excit ation -con trac tion coupling in...
TRANSCRIPT

ExcitExcitationation--concontractracttiionon
ccoupling in oupling in
cardiomyocytescardiomyocytes
Dr. Tóth AndrásDr. Tóth András

2+2+ii
„Intracellular free calcium concentration”

TopicsMajor cellular structures involved in E-C coupling
Myofilaments: The end effector of E-C coupling
Sources and sinks of activator Ca
Cardiac action potentials and ion channels*
Ca influx via sarcolemmal Ca channels
Na/Ca exchange and the sarcolemmal Ca-pump
Sarcoplasmic reticulum Ca uptake, content & release
Excitation-contraction coupling
Control of cardiac contraction by SR & SL Ca fluxes
Cardiac inotropy
Ca „mismanagement”

Similarities between cardiac and skeletal muscle EC coupling
� Activated Ca2+ channels trigger the opening of SR
Ca2+ release channels
� APs provide the excitation stimulus used to activate
plasma membrane Ca2+ channels (or DHPRs)
� Resulting elevation in intracellular Ca2+ activates the
contractile machinery
� Both muscle types are striated & contain T-tubules
and highly developed intracellular SR networks
1!

Differences between cardiac and skeletal muscle EC coupling
� The heart contains specialized excitatory tissues (e.g.
SA node) and conductive fibers (Purkinje Fibers)
� The heart is a syncytium of many cells electrically
connected at intercalated discs by gap junctions
� The ventricular AP is 100x longer (250 ms) than that
of skeletal muscle
� Cardiac muscle contains a less developed T-tubule
and SR system
2!

Summary of cardiac EC coupling
� An AP is propagated from an adjacent myocyte via
gap junctions located at the intercalated disc
� AP activates membrane Ca2+ channels causing a
substantial Ca2+ influx during a prolonged AP
� Local increase in myoplasmic Ca2+ triggers a larger
release of Ca2+ from the SR (CICR)
� The global increase in myoplasmic Ca2+ activates the
myofilaments to initiate contraction
� β1-adrenergic stimulation increases contractility by
increasing Ca2+ current, release, and reuptake
3!

Major cellular structures
involved in EC coupling

“Our hero” the cardiac ventricular myocyte
~ 100 x 25 µµµµm
4
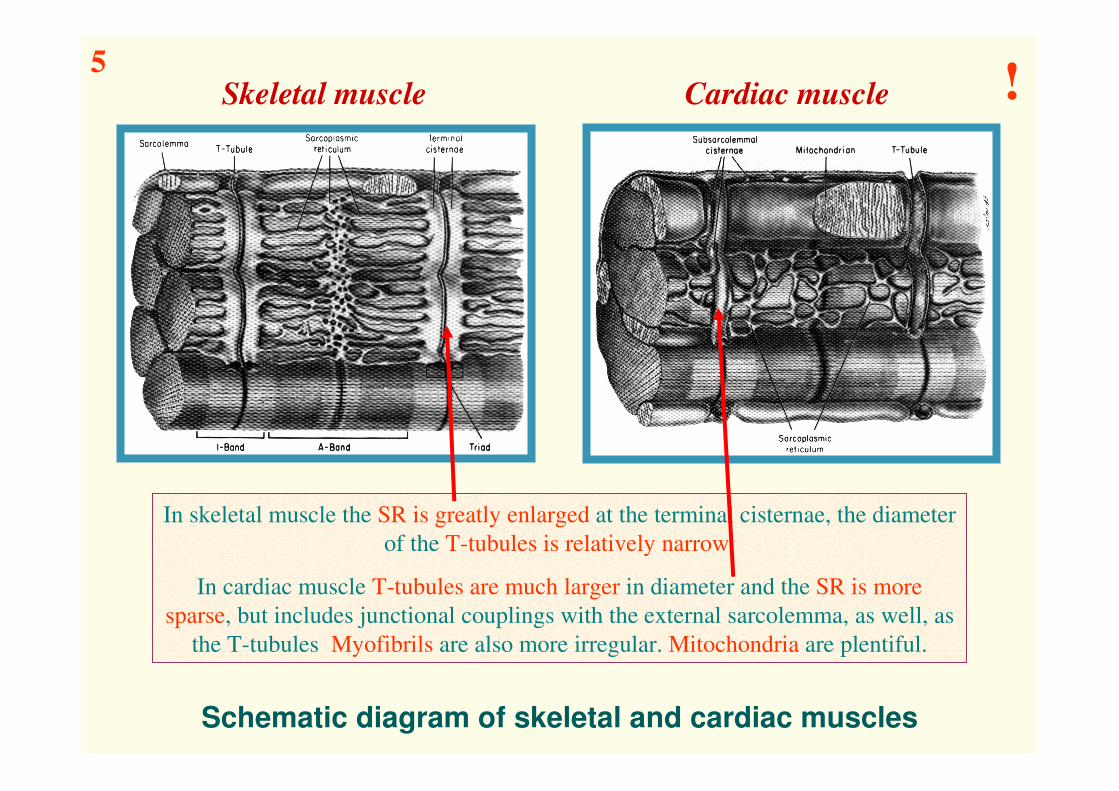
Schematic diagram of skeletal and cardiac muscles
In skeletal muscle the SR is greatly enlarged at the terminal cisternae, the diameter
of the T-tubules is relatively narrow.
In cardiac muscle T-tubules are much larger in diameter and the SR is more
sparse, but includes junctional couplings with the external sarcolemma, as well, as
the T-tubules Myofibrils are also more irregular. Mitochondria are plentiful.
Skeletal muscle Cardiac muscle5
!

The „restricted space” located between junctional SR and the sarcolemma forms a local
intracellular compartment which, has a very special role in both EC coupling and calcium
homeostasis.
In this space changes in Na+, K+ & Ca2+ concentrations are significantly greater than in
all other compartments of the cytosol. L-type Ca channel & NCX protein densities in the
junctional sarcolemma are also much higher than in any other regions of the sarcolemma.
The „restricted space”
6!
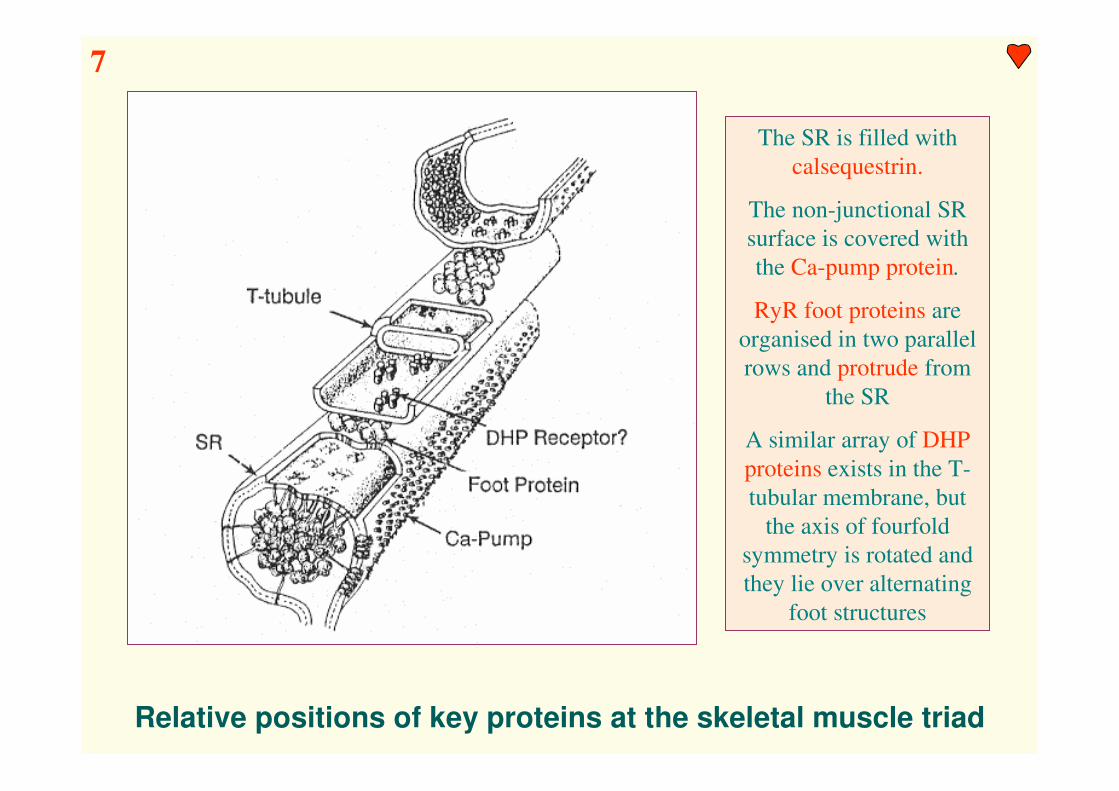
Relative positions of key proteins at the skeletal muscle triad
The SR is filled with
calsequestrin.
The non-junctional SR
surface is covered with
the Ca-pump protein.
RyR foot proteins are
organised in two parallel
rows and protrude from
the SR
A similar array of DHP
proteins exists in the T-
tubular membrane, but
the axis of fourfold
symmetry is rotated and
they lie over alternating
foot structures
7

Structural differences in skeletal & cardiac T-tubule junctions
In contrast to skeletal muscle, where DHPRs are found in very regular
structure, the DHPRs in the heart cells are sparse and less aligned.
8

Myofilaments: the end
effector of ECc

Myofilament proteins
Cardiac Troponin-C
One Ca-specific binding site
(regulation, Kd = 500 nM)
Two Ca-Mg specific binding
sites (stability)
9!

