epoxide hydrolase in the bulb mite rhizoglyphus robini: properties and induction
TRANSCRIPT

Experimental & Applied Acarology, 17 (1993) 381-392 381
Epoxide hydrolase in the bulb mite Rhizoglyphus robini: properties and induction
Ephraim Cohen, Uri Gerson and Rita Mozes-Koch Department of Entomology, Faculty of Agriculture, The Hebrew University of Jerusalem,
Rehovot 76 100, Israel (Accepted 26 October 1992)
ABSTRACT
Cohen, E., Gerson, U. and Mozes-Koch, R., 1993. Epoxide hydrolase in the bulb mite Rhizoglyphus robini: properties and induction, 1993. Exp. Appl. Acarol. 17: 381-392.
Using styrene oxide as substrate, most of the epoxide hydrolase (EH) activity monitored in the bulb mite Rhizogt)phtt~ robini was associated with the microsomal compartment. The microsomal and cytosolic EHs did not display any significant preference in hydrating trans stilbene oxide (TSO) and cis stilbene oxide (CSO). The microsomal EH, which has a Km'value of 5×IO-~M and pH optimum of 7.8, was sensitive to ethanol and its activity was inhibited to a moderate extent by 4-fluorochal- cone oxide, TSO, CSO and trans-chalcone oxide at a level of IO-4M. Microsomal EH was consider- ably induced (4--5-fold) in mites feeding garlic and onion, or ingesting TSO-impregnated filter papers. Other epoxides like CSO, 2,4-dichlorostilbene oxide, methyl chaicone oxide and heptachlor epoxide displayed moderate induction levels (1.4-2.6-fold). Of the toxicants assayed only sodium phenobar- bital was a potent inducer. Lindane, malathion and DDT did not stimulate EH activity and 3-methyl- cholanthrene was even inhibitory. A decrease in EH activity was observed with a number of phyto- chemicals tested such as sinigrin, flavone, menthol, trans-[5-carotene, chalcone, aUyl sulphide and trans-cinnarrfic acid.
INTRODUCTION
Bioconversion of epoxides catalyzed by enzymatic hydration of the oxirane ring is widespread and has been recognized as an important step in metabolism of endo- genous substrates as well as in the detoxification of xenobiotic compounds (Brooks, 1977; Mullin, 1988; Oesch, 1972). In addition to the membrane-bound microso- mat epoxide hydrolase (EH) (EC 3.3.2.3), a cytosolic enzyme with different prop- erties was also studied in mammalian and arthropod systems (Mullin et al., 1984; Wixtrom and Hammock, 1985). Similar to the other detoxifying systems such as microsomal monooxygenases, esterases and conjugating enzymes, the EHs are also induced by a variety of synthetic compounds, toxicants, and by phytochemi- cats (DePierre et al., 1984; Hammock and Ota, 1983; Meijer et al., 1982; Mullin, 1988).
© Science and Technology letters. All rights reserved.

382 E. COHl~ El" AI..
Detoxifying enzyme systems including EH and their induction were studied in mites. Higher trans EH (microsomal) and cis EH (cytosolic) activities were detected, correspondingly, in the two-spotted spider mite Tetranychus urticae and in the predatory Amblyseiusfallacis (Mullin et al., 1982). It has been speculated that herbivory in arthropods is associated with elevated microsomal EH activity (Mullin and Croft, 1984).
The easily mass-reared bulb mite Rhizoglyphus robini has been depicted as a suitable model acarine to investigate a variety of biological, physiological and bio- chemical aspects (Gerson et al.,-1991). Studies related to activities, inhibition and induction of general esterases (Capua et aL, 1990), aldrin epoxidase and glutathione S-transferase (GST) (Capua et aL, 1991) have already been carried out. This study focuses on R. robini EH, emphasizing induction of the enzyme after feeding on plant materials, various toxicants and plant allekxzhemicals.
