enzymatic responses of the riceland prawn, macrobrachium lanchesteri, to chlorpyrifos exposure
TRANSCRIPT

Biologia 67/4: 762—766, 2012Section ZoologyDOI: 10.2478/s11756-012-0061-6
Enzymatic responses of the riceland prawn,Macrobrachium lanchesteri, to chlorpyrifos exposure
Wuttipong Tongbai, Rapee Boonplueng & Praneet Damrongphol*
Department of Biology, Faculty of Science, Mahidol University, Bangkok 10400, Thailand; e-mail: [email protected]
Abstract: Cholinesterase (ChE) was characterized in whole bodies of adult riceland prawns, Macrobrachium lanches-teri, based on substrate preference and inhibitor sensitivity. M. lanchesteri ChE activity was mainly attributable toacetylcholinesterase (AChE) since it preferentially hydrolyzed an AChE-specific substrate, acetylthiocholine iodide, overs-butyrylthiocholine iodide and was sensitive to 1,5-bis-(4-allyldimethyl-ammoniumphenyl)-pentan-3-one dibromide, a spe-cific inhibitor for AChE. The effect of chlorpyrifos exposure on M. lanchesteri were also investigated. The 24, 48, 72 and 96 hLC50 values for chlorpyrifos were 3.37, 2.76, 2.58 and 2.53 µg L−1, respectively. Based on these values, adult M. lanchesteriwere exposed to 0.5, 1.5, 2.5 and 3.5 µg chlorpyrifos L−1 for 24, 48, 72 and 96 h. Chlorpyrifos appeared to induce drasticenzymatic responses in M. lanchesteri. AChE activity was inhibited after 24 h of exposure in a dose- and time-dependentmanner. The levels of lipid peroxidation increased significantly after 24 h of exposure. However, the activities of the antioxi-dant enzyme, catalase, and the detoxification enzyme, glutathione S-transferase, were reduced. In conclusion, M. lanchesteriis sensitive to chlorpyrifos and can serve as a bioindicator species for freshwater environmental assessment.
Key words: acetylcholinesterase; lipid peroxidation; catalase; glutathione S-transferase; chlorpyrifos; prawn; Macro-brachium lanchesteri
Introduction
Inhibition of cholinesterase (ChE) activity in organ-isms has been widely used as a sensitive biomarkerfor determining organophosphate and carbamate pes-ticide exposure in surrounding environment (Kirbyet al. 2000). Although pesticide residues in the en-vironments may be undetected by chemical assays,the inhibition of ChE activity in organisms maystill be detected (Fulton & Key 2001). Like verte-brates, crustaceans contain two major forms of ChE,acetylcholinesterase (AChE; EC 3.1.1.7) and butyryl-cholinesterase (BChE; EC 3.1.1.8). The predominantform of ChE present in an organism varies amongspecies and tissues. The type of ChE can be differenti-ated based on substrate preference and inhibitor sen-sitivity. AChE preferentially hydrolyzes acetylcholine(ACh) whereas BChE preferentially hydrolyzes butyryl-choline but also hydrolyzes propionylcholine and AChto a lesser extent. All forms of ChE are inhibited byeserine sulfate. However, AChE is selectively inhibitedby 1,5-bis (4-allyldimethyl-ammoniumphenyl)-pentan-3-one dibromide (BW284c51) but not by tetraisopropy-lpyrophosphoramide (iso-OMPA). BChE is selectivelyinhibited by iso-OMPA (Massoulié et al. 1993; Key &Fulton 2002; García-de la Parra et al. 2006; Xuereb etal. 2007). Inhibition of AChE by toxicants causes an ac-cumulation of ACh at cholinergic synapses resulting in
hyperactivation of postsynaptic cholinergic receptors.Behavioral abnormalities such as tremors, convulsionsand paralysis, and finally death of an organism are in-duced (Day & Scott 1990).Chlorpyrifos (O,O-diethyl 0-3,5,6-trichloro-2-pyri-
