enzymes and electron transport in microbial chlorate...
TRANSCRIPT

Faculty of Technology and ScienceChemistry
DISSERTATION
Karlstad University Studies2008:36
Jan Bohlin
Enzymes and electron transport in microbial
chlorate respiration

Karlstad University Studies2008:36
Jan Bohlin
Enzymes and electron transport in microbial
chlorate respiration

Jan Bohlin. Enzymes and electron transport in microbial chlorate respiration
DISSERTATION
Karlstad University Studies 2008:36ISSN 1403-8099 ISBN 978-91-7063-196-2
© The Author
Distribution:Faculty of Technology and ScienceChemistry651 88 Karlstad054-700 10 00
www.kau.se
Printed at: Universitetstryckeriet, Karlstad 2008

I
Abstract
Microbial chlorate respiration plays an important role in the turnover of oxochlorates in nature and in industrial waste management. This thesis deals with the characterization of the molecular components of chlorate respiration in Ideonella dechloratans. Chlorate respiration utilizes two soluble periplasmic enzymes, chlorate reductase and chlorite dismutase, to convert chlorate to chloride and oxygen. The genes encoding the enzymes participating in the chlorate degradation have been sequenced, and are found in close proximity, forming a gene cluster for chlorate metabolism. This work also includes the successful recombinant expression of three genes from Ideonella dechloratans. Two of the gene products, chlorite dismutase and the C subunit of chlorate reductase, participate in the chlorate respiration. The third gene, which is found close to the gene cluster for chlorate metabolism, encodes a soluble c-type cytochrome. The localization of the gene suggests the corresponding protein as a candidate for a role as electron donor to chlorate reductase. Also, the role of soluble periplasmic c cytochromes of Ideonella dechloratans in chlorate respiration was studied. At least one of the soluble c cytochromes was found capable of serving as electron donor for chlorate reduction. This c cytochrome, and several others, can also donate electrons to a terminal oxidase for subsequent reduction of oxygen, as required for the branched electron flow during chlorate respiration.


III
List of papers
I. Danielsson Thorell H, Karlsson J, Portelius E, Nilsson T,
Cloning, characterisation, and expression of a novel gene encoding chlorite dismutase from Ideonella dechloratans, Biochim Biophys Acta 1577 (2002) 445-451.
II. Danielsson Thorell H, Stenklo K, Karlsson J, Nilsson T, A
gene cluster for chlorate metabolism in Ideonella dechloratans, Appl Environ Microbiol 69 (2003) 5585-5592.
III. Karlsson J and Nilsson T, The C subunit of Ideonella dechloratans
chlorate reductase: expression, purification, refolding and heme reconstitution, Protein Expr Purif 41 (2005) 306-312.
IV. Bohlin J, Smedja Bäcklund A, Gustavsson N, Sara Wahlberg,
Nilsson T, Expression, refolding and reconstitution of a c-type cytochrome of Ideonella dechloratans, (2008) Manuscript.
V. Smedja Bäcklund A, Bohlin J, Gustavsson N, Nilsson T,
Periplasmic c cytochromes coupled to chlorate reduction in Ideonella dechloratans, Appl Environ Microbiol (2008) Submitted.

IV
Abbreviations
ATP Adenosine triphosphate CE Capillary electrophoresis Cld Chlorite dismutase cld The gene encoding chlorite dismutase (all genes are denoted in italic) ClO2- Chlorite ClO3- Chlorate ClO4- Perchlorate DdhA The α subunit of dimethyl sulfide dehydrogenase DMS Dimethyl sulfide DMSO Dimethyl sulfoxide DNA Deoxyribonucleic acid DTT Dithiothreitol EbdA The α subunit of ethylbenzene dehydrogenase EDTA Ethylenediaminetetraacetic acid EPR Electron paramagnetic resonance ESI Electrospray ionization Fe-S Iron-sulfur center GST Glutathione-S-Transferase HPLC High performance liquid chromatography IMAC Immobilized Metal Ion Affinity Chromatography IPTG Isopropyl-β-D-thiogalactoside ISIde1 Gene encoding an insertion sequence in I. dechloratans MALDI Matrix-assisted laser desorption ionization MBP Maltose binding protein MCD Magnetic circular dichroism MGD Molybdopterin guanine dinucleotide MobB Subunit of enzyme participating in molybdenum cofactor synthesis MS Mass spectrometry MS/MS Tandem mass spectrometry NADH Reduced nicotinamide adenine dinucleotide Nap Periplasmic respiratory nitrate reductase Nar Membrane-associated cytoplasmic respiratory nitrate reductase Nir Nitrite reductase ORF Open reading frame PAGE Polyacrylamide gel electrophoresis PCR Polymerase chain reaction PEG Polyethylene glycol RNA Ribonucleic acid SDS Sodium dodecyl sulfate SerA The α subunit of selenate reductase Tat Twin-arginine translocation pathway TOF Time of flight

Table of contents
Abstract I
List of papers III
Abbreviations IV
Introduction 1 Oxochlorates and bacterial respiration 1 Oxochlorates in the environment 1 Chlorate-reducing bacteria 2 Chlorate reductase and perchlorate reductase 3 Chlorite dismutase 4 The electron supply for chlorate reduction 4
Methods 8 Expression of recombinant proteins 8 Refolding and heme reconstitution of recombinant proteins 9 Mass spectrometry and peptide sequencing 10
Results and discussion 13 Cloning, characterisation, and expression of a novel gene encoding chlorite dismutase from Ideonella dechloratans [Paper I] 13 A gene cluster for chlorate metabolism in Ideonella dechloratans [Paper II] 14 The C subunit of Ideonella dechloratans chlorate reductase: expression, purification, refolding and heme reconstitution [Paper III] 18 Periplasmic c-cytochromes of Ideonella dechloratans [Paper IV and paper V] 20
Conclusions 25
Tack 26
References 28


1
Introduction Oxochlorates and bacterial respiration Bacterial respiration takes a significant part in the global circulation of several of the elements. Some of the reactions involved are of considerable interest for application in the biotreatment of toxic waste materials and for bioremediation [1]. The bacterial metabolism of oxyanions of compounds such as chlorine, selenium, and arsenic has therefore attracted interest lately [2, 3]. The present investigation deals with oxochlorates, oxyanions of chlorine, as electron acceptors in bacterial respiration. Oxochlorates in the environment In general, oxochlorates do not occur naturally in the environment. The only known natural source of any of these compounds is nitrate deposits in Chile which contain small amounts of perchlorate [4]. Environmental contamination by oxochlorates, such as chlorite (ClO2ֿ), chlorate (ClO3ֿ) and perchlorate (ClO4ֿ) caused by human activities has been recognized as a problem since the 1950’ies [5, 6]. The bulk of perchlorate contamination originates from its use in military (e.g. rocket propellant) and industrial applications [4, 5, 7-9]. Chlorate has been used as herbicide [10, 11], and is produced for use as starting material for production of chlorine dioxide in pulp mills [12]. Bleaching of pulp with chlorine dioxide produces chlorate as a byproduct due to the disproportionation of chlorine dioxide into chlorate and chlorite [13, 14]. The same problem arises when using chlorine dioxide as water disinfectant [15]. The environmental impact of chlorate is mainly due to its interference with nitrate uptake and/or metabolism in plants, in particular in marine species, such as brown algae [16]. This is due either to chlorate acting as competitive inhibitor to nitrate reductase, or to the conversion of chlorate to the highly reactive chlorite by the same enzyme [6, 10, 17, 18]. In mammals, toxicological studies have shown that perchlorate has a negative effect on iodide uptake by the thyroid gland [19, 20]. Perchlorate binds to the sodium-iodide symporter and inhibits the uptake of iodide. Long-term human exposure to perchlorate could ultimately result in the condition hypothyroidism [21].