MATERIALS AND METHODS
ChemicaL~ The radioactive substrate [7-3H] styrene oxide with a specific activity o f 99m Ci mmol -I was supplied by Amersham (England). 3H-trans stilbene oxide (TSO) and 3H-cis stilbene oxide (CSO), 4-fluorochalcone oxide, 4-methylchalcone oxide and chalcone oxide were synthesized and kindly provided by Bruce D. Hammock from the University of California, Davis. AUyl sulphide, trans-cinnamic acid, flavone, trans-~-carotene, isooctane (2,2,4-tri-methylpentane), menthol, sin- igrin, cis-stilbene oxide, trans-stilbene oxide, styrene oxide and Tween 80 were purchased from Sigma (St. Louis, Mo). Chalcone, chalcone oxide, trans-chloro- stilbenc oxide, 2,4-dichlorostilbene oxide and 3-methylcholanthrene were supplied by Aldrich (Milwaukee, Wisc.). Sodium phenobarbital was a product ofJ. T. Baker Chemicals (Phillipsburg, N. J.). 4-Methoxy trans-stilbene oxide was synthesized by reacting trans-4-hydroxy-stilbene oxide with sodium hydroxide followed by methylation with methyl iodide, p,p'-DDT, heptachlor epoxide, lindane (hexachlorocyclo-hexane, "c-isomer) and malathion were pure standards. All other chemicals used were of analytical grade.
The bulb mite Bulb mite Rhizoglyphus robini Clapartde (Astigmata, Acaridae) colonies were maintained in Petri plates on a peanut paste diet at 27°C as described by Gerson et aL (1991). Crowded conditions and extensive diet consumption forced the mites to migrate out of the Petri plates. This behavior facilitated massive collection of mites to be used for the various experiments. The parasitic mite Varroajacobsoni Oudemans (Mesostigmata, Varroidae) were collected from infested colonies of Italian honey bees (Apis mellifera L.). Experimental bee hives for this purpose were kept at Triwaks Bee Research Center, Faculty of Agriculture, Rehovot. The spider mite Tetranychus urtice Koch (Prostigmata: Tetranychidae) raised on Lima beans,

EPOXIDE HYDROLASE IN THE BULB MITE RHIZOGLYPHUS ROBINI PROPERTIES AND INDUCTION 383
and its predator Phytoseiu!us persimitis Athias-Henriot (Mesostigmata Phyto- seiidae) were obtained from Kibbutz Sde Eliahu Biological Control Insectaries.
EH - preparation Mites were homogenized in 10 vol (approximate volume of 1 mL) of 0.1M sodium phosphate buffer, pH 7.4 containing 1.15% KCI. The homogenization was per- formed by an Ultra-Turrax (2 min) followed by a Teflon-glass homogenizer (2 min). The homogenate was filtered through 4 layers of gauze and the filtrate was centrifuged for 25 min at 12,000 g. The postmitochondrial supernatant was filtered through glass wool and further spun at 4°C for 60 min at 150,000 g to obtain cyto- solic and microsomal fractions. The microsomal pellet was suspended in 50 mM sodium phosphate, pH 7.4 containing 0.58% KCI.
E H - radioassay R. robini EH activity was measured essentially according to the procedure described by Oesch et al. (1971a) with minor modifications. The reaction mixture in a total volume of 100~tL consisted of 70~tL 0.1M Tris-HC1 buffer solution, pH 7.8 containing 0. 1% (v/v) Tween 80, and 25~tL of the enzyme preparation. The reaction was started with 5l.tL 3H-styrene oxide containing 10-3M cold substrate dissolved in ethanol. The final concentration of styrene oxide in the reaction mixture was 5 × 10-SM. The reaction was carried out at 37°C for 10 min in a shaking bath, and stopped by adding 200ktL isooctane. Following vigorous shaking the aqueous and organic phases were separated by low-speed centrifugation at 3,000 g for 10 min. Aliquots (50ktL) of the aqueous phase were removed for radioassay using 3 mL of lnsta Gel II scintillation fluid. Control assays contained boiled enzyme preparations and protein was determined by the Bradford method (Bradford, 1976) using bovine serum albumin as a standard. The specific activity of the mite EH was calculated from the amount of the diol formed according to Wixtrom and Hammock (1985).
