ethanol inhibits excitatory neurotransmission in the nucleus accumbens of adolescent mice through...
TRANSCRIPT

Ethanol inhibits excitatory neurotransmission in thenucleus accumbens of adolescent mice throughGABAA and GABAB receptors
Devesh Mishra & Karima CherguiThe Karolinska Institute, Department of Physiology and Pharmacology, Section of Molecular Neurophysiology, Sweden
ABSTRACT adb_350 1..9
Age-related differences in various acute physiological and behavioral effects of alcohol have been demonstrated inhumans and in other species. Adolescents are more sensitive to positive reinforcing properties of alcohol than adults,but the cellular mechanisms that underlie such a difference are not clearly established. We, therefore, assessed agedifferences in the ability of ethanol to modulate glutamatergic synaptic transmission in the mouse nucleus accumbens(NAc), a brain region importantly involved in reward mechanisms. We measured field excitatory postsynapticpotentials/population spikes (fEPSP/PS) in NAc slices from adolescent (22–30 days old) and adult (5–8 months old)male mice. We found that 50 mM ethanol applied in the perfusion solution inhibits glutamatergic neurotransmissionin the NAc of adolescent, but not adult, mice. This effect is blocked by the gamma-aminobutyric acid (GABA)A receptorantagonist bicuculline and by the GABAB receptor antagonist CGP 55845. Furthermore, bicuculline applied aloneproduces a stronger increase in the fEPSP/PS amplitude in adult mice than in adolescent mice. Activation of GABAA
receptors with muscimol produces a stronger and longer lasting depression of neurotransmission in adolescent mice ascompared with adult mice. Activation of GABAB receptors with SKF 97541 also depresses neurotransmission morestrongly in adolescent than in adult mice. These results demonstrate that an increased GABA receptor functionassociated with a reduced inhibitory tone underlies the depressant action of ethanol on glutamatergic neurotransmis-sion in the NAc of adolescent mice.
Keywords Alcohol, field excitatory postsynaptic potentials, GABAA receptor, GABAB receptor, glutamate, nucleusaccumbens.
Correspondence to: Karima Chergui, The Karolinska Institute, Department of Physiology and Pharmacology, Section of Molecular Neurophysiology, VonEulers väg 8, 171 77 Stockholm, Sweden. E-mail: [email protected]
INTRODUCTION
Heavy alcohol consumption in adolescents is a majorpublic health problem in many societies. During adoles-cence, neurochemical remodeling of brain areas involvedin emotions, learning, decision-making and reward-motivated behaviors occur and may alter the sensitivityof adolescents to some of alcohol effects (Spear 2002;Clark, Thatcher & Tapert 2008; Doremus-Fitzwater, Var-linskaya & Spear 2010). Thus, several differences in thephysiological and behavioral effects of alcohol have beendemonstrated in humans and in rodent models. Forexample, adolescents are more sensitive to the rewardingproperties of alcohol than adults (Spear 2002; Philpot,Badanich & Kirstein 2003; Hefner & Holmes 2007; Clarket al. 2008; Doremus-Fitzwater et al. 2010; Spear &
Varlinskaya 2010). The cellular mechanisms that under-lie age-related alterations in brain functions produced byalcohol are not clearly determined. The nucleus accum-bens (NAc), and its glutamatergic inputs from the pre-frontal cortex, amygdala and hippocampus, play animportant role in motivation and reward mechanisms,and mediate alcohol reinforcement (Vengeliene et al.2008). Modulation of excitatory neurotransmission inthe NAc is involved in the reinforcing effects of severaladdictive substances (Hyman, Malenka & Nestler 2006;Vengeliene et al. 2008). Unfortunately, the ability ofalcohol to modulate excitatory neurotransmission in theNAc, the potential age-related differences and the under-lying mechanisms of ethanol action have not been exten-sively studied. We, therefore, examined and comparedthe effect of acute ethanol on glutamatergic synaptic
PRECLINICAL STUDY
Addiction Biologydoi:10.1111/j.1369-1600.2011.00350.x
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

transmission in the core region of the NAc of adolescentand adult mice.
MATERIALS AND METHODS
Experiments using mice were approved by our localethical committee (Stockholms norra djurförsöksetiskanämnd) and were performed as described previously(Schotanus & Chergui 2008b). Male C57BL/6 mice aged22–30 days (adolescents) and 5–8 months (adults)(Laviola et al. 2003) were anesthetized by inhalation ofisoflurane and underwent cervical dislocation followed bydecapitation under anesthesia. Their brains were rapidlyremoved and coronal brain slices (400 mm thick), con-taining the NAc, striatum and the overlying cortex wereprepared with a microslicer (VT 1000S, Leica Microsys-tem, Heppenheim, Germany). Slices were incubated for atleast 1 hour, at 32°C in oxygenated (95% O2 + 5% CO2)artificial cerebrospinal fluid (aCSF) containing (in mM)126 NaCl, 2.5 KCl, 1.2 NaH2PO4, 1.3 MgCl2, 2.4 CaCl2,10 glucose and 26 NaHCO3, pH 7.4.