2
The toxic effects of oxochlorates, together with their chemical stability and water solubility, have raised concerns about their environmental impact [10], and increased the interest in biological methods for treatment of oxochlorate-contaminated wastes. Chlorate and perchlorate can serve as respiratory electron acceptors in certain bacteria, with chloride ions as the end product [22, 23]. This reaction is the basis for biotreatment of chlorate-contaminated bleaching effluents in pulp mills [14, 24]. It has also attracted interest for biotreatment of perchlorate-contaminated wastes or bioremediation of contaminated sites [9, 25]. Chlorate-reducing bacteria Perchlorate- and chlorate-respiring bacteria have been found in a wide range of environments, e.g. in waste water sludge, aquatic sediments, forest and agriculture soils and animal waste lagoons [24, 26]. A number of bacterial species, such as Dechloromonas agitata (previously referred to as strain CKB) [26], Wolinella succinogenes [8], strain GR-1 [25], Pseudomonas chloritidismutans [27], Dechloromonas aromatica (previously referred to as strain RCB) [28], Moorella perchloratireducens [29] and Ideonella dechloratans [30], have been isolated using selection for growth on perchlorate or chlorate as the sole electron acceptor. D. aromatica strain RCB is, so far, the only known chlorate respiring species whose genome has been completely sequenced (http://www.jgi.doe.gov/). This species couples growth and benzene oxidation to the reduction of perchlorate [31]. Additional chlorate respirers have been identified by Bender et al. [32] by PCR, using primers specific for a conserved region in the gene for chlorite dismutase. This enzyme has, so far, been found in all chlorate respiring bacteria. Recently, also a method using quantitative real-time PCR has been developed in purpose of detecting pcrA, the catalytic subunit of perchlorate reductase [33]. Chlorate reduction is a two-step reaction that takes place in the periplasm. The molecular components of chlorate respiration are the enzymes chlorate reductase and chlorite dismutase, and a system for electron transfer from membrane-bound respiratory complexes (fig. 1). In the first step, chlorate is reduced to chlorite by chlorate reductase. In the second step, chlorite dismutase catalyses the decomposition of chlorite into chloride and oxygen, which is then used by a respiratory oxidase. In the case of perchlorate reduction, the same enzyme, in this case termed perchlorate reductase, reduces perchlorate to chlorate, and then to chlorite [25, 34].
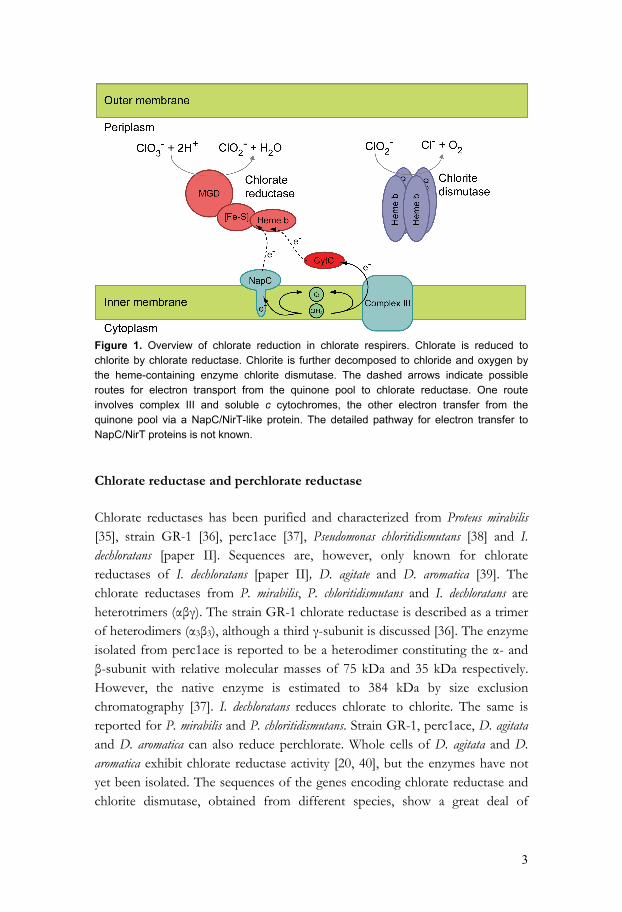
3
Figure 1. Overview of chlorate reduction in chlorate respirers. Chlorate is reduced to chlorite by chlorate reductase. Chlorite is further decomposed to chloride and oxygen by the heme-containing enzyme chlorite dismutase. The dashed arrows indicate possible routes for electron transport from the quinone pool to chlorate reductase. One route involves complex III and soluble c cytochromes, the other electron transfer from the quinone pool via a NapC/NirT-like protein. The detailed pathway for electron transfer to NapC/NirT proteins is not known. Chlorate reductase and perchlorate reductase Chlorate reductases has been purified and characterized from Proteus mirabilis [35], strain GR-1 [36], perc1ace [37], Pseudomonas chloritidismutans [38] and I. dechloratans [paper II]. Sequences are, however, only known for chlorate reductases of I. dechloratans [paper II], D. agitate and D. aromatica [39]. The chlorate reductases from P. mirabilis, P. chloritidismutans and I. dechloratans are heterotrimers (αβγ). The strain GR-1 chlorate reductase is described as a trimer of heterodimers (α3β3), although a third γ-subunit is discussed [36]. The enzyme isolated from perc1ace is reported to be a heterodimer constituting the α- and β-subunit with relative molecular masses of 75 kDa and 35 kDa respectively. However, the native enzyme is estimated to 384 kDa by size exclusion chromatography [37]. I. dechloratans reduces chlorate to chlorite. The same is reported for P. mirabilis and P. chloritidismutans. Strain GR-1, perc1ace, D. agitata and D. aromatica can also reduce perchlorate. Whole cells of D. agitata and D. aromatica exhibit chlorate reductase activity [20, 40], but the enzymes have not yet been isolated. The sequences of the genes encoding chlorate reductase and chlorite dismutase, obtained from different species, show a great deal of

4
similarity, but the arrangement of the genes differ [paper II and paper IV]. Chlorate reductase is a member of the dimethylsulfoxide (DMSO) reductase family, which is a large and diverse group of molybdenum enzymes found in archaea and bacteria. These enzymes are often involved in the anaerobic respiration, and the dissimilatory reduction of oxyanions and oxides [2]. Chlorite dismutase Chlorite dismutase has been purified from strain GR-1 [41], D. agitata [42] and I. dechloratans [43]. The three purified enzymes are all heme-containing homotetramers, with molecular masses around 100 kDa. Chlorite dismutase from I. dechloratans has been expressed recombinantly by Danielsson Thorell et al. (paper I). Recently Streit and DuBois [44] reported the successful expression of recombinant chlorite dismutase from D. aromatica. The native counterpart of D. aromatica chlorite dismutase has, however, not yet been isolated. The recombinant variant was predicted to be a homotetramer with a molecular weight of 116 kDa and was reported soluble and highly functional. Chlorite dismutase from I. dechloratans shows a high level of identity with its counterparts in D. agitata (ID: AY124796), D. aromatica (ID: CP000089), Dechlorosoma sp. KJ (ID: EU571095), Pseudomonas stutzeri (ID: EU436748), P. chloritidismutans (ID: EU436747) and Dechloromonas hortensis (ID: EU436749) at the amino acid level. Electron paramagnetic resonance (EPR) spectroscopy shows that chlorite dismutase from I. dechloratans and strain GR-1 contains a high-spin ferric heme [43, 45] and the hemochrome analyses suggest a b-type heme [paper I]. The electron supply for chlorate reduction Even though the pathway for chlorate reduction is fairly well understood, less is known about the source of electrons for the initial reduction of chlorate or perchlorate. The situation is, however, analogous to several variants of bacterial respiratory chains, where the final steps are separated from the membrane-bound respiratory complexes. The role of the respiratory chain is to convert the free energy available from a redox reaction into a proton gradient across the cell membrane. This electrochemical gradient provides the driving force for synthesis of ATP by the membrane-bound ATP-synthase.

5
Prokaryotic respiration takes many forms depending on electron donors available and the nature of the terminal electron acceptor. Most respiratory chains have in common the participation of lipid-soluble quinones as electron carriers (fig. 2). In some cases, the quinones act as electron donors in the final step of respiration. Examples are quinol oxidases such as cytochrome bo3, and Nar (respiratory nitrate reductases) [46, 47]. In other cases, a membrane-bound complex III transfers electrons from the quinone pool to water-soluble c cytochromes that can serve as electron donors to either membrane-bound complexes or soluble enzymes. Membrane-bound and terminal reductases using cytochrome c as substrate include aa3 and cbb3 cytochrome c oxidase [48, 49]. Examples of soluble enzymes are nitrite and N2O reductases and NapAB (periplasmic nitrate reductase) [50, 51]. The chlorate and perchlorate reductases also belong to this category of enzymes. Finally, there are also examples of prokaryotes that carry out their terminal reduction using oxidoreductases localized to the outer membrane. A good example is different strains of the facultative anaerobe Shewanella, where the proposed electron transfer is that CymA, a membrane anchored c-type cytochrome structurally similar to the NapC/NirT family, via a soluble c-type cytochrome, reduces the terminal reductase located in the bacterial cell wall (fig. 2)[52-55].
Figure 2. Examples of aerobic and anaerobic respiration in prokaryotes. Electrons are transferred from a dehydrogenase, via the quinone pool to finally end up in a terminal oxido-reductase. Nap, respiratory periplasmic nitrate reductase; Nar, Respiratory cytoplasmic nitrate reductase; Nir, Nitrite reductase. Different strategies for electron transport are found among the oxidoreductases of the DMSO reductase family. In the Nap system, found in the periplasm in some nitrate-reducing bacteria, electrons are passed via the NapC-protein, a membrane-anchored tetraheme c-cytochrome (fig. 3a) [47]. A different arrangement for electron transport between periplasmic enzymes and a
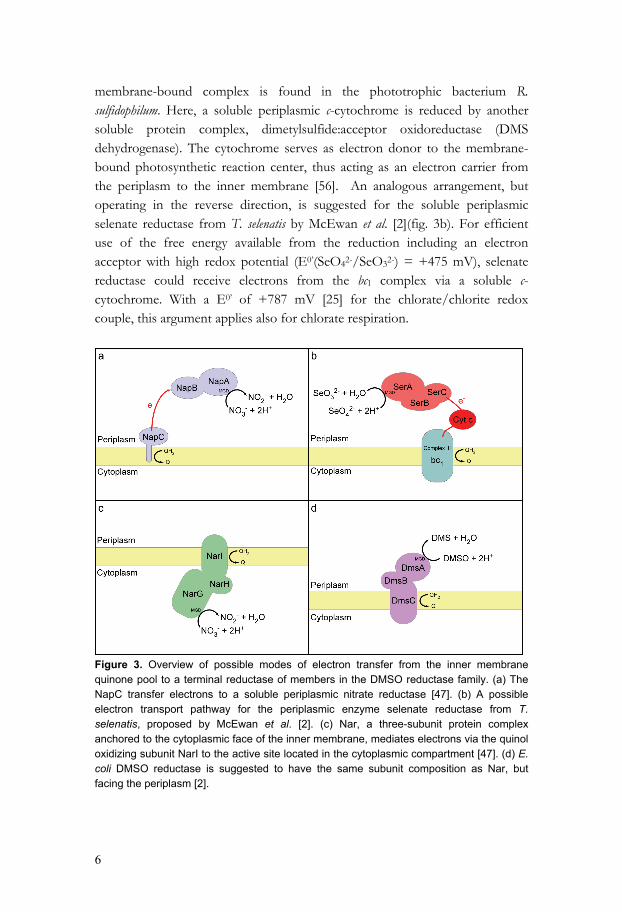
6
membrane-bound complex is found in the phototrophic bacterium R. sulfidophilum. Here, a soluble periplasmic c-cytochrome is reduced by another soluble protein complex, dimetylsulfide:acceptor oxidoreductase (DMS dehydrogenase). The cytochrome serves as electron donor to the membrane-bound photosynthetic reaction center, thus acting as an electron carrier from the periplasm to the inner membrane [56]. An analogous arrangement, but operating in the reverse direction, is suggested for the soluble periplasmic selenate reductase from T. selenatis by McEwan et al. [2](fig. 3b). For efficient use of the free energy available from the reduction including an electron acceptor with high redox potential (E0’(SeO42-/SeO32-) = +475 mV), selenate reductase could receive electrons from the bc1 complex via a soluble c-cytochrome. With a E0’ of +787 mV [25] for the chlorate/chlorite redox couple, this argument applies also for chlorate respiration.
Figure 3. Overview of possible modes of electron transfer from the inner membrane quinone pool to a terminal reductase of members in the DMSO reductase family. (a) The NapC transfer electrons to a soluble periplasmic nitrate reductase [47]. (b) A possible electron transport pathway for the periplasmic enzyme selenate reductase from T. selenatis, proposed by McEwan et al. [2]. (c) Nar, a three-subunit protein complex anchored to the cytoplasmic face of the inner membrane, mediates electrons via the quinol oxidizing subunit NarI to the active site located in the cytoplasmic compartment [47]. (d) E. coli DMSO reductase is suggested to have the same subunit composition as Nar, but facing the periplasm [2].