Feeding effects Synthetic compounds and phytochemicals: The propensity of R. robini to ingest inert cellulose paper was used in the present study for examining effects of various substances on EH activity. The compounds were dissolved in 150~tL acetone and applied on black filter paper discs (Whatman 29, 5cm in diameter) at a level of 5 mg per disc. After evaporation of the solvent, the paper was placed inside a plastic Petri dish (5 cm in diameter), and 3001aL of distilled water were added. About 100 mg of mites were introduced and the plates transferred to an incubator (28°C, 80%rh) for 48 hours. Small pieces of black filter paper, which could be seen inside the mites, indicated ingestion of the applied compounds. Following the incubation period, the mites were removed from the plates, homogenized and radioassayed as described before. Filter papers treated with distilled water served as controls.

384 E. COI.IEN l-l" At..
Plant materials: Slices of various vegetables purchased from the local super- market were placed inside a Petri plate (5 cm in diameter) into which about 200 mg of mites were introduced. The plates were transferred to an incubator (28°C, 80% rh) for a 48 hours period. The mites wcre assayed for EH activity as previously described.
RESUI.TS
To obtain membrane-free cytosolic fractions of R. robini, post-mitochondrial supernatants were centrifuged twice at 150,000 g. Using styrene oxide as substrate, the specific activity of the microsomal EH was about 5-fold higher as compared to its cytosolic counterpart (Table 1). Radiolabelled trans-stilbenc oxide (TSO) and cis-stilbene oxide (CSO) were used as substrates to successfully differentiate between microsomal and cytosolic rat liver EHs (Wixtrom and Hammock, 1985). In the mammalian system the epoxide ring in TSO is hydrolized much more effcctively by tile microsomal enzyme as compared to the soluble EH, and vise versa with respect to CSO. Unlike the mammalian EHs, the microsomal and cyto- solic cnzymes of the bulb mite hydrolized with similar efficacy both TSO and CSO (Table 1 ). Both compounds, however, were poor substrates as compared to styrene oxide.
Mullin and Croft (1984) argued that higher EH activity is linked to herbivory in arthropods. To examine this hypothesis, microsomal EH activity was measured in four different mite species (Table 2). The highest specific activity value was determined in the predatory mite P. persimilis (2.2 nmol min -t mg -~ microsomal protein) as compared to the parasitic mite V. jacobsoni which had a much lower value (0.44 nmol min -~ mg -1 protein). Intermediate activity values were detected for the phytophagous mite species (R. robini and T. urticae). Information from additional mite species is needed to draw any conclusion regarding the relation- ship between EH activity and feeding habits of mites.
TABLE 1
Activity of Rhizoglyphus robini microsomal and cytosolic epoxide hydrolam (Ell) using different substrates.
EH activity (nmol styrene diol min J mg -I protein)
Substrate Microsomes Cytosol
q l-styrene oxide 3.26x'-0.6 101)% 0.65+0.1 100% 3H-trans-stilbene oxide 0.77-+0.1 23.6% 0.25+0.03 38.5% SH-cis-stilbene oxide 0.74+_0.2 22.7% 0.16-+0.02 24.6%
Using styrene oxide as substrate the activity of the cytosolic enzyme was about 20% as compared to it~ microsomal counterpart.