dyl phosphorothioate) is an organophosphate pesticidethat has been extensively used for agricultural pest con-trol. Chlorpyrifos exerts its toxicity by inhibiting ChEactivity (Xuereb et al. 2007). In addition to its neu-rotoxicity, chlorpyrifos induces oxidative stress in anorganism either by generating reactive oxygen species(ROS) or interfering with the antioxidant defense mech-anism (Kavitha & Venkateswara Rao 2008). ROS in-cluding superoxide anion radicals, hydrogen peroxide(H2O2) and hydroxyl radicals, are highly reactive, thusdamaging biological molecules leading to lipid peroxi-dation (Livingstone 2001). Estimation of lipid peroxi-dation level by determining the production of thiobar-bituric acid reactive species (TBARS) is widely used asa biomarker for oxidative damage in organisms (Lack-ner 1998). Under normal conditions, ROS are elimi-nated by antioxidant enzymes such as catalase (CAT;EC.1.11.1.6), which decomposes H2O2 into water andO2 molecules (Diguiseppi & Fridovich 1984). An intox-icated organism may recover by the use of detoxifica-tion enzymes such as glutathione S-transferase (GST;EC.2.5.1.18), which catalyzes the conjugation of thethiol moiety of reduced glutathione with a variety of
* Corresponding author
c©2012 Institute of Zoology, Slovak Academy of Sciences

Responses of riceland prawn to chlorpyrifos 763
compounds bearing electrophilic centers, such as pesti-cides, thereby eliminating them from the cells (Ander-son 1997). Both antioxidant and detoxification enzymeactivities have been used as biomarkers for environmen-tal assessment.Crustaceans are sensitive to pesticides due to their
close phylogenetic relationship with insects. Ricelandprawns,Macrobrachium lanchesteri (de Man, 1911), aresmall freshwater prawns widely distributed in SoutheastAsia. They are commonly found in slow-flowing watersuch as ponds, canals or ricefields and serve as prey forfreshwater fish and also a common food source for localpeople. M. lanchesteri are subject to risk of pesticidecontamination owing to the heavy use of pesticides infood crops. Furthermore, M. lanchesteri can be easilyreared under laboratory conditions. It may therefore beuseful as a bioindicator species for monitoring freshwa-ter environmental contamination.The aims of the present study were to identify the
form of ChE present in adult M. lanchesteri based onsubstrate preference and inhibitor sensitivity. The effectof chlorpyrifos exposure on adult M. lanchesteri wasalso evaluated in term of the median lethal concentra-tion (LC50) values and enzymatic responses includinglipid peroxidation, AChE, CAT and GST.
Material and methods
ChemicalsAcetylthiocholine iodide (ATChI), s-butyrylthiocholine io-dide (BTChI), propionylthiocholine iodide (PTChI), eserinesulfate, BW284c51, iso-OMPA, 5,5-dithio-bis-2-nitrobenzo-ate (DTNB), 1-chloro-2,4-dinitrobenzene (CDNB), GSH, 2-thiobarbituric acid, 1,1,3,3-tetramethoxypropane and Coo-massie Brilliant Blue G-250 were obtained from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Butylated hy-droxytoluene, 30% H2O2 and trichloroacetic acid were ob-tained from BDH (Poole, England). Technical grade chlor-pyrifos (99.5% purity) was obtained from Dr. EhrenstorferGmbH (Augsburg, Germany).
Experimental prawnsAdult M. lanchesteri of 0.3–0.5 g in weight and 3.0–4.0 cmin length from canals in Supanburi province, Thailand werepurchased from a local market. They were acclimatized un-der laboratory conditions with a 12 h dark/12 h light periodin a 50×120×50 cm3 glass aquarium containing approxi-mately 200 L of water at 27 ± 1◦C for at least one week.
