environmental regulation of fertilization and fruit
TRANSCRIPT

ENVIRONMENTAL REGULATION OF FERTILIZATION
AND FRUIT SETTING IN DATE PALM (Phoenix
dactylifera L.)
Thesis is submitted to
The Robert H. Smith Faculty of Agriculture Food and
Environment for the M.Sc. in Plant Sciences
by
Filip Slavković
January 2015

This thesis was written under the supervision of Dr. Yuval Cohen and Prof.
Rina Kamenetsky of the Agriculture Research Organization, The Volcani
Center.
Filip Slavković Dr. Yuval Cohen
Prof. Rina Kamenetsky

ACKNOWLEDGEMENTS
I would like to express my gratitude to Dr. Yuval Cohen and Prof. Rina
Kamenetsky for their kind and generous support throughout the research,
great help in search for future projects, as well as for making my stay in Israel
feel at home.
I would like to thank David Birger for sharing his experiences and advice for
development of fertilization protocols.
Special thanks to Mazal Ish-Shalom and Miriam Benita for their generous
help regarding the molecular biology protocols and guidance.
I would like to thank Hanita Zemach for her practical guidance on histological
protocols, staining and microscopy.
I wish to thank Dr. Sonia Philosoph-Hadas and Dr. Shimon Meir for helpful
suggestions regarding the setup and post-harvest application of anti-ethylene
compounds.
I would like to thank everyone taking part in this project in Southern Arava
Research Center and mainly Avi Sadowsky and Amnon Greenberg for their
help in the in vivo experiment.
I would like to express my personal gratitude to the Hebrew University of
Jerusalem and the Pears Foundation for supporting my studies.
Finally, I would like to thank my mother Sofija Slavković and my family for
continuous support and encouragement.

Dedication
This thesis is dedicated to the loving memory of my father Siniša Slavković
(1955-2006) who raised me to be the person I am today.

Table of Contents 1. INTRODUCTION ..................................................................................... 1
1.1. Botanical background ...................................................................................... 1
1.2. Reproductive biology of date palm .............................................................. 2
1.3. Horticultural challenges .................................................................................. 3
1.4. Fertilization process ......................................................................................... 4
1.5. The role of hormones in post-pollination processes .............................. 6
1.6. Parthenocarpy ................................................................................................... 7
1.7. Genomic and transcriptomic research in date palm .............................. 9
2. RESEARCH OBJECTIVES ................................................................... 10
3. MATERIALS AND METHODS ............................................................... 11
3.1. Plant material ................................................................................................... 11
3.2. Development of an in vitro assay for spikelet culturing ....................... 11
3.2.1. Selection and optimization of growth media and cultivation
conditions under different temperature regimes ................................. 12
3.2.2. Prevention of fungal contamination ......................................................... 12
3.2.3. Use of ethylene inhibitors to prolong vase-life ..................................... 12
3.3. Effect of temperature regimes on fertilization and fruit setting in
modular phytotrons - in vivo ........................................................................ 13
3.4. Histology and microscopy ............................................................................ 14
3.5. Pollen tube germination and elongation in vitro and on the stigma 14
3.6. Molecular analysis .......................................................................................... 15
3.6.1. RNA extraction .............................................................................................. 16
3.6.2. Gene validation and primer design ......................................................... 16
3.6.3. Gene expression analysis .......................................................................... 17
3.7. Statistical analysis .......................................................................................... 17
4. RESULTS .............................................................................................. 18

4.1. Morpho-anatomical characterization of fertilization and fruit set in
field-grown date cultivars 'Medjoul' and 'Barhee' .................................. 18
4.1.1. 'Barhee' ............................................................................................................ 19
4.1.2. 'Medjoul' ........................................................................................................... 21
4.1.3. Comparison between 'Barhee' and 'Medjoul' ....................................... 23
4.2. Effects of temperature regimes on pollination, fertilization and fruit-
set in date palm ............................................................................................... 24
4.2.1. Development of an in vitro assay for studying date palm
fertilization ...................................................................................................... 24
4.2.2. Effect of temperature regimes on pollination and fertilization ......... 31
4.2.3. Effects of temperature regimes on fertilization and fruit set in
"modular phytotrons" - in vivo ................................................................... 33
4.3. Expression analysis of genes involved in hormonal regulation
during early fruit development of cultivars 'Medjoul' and 'Barhee' ... 41
5. DISCUSSION ........................................................................................ 46
5.1. Morpho-anatomical traits of reproductive system and fruit set in
'Medjoul' and 'Barhee' ................................................................................... 46
5.2. Temperature affects pollen germination, fertilization and fruit-set
processes .......................................................................................................... 48
5.3. Molecular analysis of genes involved in hormonal regulation during
early fruit development of cultivars 'Medjoul' and 'Barhee' ................ 54
References ................................................................................................... 58
Appendix ...................................................................................................... 67

List of Figures
Figure 1: Schematic representation of fertilization in angiosperms ................ 5
Figure 2: Experimental layout of pollination under controlled temperature
conditions in special chambers in the field ................................................... 13
Figure 3: Daily average and extreme temperatures at pollination in
Yotvata during spring periods of 2005-2012 ............................................... 14
Figure 4:Morphological characterization of early fruitlet development in
pollinated and non-pollinated 'Barhee' and 'Medjoul' during the first four
weeks after pollination .................................................................................. 18
Figure 5: Pollinated seed-bearing 'Barhee' fruitlets versus non-pollinated
single and triple parthenocarpic fruitlets ....................................................... 19
Figure 6: Histological characterization of early fruitlet development in cv.
'Barhee' during the first 5 WAP .................................................................... 20
Figure 7: Deterioration of two ovules in pollinated 'Barhee' as opposed to
uniform ovule growth in non-pollinated flower .............................................. 20
Figure 8: Histological characterization of early fruit development in
'Medjoul' during the first 40 DAP in 2012 ..................................................... 21
Figure 9: Effects of Ethylene inhibitors on spikelets "vase life" 10 days
after culturing ............................................................................................... 26
Figure 10: Effect of 1-MCP and STS on viability, flower abscission and
spikelet browning of pollinated and non-pollinated spikelets in vitro ............ 28
Figure 11: Pollinated and non-pollinated inflorescences exposed to four
temperature regimes 10 days after setup ..................................................... 29
Figure 12: Effect of four temperature regimes on pollinated and non-
pollinated inflorescences in vitro .................................................................. 30
Figure 13: Pollen tube length 9 hours after incubation at 4 constant
temperature regimes .................................................................................... 31
Figure 14: Pollen tube length (µm) in vitro in the dark under four constant
temperature treatments measured after 3, 6 and 9 hours respectively ........ 32