The “sliding filament” mechanism of contraction in cardiac cells
10!

Ca2+
*A + M � ADP � Pi
High actin affinity
A-M � ADP � Pi
ADP+Pi
ADP+Pi
A-M
Pi
ADP
*
A-M ATP
Low actin affinity
*
ATP
ATP
Resting muscle
Rigor Complex
The major steps of the crossbridge cycle in cardiac muscle
11!

The regulation of the contractile force in skeletal & cardiac muscle
The contractile force in skeletal muscle is determined by
A) Contraction summation (tetanus)
B) Activation of further fibers (recruitment)
C) Sarcomer length (myofilament overlap)
The contractile force in cardiac muscle is determined by
A) Intracellular Ca concentration (analog)
(intrinsic regulation)
B) Sarcomer length (myofilament overlap)
(extrinsic regulation)
12!

The length-tension relationship in skeletal & cardiac muscle
13!

Factors which alter cardiac myofilament Ca-sensitivity
Ca-sensitizer agents Hypoxia – ischemia
Positive inotropic agents
14

Force-velocity and force-power curves in cardiac muscle
15!

Sources, sinks and kinetics
of activator Ca
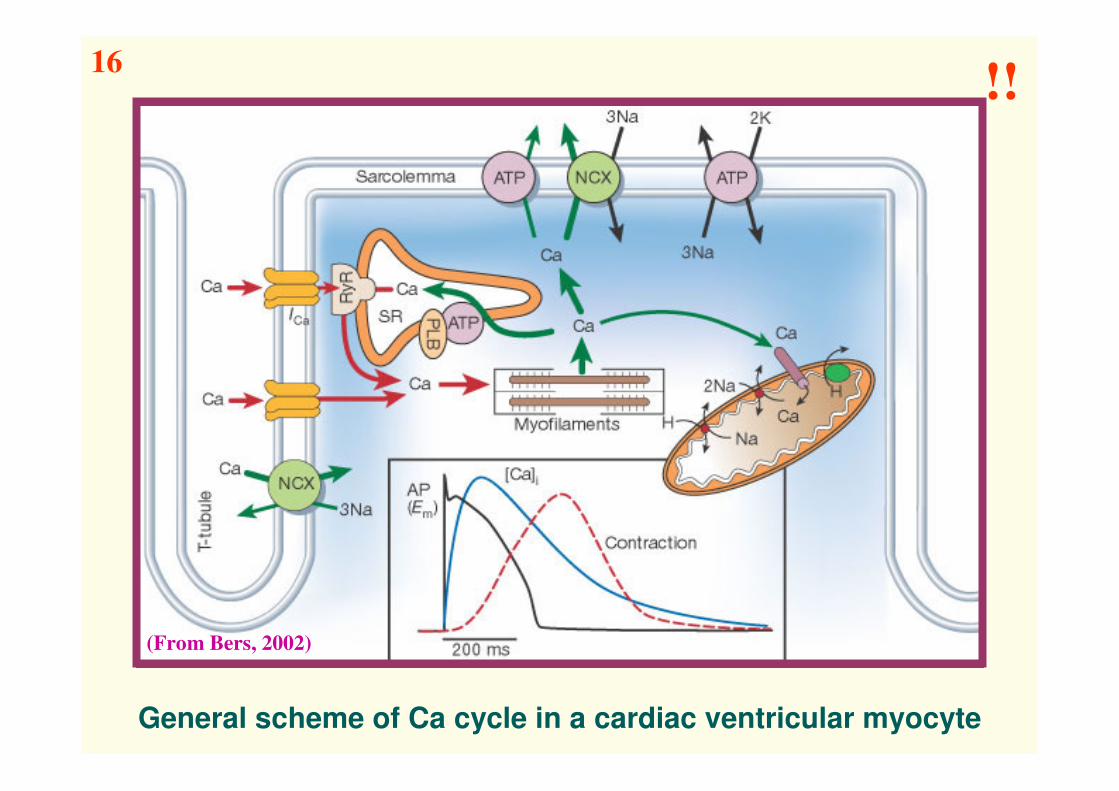
General scheme of Ca cycle in a cardiac ventricular myocyte
(From Bers, 2002)
16!!

Total Ca requirements for myofilament activation
A) Ca sensitivity of the myofilaments (F = Fmax/(1 + (Km/[Ca]i)n)
B) The amount of added total cytosolic (activator ) Ca, required to
activate contractile force
Data shown were measured in „skinned” cardiac fibers (Hill
coefficient n = 2) & intact cardiomyocytes (n = 4)
17

Dynamic Ca changes (Ca transient) during a twitch
Rabbit vetricular
myocyte model
A) Free [Ca]i and change in total cytosolic [Ca]cyt
B) Associated changes in Ca bound to different cytosolic ligands
C) Ca currents and transporter fluxes
!!!
Experimental
determination of the
“exact” [Ca]i value is
practically impossible !!!
18!

Ca content of the cytosol, Ca influx & efflux mechanisms
Extracellular space (ECS ∼ 30% total body volume)
[Ca]: 2 mmol/L ECS x 0.55 L ECS/L cytosol =1000 µmol/L cytosol
Influx: VD Ca channels, Na/Ca exchanger, „leakage” channels
Efflux: Na/Ca exchanger, sarcolemmal Ca-ATPase
Internal sarcolemmal surface
[Ca]: 60 µmol/L cytosol (no role in EC coupling!)
(following very quick removal of extracellular Ca, depolarization
does not produce measurable contraction or [Ca] increase)
Sarcoplasmic reticulum
[Ca]: 50-250 µmol/L cytosol
Influx: SR Ca-release channel (to cytosol!)
Efflux: Sr Ca ATPase (SERCA2) (to SR!)
Mitochondria
[Ca]: 10 000 µmol/L cytosol (in vitro) (PO43- – „matrix loading”)
[Ca]: 100 µmol/L cytosol (in vivo)
Influx: Na/Ca antiport (to cytosol!)
Efflux: Ca uniport (to mitochondrium!)
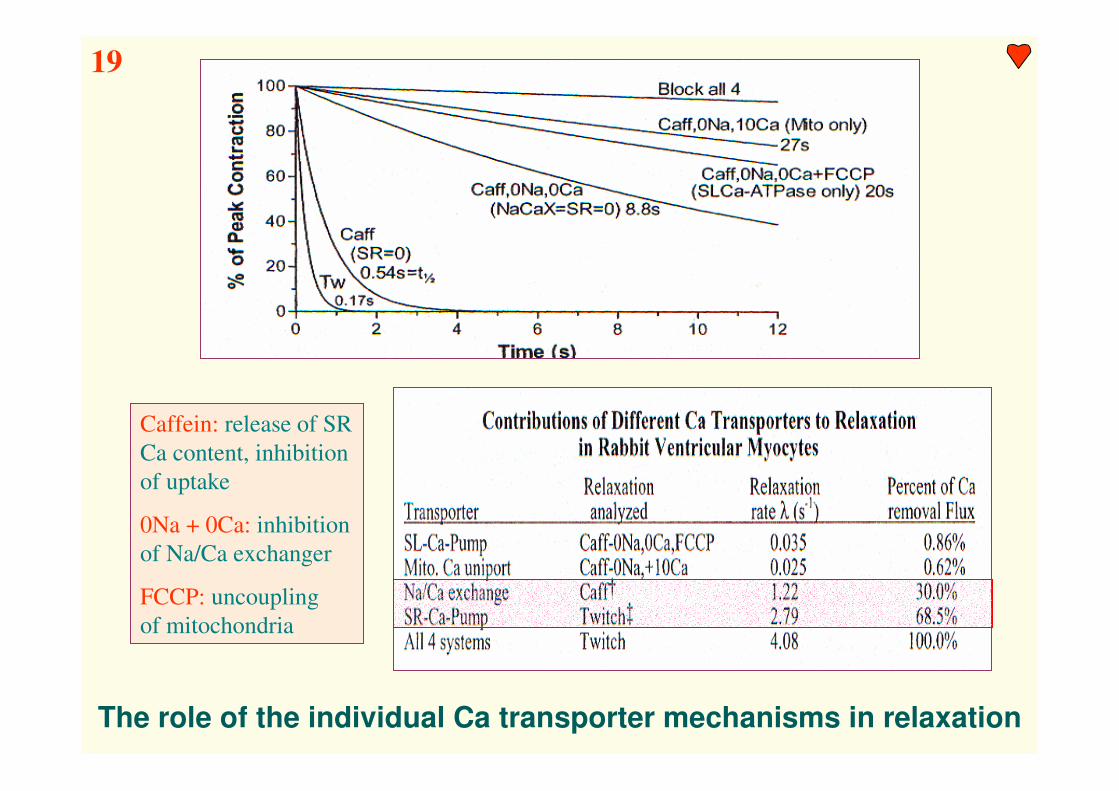
The role of the individual Ca transporter mechanisms in relaxation
Caffein: release of SR
Ca content, inhibition
of uptake
0Na + 0Ca: inhibition
of Na/Ca exchanger
FCCP: uncoupling
of mitochondria
19

The role of the mitochondrium in intracellular Ca regulation
The Ca cycle across the inner
mitochondrial membrane – changes in
[Ca2+]m are reflected in activities of the
mitochondrial dehydrogenases
Mitochondrial free Ca content
[Ca]m as a function of cytosolic Ca
concentration [Ca]i
Critical [Ca]i
≈ 500 nM
20