EPOXIDE HYDROLASE IN THE BULB MITE RtlIZOGI.YPHUS R(.)BhVI PROPERTIES AND INDUCTION 385
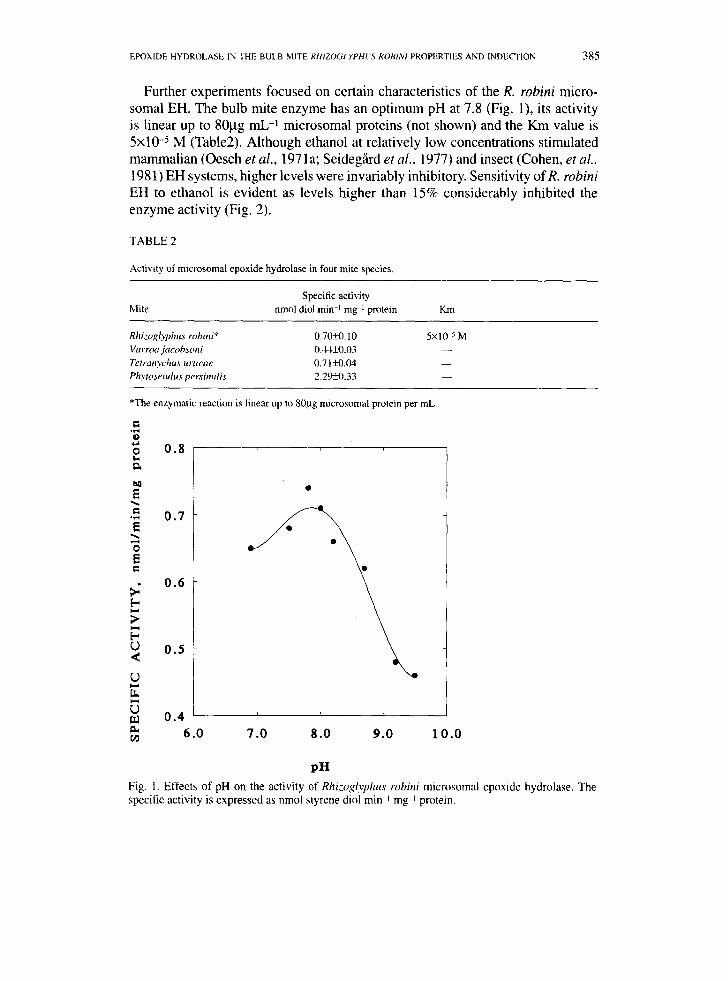
Further experiments focused on certain characteristics of the R. robini micro- somal EH. The bulb mite enzyme has an optimum pH at 7.8 (Fig. 1), its activity is linear up to 80btg mL -1 microsomal proteins (not shown) and the Km value is 5x10 -5 M (Table2). Although ethanol at relatively low concentrations stimulated mammalian (Oesch et al., 1971 a; Seideg~d et al., 1977) and insect (Cohen, et al., 1981) EH systems, higher levels were invariably inhibitory. Sensitivity of R. robini EH to ethanol is evident as levels higher than 15% considerably inhibited the enzyme activity (Fig. 2).
TABLE 2
Activity of microsomal epoxide hydrolase in four mite species.
Specific activity
Mite nmol diol min -t m g i protein Km
Rhizoglyphus robini* 0.70_+0.10 5x I 0 ~ M
Varr~m jacohaoni 0.44£-4,).03 - -
Tetranychus urticae 0.7 t M.).04 - -
Phytoseiulus persimilis 2.295-0.33 - -
*The enzymat ic reaction is linear t,p to 80ug microsomal protein per mL.
ol-,~ 0
0
E ° , "4
ff
t- C3
0 . 8
0.7
0 . 6
0 . 5 -
U
0.4 ' '
6 . 0 7 . 0 8 .0 9 . 0 10.0
pH
Fig. 1. Effects of pH on the activity of Rhizoglyphus robini microsomal epoxide hydrolase. The specilic activity is expressed as nmol styrene diol rain ~ mg i protein.

386 E . C O H E N E T A L .
Inhibition of T. urticae EH by chalcone oxide and 4-methylchaicone oxide (at a level of 0.1 mM) was reported by Mullin et al. (1984). Chalcone oxide and various other epoxides at a concentration of 0.5mM were assayed as to their in vitro effect on R. robini EH activity (Table 3). A considerable inhibitory effect (42%) was observed in particular with 4-fluoro-chalcone oxide. Compared to the above, TSO, CSO and trans-chalcone oxide were less inhibitory while no or slight EH inhibition was observed with 4-methoxystilbene oxide and 2,4-dichlorostilbene oxide.
TABLE 3
In vitro effects of various epoxides on the activity of Rhizoglyphus robini epoxide hydrolase.
Oxide* Activity _LSD** % of control
Control 1.95:L-0.38 100 trans-Stilbene oxide 1.28_+0.58 65.5 cis-Stilbene oxide 1.74i~).22 89.2 4-Methoxystilbebe oxide 1.61_+0.32 82.6 2,4-Dichlorostilbene oxide 1.92-+0.48 98.5 trans-Chalcone oxide 1.55_'!-0.22 79.5 4-Fluorochalcone oxide 1.13:L-0.27 58.0
*Final concentration in th e reaction mixture was 5 x 10 -7 M.