Preparation of tissue homogenatePrawns were euthanized by placing them at –20◦C for about10 min. The exoskeleton of each prawn was then removedand the whole body was cut into small pieces and was ho-mogenized in 5 mL homogenization buffer (0.1 M sodiumphosphate buffer, pH 7.2). Crude homogenate was cen-trifuged at 10,000 ×g, at 4◦C for 20 min. The supernatantwas collected and used immediately for analysis or kept at–80◦C before use.
Characterization of ChEM. lanchesteri ChE activity was determined by colorimet-ric assay (Ellman 1961). Three different substrates, ATChI,BTChI and PTChI were tested for substrate preference.Pooled supernatant from 20 prawns (75 µL) was added to
reaction buffer (875 µL) containing 0.35 mM DTNB in 0.1M sodium phosphate buffer, pH 8.0, and the mixture wasincubated at 25◦C for 10 min. Freshly prepared substrate(50 µL) in 0.1 M sodium phosphate buffer, pH 8.0, wasadded to make up to final concentrations of 0.0625, 0.125,0.25, 0.5, 1.0, 2.0, 3.0, 5.0, 10.0 and 15.0 mM. The ab-sorbance was measured at 412 nm every 30 s for 2 min. Fourdifferent pooled samples were tested for each substrate.
Inhibitor sensitivity was determined using three differ-ent inhibitors, eserine sulfate, BW284c51 and iso-OMPA.Eserine sulfate and iso-OMPA were prepared in ethanolwhereas BW284c51 was prepared in deionized water. Eachinhibitor (5 µL) was added to pooled supernatant (495 µL)to final concentrations of 10−9, 10−8, 10−7, 10−6, 10−5 and10−4 M and the mixture was incubated at 25◦C for 30 min.Ethanol and deionized water were also added to correspond-ing conditions as controls. The mixture (75 µL) was thenadded to the reaction buffer (875 µL) at 25◦C for 10 minprior to addition of 0.5 mM ATChI (50 µL) in 0.1 M sodiumphosphate buffer, pH 8.0, to initiate the reaction. The ab-sorbance was measured at 412 nm as described previously.Four different pooled samples were tested for each inhibitor.
Determination of LC50 valuesM. lanchesteri were exposed to 0.1, 1.0, 2.0, 3.0 and 4.0 µgchlorpyrifos L−1. Each experimental group comprised 15prawns placed in a glass aquarium (20×30×20 cm3) contain-ing about 10 L of each chlorpyrifos concentration. As stockchlorpyrifos was prepared in acetone, controls with andwithout acetone were also included. Mortality was recordedafter 24, 48, 72 and 96 h. Four replicate experiments wereconducted. The LC50 values were determined using ProbitAnalysis (Finney 1971).
Chlorpyrifos exposureBased on the 96-h LC50 value, M. lanchesteri were exposedto 0.5, 1.5, 2.5 and 3.5 µg chlorpyrifos L−1. Each experimen-tal group comprised 60 prawns placed in a glass aquarium(30×40×60 cm3) containing about 60 L of each chlorpyrifosconcentration. Controls with and without acetone were in-cluded. About 10–15 prawns were collected after 24, 48, 72,and 96 h for determination of lipid peroxidation levels, andAChE, CAT and GST activities. Four replicate experimentswere performed.
Enzyme assayPrawn supernatant was prepared and determined for AChEactivity as described previously. CAT activity was deter-mined using the method of Aebi (1984). Briefly, prawnsupernatant (100 µL) was mixed with 50 mM potassiumphosphate buffer, pH 7.0, (900 µL) and then 30 mM H2O2(500 µL) was added. The absorbance at 240 nm was imme-diately measured for 15 sec. GST activity was determinedbased on the method of Habig (1974). Briefly, prawn su-pernatant (50 µL) was added to 0.1 M potassium phos-phate, pH 6.5, (950 µL) containing 0.1% Triton-X 100, 1mM CDNB and 1 mM GSH at 25 ◦C for 1 min. The changein absorbance was monitored at 340 nm every 30 s for 3min.