7
A different approach is used by the membrane-bound nitrate reductase (NAR), which is a three-subunit complex, facing the cytoplasm and anchored to the membrane via NarI, a di-b-heme quinol dehydrogenase (fig. 3c) [47]. E. coli DMSO reductase is suggested to have a similar subunit composition but with the subunit containing the active site facing the periplasm (fig. 3d) [2]. Also, archaeal respiratory nitrate reductases found in the Haloarcula marismortui and Haloferax mediterranei are suggested to be configured in the same way [57, 58]. I. dechloratans respires with both oxygen and chlorate as terminal electron acceptors, suggesting that the electron transport includes both a periplasmic oxidoreductase and a membrane-bound terminal oxidase. I. dechloratans is reported to be cytochrome c-oxidase positive [30]. Thus, the oxygen produced by decomposition of chlorite is also used as respiratory electron acceptors. Of the six electrons needed for reduction of chlorate to chloride, two are used by chlorate reductase, and the remaining four by a membrane-bound cytochrome c oxidase. This requires a branch point where the electron flow diverges, with one branch supplying electrons for chlorate reductase, and the other supplying electrons for the terminal oxidase. The present work was undertaken with the aim of understanding in more detail the molecular basis for (per)chlorate respiration. This includes characterization of genes and proteins involved in chlorate reduction, chlorite decomposition and electron transfer. Three genes has been cloned and expressed. Two of the gene-products, chlorite dismutase and the C subunit of chlorate reductase, participate in the chlorate respiration. The third gene, which is found close to the gene cluster for chlorate metabolism, encodes a soluble c-type cytochrome. The localization of the gene makes its product a candidate for a role in electron supply to chlorate reductase. Of the soluble periplasmic c-type cytochromes of I. dechloratans, at least one serving as electron donor to chlorate reductase was identified (paper V), suggesting an electron transport route similar to that proposed in T. selenatis (fig. 3b) [2].

8
Methods Expression of recombinant proteins Several expression hosts are used for the production of recombinant proteins. However, the present work focuses on production of recombinant proteins in E. coli. This includes the insertion into the host a plasmid carrying the target gene under control of a suitable promoter. In the present study, the T7 promoter has been used. For expression using the T7 promoter [59, 60] the plasmid carrying the target gene is transformed into a host containing the gene encoding T7 RNA polymerase (λDE3 lysogen). Here, the T7 RNA polymerase gene is under the control of the lacUV5 promoter, which may be induced with IPTG. Expression from the T7 promoter is induced when the host cell expresses T7 RNA polymerase. High-level expression of the target gene can produce as much as 40-50% of the total cell protein. However, the expression levels obtained with strong promoter systems can also give solubility problems, with most of the target protein ending up in insoluble aggregates, also known as inclusion bodies. There are several ways to suppress the formation of inclusion bodies. One obvious approach is to lower the rate of protein synthesis by using a vector with a weaker promoter. A lower extent of expression can also be achieved by lowering the IPTG concentration. However, this only applies for certain expression systems. Expression at lower growth temperature can also produce more soluble protein. Other ways to enhance expression of soluble and correctly folded proteins is by co-expressing chaperones and foldases [61]. Also, growth of the host cells under osmotic stress could favour production of soluble target protein [62]. Expression of the target protein as a fusion product is a different approach for improvement of stability and solubility. Widely used fusion partners for protein expression in E. coli are glutathione S-transferase (GST) [63], maltose-binding protein (MBP) [64] and thioredoxin [65]. GST and MBP have the advantage of high-level expression followed by purification by affinity chromatography, whereas thioredoxin is chosen because of its high solubility in the E. coli cytoplasm. Several vector systems also provide the option of tagging the target protein with histidine residues for subsequent purification with immobilized metal affinity chromatography (IMAC).

9
Refolding and heme reconstitution of recombinant proteins The formation of inclusion bodies can also be turned to an advantage, because they can often be produced in large quantities and are reasonably easy to purify. Moreover, the insoluble protein often is protected against proteolytic activity. On the other hand, the inclusion bodies must be solubilized using denaturing conditions. Renaturation of the target protein, with the aim of regaining native properties can be difficult, especially when the process includes insertion of a cofactor. Initial purification of inclusion bodies usually includes several washing steps, with pelleting of the inclusion bodies by centrifugation. Even though the major part of the washed pellet is likely to be the target protein, it probably also is contaminated with membrane fragments containing host membrane proteins. Therefore, additional purification in the denatured state is usually needed after the inclusion bodies have been dissolved using denaturants such as urea or guanidine chloride. To produce native protein after purification in the denatured state, refolding is carried out. The major problem in refolding is the kinetic competition between folding and aggregation. Folding is normally a unimolecular reaction subject to first-order kinetics, whereas aggregation is of second- or higher order [66]. High dilution is therefore expected to favor folding over aggregation. However, conditions that promote refolding to the native state are individual and must be determined empirically in each case. Important parameters include protein concentration, pH, temperature, ionic strength and the redox environment [67]. Refolding is usually initiated by the removal of the denaturant by dialysis or dilution. Other possible methods are so-called column refolding by affinity chromatography or size exclusion chromatography [68]. Protein refolding is reviewed by Lilie et al. and De Bernardez Clark [69, 70]. Cofactors, such as metal ions or other non-protein components, may play essential roles for the stability of proteins. Also, the presence of correctly reconstituted cofactors is crucial for a fully functioning protein. Refolding of heme-containing protein includes the in vitro association of the heme group, either by incubating heme with the refolded apo-protein or by carrying out refolding in the presence of the cofactor. Successful reconstitution with heme has been reported for hemoglobin, myoglobin [71], cytochrome b-562 [72], synthetic peptides [73, 74], the soluble domain of cytochrome b5 (reconstituted

10
with manganese protoporphyrin IX) [75] and the spinach chloroplast cytochrome b6 [76]. Successful reconstitution of c type cytochromes has also been reported [77]. Mass spectrometry and peptide sequencing Mass spectrometry is an analytical tool used for measuring the mass of molecules with very high accuracy. For large molecules, such as biomolecules, masses can be determined within an accuracy of 0.01% of the total molecular mass of the sample [78, 79]. A mass spectrometer consists of three parts, the ion source, the analyzer and the detector. The sample is introduced into the instrument and its components are ionized by the ion source, producing ions in the gas phase. The resulting ions are then passed into the analyzer region, where they are separated according to their mass-to-charge ratios (m/z). The sample can be introduced directly into the ionization source. Alternatively MS can be used as a selective detection method after another separation method, such as high pressure liquid chromatography (HPLC). Capillary electrophoresis (CE) is also possible to use as a preseparation step for biomolecules [80]. The most common methods of ionization of biomolecules are matrix assisted laser desorption ionization (MALDI) and electrospray ionization (ESI) [81]. These provide mild ionization conditions and produce little fragmentation of the sample molecules. MALDI is a solid-state technique in which laser is used to vaporize a mixture of sample and matrix dried onto a solid support. The matrix absorbs the energy from the laser and protects the biomolecules against fragmentation. The matrix consists of crystallized molecules, such as α-cyano-4-hydroxy cinnamic acid. When laser light hits the sample a small part vaporizes, transferring the solid sample into gas phase. ESI, in contrast, is a flow-based system where liquid sample flowing into the system produces an aerosol. The small droplets are ionized as they pass through the charged sample inlet nozzle. The liquid then evaporates, resulting in ionized sample in the gas phase [82]. The source of the flowing sample can be the outlet of a chromatographic column which is coupled in-line with the ESI. In contrast, MALDI is an offline technique, in which sample is dried onto a solid support. MALDI and ESI differ in the charge state they impart on the sample ions. MALDI produces singly charged (+1) ions in most cases. ESI produces a range of differently charged ions for each sample molecule. The resulting complexity of an ESI spectrum, with a

11
collection of ions for each sample molecule, one for each charge state, lowers the signal-to-noise ratio. On the other hand, the multiple charges on the fragments make the method suitable for analysis of large sample molecules, such as non degraded proteins, since m/z will fall in a range suitable for the mass analyzer even at high molecular masses [81, 82]. When using MS for protein identification, a common method is to generate tryptic peptides, usually by in-gel digestion, followed by mass determination. A protein database is then queried for fragments with the observed masses. If the sequence of the protein of interest is not present in the database, a different approach is needed. Using a tandem mass spectrometer a peptide, selected from the MS-analysis, is usually fragmented by collision with an inert gas. The resulting fragments are separated in a second MS-step, producing a MS/MS spectrum. With knowledge about the fragmentation mechanism, it is possible to deduce the sequence of the original peptide [83]. Fragmentation usually occurs by cleavage of one of the bonds in the peptide backbone, most commonly the peptide bond (CO-NH). However, it is also possible with cleavage between the α-carbon and the carbonyl carbon (CH-CO) and between the α-carbon and the nitrogen (NH-CH). Each bond breakage give rise to two species, one neutral and one charged, in the case of singly charged ions produced by MALDI. The charge can stay on either of the two species depending on the cleavage mechanism and relative proton affinity of the fragments. Hence, there are six possible fragment ions for each amino acid residue. The fragments labeled a, b and c is ions having their charges retained at the N-terminal fragment. The x, y, and z ions are charged C-terminal fragments (fig. 4).
Figure 4. Overview of the six possible cleavage sites in the amino acid backbone, and their corresponding ions.

12
A consequence of preferred cleavage of the peptide bond is that the dominating ions in an MS-spectrum are the b and y ions. The mass difference between two adjacent ions from the same series corresponds to the mass of a particular amino acid residue. Due to the complexity of the tandem mass spectra and the difficulty to identify different ion series, the analysis often is performed with computer assistance. With high quality spectra and/or some knowledge about the sequence at hand, it can also be possible to pick out the amino acid sequence manually [81]. Derivatization or chemical labeling can also facilitate the interpretation of a complex peptide fragment pattern [84, 85].