**The specific activity expressed as nmol styrene diol formed per min per mg microsomal protein.
.,..q o ~ 0 . 8
e a
• =- 0 . 6
8 i:1 . 0 . 4
~ 0 . 2
U
~1 0 . 0
i i
_ . . J I I l I
0 5 1 0 1 5 2 0 2 5 3 0
E T H A N O L . %
Fig. 2. Inhibitory effect of ethanol on the activity ofRhizoglyphus robini microsomal epoxide hydro- lase. The specific activity is expressed as nmol styrene diol rain -j mg -I protein.

EPOXIDE HY1)ROLASE IN TIlE BIS1.B MITE RIIIZOGL}7qlUS ROBINI PROPERTIES ANI) INDUCTION 387
Being polyphagous, the bulb mite is able to grow and reproduce on a great number of substrates (Gerson et aL, 1991). Fig. 3 depicts the effects of various vegetables on the bulb mite EH activity. Similar to what was observed with GST (Capua et al., 1991), garlic and onion greatly induced EH activity by 4 and 5 fold respectively. Considerable induction of activity was also observed with carrot and zucchini. Slight or no induction was evident with other vegetables such as potato, egg plant, pepper, pumpkin and garden beet.
The propensity of the bulb mite to consume inert cellulose filter paper was exploited in the following induction experiments. R. robini consuming TSO- impregnated filter papers showed about 5-fold increase in microsomal EH activity (Figs 4, 5). In the dose-response experiment (Fig. 4) it was demonstrated that maximum EH activity was obtained at a level of 1.3 mg TSO per filter paper disc. Lesser stimulatory effects were observed at levels of 2.5 mg and 5.0 mg per disc. Fig. 5 describes the inducing effect of various epoxide containing compounds on R. robini EH activity. In addition to the above mentioned TSO, CSO and 2,4-dichlorostilbene oxide appreciably stimulated EH activity by 2.2 and 2.6-fold, respectively. Less potent were trans-chlorostilbene oxide (1.7 fold), 4-methyl- chalcone oxide (1.6-fold) and heptachlor epoxide (1.4 fold). Of the toxicants examined DDT, iindane (T-HCH) and malathion did not induce EH activity,
6 0 0
0 5 0 0 [...,
Z 0 L) 4 0 0 la, 0
0
(J
¢2 Z
3 0 0
2 0 0
100
0
0 ~ t)
e~
u E ' = N 2
P L A N T M A T E R I A L
Fig. 3. Effects of various plant materials on the induction of microsomal epoxide hydrolase in Rhyzoglyph~r robini. The mean specific activity of the control was 0.7 nmol styrene diol min -Img -] protein; the SD values for the other treatments were less than 10% of the mean.

388 E . C O H E N E T A L .
~ 7
~ 6 E
~_ 5
~ 4
~ 3
r. 2 .<
u 1
0 o 0
- - i . . . . . . r ~ - r . . . .
T I T I
L J J l L_
1 2 3 4 5 6
t r a n s - S T I L B E N E OXIDE. M G / D I S C
Fig. 4. Induction ofRhizoglyphm~ robini microsomal epoxide hydrolase by trans-stilbene oxide. The specific activity is expressed as nmol styrene diol rain - !mg -I protein.
0 b. 2: 0 U
5 0 0
4 0 0
3 0 0
a. 2 0 0 0
1 0 0
0
T S O C S O H E S O C S O D M C O C O
E P O X I D E S
Fig. 5. Induction of the microsomal epoxide hydrola~ ofRhizoglyphus robini by various epoxides. T S O - trans-stilbene oxide, C S O - cis-stilbene oxide, H E - heptachlor epoxide, S O C - trans-chioros- tilbene oxide, SOD - 2,4-dichlorostilbene oxide, MCO - 4-methylchalcone oxide, CO - chalcone oxide. The mean specific activity of the control was 1.0 nmol styrene diol min -l mg-] protein; the SD values for the tested compounds were less than 10% of the mean.