Determination of lipid peroxidationThe level of lipid peroxidation was determined by measur-ing the production of TBARS using the method describedby Buege & Aust (1978). Prawn supernatant (400 µL)was mixed with solution (800 µL) containing 3.75% tri-choroacetic acid, 0.05 N HCl, 9.25% 2-thiobarbituric acidand 0.03% butylated hydroxytoluene at 25 ◦C for 5 min.
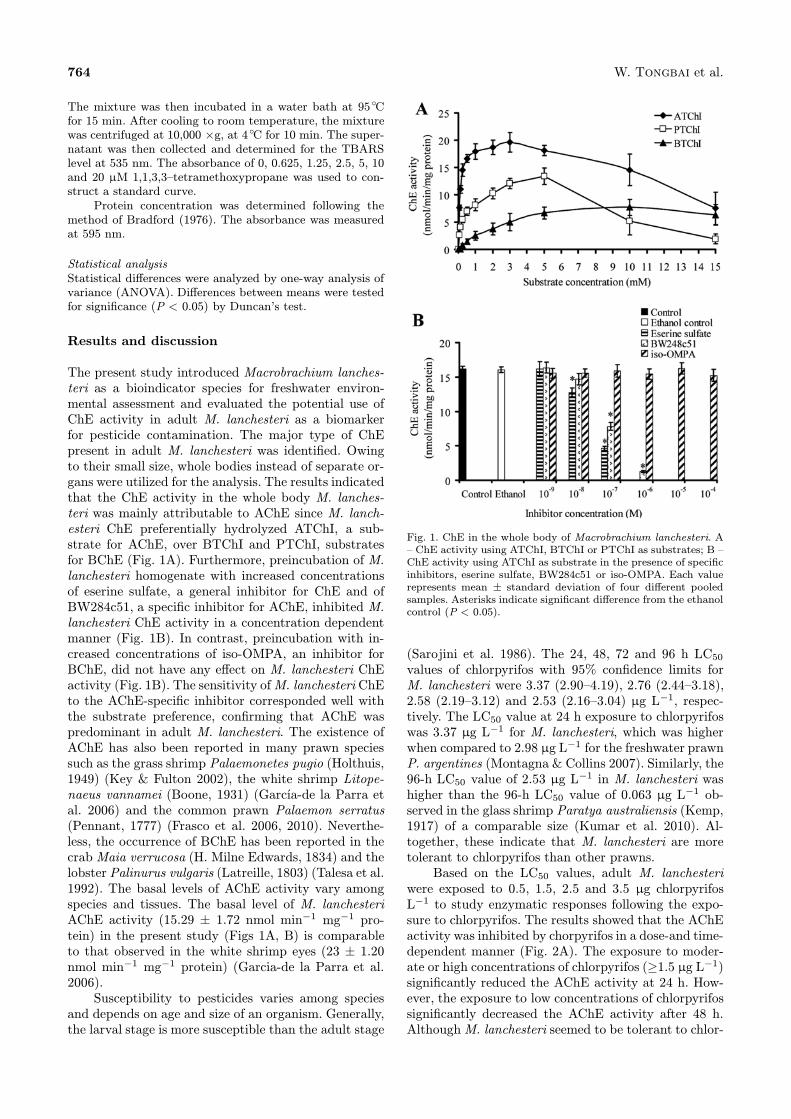
764 W. Tongbai et al.
The mixture was then incubated in a water bath at 95◦Cfor 15 min. After cooling to room temperature, the mixturewas centrifuged at 10,000 ×g, at 4◦C for 10 min. The super-natant was then collected and determined for the TBARSlevel at 535 nm. The absorbance of 0, 0.625, 1.25, 2.5, 5, 10and 20 µM 1,1,3,3–tetramethoxypropane was used to con-struct a standard curve.
Protein concentration was determined following themethod of Bradford (1976). The absorbance was measuredat 595 nm.