Slices were transferred to a recording chamber(Warner Instruments, Hamden, CT) mounted on anupright microscope (Olympus, Solna, Sweden) and werecontinuously perfused with oxygenated aCSF at 28°C.Extracellular field recordings were obtained using a glassmicropipette filled with aCSF positioned on the slicesurface. These synaptic responses were evoked by singlestimulation pulses applied every 15 seconds to the brainslice through a concentric bipolar stimulating electrode(FHC, Bowdoinham, ME) placed near the recording elec-trode. Signals were amplified 500 times via an Axopatch200B amplifier (Axon Instruments, Foster City, CA),acquired at 10 kHz, filtered at 2 kHz and recorded on a Dellcomputer (Stockholm, Sweden) using acquisition anddata analysis software from Axon Instruments (pClamp9).
Ethanol (95%, Solveco AB, Stockholm, Sweden) wasdiluted in aCSF to a final concentration of 20, 50 or100 mM. All other chemicals and drugs were purchasedfrom Sigma and Tocris Bioscience (Bristol, UK). Drugswere applied in known concentrations in the perfusionsolution by switching a three-way tap. Numerical valuesare expressed as means � standard error of the mean(SEM), with n indicating the number slices tested. Statis-tical significance of the results was assessed by using thetwo-tailed Student’s t-test for paired and for unpairedobservations and by using the two-way analysis of vari-ance followed by the Bonferroni post-test.
RESULTS
Extracellular field recordings in the NAc
To examine the modulation by ethanol of glutamatergicsynaptic transmission in the NAc, we made extracellular
field recordings in the core region of the NAc in acutebrain slices, as illustrated in Fig. 1a. As described inearlier studies (Pennartz, Boeijinga & Lopes da Silva1990; Nicola, Kombian & Malenka 1996; Schotanus &Chergui 2008a,b,c; Schotanus, Fredholm & Chergui2006), brief (0.1 ms duration) single pulse electricalstimulation of cortico-accumbens slices elicited a charac-teristic biphasic response, which follows the stimulusartifact, with two negative components (Fig. 1b). The firstnegative component, termed N1, is a non-synapticcompound action potential, which is independent ofglutamate release and activation of glutamate receptors.N1 is generated by firing activity in afferent and efferentfibers and in neurons in the NAc. The second negativecomponent, called N2 or population spike (PS) in somestudies, is mediated by endogenous glutamate releasedupon electrical stimulation of glutamate-containingfibers present in the slice. N2 is mediated by the a-amino-3-hydroxyl-5-methyl-4-isoxazolepropionic acid (AMPA)type of glutamate receptors, as demonstrated by theblockade of this component by the AMPA receptorantagonist 6-Cyano-7-nitroquinoxaline-2,3-dione (notshown, but see Schotanus et al. 2006). Thus, N2 reflectsexcitatory monosynaptic transmission in a population ofneurons in the NAc. We, therefore, termed this compo-nent ‘field excitatory postsynaptic potential/populationspike’ (fEPSP/PS) (Buckby & Lacey 2001; Schotanus et al.2006). A stimulus/response (input/output) curve wasestablished for each slice, at the beginning of the record-ing session, by measuring the amplitude of the fEPSP/PSevoked by increasing stimulation intensities. The ampli-tude of the fEPSP/PS was dependent on the intensity ofthe stimulation applied to the slice. Input/output curveswere similar in adolescent and in adult mice (Fig. 1b).
Effect of ethanol on fEPSP/PS amplitude in the NAc ofadolescent and adult mice
fEPSP/PSs were evoked by single stimulation pulsesapplied at an intensity (between 27 mA and 34 mA) yield-ing half maximal response, as assessed by an input/output curve (Fig. 1b). Once a stable baseline of thefEPSP/PS amplitude was obtained, ethanol was applied inthe perfusion solution for 20 minutes. At 20 mM, ethanoldid not produce a significant effect on the amplitude ofthe fEPSP/PS (% of baseline: 95.8 � 5.4, n = 6 slices) inthe NAc of adolescent mice. At 50 mM, ethanoldecreased the amplitude of the fEPSP/PS in adolescentmice (to 78.6 � 1.7% of baseline, n = 23 slices, Fig. 2a &d). In eight slices, we used a paired pulse protocol (twosingle stimulation pulses applied with a 20-ms interval)and found that the ratio between the second fEPSP/PSand the first fEPSP/PS was significantly increased to117.4 � 3.4% of baseline (P < 0.001, Fig. 2b). This
2 Devesh Mishra & Karima Chergui
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

result confirms previous findings that ethanol depressesglutamatergic synaptic transmission in the NAc througha presynaptic mode of action (Nie et al. 1993; Siggins,Roberto & Nie 2005). In adult mice, the depressant actionof 50 mM ethanol on the fEPSP/PS amplitude wasreduced (92.8 � 3.1% of baseline, n = 10, P < 0.001 ascompared with the effect of ethanol in adolescent mice,Fig. 2c & d). To determine whether the effect of ethanolin adolescent mice was concentration-dependent andwhether a higher concentration of ethanol could inhibitglutamatergic neurotransmission in adult mice, weexamined the effect of 100 mM ethanol on fEPSP/PSamplitude. At this concentration, the depressant effect ofethanol was significantly reduced in adolescent mice (%of baseline: 88.9 � 3.2, n = 7, P < 0.05 as comparedwith the effect of 50 mM ethanol). In adult mice,100 mM ethanol did not produce a stronger synapticdepression than that seen at 50 mM (% of baseline:93.4 � 2.3, n = 8, Fig. 2d). Thus, when applied at a phar-macologically relevant concentration (50 mM), ethanol
inhibits glutamatergic synaptic transmission morestrongly in the NAc of adolescent than of adult mice.
