641.
Borgmann, K. L., and Rodewald, A. D. 2004. Nest predation in an urbanizing landscape:
The role of exotic shrubs. Ecological Applications 14: 1757-1765.
Borovickova, L. 1992. The Effect of Soil Cultivation on the Occurrence of Weeds in
Crops Included in Crop-Rotation. Rost Vyroba 38: 611-617.
Botham, M. S., Rothery, P., Hulme, P. E., Hill, M. O., Preston, C. D., and Roy, D. B.
2009. Do urban areas act as foci for the spread of alien plant species? An
assessment of temporal trends in the UK. Diversity and Distributions 15: 338-345.
Bourchier, R. S., and Van Hezewijk, B. H. 2010. Distribution and Potential Spread of
Japanese Knotweed (Polygonum cuspidatum) in Canada Relative to Climatic
Thresholds. Invasive Plant Science and Management 3: 32-39.
Bowman, W. C., and Sanghvi, I. S. 1963. Pharmacological Actions of Hemlock (Conium
Maculatum) Alkaloids. J Pharm Pharmacol 15: 25.
Boyer, C. R., Cole, J. C., and Payton, M. E. 2008. Survey of cultural practices used in
production of wintercreeper euonymus. Horttechnology 18: 158-161.
Bradley, B. A. 2010. Assessing ecosystem threats from global and regional change:
hierarchical modeling of risk to sagebrush ecosystems from climate change, land
use and invasive species in Nevada, USA. Ecography 33: 198-208.
Brantley, S. T., and Young, D. R. 2010. Linking light attenuation, sunflecks, and canopy
architecture in mesic shrub thickets. Plant Ecol 206: 225-236.
Briggs, D., Hodkinson, H., and Block, M. 1991. Precociously Developing Individuals in
Populations of Chickweed [Stellaria-Media (L) Vill] from Different Habitat
Types, with Special Reference to the Effects of Weed-Control Measures. New
Phytologist 117: 153-164.
Brisson, J., Paradis, E., and Bellavance, M. E. 2008. Evidence of sexual reproduction in
the invasive common reed (Phragmites Australis subsp Australis; Poaceae) in
eastern Canada: A possible consequence of global warming? Rhodora 110: 225-
230.
Brod, G. 1968. Studies on Biology and Ecology Barnyard Grass Echinochloa Crus-Galli
(L) Beauv. Weed Res 8: 115.
Burgess, K. S., and Husband, B. C. 2006. Habitat differentiation and the ecological costs
of hybridization: the effects of introduced mulberry (Morus alba) on a native
congener (M-rubra). J Ecol 94: 1061-1069.
Busey, P. 2003. Cultural management of weeds in turfgrass: A review. Crop Sci 43:
1899-1911.
Call, L. J., and Nilsen, E. T. 2005. Analysis of interactions between the invasive tree-of-
heaven (Ailanthus altissima) and the native black locust (Robinia pseudoacacia).
Plant Ecol 176: 275-285.
Callaway, R. M., and Aschehoug, E. T. 2000. Invasive plants versus their new and old

80
neighbors: A mechanism for exotic invasion. Science 290: 521-523.
Callaway, R. M., and Maron, J. L. 2006. What have exotic plant invasions taught us over
the past 20 years? Trends in Ecology & Evolution 21: 369-374.
Castellano, S. M., and Boyce, R. L. 2007. Spatial patterns of Juniperus virginiana and
Lonicera maackii on a road cut in Kentucky, USA. Journal of the Torrey
Botanical Society 134: 188-198.
Catling, P. M., and Carbyn, S. 2006. Recent invasion, current status and invasion
pathway of European Common Reed, Phragmites australis subspecies australis,
in the southern Ottawa district. Can Field Nat 120: 307-312.
Catling, P. M., Oldham, M. J., Sutherland, D. A., Brownell, V. R., and Larson, B. M. H.
1997. The recent spread of autumn-olive, Elaeagaus umbellata, into southern
Ontario and its current status. Can Field Nat 111: 376-380.
Chauvel, B., Dessaint, F., Cardinal-Legrand, C., and Bretagnolle, F. 2006. The historical
spread of Ambrosia artemisiifolia L. in France from herbarium records. Journal of
Biogeography 33: 665-673.
Cheng, Y. B., Tom, E., and Ustin, S. L. 2007. Mapping an invasive species, kudzu
(Pueraria montana), using hyperspectral imagery in western Georgia. Journal of
Applied Remote Sensing 1: 013514.
Cheplick, G. P. 2005. Biomass partitioning and reproductive allocation in the invasive,
cleistogamous grass Microstegium vimineum: Influence of the light environment.
Journal of the Torrey Botanical Society 132: 214-224.
Chornesky, E. A., Bartuska, A. M., Aplet, G. H., Britton, K. O., Cummings-Carlson, J.,
Davis, F. W., Eskow, J., Gordon, D. R., Gottschalk, K. W., Haack, R. A., Hansen,
A. J., Mack, R. N., Rahel, F. J., Shannon, M. A., Wainger, L. A., and Wigley, T.
B. 2005. Science priorities for reducing the threat of invasive species to
sustainable forestry. Bioscience 55: 335-348.
Christen, D. C., and Matlack, G. R. 2009. The habitat and conduit functions of roads in
the spread of three invasive plant species. Biological Invasions 11: 453-465.
Cole, P. G., and Weltzin, J. F. 2004. Environmental correlates of the distribution and
abundance of Microstegium vimineum, in east Tennessee. Southeastern Naturalist
3: 545-562.
Colunga-Garcia, M., Haack, R. A., and Adelaja, A. O. 2009. Freight Transportation and
the Potential for Invasions of Exotic Insects in Urban and Periurban Forests of the
United States. J Econ Entomol 102: 237-246.
Crawford, P. H. C., and Hoagland, B. W. 2009. Can herbarium records be used to map
alien species invasion and native species expansion over the past 100 years?
Journal of Biogeography 36: 651-661.
Cuneo, P., Jacobson, C. R., and Leishman, M. R. 2009. Landscape-scale detection and
mapping of invasive African Olive (Olea europaea L. ssp cuspidata Wall ex G.
Don Ciferri) in SW Sydney, Australia using satellite remote sensing. Appl Veg
Sci 12: 145-154.
D'Iorio, M., Jupiter, S. D., Cochran, S. A., and Potts, D. C. 2007. Optimizing remote
sensing and GIS tools for mapping and managing the distribution of an invasive
mangrove (Rhizophora mangle) on South Molokai, Hawaii. Mar Geod 30: 125-
144.
Davis, M. A. 2003. Biotic globalization: Does competition from introduced species