Figure 15: Pollen germination on the stigma in vitro under 4 constant
temperatures ................................................................................................ 33
Figure 16: Temperature comparison in high temperature units, medium
temperature units and low temperature units ............................................... 35
Figure 17: Effects of three temperature regimes (warm – 32/18°C,
medium – 25/12°C and cool – 20/8°C) on pollen germination on the
stigma in bunches pollinated on the trees in modular phytotrons in vivo ...... 36
Figure 18: Effects of three temperature treatments (warm – 32/18°C,
medium – 25/12°C and cool – 20/8°C) on pollen germination on the
stigma in vivo ............................................................................................... 36
Figure 19: Fruitlets 5 weeks following pollination in temperature
controlled units (2013) .................................................................................. 37
Figure 20: Effect of three temperature treatments in vivo on fruitlet size 9
WAP (2014).................................................................................................. 38
Figure 21: Percentage of normal, parthenocarpic, aborted and non-
developed fruits respectively, in response to pollination and growth under
different temperatures in vivo (2013) ............................................................ 38
Figure 22: Percentage of normal, parthenocarpic, aborted and non-
developed fruits, in response to pollination and growth under different
temperatures in vivo, 10 WAP in 2014 ......................................................... 39
Figure 23: Effects of three temperature regimes (warm – 32/18°C,
medium – 25/12°C and cool – 20/8°C) on early development of pollinated
flowers / fruitlets in modular phytotrons in vivo (2013) ................................. 40
Figure 24: Heat map of EvaGreen Ct values of pollinated and non-
pollinated flowers of 'Barhee' and 'Medjoul' during the first four weeks of
fruit development .......................................................................................... 42
Figure 25: Hierarchical cluster analysis of genes expressed in pollinated
and non-pollinated flowers of cv. 'Barhee' and 'Medjoul' during four
developmental stages after pollination ......................................................... 43
Figure 26: Differential expressions of selected genes in 'Medjoul' and
'Barhee' during four weeks of early fruit development .................................. 45

List of Tables
Table 1: Percentage of parthenocarpic singlets, parthenocarpic triplets,
normal and shed fruitlets respectively in 'Barhee' at 9 WAP (2014) ............. 19
Table 2: Sizes of carpels and ovules (mm2) in pollinated vs. non-
pollinated flowers of 'Barhee' during first 35 DAP (2012) ............................. 22
Table 3: Carpel and ovule size in pollinated and non-pollinated 'Medjoul'
flowers / fruitlets (mm2) during 0-40 DAP (2012) .......................................... 22
Table 4: Size of degenerating carpels and ovules (mm2) in 'Medjoul', 13
and 21 DAP in pollinated and non-pollinated flowers / fruitlets (2012) ......... 22
Table 5: Percentage of fruitlet-drop (abscission) in 'Barhee' versus
'Medjoul' 3, 4 and 9 WAP respectively grown in the field (March-April
2014) ............................................................................................................ 23
Table 6: Effect of culture media and temperature on isolated spikelets
viability and flower abscission ...................................................................... 25
Table 7: Effects of four temperatures on pollen germination in isolated
spikelet sections in vitro ............................................................................... 33
Table 8: Variation in fruitlet weight 5 WAP (2013) in temperature
controlled units ............................................................................................. 37
Table 9: Effect of different temperature treatments in vivo on fruitlet
weight 9 WAP in the season of 2013 and 10 WAP in 2014 respectively ...... 37
Table 10: Effects of three alternating temperatures in vivo ("Modular
phytotron") on carpel and ovule size (mm2) of pollinated ‘Medjoul’ at five
points after pollination .................................................................................. 40
Table 11: Differential expression of selected genes between non-
pollinated and pollinated flowers of 'Medjoul', four weeks after pollination ... 44

ABSTRACT
Control of pollination and fertilization in date palm is essential for
development of high quality fruits. Overly high rate of fruit set may cause
excessive fruit load, requiring expensive fruit thinning to prevent reduction in
fruit size and marketability. On the other hand, inefficient pollination results in
lower yields. None-fertilized flowers may also develop into parthenocarpic
singlet or triplet fruits, which have no commercial value. Although female
flower comprises three separate carpels, only a single carpel develops into a
fruit, while two others degenerate. In addition, pollination efficiency,
environmental conditions and genetic background (different cultivars)
influence developmental processes of fertilization and fruit development.
The aim of our research is comprehensive characterization of fertilization and
early fruit development in date palm under different conditions. Specifically,
we focused on:
1. Morpho-physiological depiction of fertilization and fruit set processes in
two date cultivars 'Barhee' and 'Medjoul';
2. Study of temperature effects on fertilization and early fruit development;
3. Expression analysis of genes involved in hormonal regulation of fruit
development and carpel degeneration.
Date is a very large tree. To study environmental effects on its reproductive
biology, various techniques were applied. We combined in vitro studies with
experiments in planta in the orchard. Only limited success was achieved in
calibration of an in vitro culturing protocol for pollination of inflorescence
sections, since "vase life" of the detached flowers was very short and
senescence occurred within several days to two weeks.
Special "modular phytotrons", assembled on pollinated inflorescences of
whole date trees in the orchard, were designed for this research, enabling
modification of temperature regimes in planta. Pollen tube growth,
fertilization, fruitlet formation and carpel degeneration, as well as early
development of parthenocarpic fruits were defined and characterized by
macro- and microscopic analyses. We have shown that the two studied
cultivars varied significantly in their reproductive biology, including
development of parthenocarpic fruits, fruitlet shedding and differential

regulation of the physiological processes. Relatively low temperatures,
applied during plant fertilization, significantly decreased pollen germination
rate, enhanced formation of parthenocarpic fruits and reduced normal fruit
development.
Using histological data, we defined the 'developmental checkpoints' and
consequent stages of early fruit formation in two cultivars, and confirmed that
processes of ovule degeneration and carpel shrinkage occur earlier than we
can observe morphologically. Under our experimental conditions, ovule
degeneration could first be detected in 'Barhee' 14 days after pollination,
lagging about a week in pollinated 'Medjoul'. Moreover, carpels of pollinated
flowers were significantly larger in size in comparison to the non-pollinated
flowers already at 27 DAP in both 'Medjoul' and 'Barhee'. Carpel
degeneration in pollinated 'Medjoul' flowers was significantly faster, as
compared to non-pollinated flowers. This demonstrates that in date palm,
pollination has substantial effect on triggering a specific developmental
pattern. Comparison of fruit shedding between cultivars showed highest fruit-
drop in non-pollinated 'Medjoul'.
We performed high throughput gene expression analysis of pollinated and
non-pollinated flowers of the two date cultivars using Microfluidic Dynamic
Array (Fluidigm). Ninety six genes involved in signaling of main plant
hormones (auxin, gibberellin, cytokinin, abscissic acid, and ethylene) were
selected for the expression analysis, using the recently published date palm
transcriptome data. Relative expression of these genes was analyzed at five
developmental stages of early fruit development of two cultivars. Preliminary
results suggest differential expression patterns among the cultivars,
pollination treatments, and different developmental stages.