13
Results and discussion Cloning, characterisation, and expression of a novel gene encoding chlorite dismutase from Ideonella dechloratans [Paper I] The gene encoding the periplasmic chlorite dismutase was isolated from a genomic library using a hybridization probe generated by PCR, with degenerate primers designed from partial amino acid sequences. The complete gene was sequenced using primer walking. For heterologous expression in E. coli, the complete gene, including the sequence encoding a 22 amino acid residue signal peptide, was amplified by PCR and cloned into a pET-3a expression vector. Positive clones were confirmed by colony PCR and the sequences were analyzed and verified. To increase heme synthesis during cell growth, the heme precursor δ-aminolevulinic acid and FeSO4 were added [86, 87] to the culture. After induction with IPTG, chlorite dismutase activity could be detected, and SDS-PAGE revealed a novel protein with a molecular weight of 30 kDa. The molecular weight of native chlorite dismutase monomer is 25 kDa [43]. The higher molecular weight of the recombinant protein indicates that the signal peptide was not processed. The chlorite dismutase activity detected in intact cells did not increase noticeable upon cell disruption, suggesting most of the activity is located in the periplasmic compartment. However, only small part of the chlorite dismutase activity was found in the soluble periplasmic fraction. Instead, most of the activity was detected in the membrane fraction. Together, these results suggest that the major part of the protein is exported to the periplasm but remains associated with the membrane. Attempts to solubilize the recombinant protein from the membrane fraction using detergents, high ionic strength, butanol or urea [88] were unsuccessful. However, the target protein could be solubilized with guanidinium chloride, with loss of the activity. The denatured protein could be refolded by dialysis and reconstituted with heme, recovering some of the activity. Instead, the gene encoding the mature protein, without the signal peptide, was cloned for expression in the cytosol. Intact BL21(DE3) cells transformed with a

14
plasmid carrying the target gene showed very low activity after induction with IPTG. However, after cell disruption, the chlorite dismutase activity of the homogenate was substantially higher. The mature recombinant chlorite dismutase was purified with ammonium sulfate precipitation and hydrophobic chromatography followed by size exclusion chromatography, as described earlier [43]. SDS-PAGE analysis showed that the recombinant form had a slightly lower mobility. This is probably due to a posttranslational modification affecting the hydrodynamic radius of the denatured enzyme, rather than a difference in the molecular weight between the native and the recombinant enzyme [89]. The optical spectrum of the recombinant form differed somewhat from that of the native enzyme in the Soret region. The heme content found was about half of that of the native chlorite dismutase. In order to facilitate purification, chlorite dismutase was also cloned and expressed as a GST fusion protein. This resulted in a soluble fusion protein that could be purified in one step by affinity chromatography. The specific activity was, however, only 5% of that of native chlorite dismutase, and the heme content was very low. This fusion protein has not been investigated further, but the refolding results presented above, together with the results discussed in paper III, suggest that in vitro heme incorporation could be feasible. A gene cluster for chlorate metabolism in Ideonella dechloratans [Paper II] Chlorate reductase was isolated and purified from I. dechloratans. The purified enzyme was characterized and found to contain a molybdenum cofactor and iron. The optical spectrum and the pyridine hemochrome analysis also indicated the presence of heme b. The enzyme activity was followed by methyl viologen radical oxidation [90], using chlorate as electron acceptor. Since chlorate reductase activity could be detected in intact cells and since the electron donor, methyl viologen radical, does not enter the cytoplasmic compartment [91], a periplasmic location was suggested for the I. dechloratans chlorate reductase. In order to clone and sequence the genes encoding chlorate reductase from I. dechloratans, the α subunit was digested with trypsin and two of the resulting internal peptides were sequenced. Based on the peptide sequences, and the homology to other MGD (molybdopterin guanine dinucleotide) proteins, two degenerate internal primers were synthesized in order to produce digoxigenin-

15
labeled probes by PCR. The probes were used to screen the genomic library described in paper I, and positive clones were sequenced by primer walking. The two different clones isolated from the genomic library did, however, not cover the complete gene cluster. Further screening with additional probes, and attempts to isolate the missing sequence by inverse PCR were unsuccessful, probably due to the high GC content and the resulting difficulty to obtain sufficiently specific hybridization. In order to sequence the remaining part of the gene cluster, a Genome Walker library (Clontech) was constructed. Genome walking is a method for cloning an unknown DNA sequence adjacent to known sequence. The library is constructed by digesting genomic DNA, using restriction enzymes producing blunt ends, followed by ligation with an adaptor, provided by Clontech. Nested PCR is then carried out with two adaptor-specific and two gene-specific primers (fig. 5). In the present case, problems due to the high GC content of the template were avoided by using hot start PCR and buffers optimized for GC-rich templates. The PCR reactions resulted in an approximately 2000 base pair long product, which was cloned and partially sequenced. On basis of the new sequence information, additional primers were constructed. The PCR product obtained from these primers was then cloned and the remaining part of the fragment was sequenced.
AP1AP2
GSP1GSP2
Genomic DNA fragment Genome Walker adaptor
5'5'Genome Walker adaptor
Figure 5. Schematic figure of a Genome Walker library. Adaptors are ligated to the blunt end digested DNA. A nested PCR is then performed, with two adaptor specific- (AP) and two gene specific primers (GSP). The complete sequence was found to include four genes, with the arrangement clrABDC (fig.6). This arrangement is similar to that found in other members of the type II DMSO reductase family such as serABDC from T. selenatis [92] and ddhABDC from R. sulfidophilum [56]. However, edbABCD from the Azoarcus-like strain EbN1 [93] and the uncharacterized clrABCD from D. aromatica has a slightly different gene arrangement. In the case of D. aromatica, the clrA, clrB and clrD sequences are homologous to those found in I. dechloratans. The clrC sequence, however, shows no resemblance to its counterpart in I. dechloratans. Instead, the clrC gene shows sequence similarity to known tetraheme c-

16
cytochromes. The sequence includes four characteristic heme binding sequence motifs (CXXCH), indicative for c-cytochromes. This cytochrome is not predicted to be a membrane associated protein [39], but to serve as an electron transporter between a NapC-like system and periplasmic perchlorate reductase.
cld ISIde1 clrA clrB clrD clrC ORF1 ORF2
Figure 6. Overview of genes involved in the chlorate respiration in I. dechloratans. The gene encoding chlorite dismutase (cld) and the group of genes encoding chlorate reductase (clrABDC) are separated by an insertion element (ISIde1). ORF1 is, by Pfam, identified as a soluble cytochrome c, and ORF2 as a mobB, a protein involved in the molybdopterin biosynthesis. The present study shows that chlorate reductase from I. dechloratans contains a molybdenum cofactor. The gene encoding the A subunit includes a sequence motif characteristic for molybdenum-containing type II DMSO reductases. The gene also contain a N-terminal leader peptide for export to the periplasm via the Tat pathway [94]. Analyses of the clrB gene do not reveal any presence of a signal peptide. Instead, ClrB is suggested to be translocated as a folded protein together with ClrA, by the Tat system using the “hitchhiker” mechanism proposed by Rodrigue et al. [95]. The gene encoding ClrB contains four sequence motifs that could bind Fe-S cluster. The gene encoding ClrC has a predicted 22-residue leader peptide for export to the periplasm. This signal peptide is similar to that found in the homologue SerC and is suggested to direct export of the unfolded subunit via a secretory pathway [92, 96]. Mutagenesis shows that DdhC from R. sulfidophilum is the heme-containing subunit, and magnetic circular dichroism (MCD) suggests that the axial ligands are histidine and methionine [97]. Although alignment of I. dechloratans ClrC and its homologues (fig. 7) indicates that the C subunit is less conserved than ClrA and ClrB, it has been suggested that this subunit binds heme b [56, 58, 92, 98, 99]. Conserved methionine and histidine residues are present in DdhC, SerC and ClrC sequences (* in figure 7). The methionine is present also in the other sequences, but here no obvious candidate for the other axial ligand can be found. Archaeal ClrC homologues ORF7 from H. mediterranei and ORF8 from H. marismortui are found associated to genes involved in nitrate respiration. The corresponding polypeptides do not, however, seem to be parts of αβγ-periplasmic oxidoreductases.
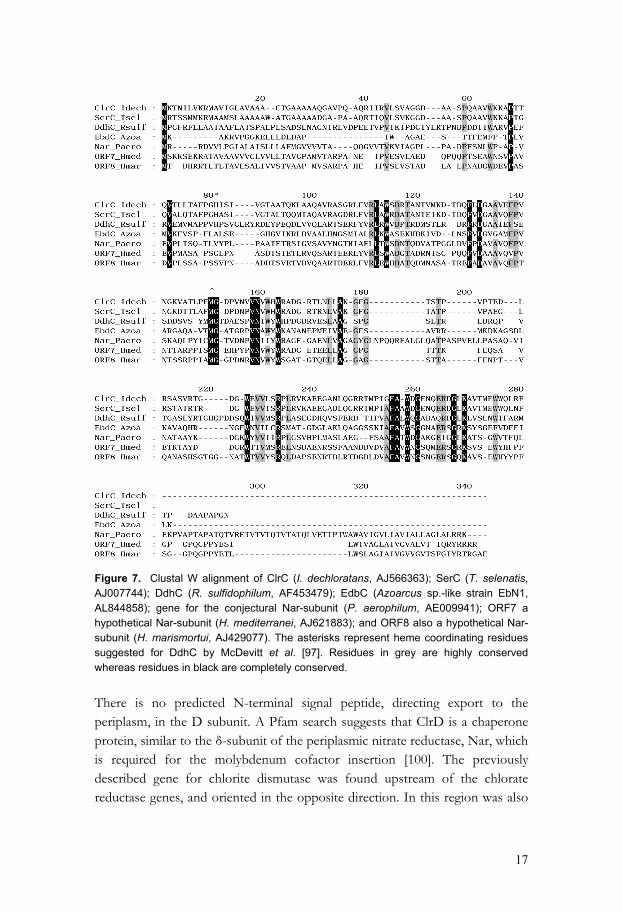
17
Figure 7. Clustal W alignment of ClrC (I. dechloratans, AJ566363); SerC (T. selenatis, AJ007744); DdhC (R. sulfidophilum, AF453479); EdbC (Azoarcus sp.-like strain EbN1, AL844858); gene for the conjectural Nar-subunit (P. aerophilum, AE009941); ORF7 a hypothetical Nar-subunit (H. mediterranei, AJ621883); and ORF8 also a hypothetical Nar-subunit (H. marismortui, AJ429077). The asterisks represent heme coordinating residues suggested for DdhC by McDevitt et al. [97]. Residues in grey are highly conserved whereas residues in black are completely conserved. There is no predicted N-terminal signal peptide, directing export to the periplasm, in the D subunit. A Pfam search suggests that ClrD is a chaperone protein, similar to the δ-subunit of the periplasmic nitrate reductase, Nar, which is required for the molybdenum cofactor insertion [100]. The previously described gene for chlorite dismutase was found upstream of the chlorate reductase genes, and oriented in the opposite direction. In this region was also