Statistical analysisStatistical differences were analyzed by one-way analysis ofvariance (ANOVA). Differences between means were testedfor significance (P < 0.05) by Duncan’s test.
Results and discussion
The present study introduced Macrobrachium lanches-teri as a bioindicator species for freshwater environ-mental assessment and evaluated the potential use ofChE activity in adult M. lanchesteri as a biomarkerfor pesticide contamination. The major type of ChEpresent in adult M. lanchesteri was identified. Owingto their small size, whole bodies instead of separate or-gans were utilized for the analysis. The results indicatedthat the ChE activity in the whole body M. lanches-teri was mainly attributable to AChE since M. lanch-esteri ChE preferentially hydrolyzed ATChI, a sub-strate for AChE, over BTChI and PTChI, substratesfor BChE (Fig. 1A). Furthermore, preincubation of M.lanchesteri homogenate with increased concentrationsof eserine sulfate, a general inhibitor for ChE and ofBW284c51, a specific inhibitor for AChE, inhibited M.lanchesteri ChE activity in a concentration dependentmanner (Fig. 1B). In contrast, preincubation with in-creased concentrations of iso-OMPA, an inhibitor forBChE, did not have any effect on M. lanchesteri ChEactivity (Fig. 1B). The sensitivity ofM. lanchesteri ChEto the AChE-specific inhibitor corresponded well withthe substrate preference, confirming that AChE waspredominant in adult M. lanchesteri. The existence ofAChE has also been reported in many prawn speciessuch as the grass shrimp Palaemonetes pugio (Holthuis,1949) (Key & Fulton 2002), the white shrimp Litope-naeus vannamei (Boone, 1931) (García-de la Parra etal. 2006) and the common prawn Palaemon serratus(Pennant, 1777) (Frasco et al. 2006, 2010). Neverthe-less, the occurrence of BChE has been reported in thecrabMaia verrucosa (H. Milne Edwards, 1834) and thelobster Palinurus vulgaris (Latreille, 1803) (Talesa et al.1992). The basal levels of AChE activity vary amongspecies and tissues. The basal level of M. lanchesteriAChE activity (15.29 ± 1.72 nmol min−1 mg−1 pro-tein) in the present study (Figs 1A, B) is comparableto that observed in the white shrimp eyes (23 ± 1.20nmol min−1 mg−1 protein) (Garcia-de la Parra et al.2006).Susceptibility to pesticides varies among species
and depends on age and size of an organism. Generally,the larval stage is more susceptible than the adult stage
Fig. 1. ChE in the whole body of Macrobrachium lanchesteri. A– ChE activity using ATChI, BTChI or PTChI as substrates; B –ChE activity using ATChI as substrate in the presence of specificinhibitors, eserine sulfate, BW284c51 or iso-OMPA. Each valuerepresents mean ± standard deviation of four different pooledsamples. Asterisks indicate significant difference from the ethanolcontrol (P < 0.05).