81
threaten biodiversity? Bioscience 53: 481-489.
Deering, R. H., and Vankat, J. L. 1999. Forest colonization and developmental growth of
the invasive shrub Lonicera maackii. Am Midl Nat 141: 43-50.
Delisle, F., Lavoie, C., Jean, M., and Lachance, D. 2003. Reconstructing the spread of
invasive plants: taking into account biases associated with herbarium specimens.
Journal of Biogeography 30: 1033-1042.
Demars, B. G. 1994. Star-of-Bethlehem, Ornithogalum-Umbellatum L (Liliaceae) - an
Invasive, Naturalized Plant in Woodlands of Ohio. Nat Area J 14: 306-307.
Demers, A. M., Backman, P. A., and Berner, D. K. 2005. Biological control of Canada
thistle (Cirsium arvense) using the rust Puccinia punctiformis. Phytopathology 95:
S156-S156.
Derr, J. F. 1989. Multiflora Rose (Rosa-Multiflora) Control with Metsulfuron. Weed
Technology 3: 381-384.
Dessaint, F., Chauvel, B., and Bretagnolle, F. 2005. Ragweed (Ambrosia artemisiifolia
L.): expansion history of a (biological pollutant) in France. M S-Med Sci 21: 207-
209.
Dickens, R., and Buchanan, G. A. 1971. Influence of Time of Herbicide Application on
Control of Kudzu. Weed Sci 19: 669.
Ding, J. Q., Wu, Y., Zheng, H., Fu, W. D., Reardon, R., and Liu, M. 2006. Assessing
potential biological control of the invasive plant, tree-of-heaven, Ailanthus
altissima. Biocontrol Sci Techn 16: 547-566.
Ede, F. J., Auger, M., and Green, T. G. A. 1997. Optimizing root cutting success in
Paulownia spp. J Hortic Sci 72: 179-185.
Edman, J. D. 2004. North American invasion, epidemic spread and control of West Nile
Virus. Abstr Pap Am Chem S 227: U73-U73.
Ehrenfeld, J. G., Kourtev, P., and Huang, W. Z. 2001. Changes in soil functions
following invasions of exotic understory plants in deciduous forests. Ecological
Applications 11: 1287-1300.
Engler, R., Guisan, A., and Rechsteiner, L. 2004. An improved approach for predicting
the distribution of rare and endangered species from occurrence and pseudo-
absence data. Journal of Applied Ecology 41: 263-274.
Epstein, A. H., Hill, J. H., and Nutter, F. W. 1997. Augmentation of rose rosette disease
for biocontrol of multiflora rose (Rosa multiflora). Weed Sci 45: 172-178.
Espenschied-Reilly, A. L., and Runkle, J. R. 2008. Distribution and changes in
abundance of Ailanthus altissima (Miller) swingle in a southwest Ohio woodlot.
Ohio J Sci 108: 16-22.
Essl, F. 2007. From ornamental to detrimental? The incipient invasion of Central Europe
by Paulownia tomentosa. Preslia 79: 377-389.
Fairbairn, J. W., and Challen, S. B. 1959. Alkaloids of Hemlock (Conium-Maculatum L)
- Distribution in Relation to the Development of the Fruit. Biochem J 72: 556-561.
Fairbrot.De, and Gray, J. R. 1972b. Microstegium-Vimineum (Trin) a-Camus (Gramineae)
in United-States. B Torrey Bot Club 99: 97.
Fasola, L., Chehebar, C., Macdonald, D. W., Porro, G., and Cassini, M. H. 2009. Do alien
North American mink compete for resources with native South American river
otter in Argentinean Patagonia? J Zool 277: 187-195.
Fernande, R. 1971. Glechoma hederacea L. Bot J Linn Soc 64: 72.