18
identified an insertion element which is termed ISIde1. Downstream of the genes encoding chlorate reductase two additional ORF’s are identified (fig. 6). ORF1 is recognized by Pfam as a gene encoding a soluble cytochrome c, and ORF2 as a gene, mobB, which encodes a protein involved in the molybdopterin cofactor maturation system. The soluble cytochrome c is of particular interest because of its possible involvement in electron transport. The C subunit of Ideonella dechloratans chlorate reductase: expression, purification, refolding and heme reconstitution [Paper III] For future study of the function, the C subunit from I. dechloratans chlorate reductase has been expressed as a GST fusion protein. Both the gene encoding the predicted mature C subunit and the complete gene including the predicted signal peptide were amplified by PCR from a Genome Walker library. DNA polymerase and buffer suitable for GC-rich templates were used. The amplified gene encoding the predicted mature protein was ligated into a pET-3A expression vector, transformed into E. coli and confirmed free from mutations by cycle sequencing. However, no signs of expression were observed, possible as the result of an unsuitable N-terminal or penultimate amino acid residue [101, 102]. Instead, the complete gene, including the signal peptide, was ligated into a pGEX-2T fusion vector and treated the same way as the gene encoding the predicted mature protein. Expression in E. coli BL21 produced a considerable amount of target protein. However, the major part of the target protein was found in insoluble aggregates as inclusion bodies, whereas only a minor part, that also seemed to be subject to proteolytic degradation, was found in the soluble fraction. Attempts to increase the fraction of soluble target protein and to inhibit the proteolytic activity were unsuccessful. On the other hand, a higher expression level increased the amount of insoluble target protein, and also decreased the extent of proteolytic degradation to an acceptable level. These results suggested a strategy of expressing the fusion protein in large quantities as inclusion bodies for subsequent purification in the denatured state. The inclusion bodies were washed in low ionic strength buffer, and solubilized in urea. The solubilized GST/ClrC fusion protein was further purified by cation exchange chromatography. About 50 mg protein per liter cell culture, sufficiently pure for refolding studies, was obtained. In order to optimize the

19
renaturation of the fusion protein, screening of different refolding conditions was performed. It was found that dilution at 4° C into Tris buffer containing polyethylene glycol (PEG), arginine, EDTA and dithiothreitol (DTT) recovered about 60% soluble protein with GST activity. The additives PEG and arginine probably serve to prevent aggregation of the protein, whereas DTT is included to prevent formation of incorrect disulfide bonds [103, 104]. Reconstitution of heme is generally performed after the refolding, with the apo-protein in its native conformation [76, 105, 106]. Incubation of heme with the refolded fusion protein did, however, not give any spectral evidence for heme incorporation. This may be due to partial aggregation of the fusion protein, or that the GST fusion partner interferes with the heme incorporation. On the other hand, the presence of an equimolar amount of heme in the refolding buffer resulted in a product with an optical spectrum reminiscent of a b-type cytochrome. Because of an apparent effect of DTT on the pyridine hemochrome spectrum of the refolded product, it was necessary to exclude DTT from the refolding buffer and instead preincubate the urea-denatured protein with DTT. The refolded and heme-reconstituted fusion protein was characterized using UV-vis spectroscopy. The optical spectrum of the recombinant fusion protein and native chlorate reductase were similar, with minor differences in the positions of the Soret bands, and the width of the α-bands. The Soret band of the native chlorate reductase is found at 426 nm in the reduced state, and at 416 nm in the oxidized state. The reduced form of the fusion protein has its Soret maximum at 424 nm, and in the oxidized form at 415 nm. The difference between the Soret band positions of native chlorate reductase and recombinant fusion protein is probably due to the presence of free or unspecifically bound heme, absorbing at 385 nm [107]. In the visible region, no difference between the positions of the α and β bands of native chlorate reductase and the reconstituted product was observed. In the reduced-minus-oxidized difference spectra in the visible region, α and β bands are found at 559 and 529 nm in both the native chlorate reductase and in the reconstituted C subunit. Broader peaks were, however, observed in the latter case. The UV-visible spectrum of the heme group in the GST/ClrC fusion suggest that this protein can provide a heme environment similar to that of the native chlorate reductase found in I. dechloratans. This supports the earlier suggestions that the C subunit of oxidoreductases in the type II DMSO reductase family is

20
the heme b-containing subunit. The recombinant version of the C subunit, equipped with a GST tag, is therefore expected to be useful for further studies of electron transfer and protein interactions. Periplasmic c-cytochromes of Ideonella dechloratans [Paper IV and paper V] As discussed earlier, two different strategies for electron transfer between membrane-bound respiratory complexes and soluble proteins of the DMSO-reductase family have been found. One is the transfer of electrons via a membrane-bound NapC/NirT complex [108]. The other strategy is the transfer of electrons via a soluble periplasmic c-type cytochrome, by analogy with the phototropic bacterium Rhodovulum sulfidophilum [56, 97]. To investigate role of soluble c-type cytochromes and their ability to serve as electron donors for reduction of chlorate and oxygen, the c cytochromes in a periplasmic extract was reduced. On addition of chlorate, in presence of chlorate reductase, partial oxidation of c cytochromes was observed. Thus, there are soluble c-type cytochromes capable of serving as electron donors to chlorate reductase present in the periplasm of I. dechloratans. From these results, it was decided to attempt the isolation of soluble periplasmic c cytochromes in order to investigate their role as electron donors. Five candidate c cytochromes, 20, 10, 9, 8 and 6 kDa in size, were identified in the periplasmic extract using SDS-PAGE and a staining method that specifically detects covalently bound heme [109]. The 10 and 6 kDa proteins were the most abundant of these as judged by the heme staining method. In order to investigate the c-type cytochromes separately, ion exchange chromatography was performed in two steps. In the first step a cation exchange column was used resulting in retention of the 10 kDa cytochrome. No other cytochromes were retained on this medium. In the second chromatography step, using an anion exchange column, the 6 kDa protein could be isolated from the other cytochromes. Our attempts to separate the c cytochromes 8, 9 and 20 kDa in size were, however, unsuccessful. When working with the C subunit of chlorate reductase [paper II and paper III] we identified two more open reading frames immediately downstream the gene cluster for chlorate metabolism. The ORF closest to the gene cluster was

21
suggested by BLAST and Pfam to encode a soluble class 1 c-type cytochrome. The ORF further downstream was identified as mobB gene, which encodes a protein that participates in the maturation process of molybdopterin, which is a co-factor of the A subunit of chlorate reductase. The identification of the gene encoding a soluble c-type cytochrome in close proximity to the gene cluster for chlorate reductase, taken together with the results from tests of reduced periplasm, prompted an investigation of the gene product as an electron donor for chlorate reductase. For heterologous expression, the gene encoding the c-type cytochrome was amplified by PCR and ligated into the expression vector pET-26b. The predicted 19-residue signal peptide was replaced by a pelB leader peptide, for export to the periplasm. Also, this construct included a C terminal 6-residue His-tag for subsequent purification by IMAC. The ligation product and pEC86, harboring genes encoding proteins responsible for the cytochrome c maturation process [110], were then co-transformated into the E. coli expression hosts NovaBlue(DE3) and BL21(DE3). Expression in NovaBlue(DE3) gave only small amount of the recombinant protein, with very low levels of incorporated heme. Expression in BL21(DE3) gave, however, considerably higher level of expression, but in this case the target apo-protein ended up as inclusion bodies in the cytoplasm. From these results, it was decided to attempt production of the gene product expressed as inclusion bodies, followed by denaturation, purification, refolding and heme-reconstitution. Using IMAC, the denatured urea-solubilized target protein could be purified in one step. The purified protein was then refolded by dilution in cold refolding buffer. In prokaryotic bacteria, c-type cytochromes are normally exported in their apo-form to the periplasm via the secretory pathway (Sec). The heme-cofactor is separately translocated over the inner-membrane and then covalently attached to the cytochrome by the ccm system [111]. Because the target protein was expressed as inclusion bodies in the cytosol, it lacks the heme-group. Therefore the heme needs to be inserted and covalently attached either during or after the refolding process. In the present case, reconstitution after refolding was the most successful approach. Reduced heme was added to the refolded protein in a 1:1 ratio. The reconstitution could be followed spectrofotomerically by observing the optical spectrum of the heme group. Immediately after the addition of heme, the α-peak was found at 556.5 nm indicating heme incorporation, but no formation of thioether bonds. During the course of seven days the α-peak shifted to 553 nm. Pyridine hemochrome analysis [112],

