evaluation of the bacterial diversity in the human tongue...
TRANSCRIPT

Research ArticleEvaluation of the Bacterial Diversity inthe Human Tongue Coating Based on Genus-Specific Primersfor 16S rRNA Sequencing
Beili Sun1 Dongrui Zhou2 Jing Tu1 and Zuhong Lu12
1State Key Laboratory of Bioelectronics Southeast University Nanjing 210096 China2Key Laboratory of Child Development and Learning Science Ministry of Education of China Southeast UniversityNanjing 210096 China
Correspondence should be addressed to Zuhong Lu zhluseueducn
Received 25 February 2017 Revised 20 June 2017 Accepted 20 July 2017 Published 20 August 2017
Academic Editor Koichiro Wada
Copyright copy 2017 Beili Sun et alThis is an open access article distributed under the Creative Commons Attribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The characteristics of tongue coating are very important symbols for disease diagnosis in traditional Chinese medicine (TCM)theory As a habitat of oral microbiota bacteria on the tongue dorsum have been proved to be the cause of many oral diseases Thehigh-throughput next-generation sequencing (NGS) platforms have been widely applied in the analysis of bacterial 16S rRNA geneWe developed a methodology based on genus-specific multiprimer amplification and ligation-based sequencing for microbiotaanalysis In order to validate the efficiency of the approach we thoroughly analyzed six tongue coating samples from lung cancerpatients with different TCM types and more than 600 genera of bacteria were detected by this platform The results showed thatligation-based parallel sequencing combined with enzyme digestion and multiamplification could expand the effective length ofsequencing reads and could be applied in the microbiota analysis
1 Introduction
The complex microbial flora living on or within the humanbody has long been proposed to contribute to the humanhealth as well as disease [1ndash8] (Eckburg et al 2005) Usingculturing or unculturing methodology over 25000 bacterialphylotypes and over 700 prevalent taxa at the species levelhave been identified in the oralmicrobiome colonizing in theoral cavity including teeth gingival sulcus tongue cheekspalates and tonsils [9ndash20] (Keijser et al 2008 Nasidze et al2009)
As a reservoir for oral microorganisms food scrapssaliva and shed epithelial cells the human tongue has beeninvestigated showing significant association with the micro-bial communities of the gut and diseases such as gastritisor halitosis [11 21ndash24] In traditional Chinese medicine(TCM) according to the color and thickness the humantongue coating can be divided into a few types consisting
of thin-white coating thick-white coating sticky-white coat-ing thin-yellow coating and so forth [25] The color andshape of tongue coating may reflect the composition of thebacteria colonizing the tongue dorsum Several researcheshave considered the association between tongue coatingmicrobiome and traditional tongue diagnosis [26ndash29] Jianget al investigated 19 gastritis patients with a TCM ColdSyndrome or TCM Hot Syndrome tongue coating and 8healthy controls by Illumina paired-end double-barcode 16SrRNA V6 tag sequencing Han et al sequenced the V2ndashV4region of 16S rRNA gene by pyrosequencing to investigate thetongue coatingmicrobiome in patients with colorectal cancerand healthy controlsTheir results indicated that the richnessof the bacterial communities in the patients with thin tonguecoating and healthy controls was higher than in the patientswith thick tongue coating
In the past few years pyrosequencing of the 16S rRNAgene and sequencing-by-synthesis of metagenomics have
HindawiBioMed Research InternationalVolume 2017 Article ID 8184160 12 pageshttpsdoiorg10115520178184160
2 BioMed Research International
(a) (b)
Figure 1 Thin-white tongue coating type (a) and white-greasy tongue coating type (b) in TCM theory
been thewidely applied technologies in the study ofmicrobialcommunities [30ndash39] Pyrosequencing technology has thebenefits of relatively long length of sequencing read andthe drawbacks of high reagent cost and high error rates inhomopolymer repeats Metagenomic analysis requires deepsequencing data mining and large amounts of sequencingreads for gene assembly and annotation [24 27 40] Ligation-based sequencing technology provides inherent error cor-rection by two-base encoding which makes the platformmuchmore accurate [34]Due to the short sequencing lengththis methodology has a few restrictions in the microbiomediversity research
In the present study we develop an effective approachusing multigroup amplification and massively parallelligation-based sequencing technology to determine thebacterial diversity In addition we compare the tonguecoating microbial diversities of different TCM tonguecoating types using this method
2 Materials and Methods
21 Samples Tongue coating samples were collected fromtwo groups of lung cancer patients in Nanjing Chest HospitalEach group represented a specific TCM tongue coating typethe thin-white type and the white-greasy type (Figure 1) Atotal of 13 subjects were collected in the morning and allthe volunteers had no breakfast before sampling All subjectshad no oral inflammation and had refrained from brushingteeth and drinking colored beverage for 16 hours beforetesting The tongue coating samples were collected withsterilized cotton swabs mixed with 1ml of Ringerrsquos solutionand stored in minus20∘C immediately until extracting the totalmicrobial genome DNA All the 13 samples were investigatedby DGGE analysis (data not shown) Based on the DGGEresults the bacteria composition was significantly differentbetween different tongue coating types Six typical samplesfrom two tongue coating types were chosen for thoroughparallel sequencing and the clinical parameters of subjectswere shown in Table 1
22 Multiple Group PCR Primers Design The multigroupPCR primers were designed based on Human Oral Micro-biome Database (HOMD) (httpwwwhomdorg) The 16SrRNA gene sequences of a genus containing 4 species atleast in HOMD were picked out as a group and aligned byMEGA (v 402) Totally 421 species belonging to 27 generawere selected The conserved regions flanking the variableregion can be used for the multigroup PCR primers designA specific group primer consisted of 22 bp of sequencescontaining Eco57I recognition site and 20 bp of conservedregion sequences (Figure 2) Table 2 indicates all the 26 pairsof group primers generated
23 DNA Extraction Amplification and Ligation SequencingLibrary Preparation All the samples were centrifuged andresuspended in 1ml of 10x TE buffer (pH 8) The suspensionwas mixed with 100120583l of lysozyme (200mgml) and incu-bated at 37∘C for 1 h Lysis solution containing 50120583l of SDS(10 vv) and 20 120583l of Proteinase K (20mgml) was addedand the mixture was incubated at 55∘C for 3 h The totalbacterial genome DNA was obtained by phenol-chloroformextraction and ethanol precipitation
The PCR amplification was carried out separately by eachof group primers for each sample with the same conditionexcept for the annealing temperature All the PCR reactionswere performed in a final volume of 25120583l containing 1xPCR buffer (Takara) 25mM MgCl
2 200120583M each dNTP
25U of Taq polymerase 20 ng of template DNA and 1 120583Meach forward and reverse primer The PCR mixtures wereinitially denatured at 95∘C for 3min followed by 30 cycles45 s at 94∘C 45 s at (119879m minus 5)
∘C and 45 s at 72∘C ThePCR amplicons were digested using the enzyme Eco57I (Fer-mentas Burlington Canada) according to manufacturerrsquosinstructions The expected fragment DNA was acquiredby agarose gel electrophoresis and purified by QIAEX IIGel Extraction Kit (Qiagen Hilden Germany) PCR ampli-con mixture was prepared by pooling approximately equalamounts of recovered fragments from the same sample Afterblunting the 31015840 protruding termini of mixtures by T4 DNA
BioMed Research International 3
Table 1 Clinical parameters and barcode primer sequences of study subjects
Subjects ID Gender Age TCM tonguecoating type Barcode sequence (51015840 to 31015840)
A4 Male 30 Thin-white CTGCCCCGGGTTCCTCATTCTCTAAGCCCCTGCTGTACGGCCAAGGCGA5 Male 58 Thin-white CTGCCCCGGGTTCCTCATTCTCTCACACCCTGCTGTACGGCCAAGGCGA6 Female 72 Thin-white CTGCCCCGGGTTCCTCATTCTCTTCCCTTCTGCTGTACGGCCAAGGCGB2 Male 57 White-greasy CTGCCCCGGGTTCCTCATTCTCTCCGATTCTGCTGTACGGCCAAGGCGB3 Female 34 White-greasy CTGCCCCGGGTTCCTCATTCTCTTCGTTGCTGCTGTACGGCCAAGGCGB4 Male 62 White-greasy CTGCCCCGGGTTCCTCATTCTCTGGGCACCTGCTGTACGGCCAAGGCG
PRPF
Eco57Idigestion
CF CR
CS CS
gDNA
Sequencing
20 <J 20 <J
6 <J6 <J
4 <J 4 <J
16 <J
16 <J 18 <J
16 <J
35 <J 35 <J
Figure 2 Schematic diagram of the primer design and librarypreparation All the 16S rRNA gene sequences from the same generawere aligned to generate 20 bp of the conserved forward sequence(CF) and conserved reverse sequence (CR) flanking the variableregion The forward primer (PF) and reverse primer (PR) weredesigned by adding 6 bp of Eco57I recognition site and 1618 bp ofauxiliary sequence at the 51015840 end ofCF andCRThemicrobial genomeDNA of each subject was amplified by all the pairs of primers fromdifferent genus producing the amplicons containing the Eco57Icutting site (CS) which was 16 bp downstream of the recognitionsite Followed by Eco57I digestion products generated by all thegenus-specific primers conserving 4 bp of each primer sequencewere mixed together to produce the sequencing library The resultsof ligation-based sequencing were millions of reads which were thefirst 35 bases at the 51015840 end from either forward strand or reversestrand of library
polymerase four sequencing libraries were constructed forthe ligation-based sequencing system
24 Massively Parallel Ligation Sequencing In order to max-imize the sequencing capacity and simplify the workflowof sample preparation all the samples were operated in asingle sequencing run with barcodes added Six barcodes
were ligated to the 31015840 end of the library templates with T4DNA ligase as unique adaptors (Table 1)
SOLiD P1 adaptors (51015840-CCACTACGCCTCCGCTTT-CCTCTCTATGGGCAGTCGGTGAT-31015840) were ligated at the51015840 end of templates followed by standard SOLiD librarypreparation protocol
25 Construction of Reference Sequences Due to the char-acteristic of 2-base encoding in SOLiD sequencing thesequencing reads are in color-space format ldquo0 1 2 3rdquowhich stands for the permutation of the adjacent basesThe brief data processing pipeline in our study wasdesigned as Figure 2 shows because the novel microbialanalysis methods are inappropriate A total of 1049433unaligned 16S rDNA sequences of both bacteria and archaeawere downloaded from Ribosomal Database Project (RDP)(httprdpcmemsuedu) resource (release 10 update 13)Using in silico analysis of amplification by the designedprimers and endonuclease digestion 26 groups of fragmentswere selected Repetitive sequences were moved to construct26 groups of reference sequences (REF-DB) Taxonomiesof REF-DB (TAXA-DB) were assigned using the originalfull-length 16S rRNA gene sequences by the RDP onlineclassifier (httprdpcmemsueduclassifierclassifierjsp) at80 confidence cut-off If the reference was yielded frommore than one full-length 16S rRNA gene sequence eachof the original sequences was assigned separately From thegenus level the taxonomy shared by three-fourthsmajority ofthe full length sequences was defined as the taxonomy of thereferenceOtherwise taxonomyof higher level was compareduntil the domain level
26 Analysis of Sequencing Data Sequencing reads weresplit into six samples according to the 4-mer barcodes inP2 adaptor In color space all the samples were alignedseparately to REF-DB by Corona Lite Program (v42httpsolidsoftwaretoolscom) with up to 3 color-space mis-matches For unique reads (uniquely placed matches) afew steps of filtering were performed Firstly the matchingposition should be at the 51015840 end of the reference sequence(forward-strand matching) or reverse compliment (reverse-strand matching) Meanwhile the top four positions ofreads were checked with one mismatch in color space tofilter out nonspecific amplification or false endonuclease-digested products Secondly the abundance of reads match-ing ldquoforward-strandrdquo and ldquoreverse-strandrdquo references was
4 BioMed Research International
Table 2 The designed multigroup PCR primers