Abbreviations
ABA: abscisic acid
AUX: auxin
CK: cytokinin
CTAB: cetyl trimethylammonium bromide
DAP: days after pollination
DDW: double distilled water
DH: dehydrogenase
EST: expressed sequence tags
ET: ethylene
FAA: formalin:acetic acid:alcohol
GA: gibberellic acid
GT: glucosyl-transferase
PCD: programmed cell death
Std Dev: standard deviation
WAP: weeks after pollination

1
1. INTRODUCTION
Date palm (Phoenix dactylifera L.) is one of the oldest and economically most
important trees in the Middle East and North Africa (Chao and Krueger, 2007),
estimated to have nearly 2,000 varieties around the world. Its economic utility is
multifold and includes staple food, beverages, ornamentals and architectural
materials with a yearly production of 7,5 million tons of fruit (Chao and Krueger,
2007; FAOSTAT, 2012).
In Israel, date palms are commercially grown all along the Jordan rift from the
Sea of Galilee in the north to the Arava valley on the south. More than 30,000
metric tons of date fruit are annually produced in Israel, out of which about 30%
was exported in 2012, mainly to Europe (Cohen and Glasner, 2014). Among
cultivars, ’Medjoul‘ has the highest commercial value, surpassing all other
varieties with regard to fruit quality and size. ’Medjoul‘ fruit quality, however, is
very sensitive to the environmental conditions (Zaid and De Wet, 2002).
1.1. Botanical background
Phoenix dactylifera is a diploid (2n = 36), perennial, and monocotyledonous
plant, of the Arecaceae (Palmaceae) family, or the palm family. The genus
Phoenix is distinguished from other genera of pinnate-leaved palms by the
upward and lengthwise folding of the pinnae, as well as by peculiarly furrowed
seeds (Nixon and Carpenter, 1978; Dransfield, 2008). The genus contains 14
species, all native to the tropical or subtropical regions of Africa and Southern
Asia; ranging from the Atlantic islands through Africa, Crete, the Middle East
and India to Hong Kong, Taiwan, Philippines, Sumatra and Malaya (Barrow,
1998; Dransfield, 2008). Most of the 14 Phoenix species are well known as
ornamentals, the highly valued is P. canariensis Chabeaud, commonly called
the Canary Island Palm (Nixon and Carpenter, 1978; Zaid and De Wet, 2002).
The name of the date palm originates from its fruit: "phoenix" from Greek means
purple or red (fruit), and "dactylifera" refers to the finger-like appearance of the
fruit (Chao and Krueger, 2007).
Since ancient time this plant has been recognized as the "tree of life" because of
its integration in human settlement, wellbeing, and food security in hot and
barren parts of the world, where only a few crops can be produced (Jain et al.,
2011). The origin of the date palm is thought to be the ancient Mesopotamia

2
area (southern Iraq) or western India (Wrigley, 1995). From the center of origin,
date cultivation spread throughout the Arabian Peninsula, North Africa and the
Middle East. The expansion of date cultivation later accompanied the expansion
of Islam and reached southern Spain and Pakistan, and was subsequently, with
the Spanish missionaries, introduced to North America and Australia (Nixon,
1951; Zaid and De Wet 2002; Chao and Krueger, 2007).
Being a monocotyledon, date palm has highly developed fibrous roots and
lignified trunk that lacks cambium, hence it cannot be grafted. It is the tallest of
the Phoenix species, however, in spite of its enormous size, date palm tree is
grass-like: rather flexible to strong desert winds. The growth form of a palm tree
is characteristic; the plant usually consists of an unbranched stem with a crown
of large leaves at the apex, reaching the height of over 20 m, and having the
crown radius of about 7-8 m. The leaves are 4-5 m long, pinnate, growing
upward in a spiral pattern. Each leaf has an auxiliary bud that may be
vegetative, floral or intermediate. A fully productive date palm tree can support
up to 30 clusters, which can carry more than 300 kg of fruits (Jain et al., 2011).
1.2. Reproductive biology of date palm
Date palm is dioecious, meaning that female and male reproductive structures
are separated to different individuals; each generating unisexual flowers.
Flowers are developing in a big cluster (inflorescence) called spadix or spike,
which consists of a central stem called rachis and several dozens of strands or
spikelets (Zaid and De Wet, 2002), each carries numerous flowers. The
developing inflorescences are enclosed in a hard, fibrous cover (the spathe) that
protects the flowers (Chao and Krueger, 2007). As many as 8,000 to 10,000
flowers may be present in a single female inflorescence and even more can be
found in a male inflorescence (Zaid and De Wet, 2002).
In general, flowers of female and male trees differ in morphology (Nixon and
Carpenter, 1978; Vandercook et al., 1980). The staminate (male) and pistillate
(female) flowers are connected to spikelet with flattened peduncle, short to
elongate, in the pistillate often elongating after fertilization (Dransfield 2008).
The female flower is globose and contains three sepals and three petals that are
fused together, so that only their tips diverge, and three carpels that are
separated; each carpel contains a single anatropous ovule with a single large

3
stigma. The male flower comprises three connate sepals in a low cupule and
three rounded petals; it is sweet-scented and normally possesses six stamens.
Pollen is ellipsoidal, bisymmetric or slightly asymmetric with the longest axis 17-
30 µm (Dransfield, 2008).
Upon pollination, only one ovule of female flower develops into a fruit, whereas
the other two carpels degenerate (Zaid and De Wet, 2002). However, when
pollination is not efficient, parthenocarpic fruits can form, in which one or all
three non-fertilized carpels develop (Reuveni, 1986). Parthenocarpic fruits may
develop from all three carpels, or, similar to the normal fruit, only single carpel
will continue its development to the parthenocarpic fruit, while the other two will
abort. Various cultivars differ in their ability to produce normal and
parthenocarpic fruits. For example, in 'Barhee', non-pollinated flowers tend to
produce triple parthenocarpic fruitlets, while single parthenocarpic fruits are
characteristic for "Medjoul'. Moreover, most parthenocarpic fruits of non-
pollinated 'Medjoul' tend to shed during development, while in 'Barhee' they
usually remain attached to the spikelets.
1.3. Horticultural challenges
From the horticultural standpoint, efficient pollination and fertilization are crucial
for successful fruit development and marketable dates. When pollination is
inefficient, female flowers are not fertilized, which leads to the development of
parthenocarpic fruits with no commercial value (Zaid and De Wet, 2002). On the
other hand, too efficient fertilization may cause excessive fruit load, which
reduces fruit size and marketability and requires expensive fruit thinning.
Therefore, optimization of the pollination process is extremely important for the
production of quality fruits.
Pollination and fertilization processes are limited by various environmental
factors. In general, 12-27˚C are optimal for the growth of date palms, while the
trees can withstand high temperatures up to 50˚C and short periods of frost at -
5˚C.They flower when the shade temperature increases to more than 18˚C, and
for fruit setting more than 25˚C are required (Zaid and de Wet, 2002a). The ideal
temperature for the growth of the date palm, during the period from pollination to
fruit ripening, ranges from 21 to 27˚C (Zaid and de Wet, 2002a). However,
temperatures in some arid regions vary drastically on a daily basis with