(Sarojini et al. 1986). The 24, 48, 72 and 96 h LC50values of chlorpyrifos with 95% confidence limits forM. lanchesteri were 3.37 (2.90–4.19), 2.76 (2.44–3.18),2.58 (2.19–3.12) and 2.53 (2.16–3.04) µg L−1, respec-tively. The LC50 value at 24 h exposure to chlorpyrifoswas 3.37 µg L−1 for M. lanchesteri, which was higherwhen compared to 2.98 µg L−1 for the freshwater prawnP. argentines (Montagna & Collins 2007). Similarly, the96-h LC50 value of 2.53 µg L−1 in M. lanchesteri washigher than the 96-h LC50 value of 0.063 µg L−1 ob-served in the glass shrimp Paratya australiensis (Kemp,1917) of a comparable size (Kumar et al. 2010). Al-together, these indicate that M. lanchesteri are moretolerant to chlorpyrifos than other prawns.Based on the LC50 values, adult M. lanchesteri
were exposed to 0.5, 1.5, 2.5 and 3.5 µg chlorpyrifosL−1 to study enzymatic responses following the expo-sure to chlorpyrifos. The results showed that the AChEactivity was inhibited by chorpyrifos in a dose-and time-dependent manner (Fig. 2A). The exposure to moder-ate or high concentrations of chlorpyrifos (≥1.5 µg L−1)significantly reduced the AChE activity at 24 h. How-ever, the exposure to low concentrations of chlorpyrifossignificantly decreased the AChE activity after 48 h.AlthoughM. lanchesteri seemed to be tolerant to chlor-

Responses of riceland prawn to chlorpyrifos 765
Fig. 2. Effects of chlorpyrifos on Macrobrachium lanchesteri. A – AChE activity; B – TBAR level; C – CAT activity; D – GST activity.Each value represents mean ± standard deviation of four different replicate experiments. Asterisks indicate significant difference(P < 0.05) from the acetone control; a indicates significant difference (P < 0.05) from the corresponding activity in the previous timeinterval.
pyrifos compared to other prawns, the enzymatic activ-ities were still sensitive to such exposure. Sensitivity ofM. lanchesteri AChE to chlorpyrifos exposure rendersit a potential biomarker for freshwater environmentalassessment.Significant levels of lipid peroxidation were induced
as detected by a marked increase (P< 0.05) in the levelsof TBARS, a lipid peroxidation product, by 24 h in adose-dependent manner. However, no further increasein the lipid peroxidation level was observed with in-creased time of exposure (Fig. 2B). Increased lipid per-oxidation levels may be due to ROS production causedby chlorpyrifos exposure.CAT was not as sensitive to chlorpyrifos exposure
as GST. The activity of CAT was not affected at 24 hbut was reduced after 48 h when exposed to moderateor high concentrations of chlorpyrifos (1.5, 2.5 or 3.5 µgL−1) or after 72 h when exposed to a low concentration(0.5 µg L−1) of chlorpyrifos. There was no further de-cline with increased time of exposure (Fig. 2C). How-ever, the GST activity was reduced within 24 h evenwhen exposed to low concentration of chlorpyrifos butthere was no significant change (P > 0.05) in the activ-ity with increased time of exposure (Fig. 2D). DecreasedCAT activity may be associated with ROS productionsince ROS has been shown to directly inhibit CAT ac-tivity (Kono & Fridovich 1982; Escobar et al. 1996).Such a decrease results in a low level of H2O2 degrada-tion thus aggravating oxidative stress inM. lanchesteri.
Decreased GST activity may be indirectly associatedwith high levels of lipid peroxidation since the end prod-ucts of lipid peroxidation, such as malondialdehyde, arereactive aldehydes which may crosslink GST (Leaver &George 1998; Martinez-Lara et al. 2002).Increased activity of antioxidant or detoxification
enzymes is a common response as a defense mechanismof an organism when exposed to a moderate concentra-tion of a toxicant. The reduced activities of CAT andGST in M. lanchesteri following the exposure to chlor-pyrifos in the present study may be due to the fact thatthe concentrations of chlorpyrifos under these experi-mental setting based on the LC50 values were high, re-sulting in the less optimal conditions for these enzymesto function properly in the cells. A lower concentrationof chlorpyrifos may possibly enchance the activity ofthese two enzymes.In conclusion, M. lanchesteri can serve as a sen-
sitive bioindicator species for freshwater environmentalassessment. AChE is the major form of ChE in wholebodies of M. lanchesteri and it is sensitive to chlorpyri-fos exposure. Moreover, exposure to chlorpyrifos in thepresent study induced high levels of lipid peroxidationand caused an inhibition of CAT and GST activities.
Acknowledgements
The authors would like to thank John R. Milne for criti-cal reading of the manuscript. This work was supported by

766 W. Tongbai et al.
the Higher Education Commission, Ministry of Education,Thailand, and a grant from Mahidol University.