Primer ID Genus name Primer sequence (51015840 - 31015840) 119879m (∘C) Amplicon length (bp)ActF Actinomyces Pa-Eb-GCGAAGAACCTTACCAAGGC 562 142ActR Actinomyces Qc-E-TGACGACAACCATGCACCACPreF Prevotella P-E-GAACCTTACCCGGGCTTGAA 542 138PreR Prevotella Q-E-TGACGACAACCATGCAGCACStrF Streptococcus P-E-AACGATAGCTAATACCGCAT 461 139StrR Streptococcus Q-E-TAATACAACGCAGGTCCATCTreF Treponema P-E-CGCGAGGAACCTTACCTGGG 537 137TreR Treponema Q-E-ACGACAGCCATGCAGCACCTLepF Leptotrichia P-E-ACGCGAGGAACCTTACCAGA 535 139LepR Leptotrichia Q-E-CAGCCATGCACCACCTGTCTNeiF Neisseria P-E-CGGGTTGTAAAGGACTTTTG 492 135NeiR Neisseria Q-E-AGTTAGCCGGTGCTTATTCTSelF Selenomonas P-E-CTTCGGATCGTAAAGCTCTG 514 142SelR Selenomonas Q-E-TAGTTAGCCGTGGCTTCCTCCapF Capnocytophaga P-E-TACGCGAGGAACCTTACCAA 513 138CapR Capnocytophaga Q-E-ACAACCATGCAGCACCTTGAPorF Porphyromonas P-E-CAGCCAAGTCGCGTGAAGGA 540 172PorR Porphyromonas Q-E-CTGGCACGGAGTTAGCCGATFusF Fusobacterium P-E-ACGCGTAAAGAACTTGCCTC 514 75FusR Fusobacterium Q-E-ACGCGTAAAGAACTTGCCTCMycF Mycoplasma P-E-GATGGAGCGACACAGCGTGC 552 187MycR Mycoplasma Q-E-GCGGCTGCTGGCACATAGTTDiaF Dialister P-E-CGGAATTATTGGGCGTAAAG 531 161DiaR Dialister Q-E-CTTTCCTCTCCGATACTCCAEubF Eubacterium P-E-GATACCCTGGTAGTCCACGC 516 146EubR Eubacterium Q-E-CTCCCCAGGTGGAATACTTAFirF Firmicutes P-E-GATACCCTGGTAGTCCACGC 516 146FirR Firmicutes Q-E-CTCCCCAGGTGGAATACTTAPepsF Peptostreptococcus P-E-TAATTCGAAGCAACGCGAAG 527 154PepsR Peptostreptococcus Q-E-CGACAACCATGCACCACCTGKinF Kingella P-E-GGAATTACTGGGCGTAAAGC 524 168KinR Kingella Q-E-AATTCTACCCCCCTCTGACAPepnF Peptoniphilus P-E-ATCACTGGGCGTAAAGGGTT 511 186PepnR Peptoniphilus Q-E-CGCATTTCACCGCTACACTALachF Lachnospiraceae P-E-CAACGCGAAGAACCTTACCA 542 146LachR Lachnospiraceae Q-E-GACGACAACCATGCACCACCPasF Aggregatibacterb P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Aggregatibacter Q-E-TTAGCCGGTGCTTCTTCTGTPasF Haemophilus P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Haemophilus Q-E-TTAGCCGGTGCTTCTTCTGTLactF Lactobacillus P-E-CCGCAACGAGCGCAACCCTT 553 103LactR Lactobacillus CCTCCGGTTTGTCACCGGCATM7F TM7 P-E-GGGCGTAAAGAGTTGCGTAG 492 185TM7R TM7 Q-E-TACGGATTTCACTCCTACACGemF Gemella AAAGCTCTGTTGTTAGGGAA 468 98GemR Gemella P-E-GGTGGCTTTCTGGTTAGGTAPseF Pseudomonas GGCGGCAGGCCTAACACATG 565 99PseR Pseudomonas P-E-TTACTCACCCGTCCGCCGCTVeiF Veillonella P-E-GTAAAGCTCTGTTAATCGGG 481 100VeiR Veillonella GTGGCTTTCTATTCCGGTACMogF Mogibacterium P-E-CACGTGCTACAATGGTCGGT 507 101MogR Mogibacterium ATCCGAACTGGGATCGGTTTCatF Catonella P-E-CATGCAAGTCGAACGGAGAT 508 101CatR Catonella GTTACTCACCCGTCCGCCACaCCAAGGCGGCCGTACG bCTGAAG cCCGACGTCGACTATCCAT
BioMed Research International 5
Table 3 Statistical results of SOLiD sequencing tags mapping with RDP references
Subject ID Raw reads Matched reads Uniquely matched readsCounts Frequency Counts Frequency
A4 12455117 2043364 1641 119454 096A5 5043037 861788 1709 63751 126A6 27658749 4965785 1795 418203 151B2 10171189 1406962 1383 160263 158B3 8938854 1968093 2202 96206 108B4 10848548 1915504 1766 119292 11
Table 4 OTUs assignment of unique tags based on RDP classifier
OTU A4 A5 A6 B2 B3 B4Number PCT () Number PCT () Number PCT () Number PCT () Number PCT () Number PCT ()
Genus 548 9320 406 9490 495 9593 464 9490 388 9490 637 9522Family 15 255 11 257 10 194 12 245 10 245 17 254Order 9 153 3 070 4 078 4 073 3 073 6 090Class 9 153 3 070 6 116 7 122 5 122 8 120Phylum 1 170 0 0 0 0 1 024 1 024 0 0Domain 6 102 5 117 1 019 3 049 2 049 1 015Total OTUs 588 428 516 491 409 669
counted separately and the reads with less than 5 countsin both strands were discarded Finally Operational Taxo-nomic Units (OTUs) were identified according to matchedsequences in the REF-DB and corresponding taxonomyinformation in TAXA-DB For the nonunique reads thatmatched the reference in more than one location all ofthe matched references were assigned at the phylum levelaccording to TAXA-DB A matched read was assigned toa specific taxonomy while all the references it matchedbelonged to accordant phylum Otherwise the read wasdenoted as ldquoundefinedrdquo Both the unique and nonuniquematched reads were summarized by a set of Perl scripts
27 Phylogenetic Analysis The original full-length sequencesof matched OTUs were aligned using ClustalW and a relaxedneighbor-joining tree was built by PHILIP 368 Weightedand unweighted UniFrac were run using the resulting treeand environment annotation of number of hits PCA wasperformed on the resulting matric of distances between eachpair of samples
28 Prediction of Metagenome from 16S rRNA Gene DataThe PICRUSt project aims to support prediction of theunobserved character states in a community of organismsfrom phylogenetic information about the organisms in thatcommunity This program was used to predict metagenomeabundance from 16S rRNA gene data
3 Results
31 Sequencing Performance of SOLiD Reads One-quarter ofslide was used to perform the ligation-based sequencing and
84399432 reads of 35-base length (275 gigabases in total)were captured after removing sequences of insufficient qual-ity Splitted by 4-mer barcodes a total number of 75115494reads (89) were generated (Table 3) The difference ofsequencing throughput among six samples was probably dueto the sample quality or the procedure of library preparation
32 Taxa Assignment of the Sequencing Reads Since SOLiDsystem employs 2-base encoding and color-space strategya single color change is a measurement error two adjacentcolor changes may result from a single nucleotide variationin base space and three color-space mismatches might implytwo adjacent variants compared to reference We used up tothree color changes in 35-base length as the alignment param-eter which implied at most two nucleotide mismatchesConsequently the sequence similarity was more than 94which corresponded to the genus level classification Alignedto REF-DB matched and uniquely matched reads weresummarized in Table 3
To increase the accuracy of taxonomy assignment twosteps of validation were performed before OTUs definitionas Methods described Altogether we discarded 3375 ofthe uniquelymatched reads leaving 647375 sequencing readsfor taxonomy analysis Compared to TAXA-DB number ofOTUswas identified ranging from409 to 669 for six samplesrespectively Most of the OTUs (more than 93) could beassigned at the genus or lower level while a very small amountof OTUs was just assigned at the phylum or higher level(Table 4)
33 Microbiome Diversity of the Tongue Coating SamplesThe dominant phyla (relative abundance gt 1) across allsamples were Firmicutes (5519 plusmn 1600) Proteobacteria
6 BioMed Research International
0102030405060708090
100
Sequ
ence
s cla
ssifi
ed to
phy
lum
leve
l (
)
A5 A6 B2 B3 B4A4
DeferribacteresCyanobacteriaAcidobacteriaChlamydiaeSpirochaetes
ChlorobiActinobacteriaBacteroidetesProteobacteriaFirmicutes
Figure 3 Relative abundances of taxonomy classification of thetongue coating microbiome at the phylum level in the six tonguecoating samples
(2611 plusmn 1485) Bacteroidetes (1480 plusmn 236) Acti-nobacteria (203 plusmn 145) and Chlorobi (117 plusmn 079)Ultimately we identified 209 genera from the six samplesand the abundance of each genus was defined in MethodsAs shown in Figure 3 92 genera were assigned to phylumFirmicutes with the maximum abundance as well PhylaProteobacteria and Bacteroidetes contained some dominantgenera in our results while only 68 and 30 genera wereassigned to these two phyla separately The fifteen mostabundant genera of each sample were illustrated in Figure 4with total abundance ranging from 702 to 82 Therewere 163 genera assigned from 79762 reads for sample A4and the most abundant genera were Haemophilus (1338Proteobacteria) Haliscomenobacter (1023 Bacteroidetes)Enterococcus (950 Firmicutes) Streptococcus (556 Fir-micutes) and Acetanaerobacterium (417 Firmicutes) Forsample A5 134 genera were identified from 41580 readshaving the most abundant genera of Haemophilus (1140)Haliscomenobacter (781) Bacillus a (640 Firmicutes)Streptococcus (550) and Enterococcus (462) There were141 genera assigned from 242085 reads for sample A6and the most abundant genera were Ralstonia (1731 Pro-teobacteria) Anaerostipes (1032 Firmicutes) Bacillus a(953 Firmicutes) Haemophilus (496) and Coprococcus(369 Firmicutes) For white-greasy tongue coating typesamples for sample B2 119 genera were found within 155093reads and the dominant genera were Roseburia (2691Firmicutes) Anaerotruncus (98 Firmicutes) Coprococ-cus (791) Sphingobacterium (581 Bacteroidetes) andDorea (348 Firmicutes) For sample B3 128 genera wereassigned from 47717 reads with the largest proportiongenera of Streptococcus (3425) Sphingobacterium (714)Prevotella (708 Bacteroidetes) Enterococcus (679) andAcetanaerobacterium (450) For sample B4 143 generawerefound within 81138 reads and the dominant genera were
Haemophilus (1903) Sphingobacterium (1065) Entero-coccus (794) Acetanaerobacterium (531) and Bacillus c(432 Firmicutes)Overall the dominantmicrobial phyla ofthe three thin-white tongue coating samples were FirmicutesProteobacteria and Bacteroidetes while the white-greasysamples B2 and B3 had the second abundant phylum ofBacteroidetes and the sample B4 had the most abundantphylum of Proteobacteria
In order tomaximize the high-throughput usable data fordeep analysis both the uniquely and nonuniquely matchedreads were classified at phylum level The percent of assignedreads was ranging from 588 to 656 Compared withthe results based on unique reads the proportion of phylaFirmicutes and Actinobacteria had dramatically increased(data not shown)
34 Shared Genera of the Same Tongue Coating Type SamplesWe compared the shared genera within the same tonguecoating type samples There were 117 genera shared betweenthe thin-white tongue coating type samples Of these genera58were fromFirmicutes 37 generawere fromProteobacteriaand 12 genera belonged to Bacteroidetes For the white-greasytongue coating type samples 96 genera were observed to beshared 48 genera belonged to Firmicutes 28 genera belongedto Proteobacteria and 11 genera belonged to Bacteroidetesrespectively
35 Comparison of Bacterial Diversity between DifferentTongue Coating Type Samples There were 77 genera sharedacross the six samples (Figure 5) 35 genera belonged toFirmicutes 23 genera belonged to Proteobacteria and 8genera belonged to Bacteroidetes We compared the generaobserved in one tongue coating type but not the otherFigure 6 showed the genera that appeared in all the thin-whitetongue coating type but existed in none white-greasy tonguecoating type sample A total of 12 genera were observed onlyin the thin-white tongue coating type samples PelomonasHaemophilus Thioalkalispira and Zoogloea (Proteobacteria)Jeotgalibacillus Granulicatella Lachnobacterium Peptostrep-tococcus Anaeromusa Parasporobacterium and Sporobac-terium (Firmicutes) and Prevotella (Bacteroidetes) Figure 7showed the genera that appeared in all the white-greasytongue coating type but did not exist in all the thin-white tongue coating type sample The white-greasy tonguecoating type samples were specifically unique for 6 gen-era Anaerostipes Lactobacillus Anaerobacter Ruminococ-caceae Incertae sedis and Oribacterium (Firmicutes) andAcidithiobacillus (Proteobacteria) Analyzed by Studentrsquos 119905-test 19 genera performed significantly different between twotongue coating types (Table 5)
Analyzed by UniFrac software the thin-white tonguecoating type and white-greasy tongue coating type wereobservably different at the genus level as shown in principalcomponent analysis (PCA) plot (Figure 8) The three thin-white tongue coating subjects had relatively similar microbialdiversity (Bonferroni-corrected 119875 value is 025) while three
BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
(a) (b)
Figure 1 Thin-white tongue coating type (a) and white-greasy tongue coating type (b) in TCM theory
been thewidely applied technologies in the study ofmicrobialcommunities [30ndash39] Pyrosequencing technology has thebenefits of relatively long length of sequencing read andthe drawbacks of high reagent cost and high error rates inhomopolymer repeats Metagenomic analysis requires deepsequencing data mining and large amounts of sequencingreads for gene assembly and annotation [24 27 40] Ligation-based sequencing technology provides inherent error cor-rection by two-base encoding which makes the platformmuchmore accurate [34]Due to the short sequencing lengththis methodology has a few restrictions in the microbiomediversity research
In the present study we develop an effective approachusing multigroup amplification and massively parallelligation-based sequencing technology to determine thebacterial diversity In addition we compare the tonguecoating microbial diversities of different TCM tonguecoating types using this method
2 Materials and Methods
21 Samples Tongue coating samples were collected fromtwo groups of lung cancer patients in Nanjing Chest HospitalEach group represented a specific TCM tongue coating typethe thin-white type and the white-greasy type (Figure 1) Atotal of 13 subjects were collected in the morning and allthe volunteers had no breakfast before sampling All subjectshad no oral inflammation and had refrained from brushingteeth and drinking colored beverage for 16 hours beforetesting The tongue coating samples were collected withsterilized cotton swabs mixed with 1ml of Ringerrsquos solutionand stored in minus20∘C immediately until extracting the totalmicrobial genome DNA All the 13 samples were investigatedby DGGE analysis (data not shown) Based on the DGGEresults the bacteria composition was significantly differentbetween different tongue coating types Six typical samplesfrom two tongue coating types were chosen for thoroughparallel sequencing and the clinical parameters of subjectswere shown in Table 1