82
Flory, S. L. 2010. Management of Microstegium vimineum Invasions and Recovery of
Resident Plant Communities. Restor Ecol 18: 103-112.
Flory, S. L., and Clay, K. 2009. Effects of roads and forest successional age on
experimental plant invasions. Biological Conservation 142: 2531-2537.
Foote, L. E., Kill, D. L., and Williams, C. S. 1970. Canada Thistle Control on Roadsides.
Weed Sci 18: 307.
Forman, J., and Kesseli, R. V. 2003. Sexual reproduction in the invasive species Fallopia
japonica (Polygonaceae). Am J Bot 90: 586-592.
Forseth, I. N., and Innis, A. F. 2004. Kudzu (Pueraria montana): History, physiology,
and ecology combine to make a major ecosystem threat. Crit Rev Plant Sci 23:
401-413.
Gadella, T. W. J. 1972. Some Notes on Ornithogalum-Umbellatum L and Ornithogalum-
Divergens Bor. Acta Bot Neerl 21: 257.
Ghersa, C. M., de la Fuente, E., Suarez, S., and Leon, R. J. C. 2002. Woody species
invasion in the Rolling Pampa grasslands, Argentina. Agr Ecosyst Environ 88:
271-278.
Goeden, R. D., and Ricker, D. W. 1982. Poison Hemlock, Conium-Maculatum, in
Southern-California - an Alien Weed Attacked by Few Insects. Ann Entomol Soc
Am 75: 173-176.
Goldstein, C. L., Williard, K. W. J., and Schoonover, J. E. 2009. Impact of an Invasive
Exotic Species on Stream Nitrogen Levels in Southern Illinois. J Am Water
Resour As 45: 664-672.
Goodell, K., McKinney, A. M., and Lin, C. H. 2010. Pollen Limitation and Local
Habitat-Dependent Pollinator Interactions in the Invasive Shrub Lonicera Maackii.
Int J Plant Sci 171: 63-72.
Graham, C. H., Ferrier, S., Huettman, F., Moritz, C., and Peterson, A. T. 2004. New
developments in museum-based informatics and applications in biodiversity
analysis. Trends in Ecology & Evolution 19: 497-503.
Grubisic, D., Neskovic, M., and Konjevic, R. 1985. Changes in Light Sensitivity of
Paulownia-Tomentosa and Paulownia-Fortunei Seeds. Plant Sci 39: 13-16.
Harris, C., Jiang, H., Liu, D. J., Brian, Z., and He, K. T. 2009. Testing the roles of species
native origin and family membership in intentional plant introductions using
nursery data across the state of Kentucky. Journal of the Torrey Botanical Society
136: 122-127.
Hartman, K. M., and McCarthy, B. C. 2004. Restoration of a forest understory after the
removal of an invasive shrub, Amur honeysuckle (Lonicera maackii). Restor Ecol
12: 154-165.
Hartman, K. M., and McCarthy, B. C. 2008. Changes in forest structure and species
composition following invasion by a non-indigenous shrub, Amur honeysuckle
(Lonicera maackii). Journal of the Torrey Botanical Society 135: 245-259.
Hayden, A. 1934. Distribution and reproduction of Canada thistle in Iowa. Am J Bot 21:
355-373.
Heisey, R. M. 1990. Evidence for Allelopathy by Tree-of-Heaven (Ailanthus-Altissima).
J Chem Ecol 16: 2039-2055.
Hershman, D. E., Bachi, P. R., Harmon, C. L., Harmon, P. F., Palm, M. E., McKemy, J.
M., Zeller, K. A., and Levy, L. 2006. First report of soybean rust caused by

83
Phakopsora pachyrhizi on Kudzu (Pueraria montana var. lobata) in Kentucky.
Plant Dis 90: 834-834.
Higgins, S. I., Richardson, D. M., Cowling, R. M., and Trinder-Smith, T. H. 1999.
Predicting the Landscape-Scale Distribution of Alien Plants and Their Threat to
Plant Diversity. Conservation Biology 13: 303-313.
Hindal, D. F., and Wong, S. M. 1988. Potential Biocontrol of Multiflora Rose, Rosa-
Multiflora. Weed Technology 2: 122-131.
Huang, C. Y., and Asner, G. P. 2009. Applications of Remote Sensing to Alien Invasive
Plant Studies. Sensors-Basel 9: 4869-4889.
Hulme, P. E. 2006. Beyond control: wider implications for the management of biological
invasions. Journal of Applied Ecology 43: 835-847.
Hunt, D. M., and Zaremba, R. E. 1992. the Northeastward spread of Microstegium-
Vimineum (Poaceae) into New-York and adjacent states. Rhodora 94: 167-170.
Hyatt, L. A., and Casper, B. B. 2000. Seed bank formation during early secondary
succession in a temperate deciduous forest. J Ecol 88: 516-527.
Inderjit, and Dakshini, K. M. M. 1998. Allelopathic interference of chickweed, Stellaria
media with seedling growth of wheat (Triticum aestivum). Can J Bot 76: 1317-
1321.
Johnson, B. J. 1979. Annual Bluegrass and Broadleaf Weed-Control in Bermudagrass
Turf. Geo Agr Exp St Re Re: 3-14.
Jones, R. L. 2005. Plant life of Kentucky: an illustrated guide to the vascular
flora.Lexington, Ky: The University Press of Kentucky.
Jongejans, E., Shea, K., Skarpaas, O., Kelly, D., Sheppard, A. W., and Woodburn, T. L.
2008. Dispersal and demography contributions to population spread of Carduus
nutans in its native and invaded ranges. J Ecol 96: 687-697.
Jongejans, E., Sheppard, A. W., and Shea, K. 2006. What controls the population
dynamics of the invasive thistle Carduus nutans in its native range? Journal of
Applied Ecology 43: 877-886.
Kaproth, M. A., and McGraw, J. B. 2008. Seed viability and dispersal of the wind-
dispersed invasive Ailanthus altissima in aqueous environments. Forest Sci 54:
490-496.
Kaufman, S. R., and Kaufman, W. 2007. Invasive plants : a guide to identification and
the impacts and control of common North American species. 1st ed.
Mechanicsburg, PA: Stackpole Books. Pp. v, 458 p.
Kelly, A. B., Small, C. J., and Dreyer, G. D. 2009. Vegetation classification and invasive
species distribution in natural areas of southern New England. Journal of the
Torrey Botanical Society 136: 500-519.
Kennedy, R. A., Barrett, S. C. H., Vanderzee, D., and Rumpho, M. E. 1980. Germination
and Seedling Growth under Anaerobic Conditions in Echinochloa-Crus-Galli
(Barnyard Grass). Plant Cell Environ 3: 243-248.
Kimothi, M. M., Anitha, D., Vasistha, H. B., Soni, P., and Chandola, S. K. 2010. Remote
sensing to map the invasive weed, Lantana camara in forests. Trop Ecol 51: 67-
74.
King, J. R., Tschinkel, W. R., and Ross, K. G. 2009. A case study of human exacerbation
of the invasive species problem: transport and establishment of polygyne fire ants
in Tallahassee, Florida, USA. Biological Invasions 11: 373-377.