Cardiac action potentials
and ion channels*
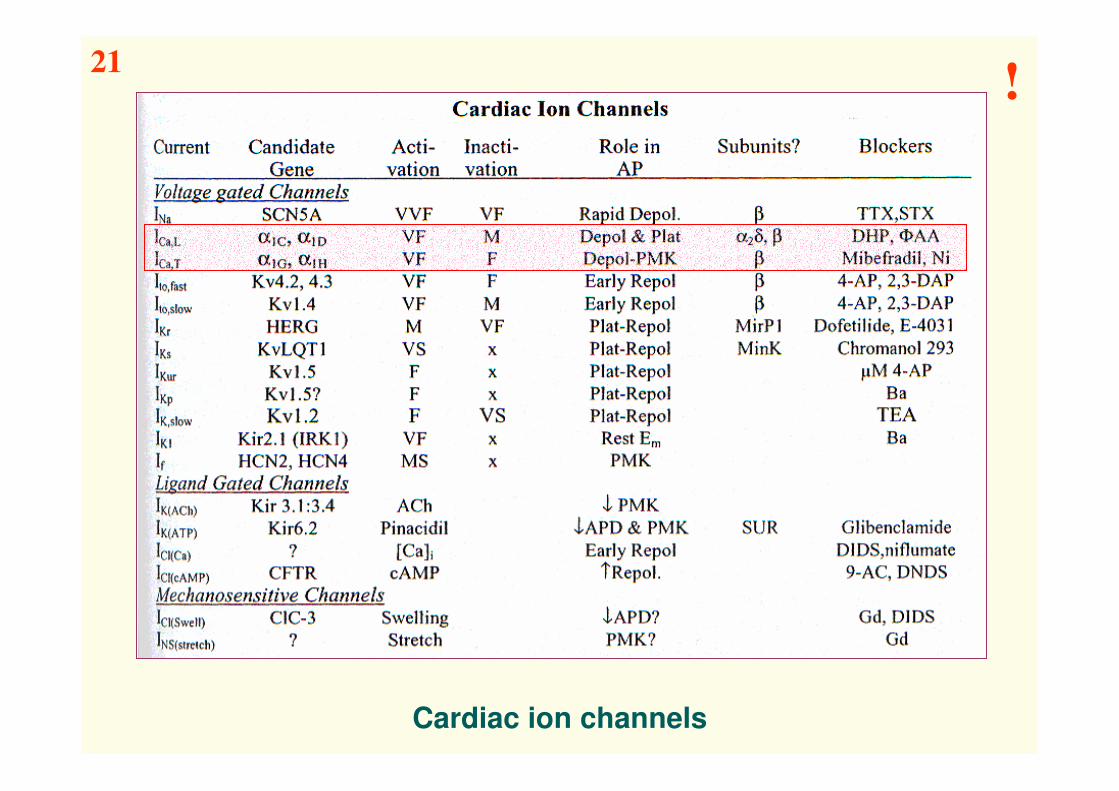
Cardiac ion channels
21!

Ca influx via sarcolemmal
Ca channels

The role of L-type Ca channels in ventricular myocytes
(From Bers, 2002)
22!

Properties of cardiac L- & T-type Ca channels
23!

Properties of the L-typeCa channel
CaL channel: ∼ 3-5/µm2
DHP receptor: ∼ 20/µm2
24
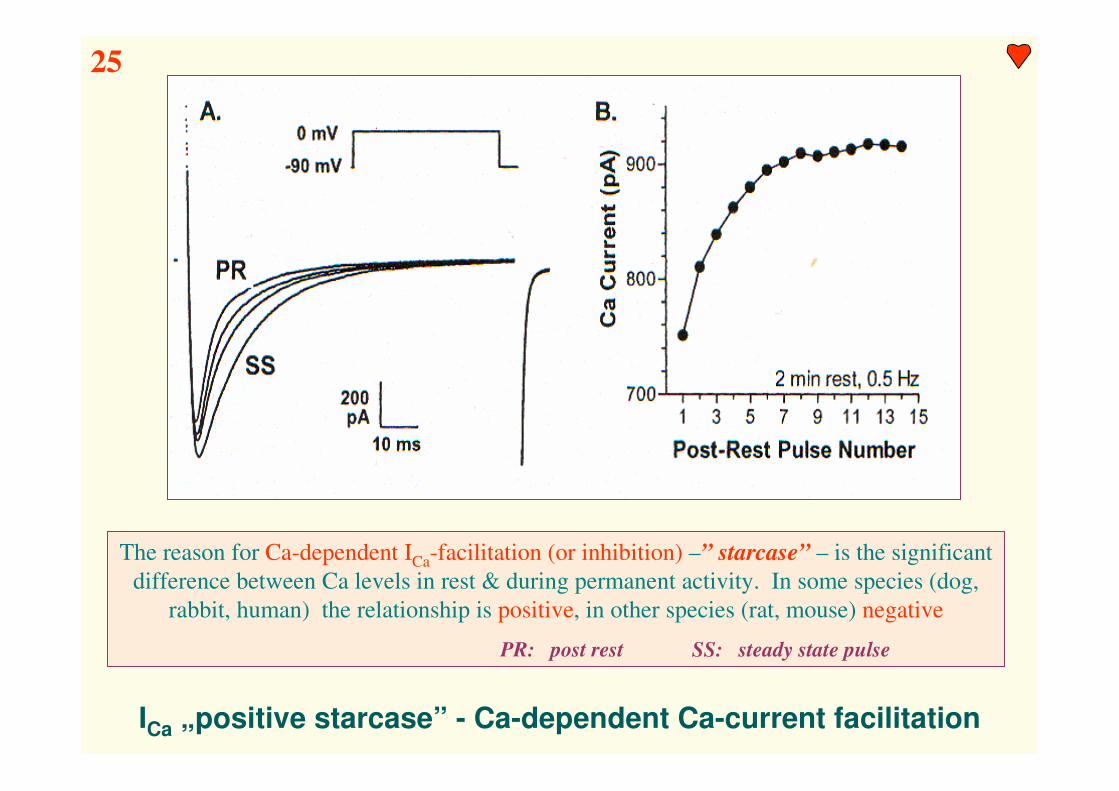
ICa „positive starcase” - Ca-dependent Ca-current facilitation
The reason for Ca-dependent ICa-facilitation (or inhibition) –” starcase” – is the significant
difference between Ca levels in rest & during permanent activity. In some species (dog,
rabbit, human) the relationship is positive, in other species (rat, mouse) negative
PR: post rest SS: steady state pulse
25

ICa during square pulse and AP-clamp
The amount of
transported Ca is
mainly determined
by the shape -
duration of the AP
Long AP: large
Ca influx
Short AP: less
Ca influx
26

Modulation of ICa by agonists and antagonists
Antagonists: Dihidropiridin (DHP)-family (nifedipine, nitrendipine,
nimodipine, nisoldipine, (+) Bay K 8644,
azidopine, iodipine)
Fenil-alkilamin (ΦAA)-family (Verapamil)
Benzothiazepin (BTX)-family (Diltiazem)
Agonists: (-) Bay K8644
(+) S-202-79, etc.
Agonists: mode 2 („permanently” open state)
(e.g. Bay K 8644 ∼ 0.6ms → ∼ 20 ms)
Antagonists: mode 0 („permanently” closed state)
27!

Dual pathways for activation of ICa by ββββ-adrenergic stimulation
1. Gs → Adenylyl-cyclase → cAMP↑ → PKA
2. Gs → direct effect (AKAP: PKA anchoring protein, PLB: phospholamban)
1.
2.
28

Conclusion
A) L-type Ca channel current (ICa) is the main route of Ca entry into
the cell (vs. leak, Na/Ca exchange, or ICa,T).
B) ICa plays a central role in cardiac EC-coupling and overall Ca
regulation and contraction.
C) The kinetics and amplitude of the ICa during the AP are critical factors
in controlling the amount of Ca released by the SR.
D) Ca which enters as ICa may also contribute directly to the activation
of the myofilaments, and to the replenishment of the SR Ca stores.
E) For a steady state to exist, the amount of Ca influx via ICa must be
extruded from the cell during the same cardiac cycle (e.g. via NCX).
Any uncompensated Ca influx could constitute a progressive Ca load
of the cell.
F) Due to the high conductance of these ion channels, a relative small
number of Ca channels which fail to inactivate (or reactivate), could
lead to substantial Ca gain (especially in depolarized cell). This can
compromize relaxation and contraction, and even be arrhytmogenic.
!