22 Multiple Group PCR Primers Design The multigroupPCR primers were designed based on Human Oral Micro-biome Database (HOMD) (httpwwwhomdorg) The 16SrRNA gene sequences of a genus containing 4 species atleast in HOMD were picked out as a group and aligned byMEGA (v 402) Totally 421 species belonging to 27 generawere selected The conserved regions flanking the variableregion can be used for the multigroup PCR primers designA specific group primer consisted of 22 bp of sequencescontaining Eco57I recognition site and 20 bp of conservedregion sequences (Figure 2) Table 2 indicates all the 26 pairsof group primers generated
23 DNA Extraction Amplification and Ligation SequencingLibrary Preparation All the samples were centrifuged andresuspended in 1ml of 10x TE buffer (pH 8) The suspensionwas mixed with 100120583l of lysozyme (200mgml) and incu-bated at 37∘C for 1 h Lysis solution containing 50120583l of SDS(10 vv) and 20 120583l of Proteinase K (20mgml) was addedand the mixture was incubated at 55∘C for 3 h The totalbacterial genome DNA was obtained by phenol-chloroformextraction and ethanol precipitation
The PCR amplification was carried out separately by eachof group primers for each sample with the same conditionexcept for the annealing temperature All the PCR reactionswere performed in a final volume of 25120583l containing 1xPCR buffer (Takara) 25mM MgCl
2 200120583M each dNTP
25U of Taq polymerase 20 ng of template DNA and 1 120583Meach forward and reverse primer The PCR mixtures wereinitially denatured at 95∘C for 3min followed by 30 cycles45 s at 94∘C 45 s at (119879m minus 5)
∘C and 45 s at 72∘C ThePCR amplicons were digested using the enzyme Eco57I (Fer-mentas Burlington Canada) according to manufacturerrsquosinstructions The expected fragment DNA was acquiredby agarose gel electrophoresis and purified by QIAEX IIGel Extraction Kit (Qiagen Hilden Germany) PCR ampli-con mixture was prepared by pooling approximately equalamounts of recovered fragments from the same sample Afterblunting the 31015840 protruding termini of mixtures by T4 DNA
BioMed Research International 3
Table 1 Clinical parameters and barcode primer sequences of study subjects
Subjects ID Gender Age TCM tonguecoating type Barcode sequence (51015840 to 31015840)
A4 Male 30 Thin-white CTGCCCCGGGTTCCTCATTCTCTAAGCCCCTGCTGTACGGCCAAGGCGA5 Male 58 Thin-white CTGCCCCGGGTTCCTCATTCTCTCACACCCTGCTGTACGGCCAAGGCGA6 Female 72 Thin-white CTGCCCCGGGTTCCTCATTCTCTTCCCTTCTGCTGTACGGCCAAGGCGB2 Male 57 White-greasy CTGCCCCGGGTTCCTCATTCTCTCCGATTCTGCTGTACGGCCAAGGCGB3 Female 34 White-greasy CTGCCCCGGGTTCCTCATTCTCTTCGTTGCTGCTGTACGGCCAAGGCGB4 Male 62 White-greasy CTGCCCCGGGTTCCTCATTCTCTGGGCACCTGCTGTACGGCCAAGGCG
PRPF
Eco57Idigestion
CF CR
CS CS
gDNA
Sequencing
20 <J 20 <J
6 <J6 <J
4 <J 4 <J
16 <J
16 <J 18 <J
16 <J
35 <J 35 <J
Figure 2 Schematic diagram of the primer design and librarypreparation All the 16S rRNA gene sequences from the same generawere aligned to generate 20 bp of the conserved forward sequence(CF) and conserved reverse sequence (CR) flanking the variableregion The forward primer (PF) and reverse primer (PR) weredesigned by adding 6 bp of Eco57I recognition site and 1618 bp ofauxiliary sequence at the 51015840 end ofCF andCRThemicrobial genomeDNA of each subject was amplified by all the pairs of primers fromdifferent genus producing the amplicons containing the Eco57Icutting site (CS) which was 16 bp downstream of the recognitionsite Followed by Eco57I digestion products generated by all thegenus-specific primers conserving 4 bp of each primer sequencewere mixed together to produce the sequencing library The resultsof ligation-based sequencing were millions of reads which were thefirst 35 bases at the 51015840 end from either forward strand or reversestrand of library
polymerase four sequencing libraries were constructed forthe ligation-based sequencing system
24 Massively Parallel Ligation Sequencing In order to max-imize the sequencing capacity and simplify the workflowof sample preparation all the samples were operated in asingle sequencing run with barcodes added Six barcodes
were ligated to the 31015840 end of the library templates with T4DNA ligase as unique adaptors (Table 1)
SOLiD P1 adaptors (51015840-CCACTACGCCTCCGCTTT-CCTCTCTATGGGCAGTCGGTGAT-31015840) were ligated at the51015840 end of templates followed by standard SOLiD librarypreparation protocol
25 Construction of Reference Sequences Due to the char-acteristic of 2-base encoding in SOLiD sequencing thesequencing reads are in color-space format ldquo0 1 2 3rdquowhich stands for the permutation of the adjacent basesThe brief data processing pipeline in our study wasdesigned as Figure 2 shows because the novel microbialanalysis methods are inappropriate A total of 1049433unaligned 16S rDNA sequences of both bacteria and archaeawere downloaded from Ribosomal Database Project (RDP)(httprdpcmemsuedu) resource (release 10 update 13)Using in silico analysis of amplification by the designedprimers and endonuclease digestion 26 groups of fragmentswere selected Repetitive sequences were moved to construct26 groups of reference sequences (REF-DB) Taxonomiesof REF-DB (TAXA-DB) were assigned using the originalfull-length 16S rRNA gene sequences by the RDP onlineclassifier (httprdpcmemsueduclassifierclassifierjsp) at80 confidence cut-off If the reference was yielded frommore than one full-length 16S rRNA gene sequence eachof the original sequences was assigned separately From thegenus level the taxonomy shared by three-fourthsmajority ofthe full length sequences was defined as the taxonomy of thereferenceOtherwise taxonomyof higher level was compareduntil the domain level
26 Analysis of Sequencing Data Sequencing reads weresplit into six samples according to the 4-mer barcodes inP2 adaptor In color space all the samples were alignedseparately to REF-DB by Corona Lite Program (v42httpsolidsoftwaretoolscom) with up to 3 color-space mis-matches For unique reads (uniquely placed matches) afew steps of filtering were performed Firstly the matchingposition should be at the 51015840 end of the reference sequence(forward-strand matching) or reverse compliment (reverse-strand matching) Meanwhile the top four positions ofreads were checked with one mismatch in color space tofilter out nonspecific amplification or false endonuclease-digested products Secondly the abundance of reads match-ing ldquoforward-strandrdquo and ldquoreverse-strandrdquo references was
4 BioMed Research International
Table 2 The designed multigroup PCR primers
Primer ID Genus name Primer sequence (51015840 - 31015840) 119879m (∘C) Amplicon length (bp)ActF Actinomyces Pa-Eb-GCGAAGAACCTTACCAAGGC 562 142ActR Actinomyces Qc-E-TGACGACAACCATGCACCACPreF Prevotella P-E-GAACCTTACCCGGGCTTGAA 542 138PreR Prevotella Q-E-TGACGACAACCATGCAGCACStrF Streptococcus P-E-AACGATAGCTAATACCGCAT 461 139StrR Streptococcus Q-E-TAATACAACGCAGGTCCATCTreF Treponema P-E-CGCGAGGAACCTTACCTGGG 537 137TreR Treponema Q-E-ACGACAGCCATGCAGCACCTLepF Leptotrichia P-E-ACGCGAGGAACCTTACCAGA 535 139LepR Leptotrichia Q-E-CAGCCATGCACCACCTGTCTNeiF Neisseria P-E-CGGGTTGTAAAGGACTTTTG 492 135NeiR Neisseria Q-E-AGTTAGCCGGTGCTTATTCTSelF Selenomonas P-E-CTTCGGATCGTAAAGCTCTG 514 142SelR Selenomonas Q-E-TAGTTAGCCGTGGCTTCCTCCapF Capnocytophaga P-E-TACGCGAGGAACCTTACCAA 513 138CapR Capnocytophaga Q-E-ACAACCATGCAGCACCTTGAPorF Porphyromonas P-E-CAGCCAAGTCGCGTGAAGGA 540 172PorR Porphyromonas Q-E-CTGGCACGGAGTTAGCCGATFusF Fusobacterium P-E-ACGCGTAAAGAACTTGCCTC 514 75FusR Fusobacterium Q-E-ACGCGTAAAGAACTTGCCTCMycF Mycoplasma P-E-GATGGAGCGACACAGCGTGC 552 187MycR Mycoplasma Q-E-GCGGCTGCTGGCACATAGTTDiaF Dialister P-E-CGGAATTATTGGGCGTAAAG 531 161DiaR Dialister Q-E-CTTTCCTCTCCGATACTCCAEubF Eubacterium P-E-GATACCCTGGTAGTCCACGC 516 146EubR Eubacterium Q-E-CTCCCCAGGTGGAATACTTAFirF Firmicutes P-E-GATACCCTGGTAGTCCACGC 516 146FirR Firmicutes Q-E-CTCCCCAGGTGGAATACTTAPepsF Peptostreptococcus P-E-TAATTCGAAGCAACGCGAAG 527 154PepsR Peptostreptococcus Q-E-CGACAACCATGCACCACCTGKinF Kingella P-E-GGAATTACTGGGCGTAAAGC 524 168KinR Kingella Q-E-AATTCTACCCCCCTCTGACAPepnF Peptoniphilus P-E-ATCACTGGGCGTAAAGGGTT 511 186PepnR Peptoniphilus Q-E-CGCATTTCACCGCTACACTALachF Lachnospiraceae P-E-CAACGCGAAGAACCTTACCA 542 146LachR Lachnospiraceae Q-E-GACGACAACCATGCACCACCPasF Aggregatibacterb P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Aggregatibacter Q-E-TTAGCCGGTGCTTCTTCTGTPasF Haemophilus P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Haemophilus Q-E-TTAGCCGGTGCTTCTTCTGTLactF Lactobacillus P-E-CCGCAACGAGCGCAACCCTT 553 103LactR Lactobacillus CCTCCGGTTTGTCACCGGCATM7F TM7 P-E-GGGCGTAAAGAGTTGCGTAG 492 185TM7R TM7 Q-E-TACGGATTTCACTCCTACACGemF Gemella AAAGCTCTGTTGTTAGGGAA 468 98GemR Gemella P-E-GGTGGCTTTCTGGTTAGGTAPseF Pseudomonas GGCGGCAGGCCTAACACATG 565 99PseR Pseudomonas P-E-TTACTCACCCGTCCGCCGCTVeiF Veillonella P-E-GTAAAGCTCTGTTAATCGGG 481 100VeiR Veillonella GTGGCTTTCTATTCCGGTACMogF Mogibacterium P-E-CACGTGCTACAATGGTCGGT 507 101MogR Mogibacterium ATCCGAACTGGGATCGGTTTCatF Catonella P-E-CATGCAAGTCGAACGGAGAT 508 101CatR Catonella GTTACTCACCCGTCCGCCACaCCAAGGCGGCCGTACG bCTGAAG cCCGACGTCGACTATCCAT
BioMed Research International 5
Table 3 Statistical results of SOLiD sequencing tags mapping with RDP references
Subject ID Raw reads Matched reads Uniquely matched readsCounts Frequency Counts Frequency
A4 12455117 2043364 1641 119454 096A5 5043037 861788 1709 63751 126A6 27658749 4965785 1795 418203 151B2 10171189 1406962 1383 160263 158B3 8938854 1968093 2202 96206 108B4 10848548 1915504 1766 119292 11
Table 4 OTUs assignment of unique tags based on RDP classifier
OTU A4 A5 A6 B2 B3 B4Number PCT () Number PCT () Number PCT () Number PCT () Number PCT () Number PCT ()
Genus 548 9320 406 9490 495 9593 464 9490 388 9490 637 9522Family 15 255 11 257 10 194 12 245 10 245 17 254Order 9 153 3 070 4 078 4 073 3 073 6 090Class 9 153 3 070 6 116 7 122 5 122 8 120Phylum 1 170 0 0 0 0 1 024 1 024 0 0Domain 6 102 5 117 1 019 3 049 2 049 1 015Total OTUs 588 428 516 491 409 669
counted separately and the reads with less than 5 countsin both strands were discarded Finally Operational Taxo-nomic Units (OTUs) were identified according to matchedsequences in the REF-DB and corresponding taxonomyinformation in TAXA-DB For the nonunique reads thatmatched the reference in more than one location all ofthe matched references were assigned at the phylum levelaccording to TAXA-DB A matched read was assigned toa specific taxonomy while all the references it matchedbelonged to accordant phylum Otherwise the read wasdenoted as ldquoundefinedrdquo Both the unique and nonuniquematched reads were summarized by a set of Perl scripts
27 Phylogenetic Analysis The original full-length sequencesof matched OTUs were aligned using ClustalW and a relaxedneighbor-joining tree was built by PHILIP 368 Weightedand unweighted UniFrac were run using the resulting treeand environment annotation of number of hits PCA wasperformed on the resulting matric of distances between eachpair of samples
28 Prediction of Metagenome from 16S rRNA Gene DataThe PICRUSt project aims to support prediction of theunobserved character states in a community of organismsfrom phylogenetic information about the organisms in thatcommunity This program was used to predict metagenomeabundance from 16S rRNA gene data
3 Results
31 Sequencing Performance of SOLiD Reads One-quarter ofslide was used to perform the ligation-based sequencing and
84399432 reads of 35-base length (275 gigabases in total)were captured after removing sequences of insufficient qual-ity Splitted by 4-mer barcodes a total number of 75115494reads (89) were generated (Table 3) The difference ofsequencing throughput among six samples was probably dueto the sample quality or the procedure of library preparation
32 Taxa Assignment of the Sequencing Reads Since SOLiDsystem employs 2-base encoding and color-space strategya single color change is a measurement error two adjacentcolor changes may result from a single nucleotide variationin base space and three color-space mismatches might implytwo adjacent variants compared to reference We used up tothree color changes in 35-base length as the alignment param-eter which implied at most two nucleotide mismatchesConsequently the sequence similarity was more than 94which corresponded to the genus level classification Alignedto REF-DB matched and uniquely matched reads weresummarized in Table 3
To increase the accuracy of taxonomy assignment twosteps of validation were performed before OTUs definitionas Methods described Altogether we discarded 3375 ofthe uniquelymatched reads leaving 647375 sequencing readsfor taxonomy analysis Compared to TAXA-DB number ofOTUswas identified ranging from409 to 669 for six samplesrespectively Most of the OTUs (more than 93) could beassigned at the genus or lower level while a very small amountof OTUs was just assigned at the phylum or higher level(Table 4)