References
Aebi H. 1984. Catalase in vitro. Methods Enzymol. 105: 121–126.DOI: 10.1016/S0076-6879(84)05016-3
Anderson M.E. 1997. Glutathione and glutathione delivery com-pounds. Adv. Pharmacol. 38: 65–78. DOI: 10.1016/S1054-3589(08)60979-5
Bradford M. 1976. A rapid and sensitive method for the quantifi-cation of microgram quantities of protein utilizing the prin-ciple of protein-dye binding. Anal. Biochem. 72 (1–2): 248–254. DOI: 10.1016/0003-2697(76)90527-3
Buege J.A. & Aust S.D. 1978. Microsomal lipid peroxida-tion. Methods Enzymol. 52: 302–310. DOI: 10.1016/S0076-6879(78)52032-6
Day K.E. & Scott I.M. 1990. Use of acetylcholinesterase activityto detect sublethal toxicity in stream invertebrates exposedto low concentrations of organophosphate insecticides. Aquat.Toxicol. 18 (2): 101–113. DOI: 10.1016/0166-445X(90)90021-G
Diguiseppi J. & Fridovich I. 1984. The toxicology of molecularoxygen. CRC Crit. Rev. Toxicology 12 (4): 315–342. DOI:10.3109/10408448409044213
Ellman G.L., Courtney K.D., Andres V. & Featherstone R.M.1961. A new and rapid colorimetric determination of acetyl-cholinesterase activity. Biochem. Pharmacol. 7 (2): 88–95.DOI: 10.1016/0006-2952(61)90145-9
Escobar J.A., Rubio M.A. & Lissi E.A. 1996. SOD and cata-lase inactivation by singlet oxygen and peroxyl radicals.Free Radic. Biol. Med. 20 (3): 285–290. DOI: 10.1016/0891-5849(95)02037-3
Finney D.J. 1971. Probit Analysis, (3rd ed). Cambridge Univer-sity Press, Cambridge, 355 pp. ISBN: 052108041X
Frasco M.F., Eržen I., Stojan J. & Guilhermino L. 2010. Lo-calization and properties of cholinesterases in the commonprawn (Palaemon serratus): a kinetic-histochemical study.Biol. Bull. 218 (1): 1–5. PMID: 20203248
Frasco M.F., Fournier D., Carvalho F. & Guilhermino L. 2006.Cholinesterase from the common prawn (Palaemon serratus)eyes: catalytic properties and sensitivity to organophosphateand carbamate compounds. Aquat. Toxicol. 77 (4): 412–421.DOI: 10.1016/j.auqatox.2006.01.011
Fulton M.H. & Key P.B. 2001. Acetylcholinesterase inhibition inestuarine fish and invertebrates as an indicator of organophos-phorus insecticide exposure and effects. Environ. Toxicol.Chem. 20 (1): 37–45. DOI: 10.1002/etc.5620200104
García-de la Parra L.M., Bautista-Covarrubias J.C., Rivera-de laRosa N., Betancourt-Lozano M. & Guilhermino L. 2006. Ef-fects of methamidophos on acetylcholinesterase activity, be-havior, and feeding rate of the white shrimp (Litopenaeusvannamei). Ecotoxicol. Environ. Safe. 65 (3): 372–380. DOI:10.1016/j.ecoenv.2005.09.001
Habig W.H., Pabst M.J. & Jakoby W.B. 1974. Glutathione S-transferases. The first enzymatic step in mercapturic acid for-mation. J. Biol. Chem. 249 (22): 7130–7139. PMID: 4436300
Kavitha P. & Venkateswara Rao J. 2008. Toxic effects of chlor-pyrifos on antioxidant enzymes and target enzyme acetyl-cholinesterase interaction in mosquito fish, Gambusia affi-nis. Envion. Toxicol. Pharmacol. 26 (2): 192–198. DOI:10.1016/j.etap.2008.03.010