4
amplitudes reaching more than 20˚C; as a result, the success of pollination,
fertilization and consequent fruit set is often bellow optimum.
1.4. Fertilization process
In order to fertilize female flowers, pollen from male trees must reach the stigma.
Naturally, wind-mediated pollination, anemophily, is common in date palms. In
commercial production, male inflorescences are collected from male trees for
artificial pollination (Chao and Krueger, 2007). Pollen harvested from a single
male tree is sufficient for pollination of 50 female trees. Therefore, an Israeli
date plantation has approximately 2 % male trees (Cohen and Glasner, 2014).
During pollination, pollen grains of angiosperms reach the female pistil and
adhere to the stigma. Upon germination, the pollen grain elongates into a pollen
tube, penetrates the stigma and heads toward the ovary by creating a path
through the female carpelate tissue. Eventually, the tube, containing two sperm
cells, enters the ovule and reaches the female gametophyte where fertilization
takes place (Figure 1). Female gametophyte is an eight-nuclei structure,
comprising the egg cell, polar nuclei, two sinergids and three antipodial cells.
After the pollen tube enters the female gametophyte, the pollen tube nucleus
disintegrates and the two sperm cells are released; one of the two sperm cells
fertilizes the egg cell, forming a diploid (2n) zygote, which then divides
repeatedly by mitosis to give rise to the seed embryo; the other sperm cell will
fuse with the two haploid polar nuclei forming triploid (3n) endosperm, which will
form the main nutrient source for the growing embryo. In angiosperms, i.e. the
flowering plants, this process is termed a double fertilization.
Fertilization leads to fruit set - the commitment of the ovary to proceed with fruit
development, also defined as the changeover from the static condition of the
flower ovary to the rapidly growing condition of the young fruit (Serrani et al.,
2007). This process is controlled by positive growth signals generated during
fertilization, e.g. auxins, gibberellins (GAs), cytokinins and ethylene (Crane
1964; Nitsch 1970; O'Neill 1997; Srivastava, 2005).

5
Figure 1 - Schematic representation of fertilization in angiosperms. From:
Brower, B. Pollination and Fertilization. Retrieved September, 2014, from
Connexions,
http://cnx.org/content/m44723/latest/?collection=col11516/latest
Note that in date palms each one of the three carpels is separated, having
its own stigma, and only one ovule is fertilized.
Fertilization leads to fruit set - the commitment of the ovary to proceed with fruit
development, also defined as the changeover from the static condition of the
flower ovary to the rapidly growing condition of the young fruit (Serrani et al.,
2007). This process is controlled by positive growth signals generated during
fertilization, e.g. auxins, gibberellins (GAs), cytokinins and ethylene (Crane
1964; Nitsch 1970; O'Neill 1997; Srivastava, 2005).
In most flowering plants, early fruit development can be divided into three
phases. The earliest phase involves the development of the ovary and the
decision to abort or to proceed with further cell division and fruit development,
i.e. the fruit set. In the second phase, fruit growth is due primarily to cell division.
The third phase begins after cell division ceases, and fruit growth continues
mostly by cell expansion, until the fruit reaches its final size (Gillaspy et al.,
1993). In monocotyledonous plants, the region of most rapid growth of the fruit
is the very base, that is, the region enclosed by the calyx (Haas and Bliss,
1935).

6
1.5. The role of hormones in post-pollination processes
Hormonal regulation plays a prominent role in fruit development of plants (Nitsch
1970, Ozga et al., 2003), including all classes of plant hormones: auxins, GAs,
cytokinins, inhibitors (for example, ABA), and ethylene.
Gibberellins (GAs) are tetracyclic diterpenoids that control a wide range of
developmental processes; they are key factors for fruit-set and development. GA
treatment of unpollinated pistils promotes fruit initiation, probably by mimicking
GA production upon ovule fertilization (Vivian-Smith and Koltunow, 1999;
Dorcey et al., 2009). In fact, upon pollination, GA biosynthesis genes are up-
regulated, and bioactive GA1 and its precursor GA20 levels increase (Ben-
Cheikh et al., 1997).
Gibberellins and auxins are considered to be the main stimulus in the induction
of fruit set, since their endogenous levels increase suddenly in ovaries after
fertilization (Gillaspy et al., 1993; Ben-Cheikh et al., 1997; Goetz et al., 2002).
Moreover, auxins and GAs are widely known for their ability to promote
fertilization-independent fruit development in several species (Barendse and
Peeters, 1995; Nitsch, 1970; Ozga and Reinecke, 2003). For example,
stimulation of fruit set by GA has been observed in pear (Pyrus communis). GA-
induced fruit set may occur in the absence of pollination, resulting in
parthenocarpic fruits.
On the other hand, following successful pollination, the presence of fertilized
ovules generally triggers the development of the ovary into a fruit. The
commitment to proceed with fruit development (fruit set) is therefore dependent
on one or more positive growth signals produced by pollen during germination
and pollen tube growth and during or after fusion of the nuclei (Gillaspy, 1993).
Sastry and Muir (1963) showed that it is not auxin, but GA that is transferred
from the germinating pollen to the ovary. Subsequently, the GA may induce an
increase of the auxin content in the ovary to levels adequate to trigger fruit
growth (Sastry and Muir, 1963; Koshioka et al., 1994). Furthermore, Vriezen et
al. (2008) showed that the mRNA levels of several ethylene biosynthesis genes
and genes involved in ethylene signaling decreased after pollination in tomato,
as well as transcript levels of ABA biosynthesis genes. Accordingly, these
findings led to a conclusion that the onset of fruit development depends on the