84
Knapp, L. B., Fownes, J. H., and Harrington, R. A. 2008. Variable effects of large
mammal herbivory on three non-native versus three native woody plants. Forest
Ecology and Management 255: 92-98.
Knight, A. P. 1987. Poison Hemlock. Comp Cont Educ Pract 9: F256-F257.
Koger, C. H., Stritzke, J. E., and Cummings, D. C. 2002. Control of sericea lespedeza
(Lespedeza cuneata) with triclopyr, fluroxypyr, and metsulfuron. Weed
Technology 16: 893-900.
Kohler, E. A., Throssell, C. S., and Reicher, Z. J. 2004a. Cultural and chemical control of
ground ivy (Glechoma hederacea). Hortscience 39: 1148-1152.
Kohler, E. A., Throssell, C. S., and Reicher, Z. J. 2004b. Ground ivy (Glechoma
hederacea) populations respond differently to 2,4-D or triclopyr. Weed
Technology 18: 566-574.
Koshy, T. K. 1969. Breeding Systems in Annual Bluegrass Poa Annua L. Crop Sci 9: 40.
Kourtev, P. S., Ehrenfeld, J. G., and Huang, W. Z. 1998. Effects of exotic plant species
on soil properties in hardwood forests of New Jersey. Water Air Soil Poll 105:
493-501.
Kowarik, I. 2003. Human agency in biological invasions: secondary releases foster
naturalisation and population expansion of alien plant species. Biological
invasions 5: 293
Kowarik, I., and Saumel, I. 2008. Water dispersal as an additional pathway to invasions
by the primarily wind-dispersed tree Ailanthus altissima. Plant Ecol 198: 241-252.
Kwong, K. L., Chan, R. K. Y., and Qiu, J. W. 2009. The Potential of the Invasive Snail
Pomacea Canaliculata as a Predator of Various Life-Stages of Five Species of
Freshwater Snails. Malacologia 51: 343-356.
Lalonde, R. G., and Roitberg, B. D. 1994. Mating System, Life-History, and
Reproduction in Canada Thistle (Cirsium-Arvense Asteraceae). Am J Bot 81: 21-
28.
Lamont, E. E., and Young, S. M. 2002. Noteworthy plants reported from the Torrey
Range - 2001. Journal of the Torrey Botanical Society 129: 363-371.
Landenberger, R. E., Kota, N. L., and McGraw, J. B. 2007. Seed dispersal of the non-
native invasive tree Ailanthus altissima into contrasting environments. Plant Ecol
192: 55-70.
Langdon, K. R., and Johnson, K. D. 1994. Additional Notes on Invasiveness of
Paulownia-Tomentosa in Natural Areas. Nat Area J 14: 139-140.
Laubhan, M. K., and Shaffer, T. L. 2006. Seed germination of Cirsium arvense and
Lepidium latifolium: Implications for management of Montane wetlands.
Wetlands 26: 69-78.
Lavoie, C., Jean, M., Delisle, F., and Letourneau, G. 2003. Exotic plant species of the St
Lawrence River wetlands: a spatial and historical analysis. Journal of
Biogeography 30: 537-549.
Lavoie, C., Jodoin, Y., and de Merlis, A. G. 2007. How did common ragweed (Ambrosia
artemisiifolia L.) spread in Quebec? A historical analysis using herbarium records.
Journal of Biogeography 34: 1751-1761.
Legere, A. 2009. Weed science at the core of 21st century issues. Phytoprotection 90: 5-
11.
Lelong, B., Lavoie, C., Jodoin, Y., and Belzile, F. 2007. Expansion pathways of the

85
exotic common reed (Phragmites australis): a historical and genetic analysis.
Diversity and Distributions 13: 430-437.
Lindgren, C. J. 2003. A brief history of Purple Loosestrife, Lythrum salicaria, in
Manitoba and its status in 2001. Can Field Nat 117: 100-109.
Liu, J., Liang, S. C., Liu, F. H., Wang, R. Q., and Dong, M. 2005. Invasive alien plant
species in China: regional distribution patterns. Diversity and Distributions 11:
341-347.
Lodge, D. M., Williams, S., MacIsaac, H. J., Hayes, K. R., Leung, B., Reichard, S., Mack,
R. N., Moyle, P. B., Smith, M., Andow, D. A., Carlton, J. T., and McMichael, A.
2006. Biological invasions: recommendations for U.S. policy and management.
Ecol Appl 16: 2035-54.
Logan, R. H., Hoveland, C. S., and Donnelly, E. D. 1969. A Germination Inhibitor in
Seedcoat of Sericea (Lespedeza Cuneata (Dumont.) G. Don.). Agronomy Journal
61: 265.
Lopez, T. A., Cid, M. S., and Bianchini, M. L. 1999. Biochemistry of hemlock (Conium
maculatum L.) alkaloids and their acute and chronic toxicity in livestock. A
review. Toxicon 37: 841-865.
Luken, J. O., and Goessling, N. 1995. Seedling Distribution and Potential Persistence of
the Exotic Shrub Lonicera-Maackii in Fragmented Forests. Am Midl Nat 133:
124-130.
Luken, J. O., and Mattimiro, D. T. 1991. Habitat-Specific Resilience of the Invasive
Shrub Amur Honeysuckle (Lonicera-Maackii) during Repeated Clipping.
Ecological Applications 1: 104-109.
Luken, J. O., and Thieret, J. W. 1996. Amur honeysuckle, its fall from grace. Bioscience
46: 18-24.
Main, C. L., Robinson, D. K., Teuton, T. C., and Mueller, T. C. 2004. Star-of-Bethlehem
(Ornithogalum umbellatum) control with postemergence herbicides in dormant
bermudagrass (Cynodon dactylon) turf. Weed Technology 18: 1117-1119.
Manchester, S. J., and Bullock, J. M. 2000. The impacts of non-native species on UK
biodiversity and the effectiveness of control. Journal of Applied Ecology 37: 845-
864.
Maun, M. A. 1977. Suppressing Effect of Soybeans on Barnyard Grass. Can J Plant Sci
57: 485-490.
McCay, T. S., McCay, D. H., and Czajka, J. L. 2009. Deposition of exotic bird-dispersed
seeds into three habitats of a fragmented landscape in the northeastern United
States. Plant Ecol 203: 59-67.
McNatty, A., Abbott, K. L., and Lester, P. J. 2009. Invasive ants compete with and
modify the trophic ecology of hermit crabs on tropical islands. Oecologia 160:
187-194.
Meadows, R. E., and Saltonstall, K. 2007. Distribution of native and introduced
Phragmites australis in freshwater and oligohaline tidal marshes of the Delmarva
Peninsula and southern New Jersey. Journal of the Torrey Botanical Society 134:
99-107.
Medley, K. E. 1997. Distribution of the non-native shrub Lonicera maackii in Kramer
Woods, Ohio. Phys Geogr 18: 18-36.
Mehrhoff, L. J. 2000. Perennial Microstegium vimineum (Poaceae): An apparent