33 Microbiome Diversity of the Tongue Coating SamplesThe dominant phyla (relative abundance gt 1) across allsamples were Firmicutes (5519 plusmn 1600) Proteobacteria
6 BioMed Research International
0102030405060708090
100
Sequ
ence
s cla
ssifi
ed to
phy
lum
leve
l (
)
A5 A6 B2 B3 B4A4
DeferribacteresCyanobacteriaAcidobacteriaChlamydiaeSpirochaetes
ChlorobiActinobacteriaBacteroidetesProteobacteriaFirmicutes
Figure 3 Relative abundances of taxonomy classification of thetongue coating microbiome at the phylum level in the six tonguecoating samples
(2611 plusmn 1485) Bacteroidetes (1480 plusmn 236) Acti-nobacteria (203 plusmn 145) and Chlorobi (117 plusmn 079)Ultimately we identified 209 genera from the six samplesand the abundance of each genus was defined in MethodsAs shown in Figure 3 92 genera were assigned to phylumFirmicutes with the maximum abundance as well PhylaProteobacteria and Bacteroidetes contained some dominantgenera in our results while only 68 and 30 genera wereassigned to these two phyla separately The fifteen mostabundant genera of each sample were illustrated in Figure 4with total abundance ranging from 702 to 82 Therewere 163 genera assigned from 79762 reads for sample A4and the most abundant genera were Haemophilus (1338Proteobacteria) Haliscomenobacter (1023 Bacteroidetes)Enterococcus (950 Firmicutes) Streptococcus (556 Fir-micutes) and Acetanaerobacterium (417 Firmicutes) Forsample A5 134 genera were identified from 41580 readshaving the most abundant genera of Haemophilus (1140)Haliscomenobacter (781) Bacillus a (640 Firmicutes)Streptococcus (550) and Enterococcus (462) There were141 genera assigned from 242085 reads for sample A6and the most abundant genera were Ralstonia (1731 Pro-teobacteria) Anaerostipes (1032 Firmicutes) Bacillus a(953 Firmicutes) Haemophilus (496) and Coprococcus(369 Firmicutes) For white-greasy tongue coating typesamples for sample B2 119 genera were found within 155093reads and the dominant genera were Roseburia (2691Firmicutes) Anaerotruncus (98 Firmicutes) Coprococ-cus (791) Sphingobacterium (581 Bacteroidetes) andDorea (348 Firmicutes) For sample B3 128 genera wereassigned from 47717 reads with the largest proportiongenera of Streptococcus (3425) Sphingobacterium (714)Prevotella (708 Bacteroidetes) Enterococcus (679) andAcetanaerobacterium (450) For sample B4 143 generawerefound within 81138 reads and the dominant genera were
Haemophilus (1903) Sphingobacterium (1065) Entero-coccus (794) Acetanaerobacterium (531) and Bacillus c(432 Firmicutes)Overall the dominantmicrobial phyla ofthe three thin-white tongue coating samples were FirmicutesProteobacteria and Bacteroidetes while the white-greasysamples B2 and B3 had the second abundant phylum ofBacteroidetes and the sample B4 had the most abundantphylum of Proteobacteria
In order tomaximize the high-throughput usable data fordeep analysis both the uniquely and nonuniquely matchedreads were classified at phylum level The percent of assignedreads was ranging from 588 to 656 Compared withthe results based on unique reads the proportion of phylaFirmicutes and Actinobacteria had dramatically increased(data not shown)
34 Shared Genera of the Same Tongue Coating Type SamplesWe compared the shared genera within the same tonguecoating type samples There were 117 genera shared betweenthe thin-white tongue coating type samples Of these genera58were fromFirmicutes 37 generawere fromProteobacteriaand 12 genera belonged to Bacteroidetes For the white-greasytongue coating type samples 96 genera were observed to beshared 48 genera belonged to Firmicutes 28 genera belongedto Proteobacteria and 11 genera belonged to Bacteroidetesrespectively
35 Comparison of Bacterial Diversity between DifferentTongue Coating Type Samples There were 77 genera sharedacross the six samples (Figure 5) 35 genera belonged toFirmicutes 23 genera belonged to Proteobacteria and 8genera belonged to Bacteroidetes We compared the generaobserved in one tongue coating type but not the otherFigure 6 showed the genera that appeared in all the thin-whitetongue coating type but existed in none white-greasy tonguecoating type sample A total of 12 genera were observed onlyin the thin-white tongue coating type samples PelomonasHaemophilus Thioalkalispira and Zoogloea (Proteobacteria)Jeotgalibacillus Granulicatella Lachnobacterium Peptostrep-tococcus Anaeromusa Parasporobacterium and Sporobac-terium (Firmicutes) and Prevotella (Bacteroidetes) Figure 7showed the genera that appeared in all the white-greasytongue coating type but did not exist in all the thin-white tongue coating type sample The white-greasy tonguecoating type samples were specifically unique for 6 gen-era Anaerostipes Lactobacillus Anaerobacter Ruminococ-caceae Incertae sedis and Oribacterium (Firmicutes) andAcidithiobacillus (Proteobacteria) Analyzed by Studentrsquos 119905-test 19 genera performed significantly different between twotongue coating types (Table 5)
Analyzed by UniFrac software the thin-white tonguecoating type and white-greasy tongue coating type wereobservably different at the genus level as shown in principalcomponent analysis (PCA) plot (Figure 8) The three thin-white tongue coating subjects had relatively similar microbialdiversity (Bonferroni-corrected 119875 value is 025) while three
BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
Table 1 Clinical parameters and barcode primer sequences of study subjects
Subjects ID Gender Age TCM tonguecoating type Barcode sequence (51015840 to 31015840)
A4 Male 30 Thin-white CTGCCCCGGGTTCCTCATTCTCTAAGCCCCTGCTGTACGGCCAAGGCGA5 Male 58 Thin-white CTGCCCCGGGTTCCTCATTCTCTCACACCCTGCTGTACGGCCAAGGCGA6 Female 72 Thin-white CTGCCCCGGGTTCCTCATTCTCTTCCCTTCTGCTGTACGGCCAAGGCGB2 Male 57 White-greasy CTGCCCCGGGTTCCTCATTCTCTCCGATTCTGCTGTACGGCCAAGGCGB3 Female 34 White-greasy CTGCCCCGGGTTCCTCATTCTCTTCGTTGCTGCTGTACGGCCAAGGCGB4 Male 62 White-greasy CTGCCCCGGGTTCCTCATTCTCTGGGCACCTGCTGTACGGCCAAGGCG
PRPF
Eco57Idigestion
CF CR
CS CS
gDNA
Sequencing
20 <J 20 <J
6 <J6 <J
4 <J 4 <J
16 <J
16 <J 18 <J
16 <J
35 <J 35 <J
Figure 2 Schematic diagram of the primer design and librarypreparation All the 16S rRNA gene sequences from the same generawere aligned to generate 20 bp of the conserved forward sequence(CF) and conserved reverse sequence (CR) flanking the variableregion The forward primer (PF) and reverse primer (PR) weredesigned by adding 6 bp of Eco57I recognition site and 1618 bp ofauxiliary sequence at the 51015840 end ofCF andCRThemicrobial genomeDNA of each subject was amplified by all the pairs of primers fromdifferent genus producing the amplicons containing the Eco57Icutting site (CS) which was 16 bp downstream of the recognitionsite Followed by Eco57I digestion products generated by all thegenus-specific primers conserving 4 bp of each primer sequencewere mixed together to produce the sequencing library The resultsof ligation-based sequencing were millions of reads which were thefirst 35 bases at the 51015840 end from either forward strand or reversestrand of library
polymerase four sequencing libraries were constructed forthe ligation-based sequencing system
24 Massively Parallel Ligation Sequencing In order to max-imize the sequencing capacity and simplify the workflowof sample preparation all the samples were operated in asingle sequencing run with barcodes added Six barcodes
were ligated to the 31015840 end of the library templates with T4DNA ligase as unique adaptors (Table 1)
SOLiD P1 adaptors (51015840-CCACTACGCCTCCGCTTT-CCTCTCTATGGGCAGTCGGTGAT-31015840) were ligated at the51015840 end of templates followed by standard SOLiD librarypreparation protocol
25 Construction of Reference Sequences Due to the char-acteristic of 2-base encoding in SOLiD sequencing thesequencing reads are in color-space format ldquo0 1 2 3rdquowhich stands for the permutation of the adjacent basesThe brief data processing pipeline in our study wasdesigned as Figure 2 shows because the novel microbialanalysis methods are inappropriate A total of 1049433unaligned 16S rDNA sequences of both bacteria and archaeawere downloaded from Ribosomal Database Project (RDP)(httprdpcmemsuedu) resource (release 10 update 13)Using in silico analysis of amplification by the designedprimers and endonuclease digestion 26 groups of fragmentswere selected Repetitive sequences were moved to construct26 groups of reference sequences (REF-DB) Taxonomiesof REF-DB (TAXA-DB) were assigned using the originalfull-length 16S rRNA gene sequences by the RDP onlineclassifier (httprdpcmemsueduclassifierclassifierjsp) at80 confidence cut-off If the reference was yielded frommore than one full-length 16S rRNA gene sequence eachof the original sequences was assigned separately From thegenus level the taxonomy shared by three-fourthsmajority ofthe full length sequences was defined as the taxonomy of thereferenceOtherwise taxonomyof higher level was compareduntil the domain level
26 Analysis of Sequencing Data Sequencing reads weresplit into six samples according to the 4-mer barcodes inP2 adaptor In color space all the samples were alignedseparately to REF-DB by Corona Lite Program (v42httpsolidsoftwaretoolscom) with up to 3 color-space mis-matches For unique reads (uniquely placed matches) afew steps of filtering were performed Firstly the matchingposition should be at the 51015840 end of the reference sequence(forward-strand matching) or reverse compliment (reverse-strand matching) Meanwhile the top four positions ofreads were checked with one mismatch in color space tofilter out nonspecific amplification or false endonuclease-digested products Secondly the abundance of reads match-ing ldquoforward-strandrdquo and ldquoreverse-strandrdquo references was
4 BioMed Research International
Table 2 The designed multigroup PCR primers
Primer ID Genus name Primer sequence (51015840 - 31015840) 119879m (∘C) Amplicon length (bp)ActF Actinomyces Pa-Eb-GCGAAGAACCTTACCAAGGC 562 142ActR Actinomyces Qc-E-TGACGACAACCATGCACCACPreF Prevotella P-E-GAACCTTACCCGGGCTTGAA 542 138PreR Prevotella Q-E-TGACGACAACCATGCAGCACStrF Streptococcus P-E-AACGATAGCTAATACCGCAT 461 139StrR Streptococcus Q-E-TAATACAACGCAGGTCCATCTreF Treponema P-E-CGCGAGGAACCTTACCTGGG 537 137TreR Treponema Q-E-ACGACAGCCATGCAGCACCTLepF Leptotrichia P-E-ACGCGAGGAACCTTACCAGA 535 139LepR Leptotrichia Q-E-CAGCCATGCACCACCTGTCTNeiF Neisseria P-E-CGGGTTGTAAAGGACTTTTG 492 135NeiR Neisseria Q-E-AGTTAGCCGGTGCTTATTCTSelF Selenomonas P-E-CTTCGGATCGTAAAGCTCTG 514 142SelR Selenomonas Q-E-TAGTTAGCCGTGGCTTCCTCCapF Capnocytophaga P-E-TACGCGAGGAACCTTACCAA 513 138CapR Capnocytophaga Q-E-ACAACCATGCAGCACCTTGAPorF Porphyromonas P-E-CAGCCAAGTCGCGTGAAGGA 540 172PorR Porphyromonas Q-E-CTGGCACGGAGTTAGCCGATFusF Fusobacterium P-E-ACGCGTAAAGAACTTGCCTC 514 75FusR Fusobacterium Q-E-ACGCGTAAAGAACTTGCCTCMycF Mycoplasma P-E-GATGGAGCGACACAGCGTGC 552 187MycR Mycoplasma Q-E-GCGGCTGCTGGCACATAGTTDiaF Dialister P-E-CGGAATTATTGGGCGTAAAG 531 161DiaR Dialister Q-E-CTTTCCTCTCCGATACTCCAEubF Eubacterium P-E-GATACCCTGGTAGTCCACGC 516 146EubR Eubacterium Q-E-CTCCCCAGGTGGAATACTTAFirF Firmicutes P-E-GATACCCTGGTAGTCCACGC 516 146FirR Firmicutes Q-E-CTCCCCAGGTGGAATACTTAPepsF Peptostreptococcus P-E-TAATTCGAAGCAACGCGAAG 527 154PepsR Peptostreptococcus Q-E-CGACAACCATGCACCACCTGKinF Kingella P-E-GGAATTACTGGGCGTAAAGC 524 168KinR Kingella Q-E-AATTCTACCCCCCTCTGACAPepnF Peptoniphilus P-E-ATCACTGGGCGTAAAGGGTT 511 186PepnR Peptoniphilus Q-E-CGCATTTCACCGCTACACTALachF Lachnospiraceae P-E-CAACGCGAAGAACCTTACCA 542 146LachR Lachnospiraceae Q-E-GACGACAACCATGCACCACCPasF Aggregatibacterb P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Aggregatibacter Q-E-TTAGCCGGTGCTTCTTCTGTPasF Haemophilus P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Haemophilus Q-E-TTAGCCGGTGCTTCTTCTGTLactF Lactobacillus P-E-CCGCAACGAGCGCAACCCTT 553 103LactR Lactobacillus CCTCCGGTTTGTCACCGGCATM7F TM7 P-E-GGGCGTAAAGAGTTGCGTAG 492 185TM7R TM7 Q-E-TACGGATTTCACTCCTACACGemF Gemella AAAGCTCTGTTGTTAGGGAA 468 98GemR Gemella P-E-GGTGGCTTTCTGGTTAGGTAPseF Pseudomonas GGCGGCAGGCCTAACACATG 565 99PseR Pseudomonas P-E-TTACTCACCCGTCCGCCGCTVeiF Veillonella P-E-GTAAAGCTCTGTTAATCGGG 481 100VeiR Veillonella GTGGCTTTCTATTCCGGTACMogF Mogibacterium P-E-CACGTGCTACAATGGTCGGT 507 101MogR Mogibacterium ATCCGAACTGGGATCGGTTTCatF Catonella P-E-CATGCAAGTCGAACGGAGAT 508 101CatR Catonella GTTACTCACCCGTCCGCCACaCCAAGGCGGCCGTACG bCTGAAG cCCGACGTCGACTATCCAT
BioMed Research International 5
Table 3 Statistical results of SOLiD sequencing tags mapping with RDP references
Subject ID Raw reads Matched reads Uniquely matched readsCounts Frequency Counts Frequency
A4 12455117 2043364 1641 119454 096A5 5043037 861788 1709 63751 126A6 27658749 4965785 1795 418203 151B2 10171189 1406962 1383 160263 158B3 8938854 1968093 2202 96206 108B4 10848548 1915504 1766 119292 11
Table 4 OTUs assignment of unique tags based on RDP classifier