7
induction of GAs and auxin responses, while ethylene and ABA responses are
attenuated.
1.6. Parthenocarpy
Parthenocarpy is generally considered as the formation of a fruit without
fertilization of the ovules, and it was introduced by Noll (1902) to designate fruit
formation without pollination or other stimulation (Nitsch, 1952). For example,
the oriental persimmon (Diospyros kaki), as well as some varieties of figs, pears,
and grapes, are often parthenocarpic. Moreover, the cultivated banana is always
parthenocarpic (D'Angremond, 1912), while the wild banana is not.
Parthenocarpic fruit development can be genetically controlled or artificially
induced by exogenous application of hormones, mostly auxin and GAs. GAs
have been reported to promote parthenocarpic fruit development in different
species such as tomatoes (Serrani et al., 2007), apples (Hayashi et al., 1968),
pears (Gil et al., 1972), as well as in various cultivars of date palm (Shaheen et
al., 1988). In addition, seedless dates were obtained in unpollinated bunches
treated with GA3. Abd-Alaal et al. (1982) found that the use of 2,4D, 2,4,5-T,
2,4,5-TP, IAA and GA3 at the concentrations of 25-100 ppm resulted in
formation of seedless dates in the 'Khadrawi' date cultivar (Shaheen et al.
1988).
Several authors reported a correlation between increased auxin and gibberellin
levels in the ovary before fertilization and parthenocarpic fruit development
(Gillaspy et al., 1993). The endogenous levels of auxins and GAs are higher in
ovaries of parthenocarpic tomato lines than in seed-producing lines (Gustafson,
1939b; Nitsch et al., 1960; Mapelli et al., 1979; Mapelli and Lombardi, 1982).
However, Fuentes et al. (2012) reported that in Arabidopsis, auxin-induced
parthenocarpy occurs entirely through GA signaling, and is dependent and
independent of functional GA signaling machinery, associated with the DELLA
proteins.
With respect to genetic manipulation, Pandolfini (2009) suggests that
parthenocarpy can be achieved in several ways: by genetic modification of auxin
synthesis, auxin sensitivity and auxin content, or by manipulating genes of the
auxin (IAA9 or ARF8), or gibberellin signal transduction (DELLA). For example,
two components of the auxin signal transduction pathway, AUXIN RESPONSE

8
FACTOR8 (ARF8) from Arabidopsis (Vivian-Smith et al., 2001; Goetz et al.,
2006) and the Aux/IAA protein IAA9 from tomato (Wang et al., 2005), have been
shown to repress ovary growth before fertilization. Aux/IAA proteins can bind to
ARF proteins to activate or inhibit the transcription of auxin responsive genes
(Ulmasov et al., 1999b; Hardtke et al., 2004; Tatematsu et al., 2004). It has
been proposed that both Arabidopsis and tomato possess ARF8- and IAA9-like
orthologs that interact and, together with potentially other as yet unknown
proteins, form a protein complex that prevents fruit set prior to fertilization
(Goetz et al., 2006; Swain and Koltunow, 2006). Namely, ARF8 is an ovule-
specific transcription factor that negatively regulates fruit set (Goetz et al.,
2006); after pollination/fertilization ARF8 gene expression is switched off.
Moreover, fruit development can be uncoupled from fertilization also by silencing
DELLA proteins, which are repressors of GA signaling (Marti et al., 2007).
DELLA proteins are a subfamily of the GRAS protein family of putative
transcription factors characterized by the conserved amino acid motif DELLA
(Thomas and Sun, 2004). These proteins are negative regulators of GA
signaling that act immediately downstream of the GA receptor. Binding of GA to
its soluble receptor, GID1, causes binding of GID1-GA to DELLAs and leads to
their degradation via the ubiquitin-proteasome pathway. DELLAs are nuclear
localized and are hypothesized to function as transcriptional regulators (Eckardt,
2007).
Cytokinins (CK) play a central role in the regulation of cell division (Frank and
Schmulling, 1999). In tomato fruit, CK levels peak during the phase of high
mitotic activity (Bohner and Bangerth, 1988). Moreover, high levels of CK were
reported to induce programmed cell death (PCD) in proliferating cells of carrot
and Arabidopsis (Carimi et al., 2002).
In many plant species, plant hormones were also reported to be associated with
female gametophyte development, as well as pollen germination and pollen tube
elongation. For example, GAs promote Arabidopsis petal, stamen and anther
development by opposing the function of the DELLA proteins RGA, RGL1 and
RGL2 (Richards et al., 2001). Before fertilization, DELLA proteins repress
growth and elongation of the ovary. On fertilization, auxin (IAA) is produced in
the ovules, inducing GA3 production in the valves. GA3 then mediates DELLA
degradation and fruit growth (Sundberg and Østergaard, 2009).In orchid

9
Phalaenopsis, ovary wall epidermal cells begin to elongate and form hair cells
two days after pollination; this is the earliest visible morphological change in
female gametophyte after pollination and prior to pollen germination, indicating
that signals associated with pollination itself trigger these changes (Zhang and
Oneill, 1993). The effects of inhibitors of ethylene biosynthesis
(aminoethoxyvinylglycine - AVG) on early morphological changes indicated that
ethylene, in the presence of auxin (NAA), is required to initiate ovary
development and, indirectly, subsequent ovule differentiation. Furthermore,
pollen germination and growth were strongly inhibited by AVG, indicating that
male gametophyte development is also regulated by ethylene.
1.7. Genomic and transcriptomic research in date palm
In the last years, much effort has been made in creating molecular information
on date palms. Two drafts of the date palm nuclear genome (cv. 'Khalas') were
published in 2009 (Al-Dous et al.; GCA_000181215.2), and 2011 (Al-Dous et al.;
GCA_000413155.1), estimating the genome size of 550-650 Mb. First, full-
genome assemblies of the two date palm organelles, plastid and mitochondrion
have been published (Yang et al.; 2012, NC_013991.2; Al-Mssallem et al.,
2013, NC_016740.1). Further, a comparative transcriptome study on mesocarps
of oil palm and date palm was performed (Bourgis et al., 2011), as well as
identification and characterization of differentially expressed ESTs in date palm
leaves affected by brittle leaf disease (Saidi et al., 2010). The genomic approach
was used to acquire massive transcriptome data for the date palm fruit at seven
different developmental stages (Al-Mssallem et al., 2013), subsequently merged
into three stages (Yang et al., 2012). Annotated isotigs of the defined fruiting
stages provide the ground information to study biological processes of interest in
fruit development.
In spite of its undeniable significance, only a few studies were performed on
date palm fertilization, fruit setting and development. Unlike other crops whose
stress-related reproductive biology is well-known, the effects of environmental
conditions i.e. temperatures, on pollen tube growth, fertilization and fruit set in
date require further study. Due to vulnerability of the reproductive stage and the
fact that temperatures in some arid regions vary drastically on a daily basis, the
consequent success of pollination, fertilization and fruit set is often bellow

11
optimum. One of the important questions is whether temperatures limit these
processes. In the present study, by using transcriptome data, we also aim to
focus on genes involved in hormonal regulation of pollination, fertilization and
early fruit development in two cultivars. Our working hypothesis suggests that in
date palm, fertilization, early fruit development and parthenocarpy are
significantly affected by environmental conditions and hormonal balance.
2. RESEARCH OBJECTIVES
The main aims of the research are as follows:
(1) Morphological and anatomical characterization of pollination, fertilization
and fruit setting processes in date palm cultivars 'Medjoul' and 'Barhee';
(2) Characterization of parthenocarpic fruit development in two cultivars
'Medjoul' and 'Barhee' and comparison to normal fruit development;
(3) Assessment of the temperature effects on fertilization and fruit setting;
(4) Expression analysis of genes involved in hormonal regulation during fruit
development in pollinated and non-pollinated flowers.