Na/Ca exchange & the
sarcolemmal Ca-pump

Sarcolemmal Ca-transport mechanisms in ventricular myocyte
(From Bers, 2002)
29!
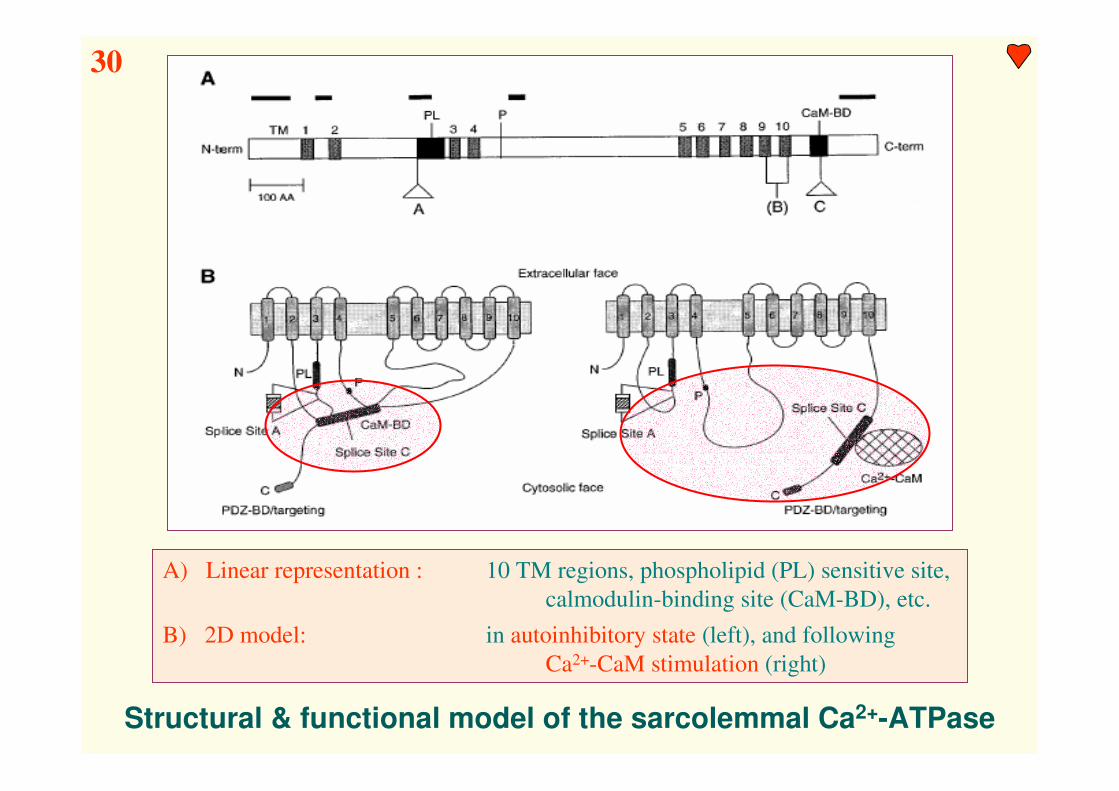
Structural & functional model of the sarcolemmal Ca2+-ATPase
A) Linear representation : 10 TM regions, phospholipid (PL) sensitive site,
calmodulin-binding site (CaM-BD), etc.
B) 2D model: in autoinhibitory state (left), and following
Ca2+-CaM stimulation (right)
30

Kinetic properties of the cardiac sarcolemmal Ca2+-ATPase
Calmodulin binding is an essential condition for physiological activity !!!
31!

A structural model for the Na/Ca exchanger (NCX)
regulation
szabályozásXIP: exchange inhibitory protein
32
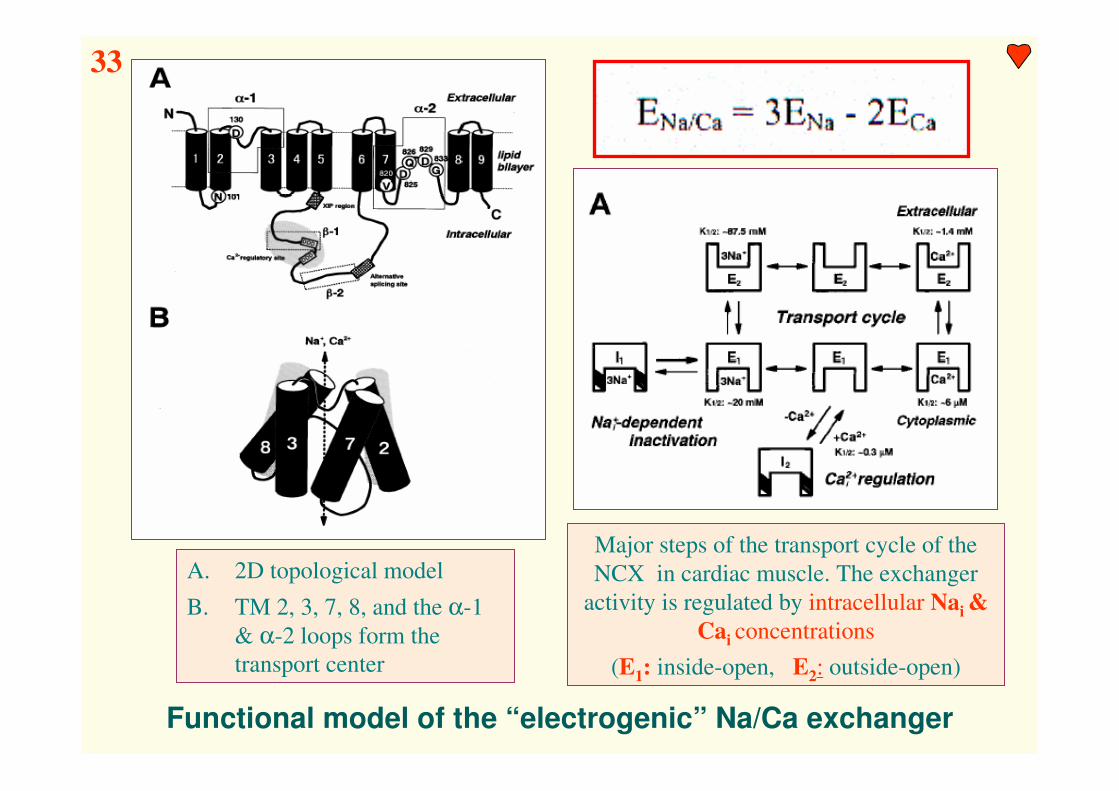
Functional model of the “electrogenic” Na/Ca exchanger
A. 2D topological model
B. TM 2, 3, 7, 8, and the α-1
& α-2 loops form the
transport center
Major steps of the transport cycle of the
NCX in cardiac muscle. The exchanger
activity is regulated by intracellular Nai &
Cai concentrations
(E1: inside-open, E2: outside-open)
33
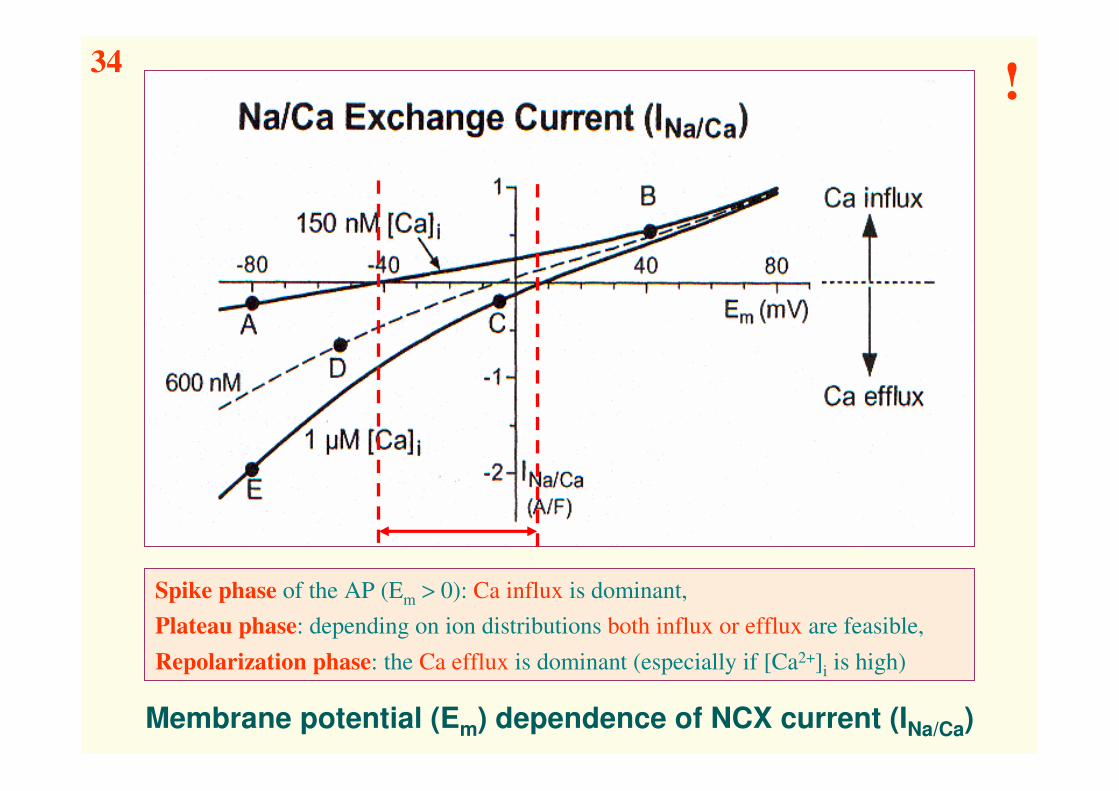
Membrane potential (Em) dependence of NCX current (INa/Ca)
Spike phase of the AP (Em > 0): Ca influx is dominant,
Plateau phase: depending on ion distributions both influx or efflux are feasible,
Repolarization phase: the Ca efflux is dominant (especially if [Ca2+]i is high)
34!
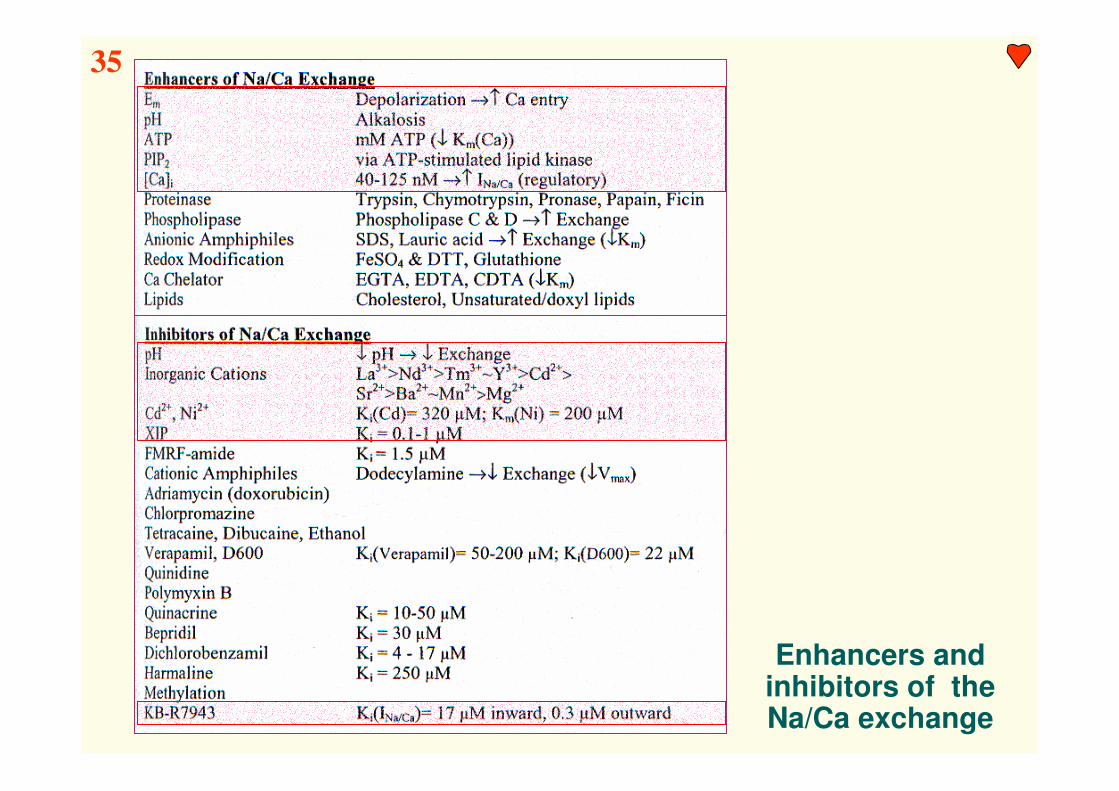
Enhancers and inhibitors of theNa/Ca exchange
35