OTU A4 A5 A6 B2 B3 B4Number PCT () Number PCT () Number PCT () Number PCT () Number PCT () Number PCT ()
Genus 548 9320 406 9490 495 9593 464 9490 388 9490 637 9522Family 15 255 11 257 10 194 12 245 10 245 17 254Order 9 153 3 070 4 078 4 073 3 073 6 090Class 9 153 3 070 6 116 7 122 5 122 8 120Phylum 1 170 0 0 0 0 1 024 1 024 0 0Domain 6 102 5 117 1 019 3 049 2 049 1 015Total OTUs 588 428 516 491 409 669
counted separately and the reads with less than 5 countsin both strands were discarded Finally Operational Taxo-nomic Units (OTUs) were identified according to matchedsequences in the REF-DB and corresponding taxonomyinformation in TAXA-DB For the nonunique reads thatmatched the reference in more than one location all ofthe matched references were assigned at the phylum levelaccording to TAXA-DB A matched read was assigned toa specific taxonomy while all the references it matchedbelonged to accordant phylum Otherwise the read wasdenoted as ldquoundefinedrdquo Both the unique and nonuniquematched reads were summarized by a set of Perl scripts
27 Phylogenetic Analysis The original full-length sequencesof matched OTUs were aligned using ClustalW and a relaxedneighbor-joining tree was built by PHILIP 368 Weightedand unweighted UniFrac were run using the resulting treeand environment annotation of number of hits PCA wasperformed on the resulting matric of distances between eachpair of samples
28 Prediction of Metagenome from 16S rRNA Gene DataThe PICRUSt project aims to support prediction of theunobserved character states in a community of organismsfrom phylogenetic information about the organisms in thatcommunity This program was used to predict metagenomeabundance from 16S rRNA gene data
3 Results
31 Sequencing Performance of SOLiD Reads One-quarter ofslide was used to perform the ligation-based sequencing and
84399432 reads of 35-base length (275 gigabases in total)were captured after removing sequences of insufficient qual-ity Splitted by 4-mer barcodes a total number of 75115494reads (89) were generated (Table 3) The difference ofsequencing throughput among six samples was probably dueto the sample quality or the procedure of library preparation
32 Taxa Assignment of the Sequencing Reads Since SOLiDsystem employs 2-base encoding and color-space strategya single color change is a measurement error two adjacentcolor changes may result from a single nucleotide variationin base space and three color-space mismatches might implytwo adjacent variants compared to reference We used up tothree color changes in 35-base length as the alignment param-eter which implied at most two nucleotide mismatchesConsequently the sequence similarity was more than 94which corresponded to the genus level classification Alignedto REF-DB matched and uniquely matched reads weresummarized in Table 3
To increase the accuracy of taxonomy assignment twosteps of validation were performed before OTUs definitionas Methods described Altogether we discarded 3375 ofthe uniquelymatched reads leaving 647375 sequencing readsfor taxonomy analysis Compared to TAXA-DB number ofOTUswas identified ranging from409 to 669 for six samplesrespectively Most of the OTUs (more than 93) could beassigned at the genus or lower level while a very small amountof OTUs was just assigned at the phylum or higher level(Table 4)
33 Microbiome Diversity of the Tongue Coating SamplesThe dominant phyla (relative abundance gt 1) across allsamples were Firmicutes (5519 plusmn 1600) Proteobacteria
6 BioMed Research International
0102030405060708090
100
Sequ
ence
s cla
ssifi
ed to
phy
lum
leve
l (
)
A5 A6 B2 B3 B4A4
DeferribacteresCyanobacteriaAcidobacteriaChlamydiaeSpirochaetes
ChlorobiActinobacteriaBacteroidetesProteobacteriaFirmicutes
Figure 3 Relative abundances of taxonomy classification of thetongue coating microbiome at the phylum level in the six tonguecoating samples
(2611 plusmn 1485) Bacteroidetes (1480 plusmn 236) Acti-nobacteria (203 plusmn 145) and Chlorobi (117 plusmn 079)Ultimately we identified 209 genera from the six samplesand the abundance of each genus was defined in MethodsAs shown in Figure 3 92 genera were assigned to phylumFirmicutes with the maximum abundance as well PhylaProteobacteria and Bacteroidetes contained some dominantgenera in our results while only 68 and 30 genera wereassigned to these two phyla separately The fifteen mostabundant genera of each sample were illustrated in Figure 4with total abundance ranging from 702 to 82 Therewere 163 genera assigned from 79762 reads for sample A4and the most abundant genera were Haemophilus (1338Proteobacteria) Haliscomenobacter (1023 Bacteroidetes)Enterococcus (950 Firmicutes) Streptococcus (556 Fir-micutes) and Acetanaerobacterium (417 Firmicutes) Forsample A5 134 genera were identified from 41580 readshaving the most abundant genera of Haemophilus (1140)Haliscomenobacter (781) Bacillus a (640 Firmicutes)Streptococcus (550) and Enterococcus (462) There were141 genera assigned from 242085 reads for sample A6and the most abundant genera were Ralstonia (1731 Pro-teobacteria) Anaerostipes (1032 Firmicutes) Bacillus a(953 Firmicutes) Haemophilus (496) and Coprococcus(369 Firmicutes) For white-greasy tongue coating typesamples for sample B2 119 genera were found within 155093reads and the dominant genera were Roseburia (2691Firmicutes) Anaerotruncus (98 Firmicutes) Coprococ-cus (791) Sphingobacterium (581 Bacteroidetes) andDorea (348 Firmicutes) For sample B3 128 genera wereassigned from 47717 reads with the largest proportiongenera of Streptococcus (3425) Sphingobacterium (714)Prevotella (708 Bacteroidetes) Enterococcus (679) andAcetanaerobacterium (450) For sample B4 143 generawerefound within 81138 reads and the dominant genera were
Haemophilus (1903) Sphingobacterium (1065) Entero-coccus (794) Acetanaerobacterium (531) and Bacillus c(432 Firmicutes)Overall the dominantmicrobial phyla ofthe three thin-white tongue coating samples were FirmicutesProteobacteria and Bacteroidetes while the white-greasysamples B2 and B3 had the second abundant phylum ofBacteroidetes and the sample B4 had the most abundantphylum of Proteobacteria
In order tomaximize the high-throughput usable data fordeep analysis both the uniquely and nonuniquely matchedreads were classified at phylum level The percent of assignedreads was ranging from 588 to 656 Compared withthe results based on unique reads the proportion of phylaFirmicutes and Actinobacteria had dramatically increased(data not shown)
34 Shared Genera of the Same Tongue Coating Type SamplesWe compared the shared genera within the same tonguecoating type samples There were 117 genera shared betweenthe thin-white tongue coating type samples Of these genera58were fromFirmicutes 37 generawere fromProteobacteriaand 12 genera belonged to Bacteroidetes For the white-greasytongue coating type samples 96 genera were observed to beshared 48 genera belonged to Firmicutes 28 genera belongedto Proteobacteria and 11 genera belonged to Bacteroidetesrespectively
35 Comparison of Bacterial Diversity between DifferentTongue Coating Type Samples There were 77 genera sharedacross the six samples (Figure 5) 35 genera belonged toFirmicutes 23 genera belonged to Proteobacteria and 8genera belonged to Bacteroidetes We compared the generaobserved in one tongue coating type but not the otherFigure 6 showed the genera that appeared in all the thin-whitetongue coating type but existed in none white-greasy tonguecoating type sample A total of 12 genera were observed onlyin the thin-white tongue coating type samples PelomonasHaemophilus Thioalkalispira and Zoogloea (Proteobacteria)Jeotgalibacillus Granulicatella Lachnobacterium Peptostrep-tococcus Anaeromusa Parasporobacterium and Sporobac-terium (Firmicutes) and Prevotella (Bacteroidetes) Figure 7showed the genera that appeared in all the white-greasytongue coating type but did not exist in all the thin-white tongue coating type sample The white-greasy tonguecoating type samples were specifically unique for 6 gen-era Anaerostipes Lactobacillus Anaerobacter Ruminococ-caceae Incertae sedis and Oribacterium (Firmicutes) andAcidithiobacillus (Proteobacteria) Analyzed by Studentrsquos 119905-test 19 genera performed significantly different between twotongue coating types (Table 5)
Analyzed by UniFrac software the thin-white tonguecoating type and white-greasy tongue coating type wereobservably different at the genus level as shown in principalcomponent analysis (PCA) plot (Figure 8) The three thin-white tongue coating subjects had relatively similar microbialdiversity (Bonferroni-corrected 119875 value is 025) while three
BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
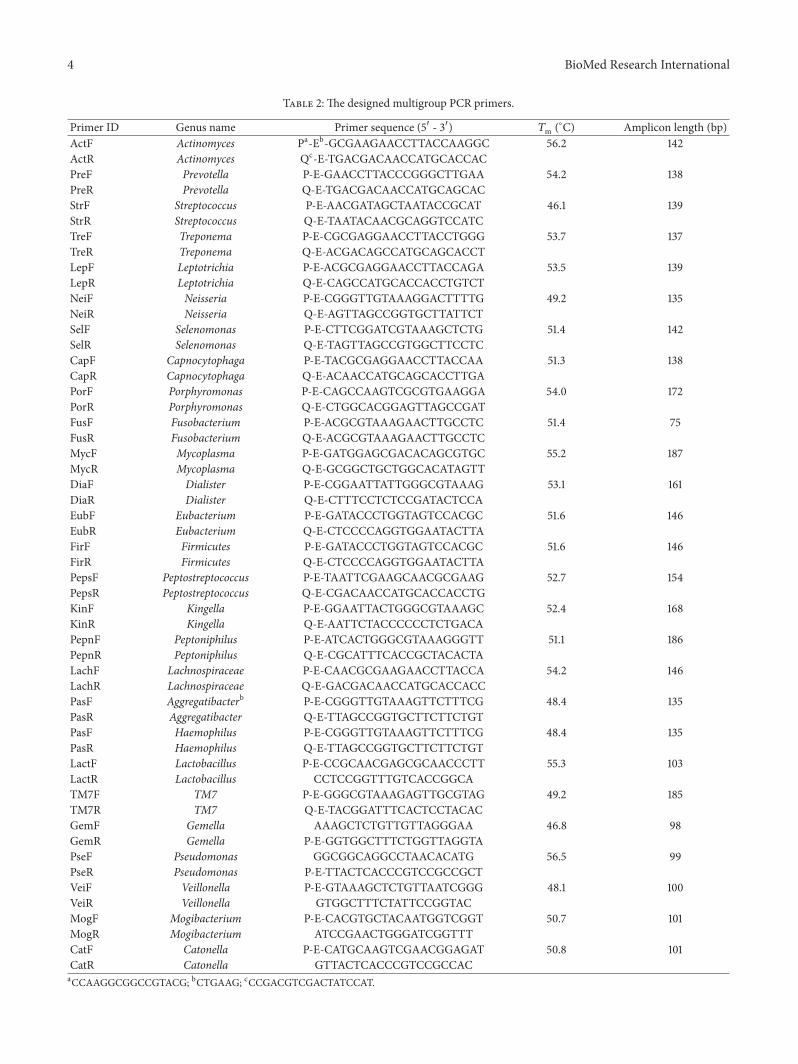
4 BioMed Research International
Table 2 The designed multigroup PCR primers
Primer ID Genus name Primer sequence (51015840 - 31015840) 119879m (∘C) Amplicon length (bp)ActF Actinomyces Pa-Eb-GCGAAGAACCTTACCAAGGC 562 142ActR Actinomyces Qc-E-TGACGACAACCATGCACCACPreF Prevotella P-E-GAACCTTACCCGGGCTTGAA 542 138PreR Prevotella Q-E-TGACGACAACCATGCAGCACStrF Streptococcus P-E-AACGATAGCTAATACCGCAT 461 139StrR Streptococcus Q-E-TAATACAACGCAGGTCCATCTreF Treponema P-E-CGCGAGGAACCTTACCTGGG 537 137TreR Treponema Q-E-ACGACAGCCATGCAGCACCTLepF Leptotrichia P-E-ACGCGAGGAACCTTACCAGA 535 139LepR Leptotrichia Q-E-CAGCCATGCACCACCTGTCTNeiF Neisseria P-E-CGGGTTGTAAAGGACTTTTG 492 135NeiR Neisseria Q-E-AGTTAGCCGGTGCTTATTCTSelF Selenomonas P-E-CTTCGGATCGTAAAGCTCTG 514 142SelR Selenomonas Q-E-TAGTTAGCCGTGGCTTCCTCCapF Capnocytophaga P-E-TACGCGAGGAACCTTACCAA 513 138CapR Capnocytophaga Q-E-ACAACCATGCAGCACCTTGAPorF Porphyromonas P-E-CAGCCAAGTCGCGTGAAGGA 540 172PorR Porphyromonas Q-E-CTGGCACGGAGTTAGCCGATFusF Fusobacterium P-E-ACGCGTAAAGAACTTGCCTC 514 75FusR Fusobacterium Q-E-ACGCGTAAAGAACTTGCCTCMycF Mycoplasma P-E-GATGGAGCGACACAGCGTGC 552 187MycR Mycoplasma Q-E-GCGGCTGCTGGCACATAGTTDiaF Dialister P-E-CGGAATTATTGGGCGTAAAG 531 161DiaR Dialister Q-E-CTTTCCTCTCCGATACTCCAEubF Eubacterium P-E-GATACCCTGGTAGTCCACGC 516 146EubR Eubacterium Q-E-CTCCCCAGGTGGAATACTTAFirF Firmicutes P-E-GATACCCTGGTAGTCCACGC 516 146FirR Firmicutes Q-E-CTCCCCAGGTGGAATACTTAPepsF Peptostreptococcus P-E-TAATTCGAAGCAACGCGAAG 527 154PepsR Peptostreptococcus Q-E-CGACAACCATGCACCACCTGKinF Kingella P-E-GGAATTACTGGGCGTAAAGC 524 168KinR Kingella Q-E-AATTCTACCCCCCTCTGACAPepnF Peptoniphilus P-E-ATCACTGGGCGTAAAGGGTT 511 186PepnR Peptoniphilus Q-E-CGCATTTCACCGCTACACTALachF Lachnospiraceae P-E-CAACGCGAAGAACCTTACCA 542 146LachR Lachnospiraceae Q-E-GACGACAACCATGCACCACCPasF Aggregatibacterb P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Aggregatibacter Q-E-TTAGCCGGTGCTTCTTCTGTPasF Haemophilus P-E-CGGGTTGTAAAGTTCTTTCG 484 135PasR Haemophilus Q-E-TTAGCCGGTGCTTCTTCTGTLactF Lactobacillus P-E-CCGCAACGAGCGCAACCCTT 553 103LactR Lactobacillus CCTCCGGTTTGTCACCGGCATM7F TM7 P-E-GGGCGTAAAGAGTTGCGTAG 492 185TM7R TM7 Q-E-TACGGATTTCACTCCTACACGemF Gemella AAAGCTCTGTTGTTAGGGAA 468 98GemR Gemella P-E-GGTGGCTTTCTGGTTAGGTAPseF Pseudomonas GGCGGCAGGCCTAACACATG 565 99PseR Pseudomonas P-E-TTACTCACCCGTCCGCCGCTVeiF Veillonella P-E-GTAAAGCTCTGTTAATCGGG 481 100VeiR Veillonella GTGGCTTTCTATTCCGGTACMogF Mogibacterium P-E-CACGTGCTACAATGGTCGGT 507 101MogR Mogibacterium ATCCGAACTGGGATCGGTTTCatF Catonella P-E-CATGCAAGTCGAACGGAGAT 508 101CatR Catonella GTTACTCACCCGTCCGCCACaCCAAGGCGGCCGTACG bCTGAAG cCCGACGTCGACTATCCAT
BioMed Research International 5
Table 3 Statistical results of SOLiD sequencing tags mapping with RDP references
Subject ID Raw reads Matched reads Uniquely matched readsCounts Frequency Counts Frequency
A4 12455117 2043364 1641 119454 096A5 5043037 861788 1709 63751 126A6 27658749 4965785 1795 418203 151B2 10171189 1406962 1383 160263 158B3 8938854 1968093 2202 96206 108B4 10848548 1915504 1766 119292 11
Table 4 OTUs assignment of unique tags based on RDP classifier
OTU A4 A5 A6 B2 B3 B4Number PCT () Number PCT () Number PCT () Number PCT () Number PCT () Number PCT ()
Genus 548 9320 406 9490 495 9593 464 9490 388 9490 637 9522Family 15 255 11 257 10 194 12 245 10 245 17 254Order 9 153 3 070 4 078 4 073 3 073 6 090Class 9 153 3 070 6 116 7 122 5 122 8 120Phylum 1 170 0 0 0 0 1 024 1 024 0 0Domain 6 102 5 117 1 019 3 049 2 049 1 015Total OTUs 588 428 516 491 409 669
counted separately and the reads with less than 5 countsin both strands were discarded Finally Operational Taxo-nomic Units (OTUs) were identified according to matchedsequences in the REF-DB and corresponding taxonomyinformation in TAXA-DB For the nonunique reads thatmatched the reference in more than one location all ofthe matched references were assigned at the phylum levelaccording to TAXA-DB A matched read was assigned toa specific taxonomy while all the references it matchedbelonged to accordant phylum Otherwise the read wasdenoted as ldquoundefinedrdquo Both the unique and nonuniquematched reads were summarized by a set of Perl scripts