Effect of gamma-aminobutyric acid (GABA)A andGABAB receptor antagonists
We sought to investigate the cellular mechanism thatcould underlie this age-related difference in the effect of50 mM ethanol. Several lines of evidence stronglysuggest that an increased GABAergic inhibition mediatesthe acute behavioral and cognitive effects of ethanol(Weiner & Valenzuela 2006; Vengeliene et al. 2008). We,therefore, examined the involvement of GABA in theobserved depressant action of ethanol on glutamatergicneurotransmission in the NAc. In the presence of eitherthe GABAA receptor antagonist bicuculline (5 mM,Fig. 3a) or the GABAB receptor antagonist CGP 55845(1 mM, Fig. 3b), ethanol did not decrease the amplitudeof the fEPSP/PS in the NAc of adolescent mice(92.9 � 1.7% of baseline, n = 9 for bicuculline, and
fEPSP/PS
0 10 20 30 40 50 60 70 800.0
0.2
0.4
0.6
0.8
1.0 AdultAdolescent
Stimulation intensity (µA)
fEP
SP
/PS
am
pli
tud
e (m
V)
N1
3 ms
0.2 mV
3 ms
Artifact
(b)
20 µA 80 µA
0.2 mV
(a)
Figure 1 Extracellular field recordings in the nucleus accumbens (NAc). Glutamatergic synaptic transmission was examined in the mouseNAc by recording extracellular field excitatory postsynaptic potentials/population spikes (fEPSP/PSs) evoked by electrical stimulation ofglutamatergic fibers in brain slices. (a) Schematic of a brain slice containing the NAc.The tips of the stimulating and recording electrodes wereplaced close to each other below the anterior commissure (AC), in the core part of the NAc (AcbC), as indicated by the gray area. (b) Upperpanel shows records of responses measured in one slice and evoked by single stimulation pulses at two intensities: 20 mA and 80 mA.Extracellular responses are composed of two negative components that follow the stimulus artifact: N1 is a non-synaptic component, thesecond component has a synaptic origin and is termed fEPSP/PS. For purposes of clarity, the stimulus artifact is removed from the tracespresented in all the other figures. Graph illustrates averaged input/output curves, i.e. the amplitude of the fEPSP/PS (mean � SEM) evoked byincreasing stimulation intensities of single stimulation pulses, measured in n = 16 and n = 5 slices from adolescent (open squares) and adult (filledcircles) mice, respectively
Ethanol and neurotransmission inhibition 3
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

96.5 � 1.5% of baseline, n = 6 for CGP 55845,P < 0.001 versus control slices with ethanol alone).Thus, ethanol inhibits neurotransmission in the NAc ofadolescent mice through the activation of GABAA andGABAB receptors.
To determine whether the basal GABAergic tone dif-fered between adolescent and adult mice, we comparedthe effect of bicuculline on the fEPSP/PS amplitudebetween adolescent and adult mice. As observed in previ-ous studies (Buckby & Lacey 2001; Schotanus & Chergui2008a), bicuculline (5 mM) applied alone, for 25minutes, increased the amplitude of the fEPSP/PS in theNAc of adolescent and adult mice. However, we foundthat this effect was significantly (P < 0.05) more pro-nounced in adult (% of baseline: 184.5 � 17.6, n = 7)
than in adolescent (% of baseline: 141.3 � 8.5, n = 7,Fig. 3c) mice. CGP 55845 (1 mM) had no effect on thefEPSP/PS amplitude in adolescent (n = 6 slices) and inadult (n = 5 slices) mice (Fig. 3d). These results suggestthat spontaneously released GABA acting on GABAA
receptors more strongly regulates glutamatergic neu-rotransmission in the NAc of adult mice than of adoles-cent mice.