Changes in ENa/Ca and INa/Ca during an action potential in rabbit ventricular myocyte
„Reverse mode”: Ca2+ influx, Na+ efflux – early phase of AP
„Forward mode”: Ca2+ efflux, Na+ influx – late phase of AP
36!

It is so simple to approximate the exchanger current !!!
37

Competition among Ca transport-mechanisms during relaxation
Heart failure:
INCX ↑↑↑↑↑↑↑↑↑↑↑↑
ISERCA ↓↓↓↓↓↓↓↓↓↓↓↓
38

“Rest decay” of SR Ca content
The rate of the rest decay
is mainly depending on
the ratio of the leakage
currents of the SR and SL
and the activity of the
Na/Ca exchanger
The rate of the rest decay
is quite species
dependent: it is small in
rat, but rather significant
in rabbit
Resting state is not a
physiological state for
the cardiac cell !!!
39

Conclusion
A) Na/Ca exchanger mechanism is essential in myocardial intracellular
Ca regulation
B) Na/Ca exchange is the main means by which Ca (entering the cell
via L-type Ca channels) is extruded from the cell, during both relax-
ation & diastole. By comparison the sarcolemmal Ca-pump (SLCP)
seems relatively unimportant in cardiac muscle.
C) Na/Ca exchange can even compete with the powerful SR Ca-pump
(SERCA) for cytoplasmic Ca (~ 1:2), thus contributing to relaxation
D) Na/Ca exchange can also mediate Ca influx sufficient to activate cell
contraction, but this probably does not occur under normal physio-
logical conditions (where its main role is Ca extrusion).
E) In order for a steady state to be achieved the average amount of Ca
extruded during each cardiac cycle should equal the amount of Ca
influx by L-type Ca channels.
F) Since Na/Ca exchange is the main means by which the cell extrudes
Ca, anything which prevents this Ca extrusion will increase cellular
Ca loading and can lead to Ca overload.
!

Sarcoplasmic reticulum - Ca
uptake, content and release

SR Ca-transport mechanisms in ventricular myocytes
(From Bers, 2002)
40!

SR Ca-pump (SERCA2) structure & steps of Ca transport
Structure:
10 transmembrane spans. 70% of
the protein is on the cytoplasmic
side of the SR membrane (β-strand,
phosphorilation & nucleotid
binding sites, stalk domains and a
hinge)..
A: Ca2+ uptake from the cytosol
B: Ca2+ release to SR lumen
Steps of Ca transport:
E1: 2 high affinity Ca2+ binding sites,
Ca & ATP binding, phosphorilation,
transition to E2 state
E2: lower affinity state, Ca2+ release
to SR, 2 H+ transported to cytosol,
transition to E1 state
A B
41

Phospholamban structure & its effect on SR Ca transport
PLB-SERCA2
interaction:
heterodimer PLB-
SERCA inhibits Ca
transport –
phosphorilation or
Ca binding reduce
inhibition
Ratio: 2(-3) PLB monomer/SERCA2 (non-saturated)
PLB
42

Pharmacological inhibitors of the SR Ca-ATPase (SERCA2)
Thapsigargin (TG) (Kd < 2 pM)
Cyclopiazonic acid (CPA)
2,5-di(tert-butyl)-1,4-benzohydroquinone (TBQ)
Major (patho)physiological regulatory factors of the SR Ca-ATPase
Ca: Normally [Ca]i is the limiting substrate for SERCA, thus the amount of
available Cai is the main factor which regulate pump activity
pH: Optimal pH for the SERCA is slightly alkaline (∼ 8). Decrease in pH
(especially pH < 7,4, e.g. acidosis associated with ischemia)
depress the rate of SR Ca-pumping and elongates relaxation
ATP: SERCA has a high affinity ATP site (Kd ∼ 1 µM) which is the substrate
site and a second, lower ATP affinity site (Kd ∼ 200 µM) which
serves a regulatory role. ATP normally is not a limiting factor.
Mg: The actual substrate of SERCA is probably Mg-ATP, thus a decrease in
intracellular Mg concentration may depress its activity
Inhibitors & regulators of the SR Ca-pump
43!

Major factors influencing SR Ca content
SOC: store operated channels
triadin, junctin:
SR structure proteins
44!

Properties of the SR Ca release channel (ryanodine receptor)
MW = 2 260 000 Da
According to some
hypotheses the output
of the RyR Ca channel
is located at the side of
the molecule, thus Ca
ions from the SR may
directly enter the
„restricted space”
45

Ca sparks in isolated ventricular myocytes
A) Two Ca sparks
(2D confocal
fluorescence)
B) Single Ca spark
(line-scan image)
C) [Ca]i computed
from the image
D) Surface plot of
[Ca]i during a Ca
spark
The elementary event of Ca-release from the SR is the local „spark”, which often
occurs during rest in a stochastic manner. 6-20 RyRs contribute to a single spark, which
starts at the T-tubule and increases [Ca2+]i in ∼ 10 ms to a peak value of 200-300 nmol.
The reason for its time dependent decrease is Ca diffusion and Ca reuptake.
Fusion of a large
number of sparks
leads to Ca-transient
& contraction !!!
46

Factors which alter
SR Ca release
The RyR macromolecular „signaling
komplex” serves as „final
integrator” site for a number of Ca
regulatory mechanisms !!!
47

Kiriazis 2000
Effects of genetical modulation of Ca-transport mechanisms
48

Conclusion
The SR can accumulate sufficient Ca and release it fast
enough to activate cardiac muscle contraction
Some typical values
In a typical ventricular myocyte there are ~ 2.5*105 DHPR, ~ 1.5-2.5*106
RyR & ~ 0.75-1.25*109 SR Ca-ATPase molecules
Typical Ca spark activity in rest is ~ 50/s for this level of spark activity the
activation of ~ 1000/s RyR is needed (only ~ 0.02% of RyRs)
For peak SR Ca release (~ 3 mM/s) ~ 40 000/s RyR is needed
(only ~ 4% of RyRs)
For total SR Ca release (~ 50 µmol/L citosol) ~ 7500 spark is needed
(only ~ 5% of RyRs)
For the measured Ca influx current via L-type Ca channels (max.1 nA)
~ 2-3% of the channels (DHPRs) are needed (only ~ 5000)
!