27 Phylogenetic Analysis The original full-length sequencesof matched OTUs were aligned using ClustalW and a relaxedneighbor-joining tree was built by PHILIP 368 Weightedand unweighted UniFrac were run using the resulting treeand environment annotation of number of hits PCA wasperformed on the resulting matric of distances between eachpair of samples
28 Prediction of Metagenome from 16S rRNA Gene DataThe PICRUSt project aims to support prediction of theunobserved character states in a community of organismsfrom phylogenetic information about the organisms in thatcommunity This program was used to predict metagenomeabundance from 16S rRNA gene data
3 Results
31 Sequencing Performance of SOLiD Reads One-quarter ofslide was used to perform the ligation-based sequencing and
84399432 reads of 35-base length (275 gigabases in total)were captured after removing sequences of insufficient qual-ity Splitted by 4-mer barcodes a total number of 75115494reads (89) were generated (Table 3) The difference ofsequencing throughput among six samples was probably dueto the sample quality or the procedure of library preparation
32 Taxa Assignment of the Sequencing Reads Since SOLiDsystem employs 2-base encoding and color-space strategya single color change is a measurement error two adjacentcolor changes may result from a single nucleotide variationin base space and three color-space mismatches might implytwo adjacent variants compared to reference We used up tothree color changes in 35-base length as the alignment param-eter which implied at most two nucleotide mismatchesConsequently the sequence similarity was more than 94which corresponded to the genus level classification Alignedto REF-DB matched and uniquely matched reads weresummarized in Table 3
To increase the accuracy of taxonomy assignment twosteps of validation were performed before OTUs definitionas Methods described Altogether we discarded 3375 ofthe uniquelymatched reads leaving 647375 sequencing readsfor taxonomy analysis Compared to TAXA-DB number ofOTUswas identified ranging from409 to 669 for six samplesrespectively Most of the OTUs (more than 93) could beassigned at the genus or lower level while a very small amountof OTUs was just assigned at the phylum or higher level(Table 4)
33 Microbiome Diversity of the Tongue Coating SamplesThe dominant phyla (relative abundance gt 1) across allsamples were Firmicutes (5519 plusmn 1600) Proteobacteria
6 BioMed Research International
0102030405060708090
100
Sequ
ence
s cla
ssifi
ed to
phy
lum
leve
l (
)
A5 A6 B2 B3 B4A4
DeferribacteresCyanobacteriaAcidobacteriaChlamydiaeSpirochaetes
ChlorobiActinobacteriaBacteroidetesProteobacteriaFirmicutes
Figure 3 Relative abundances of taxonomy classification of thetongue coating microbiome at the phylum level in the six tonguecoating samples
(2611 plusmn 1485) Bacteroidetes (1480 plusmn 236) Acti-nobacteria (203 plusmn 145) and Chlorobi (117 plusmn 079)Ultimately we identified 209 genera from the six samplesand the abundance of each genus was defined in MethodsAs shown in Figure 3 92 genera were assigned to phylumFirmicutes with the maximum abundance as well PhylaProteobacteria and Bacteroidetes contained some dominantgenera in our results while only 68 and 30 genera wereassigned to these two phyla separately The fifteen mostabundant genera of each sample were illustrated in Figure 4with total abundance ranging from 702 to 82 Therewere 163 genera assigned from 79762 reads for sample A4and the most abundant genera were Haemophilus (1338Proteobacteria) Haliscomenobacter (1023 Bacteroidetes)Enterococcus (950 Firmicutes) Streptococcus (556 Fir-micutes) and Acetanaerobacterium (417 Firmicutes) Forsample A5 134 genera were identified from 41580 readshaving the most abundant genera of Haemophilus (1140)Haliscomenobacter (781) Bacillus a (640 Firmicutes)Streptococcus (550) and Enterococcus (462) There were141 genera assigned from 242085 reads for sample A6and the most abundant genera were Ralstonia (1731 Pro-teobacteria) Anaerostipes (1032 Firmicutes) Bacillus a(953 Firmicutes) Haemophilus (496) and Coprococcus(369 Firmicutes) For white-greasy tongue coating typesamples for sample B2 119 genera were found within 155093reads and the dominant genera were Roseburia (2691Firmicutes) Anaerotruncus (98 Firmicutes) Coprococ-cus (791) Sphingobacterium (581 Bacteroidetes) andDorea (348 Firmicutes) For sample B3 128 genera wereassigned from 47717 reads with the largest proportiongenera of Streptococcus (3425) Sphingobacterium (714)Prevotella (708 Bacteroidetes) Enterococcus (679) andAcetanaerobacterium (450) For sample B4 143 generawerefound within 81138 reads and the dominant genera were
Haemophilus (1903) Sphingobacterium (1065) Entero-coccus (794) Acetanaerobacterium (531) and Bacillus c(432 Firmicutes)Overall the dominantmicrobial phyla ofthe three thin-white tongue coating samples were FirmicutesProteobacteria and Bacteroidetes while the white-greasysamples B2 and B3 had the second abundant phylum ofBacteroidetes and the sample B4 had the most abundantphylum of Proteobacteria
In order tomaximize the high-throughput usable data fordeep analysis both the uniquely and nonuniquely matchedreads were classified at phylum level The percent of assignedreads was ranging from 588 to 656 Compared withthe results based on unique reads the proportion of phylaFirmicutes and Actinobacteria had dramatically increased(data not shown)
34 Shared Genera of the Same Tongue Coating Type SamplesWe compared the shared genera within the same tonguecoating type samples There were 117 genera shared betweenthe thin-white tongue coating type samples Of these genera58were fromFirmicutes 37 generawere fromProteobacteriaand 12 genera belonged to Bacteroidetes For the white-greasytongue coating type samples 96 genera were observed to beshared 48 genera belonged to Firmicutes 28 genera belongedto Proteobacteria and 11 genera belonged to Bacteroidetesrespectively
35 Comparison of Bacterial Diversity between DifferentTongue Coating Type Samples There were 77 genera sharedacross the six samples (Figure 5) 35 genera belonged toFirmicutes 23 genera belonged to Proteobacteria and 8genera belonged to Bacteroidetes We compared the generaobserved in one tongue coating type but not the otherFigure 6 showed the genera that appeared in all the thin-whitetongue coating type but existed in none white-greasy tonguecoating type sample A total of 12 genera were observed onlyin the thin-white tongue coating type samples PelomonasHaemophilus Thioalkalispira and Zoogloea (Proteobacteria)Jeotgalibacillus Granulicatella Lachnobacterium Peptostrep-tococcus Anaeromusa Parasporobacterium and Sporobac-terium (Firmicutes) and Prevotella (Bacteroidetes) Figure 7showed the genera that appeared in all the white-greasytongue coating type but did not exist in all the thin-white tongue coating type sample The white-greasy tonguecoating type samples were specifically unique for 6 gen-era Anaerostipes Lactobacillus Anaerobacter Ruminococ-caceae Incertae sedis and Oribacterium (Firmicutes) andAcidithiobacillus (Proteobacteria) Analyzed by Studentrsquos 119905-test 19 genera performed significantly different between twotongue coating types (Table 5)
Analyzed by UniFrac software the thin-white tonguecoating type and white-greasy tongue coating type wereobservably different at the genus level as shown in principalcomponent analysis (PCA) plot (Figure 8) The three thin-white tongue coating subjects had relatively similar microbialdiversity (Bonferroni-corrected 119875 value is 025) while three
BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
Table 3 Statistical results of SOLiD sequencing tags mapping with RDP references
Subject ID Raw reads Matched reads Uniquely matched readsCounts Frequency Counts Frequency
A4 12455117 2043364 1641 119454 096A5 5043037 861788 1709 63751 126A6 27658749 4965785 1795 418203 151B2 10171189 1406962 1383 160263 158B3 8938854 1968093 2202 96206 108B4 10848548 1915504 1766 119292 11
Table 4 OTUs assignment of unique tags based on RDP classifier
OTU A4 A5 A6 B2 B3 B4Number PCT () Number PCT () Number PCT () Number PCT () Number PCT () Number PCT ()
Genus 548 9320 406 9490 495 9593 464 9490 388 9490 637 9522Family 15 255 11 257 10 194 12 245 10 245 17 254Order 9 153 3 070 4 078 4 073 3 073 6 090Class 9 153 3 070 6 116 7 122 5 122 8 120Phylum 1 170 0 0 0 0 1 024 1 024 0 0Domain 6 102 5 117 1 019 3 049 2 049 1 015Total OTUs 588 428 516 491 409 669
counted separately and the reads with less than 5 countsin both strands were discarded Finally Operational Taxo-nomic Units (OTUs) were identified according to matchedsequences in the REF-DB and corresponding taxonomyinformation in TAXA-DB For the nonunique reads thatmatched the reference in more than one location all ofthe matched references were assigned at the phylum levelaccording to TAXA-DB A matched read was assigned toa specific taxonomy while all the references it matchedbelonged to accordant phylum Otherwise the read wasdenoted as ldquoundefinedrdquo Both the unique and nonuniquematched reads were summarized by a set of Perl scripts
27 Phylogenetic Analysis The original full-length sequencesof matched OTUs were aligned using ClustalW and a relaxedneighbor-joining tree was built by PHILIP 368 Weightedand unweighted UniFrac were run using the resulting treeand environment annotation of number of hits PCA wasperformed on the resulting matric of distances between eachpair of samples
28 Prediction of Metagenome from 16S rRNA Gene DataThe PICRUSt project aims to support prediction of theunobserved character states in a community of organismsfrom phylogenetic information about the organisms in thatcommunity This program was used to predict metagenomeabundance from 16S rRNA gene data
3 Results
31 Sequencing Performance of SOLiD Reads One-quarter ofslide was used to perform the ligation-based sequencing and
84399432 reads of 35-base length (275 gigabases in total)were captured after removing sequences of insufficient qual-ity Splitted by 4-mer barcodes a total number of 75115494reads (89) were generated (Table 3) The difference ofsequencing throughput among six samples was probably dueto the sample quality or the procedure of library preparation
32 Taxa Assignment of the Sequencing Reads Since SOLiDsystem employs 2-base encoding and color-space strategya single color change is a measurement error two adjacentcolor changes may result from a single nucleotide variationin base space and three color-space mismatches might implytwo adjacent variants compared to reference We used up tothree color changes in 35-base length as the alignment param-eter which implied at most two nucleotide mismatchesConsequently the sequence similarity was more than 94which corresponded to the genus level classification Alignedto REF-DB matched and uniquely matched reads weresummarized in Table 3
To increase the accuracy of taxonomy assignment twosteps of validation were performed before OTUs definitionas Methods described Altogether we discarded 3375 ofthe uniquelymatched reads leaving 647375 sequencing readsfor taxonomy analysis Compared to TAXA-DB number ofOTUswas identified ranging from409 to 669 for six samplesrespectively Most of the OTUs (more than 93) could beassigned at the genus or lower level while a very small amountof OTUs was just assigned at the phylum or higher level(Table 4)
33 Microbiome Diversity of the Tongue Coating SamplesThe dominant phyla (relative abundance gt 1) across allsamples were Firmicutes (5519 plusmn 1600) Proteobacteria
6 BioMed Research International
0102030405060708090
100
Sequ
ence
s cla
ssifi
ed to
phy
lum
leve
l (
)
A5 A6 B2 B3 B4A4
DeferribacteresCyanobacteriaAcidobacteriaChlamydiaeSpirochaetes
ChlorobiActinobacteriaBacteroidetesProteobacteriaFirmicutes
Figure 3 Relative abundances of taxonomy classification of thetongue coating microbiome at the phylum level in the six tonguecoating samples
(2611 plusmn 1485) Bacteroidetes (1480 plusmn 236) Acti-nobacteria (203 plusmn 145) and Chlorobi (117 plusmn 079)Ultimately we identified 209 genera from the six samplesand the abundance of each genus was defined in MethodsAs shown in Figure 3 92 genera were assigned to phylumFirmicutes with the maximum abundance as well PhylaProteobacteria and Bacteroidetes contained some dominantgenera in our results while only 68 and 30 genera wereassigned to these two phyla separately The fifteen mostabundant genera of each sample were illustrated in Figure 4with total abundance ranging from 702 to 82 Therewere 163 genera assigned from 79762 reads for sample A4and the most abundant genera were Haemophilus (1338Proteobacteria) Haliscomenobacter (1023 Bacteroidetes)Enterococcus (950 Firmicutes) Streptococcus (556 Fir-micutes) and Acetanaerobacterium (417 Firmicutes) Forsample A5 134 genera were identified from 41580 readshaving the most abundant genera of Haemophilus (1140)Haliscomenobacter (781) Bacillus a (640 Firmicutes)Streptococcus (550) and Enterococcus (462) There were141 genera assigned from 242085 reads for sample A6and the most abundant genera were Ralstonia (1731 Pro-teobacteria) Anaerostipes (1032 Firmicutes) Bacillus a(953 Firmicutes) Haemophilus (496) and Coprococcus(369 Firmicutes) For white-greasy tongue coating typesamples for sample B2 119 genera were found within 155093reads and the dominant genera were Roseburia (2691Firmicutes) Anaerotruncus (98 Firmicutes) Coprococ-cus (791) Sphingobacterium (581 Bacteroidetes) andDorea (348 Firmicutes) For sample B3 128 genera wereassigned from 47717 reads with the largest proportiongenera of Streptococcus (3425) Sphingobacterium (714)Prevotella (708 Bacteroidetes) Enterococcus (679) andAcetanaerobacterium (450) For sample B4 143 generawerefound within 81138 reads and the dominant genera were