11
3. MATERIALS AND METHODS
3.1. Plant material
Trees of date palms (Phoenix dactylifera L.) cv. 'Medjoul' and 'Barhee' from
Southern Arava Research Center, Kibbutz Samar and Mitzpe Shalem were
used in this research. The inflorescences of Canary palm (Phoenix canariensis)
trees, grown in the campus of Agricultural Research Organization, the Volcani
Center in Bet Dagan were employed for calibration of the in vitro assay and
treatments with anti-ethylene compounds.
3.2. Development of an in vitro assay for spikelet culturing
For in vitro studies, inflorescences of date palm 'Medjoul' were brought from the
experimental orchard of Southern Arava Research Center (February – April of
2013) and from Mitzpe Shalem orchard (March 2014).
Inflorescences, enclosed in their spathes, were cut, wrapped in wet paper,
placed in paper bags and were immediately delivered to the laboratory in cooled
containers. In the lab, the spathes were gently open, and single spikelets
carrying flowers were cut to the length of approximately 15cm (2012).
Alternatively, the central parts of the bunch, comprising approximately 15-20
spikelets were used (2013). To prevent embolism, spikelet base was cut under
water. Flowers were carefully pollinated with normal pollen with a small
paintbrush, and inflorescence sections were incubated in growth chambers at
constant temperatures: 15°C, 20°C, 25°C and 30°C and a 12-h photoperiod at
the Volcani Center (2012) or at different temperature regimes (34/28˚C,
28/22˚C, 22/16˚C and 16/10˚C, day/night) at the Phytotron of the Faculty of
Agriculture of the Hebrew University of Jerusalem (2013). For sampling, four
replicates were used per each time point (1, 2, 3, 4, 6, and 7 days after
pollination), so that a total of 72 samples were collected from in vitro fertilization
under constant temperature regimes (4 replicates x 6 time points x 3
temperature treatments). Spikelets with flowers that were not pollinated were
used as negative control. Pollinated flowers (or control) were fixed in FAA for
macro- and microscopic studies.
In the in vitro assay, each sample (spikelet or inflorescence section) was scored
twice a week for: (a) solution or media contamination (fungi), (b) spikelet

12
browning, (c) spikelet drying, (d) flower/fruitlet browning, (f) flower/fruitlet drop,
(g) stigma browning, and (h) general senescence.
Calibration of the assay for spikelet culturing in vitro comprised the evaluation of
various media, preserve compounds, fungicides and ethylene inhibitors:
3.2.1. Selection and optimization of growth media and cultivation conditions
under different temperature regimes
The spikelets were grown either on solid (agar) or liquid media. Each treatment
was performed in 8 repetitions.
For agar plates, components used were: 3% sucrose, Murashige Skoog (MS)
medium (Getter M0222), casein hydrolydase (Getter YB- C1301), plant agar
(0.8%, Getter YM-P1001) and active charcoal (0.25%, Getter YB-C1302). pH
was set to 5.7. In half of the plates with agar, 5 ml of water were added over a
cotton plug to keep humidity of the chamber.
To test the liquid media, the spikelets were placed in the tubes with 10 ml water
(control) or water solutions of “TOG6”, TOG6 + 2% sucrose, “Longlife” (GADOT)
(liquid). All media were replaced on a weekly schedule. Prior to replacing, the
base of each spikelet was cut to remove damaged tissue with clogged water
vessels.
3.2.2. Prevention of fungal contamination
In order to prevent fungal contamination, 0.2% Marpan fungicide was added
(dipped) to the liquid media with spikelets which was then replaced every 7
days.
3.2.3. Use of ethylene inhibitors to prolong vase-life
To prevent senescence and rapid deterioration of plant material during in vitro
culturing, spikelets (from Canary palm inflrorecences) cultivated in liquid solution
were pulse-treated with 0.2% Silver thiosulfate (STS) for 4, 8 and 16 hours
respectively, at 20°C. Alternatively, spikelets placed in sealed glass chambers at
20°C, and incubated with a total concentration of 500 ppb 1-methylcyclopropene
(1-MCP). The gaseous 1-MCP was prepared in a closed Florence flask with 1%
KOH, and was then injected into the chambers. The chambers remained sealed
for 4 hours. Incubation with 1-MCP was performed just before pollination, or 1 or
2 days after pollination (DAP).

13
Figure 2 Left: Experimental layout of pollination under controlled temperature
conditions in special chambers in the field (a); right: modular phytotrons assembled
on pollinated bunches on a tree (2013) (b). Three alternating temperature regimes
were applied in four replicates, using one bunch per replicate.
20/8°C
32/18°C
25/12°C
20/8°C
25/12°
32/18°
20/8°C
32/18°
25/12°
32/18° 20/8°
25/12°
C
a) b)
3.3. Effect of temperature regimes on fertilization and fruit setting in
modular phytotrons - in vivo
For in vivo studies, we used ten years old intact date palm trees 'Medjoul',
grown in an orchard at Southern Arava R&D, Yotvata, Israel. Twelve bunches
(three bunches per tree in four adjacent trees) were pollinated with a pollen
mixture (50% viable pollen + 50% inertial material made of potato flour and
charcoal). Bunches were enclosed in special temperature controlled units
designed by Crystal Vision (Kibbutz Samar, Israel) with the aim to create a
specific temperature regime. The units were regulated by a computerized
system to induce three different temperature regimes: cold 20/8°C, medium
25/12°C and warm temperature 32/18°C (day/night, respectively) (Figure 2).
Figure 3 represents daily average and extreme temperature during flowering of
date palm in Yotvata. In the temperature units, temperatures were lowest at
05:00, increased gradually to highest level at 15:00, and then gradually
decreased. For control, four additional bunches were pollinated, but were
exposed to outdoor temperatures instead of being enclosed in chambers. Units
were installed on the trees around individual inflorescences and sealed during
the period of 14.03-22.04, 2013 and 18.03-01.04, 2014. Then, units were
removed and further fruitlet development was followed under local weather
conditions.