Key P.B. & Fulton M.H. 2002. Characterization of cholinesteraseactivity in tissues of the grass shrimp (Palaemonetes pugio).Pest. Biochem. Physiol. 72 (3): 186–192. DOI: 10.1016/S0048-3575(02)00006-8
Kirby M.F., Morris S., Hurst M., Kirby S.J., Neall P., Tylor T. &Fagg A. 2000. The use of cholinesterase activity in flounder(Platichthys flesus) muscle tissue as a biomarker of neurotoxiccontamination in UK estuaries. Mar. Pollut. Bull. 40 (9):780–791. DOI: 10.1016/S0025-326X(00)00069-2
Kono Y. & Fridovich I. 1982. Superoxide radical inhibits catalase.J. Biol. Chem. 257 (10): 5751–5754. PMID: 6279612
Kumar A., Correll R., Grocke S. & Bajet C. 2010. Toxicity of se-lected pesticides to freshwater shrimp, Paratya australiensis(Decapoda: Atyidae): use of time series acute toxicity data topredict chronic lethality. Ecotoxicol. Environ. Safe. 73 (3):360–369. PMID: 19914717
Lackner R. 1998. “Oxidative Stress” in fish by environmentalpollutants, pp. 203–224. In: Braunbeck T., Hinton D.E. &Streit B. (eds), Fish Ecotoxicology, Birkhauser Verlag, Basel,Switzerland, 396 pp. ISBN: 376435819X, 9783764358198,081765819X, 9780817658199
Leaver M.J. & George S.G. 1998. A piscine glutathione S-transferase which efficiently conjugates the end-products oflipid peroxidation. Mar. Environ. Res. 46 (1–5): 71–74. DOI:10.1016/S0141-1136(97)00071-8
Livingstone D.R. 2001. Contaminant-stimulated reactive oxygenspecies production and oxidative damage in aquatic organ-isms. Mar. Pollut. Bull. 42 (8): 656–666. DOI: 10.1016/S0025-326X(01)00060-1
Martinez-Lara E., Leaver M. & George S.G. 2002. Evidence fromheterologous expression of glutathione S-transferases A andA1 of the plaice (Pleuronectes platessa) that their endoge-nous role is in detoxification of lipid peroxidation products.Mar. Environ. Res. 54 (3–5): 263–266 DOI: 10.1016/S0141-1136(02)00188-5
Massoulié J., Pezzementi L., Bon S., Krejci E. & Vallette F.M.1993. Molecular and cellular biology of cholinesterases. Prog.Neurobiol. 41 (1): 31–91. DOI: 10.1016/0301-0082(93)90040-Y
Montagna M.C. & Collins P.A. 2007. Survival and growth ofPalaemonetes argentinus (Decapoda; Caridea) exposed toinsecticides with chlorpyrifos and endosulfan as active el-ement. Arch. Environ. Contam. Toxicol. 53 (3): 371–378.DOI: 10.1007/s00244-006-0209-x
Sarojini R., Nagabhushanam, R. & Avelin Mary S. 1986. Effect offenithrothion on reproduction of the freshwater prawn Mac-robrachium lamerrii. Ecotoxicol. Environ. Safe. 11 (3): 243–250. DOI: 10.1016/0147-6513(86)90098-9
Talesa V., Contenti S., Principato G.B., Pascolini R., Giovan-nini E. & Rosi G. 1992. Cholinesterases from Maia verru-cosa and Palinurus vulgaris: A comparative study. Comp.Biochem. Physiol. 101(3): 499–503. DOI: 10.1016/0742-8413(92)90077-K
Xuereb B., Noury P., Felten V., Garric J. & Geffard O. 2007.Cholinesterase activity in Gammarus pulex (Crustacea Am-phipoda): Characterization and effects of chlorpyrifos. Toxi-cology 236 (3): 178–189. DOI: 10.1016/j.tox.2007.04.010
Received February 9, 2011Accepted March 30, 2012