EPOXIDE HYDROLASE IN THE BULB MITE RHIZOGLYPHUS ROBIN1 PROPERTIES AND INDUCTION 389
whereas ingestion of 3-methylcholanthrene resulted in a considerable decrease in EH activity (Fig. 6). Sodium phenobarbital is an effective inducer, increasing R. robini EH activity by 2.6-fold as compared to the control. Most of the allelo- chemicals tested caused a decrease in R. robini EH activity (Fig. 7). Menthol, flavone, trans-carotene and chalcone drastically inhibited the enzyme activity. Less inhibitory were trans-cinnamic acid and allyl sulfide, while control-level EH activity was observed with the glycoside sinigrin.
300
25O
0 200 [- Z
150
t~ O
100
50
o PB MC DDT LIN MAL DEL
Fig. 6. Effect of various pesticides and toxicants as possible inducers of the microsomal epoxide hydrolase of Rhizoglyphus robini. PB - phenobarbital, MC - 3-methyl cholanthrene, DDT - p , p ' - DDT, LIN - lindane, MAL - malathion. Del - deltamethrin. The mean specific activity of the con- trol and the SD values as in Fig. 5.
120
9o 0
o t)
0
60
3O
0 A.S CAR CHA CIN FLA SIN M E N
ALLELOCHEMICALS
Fig. 7. Effects of various allelochemicals ingested byRhizoglyph~s" robini on the activity of its micro- somal epoxide hydrolase. AS - allyl sulfide, CAR - trans-carotene, CHA - chalcone, CIN - trans- cinnamic acid, FLA - flavone, SIN - sinigrin, MEN - menthol. The mean specific activity of the control and the SD values as in Fig. 5.

390 ~:,. COHEN ET AL.
DISCUSSION
The present study focused on R. robini EH with the intention of adding this system to the previously investigated detoxifying enzymes such as glutathione S-transferase (GST) (Capua et aL, 1991), esterases and monooxygenases (Capua et al., 1990) in the same mite species. As also observed in this study, EH of another mite species, T. urticae, is mostly associated with the microsomai fraction (Mullin et al., 1984). Unlike mammalian systems (Wixtrom and Hammock, 1985), using tritiated TSO and CSO as substrates did not differentiate between microsomal and cytosolic EHs of the bulb mite (Table 1).
The observation that metabolizing enzyme systems such as GST (Capua et al., 1991) and EH (this study) are capable of being induced to an appreciable degree by plant materials is of great significance. Induction of detoxification enzymes such as MFO, trans and cis EH and general esterases in the spider mite T. urticae was found to be host-related (Mullin and Croft, 1983). Although not hereditary in nature, the induction phenomenon may increase tolerance of pests to applied pes- ticides. It could perhaps explain different sensitivities of arthropod pests to pesti- cides depending on the infested host plant. In this regard, the early observation that T. urticae resistance to the organophosphorus (OP) tetraethyl pyrophosphate is more manifested in relation to certain plants (Saba, 1961 ), is noteworthy. The same mite species, selected for survival on a resistant cucumber variety, was also more tolerant to the OP pesticides as compared to a non-selected mite population (Gould et al., 1982). Increased tolerance to pesticides, which is related to plants or their secondary metabolites, was observed in insects. Lyophilized cotton flower buds and the allelochemical gossypol induced higher tolerance of bollworm (Heliothis Zea) larvae to methyl-parathion (Muehleisen et aL, 1989). Peppermint-fed larvae of the cut-worm Peridroma saucia displayed increased tolerance to OP and carbamate insecticides (Berry et aL, i 980). Phytochemicals such as peppermint oil and indole 3-carbinol induced midgut EH in the Southern armyworm Spodoptera eridania (Mullin, 1988).