14
In a negative control, bunches were not pollinated; instead, they were closed in
paper bags to prevent pollination. The developing flowers from each bunch were
sampled ten times, within first six weeks from pollination.
3.4. Histology and microscopy
Flowers and young fruitlets were collected from the spikelets and immediately
fixed in FAA (10% formaldehyde: 5% acetic acid: 50% ethanol, v/v).
Morphological and histological studies were performed using stereoscope
(DMLB, Leica) and light microscope (MZFLIII, Leica). For histological studies,
FAA fixed samples were gradually dehydrated in alcohol (50%, 70%, 90%, 95%
and 100% ethanol), cleared with xylene (Histo-clear) and paraffin embedded by
placing in liquid paraffin using Paraplast Plus – Tissue Embedding Medium
(8889502004). Samples were sectioned in 15 μm using a Leica RM2245
microtome and stained with Safranin / Fast Green (Ruzin, 1999).
3.5. Pollen tube germination and elongation in vitro and on the
stigma
Pollen was germinated in vitro at different constant temperatures (15°C, 20°C,
25°C and 30°C) for 3 hours in a solution of 10% sucrose and 500mg\L Boric
Acid (Bernestein, 2004). Pollen grains were visualized under a microscope
(MZFLIII, LEICA), photographed (Nikon DS-Fi1) and their tube length was
measured using the NIS-Elements BR 3.1 Program.
Figure 3 - Daily average and extreme temperatures at pollination in Yotvata
during spring periods of 2005-2012.

15
Analysis of pollen tube elongation in stigmas was adapted from Reuveni et al.
(1986) and Cohen et al. (2004). Prior to histological evaluation, FAA fixed
flowers were washed three times in double distilled water (DDW) and ethanol
(100%) 1:1; each washing lasted 10 minutes. Samples were gently stirred
during this time. Washing was repeated five more times by using DDW. Using
stereoscopic microscope, flowers were dissected, and the three carpels were
separated. Stigmas with the surrounding tissue were cut off from each carpel in
order to be assessed individually. At this point, in order to prevent from
shriveling, carpels were drenched with water droplets and were kept constantly
wet. Stigmas were then cleared in scintillation vials containing 1 ml of 10M
NaOH for two hours with the aim to slightly bleach the sample and make the
tissue softer and easier to manipulate. After being cleared with NaOH, washing
with DDW was repeated five more times.
Stigmas were placed on the glass slides, stained with aniline blue (0.4% in
0.35% K3PO4 solution), covered with a covering glass, and examined
immediately under fluorescence microscope (MZFLIII, LEICA) with a UV
excitation filter set (340-380/400/425 nm). Pollen germination on the stigma was
estimated on a five-point scale: 0 – no germination; 1 – low or sporadic
germination; 2 – moderate germination; 3 – pollen germination covers most of
the stigma; and 4 – high germination.
3.6. Molecular analysis
For molecular analysis, we used date palm cultivars 'Medjoul' and 'Barhee',
grown in the orchard at Kibutz Samar, Israel.
Flowers were collected at 5 different time points: before pollination (0), 1, 2, 3
and 4 weeks following pollination. Additional inflorescences were opened and
immediately covered with paper bags to prevent pollination. Flowers were
collected from these inflorescences at the same time points as non-pollinated
controls. Four replicates were used per treatment, per time point. Upon
sampling, flowers were manually removed at the orchard from the spikelet,
immediately frozen in liquid nitrogen, transferred in dry ice to the laboratory, and
kept in -80°C until use.

16
3.6.1. RNA extraction
RNA was extracted from 2 g flower tissue that was ground in liquid nitrogen
according to the CTAB-based method (Chang, 1993). 20-ml preheated
extraction buffer (65˚C) was quickly added to suspend the RNA and the mix was
extracted with an equal volume of chloroform: isoamyl alcohol (24:1) and
precipitated overnight at 4˚C LiCl at final concentration of 2.5 M. Nucleic acids
were pelleted using centrifugation (13,000 rpm for 20 min), washed with 70%
ethanol and dissolved in 500 µl of SSTE buffer (10 mM Tris–HCl, pH 8.0, 1 mM
EDTA, 1M NaCl, 0.5 % (w/v) SDS). Following another extraction with equal
volume of phenol: chloroform: isoamyl alcohol (25:24:1), RNA was finally
precipitated with two volumes of ice cold absolute ethanol overnight at -20˚C.
After centrifugation (13,000 rpm for 20 min) and another washing with 70%
ethanol, pellet was dried and dissolved in 30 μl of ultra-pure water ("Biological
industries"). DNA traces were digested using 1 unit of RQ1 RNase free DNase
("Promega"), in the presence of 40 units Ribolock RNase inhibitor (Thermo
Fisher Scientific) for 60 min at 37˚C. Then, RNA was re-extracted using phenol:
chloroform: isoamyl alcohol, (25:24:1), extraction and precipitation with
isopropanol and glycogen (Thermo Fisher Scientific). Eventually, it was
centrifuged, washed with ethanol and dissolved in ultra-pure water. The quality
and purity of RNA were examined by a Thermo Scientific NanoDrop™ 1000
Spectrophotometer and by running samples on 1.5% Agarose gel (sampls were
denaturing at 70°C for 10 minutes with 2X RNA loading dye (Thermo Fisher
Scientific). For cDNA synthesis, Thermo Scientific Verso cDNA Synthesis Kit
was used with random hexamer primers, according to the manufacturer
instructions. cDNA synthesis was performed at 42°C (1 hour) followed by
enzyme inactivation at 95°C for 2 minutes.
3.6.2. Gene validation and primer design
The transcriptome of the 'Khalas' cultivar (Al-Mssallem 2013, WGS-
ACYX02000001-ACYX02142304) was used as a reference to identify genes
involved in hormonal biosynthesis and regulation. Only genes active at early
early stages of fruit development (1-30 DAP) were considered. Kyoto
Encyclopedia of Genes and Genomes (KEGG) as well as National Center for
Biotechnology Information (NCBI) databases were used for data mining and