Effect of GABAA and GABAB receptor agonists
We then tested whether activation of GABAA and GABAB
receptors differentially modulates glutamatergic neu-rotransmission in adolescent as compared with adultmice by examining the effect of the GABAA receptor
5 ms
0.4 mV
Ethanol (50 mM)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120
Adolescent
23
1
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120Ethanol (50 mM)
Adult
2 31
Time (minutes)
fEP
SP
/PS
am
pli
tud
e(%
of
bas
elin
e)
-20 -10 0 10 20 30 40 50 600.5
0.6
0.7
0.8
0.9
1.0Ethanol (50 mM)
Adolescent
Time (minutes)
PP
R
###
50 mM 100 mM50
60
70
80
90
100
110
120AdolescentAdult
#
**** *
*
fEP
SP
/PS
am
pli
tud
e(%
of
bas
elin
e)
Baseline (1) Ethanol (2) Washout (3) Baseline (1) Ethanol (2) Washout (3)
Ethanol
Baseline
fEPSP/PS 1 fEPSP/PS 2
0.2 mV5 ms 5 ms
0.2 mV
(a)
(d)
(c)
(b)
Figure 2 Ethanol produces a stronger inhibition of glutamatergic synaptic transmission in the nucleus accumbens (NAc) of adolescent miceas compared with adult mice. Graphs show the time course of the effect of ethanol (50 mM), applied in the perfusion solution at the timeindicated by the horizontal bars, on the amplitude of the fEPSP/PS (mean � SEM) in adolescent (a) and adult (c) mice and on the paired-pulseratio (PPR = ratio between the second fEPSP/PS and the first fEPSP/PS in a paired-pulse stimulation protocol) in adolescent mice (b).Representative records of fEPSP/PS measured in different slices, at the time points indicated on the graphs, are illustrated above each graph.(d) Bar graph illustrates the maximal depressant effect of ethanol (50 mM and 100 mM) measured in individual slices in adolescent and in adultmice. *P < 0.05, ***P < 0.001 (paired t-test), #P < 0.05, ###P < 0.001 (analysis of variance)
4 Devesh Mishra & Karima Chergui
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

agonist muscimol, and of the GABAB receptor agonistSKF 97541 on glutamatergic synaptic transmission inthe NAc. Muscimol (0.5 mM), applied for 20 minutes,decreased the amplitude of the fEPSP/PS in the NAc ofadolescent mice to 72.2 � 2.1% of baseline (measured atthe end of the muscimol application, ‘early’ time point,n = 9, Fig. 4a & d). Muscimol produced a significantlysmaller depression of the fEPSP/PS amplitude in adultmice (82.6 � 2.7%, n = 8, P < 0.05 as compared withthe early effect of muscimol in adolescent mice, Fig. 4c& d). In addition, the depressant effect of muscimolwas long lasting in adolescent mice (depression to74.9 � 3.8%, n = 9, 1 hour after the start of perfusionwith muscimol, ‘late’ time point, Fig. 4a & d). In an addi-tional five slices, we recorded the fEPSP/PS for up to 2hours after washout of muscimol to determine whether arecovery occurred at a later time point. We found that thefEPSP/PS remained depressed in these slices 2 hoursfollowing bath application of muscimol (depressionto 82.8 � 2.6%, n = 5, Fig. 4b). In adult mice, the
amplitude of the fEPSP/PS rapidly returned to baselinevalues following muscimol (0.5 mM) application(106.8 � 3.1%, n = 8, P < 0.001, as compared with thelate effect of muscimol in adolescent mice, Fig. 4c & d). Totest if the difference in the amplitude and duration of theeffect of muscimol between adolescent and adult micewas dependent on the concentration of this agonist, weexamined the effect of 1 mM muscimol on the fEPSP/PSamplitude. We found that the early effect of muscimolwas similar in adolescent (depression to 75.5 � 3.8%,n = 4) and in adult mice (depression to 78.7 � 5.7%,n = 5, Fig. 4d). However, the long lasting effect of 1 mMmuscimol was only observed in adolescent mice (% ofbaseline: 78.1 � 6.4, n = 4, P < 0.05 as compared withadult mice: 101.0 � 4.5, n = 5, Fig. 4d).