Haemophilus (1903) Sphingobacterium (1065) Entero-coccus (794) Acetanaerobacterium (531) and Bacillus c(432 Firmicutes)Overall the dominantmicrobial phyla ofthe three thin-white tongue coating samples were FirmicutesProteobacteria and Bacteroidetes while the white-greasysamples B2 and B3 had the second abundant phylum ofBacteroidetes and the sample B4 had the most abundantphylum of Proteobacteria
In order tomaximize the high-throughput usable data fordeep analysis both the uniquely and nonuniquely matchedreads were classified at phylum level The percent of assignedreads was ranging from 588 to 656 Compared withthe results based on unique reads the proportion of phylaFirmicutes and Actinobacteria had dramatically increased(data not shown)
34 Shared Genera of the Same Tongue Coating Type SamplesWe compared the shared genera within the same tonguecoating type samples There were 117 genera shared betweenthe thin-white tongue coating type samples Of these genera58were fromFirmicutes 37 generawere fromProteobacteriaand 12 genera belonged to Bacteroidetes For the white-greasytongue coating type samples 96 genera were observed to beshared 48 genera belonged to Firmicutes 28 genera belongedto Proteobacteria and 11 genera belonged to Bacteroidetesrespectively
35 Comparison of Bacterial Diversity between DifferentTongue Coating Type Samples There were 77 genera sharedacross the six samples (Figure 5) 35 genera belonged toFirmicutes 23 genera belonged to Proteobacteria and 8genera belonged to Bacteroidetes We compared the generaobserved in one tongue coating type but not the otherFigure 6 showed the genera that appeared in all the thin-whitetongue coating type but existed in none white-greasy tonguecoating type sample A total of 12 genera were observed onlyin the thin-white tongue coating type samples PelomonasHaemophilus Thioalkalispira and Zoogloea (Proteobacteria)Jeotgalibacillus Granulicatella Lachnobacterium Peptostrep-tococcus Anaeromusa Parasporobacterium and Sporobac-terium (Firmicutes) and Prevotella (Bacteroidetes) Figure 7showed the genera that appeared in all the white-greasytongue coating type but did not exist in all the thin-white tongue coating type sample The white-greasy tonguecoating type samples were specifically unique for 6 gen-era Anaerostipes Lactobacillus Anaerobacter Ruminococ-caceae Incertae sedis and Oribacterium (Firmicutes) andAcidithiobacillus (Proteobacteria) Analyzed by Studentrsquos 119905-test 19 genera performed significantly different between twotongue coating types (Table 5)
Analyzed by UniFrac software the thin-white tonguecoating type and white-greasy tongue coating type wereobservably different at the genus level as shown in principalcomponent analysis (PCA) plot (Figure 8) The three thin-white tongue coating subjects had relatively similar microbialdiversity (Bonferroni-corrected 119875 value is 025) while three
BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
0102030405060708090
100
Sequ
ence
s cla
ssifi
ed to
phy
lum
leve
l (
)
A5 A6 B2 B3 B4A4
DeferribacteresCyanobacteriaAcidobacteriaChlamydiaeSpirochaetes
ChlorobiActinobacteriaBacteroidetesProteobacteriaFirmicutes
Figure 3 Relative abundances of taxonomy classification of thetongue coating microbiome at the phylum level in the six tonguecoating samples
(2611 plusmn 1485) Bacteroidetes (1480 plusmn 236) Acti-nobacteria (203 plusmn 145) and Chlorobi (117 plusmn 079)Ultimately we identified 209 genera from the six samplesand the abundance of each genus was defined in MethodsAs shown in Figure 3 92 genera were assigned to phylumFirmicutes with the maximum abundance as well PhylaProteobacteria and Bacteroidetes contained some dominantgenera in our results while only 68 and 30 genera wereassigned to these two phyla separately The fifteen mostabundant genera of each sample were illustrated in Figure 4with total abundance ranging from 702 to 82 Therewere 163 genera assigned from 79762 reads for sample A4and the most abundant genera were Haemophilus (1338Proteobacteria) Haliscomenobacter (1023 Bacteroidetes)Enterococcus (950 Firmicutes) Streptococcus (556 Fir-micutes) and Acetanaerobacterium (417 Firmicutes) Forsample A5 134 genera were identified from 41580 readshaving the most abundant genera of Haemophilus (1140)Haliscomenobacter (781) Bacillus a (640 Firmicutes)Streptococcus (550) and Enterococcus (462) There were141 genera assigned from 242085 reads for sample A6and the most abundant genera were Ralstonia (1731 Pro-teobacteria) Anaerostipes (1032 Firmicutes) Bacillus a(953 Firmicutes) Haemophilus (496) and Coprococcus(369 Firmicutes) For white-greasy tongue coating typesamples for sample B2 119 genera were found within 155093reads and the dominant genera were Roseburia (2691Firmicutes) Anaerotruncus (98 Firmicutes) Coprococ-cus (791) Sphingobacterium (581 Bacteroidetes) andDorea (348 Firmicutes) For sample B3 128 genera wereassigned from 47717 reads with the largest proportiongenera of Streptococcus (3425) Sphingobacterium (714)Prevotella (708 Bacteroidetes) Enterococcus (679) andAcetanaerobacterium (450) For sample B4 143 generawerefound within 81138 reads and the dominant genera were
Haemophilus (1903) Sphingobacterium (1065) Entero-coccus (794) Acetanaerobacterium (531) and Bacillus c(432 Firmicutes)Overall the dominantmicrobial phyla ofthe three thin-white tongue coating samples were FirmicutesProteobacteria and Bacteroidetes while the white-greasysamples B2 and B3 had the second abundant phylum ofBacteroidetes and the sample B4 had the most abundantphylum of Proteobacteria
In order tomaximize the high-throughput usable data fordeep analysis both the uniquely and nonuniquely matchedreads were classified at phylum level The percent of assignedreads was ranging from 588 to 656 Compared withthe results based on unique reads the proportion of phylaFirmicutes and Actinobacteria had dramatically increased(data not shown)
34 Shared Genera of the Same Tongue Coating Type SamplesWe compared the shared genera within the same tonguecoating type samples There were 117 genera shared betweenthe thin-white tongue coating type samples Of these genera58were fromFirmicutes 37 generawere fromProteobacteriaand 12 genera belonged to Bacteroidetes For the white-greasytongue coating type samples 96 genera were observed to beshared 48 genera belonged to Firmicutes 28 genera belongedto Proteobacteria and 11 genera belonged to Bacteroidetesrespectively
35 Comparison of Bacterial Diversity between DifferentTongue Coating Type Samples There were 77 genera sharedacross the six samples (Figure 5) 35 genera belonged toFirmicutes 23 genera belonged to Proteobacteria and 8genera belonged to Bacteroidetes We compared the generaobserved in one tongue coating type but not the otherFigure 6 showed the genera that appeared in all the thin-whitetongue coating type but existed in none white-greasy tonguecoating type sample A total of 12 genera were observed onlyin the thin-white tongue coating type samples PelomonasHaemophilus Thioalkalispira and Zoogloea (Proteobacteria)Jeotgalibacillus Granulicatella Lachnobacterium Peptostrep-tococcus Anaeromusa Parasporobacterium and Sporobac-terium (Firmicutes) and Prevotella (Bacteroidetes) Figure 7showed the genera that appeared in all the white-greasytongue coating type but did not exist in all the thin-white tongue coating type sample The white-greasy tonguecoating type samples were specifically unique for 6 gen-era Anaerostipes Lactobacillus Anaerobacter Ruminococ-caceae Incertae sedis and Oribacterium (Firmicutes) andAcidithiobacillus (Proteobacteria) Analyzed by Studentrsquos 119905-test 19 genera performed significantly different between twotongue coating types (Table 5)
Analyzed by UniFrac software the thin-white tonguecoating type and white-greasy tongue coating type wereobservably different at the genus level as shown in principalcomponent analysis (PCA) plot (Figure 8) The three thin-white tongue coating subjects had relatively similar microbialdiversity (Bonferroni-corrected 119875 value is 025) while three
BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7
23 27
10
86333
33
22
22
1 1
333
35
5
545
44
55
7
8
44
4
4
46
6
9
11
2023
18
7 7
7
22
22
111
33
33 3 33
2
22
22
2
2917
10
102
2
1
33
3
3
22
12
28
14181083
1006589
441380368
257256253
225188
188187
178
2982A4
A5 B3
B4A6
B2
HaemophilusEnterococcusAcetanaerobacteriumKingellaPasteuriaceae Incertae sedisCatonella
MethylibiumChloroherpetonHaliscomenobacterStreptococcusPrevotella
LactobacillusHalocellaRothiaMogibacteriumOthers
RoseburiaSphingobacteriumEnterococcusSubdoligranulumSporobacterOthers
AnaerotruncusDoreaAcetivibrioPrevotellaBacteroides
CoprococcusFaecalibacteriumEthanoligenensPaenibacillusPeptococcus
HaemophilusBacillus aEnterococcusHydrogenovibrio
DoreaPrevotella
Bacillus cCatonellaHaliscomenobacterStreptococcusSyntrophococcus
AcetanaerobacteriumPasteuriaceae Incertae sedisPeptostreptococcaceae Incertae sedisOthers
Methylibium
Bacillus cEnterococcus
EscherichiaLactobacillus
Streptococcus
Others
Actinomyces
Sphingobacterium
ChloroherpetonPeptococcus
Acetanaerobacterium Bacillus a
Cryptomonadaceae
PapillibacterPolynucleobacter
Prevotella
Haemophilus
ShigellaPolynucleobacter
OthersBurkholderia
Acetanaerobacterium Bacillus cSerratia
Sphingobacterium
ChloroherpetonStreptococcus
Enterococcus
KingellaEscherichiaMethylophilusPrevotellaBacillus a
Ralstonia
CoprococcusPorphyromonasHaliscomenobacterMethylibium
Haemophilus
Pseudomonas
Streptococcus
AnaerostipesPasteuriaceae Incertae sedis
Enterococcus
StaphylococcusPaludibacterOxobacter
Others
Figure 4 Dominant genera (top 15) assigned in the six samples
white-greasy tongue coating subjects behaved clear deviation(Bonferroni-corrected 119875 value all gt05)
36 Metagenome Prediction PICRUSt was used to predicta microbial community metagenome based on 16S genedata PICRUSt results were then analyzed using LEfSe toidentify microbial functions that were significantly differ-ent in their relative abundance among groups The top ofdifferentially abundant bacterial functions were ldquooxidative
phosphorylationrdquo ldquoribosomerdquo ldquoamino sugar and nucleotidesugar metabolismrdquo and ldquosecretion systemrdquo
4 Discussion
In this study we explored a method to detect the microbialdiversity using 16S rRNA gene by high-throughput SOLiDsequencing systemThe ligation-based system features 2-baseencoding which is a proprietarymechanism that interrogates
8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
Peptoniphilus
Ethanoligenens
Trichococcus
Lonepinella
Papillibacter
Azovibrio
Oxobacter
Melissococcus
Faecalibacterium
Cloacibacterium
Desulfitibacter
Marinimicrobium
Pediococcus
Gp4
Gracilibacter
Parabacteroides
Lachnospiraceae Incertae sedis
Aerococcus
Curvibacter
Pseudomonas
Dorea
Gelria
Johnsonella
Alkalispirillum
Dechloromonas
Coprococcus
Syntrophococcus
Weissella
Relative abundance of assigned taxonomyA4A5A6
B2B3B4
Figure 5 Relative abundance of assigned taxonomy at genus level shared in the six tongue coating samples
each base twice The 2-base encoding algorithm filtered rawerrors after sequencing providing built-in error correctionThe output of sequencing reads is in the format of color spacewhich means that the output reads must align to color-spacereference Although the short length is the main drawback ofSOLiD platform compared to other sequencing technologiesour method could extend the effective length of sequencingread to more than 100 bp First the sequencing length wasextended to 51 bp by adding 16 nucleotides digested byEco57I Second concerning that the two strands of libraryDNA could be sequenced in 51015840 to 31015840 direction the usablesequencing length was equivalent to 102 bp which was close
to the length of one hypervariable region of 16S rRNA geneMeanwhile the validation of first four sequenced nucleotideswhich were conserved from the designed primers couldenhance the sequencing accuracy in the validation step
To evaluate the effect of this methodology six tonguecoating samples from different TCM tongue coating typeswere investigated In TCM theories the tongue coatingreflects the status of physiological and clinicopathologicalchanges of inner parts of body As a common type thin-white tongue coating is a symbol of good health A white-greasy tongue coating like powder indicates turbidity andexternal pathogenic heat The abundant bacterial groups
BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
102
8
163
106
104
8
47
44
39
21
20
9
456
36
73
79
71
52
31
19
25
13
15
15
6
202
51
47
20
47
24
16
14
15
11
11
Jeotgalibacillus
Granulicatella
Pelomonas
Lachnobacterium
Haemophilus
Peptostreptococcaceae Incertae sedis
Thioalkalispira
Zoogloea
Anaeromusa
Prevotella
Parasporobacterium
Sporobacterium
Relative abundance of assigned taxonomy
A4A5A6
Figure 6 Relative abundance of assigned taxonomy at genus level shared only in the thin-white tongue coating samples
found in our study are similar to those found in most otherstudies Several studies completing microbial analysis of thehealthy human tongue using 16S rRNA sequencing showedthe most abundant phyla to be Fusobacteria ActinobacteriaFirmicutes Proteobacteria and Bacteroidetes Jiang et alinvestigated 27 tongue coating samples by Illumina technol-ogy and identified 715 differentially abundant species-levelOTUs on tongue coatings of the enrolled patients comparedto healthy controls Furthermore 123 and 258 species-levelOTUs were identified in patients with ColdHot SyndromeIn Jiang et alrsquos report the dominant phyla in Chinese tonguecoating microflora samples were Firmicutes BacteroidetesProteobacteria Actinobacteria and Fusobacteria The dom-inant phyla in our six tongue coating samples were Fir-micutes Proteobacteria Bacteroidetes Actinobacteria and
Chlorobi The similar results represented the effectiveness ofour methodology compared with other sequencing systemsHan et al sequenced the V2ndashV4 region of 16S rRNA geneby pyrosequencing to investigate the tongue coating micro-biome in patients with colorectal cancer and healthy controlsPrevotella Haemophilus and Streptococcus were dominantin Han et alrsquos samples which is the same result in ourstudy Based on these conclusions the method combiningligation-based sequencing and Eco57I digestion exhibitedequivalent effect compared with Illumina or pyrosequencingtechnologies
Using the 16S rRNA gene the core function of tonguecoating microbiome could be predicted by the PICRUStsoftware Pathways encoding for carbohydrate metabolismand oxidative phosphorylation metabolism were detected
10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
Table 5 Relative abundance comparison of significantly different genera between thin-white and white-greasy tongue coating types
Phylum Genus A4 A5 A6 B2 B3 B4 119875
Bacteroidetes Proteiniphilum 85 71 60 0 0 12 0003Bacteroidetes Salinibacter 228 274 328 68 25 13 0004Proteobacteria Thiobacter 166 199 226 0 0 10 0006Firmicutes Cryptanaerobacter 219 320 257 42 16 0 0007Firmicutes Peptococcus 439 295 338 26 16 0 0013Firmicutes Veillonella 2047 2501 1434 383 861 111 0019Bacteroidetes Bacteroides 219 313 188 111 16 26 0019Firmicutes Anaerostipes 0 0 0 100 168 158 0021Firmicutes Sporobacterium 9 15 11 0 0 0 0022Firmicutes Fastidiosipila 11 15 6 31 23 21 0026Bacteroidetes Prevotella 21 13 15 0 0 0 0027Firmicutes Parasporobacterium 20 15 11 0 0 0 0028Proteobacteria Smithella 23 19 22 0 11 0 0028Firmicutes Anaerobacter 0 0 0 31 39 20 0033Proteobacteria Thioalkalispira 47 31 24 0 0 0 0038Firmicutes Trichococcus 127 171 81 4652 5795 9135 0041Firmicutes Faecalibacterium 1504 2974 1936 95 47 232 0042Firmicutes Lachnobacterium 106 79 47 0 0 0 0046Bacteroidetes Subsaxibacter 0 17 0 26 54 65 0047
100
149
31
42
85
7
168
33
39
23
12
11
158
133
20
20
24
7
Anaerostipes
Lactobacillus
Anaerobacter
Ruminococcaceae Incertae sedis
Oribacterium
Acidithiobacillus
Relative abundance of assigned taxonomy
B2B3B4
Figure 7 Relative abundance of assigned taxonomy at genus levelshared only in the white-greasy tongue coating samples
The oral cavity is a major gateway to the human bodyMicroorganisms colonizing in the oral cavity have a sig-nificant probability of spreading to the stomach lung andintestinal tractThemetagenomeprediction validated the roleof tongue coating
In our results except for some common genera in humanbody a few environmental bacteria were observed as well
PCA P1 versus P2
A4
A5
A6
B2B3
B4
minus010 minus005minus015 005 010 015 020000P1 percent variation explained 6472
minus015
minus010
minus005
000
005
010
015
020
P2 p
erce
nt v
aria
tion
expl
aine
d 1
905
Figure 8 Principal component analysis of microbial diversities ofthin-white tongue coating patients (blue squares) and white-greasytongue coating patients (red circles)
One of the explanations is that the sequencing results arenot precise The other hypothesis is that these genera are stillunknown bacteria which have similar sequences with theenvironmental bacteria from databaseThemore bacteria aresequenced the more affirmatory genera could be defined
The composition of the microbial communities on thetongue coating varies between individuals UniFrac principalcoordinates analysis showed no apparent clustering of micro-bial communities between two types of tongue coating Thismay be indicative of the fact that despite the many different
BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 11
habitats on the human tongue many bacterial species areshared among those habitats
Conflicts of Interest
The authors declare that there are no conflicts of interestregarding the publication of this paper
Acknowledgments
The Hi-Tech Research and Development Program of China(Project no 2006AA020702) and the National Natural Sci-ence Foundation of China (Project no 30600152) supportedthis work
References
[1] A F Andersson M Lindberg H Jakobsson F Backhed PNyren and L Engstrand ldquoComparative analysis of human gutmicrobiota by barcoded pyrosequencingrdquo PLoS ONE vol 3 no7 article e2836 2008
[2] E M Bik P B Eckburg S R Gill et al ldquoMolecular analysis ofthe bacterial microbiota in the human stomachrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 3 pp 732ndash737 2006
[3] W Deng D Xi H Mao and M Wanapat ldquoThe use ofmolecular techniques based on ribosomal RNA and DNAfor rumen microbial ecosystem studies a reviewrdquo MolecularBiology Reports vol 35 no 2 pp 265ndash274 2008
[4] N Segata S Kinder Haake P Mannon et al ldquoComposition ofthe adult digestive tract bacterial microbiome based on sevenmouth surfaces tonsils throat and stool samplesrdquo GenomeBiology vol 13 no 6 pp 1ndash18 2012
[5] M H Tschop P Hugenholtz and C L Karp ldquoGetting to thecore of the gut microbiomerdquo Nature Biotechnology vol 27 no4 pp 344ndash346 2009
[6] P J Turnbaugh M Hamady T Yatsunenko et al ldquoA core gutmicrobiome in obese and lean twinsrdquoNature vol 457 no 7228pp 480ndash484 2009
[7] T H M P Consortium ldquoStructure function and diversity ofthe healthy human microbiomerdquo Nature vol 486 no 7402 pp207ndash214 2012
[8] P J Turnbaugh R E Ley M Hamady C M Fraserliggett RKnight and J I Gordon ldquoThe human microbiome projectrdquoIndian Journal of Microbiology vol 52 no 3 pp 804ndash810 2012
[9] J A Aas B J Paster L N Stokes I Olsen and F E DewhirstldquoDefining the normal bacterial flora of the oral cavityrdquo Journalof Clinical Microbiology vol 43 no 11 pp 5721ndash5732 2005
[10] E M Bik C D Long G C Armitage et al ldquoBacterial diversityin the oral cavity of 10 healthy individualsrdquo ISME Journal vol4 no 8 pp 962ndash974 2010
[11] A Bordas R McNab A M Staples J Bowman J KanapkaandM P Bosma ldquoImpact of different tongue cleaning methodson the bacterial load of the tongue dorsumrdquo Archives of OralBiology vol 53 pp S13ndashS18 2008
[12] WCrielaard E Zaura A A Schuller SMHuse R CMontijnand B J F Keijser ldquoExploring the oral microbiota of children atvarious developmental stages of their dentition in the relationto their oral healthrdquo BMC Medical Genomics vol 4 article 222011
[13] V Lazarevic K Whiteson D Hernandez P Francois and JSchrenzel ldquoStudy of inter- and intra-individual variations in thesalivary microbiotardquo BMC Genomics vol 11 no 523 2010
[14] V Lazarevic KWhiteson S Huse et al ldquoMetagenomic study ofthe oral microbiota by Illumina high-throughput sequencingrdquoJournal of Microbiological Methods vol 79 no 3 pp 266ndash2712009
[15] S Pushalkar S P Mane X Ji et al ldquoMicrobial diversity insaliva of oral squamous cell carcinomardquo FEMS Immunology andMedical Microbiology vol 61 no 3 pp 269ndash277 2011
[16] R Watanabe K Hojo S Nagaoka K Kimura T Ohshima andN Maeda ldquoAntibacterial activity of sodium citrate against oralbacteria isolated from human tongue dorsumrdquo Journal of OralBiosciences vol 53 no 1 pp 87ndash92 2011
[17] E Zaura B J Keijser S M Huse and W Crielaard ldquoDefiningthe healthy core microbiome of oral microbial communitiesrdquoBMCMicrobiology vol 9 no 259 2009
[18] M T Abreu and R M Peek Jr ldquoGastrointestinal malignancyand themicrobiomerdquoGastroenterology vol 146 no 6 pp 1534ndash1546 2014
[19] F E Dewhirst T Chen J Izard et al ldquoThe human oralmicrobiomerdquo Journal of Bacteriology vol 192 no 19 pp 5002ndash5017 2010
[20] H E Jakobsson C Jernberg A F Andersson M Sjolund-Karlsson J K Jansson and L Engstrand ldquoShort-term antibiotictreatment has differing long-term impacts on the human throatand gutmicrobiomerdquo PLoSOne vol 5 no 3 article e9836 2010
[21] C E Kazor P M Mitchell A M Lee et al ldquoDiversity ofbacterial populations on the tongue dorsa of patients withhalitosis and healthy patientsrdquo Journal of Clinical Microbiologyvol 41 no 2 pp 558ndash563 2003
[22] R Li T Ma G Jin X Liang and S Li ldquoImbalanced networkbiomarkers for traditional Chinese medicine Syndrome ingastritis patientsrdquo Scientific Reports vol 3 no 3 pp 1543-15432013
[23] B J Marsland and E S Gollwitzer ldquoHost-microorganisminteractions in lung diseasesrdquo Nature Reviews Immunology vol14 no 12 pp 827ndash835 2014
[24] P Xu and J Gunsolley ldquoApplication of metagenomics in under-standing oral health and diseaserdquo Virulence vol 5 no 3 pp424ndash432 2014
[25] J L Tang B Y Liu and K W Ma ldquoTraditional Chinesemedicinerdquo Lancet vol 372 no 9654 pp 1938ndash1940 2008
[26] S Han Y Chen J Hu and Z Ji ldquoTongue images and tonguecoating microbiome in patients with colorectal cancerrdquo Micro-bial Pathogenesis vol 77 pp 1ndash6 2014
[27] B Jiang X Liang Y Chen et al ldquoIntegrating next-generationsequencing and traditional tongue diagnosis to determinetongue coating microbiomerdquo Scientific Reports vol 2 article936 2012
[28] Z Sun J Zhao P Qian et al ldquoMetabolicmarkers andmicroeco-logical characteristics of tongue coating in patients with chronicgastritisrdquo BMC Complementary and Alternative Medicine vol13 article 227 2013
[29] Y Zhao X Gou J Dai et al ldquoDifferences in metabolites ofdifferent tongue coatings in patients with chronic hepatitis BrdquoEvidence-Based Complementary and Alternative Medicine vol2013 Article ID 204908 12 pages 2013
[30] D R Bentley S Balasubramanian H P Swerdlow et alldquoAccurate whole human genome sequencing using reversibleterminator chemistryrdquo Nature vol 7218 pp 53ndash59 456
12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 BioMed Research International
[31] T D Harris P R Buzby H Babcock et al ldquoSingle-moleculeDNA sequencing of a viral genomerdquo Science vol 320 no 5872pp 106ndash109 2008
[32] M EHudson ldquoSequencing breakthroughs for genomic ecologyand evolutionary biologyrdquo Molecular Ecology Resources vol 8no 1 pp 3ndash17 2008
[33] M Margulies M Egholm W E Altman S Attiya J S Baderand L A Bemben ldquoGenome sequencing in microfabricatedhigh-density picolitre reactorsrdquo Nature vol 437 no 7057 pp376ndash380 2005
[34] A Valouev J Ichikawa T Tonthat et al ldquoA high-resolutionnucleosome position map of C elegans reveals a lack of uni-versal sequence-dictated positioningrdquoGenome Research vol 18no 7 pp 1051ndash1063 2008
[35] M J Claesson S Cusack O OrsquoSullivan et al ldquoCompositionvariability and temporal stability of the intestinal microbiota ofthe elderlyrdquo Proceedings of the National Academy of Sciences ofthe United States of America vol 108 no 1 pp 4586ndash4591 2011
[36] T Ding and P D Schloss ldquoDynamics and associations ofmicrobial community types across the human bodyrdquo Naturevol 509 no 7500 pp 357ndash360 2014
[37] J A Huber D B MarkWelch H G Morrison et al ldquoMicrobialpopulation structures in the deep marine biosphererdquo Sciencevol 318 no 5847 pp 97ndash100 2007
[38] CHuttenhower DGevers R Knight et al ldquoStructure functionand diversity of the healthy human microbiomerdquo Nature vol486 no 7402 pp 207ndash214 2011
[39] J J Qin Y R Li Z M Cai et al ldquoA metagenome-wideassociation study of gut microbiota in type 2 diabetesrdquo Naturevol 490 no 7418 pp 55ndash60 2012
[40] J Ye X Cai and P Cao ldquoProblems and prospects of cur-rent studies on the microecology of tongue coatingrdquo ChineseMedicine vol 9 no 1 article 9 2014
Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpswwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 201
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology