evaluating stem cell response to a spider silk scaffold
TRANSCRIPT

Clemson UniversityTigerPrints
All Theses Theses
5-2017
Evaluating Stem Cell Response to a Spider SilkScaffoldKatherine Lee HafnerClemson University, [email protected]
Follow this and additional works at: https://tigerprints.clemson.edu/all_theses
This Thesis is brought to you for free and open access by the Theses at TigerPrints. It has been accepted for inclusion in All Theses by an authorizedadministrator of TigerPrints. For more information, please contact [email protected].
Recommended CitationHafner, Katherine Lee, "Evaluating Stem Cell Response to a Spider Silk Scaffold" (2017). All Theses. 2630.https://tigerprints.clemson.edu/all_theses/2630

EVALUATING STEM CELL RESPONSE TO A SPIDER SILK SCAFFOLD
_______________________________________________
A Thesis Presented to
the Graduate School of Clemson University
_______________________________________________
In Partial Fulfillment of the Requirements for the Degree
Master of Science Bioengineering
_______________________________________________
by Katherine Lee Hafner
May 2017 ________________________________________________
Accepted by: Dr. Delphine Dean, Committee Chair
Dr. Marian Kennedy, Co-Committee Chair Dr. Melinda Harman

ii
ABSTRACT Micropatterning on a surface using fibers, channels, and troughs, can act as an
effective means of inducing cell attachment and alignment. These morphological and
pattern changes as a response to physical cues can impact the potential that a cell has to
differentiate into a different cell line. This thesis evaluated the response of human dental
pulp stem cells (DPSCs), and other cell types, to spider dragline silk fibers, a potential
scaffold material for tissue regeneration, and further observed the effects of morphology,
orientation, and composition of silk on the adherence of cells.
Several cell lines were studied in this thesis, including adipose derived stem cells
(ADSCs), osteoblasts (7F2s), and fibroblasts (3T3s), but DPSCs were the main cell type
of interest. This is due to the fact that DPSCs are a proposed source of stem cells for nerve
regeneration based on their close embryonic origin to neurons and the ease with which
DPSCs can be obtained from a donor. The cells’ morphologies and spread patterns were
characterized after they were plated onto Nephila clavipes dragline fibers in media. The
inclusion of 3T3s and 7F2s in this study allowed for both direct comparisons to prior
published work and a qualitative comparison to the morphology of the DPSCs. After
twelve days, the DPSCs exhibited greater relative alignment and adherence to the spider
dragline fibers than the 3T3s and 7F2s when silk was wrapped in an aligned orientation
rather than a random orientation.
The impact of a common sterilization method (ultraviolet light) on the spider
dragline fiber surface and subsequent cell response to this modified surface was also
characterized. Exposure of the silk to ultraviolet light did not have a measureable effect on
cell alignment, but it did eliminate bacterial growth and changed fiber surface roughness.

iii
Spiders’ exposure to stressful environments did not have an effect on silk to impair cell
alignment or adhesion, and synthetic recombinant protein silk fibers did not act as a
scaffold for cell adhesion or alignment. However, cells remained viable and proliferative
in recombinant silk hydrogels, suggesting that surface characteristics and large diameter
had a negative impact on cell interaction on ‘synthetic’ silk fibers. In all cases, natural
drawn spider silk acted as an effective scaffold for cellular adhesion and alignment of
DPSCs and could be used in neural differentiation applications. Collectively, this thesis
indicates that spider silk from spiders under any extent of stress can be rendered sterile by
UV radiation and act as an effective means of cellular adhesion and alignment when silk is
organized in an aligned orientation.

iv
DEDICATION
This work is dedicated to my family and friends. They have stood by me through
the highs and lows, and I would not have been able to complete this without their help.
Also, special recognition is due to my parents, Greg and Mel, for their constant support
and patience when listening to my research grievances. Without them and their words of
encouragement, I would not have had the drive for knowledge and motivation to achieve
the goals I set my mind to.

v
ACKNOWLEDGEMENTS
I would like to thank my chair advisors, Dr. Delphine Dean and Dr. Molly Kennedy,
who have always gave me positive reinforcement no matter the situation. In my 10
semesters at Clemson, I have spent 9 of them working in a lab for Dr. Dean and Dr.
Kennedy and I cannot imagine having two greater people to model myself after. I would
like to also thank my other committee member, Dr. Melinda Harman, who acted as my
advisor in my undergraduate years and has helped to encourage my understanding and
experience in the more industrial side of bioengineering where I would like to end up.
I also gratefully acknowledge experimental help and insight from others at Clemson
University including the wonderful individuals in my research teams with Dr. Dean’s
lab and Dr. Kennedy’s lab, especially Hannah Maeser, John Catoe, and Olivia Ross.
Along with my Clemson family, I would like to thank Dallas Montag, our summer REU
student from Marietta College. Without all of your assistance, encouragement, and
expertise, I would not have accomplished my project.

vi
TABLE OF CONTENTS
Page
TITLE PAGE ....................................................................................................................... i
ABSTRACT ........................................................................................................................ ii
DEDICATION ................................................................................................................... iv
ACKNOWLEDGEMENTS ................................................................................................ v
LIST OF TABLES ........................................................................................................... viii
LIST OF FIGURES ........................................................................................................... ix
CHAPTER ONE - INTRODUCTION................................................................................ 1
1.1 MOTIVATION ................................................................................................. 1
1.2 RESEARCH AIMS ........................................................................................... 2
1.3 SIGNIFICANCE ............................................................................................... 3
CHAPTER TWO - PROLIFERATION OF STEM CELLS AND EFFECT OF CELL
TYPE ON ADHERANCE TO SPIDER SILK TREATED WITH ULTRAVIOLET
RADIATION ...................................................................................................................... 4
2.1 INTRODUCTION ............................................................................................ 4
2.2 SPIDER SILK OVERVIEW............................................................................. 4
2.3 CELL TYPES - DENTAL PULP STEM CELLS, ADIPOSE-DERIVED
STEM CELLS, OSTEOBLASTS, AND FIBROBLASTS .................................... 7
2.4 MATERIALS AND METHODS .................................................................... 11
2.5 EFFECT OF SLIDE PREPARATION METHOD AND RESPONSE OF
CELLS TO ULTRAVIOLET RADIATION ........................................................ 17
2.6 RESPONSE OF CELL TYPES TO SILK ...................................................... 20
2.7 CONCLUSIONS............................................................................................. 22

vii
CHAPTER THREE - CELLULAR RESPONSE TO SILK REELED UNDER
ENVIRONMENTAL STRESS AND RESPONSE TO SILK PROPERTIES AND
ORIENTATION ............................................................................................................... 25
3.1 INTRODUCTION .......................................................................................... 25
3.2 EFFECTS ENVIRONMENTAL STRESSES ON SPIDER SILK AND
RESPECTIVE EFFECTS OF MORPHOLOGY AND ORIENTATION ON
CELL ADHESION TO SILK ............................................................................... 25
3.3 MATERIALS AND METHODS .................................................................... 30
3.4 EFFECTS OF SILK ORIENTATION ON CELLS ........................................ 34
3.5 RESPONSE OF CELLS OF DIFFERENT PROPERTIES AND
DIAMETERS ........................................................................................................ 35
3.6 CONCLUSIONS............................................................................................. 38
CHAPTER FOUR – FUTURE WORK WITH NEUROGENIC REGENERATION AND
RECOMMENDATIONS .................................................................................................. 41
5.1 BACKGROUND FOR NEUROGENIC REGENERATION ......................... 41
5.2 RECOMMENDATIONS ................................................................................ 42
APPENDICES .................................................................................................................. 44
Appendix A MODELING CELL ROUNDNESS IN IMAGEJ ........................................ 45
Appendix B CHARACTERIZING REELING MECHANISM ....................................... 48
REFERENCES ................................................................................................................. 50

viii
LIST OF TABLES
Page
Table 1: Comparison of silk types, the gland and spinneret they are derived from, and
their function and composition [20]. ................................................................................... 5
Table 2: Roughness values (Root-Mean-Square, or RMS) of dragline silk exposed to UV
irradiation for 15 or 60 minutes compared to a control sample. ....................................... 15
Table 3: Growth media solutions in which cells and silks were plated. ........................... 16
Table 4: Average roundness varied by the three cell types (osteoblasts, fibroblasts and
DPSCs). These were calculated using the ImageJ roundness analysis. ........................... 21
Table 5: Matrix showing variation in the spider enclosures/environmental conditions
prior to silking and the population naming systems referenced within this thesis. .......... 31

ix
LIST OF FIGURES
Page
Figure 1: Diagram representing the types of silk that make up a web [20]. ....................... 5
Figure 2: Diagram of the secondary structure of dragline silk. A) Crystalline region, B)
Oriented amorphous region, C) Amorphous region [20]. ................................................... 6
Figure 3: The anatomical location for the derivation of DPSCs. ........................................ 8
Figure 4: Multilineage differentiation potential for ADSCs [40]. ...................................... 9
Figure 5: Morphology of fibroblasts (3T3s) when in two-dimensional culture. Note
spread body and lammellipodia. ....................................................................................... 10
Figure 6: Wrapping of silk around cylindrical arrangement of nails on spool. ................ 12
Figure 7: (a) shows spider silk reeling set-up with spider pinned. (b) aligned silk on glass
slide. .................................................................................................................................. 13
Figure 9: Release of gas 'bubbles' from spider silk on elastomer coated scaffold trial. This
image was taken 3 days after plating DPSCs and no cells were observed near or upon the
spider silks. ....................................................................................................................... 18
Figure 8: Development of contamination upon hand-wrapped, elastomer coated spider
silk slides. Top images show bacterial contamination. Bottom images show fungal
contamination. ................................................................................................................... 18
Figure 10: Images show DPSCs forming a general trend of adhesion to the spider silk,
proliferation, and an eventual overall alignment of the cells to the silk line over time.
Images were taken by light microscopy at 10X magnification. ........................................ 18
Figure 11: Fluoroscopy images taken after two weeks of growth show alignment and
adhesion of cells to the spider silk in comparison with exposure time to ultraviolet
radiation. The red stain (phalloidin) shows the f-actin cytoskeletal fibers, the blue DAPI

x
stain indicates the nuclei and shows the number of cells, and the green autofluorescence
indicates the silk. The 0 min UV light trial for osteoblasts experienced bacterial
contamination, resulted in very limited cell growth on and around the silk. DPSCs (top
row) showed the best alignment in individual cells and in cell populations as a whole.
Osteoblasts (middle row) had some good individual cell adherence and alignment, but did
not show bulk population alignment. Fibroblasts were mostly observed clumping around
the silk and on the scaffolds in general with very little organized individual or population
alignment........................................................................................................................... 19
Figure 12: ImageJ analysis to create threshold microscopy image to determine cell
roundness. *all scale bars same, 400 μm * ....................................................................... 21
Figure 13: Cell roundness distribution from ImageJ roundness analysis. The ‘*’ indicates
a statistical significance (p<0.05) between one group, ** indicates significance between
two groups. ........................................................................................................................ 22
Figure 14: Diagram of spider spinneret and valve [20]. ................................................... 26
Figure 15: Effect of surface energy (hydrophilicity) on contact angle. ............................ 28
Figure 16: Cell response on different diameter silks - A) Cross sectional view showing
spreading and wrapping, B) Topographical view showing spreading and wrapping. ...... 29
Figure 17: Transformation of spider silk genetic information into recombinant spider silk
proteins using plant-based transformation. ....................................................................... 30
Figure 18: Random orientation of silks when handwrapped. Images were taken 3 days
after plating cells and no cells were present near or on silks. ........................................... 34

xi
Figure 19: Effects of handwrapping silks and creation of multiple planes of view. These
images were taken at day 3 after plating and cells only appear adhered to the glass slide
in the bottom plane of view rather than upon the spider silks. ......................................... 34
Figure 20: Motor wrapped silk slides show better initial adherence of ADSCs under
optical microscopy after 5 days than randomly oriented handwrapped silks. .................. 35
Figure 21: Fluorescent microscopy depicts DPSCs after 2 weeks of growth on spider silk
drawn from different environments – A&B = ‘control spider population’, C = ‘boxed
spider population’ (Limited Space), D = ‘limited light exposure population’. ................ 36
Figure 22: Fluorescent microscopy depicts DPSCs after 2 weeks of growth on spider silk
of different materials – Left = Naturally draw dragline spider silk (control) and Right =
Hand drawn recombinant spider protein silk (synthetic). There is no cell adhesion or
alignment on the synthetic silk. ........................................................................................ 37
Figure 23: DPSCs on hydrogels of different spidroin compositions. sp 1 is made from
Spidroin 1 mimics, sp 2 from Spidroin 2 mimics, and sp 1/2 from Spidroin 1 and Spidroin
2 mimics in a 70:30 v/v ratio. All mimics were combined with chitosan and gellan gum.
........................................................................................................................................... 38
Figure 24: Physical cues and the induction of differentiation based on surface micro-
pattern [97]. ....................................................................................................................... 41
Figure 25: Typical examples of electrospinning methods. Traditional electrospinning
method to prepare nonwoven fabric scaffolds (a), the electrospinning method to prepare
cotton ball-like scaffolds (b), the electrospinning method to prepare double-layered 2-D
architectures (crosshatch pattern) using a 3-D stage (c), and the Nanospider
electrospinning method (d). [51]....................................................................................... 43

1
CHAPTER ONE - INTRODUCTION
1.1 MOTIVATION
Natural materials are often considered a golden standard in regards to biomedical
scaffold due to their general biocompatibility in the body. An advantage of natural
materials compared to synthetic materials for implants is that they are similar to materials,
and are familiar to the body systems. In addition, these natural materials generally do not
experience the issues of toxicity like their synthetic counterparts. Also, in some cases they
contain specific protein binding sites and other biochemical signals that influence the way
that cells interact with the material to help integration of tissue into a scaffold for healing
[1]. There is often a challenge in creating a more widely available synthetic material to
mimic natural materials and material properties generally suffer. However, there is a drive
to use natural materials, or synthetic materials mimicking their natural models, in medicine
as implants and scaffolds. For instance, use of fibers with the proper surface morphology
and chemistry can promote cell attachment and alignment that can influence differentiation
later on. Recent studies have examined silkworm silk as a potential option for a natural
fiber material, but spider silk is rising as a key player in the natural material fiber field due
to its high strength and biocompatibility. If a relationship between cell adherence and
alignment can be drawn to spider silk, and a synthetic version of silk can be created, there
is a greater ability to integrate this natural biomaterial into modern medicine.
The purpose of this study was to evaluate and analyze the way that different cell
types adhered and aligned to a spider silk scaffold to determine if their changes in
morphology could act as a potential influence for differentiation. After determining that

2
spider silk, sterilized by ultraviolet radiation, could act as a two-dimensional scaffold to
improve aligned orientation and elongated morphology more variable were introduced to
silk types. Within this evaluation, the study also monitored cellular responses to silks
treated with different exposures to ultraviolet light, spider silk reeled in different stressful
environments, and on recombinant silk scaffolds.
1.2 RESEARCH AIMS
Aim 1: Evaluate adhesion and alignment of cells on a spider silk scaffold treated with an
appropriate sterilization method
Spider silk was reeled and exposed to ultraviolet (UV) light as a means of
sterilization to prevent contamination. In a pilot study, silk slides were exposed to varying
amounts of UV radiation to determine if there were any negative effects of radiation on
silk or the cells plated on it. Silk slides were then subjected to a standard sterilization cycle
of UV radiation based on pilot study results, and varying cell types were plated in order to
determine the adhesion and alignment trends of each cell type.
Aim 2: Evaluate response of cells to silks reeled under stress, recombinant silk treads, and
recombinant silk hydrogels in comparison to normal condition natural silk
After successfully determining a reeling mechanism, sterilization method, and
suitable cell line for adhesion and alignment, variation to silk was creating.
Environmental stress was put onto spiders to observe changes in silk that may impact its
interaction with cells. Recombinant spider proteins, grown in tobacco leaves, were also
used to make recombinant silk hydrogels and fibers to evaluate response of cells on both

3
materials. This was intended to compare response of cells on natural silk versus
‘synthetic’ recombinant silk.
1.3 SIGNIFICANCE
This study describes the steps taken to develop an effective reeling and sterilization
method of natural silk to use for cell response evaluation. Providing a scaffold for cells to
align to and elongate against creates the proper physical cues and stimuli to help influence
neurogenic differentiation. Traumatic wounds to the central and peripheral nervous system
are difficult to treat in most part due to the inability of neurons to naturally replication, so
creating a scaffold that promotes differentiation of easily attainable mesenchymal stem
cells stands as an option for an implant to treat these type of wounds. This study also
compared the response of cells to natural silk versus silk reeled from spiders under stress
and synthetic silk. The development of a synthetic silk that mimics the properties of natural
silk allows for easier fabrication of scaffolds with different morphologies and sizes. This
variability in structure that is possible in synthetic materials can improve the way that an
implant can integrate with a body and act as a better biomaterial.

4
CHAPTER TWO - PROLIFERATION OF STEM CELLS AND EFFECT OF CELL TYPE ON ADHERANCE TO SPIDER SILK TREATED WITH
ULTRAVIOLET RADIATION 2.1 INTRODUCTION
Cells have been shown to respond to surface and topographical cues by changing
their morphology. These morphological changes lead to alterations in the ways that
biomechanical responses are transmitted and can thus influence the way that stem cells
differentiate [2], [3]. Mesenchymal stem cells (MSCs) have been shown to differentiate
into distinctive lineages. A subset of MSCs includes dental pulp stem cells (DPSCs), which
can differentiate into several lineages including neural, odontoblastic, and osteoblastic
lineages [4]–[6]. This study will look at the influence that protein based silk fibers have on
the phenotype of DPSCs and other cell types in culture.
2.2 SPIDER SILK OVERVIEW
Silk fibers are produced by silkworms and spiders for a variety of functions
including catching prey, structural support for webs, and protection from natural harsh
conditions like rain and direct sunlight [[7],[8]]. The spider produced silk fibers are
considered to be bioinert and exhibit both relatively high specific strength and toughness
[9]. Many research groups have been interested in leveraging this unique combination of
properties as scaffolds for cellular growth [[10]–[13]]. Previous studies have shown how
silks from both worms and spiders can influence cellular growth and attachments of some
types of stem cells [14]–[17]. In this work, we will focus on dragline silks produced by
spiders since these fibers are free of an immunogenic sericin coat (such as those present on
native silkworm silks that have not been degummed [17]) and are shown to be
biocompatible. Prior studies have shown that dragline fibers cause little immunogenic

5
response when implanted subcutaneously in pigs [[18], [19]]. These silks are also stable
at high temperatures up to 230 °C and, while they are insoluble in most solvents, they
eventually degrade into bioinert components, suggesting they would survive well in the
body [[20]–[23]]. Other studies have suggested that silk proteins can promote cell
attachment and growth [[17], [24], [25]].
Each spider has multiple glands and spinnerets that can produce different types of
silk for a range of functions. Several research groups have explored the range of mechanical
properties between the types of spider silk [11]. The most commonly studied spider silk,
Figure 1: Diagram representing the types of silk that make up a web [20].
Table 1: Comparison of silk types, the gland and spinneret they are derived from, and their function and composition [20].

6
dragline, is produced by the major ampullate gland and can function as the orb web frame,
radii, and safety line for orb weaver spiders [26]–[28]. These fibers have been shown to
have high specific tensile strength, comparable to that of Kevlar, and high resistance to
breakage compared to other fibers [9], [11], [27]. Besides proving to exist as some of the
strongest organic fibers known, spider silks can also experience over 15% elongation
before rupture, making them suitable for a dynamic environment [29]. The mechanical
properties of dragline fibers are attributed to the nano- and microstructure within the fiber,
namely repeated iteration of alanine- and glycine-rich regions as shown in Table 1 [[30]–
[32]]. Arg-Gly-Asp sequences have been indicated to act as biological recognition signals
that can promote cell adhesion, and the high percentages of glycine and polar amino acids
present in silk make spider silk a candidate for good cell attachment [33]. These Arg-Gly-
Asp regions stack together to form rigid crystals, so silk consists of a semi-crystalline
polymeric structure with both crystallite and amorphous regions as shown in Figure 2 [34]–
[37].
Spider species can have an influence on silk properties often due to the differences
in the uses of the webs. For instance, families like Araneus, Nephila include orb-weaving
spiders (constructing the traditionally ideal two dimensional spiral spider web), while other
Figure 2: Diagram of the secondary structure of dragline silk. A) Crystalline region, B) Oriented amorphous region, C) Amorphous region [20].

7
families of spiders will construct sheet webs or three dimensional tangle webs [23]. This
difference in structure impacts the performance and strengths of the webs. In three
dimensional webs, silks can be weaker because a moving insect is mostly stopped because
the energy required to stop it is dissipated by breaking through layers of weak strands. In
a two dimensional orb web, an insect is stopped when it is caught in the strong and stretchy
spiral threads – if not for their strength and elasticity, the threads would break and the
insects would not be captured [38]. Oftentimes, multiple strands of the strong dragline
silks are laid in the orb webs to withstand the high force of impact experienced by moving
insects.
2.3 CELL TYPES - DENTAL PULP STEM CELLS, ADIPOSE-DERIVED STEM
CELLS, OSTEOBLASTS, AND FIBROBLASTS
Dental Pulp Stem Cells
In order for a stem cell to be capable of use in regenerative medical applications, it
should meet several criteria: it can i) be found in abundant quantities, ii) be collected or
harvested by a minimally invasive procedure, iii) be differentiated into multiple cells lines
in a reproducible way, and iv) be safely and effectively transferred to a host [39]. Dental
pulp stem cells are a viable form of MSC that can be easily derived from dental pulp
extracted from teeth of animals and humans. At this point, MSCS are considered one of
the most widely available autologous source of stem cells to be used for practical and
clinical applications, and currently there have no reports that MSCs have a tendency of
differentiating into tumors the way that more ethically questionable embryonic stem cells
(ESCs) or induced pluripotent stem cells (iPSCs) do [40]. When DPSCs are not stimulated
by topographical or chemical cues, they normally generate a dentin-pulp-like tissue and

8
were capable of forming a mineralized matrix [5]. However, under the proper
circumstances of altered structural and mechanical properties, DPSCs can be induced into
other lines of differentiation than odontoblastic [41]. Microtechnology can allow for the
organization and manipulation of DPSCs in their relative scale to enable control over their
functions and phenotypes [42]. Normal DPSC morphology take on a long (>50um) and
narrow (<20um) spindle shape that elongates upon surfaces and this is expected to be the
case when plated onto spider silk.
DPSCs are known to elongate when plated on a flat surface and have been shown
to respond to substrate micropatterning on the culture surface [43]. Micropatterning of
surfaces onto which cells in media are plated, in addition to other culture conditions, can
initiate differentiation of DPSCs. Using cell aligning methods, such as collagen gel
tethering to initiate tension-induced alignment and adhesive collagen troughs in the
substrates, can help stabilize the generation of neural tissue [44]–[47]. Similarly to these
Figure 3: The anatomical location for the derivation of DPSCs.

9
surface changes, fibers can also be used as scaffolds that cells can adhere to. For instance,
due to their unique structure and properties, silk fibers have been shown to guide cell
phenotype and differentiation [14], [15]. The potential of DPSCs to differentiate into
neural cells is the reason that they are given special attention in this thesis.
Adipose Derived Stem Cells
Adipose-derived stem cells (ADSCs) are another type of adult mesenchymal stem
cell. Like DPSCs, ADSCs have high proliferation abilities, can maintain their phenotypes
over a long time of passaging, have variable differentiation pathway potential, and continue
to have a strong differential potential even after 25 passages [48]. ADSCs are
mesodermally derived and contain supportive stroma that can easily be isolated by a non-
invasive, normal procedure. Studies have shown that nearly 5x105 stem cells can be
extracted from 400-600mg of adipose tissue, and considering there are nearly 400,000
liposuctions performed in the US that yield from 100mL to >3L of adipose tissue, this can
serve as an adequate source for stem cell isolation [48]–[51]. The morphology of ADSCs
Figure 4: Multilineage differentiation potential for ADSCs [40].

10
in their natural state (unaltered by chemical or micropatterning cues) is one of an elongated
spindle shape. Also similar to DPSCs, ADSCs show low immunogenicity and have the
potential to develop into neurogenic lineages based on chemical and topographical cues
[52], [53] and thus can be expected to behave similarly.
Fibroblasts
Fibroblasts (3T3s) are a type of mesenchymally derived cell that is present in
connective tissue. Fibroblasts are responsible for synthesizing extracellular matrix (ECM)
and play a critical role in wound healing [54]. Fibroblasts are derived from human
connective tissues and can be harvested in mostly non-invasive ways, like from skin during
cosmetic surgeries or from joint tissue during arthroscopic surgeries [55], [56]. In two-
dimensional culture, fibroblasts will normally present a flat morphology with
lammellipodia (as seen in Figure 5) and they will migrate and proliferate without many
borders, and when in a three-dimensional culture, they will present a spindle-shaped
morphology [57]–[59]. When fibroblasts can align and exhibit this spindle-like shape and
Figure 5: Morphology of fibroblasts (3T3s) when in two-dimensional culture. Note spread body and lammellipodia.

11
align, they present a greater potential for wound healing of connective tissue by the
synthesis of a fresh ECM which can stand as a potential source for repairing injuries like
torn anterior cruciate ligaments [56]. A stiff surface is required to encourage ECM
remodeling for ligament repair, and linearly oriented fibers can act as a means of
encouraging fibroblasts to achieve their spindle-shaped morphology [56].
Osteoblasts
Osteoblasts (7F2s), are the final type of cell used in this cell type comparison study.
Like the other cells in this study, osteoblasts are derived from MSCs. Unlike the mostly
non-invasive methods of harvesting DPSCs, ADSCs, and 3T3s, osteoblasts are harvested
from cortical bone samples or else must be differentiated from MSCs. Micropatterning on
a surface can help with the differentiation of MSCs toward osteoblasts, and can also
promote bone synthesis when osteoblasts exist [60]. Osteoblasts are a component of bone
and are responsible for secreting bone matrix necessary to form bones. When bone is
forming, the crystalized minerals tend to orient themselves with their axis parallel to one
another [61]–[63], so creating an oriented matrix should help provide for an improvement
in the formation of bone crystals. Another key element to bone synthesis is hydroxyapatite,
a key mineral in bone matrix, will not be used to promote bone growth in this study, and
rather osteoblasts will be used a cell type to compare behavior and morphology trends of
DPSCs, ADSCs, and 3T3s to since it is not expected to vastly change in morphology.
2.4 MATERIALS AND METHODS
Pre-Drawn Silk on Elastomer Coated Slides

12
Initially, spider silks from Nephila clavipes spiders that had previously been drawn
onto metal spools were used for preparing scaffolds for cell plating. Silk spools had been
prepared by a previous research group and existed as a series of threads spread out over a
cylindrical arrangement of nails so that there was open space to collect silk threads as
demonstrated in Figure 6. 20mm x 20mm cover slip slides were prepared for silk wrapping
by undergoing a generic ethanol rinse and sitting under ultraviolet light for 30 minutes.
The slides were then coated in the 184 Silicone Elastomer Kit (Sylgard) in order to help
silk adhere and stick to the surface. Silk was manually collected from the spool with
forceps and wrapped around 20mm x 20mm glass slide, then left for 30 minutes to allow
the elastomer to cure and harden. Porcine DPSCs that had previously been harvested from
pig molars were thawed and cultured in a T75 flask to prepare them for plating onto the
wrapped silk scaffold. After the cells reached confluence in the T75 flask, they were
Figure 6: Wrapping of silk around cylindrical arrangement of nails on spool.

13
trypsinized and passaged to be seeded onto the wrapped silk scaffolds at a density of 25,000
cells per 20mm x 20mm slide. Cells were maintained in a standard media (Table 2) and
allowed to culture for 5 days. Due to the presence of bacterial and/or fungal growth on all
specimens, the plating method was re-evaluated to include a step for sterilization of the
spider silk, this will be further discussed later in this chapter. Exposure to ultraviolet (UV)
radiation was selected a means of sterilization of silk prior to plating and trials with fresh
silk from live spiders began.
Drawing Silk by Reeling Mechanism with Live Spiders
Dragline silk was collected from five Nephila clavipes spiders (commonly referred
to as golden silk orb-weavers [64]) gathered within the upstate of South Carolina. Once the
spiders were brought into the laboratory, they were kept within either a large, communal 9
ft by 10 ft by 7 ft camping tent or individual isolated boxes. The spiders were fed pinhead
crickets by placing the crickets within their webs and watered by misting each web with a
spray bottle. Due to the difficulty of hand-wrapping slides with silks and the poor results
that these slides rendered, an alternative silking method was developed that used motors to
Figure 7: (a) shows spider silk reeling set-up with spider pinned. (b) aligned silk on glass slide.

14
rotate a slide at a steady rate to allow for stability and alignment of silk on the slides. Prior
to silking, a dilute 70% v/v ethanol solution was used to clean the setup area, tools, and
containers for the slides. The silking process was continued by placing the spider into a 4
°C refrigerator for 45-60 min for sedation. Once a spider was removed from the
refrigerator, it was positioned onto its back on a piece of foam and then secured between
the foam and a Kimwipe® placed over the spider using pins (Figure 7(a) - left). The tissue
was ripped slightly to expose the spinnerets and the secured spider placed next to the motor
setup. The area around each of the spinnerets was opened with tweezers, and a single silk
thread was pulled and attached to the edge of a sterilized coverslip (20 mm side length)
with adhesive before starting the motors. Forced silking of spiders under conditions that
do not use an anesthetic condition is shown to result in properties that are consistent with
those of normally spun silk, so the reeling mechanism specified should have no negative
effects of silk properties [65]. Silk was collected at 10 mm/s (1cm/s) from each spider to
mimic natural spinning speed [66]. The rate of collection was controlled using winding
devices that pull the silk from the spider’s spinneret. Two STM17Q-2AN stepper motors
were used to run the silk collection system. Machined aluminum parts were designed so
one motor could hold and turn a coverslip for silk collection while the other motor turned
the handle for the microscope displacement stage on which the first motor was mounted.
Control of the motor speeds allowed for even spacing of silk on the coverslips for cell
plating. Cells were allowed to culture for 5+ days and images were taken by optical
microscopy.
Sterilization of Spider Silk

15
Since the silk is derived from a non-sterile source it is imperative to treat the
surfaces to discourage bacterial and fungal growth. After examining multiple forms of
sterilization, 254 nm ultraviolet (UV) light was chosen because, unlike autoclaving or
ethylene oxide, short duration UV light was found to cause minimal damage to the silk
structure [67]. Silk wrapped slides were exposed to 0, 5, 15, and 60 min of UV light. Non-
contact optical profilometry was used to check changing in surface morphology (Table 2)
and denaturation of the silk before cell plating. Cells were plated on these slides and
allowed to proliferate for two weeks. Samples were stained and imaged to assess
differences in the phenotype of cells grown on the silks exposed to different amounts of
UV light exposure.
Cell Type Effect on Alignment
Three different cell types were chosen for culture; human DPSCs (Lonza Group,
Basel, Switzerland), mouse fibroblasts (3T3s) (NIH/3T3, ATCC® CRL-1658TM) and
mouse osteoblasts (7F2s) (7F2, ATCC® CRL-12557TM). The DPSCs were collected from
adult 3rd molars and are positive for CD105, CD166, CD29, CD90, and CD73 and do not
express CD34, CD45, and CD133 as per the manufacturer (Lonza). The DPSCs,
fibroblasts, and osteoblasts were initially cultured in T75 flasks. Each cell line was cultured
in a growth media solution conducive for the growth of that type of cell (see Table 3). Each
cell type was then plated individually at a density of 10,000 cells per 9.5 cm2 into a six well
Table 2: Roughness values (Root-Mean-Square, or RMS) of dragline silk exposed to UV irradiation for 15 or 60 minutes compared to a control sample.

16
plate tray for each trial. Each well was monitored over twelve days to assess proliferation,
alignment, and adherence. Prior to staining, cell adhesion and alignment was monitored by
optical microscopy from the first plating through the two-week period of incubation. They
were then stained with phalloidin and DAPI (4',6-diamidino-2-phenylindole) following the
kit staining procedures. Images were then evaluated to monitor the alignment of actin in
the cell along the spider silk. Fluorescent microscopy was done to evaluate the adherence
and alignment of the cells.
Alignment was evaluated using the particle roundness tool in ImageJ [68].
Roundness was calculated by using Eq. 1 with a value of 1.0 indicating a perfect circle and
a value approaching 0.0 indicating an increasingly elongated shape.
Roundness = 4[Area]π[Major axis]2 = 1
Aspect Ratio= Minor Axis
Major Axis (1)
Images of each of the three cell lines (DPSCs, 7F2, and 3T3s) were analyzed and data was
taken by calculating the percentage of the total cells that fell into a range of roundness
values and comparing average roundness data for each cell type. Low roundness values
were implicative of an irregular (in this case flat and aligned) shape, high roundness values
implied that the cells were in a circular morphology.
Table 3: Growth media solutions in which cells and silks were plated.

17
Roundness data was taken for each cell type at increments of 0.02 roundness units.
Each of the cell types were compared against each other to determine the percentage of
cells with roundnesses ranging from 0.00-0.32, 0.32-0.68, and 0.68-1.00 using a two-tailed
t-test assuming equal variance. Comparisons where p < 0.05 indicated significantly
different roundnesses between two groups.
2.5 EFFECT OF SLIDE PREPARATION METHOD AND RESPONSE OF CELLS TO
ULTRAVIOLET RADIATION
Pre-Drawn Silk on Elastomer Coated Slides
Initial trials on the hand-wrapped silk slides using elastomer and no UV radiation
experiences severe contamination (Figure 8) due to the exposure of spider silk to a non-
sterile environment. This prompted the use of a sterilization method on the silk for future
trials in an effort to reduce contaminants that would kill cells. Along with issues from
contamination, the initial elastomer solution used to adhere the silk to the slides had adverse
reactions with the silk and cells, potentially due to issues with curing of the polymer and
the constant exposure to a liquid media. Figure 9 shows the spider silk released ‘bubbles’
that became captured in the elastomer and made the silk difficult to adhere to. It is
speculated that the gas ‘bubbles’ were forced out from the spider silks as they went from a
natural to super contracted state (from being introduced to the mostly liquid, uncured
elastomer) and then remained in the more cured elastomer resin. From this point, it was
determined that a very minimal amount of adhesive needed to be used in future trials, which

18
led to the usage of small amount of super glue when wrapping silks with the motorized
wrapping unit later on.
Interaction with Silk from Machine Reeling of Live Spiders
Cell adhesion and alignment was monitored by optical microscopy from the first
plating through the two-week period of incubation. In this manuscript, 24 hrs after plating
will be referred to as Day 1. Figure 10 shows a progression of the proliferation and
alignment of DPSCs on their spider silk matrix. During Day 1, cells still exhibit some
Figure 8: Release of gas 'bubbles' from spider silk on elastomer coated scaffold trial. This image was taken 3 days after plating DPSCs and no cells were observed near or upon the spider silks.Figure 9: Development of contamination upon hand-wrapped,
elastomer coated spider silk slides. Top images show bacterial contamination. Bottom images show fungal contamination.
Figure 10: Images show DPSCs forming a general trend of adhesion to the spider silk, proliferation, and an eventual overall alignment of the cells to the silk line over time. Images were taken by light microscopy at 10X magnification.

19
roundness because they had not yet adhered to a surface, but by Days 5, 7, and 12, cells
show greater spreading, higher numbers, and a more aligned orientation as a population to
the silk.
Impact of Sterilization by UV Radiation and Cell Type Influence on Adherence and
Alignment
Figure 11: Fluoroscopy images taken after two weeks of growth show alignment and adhesion of cells to the spider silk in comparison with exposure time to ultraviolet radiation. The red stain (phalloidin) shows the f-actin cytoskeletal fibers, the blue DAPI stain indicates the nuclei and shows the number of cells, and the green autofluorescence indicates the silk. The 0 min UV light trial for osteoblasts experienced bacterial contamination, resulted in very limited cell growth on and around the silk. DPSCs (top row) showed the best alignment in individual cells and in cell populations as a whole. Osteoblasts (middle row) had some good individual cell adherence and alignment, but did not show bulk population alignment. Fibroblasts were mostly observed clumping around the silk and on the scaffolds in general with very little organized individual or population alignment.

20
Cells were plated on scaffolds treated with different levels of UV light exposure to
determine if the ultraviolet radiation would cause any effect in the way that the silk
interacted with the cells. Like the environmental stress trials, the UV light exposure trials
were given two weeks for proliferation before staining for nuclei and actin. Figure 11
shows a side-by-side comparison of different cell type responses on scaffolds exposed to
different levels of UV light. Actin filaments are most aligned to the silk strands in the
DPSC trials both in individual cells and cells as a population. In osteoblasts, some
individual cells near the silk showed actin alignment, but cells further away from the silk
have unaligned actin. The same can be seen in fibroblasts that, like in their natural state,
were shown to have more of a ‘clumping’ morphology around the silk. In the DPSC and
osteoblast trials, there was no difference in cell proliferation, morphology, or behavior
depending on the level of UV light exposure. In the fibroblast trial, there was bacterial
growth on the silk in scaffold with 0 min of UV light exposure that terminated most of the
cells on these scaffolds. There was no difference in cellular response in fibroblasts on
scaffolds with 5 or 30 min of UV light. The blue DAPI stained nuclei are used to indicate
how many cells are present and show that there was proliferation in all three cell types.
2.6 RESPONSE OF CELL TYPES TO SILK
The alignment responses of the three cell types to the spider fibers were compared,
and the roundness of cells was assessed using the Roundness function in ImageJ. This
function compares the intensity within neighboring pixels to identify boundaries of cells
and later calculate the cell diameters. Figure 12 shows an example of the software
analyzing an optical image of cells. This figure clearly shows the overlay of the ImageJ

21
selected areas over the original optical image of the cells, confirming that the results are
accurate representations of the cell shapes. The average roundness of cells within each cell
type calculated with the ImageJ software are shown in Table 4. Osteoblasts (Osteo) have
the highest roundness value of 0.492, meaning they are the most circular in nature.
Following the osteoblasts are the fibroblasts with an average roundness of 0.331. DPSCs
had the lowest average roundness at 0.114, indicating that they are the most elongated of
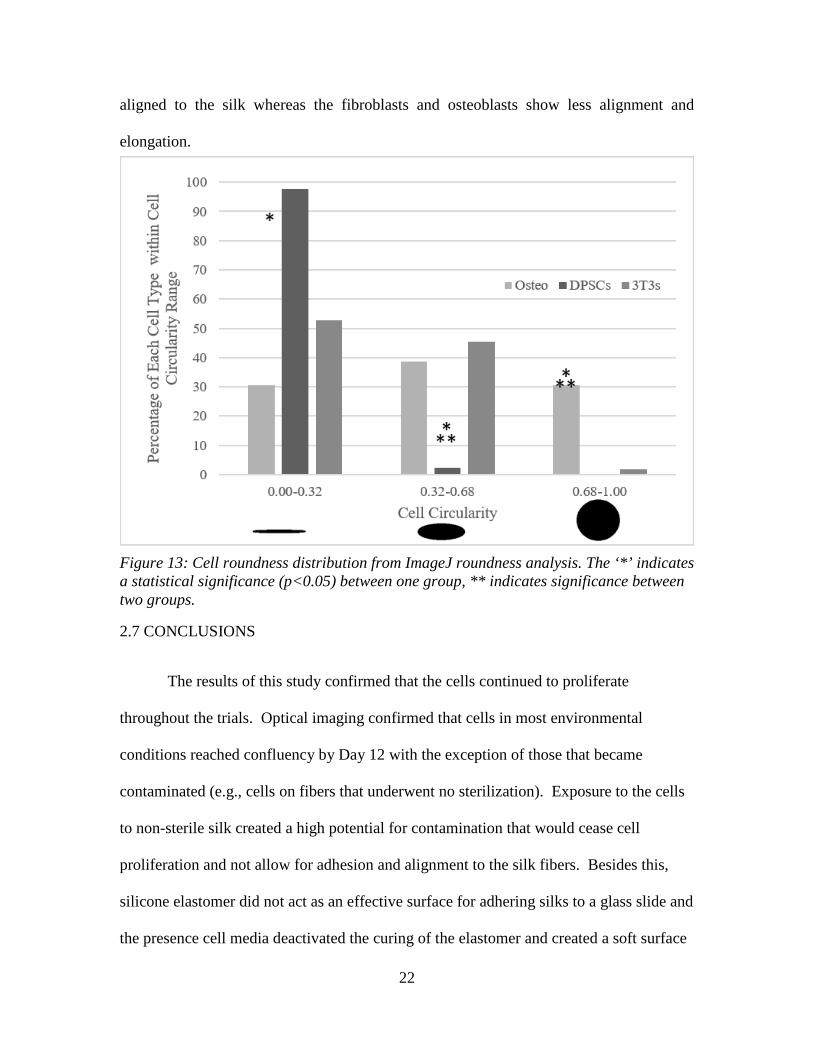
the cell types. Figure 13 shows the distribution of cells at each roundness range. As shown
by the average roundness and roundness distribution, DPSCs have the majority of their
cells falling into an elongated state (0.00-0.32) and the osteoblasts and fibroblasts show
greater percentages of cell in a more round morphology (0.32-1.00). This is representative
of fluoroscopy images where DPSCs showed a greater population trend of elongated and
Figure 12: ImageJ analysis to create threshold microscopy image to determine cell roundness. *all scale bars same, 400 μm *
Table 4: Average roundness varied by the three cell types (osteoblasts, fibroblasts and DPSCs). These were calculated using the ImageJ roundness analysis.
22
aligned to the silk whereas the fibroblasts and osteoblasts show less alignment and
elongation.
2.7 CONCLUSIONS
The results of this study confirmed that the cells continued to proliferate
throughout the trials. Optical imaging confirmed that cells in most environmental
conditions reached confluency by Day 12 with the exception of those that became
contaminated (e.g., cells on fibers that underwent no sterilization). Exposure to the cells
to non-sterile silk created a high potential for contamination that would cease cell
proliferation and not allow for adhesion and alignment to the silk fibers. Besides this,
silicone elastomer did not act as an effective surface for adhering silks to a glass slide and
the presence cell media deactivated the curing of the elastomer and created a soft surface
Figure 13: Cell roundness distribution from ImageJ roundness analysis. The ‘*’ indicates a statistical significance (p<0.05) between one group, ** indicates significance between two groups.

23
that cells could not adhere to. For this reason, a sterilization method (UV radiation) and
motorized reeling mechanism relying on small amounts of super glue was used in order
to allow for successful cell culture on spider silk slides.
The application of sterilization by UV light did not alter the cell proliferation and was
beneficial to the research team since it eliminated evidence of the bacterial and fungal
growth within the plates. Therefore, UV radiation can be used as a method of sterilizing
spider silk for cell culture without causing negative effect to cell proliferation, morphology,
or adhesion and alignment to silk. Besides this, the prevention of the spread of bacteria
and fungus better enables cellular growth, so the use of UV radiation as a sterilization
method was beneficial to cell adherence and alignment on silk in comparison to untreated
silk.
In regards to response of cell types to silk, there were most obvious differences
observed when looking at an individual cell scale (looking at the adhesion and alignment
of individual cells directly contacting silk) and a population scale (looking at the orientation
and alignment of cells that were not directly contact silk). On an individual cell scale, there
were instances of the osteoblasts and DPSCs adhering and aligning to the spider silk. These
cells were in direct contact with the silk fibers and their alignment indicates that the silk
fibers were an effective scaffold. Contrastingly, fibroblasts were found to gather around
the silk fibers, but showed no alignment of actin or in general morphology to the silk fibers.
With respect to cells that were not in direct contact with the silk fibers, the alignment
depended on the cell type. DPSCs aligned and elongated in the same orientation as those
in direct contact with the silk. However, the fluoroscopy images and ImageJ roundness
analysis showed that the fibroblasts and osteoblasts not in direct contact with the fibers did

24
not have the same elongation or orientation as cells in direct contact. Roundness measure
results aligned with the qualitative results of the fluoroscopy images and confirmed that
the DPSCs showed significantly more elongation than the other cell types. This suggests
that the spider silk scaffold can act as a significant aligning tool for large DPSC populations
and not just on a cell-by-cell basis. This trend also implies that these fiber scaffolds might
also act as an effective scaffold for differentiation of DPSCs along more elongated cell
morphology lineages (e.g., neural lineages) given the proper growth environment.

25
CHAPTER THREE - CELLULAR RESPONSE TO SILK REELED UNDER ENVIRONMENTAL STRESS AND RESPONSE TO SILK PROPERTIES AND
ORIENTATION 3.1 INTRODUCTION
Spiders are also known to produce silk of varying amino acid content when exposed
to stressful situations like being underfed or enclosed [69]. In addition to their mechanical
properties, these fibers are often studied to understand their ‘smart’ response to
environment. Dragline fibers have been shown to contract in high humidity while also
being insoluble for long periods in most aqueous solvents like those found in cell culture
and naturally in the body [23]. There is high potential for treatment of wounds and injuries
using polymers of natural origin (like spider silk), so there has been a recent drive to
manipulate natural polymers to recreate recombinant versions to electrospin and form into
hydrogel scaffolds [70]. Besides creating a wider variety of scaffold morphologies, use of
recombinant silk eliminates the problem of needing to have live spiders that silk very small
amounts of silk at a time. If a ‘synthetic’ recombinant silk can be created with the same
beneficial properties of natural silk, then it will be more feasible to use in treatments.
3.2 EFFECTS ENVIRONMENTAL STRESSES ON SPIDER SILK AND
RESPECTIVE EFFECTS OF MORPHOLOGY AND ORIENTATION ON CELL
ADHESION TO SILK
Environmental Stresses on Spiders and Silks
Spiders are naturally exposed to a wide varieties of environments and have been
shown to adapt to well to changes in their environment [71]. Previous studies have shown
the effects of heat and toxins on biochemical markers like Hsp70, an indicator that a stress

26
threshold has been exceeded, in spiders (not necessarily their silk) and have concluded that
induced stress on spiders can alter the biochemical signals of their body cells [71]–[73].
Additional studies have shown that changes in diet (or induced stress by starvation/cut back
of diet) have an effect on the amino acid content of spider silk, thus affecting spider silk
mechanical properties [74], [75]. For instance, in a study comparing major ampullate silk
of Nephila pilipes spiders that were fed crickets or flies, silks were shown to have greater
stiffness and thicker diameters when the spiders were fed crickets. This was attributed to
the change in the amino acid and spidroin proteins present in the silks [74].
There are not many current studies that evaluate the effects of environmental
stresses on spiders, so this thesis will aim to investigate whether environmental stresses to
spiders will create significant enough change to silk properties to improve or deter cell
adhesion. Major ampullate silk is expected to provide for much of the structure in a web,
and thus is expected to hold up to many stresses after being drawn. This study will evaluate
the effects of enclosing spiders in a small space, thus not allowing them to form normal
webs, and the effects of not exposing spiders to light, thus providing a ‘nighttime’
environment at all times. These silks will be evaluated for changes in physical properties
like diameter and also for effects on cell adhesion response.
Figure 14: Diagram of spider spinneret and valve [20].

27
Effect of Diameter, Spacing, and Orientation on Cell Interactions
There are several factors impacting silk diameter during spinning, especially in
terms of environment and spider size. Normally, a mature Nephila will produce a dragline
silk at a rate of 1cm/sec during web construction (but can produce up to ten times faster
when created an escape line to drop from a web) [76]. Silk diameter is controlled and
modified by a valve at the end of the duct of a spinneret, the size and shape of this valve
will impact the produced silk diameter and surfaces [30]. Dragline silk has an average
diameter of approximately 7um and a circular cross section, but the diameter can be
influenced by spider weight within a small range [77]. Spinning speed does not have a
significant effect on silk diameter, but can play a role in the strength and toughness of the
fiber produced. Another factor influencing diameter of silk is the environment that it is
placed into. Spiders’ silks are resistant to most solvents, but they will undergo
supercontraction up to 60-80% when introduced to a polar solvent like water or cell media
[23]. This contraction can induce a morphological change in conformation and orientation
of molecular chains in the silk as hydrogen bonds are broken [36], [78]. This generally
causes an increase in diameter and elasticity, but a decrease in strength of the silk [79].
Diameter of the spider silks can impact the way that cells adhere and align to them, and
this alteration in micropatterning can influence the way that cells behave and potentially
differentiate.
Previous studies have conflicting results in terms of which diameters of silk
produce the best results when culturing cells, especially when considering a nanoscale
surface topography [33], [80]. Small dimensions of fibers can be used to mimic features
of extra cellular matrix (ECM) fibers that cell adhere to. Adherence of cells to a surface is

28
also dictated by surface energy of the material, for instance, a hydrophilic material has a
higher surface energy and results in more spread out, spindle-shaped cells in comparison
to rounder cells on low surface energy hydrophobic surfaces [80]. When dealing in fibers
that ranged from 2nm to 10nm, the 10nm fibers experienced the best seeding and
attachment, but when dealing in fibers that ranged from 400nm to 1200nm, the optimal
alignment occurred in fibers that were 400nm [33]. This indicates that there is probably
an optimal zone for cellular adhesion and alignment, at least in regard to a nanoscale. This
study will evaluate silk fibers (that will be in supercontraction due to a liquid media
environment) that are in a microscale as opposed to a nanoscale like the electrospun fibers
from the previous studies cited. Previous studies indicate that as a fiber diameter
approaches a cell’s diameters, then the limiting effects of having a severe fiber curvature
becomes less predominant and more cells can adhere and align to a fiber surface. Spacing
between micropatterns and fibers can also impact the way that cells align and spread, where
more space between fibers creates a less spindle-like morphology [81]. Stem cells were
Figure 15: Effect of surface energy (hydrophilicity) on contact angle.

29
also shown to prefer an aligned silk orientation over a random orientation when plating on
Tussah silk worm silk [33].
Recombinant Spider Silk
The properties that make spider silk so beneficial to use as a biomaterial are
somewhat offset by the difficulty of obtaining natural silk. However, there has been
evidence of recombinant silk, created from natural silk DNA, acting as a viable source for
cell adherence and growth [70], [82], [83]. As aforementioned, the spider silk consists of
alternating crystalline and amorphous regions on a molecular level, with crystalline regions
forming β-sheets that are high in stiffness and rich in alanine, glycine-alanine, and glycine-
Figure 16: Cell response on different diameter silks - A) Cross sectional view showing spreading and wrapping, B) Topographical view showing spreading and wrapping.

30
alanine-serine repeats [84]. These β-sheets are present in both the major ampullate
spidroin 1 and 2 (sp 1 and sp 2) of ampullate silk [30], [31], [85]. These sheets come
together to form spider proteins that are enormous in molecular weight, weighing several
hundred kDa [86]. This size and weight is what is assumed to be the source of excellent
toughness and tensile strength in spider silks, and these sizes of proteins are able to be
transformed into recombinant proteins in plants – especially tobacco plants [84], [87], [88].
The creation of recombinant silk also allows for the opportunity to add new genetic motifs,
in addition to those that naturally exist in the silk that promote adherence, that can influence
the interaction of the cells with the recombinant material [70], [89]. For this reason,
recombinant silk threads and hydrogels created from transgenic tobacco leaves are
expected to act similarly to natural silk in the way that the influence cells to adhere and
align.
3.3 MATERIALS AND METHODS
Figure 17: Transformation of spider silk genetic information into recombinant spider silk proteins using plant-based transformation.

31
Environmental Stress
The tent was positioned next to a window to ensure natural lighting conditions and
was limited to up to four other spiders inhabiting the space at any given time. The spiders
within these conditions will be referred to as the ‘control spider population’. These
enclosures allowed the spiders to form full orb webs. Those spiders held within the 18.50
in by 18.25 in by 7.50 in Plexiglas boxes will be referred to as ‘boxed spider population’.
These spiders were provided with sticks to support web building.
For some silk collections, the exposure of the spider to light one hour prior to
silking was also controlled. This would better simulate how silk might be produced during
a nighttime period. Spiders removed from the control spider population and then kept in a
dark environment for 45-60 min prior to silking will be referred to as ‘limited light exposure
population’ within this thesis. Human DPSCs were thawed and cultured in a T75 flask to
prepare them for plating onto the wrapped silk scaffold. After the cells reached confluence
in the T75 flask, they were trypsinized and passaged to be seeded onto the wrapped silk
Table 5: Matrix showing variation in the spider enclosures/environmental conditions prior to silking and the population naming systems referenced within this thesis.

32
scaffolds at a density of 25,000 cells per 20mm x 20mm slide. Cells were maintained in a
standard media (Table 2) and allowed to culture for 2 weeks.
Collection environment testing was set up primarily to monitor opposing
environments; control vs. limited space and limited light as described in Table 5. The
environments were established to introduce varying levels of environmental stress to the
spiders before silk collection.
Recombinant Silk Fibers
As a comparison to the natural control environment spider silk, a ‘synthetic’ silk
created from recombinant silk protein of non-animal or human origin was also used in the
silk trial comparisons. Synthetic constructs containing native Nephila clavipes dragline
spidroin 1 or 2 N- and C-terminal domains that flank 8 consensus repeat domains were
transformed into tobacco each separately. The proteins were produced in the leaves of
tobacco plants and were purified via single step affinity purification [90]. The proteins were
concentrated and dialyzed against 5 mM ammonium bicarbonate and freeze-dried into
viscose liquids. Crosslinking was performed by adding 0.2 µl of acetic acid (25%) and 5
µl of glutaraldehyde (25% aqueous solution) sequentially to approximate 25 mg
recombinant spider silk protein. The mixture was maintained at ambient temperature for
more than 5 hours and then diluted to spinning solution by 10 mM phosphate buffer (pH
7). Recombinant spidroins 1 and 2 were mixed at a 70:30 ratio and overlaid with 0.5%
gellan gum solution that was kept at 55 °C. Fibers were pulled from the interface of the
recombinant spidroins and gellan gum and air-dried [91]. Human DPSCs were thawed and
cultured in a T75 flask to prepare them for plating onto the wrapped silk scaffold. After
the cells reached confluence in the T75 flask, they were trypsinized and passaged to be

33
seeded onto the wrapped silk scaffolds at a density of 25,000 cells per 20mm x 20mm slide.
Cells were maintained in a standard media (Table 2) and allowed to culture for 2 weeks.
Recombinant Silk Hydrogels
Hydrogel samples were developed in a separate lab using a gellan gum and chitosan
solution. Nephila clavipes dragline spidroin 1 or 2 mimics that containing the N- and C-
terminal domains that flank 8 consensus repeat domains were produced from transgenic
tobacco. The proteins were purified via single step affinity purification [90]. The proteins
were concentrated and dialyzed against 5 mM ammonium bicarbonate and freeze-dried
into viscose liquids. The concentrated proteins were further processed as described in Peng
et al., 2017 [92]. In summary, 0.2 µl of acetic acid (25%) and 5 µl of glutaraldehyde (25%
aqueous solution) were sequentially added to approximate 25 mg recombinant spider silk
protein. The crosslinking reaction was carried out at ambient temperature for more than 5
hours and then diluted by 10 mM phosphate buffer (pH 7). For hydrogels containing single
spidroin mimics, each crosslinked spidroin mimics were mixed with chitosan (5 mg/mL in
1% acetic acid) solution in volume to volume ratio 1:2. (medium molecular weight, Sigma-
Aldrich Cat. No. 448877). For hydrogels containing two spidroin (Spidroin 1 and 2)
mimics, spidroin 1 and 2 mimics were mixed at a 70: 30 ratio and then mixed with twice
of the total volume of chitosan solution. The spidroin mimics and chitosan mixture was
then overlaid with 0.5% gellan gum solution. Hydrogels were formed when the mixture
remain undisturbed in about 5 hours. Fibroblasts were then plated directly onto the
prepared hydrogels at a density of 20,000 cells per hydrogel well. 3T3s were treated with
their appropriate media and cells were allowed to proliferate for 7 days before staining with
actin and DAPI and imaging.

34
3.4 EFFECTS OF SILK ORIENTATION ON CELLS
Handwrapping silk slides created a random alignment of fibers, as indicated in
Figure 18, and this effected the way that cells were able to adhere and align to the silks.
When the silks were in a random orientation, it was less likely for cells to bind to them.
The looseness of the handwrapped silks also created multiple planes of view and allowed
cells to adhere to the glass slide rather than the spider silks themselves, as seen in Figure
19. Once the motorized wrapping mechanism was used to wrap silk directly onto the glass
slides in an aligned, tight fashion, cells were more successful in finding and aligning to the
silk. Figure 20 shows these results of adherence of ADSCs happening in triplicate on motor
wrapped silk slides. The morphology of the cells on the silks mimicked those of that were
Figure 18: Random orientation of silks when handwrapped. Images were taken 3 days after plating cells and no cells were present near or on silks.
Figure 19: Effects of handwrapping silks and creation of multiple planes of view. These images were taken at day 3 after plating and cells only appear adhered to the glass slide in the bottom plane of view rather than upon the spider silks.

35
not introduced to silk and left to culture normally in a T75 flask for 5 days. Overall, there
was no instance of cell adherence or alignment observed on the handwrapped, randomly
oriented silk slides and the cells preferred to adhere to the standard glass surface.
3.5 RESPONSE OF CELLS OF DIFFERENT PROPERTIES AND DIAMETERS
Effects of Environmental Stresses on Spider Silk and Interaction with Cells
Trials were also run to determine if the environment of silk drawing had any effect
on the material composition or if surface properties of the spider silk caused an impact on
their cellular response. Only DPSCs were used in these trials when comparing silk
collected from a spider in different, stress-inducing environments versus the silk collected
in normal conditions. After two weeks of proliferation, the scaffolds were stained to show
#03 #02 #01 T75 Flask Silk Cells Silk Cells Silk Cells Cells
Day 0
Day 3
Day 5
Figure 20: Motor wrapped silk slides show better initial adherence of ADSCs under optical microscopy after 5 days than randomly oriented handwrapped silks.

36
actin and nuclei in the cells. The spider silk autofluoresced in green and can be noted by
its green threadlike structure. The red phalloidin stain shows the alignment of the actin in
the cytoskeleton of the cells, indicating the way that they are spread along the silk or not
along the silk. Figure 21 compares ‘control spider population’ vs. ‘boxed spider
population’ and ‘limited light exposure population’ environments. In these trials there was
no significant difference in the response of the DPSCs plated on the scaffolds. In either
case, the stains showed that the actin of the cells was aligning to the silk regardless of
environment of silk drawing. In each case, it was evident by the number of blue nuclei that
there was no negative effect of the surfaces on cell proliferation near and away from the
silks drawn from spiders in any condition.
Figure 21: Fluorescent microscopy depicts DPSCs after 2 weeks of growth on spider silk drawn from different environments – A&B = ‘control spider population’, C = ‘boxed spider population’ (Limited Space), D = ‘limited light exposure population’.

37
Cell Response to Recombinant Silk Hydrogels and Fibers
In comparing the control, natural spider silk, and the recombinant protein
‘synthetic’ silk, it can be discerned from the staining (Figure 22) that the synthetic silk
fiber’s diameter is noticeably larger and varies along the fiber length. Natural silk has a
near constant diameter of approximately 5-10 μm while the recombinant silk (which
autofluoresced in green, red, and blue, resulting in a yellowish color in the fluoroscopy
images) has a much more varied morphology and diameter closer to 75-125 μm. However,
the material of the synthetic silk is near identical to that of the recombinant silk hydrogels,
which cells remained viable on as is seen in Figure 23. This implies that material
composition was not responsible for the lack of cell adherence to the recombinant silk
fibers, and rather that surface morphologies and large diameter were responsible for lack
of cell interaction. Besides a difference in morphology of the fibers of the natural and
recombinant silk, the stained scaffolds showed that there was DPSC adhesion and
alignment to the natural silk, but there were no cells observed adhered to or near the
recombinant silk. Other cell types (3T3s and 7F2s) responded similarly. While cells
Figure 22: Fluorescent microscopy depicts DPSCs after 2 weeks of growth on spider silk of different materials – Left = Naturally draw dragline spider silk (control) and Right = Hand drawn recombinant spider protein silk (synthetic). There is no cell adhesion or alignment on the synthetic silk.

38
remained viable in the three dimensional hydrogel, they did not take on an elongated shape
or have any specific orientation. These result indicate that there is a need to develop a
more regulated recombinant fiber surface and diameter (closer to the natural silk diameter)
to enhance cell interaction and alignment.
3.6 CONCLUSIONS
Once an effective plating method was selected, the aligned orientation created by
the motorized wrapping mechanism allowed for better cell adhesion and alignment than
the randomly oriented silk slides that had minimal cell adhesion and alignment. There were
not any significant differences between cells plated on silk produced by spiders within
selected environmental conditions. Fluorescent imaging showed that cell adhesion and
alignment was evident for all of the spider produced silks, regardless of the stress that they
were under. This demonstrates that environmental stress induction did not create a
significant enough change in amino acid composition of the produced spider silks that it
would negatively impact cellular interaction. Due to success in growth and viability of
cells on recombinant silk hydrogels of the same material as the recombinant silk fibers, it
can be concluded that the composition of the recombinant silk itself was not responsible
sp 1 sp 1/2 sp 2
Figure 23: DPSCs on hydrogels of different spidroin compositions. sp 1 is made from Spidroin 1 mimics, sp 2 from Spidroin 2 mimics, and sp 1/2 from Spidroin 1 and Spidroin 2 mimics in a 70:30 v/v ratio. All mimics were combined with chitosan and gellan gum.

39
for the lack of cell interaction on recombinant fibers. Comparison of cell response to these
spider produced silk fibers with the drawn ‘synthetic’ recombinant silk fibers suggested
that the recombinant fiber surface in contact with the cells (the outer layer) might either be
of a composition not recognized by the cells or that their relative size and shape adversely
affects cell adhesion and growth. Other groups have shown conflicting results.
Recombinant spider silk proteins drawn into fibers were reported to support fibroblast cell
adhesion and growth [93]. One study reported that their recombinant silk proteins pressed
into films or electrospun into nanofibers promoted neural stem cell differentiation and
mesenchymal stem cell growth [94], [95]. To improve the results of the recombinant silk
fiber trials, it is imperative to create a fiber that is close in diameter to natural silk (more
optimal grown on diameters 5-10 μm than 75-100 μm) with a similar surface roughness.
More importantly, these results suggest that spiders can be housed under a number
of conditions that will not impact silk composition so severely that it will influence the way
that cells adhere and align to natural silk fibers. This implies that other housing situations
used in other labs’ research should not impact the cellular interaction with silk and that
these environmental factors can be dismissed when comparing research. The results also
stress the significance of constructing a ‘synthetic’ recombinant silk with the appropriate
surface properties and diameter. Since cells were able to survive and replicate in a
recombinant silk hydrogel material, similar results were expected in the recombinant silk
fiber trials. However, the significantly different diameters and surface morphologies are
suspected to have contributed to the lack of success in inducing cell adhesion and
alignment. Chapter four will discuss further suggestions to improve this portion of the
study using electrospinning to make individual fibers or fiber mats that can be seeded with

40
the intent of implantation. Assuming cells interact more appropriately with a regulated
recombinant silk fiber, this material could serve as an effective scaffold for nerve
regeneration.

41
CHAPTER FOUR – FUTURE WORK WITH NEUROGENIC REGENERATION AND RECOMMENDATIONS
5.1 BACKGROUND FOR NEUROGENIC REGENERATION
Traumatic injuries to the peripheral and central nervous system require some type
of graft to allow for regeneration of viable cells to bridge the gap between existing cells.
Due to the nature of adhesion and alignment of cells to a spider silk scaffold, there is good
potential for silks to act as a surface to promote neurogenic differentiation. Since neurons
are unable to regenerate themselves, there is a need for the development of a method to
culture neurons through differentiation using other types of cells like ADSCs and DPSCs
[52], [53]. Micropatterning, physical cues, and with biological cues can induce
differentiation in these cells lines to promote a neurogenic lineage [96]–[98]. Stiffness and
toughness of biomaterials can cause cells to express morphologies of other cell lines, and
the alignment of cells on the silk scaffolds is expected to promote neurogenic
Figure 24: Physical cues and the induction of differentiation based on surface micro-pattern [97].

42
differentiation [98]. Based on results seen in other studies, using spider silk as a route to
neurogenic differentiation is assumed to have positive outcomes and may work well on the
natural silk scaffold or recombinant scaffolds like those used in the studies of this thesis.
5.2 RECOMMENDATIONS
These studies showed that there should be no significant effect of living and reeling
environment on that will interfere with cellular interaction. The studies also demonstrated
that ultraviolet radiation can function as an effective means of sterilizing naturally drawn
silk for cell culture to prevent the presence of bacterial and fungal contamination.
Elastomer coatings were ineffective in constructing silk scaffolds, and randomly wrapped
silk could not sustain cell adhesion and alignment as well as parallel aligned fibers. Plant-
based recombinant ydrogels were able to maintain cell viability, but did not allow for any
alignment or elongated morphology of cells, while recombinant silk fibers were either too
thick in diameter or too variable in surface to sustain cell adhesion and alignment. Overall,
these studies characterized the behavior of different cell types on natural spider silk,
recombinant silk hydrogels, and recombinant silk fibers.
While this project laid the groundwork for developing an understanding of the
reaction and behavior of cells on natural spider silk scaffolds, there is a lot of room to
further develop a synthetic recombinant silk and/or move toward differentiating cells using
silk as a micropattern. Since spider silk acted as an effective means of adhering and
aligning cells, it would be beneficial to be able to create a recombinant silk fiber that has
more equivalent physical properties to those of natural silk fibers. Other studies have
referenced the use of electrospinning to create smaller diameter fibers, and this would

43
create a more controlled, smoother recombinant fiber for a future study. Developing an
electrospinning protocol that would regulate fiber diameter while also creating an aligned
orientation could unlock a potential to make a more three dimensional scaffold for cell
seeding.
Along with simply manipulating the physical morphology of the silk fibers
themselves, there are still a wide variety of options to influence differentiation of cells
using biological markers. Previous studies cite using growth factors within cell media to
direct differentiation, while other papers have suggested that the addition of different amino
acid motifs to a transgenic, recombinant proteins can enhance cell differentiation.
Furthering these studies to include electrospun recombinant silk fibers, or including a
growth factor based media to induce neurogenic differentiation has the potential to open
the door to functionalizing spider silk as a biomaterial for use in wound healing and nerve
regeneration.
Figure 25: Typical examples of electrospinning methods. Traditional electrospinning method to prepare nonwoven fabric scaffolds (a), the electrospinning method to prepare cotton ball-like scaffolds (b), the electrospinning method to prepare double-layered 2-D architectures (crosshatch pattern) using a 3-D stage (c), andthe Nanospider electrospinning method (d). [51]

44
APPENDICES

45
Appendix A MODELING CELL ROUNDNESS IN IMAGEJ
ImageJ was used to analyze roundness off cells along spider silk. Roundness is a
way of comparing the aspect of major and minor axis, when the major and minor axes are
similar in value, the roundness value is closer to 1.0 meaning more circular. A smaller,
decimal value for roundness means that the particle (or cell in this case) is more elongated.
The equation for roundness is essentially just the inverse of Aspect Ratio: Roundness =
4[Area]π[Major axis]2 = 1
Aspect Ratio= Minor Axis
Major Axis
After fluorescent microscopy images were taken, .tif files were selected for analysis
in ImageJ. A fitting image for analysis would focus around one silk fiber and have cells
that had distinguishable shapes, like the image shown in Appendix Figure 1. The image is
opened in ImageJ and converted to an RGB Stack (Image>Type>RGB Stack) to allow for
a threshold to be overlaid on the image. The resulting image appears as is shown in
Appendix Figure 2. Once the image was converted to an RBG stack, a threshold could be
applied (Image>Adjust>Threshold>Apply>check ‘Black Background’ and ‘Only Convert
Current Image’) to create distinct shapes for ImageJ to distinguish as individual particles,
shown in Appendix Figure 3. It is important to select threshold values that create edges on
Appendix Figure 1: Example Fluoroscopy Image for Roundness Analysis in ImageJ
Appendix Figure 2: Fluoroscopy Image Converted to RGB Stack in ImageJ

46
each cell, but do not cause cells to run together. Once the threshold is set and paricles are
distinguished, an algorithm in the program can measure cell circularity (Analyze>Analyze
Particles…) and this will show the drawn outlines of cells and provide a chart of their
roundness values as seen in Appendix Figure 4 and Appendix Table 1. In order to get
smaller sample sizes for roundness ranges, cells were evaluated at increments of 0.02 units
of roundness were taken, then cells were grouped into larger ranges to express their general
roundness (Appendix Table 2). Analyzed particles were all given numbers, so ‘particles’
like the silk itself and the scale bar could be exempt from calculations by disregarding those
particle numbers. Cells were then organized by roundness range and graphed based on the
Appendix Figure 3: Threshold overlay on fluoroscopy image.
Appendix Figure 4: Analyze Particles... in ImageJ for roundness
Appendix Table 1: Values given from Analyze Particles... in ImageJ for roundnesses 0.00-1.00. To distinguish roundness areas, range will have to be changed.

47
percentage distribution of their roundness values (Appendix Table 3). This data was
reported in the thesis Chapter 2.
Appendix Table 3: Distribution of cells that fall into roundness ranges to describe the general trends of elongation or roundness of cells as analyzed by ImageJ.
Appendix Table 2: Evaluation of roundness on 0.02 unit increment to give average roundness and percentage of cells at each roundness increment.

48
Appendix B CHARACTERIZING REELING MECHANISM
In order to give an even spacing between silk fibers when reeling, a sheet was set
up to allow for predictable spacing to be programmed into the STM17Q-2AN motors
(Appendix Figure 1) being used to turn a slide. The system worked with two motors, one
which rotated the silk slide to allow for reeling, and another that moved back and forth to
control spacing of the silk. An Excel spreadsheet was set up to calculate desired speeds
for either motor (Appendix Figure 2). The ‘clip motor’ was responsible for rotating the
slide and the ‘stage motor’ moved the slide back and forth for the spacing. This allowed
for alterations in slide size and could account for variable reeling speeds for intended
Appendix Figure 1: STM17Q-2AN Motor used for reeling silk onto slide
Appendix Figure 2: Excel spreadsheet to calculate motor speeds needed for controlled spacing of silk.

49
spacing between silk strands. The following calculations explain how each motor speed
was determined:
𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷 𝐴𝐴𝐴𝐴𝐴𝐴𝐴𝐴𝐷𝐷𝐴𝐴 𝑆𝑆𝑆𝑆𝐷𝐷𝐴𝐴𝐷𝐷[𝑚𝑚𝑚𝑚]
= 2 ∗ (𝑆𝑆𝑆𝑆𝐷𝐷𝐴𝐴𝐷𝐷 𝑊𝑊𝐷𝐷𝐴𝐴𝐷𝐷ℎ[𝑚𝑚𝑚𝑚]) + 2 ∗ (𝑆𝑆𝑆𝑆𝐷𝐷𝐴𝐴𝐷𝐷 𝑇𝑇ℎ𝐷𝐷𝐷𝐷𝑖𝑖𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷[𝑚𝑚𝑚𝑚])
𝐶𝐶𝑆𝑆𝐷𝐷𝐶𝐶 𝑀𝑀𝐴𝐴𝐷𝐷𝐴𝐴𝐴𝐴 𝑆𝑆𝐶𝐶𝐷𝐷𝐷𝐷𝐴𝐴[𝐴𝐴𝐷𝐷𝑟𝑟𝐷𝐷
] = 𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐴𝐴𝐷𝐷𝐴𝐴 𝑆𝑆𝐷𝐷𝑆𝑆𝑖𝑖𝐷𝐷𝐷𝐷𝑆𝑆 𝑆𝑆𝐶𝐶𝐷𝐷𝐷𝐷𝐴𝐴 [𝑚𝑚𝑚𝑚𝐷𝐷 ]
𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷 𝐷𝐷𝐴𝐴𝐴𝐴𝐴𝐴𝐷𝐷𝐴𝐴 𝐷𝐷ℎ𝐷𝐷 𝐷𝐷𝑆𝑆𝐷𝐷𝐴𝐴𝐷𝐷 [𝑚𝑚𝑚𝑚]
𝐷𝐷𝐷𝐷𝐷𝐷𝐶𝐶𝐷𝐷𝐷𝐷𝐷𝐷𝑚𝑚𝐷𝐷𝐷𝐷𝐷𝐷 𝑁𝑁𝐷𝐷𝐷𝐷𝐴𝐴𝐷𝐷𝐴𝐴 𝐻𝐻𝐴𝐴𝐴𝐴𝐷𝐷𝐻𝐻𝐴𝐴𝐷𝐷𝐷𝐷𝐷𝐷𝑆𝑆𝑆𝑆𝐻𝐻 [𝐴𝐴𝑚𝑚]
= 𝐶𝐶𝑆𝑆𝐷𝐷𝐶𝐶 𝑀𝑀𝐴𝐴𝐷𝐷𝐴𝐴𝐴𝐴 𝑆𝑆𝐶𝐶𝐷𝐷𝐷𝐷𝐴𝐴 �𝐴𝐴𝐷𝐷𝑟𝑟𝐷𝐷� ∗ 𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐴𝐴𝐷𝐷𝐴𝐴 𝑆𝑆𝐶𝐶𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝐷𝑆𝑆 𝐵𝐵𝐷𝐷𝐷𝐷𝐵𝐵𝐷𝐷𝐷𝐷𝐷𝐷 𝑆𝑆𝐷𝐷𝐴𝐴𝐷𝐷𝐷𝐷𝐴𝐴𝐷𝐷 [𝐴𝐴𝑚𝑚]
𝑆𝑆𝐷𝐷𝐷𝐷𝑆𝑆𝐷𝐷 𝑀𝑀𝐴𝐴𝐷𝐷𝐴𝐴𝐴𝐴 𝑆𝑆𝐶𝐶𝐷𝐷𝐷𝐷𝐴𝐴 �𝐴𝐴𝐷𝐷𝑟𝑟𝐷𝐷� =
𝐷𝐷𝐷𝐷𝐷𝐷𝐶𝐶𝑆𝑆𝐷𝐷𝐷𝐷𝐷𝐷𝑚𝑚𝐷𝐷𝐷𝐷𝐷𝐷 𝑁𝑁𝐷𝐷𝐷𝐷𝐴𝐴𝐷𝐷𝐴𝐴 𝐻𝐻𝐴𝐴𝐴𝐴𝐷𝐷𝐻𝐻𝐴𝐴𝐷𝐷𝐷𝐷𝐷𝐷𝑆𝑆𝑆𝑆𝐻𝐻 [𝐴𝐴𝑚𝑚/𝐷𝐷]50𝐴𝐴𝑚𝑚
This allowed for predictable spacing given the variables that could be changed
within the programmable motor’s settings. Control over displacement of silk strands
allowed for fibers to be in an aligned orientation.

50
REFERENCES
[1] P. Fratzl, “Biomimetic materials research: what can we really learn from nature’s
structural materials?,” J. R. Soc. Interface, vol. 4, no. March, pp. 637–642, 2007.
[2] S. H. Lim, X. Y. Liu, H. Song, K. J. Yarema, and H. Mao, “Biomaterials The
effect of nano fi ber-guided cell alignment on the preferential differentiation of
neural stem cells,” Biomaterials, vol. 31, no. 34, pp. 9031–9039, 2010.
[3] M. Georgiou, J. P. Golding, A. J. Loughlin, P. J. Kingham, and J. B. Phillips,
“Engineered neural tissue with aligned, differentiated adipose-derived stem cells
promotes peripheral nerve regeneration across a critical sized defect in rat sciatic
nerve,” Biomaterials, vol. 37, pp. 242–251, 2015.
[4] C. C. Chang, K. C. Chang, S. J. Tsai, H. H. Chang, and C. P. Lin, “Neurogenic
differentiation of dental pulp stem cells to neuron-like cells in dopaminergic and
motor neuronal inductive media,” J. Formos. Med. Assoc., vol. 113, no. 12, pp.
956–965, 2014.
[5] S. Gronthos et al., “Stem cell properties of human dental pulp stem cells,” J Dent
REs, vol. 81, no. 8, pp. 521–535, 2002.
[6] S. Gronthos, M. Mankani, J. Brahim, P. G. Robey, and S. Shi, “Postnatal human
dental pulp stem cells ( DPSCs ) in vitro and in vivo,” PNAS, vol. 97, no. 25, pp.
13625–13630, 2000.
[7] T. Vehoff, a Glisović, H. Schollmeyer, a Zippelius, and T. Salditt, “Mechanical
properties of spider dragline silk: humidity, hysteresis, and relaxation.,” Biophys.

51
J., vol. 93, no. 12, pp. 4425–4432, 2007.
[8] J. M. Gosline, P. A. Guerette, C. S. Ortlepp, and K. N. Savage, “The Mechanical
Design Of Spider Silks: From Fibroin Sequence To Mechanical Function,” J. Eng.
Biol., vol. 3303, pp. 3295–3303, 1999.
[9] S. Lin et al., “Predictive modelling-based design and experiments for synthesis and
spinning of bioinspired silk fibres,” Nat. Commun., vol. 6, no. May, pp. 1–12,
2015.
[10] J. E. Trancik, J. T. Czernuszka, F. I. Bell, and C. Viney, “Nanostructural features
of a spider dragline silk as revealed by electron and X-ray diffraction studies,”
Polymer (Guildf)., vol. 47, no. 15, pp. 5633–5642, 2006.
[11] C. Wong Po Foo and D. L. Kaplan, “Genetic engineering of fibrous proteins:
Spider dragline silk and collagen,” Adv. Drug Deliv. Rev., vol. 54, no. 8, pp. 1131–
1143, 2002.
[12] F. Vollrath, P. Barth, A. Basedow, W. Engtrom, and H. List, “Local tolerance to
spider silks and protein polymers in vivo,” In Vivo (Brooklyn)., vol. 16, no. 4, pp.
229–234, 2002.
[13] S. A. Fossey and S. Tripathy, “Atomistic modeling of interphases in spider silk
fibers,” Int. J. Macromol., vol. 24, pp. 119–125, 1999.
[14] C. Radtke et al., “Spider silk constructs enhance axonal regeneration and
remyelination in long nerve defects in sheep,” PLoS One, vol. 6, no. 2, 2011.
[15] F. Roloff, S. Strauß, P. M. Vogt, G. Bicker, and C. Radtke, “Spider silk as guiding

52
biomaterial for human model neurons,” Biomed Res. Int., vol. 2014, 2014.
[16] B. Cao and C. Mao, “Oriented nucleation of hydroxylapatite crystals on spider
dragline silks,” Langmuir, vol. 23, no. 21, pp. 10701–10705, 2007.
[17] H. Wendt et al., “Artificial Skin - Culturing of different skin cell lines for
generating an artificial skin substitute on Cross-Weaved spider silk fibres,” PLoS
One, vol. 6, no. 7, 2011.
[18] M. Santin, A. Motta, G. Freddi, and M. Cannas, “In vitro evaluation of the
inflammatory potential of the silk fibroin,” J Biomed Mat Res, vol. 46, pp. 382–
389, 1999.
[19] F. Vollrath and D. P. Knight, “Liquid crystalline spinning of spider silk,” Nature,
vol. 410, 2001.
[20] P. M. Cunniff et al., “Mechanical and Thermal Properties of Dragline Silk from
the Spider Nephila clavipes,” Polym. Adv. Technol., vol. 5, no. March, pp. 401–
410, 1994.
[21] R. L. Horan et al., “In vitro degradation of silk fibroin,” Biomaterials, vol. 26, pp.
3385–3393, 2005.
[22] Y. Wang et al., “In vivo degradation of three-dimensional silk fibroin scaffolds,”
Biomaterials, vol. 29, pp. 3415–3428, 2008.
[23] D. Saravanan, “Spider Silk - Structure, Properties and Spinning,” J. Text. Appar.
Technol. Manag., vol. 5, no. 1, 2006.
[24] J. W. Kuhbier et al., “Interactions between Spider Silk and Cells – NIH / 3T3

53
Fibroblasts Seeded on Miniature Weaving Frames,” PLoS One, vol. 5, no. 8, pp.
1–9, 2010.
[25] N. Minoura, S. Aiba, Y. Gotoh, M. Tsukada, and Y. Imai, “Attachment and growth
of cultured fibroblast cells on silk protein matrices,” J. Biomed. Mater. Res., vol.
29, no. 10, pp. 1215–1221, 1995.
[26] S. P. Patil, B. Markert, and F. Gr??ter, “Rate-dependent behavior of the amorphous
phase of spider dragline silk,” Biophys. J., vol. 106, no. 11, pp. 2511–2518, 2014.
[27] M. B. Hinman, J. A. Jones, and R. V Lewis, “Synthetic spider silk : a modular
fiber,” TIBTECH, vol. 18, no. September, pp. 1701–1703, 2000.
[28] F. Vollrath, “Strength and structure of spiders ’ silks,” Mol. Biotechnol., vol. 74,
pp. 67–83, 2000.
[29] J. C. Zemlin, “A Study of the Mechanical Behavior of Spider Silks,” Collab. Res.
Inc., 1968.
[30] M. Xu and R. V Lewis, “Structure of a protein superfiber : Spider dragline silk J |,”
PNAS, vol. 87, no. September, pp. 7120–7124, 1990.
[31] I. S. A. T. Fiber, M. B. Hinman, and V. Lewis, “Isolation of a Clone Encoding a
Second Dragline Silk Fibroin The first clone isolated encoded a,” J. Biol. Chem.,
vol. 267, no. 27, pp. 19320–19324, 1992.
[32] P. Guerette, D. Ginzinger, B. Weber, and J. Gosline, “Silk properties determined
by gland-specific expression of a spider fibroin gene family,” Science (80-. )., vol.
272, no. 5258, 1996.

54
[33] J. Wang et al., “The effects of electrospun TSF nanofiber diameter and alignment
on neuronal differentiation of human embryonic stem cells,” J. Biomed. Mater.
Res. - Part A, vol. 100 A, no. 3, pp. 632–645, 2012.
[34] W. Engström and G. Hjälm, “Major ampullate spidroins from Euprosthenops
australis : multiplicity at protein , mRNA and gene levels,” Insect Mol. Biol., vol.
16, pp. 551–561, 2007.
[35] J. Gatesy, C. Hayashi, D. Motriuk, J. Woods, and R. Lewis, “Extreme diversity,
conservation, and convergence of spider silk fibroin sequences,” Science (80-. ).,
vol. 291, pp. 2603–2605, 2001.
[36] A. D. Parkhe, S. K. Seeley, K. Gardner, L. Thompson, and R. V Lewis, “Structural
Studies of Spider Silk Proteins in the Fiber,” J. Mol. Recognit., vol. 10, pp. 1–6,
1997.
[37] H. Alexandra, A. Carl, and W. Lynn, “Molecular orientation and two-component
nature of the crystalline fraction of spider dragline silk,” Science (80-. )., vol. 271,
no. January, 1996.
[38] N. Becker et al., “Molecular nanosprings in spider capture-silk threads,” Nat.
Mater., vol. 2, no. April, 2003.
[39] J. M. Gimble, “Adipose tissue-derived therapeutics,” Expert Opin Biol Ther, vol.
3, no. 5, pp. 705–713, 2003.
[40] A. Higuchi, Q. Ling, S. Hsu, and A. Umezawa, “Biomimetic Cell Culture Proteins
as Extracellular Matrices for Stem Cell Differentiation,” ACS, vol. 112, 2012.

55
[41] H. Jin, J. Chen, V. Karageorgiou, G. H. Altman, and D. L. Kaplan, “Human bone
marrow stromal cell responses on electrospun silk fibroin mats,” Biomaterials, vol.
25, pp. 1039–1047, 2004.
[42] Y. Toh, K. Blagovic, and J. Voldman, “Advancing stem cell research with
microtechnologies : opportunities and challenges,” Integr. Biol., vol. 2, no. 7,
2010.
[43] G. Chen, “Substrate topography design and fabrication for osteoblast and dental
pulp stem cells studies,” Tiger Prints, 2011.
[44] S. Oh et al., “Stem cell fate dictated solely by altered nanotube dimension,” PNAS,
vol. 106, no. 7, 2009.
[45] K. Saha et al., “Substrate Modulus Directs Neural Stem Cell Behavior,” Biophys.
J., vol. 95, no. November, pp. 4426–4438, 2008.
[46] A. Solanki, S. T. D. Chueng, P. T. Yin, R. Kappera, M. Chhowalla, and K. B. Lee,
“Axonal alignment and enhanced neuronal differentiation of neural stem cells on
graphene-nanoparticle hybrid structures,” Adv. Mater., vol. 25, no. 38, pp. 5477–
5482, 2013.
[47] S. Winkler and D. L. K. U, “Molecular biology of spider silk,” Mol. Biotechnol.,
vol. 74, pp. 85–93, 2000.
[48] Y. Shu, T. Liu, K. Song, X. Fan, X. Ma, and Z. Ciu, “Adipose-derived stem cell: a
better stem cell than BMSC,” Cell Biochem. Funct., vol. 26, pp. 664–675, 2008.
[49] B. A. Bunnell, M. Flaat, C. Gagliardi, B. Patel, and C. Ripoll, “Adipose-derived

56
stem cells: Isolation, expansion and differentiation,” Methods, vol. 45, no. 2, pp.
115–120, 2008.
[50] A. J. Katz, R. Llull, M. H. Hendrick, and J. W. Futrell, “Emerging approaches to
the tissue engineering of fat.,” Clin Plast Surf, vol. 26, no. 4, pp. 587–603, 1999.
[51] J. M. Gimble, A. J. Katz, and B. A. Bunnell, “Adipose-derived stem cells for
regenerative medicine,” Circ. Res., vol. 100, no. 9, pp. 1249–1260, 2007.
[52] P. G. Di Summa, P. J. Kingham, W. Raffoul, M. Wiberg, G. Terenghi, and D. F.
Kalbermatten, “Adipose-derived stem cells enhance peripheral nerve
regeneration,” J. Plast. Reconstr. Aesthetic Surg., vol. 63, no. 9, pp. 1544–1552,
2010.
[53] R. F. Masgutov et al., “Human adipose-derived stem cells stimulate
neuroregeneration,” Clin. Exp. Med., vol. 16, no. 3, pp. 451–461, 2016.
[54] M. L. Previtera, K. L. Trout, D. Verma, U. Chippada, R. S. Schloss, and N. A.
Langrana, “Fibroblast morphology on dynamic softening of hydrogels,” Ann.
Biomed. Eng., vol. 40, no. 5, pp. 1061–1072, 2012.
[55] K. Bott et al., “The effect of matrix characteristics on fibroblast proliferation in 3D
gels,” Biomaterials, vol. 31, no. 32, pp. 8454–8464, 2010.
[56] C. A. Bashur, L. A. Dahlgren, and A. S. Goldstein, “Effect of fiber diameter and
orientation on fibroblast morphology and proliferation on electrospun poly(d,l-
lactic-co-glycolic acid) meshes,” Biomaterials, vol. 27, no. 33, pp. 5681–5688,
2006.

57
[57] E. Cukierman, “Taking Cell-Matrix Adhesions to the Third Dimension,” Science,
vol. 294, no. 5547. pp. 1708–1712, 2001.
[58] K. M. Yamada and E. Cukierman, “Modeling Tissue Morphogenesis and Cancer
in 3D,” Cell, vol. 130, no. 4, pp. 601–610, 2007.
[59] P. Friedl and E. B. Brocker, “The biology of cell locomotion within three-
dimensional extra cellular matrix,” Cell Mol. Life Sci., vol. 57, no. 1, pp. 41–64,
2000.
[60] X. Shen et al., “Regulation of the biological functions of osteoblasts and bone
formation by Zn-incorporated coating on microrough titanium,” ACS Appl. Mater.
Interfaces, vol. 6, no. 18, pp. 16426–16440, 2014.
[61] J. Venugopal, S. Low, A. T. Choon, T. S. Sampath Kumar, and S. Ramakrishna,
“Mineralization of osteoblasts with electrospun collagen/hydroxyapatite
nanofibers,” J. Mater. Sci. Mater. Med., vol. 19, no. 5, pp. 2039–2046, 2008.
[62] S. Weiner and W. Traub, “Crystal size and organization in bone,” Connect Tissue
Res, vol. 21, no. 1–4, pp. 259–265, 1989.
[63] W. J. Landis, M. J. Song, A. Leith, L. McEwen, and B. F. McEwen, “Mineral and
organic matrix interaction in normally calcifying tendon visualized in three
dimensions by high-voltage electron microscopic tomography and graphic image
reconstruction.,” J. Struct. Biol., vol. 110, no. 1, pp. 39–54, 1993.
[64] P. A. Selden and C. Shih, “A golden orb-weaver spider ( Araneae : Nephilidae :
Nephila ) from the Middle Jurassic of China,” Biol. Lett., no. April, pp. 775–778,

58
2011.
[65] R. W. Work, “The Force-Elongation Behavior of Web Fibers and Silks Forcibly
Obtained from Orb-Web-Spinning Spiders,” Text. Res. J., vol. 46, 1976.
[66] L. Römer and T. Scheibel, “The elaborate structure of spider silk: Structure and
function of a natural high performance fiber,” Prion, vol. 2, no. 4, pp. 154–161,
2008.
[67] P. Journal, M. Science, R. March, A. May, and P. August, “Evaluation of the
Resistance of Spider Silk to Ultraviolet Irradiation,” Polym. J., vol. 36, no. 8, pp.
623–627, 2004.
[68] W. S. Rasband, “ImageJ,” U.S. Natl. Institutes Heal. Bethesda, Maryland, USA.
[69] C. Craig, C. Riekel, M. Herberstein, R. Weber, D. L. Kaplan, and N. Pierce,
“Evidence for Diet Effects on the Composition of Silk Proteins Produced by
Spiders,” Mol Biol Evol, vol. 17, no. 12, pp. 1904–1913, 2000.
[70] M. M. Moisenovich et al., “Recombinant 1F9 Spidroin Microgels,” Biochem.
Biophys. Mol. Biol., vol. 466, no. 1, pp. 105–108, 2016.
[71] A. Bednarek, M. Sawadro, and A. Babczyńska, “Modulation of the response to
stress factors of Xerolycosa nemoralis (Lycosidae) spiders living in contaminated
environments,” Ecotoxicol. Environ. Saf., vol. 131, pp. 1–6, 2016.
[72] J. F. McCarthy and L. R. Shugart, “Biological Markers of Environmental and
Ecological Contamination,” Risk Anal., vol. 12, no. 3, pp. 353–360, 1992.
[73] Kammenga, “Progress Report 1995 of BIOPRINT.Biochemical fingerprint

59
techniques as versatile tools for the risk assessment of chemicals in terrestrial
invertebrates.,” Minist. Environ. Energy, vol. 40, pp. 5–6, 1996.
[74] I. M. Tso, S. Y. Chiang, and T. A. Blackledge, “Does the giant wood spider
Nephila pilipes respond to prey variation by altering web or silk properties?,”
Ethology, vol. 113, no. 4, pp. 324–333, 2007.
[75] S. J. Blamires et al., “Mechanical Performance of Spider Silk Is Robust to
Nutrient-Mediated Changes in Protein Composition,” Biomacromolecules, vol. 16,
no. 4, pp. 1218–1225, 2015.
[76] K. B. Guess and C. Viney, “Thermal analysis of major ampullate ( drag line )
spider silk : the effect of spinning rate on tensile modulus 1,” Thermomechanics
Acta, vol. 315, pp. 61–66, 1998.
[77] L. Lin, D. Edmonds, and F. Vollrath, “Structural engineering of a orb-spider’s
web,” Nature, vol. 373, pp. 146–148, 1995.
[78] Z. Shao and F. Vollrath, “The effect of solvents on the contraction and mechanical
properties of spider silk,” Polymer (Guildf)., vol. 40, pp. 1799–1806, 1999.
[79] G. Holland, J. Jenkins, M. Creager, R. V Lewis, and J. Yarger, “Quantifying the
fraction of glycine and alanine in β-sheet and helical conformations in spider
dragline silk using solid-state NMR,” Chem Commun, no. 43, pp. 5568–5570,
2009.
[80] L. Moroni, R. Licht, J. De Boer, J. R. De Wijn, and C. A. Van Blitterswijk, “Fiber
diameter and texture of electrospun PEOT / PBT scaffolds influence human

60
mesenchymal stem cell proliferation and morphology , and the release of
incorporated compounds,” Biomaterials, vol. 27, pp. 4911–4922, 2006.
[81] B. D. Lawrence, J. K. Marchant, M. A. Pindrus, F. G. Omenetto, and D. L. Kaplan,
“Silk film biomaterials for cornea tissue engineering,” Biomaterials, vol. 30, pp.
1299–1308, 2009.
[82] M. M. Moisenovich et al., “Tissue regeneration in vivo within recombinant
spidroin 1 scaffolds,” Biomaterials, vol. 33, no. 15, pp. 3887–3898, 2012.
[83] B. B. Mandal, J. K. Mann, and S. C. Kundu, “Silk fibroin / gelatin multilayered
films as a model system for controlled drug release,” Eur. J. Pharm. Sci., vol. 37,
pp. 160–171, 2009.
[84] V. Hauptmann, N. Weichert, M. Rakhimova, and U. Conrad, “Spider silks from
plants - A challenge to create native-sized spidroins,” Biotechnol. J., vol. 8, no. 10,
pp. 1183–1192, 2013.
[85] V. G. Bogush et al., “Erratum to: ‘Recombinant analogue of Spidroin 2 for
biomedical materials,’” Dokl. Biochem. Biophys., vol. 443, no. 1, pp. 130–130,
2012.
[86] U. J. Kim, J. Park, C. Li, H. J. Jin, R. Valluzzi, and D. L. Kaplan, “Structure and
properties of silk hydrogels,” Biomacromolecules, vol. 5, no. 3, pp. 786–792,
2004.
[87] A. Sponner, B. Schlott, F. Vollrath, E. Unger, F. Grosse, and K. Weisshart,
“Characterization of the Protein Components of Nephila cla V ipes Dragline Silk

61
†,” Biochemistry, vol. 44, no. 12, pp. 4727–4736, 2005.
[88] X. Xia, Z. Qian, C. Seok, Y. Hwan, D. L. Kaplan, and S. Yup, “Native-sized
recombinant spider silk protein produced in metabolically engineered Escherichia
coli results in a strong fiber,” PNAS, vol. 107, no. 32, pp. 14059–14063, 2010.
[89] A. Teplenin et al., “Functional analysis of the engineered cardiac tissue grown on
recombinant spidroin fiber meshes,” PLoS One, vol. 10, no. 3, pp. 1–15, 2015.
[90] C. A. Peng, J. Russo, C. Gravgaard, H. McCartney, W. Gaines, and W. R. J.
Marcotte, “Spider silk-like proteins derived from transgenic Nicotiana tabacum,”
Transgenic Res, vol. 25, no. 4, 2016.
[91] C. A. Peng, J. Russo, T. A. Lyda, and W. R. J. Marcotte, “Polyelectrolyte fiber
assembly of plant-derived spider silk-like proteins,” Biomacromolecules (In Rev.,
2017.
[92] C. A. Peng, J. Russo, T. A. Lyda, and W. R. J. Marcotte, “Polyelectrolyte Fiber
Assembly of Plant-Derived Spider Silk-like Proteins,” Biomacromolecules, vol.
18, no. 3, pp. 740–746, 2017.
[93] M. Widhe et al., “Biomaterials Recombinant spider silk as matrices for cell
culture,” Biomaterials, vol. 31, no. 36, pp. 9575–9585, 2010.
[94] M. Lewicka, O. Hermanson, and A. U. Rising, “Recombinant spider silk matrices
for neural stem cell cultures,” Biomaterials, vol. 33, no. 31, pp. 7712–7717, 2012.
[95] E. Bini et al., “RGD-Functionalized Bioengineered Spider Dragline Silk
Biomaterial,” Biomacromolecules, vol. 7, no. 11, pp. 3139–3145, 2006.

62
[96] M. Pittenger and M. Mackay, “Multilineage potential of adult human
mesenchymal stem cells.,” Science (80-. )., vol. 284, no. 5411, pp. 143–7, 1999.
[97] K. Alberti et al., “Functional immobilization of signaling proteins enables control
of stem cell fate.,” Nature methods, vol. 5, no. 7. pp. 645–50, 2008.
[98] A. Higuchi, Q. Ling, Y. Chang, S. Hsu, and A. Umezawa, “Physical Cues of
Biomaterials Guide Stem Cell Differentiation Fate,” ACS, vol. 113, pp. 3297–
3328, 2013.