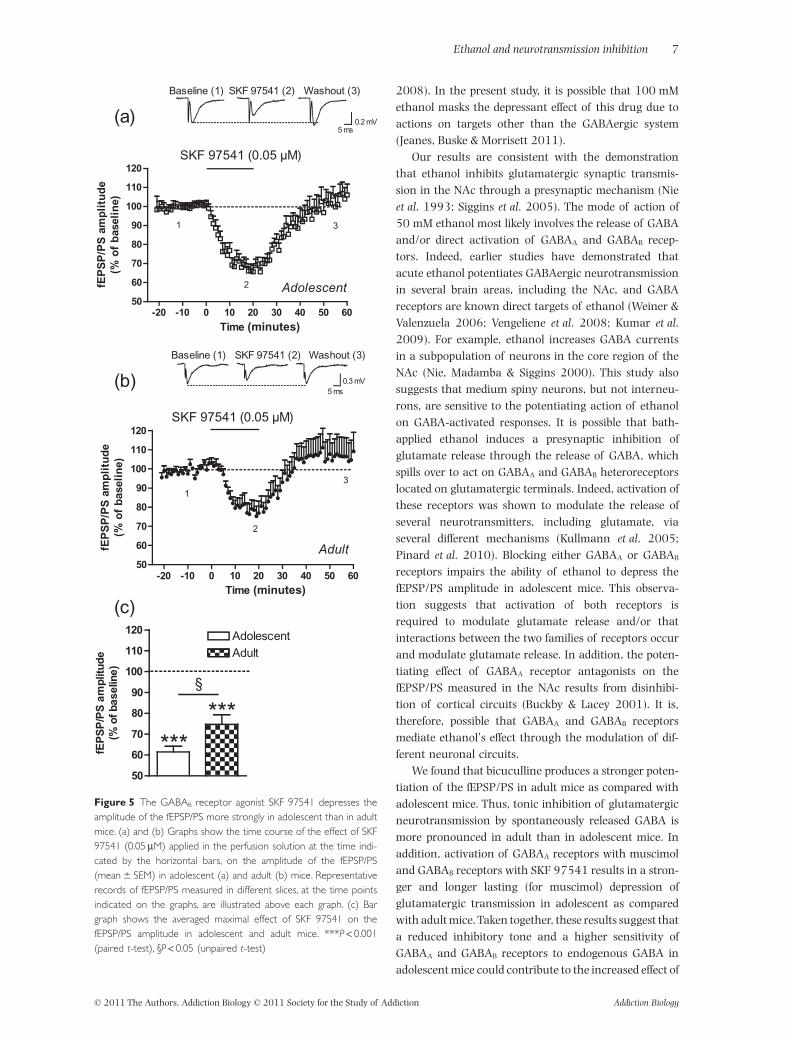
SKF 97541 (0.05 mM) also depressed the amplitude ofthe fEPSP/PS, but this effect was more pronounced inadolescent than in adult mice (61.4 � 2.7%, n = 10,and 74.7 � 4.6% n = 9, respectively, P < 0.05, Fig. 5).These results show that activation of GABAA and GABAB
Ethanol (50 mM)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120
CGP 55845 (1 µM)
Adolescent
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
Ethanol (50 mM)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120
Bicuculline (5 µM)
Adolescent
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
-20 -10 0 10 20 30 40 50 6050
75
100
125
150
175
200
Bicuculline (5 µM)
Adult
Adolescent
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
CGP 55845 (1 µM)
-20 -10 0 10 20 30 40 5050
75
100
125
150AdultAdolescent
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
(a)
(b)
(c)
(d)
Figure 3 Involvement of GABAA and GABAB receptors in the depressant action of ethanol. (a) and (b) The GABAA receptor antagonistbicuculline (5 mM) and the GABAB receptor antagonist CGP 55845 (1 mM) blocked the inhibitory effect of ethanol in adolescent mice.Thesetwo antagonists were applied in the perfusion solution 27–28 minutes before the start of the perfusion with ethanol, in combination withethanol and during ethanol washout, as indicated by the horizontal bar. (c) and (d) Graphs show the time course of the effect of bicuculline(5 mM) and CGP 55845 (1 mM) applied alone on the amplitude of the fEPSP/PS measured in adolescent (open squares) and adult (filledcircles) mice
Ethanol and neurotransmission inhibition 5
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

receptors inhibits glutamatergic synaptic transmission inthe NAc more strongly in adolescent than in adult mice.
DISCUSSION
This study demonstrates that ethanol inhibitsglutamatergic synaptic transmission in the mouse NAc inan age-dependent manner, i.e. depression is more pro-nounced in adolescents than in adults. In addition, thisstudy shows that GABAA and GABAB receptors mediatethe depressant action of ethanol. We also identify an age-dependent inhibitory effect of GABAA and GABAB recep-tor activation associated with a difference in tonicinhibition, which could underlie the larger depressantaction of ethanol seen in adolescent mice as comparedwith adult mice.
The present study examined the effect of ethanol inthe core part of the NAc. Although the precise role of the
core and of the shell parts of the NAc in reward-dependent behaviors is still a matter of debate and inves-tigation (Gonzales, Job & Doyon 2004), excitatoryneurotransmission in the NAc core was shown to play animportant role in the expression of behavioral sensitiza-tion to drugs of abuse (Wolf et al. 2004). Further studiesshould determine whether ethanol differentially affectsglutamatergic neurotransmission in the NAc core versusthe NAc shell. The present study demonstrates that in theNAc core of adolescent mice, the depressant action ofethanol is significant at a pharmacologically relevantconcentration (50 mM), but this effect is reduced at ahigher concentration (100 mM). In humans, a bloodconcentration of ethanol of 50 mM corresponds toaround three to four times that of legal driving limit inseveral countries. Such a concentration produces intoxi-cation, while human lethal blood alcohol concentrationis around 80 mM (Olsen et al. 2007; Vengeliene et al.