Excitation-contraction
coupling (ECc)
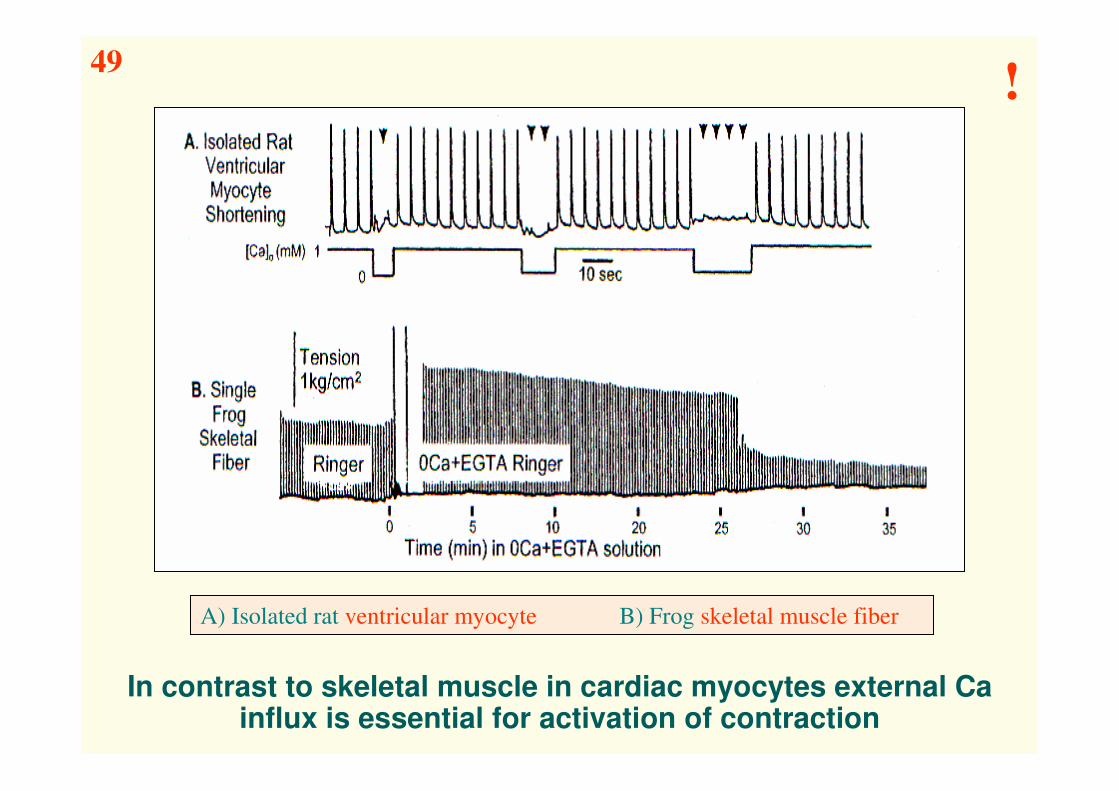
In contrast to skeletal muscle in cardiac myocytes external Ca influx is essential for activation of contraction
A) Isolated rat ventricular myocyte B) Frog skeletal muscle fiber
49!

Possible
activators of
cardiac SR
Ca release
50
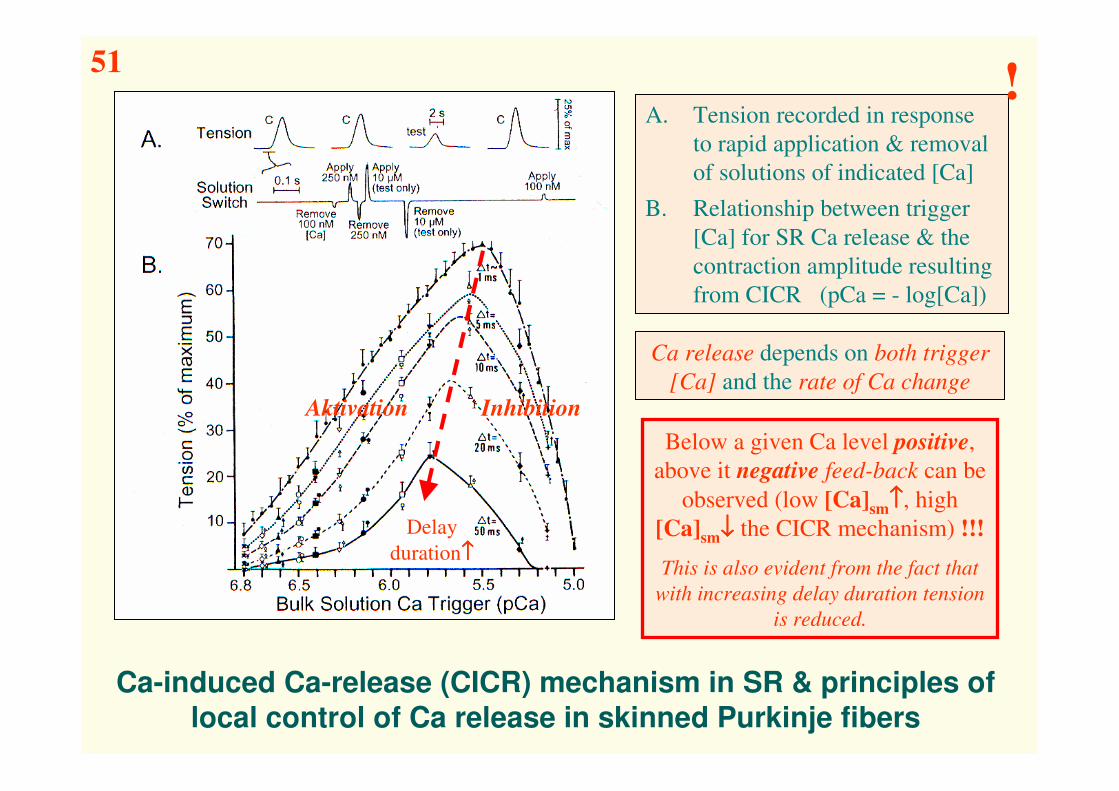
Ca-induced Ca-release (CICR) mechanism in SR & principles of
local control of Ca release in skinned Purkinje fibers
A. Tension recorded in response
to rapid application & removal
of solutions of indicated [Ca]
B. Relationship between trigger
[Ca] for SR Ca release & the
contraction amplitude resulting
from CICR (pCa = - log[Ca])
Below a given Ca level positive,
above it negative feed-back can be
observed (low [Ca]sm↑↑↑↑, high
[Ca]sm↓↓↓↓ the CICR mechanism) !!!
This is also evident from the fact that
with increasing delay duration tension
is reduced.
Aktivation Inhibition
Delay
duration↑↑↑↑
Ca release depends on both trigger
[Ca] and the rate of Ca change
51!

Diagram of Ca-induced Ca-release (CICR) in cardiac muscle
The two Ca-binding sites of the RyR bind Ca with different kinetics (1: fast, low
affinity binding site, 2: slow, high affinity binding site). Thus, following fast activation
of the RyR receptor its slow inactivation may also be induced by the Ca influx
52!

SR
+
TT
Ca2+
SRTT
Ca2+ Channel Release Channel
+ +Vm
Excitation-contraction coupling in cardiac muscle(Ca2+-Induced-Ca2+-Release)
53

„Local control” theory of EC-coupling in cardiac muscle
Observations: The rate of [Ca] change in the RyR environment either activate
or inactivate SR Ca release (i.e. the RyR).
Apparent junctional colocalization of DHPR & RyR
INa → ([Na]sm↑ → [Ca]sm↑) → SR Ca/release
Observation of localized SR Ca-release events (Ca sparks)
”Common Ca-pool” models could not explain the graded CICR
Hypothesis: RyR activity is modulated by the „fuzzy space” (ie. [Ca]sm)
„Ca-synapse” theory →→→→ 1 DHPR triggers only 1 RyR
„Cluster bomb” theory →→→→ 1 DHPR triggers a cluster of RyRs
Features: Either model could explain graded Ca release and high gain, but the
cluster bomb model does not require an extra large „single RyR” Ca-flux.
Within a cluster of RyRs (couplon) Ca release can be effectively all or
none, the release can be regenerative.
CICR gradation comes largely from recruitment of RyR clusters rather
than varying their Ca flux.
Validity: The local control theory was developed for Ica-induced SR Ca-
release and its validity is unproven for SR Ca-release induced
by different Ca triggers less localized to the junctional region
(NCX, „caged” Ca).
54!!

!
Comparison of EC-coupling in skeletal & cardiac muscle
In skeletal muscle
The physical link between DHPR & RyR is critical for VDCR
Influx of external Ca (ICa) is not required
In cardiac muscle
The physical link between DHPR & RyR is not critical for CICR.
Influx of external Ca (ICa) is crucial
55

Conclusion
A) In a simplified manner the 3 muscle types can serve as models for
the 3 major mechanisms of SR Ca-release (VDCR: skeletal muscle;
CICR: cardiac muscle; IP3ICR: smooth muscle) This is an oversimp-
liplification since all 3 mechanism may be present and functional in all 3
muscle types.
B) In skeletal muscle VDCR seems to be the crucial initiating process,
however, CICR may be very important in recruiting RyRs (∼ 50%)
which are not physically coupled to T-tubule tetrads. IP3 can also
induce Ca release (IP3ICR), but its significance is not yet clear.
C) In cardiac muscle CICR is the essential EC-coupling mechanism.
IP3 may modulate cardiac Ca release. There is also some evidence for a
functional direct link between the SL and the SR (and possibly VDCR). The
significance of this link is not yet clear.
D) In smooth muscle there is compelling evidence for both IP3ICR &
CICR. There is also evidence for that the IP3ICR interacts with a different
plasma membrane Ca channel (TRP), involved in CCE where the signal is
retrograde from IP3R to TRP.
!

Control of cardiac contraction
by SR & SL Ca fluxes
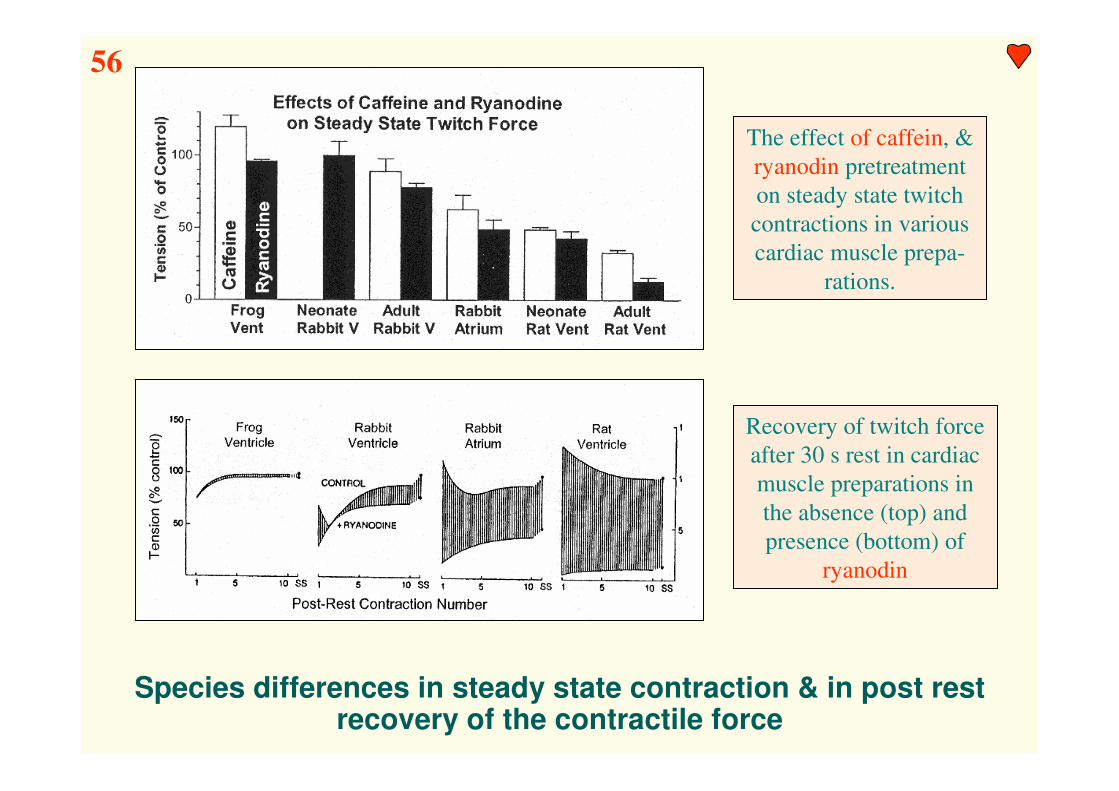
56
Species differences in steady state contraction & in post rest recovery of the contractile force
The effect of caffein, &
ryanodin pretreatment
on steady state twitch
contractions in various
cardiac muscle prepa-
rations.
Recovery of twitch force
after 30 s rest in cardiac
muscle preparations in
the absence (top) and
presence (bottom) of
ryanodin

A) [Ca]i-dependence of Ca transport in myocytes
B) Relative Ca-fluxes in ventricular preparations
C) Integrated Ca-fluxes during twitch relaxation
D) Fraction of activator Ca from ICa & SR Ca release
Analysis of cell Ca-fluxes in different species
57

Force-frequency relationship in cardiac muscle
Frequency-dependent changes in contractile force in cardiac myocyte
Force-frequency relationship in rat, rabbit, guinea-pig and human venricular myocyte
58

Conclusion
A) There is a great variation in details of [Ca]i regulation in different
cardiac muscle preparations and conditions. This apparent complexity
can be better understood by considering a small number of common systems
which interact and a few key functional properties that differ among cardiac
preparations.
B) Ca-influx can activate contractions in some hearts, but under normal
conditions in adult mammalian cardiac muscle the SR is the major
source of activator Ca. Ca influx can serve to trigger SR Ca release
and contribute to SR loading for the next contraction.
C) Ca released from the SR can be reaccumulated in the SR, or extruded
by the NCX. In steady state Ca-influx should always be balanced with
Ca-efflux during the carciac cycle.
D) During rest the Ca content of the SR can be gradually depleted by the
NCX & can also be quickly refilled during post rest activity (ICa) in
5-10 contraction). Depending on trans-sarcolemmal [Na]-gradient, rest
can either deplete or fill the SR Ca pool.
E) A dynamic yet delicate balance exists in the control of cardiac [Ca]i
and change in this system can lead to inotrópic & lusitropic effects.
!

Cardiac inotropy

Physiologic regulation of the inotropic state
Major regulatory mechanisms of cardiac muscle inotropy:
1. Sympathetic nerve system 2. Frank-Starling mechanism
3. Force-frequency relationship 4. Adrenergic regulation
5. Vascular function
59!

Hormone receptors and ion transporters in cardiac muscle 1.

Hormone receptors and ion transporters in cardiac muscle 2.

ββββ-adrenergic receptor signaling in ventricular myocytes
Top: activation, desensitization & down-regulation of the β1-receptor
Bottom: differences in G-protein coupling of the three β-receptor types
β-adrenergic mechanisms (through β2,3-receptors) may also mediate inhibitory
(cardioprotective) effects (decreasing contractility by the NO pathway) !!!
60

αααα-adrenergic transduction pathway in ventricular myocytes
αααα-adrenergic regulatory pathways: → G-protein → PLC (& PLD) → IP3+DAG →These products have divergent effects leading to positive inotropy & hypertrophy
61!