22
produced an α-peak at 551 nm which is characteristic for c-type cytochromes where both the vinyl groups have reacted with cysteine residues producing thioether bonds [113, 114]. The heme content was 28%. In order to test the recombinant protein and the isolated or partly isolated native c cytochromes as electron donors to periplasmic chlorate reductase, the proteins were treated with nitrogen and completely reduced with small portions of dithionite. As source of chlorate reductase activity, catalytic amounts of purified native chlorate reductase or I. dechloratans cell homogenate was added to the reduced samples. Chlorate dependent oxidation of reduced c-type cytochromes was followed with UV/VIS spectroscopy by observing the heme α-band. The effect of oxygen on the oxidation state of the c cytochromes was also investigated. As mentioned in the introduction, I. dechloratans is reported to be cytochrome c-oxidase positive with a strictly respiratory metabolism. Thus, adding cell homogenate to the reduced sample we also add the membrane-bound terminal oxidase and reconstruct the aerobic electron transport pathway. If any of the reduced c-type cytochromes delivers electrons to the terminal oxidase, they will be oxidized upon addition of oxygen. Figure 8 shows the chlorate-dependent and the oxygen-dependent oxidation of soluble periplasmic c-type cytochromes. The 6 kDa protein was oxidized on addition of chlorate and on addition of oxygen, suggesting that this protein is able to deliver electrons both to chlorate reductase and the terminal oxidase. The 10 kDa protein, on the other hand, was only oxidized upon addition of oxygen, suggesting that this protein donates electrons to the terminal oxidase only. The mixture of 6, 8, 9, 10 and 20 kDa protein was fully oxidized when oxygen was added, suggesting electron delivery to membrane-bound terminal oxidase by all the components present. Addition of chlorate produced oxidation of 45 % of the c cytochromes in the mixture. The 6 kDa c cytochrome is the dominating component in this mixture, and the result can be accounted for by the presence of this protein. Due to the difficulties of isolate the 8-, 9- and 20 kDa proteins, it was not possible to verify their individual ability to serve as electron donor to chlorate reductase or to the terminal oxidase. The recombinant c-type cytochrome was found to be very sensitive to air oxidation. It was therefore not possible to investigate its role as electron donor to the terminal oxidase. The addition of chlorate to the recombinant c cytochrome, in the presence of chlorate reductase, did not produce any spectral changes indicating electron transport to periplasmic chlorate reductase.
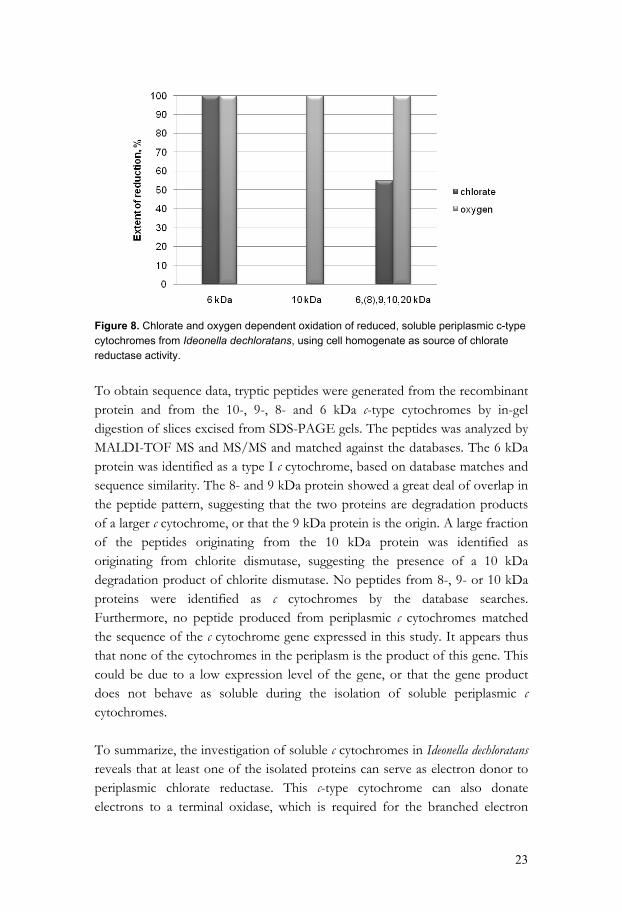
23
Figure 8. Chlorate and oxygen dependent oxidation of reduced, soluble periplasmic c-type cytochromes from Ideonella dechloratans, using cell homogenate as source of chlorate reductase activity. To obtain sequence data, tryptic peptides were generated from the recombinant protein and from the 10-, 9-, 8- and 6 kDa c-type cytochromes by in-gel digestion of slices excised from SDS-PAGE gels. The peptides was analyzed by MALDI-TOF MS and MS/MS and matched against the databases. The 6 kDa protein was identified as a type I c cytochrome, based on database matches and sequence similarity. The 8- and 9 kDa protein showed a great deal of overlap in the peptide pattern, suggesting that the two proteins are degradation products of a larger c cytochrome, or that the 9 kDa protein is the origin. A large fraction of the peptides originating from the 10 kDa protein was identified as originating from chlorite dismutase, suggesting the presence of a 10 kDa degradation product of chlorite dismutase. No peptides from 8-, 9- or 10 kDa proteins were identified as c cytochromes by the database searches. Furthermore, no peptide produced from periplasmic c cytochromes matched the sequence of the c cytochrome gene expressed in this study. It appears thus that none of the cytochromes in the periplasm is the product of this gene. This could be due to a low expression level of the gene, or that the gene product does not behave as soluble during the isolation of soluble periplasmic c cytochromes. To summarize, the investigation of soluble c cytochromes in Ideonella dechloratans reveals that at least one of the isolated proteins can serve as electron donor to periplasmic chlorate reductase. This c-type cytochrome can also donate electrons to a terminal oxidase, which is required for the branched electron

24
flow during chlorate respiration [paper V]. This is the first study reporting chlorate reduction with soluble c cytochromes as electron transporter between (per)chlorate reductase and membrane-bound respiratory complexes. The native counterpart of the recombinant c-type cytochrome could not be identified by MS. Also, no chlorate-dependent oxidation the recombinant version of the protein could be detected. Although the inability of the recombinant c cytochrome to serve as electron donor could be due to incorrect refolding or heme incorporation, the role of this c cytochrome in the Ideonella dechloratans respiration, and the significance of the localization of the corresponding gene, close to the gene cluster for chlorate metabolism, still remains to be clarified.

25
Conclusions Chlorate reductase and chlorite dismutase have been described at the gene level, and are found in close proximity constituting a gene cluster for chlorate metabolism. The gene encoding chlorite dismutase has, for the first time, been expressed recombinantly as a functional unit. The C subunit of chlorate reductase has been cloned and expressed as inclusion bodies, followed by purification in the denatured state and refolding. The heme cofactor was reconstituted and the recombinant subunit showed native-like spectral properties. This result confirms the notion of the C subunit as the heme containing subunit. The investigation of the soluble periplasmic c cytochromes of Ideonella dechloratans shows that at least one of the cytochromes under study is able to deliver electrons to the periplasmic chlorate reductase. Adjacent to the gene cluster for chlorate metabolism a gene encoding a soluble c-type cytochrome was found. The protein corresponding to this gene could not be detected in the periplasmic extract, and a recombinant reconstituted version of the protein could not be demonstrated to function as electron donor to periplasmic chlorate reductase. Further investigations are needed to find out the role of this cytochrome in the Ideonella dechloratans respiration.

26
Tack Under min tid på Karlstads universitet har jag kanske inte alltid varit den hängivne forskarstuderande som man kan önska. Mitt fokus har ibland flyttats utanför ”murarna” på grund av exempelvis barnafödande, husbygge och ambitioner som egenföretagare. Ändock sitter jag här idag med en avhandling färdig för tryck och utan allt för många gråa hårstrån i skägget. Det är många jag vill tacka som på olika sätt bidragit till detta. Först av allt vill jag rikta mitt stora tack till min handledare Thomas Nilsson. Ditt stöd och engagemang har varit ovärderligt för mig. Mina biträdande handledare, Birgitta Sundström och Leif Jönsson, vill jag tacka för goda råd, både praktiska och teoretiska. Anna, tack för gott samarbete. Nu när jag lämnar skutan får du ensam brottas med vår lilla “bagge” nere på labbet. Jag hoppas att du ändå hinner med många trevliga kryssningar över Östersjön. Jag vill också speciellt tacka min forne doktorandkollega Helena. På något sätt känns det som att det var du som uppfostrade mig till att bli en, förhoppningsvis, hyfsat bra forskarstuderande. Tack alla vänner och kollegor vid Avdelningen för kemi, speciellt mina forna och nuvarande doktorandkollegor, Mårten, Björn E, Tina, Björn A, Sandra, Jeanette, Marcus, Rozbeh och Zeki. Mina svärpäron Alf och Barbro, ni finns där för oss, och det är jag oerhört tacksam för. Tack Mamma! Jag hade tänkt skriva ett eget kapitel till dig – “Kloratrespiration för Mamma”. Tyvärr hann jag inte det. Jag hoppas att jag någon annan gång ska hinna förklara vad jag egentligen gjort. Mina bröder tillika kompanjoner Jens, Jonas och Jörgen. Mycket har hänt på ett par år, och framtiden ser ljus ut.

27
Maria, du är klippan jag vilar mitt liv mot. Utan ditt stöd och Din förståelse hade jag nog inte klarat det här. Jag älskar dig så! Alva och Julia, mina två irrbloss, vad vore jag utan er? Slutligen vill jag dedikera denna avhandling till min Far. Även om han gick bort för över tio år sedan finns han ändå alltid i mitt hjärta.

28
References
[1] Lovley D R, Coates J D (2000), Novel forms of anaerobic respiration of
environmental relevance, Curr Opin Microbiol 3 252-256.
[2] McEwan A G, Ridge J P, McDevitt C A, Hugenholtz P (2002), The DMSO Reductase Family of Microbial Molybdenum Enzymes; Molecular Properties and Role in the Dissimilatory Reduction of Toxic Elements, Geomicrobiol J 19 3-19.
[3] Krafft T, Macy J M (1998), Purification and characterization of the respiratory arsenate reductase of Chrysiogenes arsenatis, Eur J Biochem 255 647-653.
[4] Urbansky E (2002), Perchlorate as an Environmental Contaminant, Environ Sci Regul Int 9 187-192.
[5] Urbansky E T (1998), Perchlorate Chemistry: Implications for Analysis and Remediation, Bioremediat J 2 81-95.
[6] Goksöyr J (1952), On the effect of chlorate upon the nitrate reduction of plants II. The effect upon the nitrate-reducing system in Escherichia coli, Physiol Plant 5 228-240.
[7] Motzer W E (2001), Perchlorate: Problems, Detection, and Solutions, Environ Forensics 2 301-311.
[8] Wallace W, Ward T, Breen A, Attaway H (1996), Identification of an anaerobic bacterium which reduces perchlorate and chlorate as Wollinella succinogenes, J Ind Microbiol 16 68-72.
[9] Coates J D, Achenbach L A (2004), Microbial Perchlorate Reduction: Rocket-Fuelled Metabolism, Nature Reviews: Microbiology 2 569-580.
[10] van Wijk D J, Hutchinson T H (1995), The ecotoxicity of chlorate to aquatic organisms: a critical review, Ecotoxicol Environ Saf 32 244-253.
[11] Åslander A (1928), Experiments on the eradication of canada thistle, Cirsum arvense, with chlorates and other herbicides, J Agricult Res 36 915-934.
[12] Auchter R J, Preparation of bleacing chemicals, in: Rapson W H (Ed.), The bleaching of pulp, TAPPI, New York, 1963, pp. 379-403.
[13] Bergnor E, Germgård U, Kolar J J, Lindgren B O (1987), Formation of chlorate in chlorine dioxide bleaching, Cellulose Chem Technol 21 307-314.
[14] Germgård U, Teder A, Tormund D (1981), Chlorate formation during chlorine dioxide bleaching of softwood kraft pulp, Pap Puu 63 127-133.
[15] Condie L W (1986), Toxicological Problems Associated With Chlorine Dioxide, Research technol 73-78.