86
misidentification? Journal of the Torrey Botanical Society 127: 251-254.
Meiners, S. J., and LoGiudice, K. 2003. Temporal Consistency in the Spatial Pattern of
Seed Predation across a Forest: Old Field Edge. Plant Ecol 168: 45-55.
Merriam, R. W. 2003. The abundance, distribution and edge associations of six non-
indigenous, harmful plants across North Carolina. Journal of the Torrey Botanical
Society 130: 283-291.
Metcalf, C. J. E., Rees, M., Buckley, Y. M., and Sheppard, A. W. 2009. Seed predators
and the evolutionarily stable flowering strategy in the invasive plant, Carduus
nutans. Evol Ecol 23: 893-906.
Miller, J. a. E., B. . 1983. Kudzu: Where did it come from? And how can we stop it?
Southern Journal of Applied Forestry 7: 165.
Miller, K. E., and Gorchov, D. L. 2004. The invasive shrub, Lonicera maackii, reduces
growth and fecundity of perennial forest herbs. Oecologia 139: 359-375.
Miller, R. J., Carroll, A. D., Wilson, T. P., and Shaw, J. 2009. Spatiotemporal Analysis of
Three Common Wetland Invasive Plant Species Using Herbarium Specimens and
Geographic Information Systems. Castanea 74: 133-145.
Minchinton, T. E. 2006. Rafting on wrack as a mode of dispersal for plants in coastal
marshes. Aquatic Botany 84: 372-376.
Mitich, L. W. 1994. Ground Ivy. Weed Technology 8: 413-415.
Miura, R., and Kusanagi, T. 2001. Variation in the factors determining flowering time in
the Stellaria media complex. Weed Res 41: 69-81.
Molocznik, A. 2004. Time of farmers' exposure to biological factors in agricultural
working environment. Ann Agr Env Med 11: 85-89.
Moore, J. E., and Lacey, E. P. 2009. A Comparison of Germination and Early Growth of
Four Early Successional Tree Species of the Southeastern United States in
Different Soil and Water Regimes. Am Midl Nat 162: 388-394.
Moore, W. B., Doyle, C. J., and Rahman, A. 1989. Economics of Controlling Carduus-
Nutans on Grazed Pasture in New-Zealand. Crop Prot 8: 16-24.
Moret, J., Favereau, Y., and Gorenflot, R. 1991. A Biometric Study of the Ornithogalum-
Umbellatum (Hyacinthaceae) Complex in France. Plant Syst Evol 175: 73-86.
Morse, L. E., Kartesz, J. T., and Kutner, L. S. . 1995. Vascular plants of the United States.
Washington, DC.: U.S. Department of the Interior, National Biological Service.
200-205 p.
Mueller, J. P., Luginbuhl, J. M., and Bergmann, B. A. 2001. Establishment and early
growth characteristics of six Paulownia genotypes for goat browse in Raleigh, NC,
USA. Agroforestry Systems 52: 63-72.
Nakatsubo, T. 1995. Factors Limiting the Distribution of the Annual Legume
Kummerowia-Striata in a River Floodplain. Ecol Res 10: 179-187.
Newton, C. H., Nelson, L. R., Dewalt, S. J., Mikhailova, E. A., Post, C. J., Schlautman,
M. A., Cox, S. K., Bridges, W. C., and Hall, K. C. 2008. Solarization for the
control of Pueraria montana (kudzu). Weed Res 48: 394-397.
Noe, M. D. 2009. White Mulberry Economics in Willa Cather's Nebraska. Anq-Q J Short
Art N 22: 30-36.
Orr, M. S., Goodsaid, F., Amur, S., Rudman, A., and Frueh, F. W. 2007. The experience
with voluntary genomic data submissions at the FDA and a vision for the future of
the Voluntary Data Submission Program. Clin. Pharmacol. Ther. 81: 294-297.