Muscimol (2) Washout (3)
0.3 mV5 ms
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120Muscimol (0.5 µM)
Adult
2
31
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
Muscimol (0.5 µM)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120
Adolescent
2 3
1
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e) 5 ms
-20 0 20 40 60 80 100 12050
60
70
80
90
100
110
120Muscimol (0.5 µM)
Adolescent
Time (minutes)
fEP
SP
/PS
am
plit
ud
e(%
of
bas
elin
e)
50
60
70
80
90
100
110
120
AdolescentAdult
#
0.5 µMEarly
0.5 µMLate
1 µMLate
1 µM Early
####
****
*** ***
###
#
*fE
PS
P/P
S a
mp
litu
de
(% o
f b
asel
ine)
0.3 mV
Baseline (1) Muscimol (2) Washout (3) Baseline (1)(a)
(b)
(c)
(d)
Figure 4 The GABAA receptor agonist muscimol produces a long-lasting depression of the amplitude of the fEPSP/PS in adolescent but notin adult mice. (a) to (c) Graphs show the time course of the effect of muscimol (0.5 mM) applied in the perfusion solution at the time indicatedby the horizontal bars, on the amplitude of the fEPSP/PS (mean � SEM) in adolescent (a) and (b), and adult (c) mice. Graphs in (a) and (b)show the results obtained for a 1-hour and a 2-hour washout of muscimol, respectively. Representative records of fEPSP/PS measured indifferent slices, at the time points indicated on the graphs, are illustrated above the graphs. (d) Bar graph summarizes the results of theexperiments in (a) and (c) as well as the effect of 1 mM muscimol in adolescent and adult mice. *P < 0.05, **P < 0.01, ***P < 0.001 (pairedt-test), #P < 0.05, ###P < 0.001 (analysis of variance)
6 Devesh Mishra & Karima Chergui
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology
2008). In the present study, it is possible that 100 mMethanol masks the depressant effect of this drug due toactions on targets other than the GABAergic system(Jeanes, Buske & Morrisett 2011).
Our results are consistent with the demonstrationthat ethanol inhibits glutamatergic synaptic transmis-sion in the NAc through a presynaptic mechanism (Nieet al. 1993; Siggins et al. 2005). The mode of action of50 mM ethanol most likely involves the release of GABAand/or direct activation of GABAA and GABAB recep-tors. Indeed, earlier studies have demonstrated thatacute ethanol potentiates GABAergic neurotransmissionin several brain areas, including the NAc, and GABAreceptors are known direct targets of ethanol (Weiner &Valenzuela 2006; Vengeliene et al. 2008; Kumar et al.2009). For example, ethanol increases GABA currentsin a subpopulation of neurons in the core region of theNAc (Nie, Madamba & Siggins 2000). This study alsosuggests that medium spiny neurons, but not interneu-rons, are sensitive to the potentiating action of ethanolon GABA-activated responses. It is possible that bath-applied ethanol induces a presynaptic inhibition ofglutamate release through the release of GABA, whichspills over to act on GABAA and GABAB heteroreceptorslocated on glutamatergic terminals. Indeed, activation ofthese receptors was shown to modulate the release ofseveral neurotransmitters, including glutamate, viaseveral different mechanisms (Kullmann et al. 2005;Pinard et al. 2010). Blocking either GABAA or GABAB
receptors impairs the ability of ethanol to depress thefEPSP/PS amplitude in adolescent mice. This observa-tion suggests that activation of both receptors isrequired to modulate glutamate release and/or thatinteractions between the two families of receptors occurand modulate glutamate release. In addition, the poten-tiating effect of GABAA receptor antagonists on thefEPSP/PS measured in the NAc results from disinhibi-tion of cortical circuits (Buckby & Lacey 2001). It is,therefore, possible that GABAA and GABAB receptorsmediate ethanol’s effect through the modulation of dif-ferent neuronal circuits.
We found that bicuculline produces a stronger poten-tiation of the fEPSP/PS in adult mice as compared withadolescent mice. Thus, tonic inhibition of glutamatergicneurotransmission by spontaneously released GABA ismore pronounced in adult than in adolescent mice. Inaddition, activation of GABAA receptors with muscimoland GABAB receptors with SKF 97541 results in a stron-ger and longer lasting (for muscimol) depression ofglutamatergic transmission in adolescent as comparedwith adult mice. Taken together, these results suggest thata reduced inhibitory tone and a higher sensitivity ofGABAA and GABAB receptors to endogenous GABA inadolescent mice could contribute to the increased effect of
0.3 mV5 ms
0.2 mV5 ms
SKF 97541 (0.05 µM)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120
Adolescent2
31
Time (minutes)
fEP
SP
/PS
am
pli
tud
e(%
of
bas
elin
e)
SKF 97541 (0.05 µM)
-20 -10 0 10 20 30 40 50 6050
60
70
80
90
100
110
120
Adult
2
3
1
Time (minutes)
fEP
SP
/PS
am
pli
tud
e(%
of
bas
elin
e)
Baseline (1) SKF 97541 (2) Washout (3)
Baseline (1) SKF 97541 (2) Washout (3)
(a)
(b)
50
60
70
80
90
100
110
120 AdolescentAdult
******
§
fEP
SP
/PS
am
plitu
de(%
of b
asel
ine)
(c)
Figure 5 The GABAB receptor agonist SKF 97541 depresses theamplitude of the fEPSP/PS more strongly in adolescent than in adultmice. (a) and (b) Graphs show the time course of the effect of SKF97541 (0.05 mM) applied in the perfusion solution at the time indi-cated by the horizontal bars, on the amplitude of the fEPSP/PS(mean � SEM) in adolescent (a) and adult (b) mice. Representativerecords of fEPSP/PS measured in different slices, at the time pointsindicated on the graphs, are illustrated above each graph. (c) Bargraph shows the averaged maximal effect of SKF 97541 on thefEPSP/PS amplitude in adolescent and adult mice. ***P < 0.001(paired t-test), §P < 0.05 (unpaired t-test)
Ethanol and neurotransmission inhibition 7
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

ethanol as compared with adult mice. Differencesbetween adolescent and adult mice in the pharmacologi-cal and physiological properties, as well as in the level ofexpression of GABAA and GABAB receptors or in theirsubunit composition could also explain our results(Laurie, Wisden & Seeburg 1992). For example, GABAA
receptors containing the delta subunit are highly sensi-tive to GABA and to ethanol, and display continuous acti-vation (Stórustovu & Ebert 2006; Santhakumar, Wallner& Otis 2007). It remains to be determined whether thesubunit composition and/or the level of expression ofGABAA and GABAB receptors differ in the NAc of adoles-cents versus adults and whether these differencesunderlie the increased sensitivity of excitatory neu-rotransmission to acute ethanol in the NAc of adoles-cents. Channel properties of GABAA receptors could beresponsible for the long lasting effect of muscimol in theNAc of adolescent mice described here. Alternatively, thiseffect could underlie a form of long-term synaptic plastic-ity, i.e. long-term depression, as shown previously in thehippocampus (Akhondzadeh & Stone 1995).
Given the involvement of glutamatergic neurotrans-mission in the NAc in reward mechanisms, the effect ofethanol described here in adolescent mice could consti-tute a major mechanism through which alcohol mediatesits reinforcing properties. This study identifies a criticalrole for GABAA and GABAB receptors in ethanol-induced synaptic depression, and suggests a possiblemechanism by which alcohol influences brain functionsin adolescents.
Acknowledgements
This study was supported by the Swedish ResearchCouncil (grant 2008–2636) and by the Alcohol ResearchCouncil of the Swedish Alcohol Retailing Monopoly(SRA).
Financial Disclosures
The authors declare no conflict of interest.
Authors Contribution
DM performed the experiments, analyzed the data andmade the figures. KC supervised the project and wrote themanuscript. All authors critically reviewed content andapproved final version of the manuscript for publication.
References
Akhondzadeh S, Stone T (1995) Induction of a novel form ofhippocampal long-term depression by muscimol: involvementof GABAA but not glutamate receptors. Br J Pharmacol115:527–533.
Buckby LE, Lacey MG (2001) Epileptiform activity in the nucleusaccumbens induced by GABA(A) receptor antagonists in ratforebrain slices is of cortical origin. Exp Brain Res 141:146–152.
Clark DB, Thatcher DL, Tapert SF (2008) Alcohol, psychologicaldysregulation, and adolescent brain development. AlcoholClin Exp Res 32:375–385.
Doremus-Fitzwater TL, Varlinskaya EI, Spear LP (2010) Motiva-tional systems in adolescence: possible implications for agedifferences in substance abuse and other risk-taking behav-iors. Brain Cogn 72:114–123.
Gonzales RA, Job MO, Doyon WM (2004) The role of mesolimbicdopamine in the development and maintenance of ethanolreinforcement. Pharmacol Ther 103:121–146.
Hefner K, Holmes A (2007) An investigation of the behavioralactions of ethanol across adolescence in mice. Psychophar-macology 191:311–322.
Hyman SE, Malenka RC, Nestler EJ (2006) Neural mechanismsof addiction: the role of reward-related learning and memory.Annu Rev Neurosci 29:565–598.
Jeanes Z, Buske T, Morrisett R (2011) In vivo chronic intermit-tent ethanol exposure reverses the polarity of synaptic plastic-ity in the nucleus accumbens shell. J Pharmacol Exp Ther336:155–164.
Kullmann DM, Ruiz A, Rusakov DM, Scott R, Semyanov A,Walker MC (2005) Presynaptic, extrasynaptic and axonalGABAA receptors in the CNS: where and why? Prog BiophysMol Biol 87:33–46.