29
[16] Rosemarin A, Lehtinen K-J, Notini M, Mattson J (1994), Effects of pulp mill chlorate on baltic sea algae, Environ Pollut 85 3-13.
[17] Hofstra J J (1977), Chlorate Toxicity and Nitrate Reductase Activity in Tomato Plants, Physiol Plant 41 65-69.
[18] Liljeström S, Åberg B (1966), Studies on the Mechanism of Chlorate Toxicity, Lantbrukshögsk Ann Sweden 32 93-107.
[19] Stanbury J B, Wyngaarden J B (1952), Effect of perchlorate on the human thyroid gland, Metabolis 1 533-539.
[20] Achenbach L A, Michaelidou U, Bruce R A, Fryman J, Coates J D (2001), Dechloromonas agitata gen. nov., sp. nov. and Dechlorosoma suillum gen. nov., sp. nov., two novel environmentally dominant (per)chlorate-reducing bacteria and their phylogenetic position, Int J Syst Evol Microbiol 51 527-533.
[21] Wolff J (1998), Perchlorate and the Thyroid Gland, Pharmacol Rev 50 89.
[22] Logan B E (1998), A review of chlorate- and perchlorate-respiring microorganisms, Biorem J 2 69-79.
[23] Malmqvist Å, Welander T, Gunnarsson L (1991), Anaerobic growth of microorganisms with chlorate as an electron acceptor, Appl Environ Microbiol 57 2229-2232.
[24] Malmqvist Å, Welander T (1992), Anaerobic removal of chlorate from bleach effluents, Wat Sci Tech 25 237-242.
[25] Rikken G B, Kroon A G, van Ginkel C G (1996), Transformation of (per)chlorate into chloride by a newly isolated bacterium: reduction and dismutation, Appl Microbiol Biotech 45 420-426.
[26] Bruce R A, Achenbach L A, Coates J D (1999), Reduction of (per)chlorate by a novel organism isolated from paper mill waste, Environ Microbiol 1 319-329.
[27] Wolterink A F, Jonker A B, Kengen S W, Stams A J (2002), Pseudomonas chloritidismutans sp. nov., a non-denitrifying, chlorate-reducing bacterium, Int J Syst Evol Microbiol 52 2183-2190.
[28] Coates J D, Chakraborty R, Lack J G, O'Connor S M, Cole K A, Bender K S, Achenbach L A (2001), Anaerobic benzene oxidation coupled to nitrate reduction in pure culture by two strains of Dechloromonas, Nature 411 1039-1043.
[29] Balk M, van Gelder T, Weelink S A, Stams A J M (2008), (Per) chlorate Reduction by the Thermophilic Bacterium Moorella perchloratireducens sp. nov., Isolated from Underground Gas Storage, Appl Environ Microbiol 74 403-409.

30
[30] Malmqvist Å, Welander T, Moore E, Ternström A, Molin G, Stenström I (1994), Ideonella dechloratans gen.nov., sp.nov., a new bacterium capable of growing anaerobically with chlorate as an electron acceptor, System Appl Microbiol 17 58-64.
[31] Chakraborty R, Coates J D (2004), Anaerobic degradation of monoaromatic hydrocarbons, Appl Microbiol Biotechnol 64 437-446.
[32] Bender K S, Rice M R, Fugate W H, Coates J D, Achenbach L A (2004), Metabolic primers for detection of (Per)chlorate-reducing bacteria in the environment and phylogenetic analysis of cld gene sequences, Appl Environ Microbiol 70 5651-5658.
[33] Nozawa-Inoue M, Jien M, Hamilton N S, Stewart V, Scow K M, Hristova K R (2008), Quantitative Detection of Perchlorate-Reducing Bacteria by Real-Time PCR Targeting the Perchlorate Reductase Gene?, Appl Environ Microbiol 74 1941-1944.
[34] Herman D C, Frankenberger W T J (1999), Bacterial Reduction of Perchlorate and Nitrate in Water, J Environ Qual 28 1018-1024.
[35] Oltmann L F, Reijnders W N, Stoughamer A H (1976), Characterization of purified nitrate reductase A and chlorate reductase C from Proteus mirabilis, Arch Microbiol 111 25-35.
[36] Kengen S W, Rikken G B, Hagen W R, van Ginkel C G, Stams A J (1999), Purification and characterization of (per)chlorate reductase from the chlorate-respiring strain GR-1, J Bacteriol 181 6706-6711.
[37] Okeke B C, Frankenberger W T (2003), Molecular analysis of a perchlorate reductase from a perchlorate-respiring bacterium Perc1ace, Microbiol Res 158 337-344.
[38] Wolterink A F, Schiltz E, Hagedoorn P L, Hagen W R, Kengen S W, Stams A J (2003), Characterization of the chlorate reductase from Pseudomonas chloritidismutans, J Bacteriol 185 3210-3213.
[39] Bender K S, Shang C, Chakraborty R, Belchik S M, Coates J D, Achenbach L A (2005), Identification, Characterization, and Classification of Genes Encoding Perchlorate Reductase, J Bacteriol 187 5090-5096.
[40] Chakraborty R, O'Connor S M, Chan E, Coates J D (2005), Anaerobic Degradation of Benzene, Toluene, Ethylbenzene, and Xylene Compounds by Dechloromonas Strain RCB, Appl Environ Microbiol 71 8649.
[41] van Ginkel C G, Rikken G B, Kroon A G, Kengen S W (1996), Purification and characterization of chlorite dismutase: a novel oxygen-generating enzyme, Arch Microbiol 166 321-326.

31
[42] Coates J D, Michaelidou U, Bruce R A, O'Connor S M, Crespi J N, Achenbach L A (1999), Ubiquity and diversity of dissimilatory (per)chlorate-reducing bacteria, Appl Environ Microbiol 65 5234-5241.
[43] Stenklo K, Danielsson Thorell H, Bergius H, Aasa R, Nilsson T (2001), Chlorite dismutase from Ideonella dechloratans, J Biol Inorg Chem 6 601-607.
[44] Streit B R, DuBois J L (2008), Chemical and Steady-State Kinetic Analyses of a Heterologously Expressed Heme Dependent Chlorite Dismutase, Biochemistry 47 5271-5280.
[45] Hagedoorn P L, De Geus D C, Hagen W R (2002), Spectroscopic characterization and ligand-binding properties of chlorite dismutase from the chlorate respiring bacterial strain GR-1, Eur J Biochem 269 4905-4911.
[46] Welter R, Gu L Q, Yu L, Yu C A, Rumbley J, Gennis R B (1994), Identification of the ubiquinol-binding site in the cytochrome bo3-ubiquinol oxidase of Escherichia coli, J Biol Chem 269 28834-28838.
[47] Richardson D J, Berks B C, Russell D A, Spiro S, Taylor C J (2001), Functional, biochemical and genetic diversity of prokaryotic nitrate reductases, Cell Mol Life Sci 58 165-178.
[48] Garcia-Horsman J A, Barquera B, Rumbley J, Ma J, Gennis R B (1994), The superfamily of heme-copper respiratory oxidases, J Bacteriol 176 5587-5600.
[49] Carr G J, Page M D, Ferguson S J (1989), Distinction from the nitrite reductase that catalyses synthesis of nitric oxide and evidence from trapping experiments for nitric oxide as a free intermediate during denitrification, Eur J Biochem 179 683-692.
[50] Nicholls D G, Ferguson S J, Bioenergetics 2, Academic Press 1982.
[51] Richardson D J, Watmough N J (1999), Inorganic nitrogen metabolism in bacteria, Curr Opin Chem Biol 3 207-219.
[52] Moser D P, Nealson K H (1996), Growth of the Facultative Anaerobe Shewanella putrefaciens by Elemental Sulfur Reduction, Appl Environ Microbiol 62 2100.
[53] Beliaev A S, Saffarini D A (1998), Shewanella putrefaciens mtrB Encodes an Outer Membrane Protein Required for Fe (III) and Mn (IV) Reduction, J Bacteriol 180 6292.
[54] Schwalb C, Chapman S K, Reid G A (2002), The membrane-bound tetrahaem c-type cytochrome CymA interacts directly with the soluble fumarate reductase in Shewanella, Biochem Soc Trans 30 658-662.
[55] Schwalb C, Chapman S K, Reid G A (2003), The tetraheme cytochrome CymA is required for anaerobic respiration with dimethyl sulfoxide and nitrite in Shewanella oneidensis, Biochemistry (Washington) 42 9491-9497.

32
[56] McDevitt C A, Hugenholtz P, Hanson G R, McEwan A G (2002), Molecular analysis of dimethyl sulphide dehydrogenase from Rhodovulum sulfidophilum: its place in the dimethyl sulphoxide reductase family of microbial molybdopterin-containing enzymes, Mol Microbiol 44 1575-1587.
[57] Lledo B, Martinez-Espinosa R M, Marhuenda-Egea F C, Bonete M J (2004), Respiratory nitrate reductase from haloarchaeon Haloferax mediterranei: biochemical and genetic analysis, Biochim Biophys Acta 1674 50-59.
[58] Yoshimatsu K, Iwasaki T, Fujiwara T (2002), Sequence and electron paramagnetic resonance analyses of nitrate reductase NarGH from a denitrifying halophilic euryarchaeote Haloarcula marismortui, FEBS Lett 516 145-150.
[59] Studier F W (1991), Use of bacteriophage T7 lysozyme to improve an inducible T7 expression system, J Mol Biol 219 37-44.
[60] Dubendorff J W, Studier F W (1991), Controlling basal expression in an inducible T7 expression system by blocking the target T7 promoter with lac repressor, J Mol Biol 219 45-59.
[61] Georgiou G, Valax P (1996), Expression of correctly folded proteins in Escherichia coli, Curr Opin Biotechnol 7 190-197.
[62] Blackwell J R, Horgan R (1991), A novel strategy for production of a highly expressed recombinant protein in an active form, FEBS Lett 295 10-12.
[63] Smith D B, Johnson K S (1988), Single-step purification of polypeptides expressed in Escherichia coli as fusions with glutathione S-transferase, Gene 67 31-40.
[64] di Guan C, Li P, Riggs P D, Inouye H (1988), Vectors that facilitate the expression and purification of foreign peptides in Escherichia coli by fusion to maltose-binding protein, Gene 67 21-30.
[65] LaVallie E R, DiBlasio E A, Kovacic S, Grant K L, Schendel P F, McCoy J M (1993), A thioredoxin gene fusion expression system that circumvents inclusion body formation in the E. coli cytoplasm, Biotechnology (N Y) 11 187-193.
[66] Kiefhaber T, Rudolph R, Kohler H H, Buchner J (1991), Protein aggregation in vitro and in vivo: a quantitative model of the kinetic competition between folding and aggregation, Biotechnology (N Y) 9 825-829.
[67] Cardamone M, Puri N K, Brandon M R (1995), Comparing the refolding and reoxidation of recombinant porcine growth hormone from a urea denatured state and from Escherichia coli inclusion bodies, Biochemistry 34 5773-5794.
[68] Middelberg A P (2002), Preparative protein refolding, Trends Biotechnol 20 437-443.