87
Pejchar, L., and Mooney, H. A. 2009. Invasive species, ecosystem services and human
well-being. Trends in Ecology & Evolution 24: 497-504.
Peschken, D. P., and Wilkinson, A. T. S. 1981. Biocontrol of Canada Thistle (Cirsium-
Arvense) - Releases and Effectiveness of Ceutorhynchus-Litura (Coleoptera,
Curculionidae) in Canada. Can Entomol 113: 777-785.
Pichlova-Ptacnikova, R., and Vanderploeg, H. A. 2009. The invasive cladoceran
Cercopagis pengoi is a generalist predator capable of feeding on a variety of prey
species of different sizes and escape abilities. Fund Appl Limnol 173: 267-279.
Piggott, M. P., Wilson, R., Banks, S. C., Marks, C. A., Gigliotti, F., and Taylor, A. C.
2008. Evaluating exotic predator control programs using non-invasive genetic
tagging. Wildlife Res 35: 617-624.
Pimentel, D., Zuniga, R., and Morrison, D. 2005. Update on the environmental and
economic costs associated with alien-invasive species in the United States.
Ecological Economics 52: 273-288.
Ponder, W. F., Carter, G. A., Flemons, P., and Chapman, R. R. 2001. Evaluation of
museum collection data for use in biodiversity assessment. Conservation Biology
15: 648-657.
Popay, A. I., Butler, J. H., and Meeklah, F. A. 1989. Chemical Control of Nodding
Thistle (Carduus-Nutans L.) in New-Zealand Pastures. Weed Res 29: 21-28.
Pysek, P., and Richardson, D. M. 2006. The biogeography of naturalization in alien
plants. Journal of Biogeography 33: 2040-2050.
Rauschert, E. S. J., Mortensen, D. A., Bjornstad, O. N., Nord, A. N., and Peskin, N. 2010.
Slow spread of the aggressive invader, Microstegium vimineum (Japanese
stiltgrass). Biological Invasions 12: 563-579.
Reece, P. E., and Wilson, R. G. 1983. Effect of Canada Thistle (Cirsium-Arvense) and
Musk Thistle (Carduus-Nutans) Control on Grass Herbage. Weed Sci 31: 488-
492.
Rehder, A. 1993. Manual of cultivated trees and shrubsPortland, OR: Dioscorides Press.
Reichard, S. H., and White, P. 2001. Horticulture as a pathway of invasive plant
introductions in the United States. Bioscience 51: 103-113.
Reid, A. M., Morin, L., Downey, P. O., French, K., and Virtue, J. G. 2009. Does invasive
plant management aid the restoration of natural ecosystems? Biological
Conservation 142: 2342-2349.
Rejmanek, M. 1996. A theory of seed plant invasiveness: The first sketch. Biological
Conservation 78: 171-181.
Rejmanek, M., and Richardson, D. M. 1996. What attributes make some plant species
more invasive? Ecology 77: 1655-1661.
Ries, P., Dix, M. E., Ielmini, M., and Thomas, D. 2004. National Strategy and
Implementation Plan for Invasive Species Management. Washington DC. : USDA
Forest Service. 17 p.
Robertson, D. J., Robertson, M. C., and Tague, T. 1994. Colonization dynamics of 4
exotic plants in a northern piedmont natural area. B Torrey Bot Club 121: 107-
118.
Rossman, A. Y. 2001. A special issue on global movement of invasive plants and fungi.
Bioscience 51: 93-94.
Ruiz, G. M., and Carlton, J. T. 2003. Invasive species : vectors and management

88
strategiesWashington, DC: Island Press. Pp. xiii, 518 p.
Schierenbeck, K. A. 2004. Japanese honeysuckle (Lonicera japonica) as an invasive
species; History, ecology, and context. Crit Rev Plant Sci 23: 391-400.
Schmida, T. O. 1974. Poison Hemlock. Western J Med 120: 77-78.
Second, G., and Rouhan, G. 2008. Human-Mediated Emergence as a Weed and Invasive
Radiation in the Wild of the CD Genome Allotetraploid Rice Species (Oryza,
Poaceae) in the Neotropics. PLoS One 3(7): e2613.
Shea, K., Sheppard, A., and Woodburn, T. 2006. Seasonal life-history models for the
integrated management of the invasive weed nodding thistle Carduus nutans in
Australia. Journal of Applied Ecology 43: 517-526.
Shields, F. D., Bowie, A. J., and Cooper, C. M. 1995. Control of Streambank Erosion
Due to Bed Degradation with Vegetation and Structure. Water Resour Bull 31:
475-489.
Shotwell, R. 1993. The History and Folklore of North-American Wildflowers. Libr J 118:
90-90.
Snyder, B. A., Boots, B., and Hendrix, P. F. 2009. Competition between invasive
earthworms (Amynthas corticis, Megascolecidae) and native North American
millipedes (Pseudopolydesmus erasus, Polydesmidae): Effects on carbon cycling
and soil structure. Soil Biol Biochem 41: 1442-1449.
Stadler, J., Mungai, G., and Brandl, R. 1998. Weed invasion in East Africa: insights from
herbarium records. Afr J Ecol 36: 15-22.
Steavenson, H. A. 1946. Multiflora Rose for Farm Hedges. Journal of Wildlife
Management 10: 227.
Stewart, C. N., Tranel, P. J., Horvath, D. P., Anderson, J. V., Rieseberg, L. H., Westwood,
J. H., Mallory-Smith, C. A., Zapiola, M. L., and Dlugosch, K. M. 2009. Evolution
of Weediness and Invasiveness: Charting the Course for Weed Genomics. Weed
Sci 57: 451-462.
Storkey, J., and Cussans, J. W. 2000. Relationship between temperature and the early
growth of Triticum aestivum and three weed species. Weed Sci 48: 467-473.
Strayer, D. L., Eviner, V. T., Jeschke, J. M., and Pace, M. L. 2006. Understanding the
long-term effects of species invasions. Trends in Ecology & Evolution 21: 645-
651.
Strecker, A. L., and Arnott, S. E. 2008. Invasive predator, Bythotrephes, has varied
effects on ecosystem function in freshwater lakes. Ecosystems 11: 490-503.
Strubbe, D., and Matthysen, E. 2009. Experimental evidence for nest-site competition
between invasive ring-necked parakeets (Psittacula krameri) and native
nuthatches (Sitta europaea). Biological Conservation 142: 1588-1594.
Suckling, D. M., and Brockerhoff, E. G. 2010. Invasion Biology, Ecology, and
Management of the Light Brown Apple Moth (Tortricidae). Annu Rev Entomol
55: 285-306.
Sun, J. H., Liu, Z. D., Britton, K. O., Cai, P., Orr, D., and Hough-Goldstein, J. 2006.
Survey of phytophagous insects and foliar pathogens in China for a biocontrol
perspective on kudzu, Pueraria montana var. lobata (Willd.) Maesen and S.
Almeida (Fabaceae). Biol Control 36: 22-31.
Swab, R. M., Zhang, L., and Mitsch, W. J. 2008. Effect of hydrologic restoration and
Lonicera maackii removal on herbaceous understory vegetation in a bottomland