It appears that detoxification systems in R. robini were distinctively induced. TSO, which considerably enhanced EH activity (this study) and GST (Capua, et al., 1991 ), had no such effect on aldrin epoxidation in the bulb mite (Capua et al., 1991). Based on studies with mammalian liver enzymes, it has been claimed that TSO differentially induces phase 11 detoxification enzyme systems such as EH and GST (DePiere et al., 1984; Di Simplicio et aL, 1989; Kuo et al., 1981). Others, however, regard TSO as a selective inducer since it was found to affect, although to a lesser extent, also the MFO system (Meijer et al., 1982; SeidegMd et aL, 1981 ; Schmassmann and Oesch, 1978). Phenobarbital appears to be a potent inducer of all detoxifying enzymes including microsomal EHs (Wixtrom and Hammock, 1985). It was found to considerably enhance transcription levels o f EH and cyto- chrome P-450 genes in rat liver (Hardwick et al., 1983). Sodium phenobarbital is an efficient inducer of R. robini EH (Fig. 6), and a close analog (pentobarbital)

EPOXIDE HYDROLASE IN TIfE BULB MITE RIIIZOGLYFHUS ROBINI PROPERTIE.S AND INDUCTION 391
significantly induced both GST and aldrin epoxidation in the same mite species (Capua et al., 1991). On the other hand, 3-methylcholanthrene, which induced rat liver microsomal EH (DePierre et aI., 1984; see also review of Wixtrom and Hammock , 1985), inhibited the bulb mite enzyme activity.
Induction ofRhizoglyphus detoxifying enzymes by allelochemicals is evident as flavone and chalcone increased G S T activity, whereas the former and trans-~- carotene stimulated aldrin epoxidation (Capua et al., 1991 ). These allelochemicals as well as others inhibited the microsomal EH activity. The above observations in conjunction with the already mentioned reports on the effect of TSO, clearly indi- cate that the major detoxifying enzyme systems in organisms are selectively induced by a variety of exogenously-applied compounds.
ACKNOWLEDGEMENTS
The tritiated TSO and CSO as well as various chalcone oxides were kindly provided by Dr. B.D. H a m m o c k from the Department of Entomology, University of California, Davis. The technical assistance of Mrs. Tamar Joseph is greatly appreciated. We thank Mr. Akivas Falk f rom Kibbutz Sde Eliahu for the supply of T. urticae and P. persimilis.
REFERENCES
Berry, R.E., Yu, S.J. and Terriere, L.C., 1980. Influence of host plants on insecticide metabolism and management of variegated cutworm. J. Econ. Entomol., 73:771-774.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein, utilizing the principle of protein-dye binding. Analyt. Biochem.. 72: 248-254.
Brooks, G.T., 1977. Epoxide hydratase as a modifier of biotransformation and biological activity. Gen. Pharmac., 8: 221-226.
Capua, S., Cohen, E. and Gerson, U., 1990. Non-specific esterases in mites: A comparative study. Comp. Biochem. Physiol. 96C: 125-130.
Capua, S., Cohen, E. and Gerson, U., 1991. Induction of aldrin epoxidation and glutathione S-trans- ferase in the mite Rhizoglyphus robini, Entomol. Exp. Appl. 59: 43-50,
Cohen, E., 1981. Epoxide hydrase activity in the flour beetle Tribolium castaneum (Coleoptera, Tene- brionidae). Comp. Biochem. Physiol., 69B: 29-34.
DePierre, J.W., Seideg~d, J., Morgenstern, R., Balk, J., Meijer, J., Astr6m, A., Nor61ius I. and Ersmer, L., 1984. Induction of cytosolic glutathione transferase and microsomal epoxide hydrolase activities in extrahepatic organs of the rat by phenobarbital, 3-methylcholanthrene and trans-stilbene oxide. Xenobiotica, 14: 295-301.
Di Simplicio P., Jennson. H. and Mannervik B., 1989. Effects of inducers of drug metabolism on basic hepatic forms of mouse glutathione transferase. Biochem. J., 263: 679-685.
Gerson, U., Cohen, E. and Capua S., 1991. Bulb mite, Rhizogtyphus robini (Astigmata: Acaridae) as an experimental animal. Exp. Appl. Acarol., 12:t03-110.
Gould, F., Carroll, C.R. and Futuyma, D.Y., 1982. Cross-resistance to pesticides and plant defenses: A study to the two-spotted spider mite. Ent. Exp. Appl., 31: 175-180.
Hammock, B.A. and Ota, K., 1983. Differential induction of cytosolic epoxide hydrolase, microso- mal epoxide hydrolase, and glutathione S-transferase activities. Toxicol. Appl. Pharrnacol., 71: 254-265.
Hardwick, J.P., Gonzalez, F.J. and Kasper. C.B.. 1983. Transcriptional regulation of rat liver

3 9 2 E. COHEN ET AL.
epoxide hydratase, NADPH-cytochrome P-450 oxidoreductase, and cytochrome P-450b genes by phenobarbital..I. Biol. Chem., 258:8081-8085.