Kumar S, Porcu P, Werner D, Matthews D, Diaz-Granados J,Helfand R, Morrow A (2009) The role of GABA(A) receptorsin the acute and chronic effects of ethanol: a decade ofprogress. Psychopharmacology 205:529–564.
Laurie D, Wisden W, Seeburg P (1992) The distribution of thir-teen GABAA receptor subunit mRNAs in the rat brain. III.Embryonic and postnatal development. J Neurosci 12:4151–4172.
Laviola G, Macrì S, Morley-Fletcher S, Adriani W (2003) Risk-taking behavior in adolescent mice: psychobiological determi-nants and early epigenetic influence. Neurosci Biobehav Rev27:19–31.
Nicola SM, Kombian SB, Malenka RC (1996) Psychostimulantsdepress excitatory synaptic transmission in the nucleusaccumbens via presynaptic D1-like dopamine receptors. JNeurosci 16:1591–1604.
Nie Z, Madamba SG, Siggins GR (2000) Ethanol enhancesgamma-aminobutyric acid responses in a subpopulation ofnucleus accumbens neurons: role of metabotropic glutamatereceptors. J Pharmacol Exp Ther 293:654–661.
Nie Z, Yuan X, Madamba SG, Siggins GR (1993) Ethanoldecreases glutamatergic synaptic transmission in rat nucleusaccumbens in vitro: naloxone reversal. J Pharmacol Exp Ther266:1705–1712.
Olsen RW, Hanchar HJ, Meera P, Wallner M (2007) GABAAreceptor subtypes: the ‘one glass of wine’ receptors. Alcohol41:201–209.
Pennartz CM, Boeijinga PH, Lopes da Silva FH (1990) Locallyevoked potentials in slices of the rat nucleus accumbens:NMDA and non-NMDA receptor mediated components andmodulation by GABA. Brain Res 529:30–41.
Philpot RM, Badanich KA, Kirstein CL (2003) Place condition-ing: age-related changes in the rewarding and aversive effectsof alcohol. Alcohol Clin Exp Res 27:593–599.
Pinard A, Seddik R, Bettler B, Thomas PB (2010) GABAB recep-tors: physiological functions and mechanisms of diversity. In:
8 Devesh Mishra & Karima Chergui
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology

Blackburn TP, Enna SJ, eds. Advances in Pharmacology, pp.231–255. Maryland Heights, MO: Academic Press.
Santhakumar V, Wallner M, Otis TS (2007) Ethanol acts directlyon extrasynaptic subtypes of GABAA receptors to increasetonic inhibition. Alcohol 41:211–221.
Schotanus SM, Chergui K (2008a) Dopamine D1 receptors andgroup I metabotropic glutamate receptors contribute to theinduction of long-term potentiation in the nucleus accum-bens. Neuropharmacology 54:837–844.
Schotanus SM, Chergui K (2008b) Long-term potentiation in thenucleus accumbens requires both NR2A- and NR2B-containing N-methyl-d-aspartate receptors. Eur J Neurosci27:1957–1964.
Schotanus SM, Chergui K (2008c) NR2A-containing NMDAreceptors depress glutamatergic synaptic transmission andevoked-dopamine release in the mouse striatum. J Neurochem106:1758–1765.
Schotanus SM, Fredholm BB, Chergui K (2006) NMDA depressesglutamatergic synaptic transmission in the striatum throughthe activation of adenosine A1 receptors: evidence fromknockout mice. Neuropharmacology 51:272–282.
Siggins GR, Roberto M, Nie Z (2005) The tipsy terminal: presyn-aptic effects of ethanol. Pharmacol Ther 107:80–98.
Spear LP (2002) The adolescent brain and the college drinker:biological basis of propensity to use and misuse alcohol. J StudAlcohol 14:71–81.
Spear LP, Varlinskaya EI (2010) Sensitivity to ethanol and otherhedonic stimuli in an animal model of adolescence: implica-tions for prevention science? Dev Psychobiol 52:236–243.
Stórustovu S, Ebert B (2006) Pharmacological Characterizationof agonists at delta-containing GABAA receptors: functionalselectivity for extrasynaptic receptors is dependent on theabsence of gamma 2. J Pharmacol Exp Ther 316:1351–1359.
Vengeliene V, Bilbao A, Molander A, Spanagel R (2008) Neurop-harmacology of alcohol addiction. Br J Pharmacol 154:299–315.
Weiner JL, Valenzuela CF (2006) Ethanol modulation ofGABAergic transmission: the view from the slice. PharmacolTher 111:533–554.
Wolf ME, Sun X, Mangiavacchi S, Chao SZ (2004) Psychomotorstimulants and neuronal plasticity. Neuropharmacology 47(Suppl 1):61–79.
Ethanol and neurotransmission inhibition 9
© 2011 The Authors, Addiction Biology © 2011 Society for the Study of Addiction Addiction Biology