89
hardwood forest. Restor Ecol 16: 453-463.
Swearingen, J. 2009. WeedUS Database of Plants Invading Natural Areas in the United
States: Climbing Euonymus (Euonymus fortunei). Available at
http://www.invasive.org/weedus/subject.html?sub=3024.
Swingle, W. T. 1916. The early European history and the botanical name of the tree-of-
heaven, Ailanthus altissima. Journal of the Washington Academy of Sciences 6:
409-498.
Szafoni, R. E. 1991. Vegetation management guideline: multiflora rose. Nat Area J 11:
215-216.
T'ulbure, M. G., Johnston, C. A., and Auger, D. L. 2007. Rapid invasion of a Great Lakes
coastal wetland by non-native Phragmites australis and Typha. J Great Lakes Res
33: 269-279.
Tlig-Zouari, S., Rabaoui, L., Irathni, I., and Ben Hassine, O. K. 2009. Distribution,
habitat and population densities of the invasive species Pinctada radiatia
(Molluca: Bivalvia) along the Northern and Eastern coasts of Tunisia. Cah Biol
Mar 50: 131-142.
Tuiten, W., Koenraadt, C. J. M., McComas, K., and Harrington, L. C. 2009. The Effect of
West Nile Virus Perceptions and Knowledge on Protective Behavior and
Mosquito Breeding in Residential Yards in Upstate New York. Ecohealth 6: 42-
51.
Turkington, R., Kenkel, N. C., and Franko, G. D. 1980. The Biology of Canadian
Weeds .42. Stellaria-Media (L) Vill. Can J Plant Sci 60: 981-992.
USDA, N. 2010. The PLANTS Database (http://plants.usda.gov, 16 April 2010).
National Plant Data Center, Baton Rouge, LA 70874-4490 USA.: NRCS.
Vail, G. D., and Oliver, L. R. 1993. Barnyardgrass (Echinochloa crus-galli) Interference
in Soybeans (Glycine max). Weed Technology 7: 220-225.
Van Strien, A. J., Van de Pavert, R., Moss, D., Yates, T. J., Van Swaay, C. A. M., and
Vos, P. 1997. The statistical power of two butterfly monitoring schemes to detect
trends. Journal of Applied Ecology 34: 817-828.
Vila, M., and Weiner, J. 2004. Are invasive plant species better competitors than native
plant species? evidence from pair-wise experiments. Oikos 105: 229-238.
Wang, C. Z., Zhou, B., and Palm, H. L. 2008. Detecting invasive sericea lespedeza
(Lespedeza cuneata) in Mid-Missouri pastureland using hyperspectral imagery.
Environ Manage 41: 853-862.
Ward, S. M., and Jasieniuk, M. 2009. Review: Sampling Weedy and Invasive Plant
Populations for Genetic Diversity Analysis. Weed Sci 57: 593-602.
Wardle, D. A., Nicholson, K. S., Ahmed, M., and Rahman, A. 1995. Influence of Pasture
Forage Species on Seedling Emergence, Growth and Development of Carduus
Nutans. Journal of Applied Ecology 32: 225-233.
Weber, E. 1998. The dynamics of plant invasions: a case study of three exotic goldenrod
species (Solidago L.) in Europe. Journal of Biogeography 25: 147-154.
Wedekind, I. 1991. Response of Canada Thistle (Cirsium-Arvense (L) Scop) to Various
Doses of Chlorsulfuron with and without Splitting in Field Trials. Z Pflanzenk
Pflanzen 98: 323-330.
Westbrooks, R. G., United States. Federal Interagency Committee for the Management of
Noxious and Exotic Weeds., and National Wildlife Refuge System (U.S.). 1998.

90
Invasive plants : changing the landscape of America : fact bookWashington, D.C.:
Federal Interagency Committee for the Management of Noxious and Exotic
Weeds. Pp. [vi], 109 p.
Weston, L. A., Barney, J. N., and DiTommaso, A. 2005. A review of the biology and
ecology of three invasive perennials in New York State: Japanese knotweed
(Polygonum cuspidatum), mugwort (Artemisia vulgaris) and pale swallow-wort
(Vincetoxicum rossicum). Plant and Soil 277: 53-69.
White, P. C. L., Ford, A. E. S., Clout, M. N., Engeman, R. M., Roy, S., and Saunders, G.
2008. Alien invasive vertebrates in ecosystems: pattern, process and the social
dimension. Wildlife Res 35: 171-179.
Wilfong, B. N., Gorchov, D. L., and Henry, M. C. 2009. Detecting an invasive shrub in
deciduous forest understories using remote sensing. Weed Sci 57: 512-520.
Williams, S. L., and Grosholz, E. D. 2008. The invasive species challenge in estuarine
and coastal environments: Marrying management and science. Estuar Coast 31: 3-
20.
Wilson, J. B., Chiarucci, A., Diaz, S., and White, P. S. 2008. Invasive species,
management for conservation and remote sensing. Appl Veg Sci 11: 1-2.
Wilson, R. G. 1981. Effect of Canada Thistle (Cirsium-Arvense) Residue on Growth of
Some Crops. Weed Sci 29: 159-164.
Wittenberg, R., and Cock, M. J. W. 2001. Invasive alien species : a toolkit of best
prevention and management practices. Wallingford, Oxon, UK ; New York:
CABI Pub. Pp. xii, 228 p.
Wonham, M. J., de-Camino-Beck, T., and Lewis, M. A. 2004. An epidemiological model
for West Nile virus: invasion analysis and control applications. P Roy Soc Lond B
Bio 271: 501-507.
Wu, S. H., Rejmanek, M., Grotkopp, E., and DiTomaso, J. M. 2005. Herbarium records,
actual distribution, and critical attributes of invasive plants: genus Crotalaria in
Taiwan. Taxon 54: 133-138.
Yates, E. D., Levia, D. F., and Williams, C. L. 2004. Recruitment of three non-native
invasive plants into a fragmented forest in southern Illinois. Forest Ecology and
Management 190: 119-130.
Yurkonis, K. A., Meiners, S. J., and Wachholder, B. E. 2005. Invasion impacts diversity
through altered community dynamics. J Ecol 93: 1053-1061.
Zangerl, A. R., and Berenbaum, M. R. 2005. Increase in toxicity of an invasive weed
after reassociation with its coevolved herbivore. Proceedings of the National
Academy of Sciences of the United States of America 102: 15529-15532.
Zidack, N. K., and Backman, P. A. 1996. Biological control of Kudzu (Pueraria lobata)
with the plant pathogen Pseudomonas syringae pv phaseolicola. Weed Sci 44:
645-649.
Zika, P. F., and Jacobson, A. L. 2003. An overlooked hybrid Japanese knotweed
(Polygonum cuspidatum x Sachalinense; Polygonaceae) in North America.
Rhodora 105: 143-152.
Zimdahl, R. L., and Foster, G. 1993. Canada Thistle (Cirsium-Arvense) Control with
Disking and Herbicides. Weed Technology 7: 146-149.
Zipper, C. E., Burger, J. A., McGrath, J. M., and Amichev, B. 2007. Carbon
accumulation potentials of post-SMCRA coal-mined lands. the 2007 National