Kuo, C.-H., Hook, J.B. and Bemstein, J., 1981. Induction of drug-metabolizing enzymes and toxic- ity of trans-stilbene oxide in rat liver and kidney. Toxicology, 22: 149-160.
Meijer, J., Astrtim, A., DePierre, J.W., Guengerich, F.P. and Erstner, L., 1982. Characterization of the microsomal cytochrome P-450 species induced in rat liver by trans-stilbene oxide. Biochem. Pharmac., 31 : 3907-3916.
Muehleisen, D.P., Benedict, J.H., Plapp, Jr., F.W. and Carino, F.A., 1989. Effects of cotton allelo- chemicals on toxicity of insecticides and induction of detoxifying enzymes in bollworm (Lepidoptera: Noctuidae). J. Econ. Entomol., 82: 1554-1558.
Mullin, C.A., 1988. Adaptive relationships of epoxide hydrolase in herbivorous arthropods. J. Chem. Ecol., 14: 1867-1888.
Mullin, C.A. and Croft, B.A., 1983. Host-related alterations of detoxification enzymes in Tetrany- thus urticae (Acari: Tetranychidae). Environ. Entomol. 12:1278-1282.
Mullin, C.A. and Croft, B.A., 1984. Trans-epoxide hydrolase: A key indicator enzyme for herbivory in arthropods. Experientia, 40: 176-178.
Mullin, C,A. and Hammock, B.D. 1982. Chalcone oxides - potent selective inhibitors of cytosolic epoxide hydrolase. Arch. Biochem. Biophys., 216: 423-439.
Mullin, C.A., Croft, B.A., Strickler, K., Matsumura, F. and Miller, J.R., 1982. Detoxification enzyme differences between a herbivorous and predetary mite. Science, 217:1270--1271.
Mullin, C.A., Matsumura, F. and Croft, B.A., 1984. Epoxide forming and degrading enzymes in the spider mite Tetranychus urticae. Comp. Biochem. Physiol., 79C: 85-92.
Oesch, F., 1972. Mammalian epoxide hydra~s: Inducible enzymes catalysing the inactivation of car- cinogenic and cytotoxic metabolites derived from aromatic and olefinic compounds. Xenobiotica 3: 305-340.
f Oesch, F., Jerina, D.M. and Daly, J.,. 1971. A radiometric assay,for hepatic epoxide hydrase activity
with [7-~H]styrene oxide. Biochim. Biophys. Acta, 227: 685~591. Oesch, F., Kaubisch, N., Jerina, D.M. and Daly, J.W., 1971a. Hepatic epoxide hydrase: Structure-
activity relationship for substrates and inhibitors. Biochemistry, 10: 4858--4866. Saba, F., 1961. Uber Entwicklung und Ruckgang der Giftresistenz bei Tetranychus urticae Koch und
deren Abhangigkeit vonder Wirksplanze. Z. Angew. Entomol., 48: 265-293. Schmassmann, H. and Oesch, F., 1978. Trans-stilbene oxide: A selective inducer of rat liver epox-
ide hydratase. Mol. Pharmac., 14: 834-847. Seideg~d, J., DePierre, J.W., Moron, M.S., Johannesen, K.A.M. and Ersmer, L., 1977. Characteri-
zation of rat lung epoxide (styrene oxide) hydrase with modified radioactive assay of improved sensitivity. Cancer Res. 37: 1075-1(182.
Seidegz~rd, J., DePierre, J.W., Morgenstern, R., Pilotti, A. and Erstner, L., 1981. Induction of drug- metabolizing systems and related enzymes with metabolites and structural analogues of stilbene. Biochim. Biophys. Acta, 672: 65-78.
Wixtrom, R.N. and Hammock, B.D., 1985. Membrane-bound and soluble-fraction epoxide hydro- lases: Methodological Aspects. In: D. Zakim and D. A. Vessey (Editors), Biochemical Pharma- cology and Toxicology: Methodological Aspects of Drug Metabolizing Enzymes. Vol. 1. John Wiley, New York, pp. 1-93.