33
[69] Clark E D B (1998), Refolding of recombinant proteins, Curr Opin Biotechnol 9 157-163.
[70] Lilie H, Schwarz E, Rudolph R (1998), Advances in refolding of proteins produced in E. coli, Curr Opin Biotechnol 9 497-501.
[71] Hargrove M S, Barrick D, Olson J S (1996), The association rate constant for heme binding to globin is independent of protein structure, Biochemistry 35 11293-11299.
[72] Robinson C R, Liu Y, Thomson J A, Sturtevant J M, Sligar S G (1997), Energetics of heme binding to native and denatured states of cytochrome b562, Biochemistry 36 16141-16146.
[73] Rau H K, DeJonge N, Haehnel W (1998), Modular synthesis of de novo-designed metalloproteins for light-induced electron transfer, Proc Natl Acad Sci U S A 95 11526-11531.
[74] Robertson D E, Farid R S, Moser C C, Urbauer J L, Mulholland S E, Pidikiti R, Lear J D, Wand A J, DeGrado W F, Dutton P L (1994), Design and synthesis of multi-haem proteins, Nature 368 425-432.
[75] Gruenke L D, Sun J, Loehr T M, Waskell L (1997), Resonance Raman spectral properties and stability of manganese protoporphyrin IX cytochrome b5, Biochemistry 36 7114-7125.
[76] Kroliczewski J, Szczepaniak A (2002), In vitro reconstitution of the spinach chloroplast cytochrome b6 protein from a fusion protein expressed in Escherichia coli, Biochim Biophys Acta 1598 177-184.
[77] Daltrop O, Ferguson S J (2003), Cytochrome c Maturation. The in vitro reactions of horse heart apocytochrome c and Paracoccus dentrificans apocytochrome c550 with heme., J Biol Chem 278 4404-4409.
[78] Cohen S L, Chait B T (1997), Mass Spectrometry of Whole Proteins Eluted from Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis Gels, Anal Biochem 247 257-267.
[79] Stutz H (2005), Advances in the analysis of proteins and peptides by capillary electrophoresis with matrix-assisted laser desorption/ionization and electrospray-mass spectrometry detection, Electrophoresis 26 1254-1290.
[80] Bayer E (2001), Recent advances in capillary electrophoresis/electrospray-mass spectrometry, Electrophoresis 22 1251-1266.
[81] Mann M, Hendrickson R C, Pandey A (2001), Analysis of proteins and proteomes by mass spectrometry, Annu Rev Biochem 70 437-473.
[82] Herbert C G, Johnstone R A W, Mass Spectrometry Basics, CRC Press 2003.
[83] Roepstorff P, Fohlman J (1984), Proposal for a common nomenclature for sequence ions in mass spectra of peptides, Biomed Mass Spectrom 11 601.

34
[84] Chen P, Nie S, Mi W, Wang X C, Liang S P (2004), De novo sequencing of tryptic peptides sulfonated by 4-sulfophenyl isothiocyanate for unambiguous protein identification using post-source decay matrix-assisted laser desorption/ionization mass spectrometry, Rapid Commun Mass Spectrom 18 191-198.
[85] Canas B, Lopez-Ferrer D, Ramos-Fernandez A, Camafeita E, Calvo E (2006), Mass spectrometry technologies for proteomics, Brief Funct Genomic Proteomic 4 295-320.
[86] Kery V, Elleder D, Kraus J P (1995), Delta-aminolevulinate increases heme saturation and yield of human cystathionine beta-synthase expressed in Escherichia coli, Arch Biochem Biophys 316 24-29.
[87] Rivera M, Qiu F, Bunce R A, Stark R E (1999), Complete isomer-specific 1H and 13C NMR assignments of the heme resonances of rat liver outer mitochondrial membrane cytochrome b5, J Biol Inorg Chem 4 87-98.
[88] Scopes R K, Protein Purification: principles and practise - 3rd ed., Springer-Verlag New York, Inc. 1994.
[89] Danielsson Thorell H, Beyer N H, Heegaard N H, Ohman M, Nilsson T (2004), Comparison of native and recombinant chlorite dismutase from Ideonella dechloratans, Eur J Biochem 271 3539-3546.
[90] Kristjansson J K, Hollocher T C (1980), First practical assay for soluble nitrous oxide reductase of denitrifying bacteria and a partial kinetic characterization, J Biol Chem 255 704-707.
[91] Jones R W, Garland P B (1977), Sites and specificity of the reaction of bipyridylium compounds with anaerobic respiratory enzymes of Escherichia coli, Biochem J 164 199-211.
[92] Krafft T, Bowen A, Theis F, Macy J M (2000), Cloning and sequencing of the genes encoding the periplasmic-cytochrome b-containing selenate reductase of Thauera selenatis, DNA Seq. 10 365-377.
[93] Rabus R, Kube M, Beck A, Widdel F, Reinhardt R (2002), Genes involved in the anaerobic degradation of ethylbenzene in a denitrifying bacterium, strain EbN1, Arch Microbiol 178 506-516.
[94] Voordouw G (2000), A universal system for the transport of redox proteins: early roots and latest developments, Biophys Chem 86 131-140.
[95] Rodrigue A, Chanal A, Beck K, Muller M, Wu L F (1999), Co-translocation of a periplasmic enzyme complex by a hitchhiker mechanism through the bacterial tat pathway, J Biol Chem 274 13223-13228.
[96] Pugsley A P (1993), The complete general secretory pathway in gram-negative bacteria, Microbiol Rev 57 50-108.

35
[97] McDevitt C A, Hanson G R, Noble C J, Cheesman M R, McEwan A G (2002), Characterization of the redox centers in dimethyl sulfide dehydrogenase from Rhodovulum sulfidophilum, Biochemistry 41 15234-15244.
[98] Kniemeyer O, Heider J (2001), Ethylbenzene dehydrogenase, a novel hydrocarbon-oxidizing molybdenum/iron-sulfur/heme enzyme, J Biol Chem 276 21381-21386.
[99] Afshar S, Johnson E, de Vries S, Schroder I (2001), Properties of a thermostable nitrate reductase from the hyperthermophilic archaeon Pyrobaculum aerophilum, J Bacteriol 183 5491-5495.
[100] Blasco F, Dos Santos J P, Magalon A, Frixon C, Guigliarelli B, Santini C L, Giordano G (1998), NarJ is a specific chaperone required for molybdenum cofactor assembly in nitrate reductase A of Escherichia coli, Mol Microbiol 28 435-447.
[101] Tobias J W, Shrader T E (1991), The N-end rule in bacteria, Science 254 1374-1377.
[102] Hirel P H, Schmitter M J, Dessen P, Fayat G, Blanquet S (1989), Extent of N-terminal methionine excision from Escherichia coli proteins is governed by the side-chain length of the penultimate amino acid, Proc Natl Acad Sci U S A 86 8247-8251.
[103] Cleland J L, Randolph T W (1992), Mechanism of polyethylene glycol interaction with the molten globule folding intermediate of bovine carbonic anhydrase B, J Biol Chem 267 3147-3153.
[104] Hevehan D L, De Bernardez Clark E (1997), Oxidative renaturation of lysozyme at high concentrations, Biotechnol Bioeng 54 221-230.
[105] Lee G Y, Zhu J, Yu L, Yu C A (1998), Reconstitution of cytochrome b-560 (QPs1) of bovine heart mitochondrial succinate-ubiquinone reductase, Biochim Biophys Acta 1363 35-46.
[106] Varadarajan R, Szabo A, Boxer S G (1985), Cloning, expression in Escherichia coli, and reconstitution of human myoglobin, Proc Natl Acad Sci U S A 82 5681-5684.
[107] Ozols J, Strittmatter P (1964), The Interaction of Porphyrins and Metalloporhyrins with Apocytochrome Beta-5, J Biol Chem 239 1018-1023.
[108] Roldan M D, Sears H J, Cheesman M R, Ferguson S J, Thomson A J, Berks B C, Richardson D J (1998), Spectroscopic Characterization of a Novel Multiheme c-Type Cytochrome Widely Implicated in Bacterial Electron Transport, J Biol Chem 273 28785-28790.
[109] Thomas P E, Ryan D, Levin W (1976), An improved staining procedure for the detection of the peroxidase activity of cytochrome P-450 on sodium dodecyl sulfate polyacrylamide gels, Anal Biochem 75 168-176.

36
[110] Arslan E, Schulz H, Zufferey R, Künzler P, Thöny-Meyer L (1998), Overproduction of theBradyrhizobium japonicum c-Type Cytochrome Subunits of thecbb3Oxidase inEscherichia coli, Biochem Biophys Res Commun 251 744-747.
[111] Thony-Meyer L (1997), Biogenesis of respiratory cytochromes in bacteria, Microbiol Mol Biol Rev 61 337-376.
[112] Berry E A, Trumpower B L (1987), Simultaneous determination of hemes a, b, and c from pyridine hemochrome spectra, Anal Biochem 161 1-15.
[113] Bartsch R G (1971), Cytochromes: bacterial, Methods Enzymol 23 344–363.
[114] Falk J E, Porphyrins and metalloporphyrins: their general, physical and coordination chemistry, and laboratory methods, Elsevier Amsterdam: 1964.

Karlstad University StudiesISSN 1403-8099
ISBN 978-91-7063-196-2
Enzymes and electron transport in microbial chlorate respiration
Microbial chlorate respiration plays an important role in the turnover of oxochlorates in nature and in industrial waste management. This thesis deals with the characterization of the molecular components of chlorate respiration in Ideonella dechloratans. Chlorate respiration utilizes two soluble periplasmic enzymes, chlorate reductase and chlorite dismutase, to convert chlorate to chloride and oxygen. The genes encoding the enzymes participating in the chlorate degradation have been sequenced, and are found in close proximity, forming a gene cluster for chlorate metabolism. This work also includes the successful recombinant expression of three genes from Ideonella dechloratans. Two of the gene products, chlorite dismutase and the C subunit of chlorate reductase, participate in the chlorate respiration. The third gene, which is found close to the gene cluster for chlorate metabolism, encodes a soluble c-type cytochrome. The localization of the gene suggests the corresponding protein as a candidate for a role as electron donor to chlorate reductase. Also, the role of soluble periplasmic c cytochromes of Ideonella dechloratans in chlorate respiration was studied. At least one of the soluble c cytochromes was found capable of serving as electron donor for chlorate reduction. This c cytochrome, and several others, can also donate electrons to a terminal oxidase for subsequent reduction of oxygen, as required for the branched electron flow during chlorate respiration.