Comparison of αααα- and ββββ-adrenergic inotropy
α-adrenergic stimulation
increases Ca-transients to much
less extent than β-adrenergic
stimulation
β-adrenergic stimulation
decreases, α-adrenergic
stimulation increases
myofilament Ca-sensitivity
62!
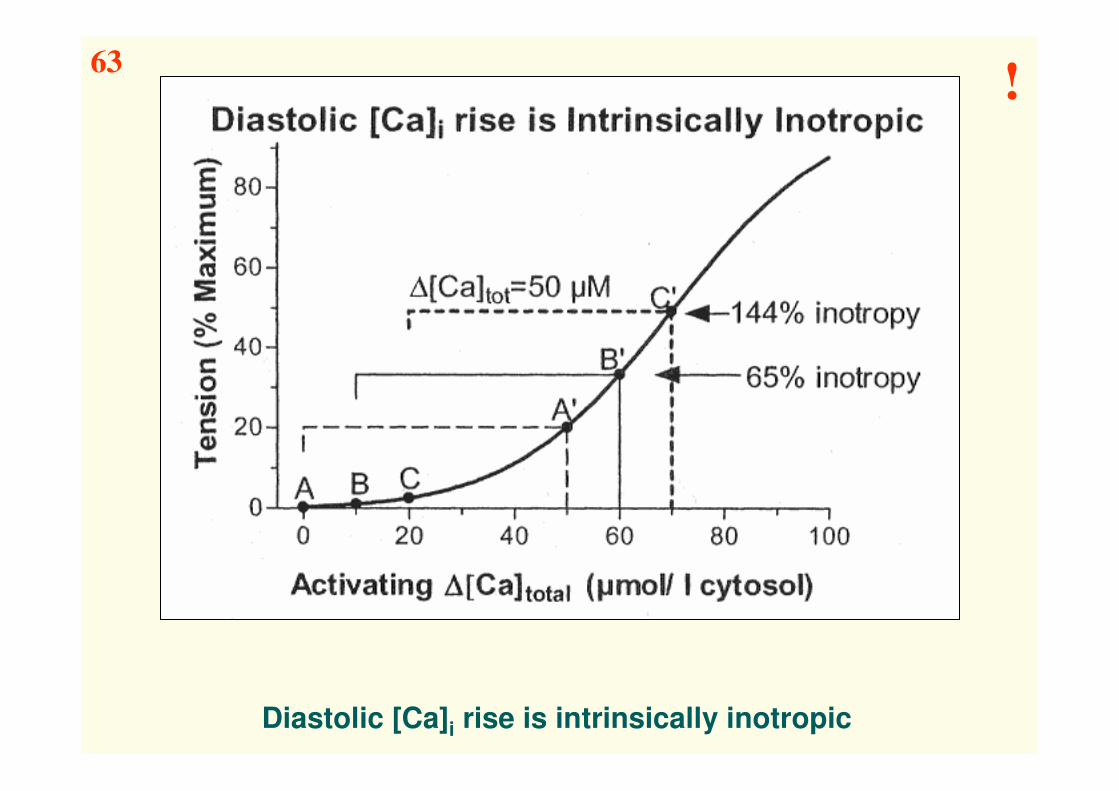
Diastolic [Ca]i rise is intrinsically inotropic
63!
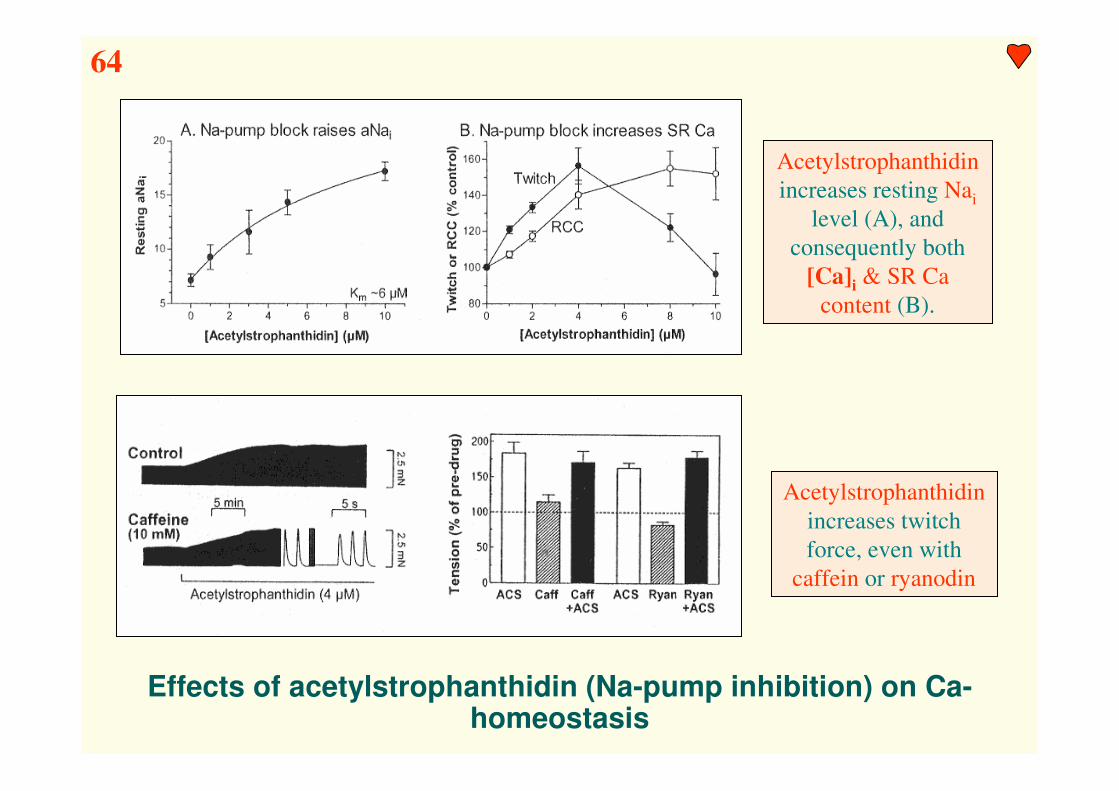
Effects of acetylstrophanthidin (Na-pump inhibition) on Ca-homeostasis
Acetylstrophanthidin
increases resting Nai
level (A), and
consequently both
[Ca]i & SR Ca
content (B).
Acetylstrophanthidin
increases twitch
force, even with
caffein or ryanodin
64
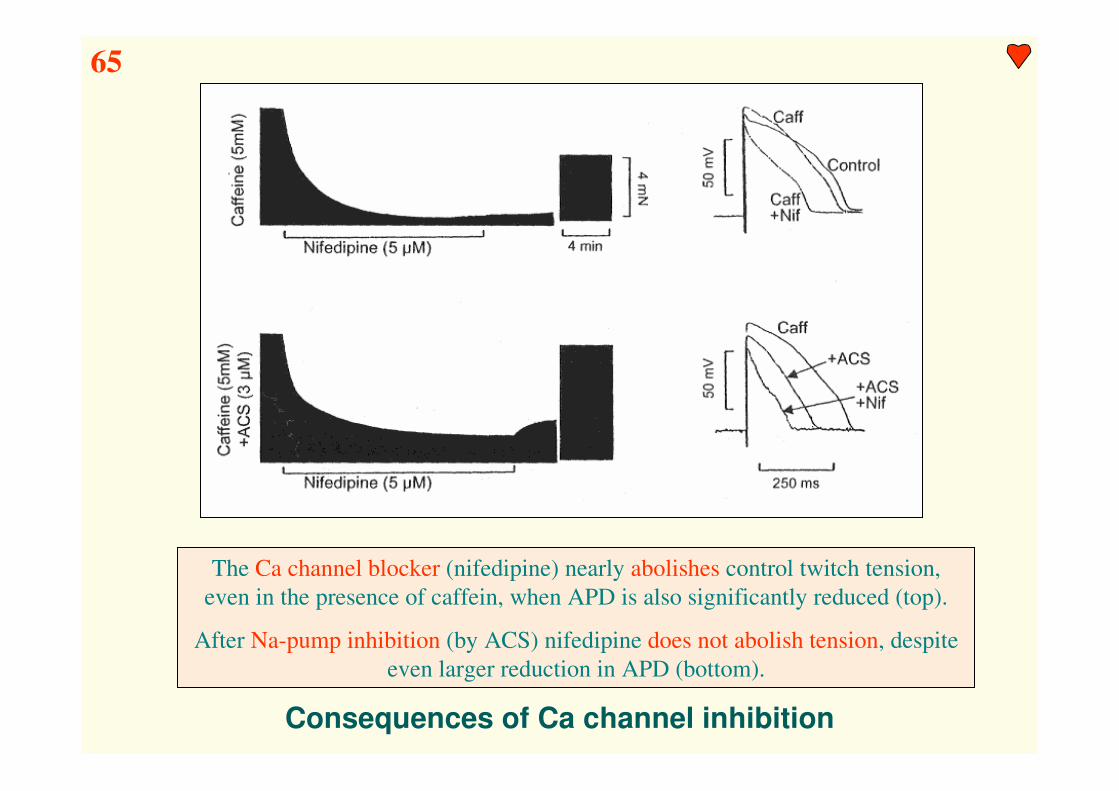
Consequences of Ca channel inhibition
The Ca channel blocker (nifedipine) nearly abolishes control twitch tension,
even in the presence of caffein, when APD is also significantly reduced (top).
After Na-pump inhibition (by ACS) nifedipine does not abolish tension, despite
even larger reduction in APD (bottom).
65

Conclusion
A) A number of mechanisms are available to increase cardiac inotropy
- hypotermia (experimental)
- β-adrenergic activation (physiological)
- α-adrenergic activation (physiological)
- CaMKII (Ca-CaM-dependent protein kinase) (physiological)
- cardioactive steroids (cardiac glycosids) (therapeutical)
B) From physiological viewpoint the role of β1-adrenergic activation (ANS) is
particularly important (inotropic, chronotropic, lusitropic, etc. effects), but
β2,3-receptors often mediate cardioprotective, inhibitory mechanisms (NO).
C) The significance of α1-adrenergic activation in increasing inotropy in the
human heart is moderate. However, it has an important role in induction of
cardiac hypertrophy (PKC). α1-AR activation enhances Ca sensitivity of the
myofilaments, but does not accelerate relaxation & typically prolong APD.
D) The role of CaMKII is less understood. As activated CaMKII also becomes
autophosphorylated, it may have the ability to integrate [Ca]i signals.
E) Digitalis is the oldest (1785) cardiac inotropic agent and the related cardio-
protective steroids are still among the most efficacious inotropic agents. By
inhibiting Na/K-ATPase, it shifts Na/Ca exchange, increases Ca influx &
decreases Ca efflux. Elevated [Ca]i intrinsically increases inotropy. Over-
dose, however, may cause negative inotropic & arrhythmogenic effects.
!

Ca “mismanagement” &
negative inotropy

Spontaneous Ca release & afterpotentials in cardiac myocytes
EADs may develop as a consequence of ICa reactivation, especially in cases when
APD is significantly elongated (e.g. LQT syndrome)
The main reason for DADs is the activation of Ca-dependent ion channels (e.g.
INA/Ca, ICl(Ca), INS(Ca)) by spontaneous spark activity generated by SR Ca overload
66!

The effect of acidosis on cardiac EC- coupling
C
D
In papillary muscle prepar-
ation pHo was shifted from
7.4 → 6.2. Contractile force
decreased, but Ca-transient
increased (A+B).
Decreased pH substantially
depresses maximal contrac-
tile force (C).
Acid transporters involved in
pHi regulation (D).
67

Some changes which occur during ischemia & reperfusion
68

Hyperthrophic signaling cascades
69

Major changes in EC-coupling during heart failure
Structures with altered
function in heart failure:
NCX (reverse & forward)
Voltage sensor
Phospholamban
SR-Ca-ATPase (SERCA2a)
Relative importance of NCX
increases significantly
Relative importance of
SERCA decreases
significantly.
70!

Positive feedback of maladaptive gene expression
Decreased pump function
is compensated by
(physiological) negative
feedback mechanisms.
Major consequences of
these compensatory
mechanisms are increased
[Ca]i & Ca transient.
[Ca]i increase induces
maladaptive gene-
expression, leading to
further depressed cardiac
pump function (circulus
viciosus).
!!!
!!!
- feedback
+ feedback
71!

Alteration of expression and function in human heart failure
72

Contractile dysfunction and arrhythmogenesis in heart failure
73

Altered calcium handling gene expression in end-stage human HF
74

Conclusion
A) The heart (ventricular myocyte) is a remarkably well tuned system, which can
rapidly vary its contractile output in response to a wide variety of physiologi-
cal stimuli by changing ion currents, Ca handling & myofilament properties.
There are some redundancies in this system, but any major perturbation in
normal [Ca] handling mechanisms still may lead to severe negative inotropy.
B) Ca overload often leads to spontaneous SR Ca-release & Ca-waves, which - if
randomly generated in a large number of cardiac cells - may also substantially
decrease contractile force.
C) Iti (transient inward current: INa/Ca+ ICl(Ca) + INS(Ca)) activated by SR Ca-release
is a major factor in eliciting delayed afterpotentials & aftercontractions.
D) Acidosis is a major consequence of myocardial ischemia & depression at low
pHi of several Ca-transport systems (NCX, SERCA) substantially contribute
to ischemic cardiodepression (decreased contractile force & Ca sensitivity).
E) In progression of the extremely komplex, multifactorial pathomechanisms of
hypoxia, ischemia, reperfusion & heart failure cellular Ca mismanagement
appears to be a common endpoint. Permanently increasing disturbances in Ca-
homeostasis may drastically depress cardiac contractile force & gradually
disables the heart in providing sufficient cardiac output to supply the metabolic
demands of the organism.
!