91
Meeting of the American Society of Mining and Reclamation, 30 Years of
SMCRA and Beyond Gillette, WY.

92
VITA
Yu Liang
Birth Place: Shenyang, Liaoning Province, China P.R.
Birth Date: June, 2nd
1985
EDUCATION
B.S. in Forestry, Beijing Forestry University, 2008 GPA: 3.7
Thesis: Analysis on Spatial Structure of Secondary Betula platyphylla and
Populus tremuloides Forest in Wuling Mountain nature reserve in Beijing
Advisor: Dr. Xiaoxian Zheng (Professor in Forestry Management)
National Computer Rank Examination Certificate (2006.11)
HONORS
College of Ag Richards Scholarship 2009-2010
Phil E. Richards In-State Tuition Scholarship 2008-2009
Research Assistantship 2008-2010
Outstanding Undergraduate Student Fellowship 2005-2007
“Three Goods” Student Fellowship 2005-2007
WORK EXPERIENCE
(1)Research Assistant, Department of Forestry, University of Kentucky.
2008.10-present
Project: Analysis of Temporal and Spatial Hotspots and Dispersal Corridors of
Invasive Species.
Create and manage a digital database of invasive species in Kentucky with
Access;
Identify the relationship between land use type and invasive species habitat using
SAS;
Identify the hotspots and dispersal corridors of invasive species using ArcGIS
(2)Teaching assistant, FOR599 GIS in Natural Resource, Lexington, KY
2009.8.23-12.15
Project: FOR599 GIS in Natural Resource.
Instruct students to use GPS Unit (Garmin, Trimble) and GIS software such as
ArcGIS 9.3.
Grade homework and tests, instruct students to improve their map creation
skill.
Tutor students with the feature analysis final projects.
(3)Research Assistant, the Key Laboratory for Silviculture and Conservation
of Ministry of Education 2006.10-2008.5

93
Project: Research on Forest Sustainable Management in Beijing Mountain Area
Field investigation in Badaling Forest Farm Beijing
The Execution of stock map based on RS and GIS Technology with Arcview;
Project: Research of Forest Certification
Translation and collation of Australia Forest Materials
(4)Intern/team leader, Eco-environment Practice in Dongzelinghe Forest
Management Station in Xiaoxing’an Mountain of China.2007.7.2-7.29
Project: The Study of the Biodiversity of Two Kinds of Broad-Leaved Korean Pine
Mixed Forest and edited local Forest Enterprise Management Plan.
Field investigation, data analysis, forest growing model rectification
(5)Research Assistant, the Laboratory of Wild Plant Protection. 2007.3 -2007.6
Project: Study o f Embryo Abortion Mechanism of Ovule in Magnolia
artificial pollination on Magnolia and isolation with pockets
blossom and pollination recording
make Magnolia androecium paraffin section
(6)Team leader, Beijing Forestry Bureau, 2006.7.24-8.1
Project: The Study of the Indigenous Tree Species and a Perfect Tree/Shrub/Grass
Combination in Beijing.
Field investigation, data analysis
(7)Intern, Identification Practice in Forestry in Longmen Forest in the suburbs
of Beijing China.2006.7.10-7.22
Project: Study o f causation of Melanconium jugandinum in Longmen Forest
Field investigation, data analysis and compose thesis
(8)Co-author, Editorial Group of “Flora of Beijing Forestry University”.2005.4-
2005.10
Edit, compose and photograph in Flora of Beijing Forestry University
THESIS AND PUBLICATION
Y. Liang, J. Hast 2008 Spindletop Recreation and Trail Map. Department of Forestry,
University of Kentucky. 2008.12
Ning, Y., X. Zheng, Y. Liang, S. Liu. 2008. Study on the Structure of Aritificial Pinus
Tabulaeformis Stand. China Forestry Science and technology, 22:47-49
Wang, W., X. Zheng, Y. Liang, S. Liu. 2007. Spatial Structure of Scenic Forest of
Cotinus coggygria in Badaling Forest Farm of Beijing. Journal of Northeast Forestry
University 36:25-26,39
Wu, L., F. Yang, X. Zheng, D. Shi, Y. Liang, S. Liu, F. Yang. 2007. Forest Management
Plan in Australia. World Forestry Research 20:62-66

94
PRESENTATION:
2009. Y. Liang. Identification of temporal and spatial hotspots and dispersal corridors of
invasive species. Department of Forestry Workshop Series. University of Kentucky,
Lexington, KY.
2010. Y.Liang. Exotic Invasive Plants in Kentucky. 2010 Spring Research Symposium.
University of Kentucky, Lexington, KY.
LANGUAGES: Chinese (native), English (Fluent)
SKILL:
Experience in collaborating with agencies, organizations, on forestry and natural
resource related projects.
Proven knowledge of GIS with an emphasis on ArcGis9 and ERDAS IMAGINE 9.0.
Experience with creating databases to store and manipulate data using Microsoft Access.
Proficient in statistical analyst in SAS, R, and Microsoft Excel.
Proficient knowledge of programming using Visual Basic, and office software like
Microsoft Office