17
gene validation in order to select a total of ninety six transcript sequences
Genes of the main hormone families: GAs (including DELLA proteins), auxins,
cytokinins, ABA, and ethylene were selected. In addition, genes related to
programmed cell death, and senescence-associated proteins were also
selected. With respect to genes coding for same proteins / enzymes, we
included sequences with both high and low relative expression, according to the
transcriptome. Multiple alignments were performed using DNAman software and
specific primers were designed for the selected genes. As house-keeping
genes, actin, elongation factor, F-box and glucose-6-phosphate dehydrogenase
(G6PD) were selected (Appendix 1). Primers were designed using Primer3
Software (http://primer3.ut.ee/), IDT Primer Quest tool
(http://eu.idtdna.com/Primerquest/Home/Index), and BioEdit Software (Ver. 7.2).
3.6.3. Gene expression analysis
cDNA samples from 72 plant samples of 'Barhee' and 'Medjoul' (four biological
repeats per treatment) were placed in two plates, as two technical repeats.
Calibration curves were made of cDNA mixtures of 'Barhee' and 'Medjoul'
samples collected at 0, 2 and 4 WAP and diluted at ratio 1:1, 1:4, 1:16, 1:64,
1:256 and 1:1024. Gene expression analysis was performed using Fluidigm
Real-time PCR analysis software version 4.1.2. Upon selecting actin as a
reference gene, and a ('time zero') sample before pollination as a reference
sample, the heat map of EvaGreen Ct values was obtained. Moreover, data was
measured as the ΔΔCT value - the fold change in gene expression normalized
to an endogenous reference gene (actin), and relative to the untreated (non-
pollinated) control. Finally, in order to acquire relative expression, data was
processed using the 2-ΔΔCT method.
3.7. Statistical analysis
Data was processed by using Jump software (JMP Ver.9). Analysis of
variance (ANOVA) was used as statistical method for comparison of the means
with Tukey-Kramer HSD and Student test. Moreover, for the gene expression
analysis, hierarchical cluster analysis was performed.
18
4. RESULTS
4.1. Morpho-anatomical characterization of fertilization and fruit set
in field-grown date cultivars 'Medjoul' and 'Barhee'
Morphological (2012, 2014) and anatomical (2012) characterization of date
flowers and fruitlets of cv. 'Barhee' and 'Medjoul' was analyzed during the first
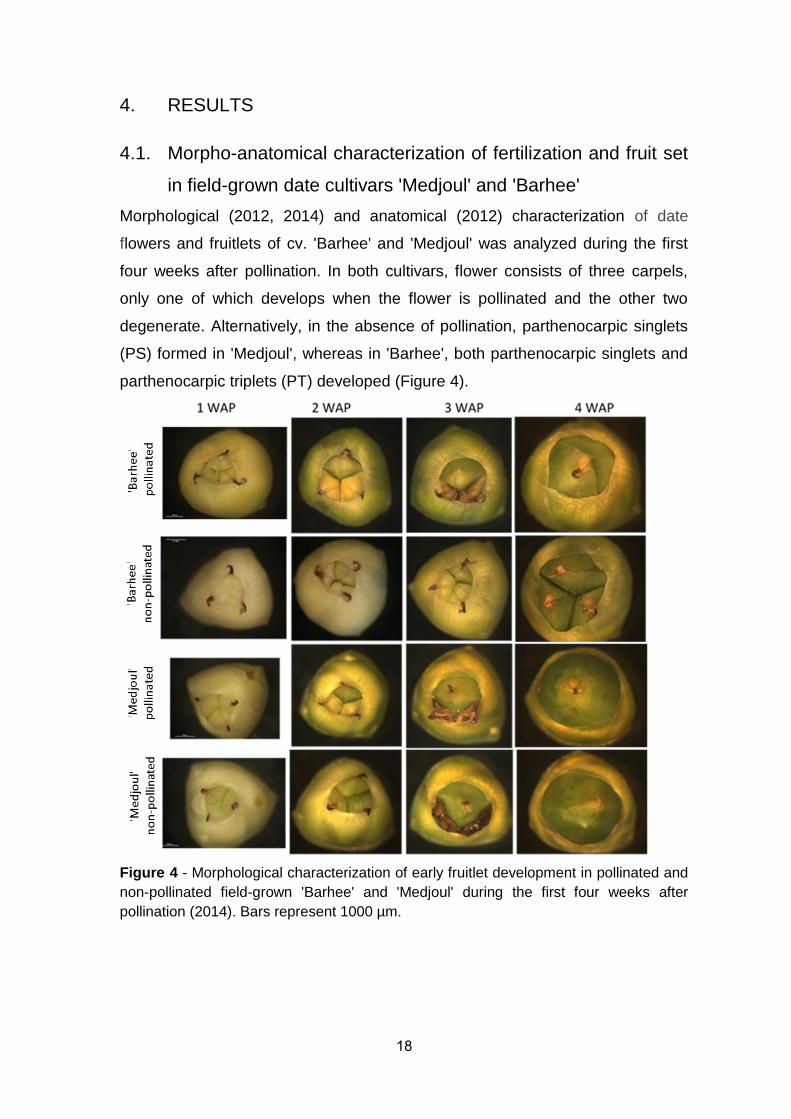
four weeks after pollination. In both cultivars, flower consists of three carpels,
only one of which develops when the flower is pollinated and the other two
degenerate. Alternatively, in the absence of pollination, parthenocarpic singlets
(PS) formed in 'Medjoul', whereas in 'Barhee', both parthenocarpic singlets and
parthenocarpic triplets (PT) developed (Figure 4).
Figure 4 - Morphological characterization of early fruitlet development in pollinated and
non-pollinated field-grown 'Barhee' and 'Medjoul' during the first four weeks after
pollination (2014). Bars represent 1000 µm.

19
4.1.1. 'Barhee'
In 'Barhee', in non-pollinated inflorescences, both PS and PT fruitlets developed.
Representative spikelets of pollinated and non-pollinated inflorescences, and a
cross section through normal, PS and PT fruitlets are presented in Figure 5. The
share of normal (seed-bearing), PS and PT, as well as shed fruitlets were
counted. In pollinated bunches only low levels of PS and PT were detected.
(Table 1, 2014). In non-pollinated flowers, high levels of parthenocarpic fruits
were detected. No difference was observed in the percentage of PS and PT.
Figure 5 - Left: Pollinated seed-bearing 'Barhee' fruitlets (a) versus non-pollinated
single and triple parthenocarpic fruitlets (b); right: cross sections of a parthenocarpic
triplet (c), parthenocarpic singlet (d), and a normal seed-bearing fruitlets (e) at 9 WAP
(2014).
Table 1 - Percentage of parthenocarpic singlets, parthenocarpic triplets, normal and
shed fruitlets respectively in 'Barhee' at 9 WAP (2014).
Shed fruitlets Normal Parthenocarpic
triple
Parthenocarpic
single 'Barhee'
51.03 ± 4.53 AB 7.14 ± 2.49 D 21.58 ± 3.84 C 20.23 ± 3.49 C No pollination
40.79 ± 3.02 B 57.00 ± 2.70 A 0.18 ± 0.18 D 0.87 ± 0.87 D Pollination

21
Figure 7 - Deterioration of two ovules in pollinated
'Barhee' as opposed to uniform ovule growth in
non-pollinated flower.
Figure 6 - Histological characterization of early fruitlet development in cv. 'Barhee'
during the first 5 WAP. Bar represents 1111 µm.
Histological analysis of pollinated and non-pollinated fruitlets revealed earlier
differences in development. In pollinated 'Barhee', we could detect a dominant
enlargement of the "chosen" carpel over the other two at 14 DAP. However, in
non-pollinated fruitlets, at this stage, all three carpels were developing at similar
rates leading to the formation of PTs. Moreover, at this stage, we could observe
deterioration of two of the ovules (in pollinated flowers) or delay in the
development of all three ovules (in non-pollinated fruitlets) (Figure 6 and 7).
21
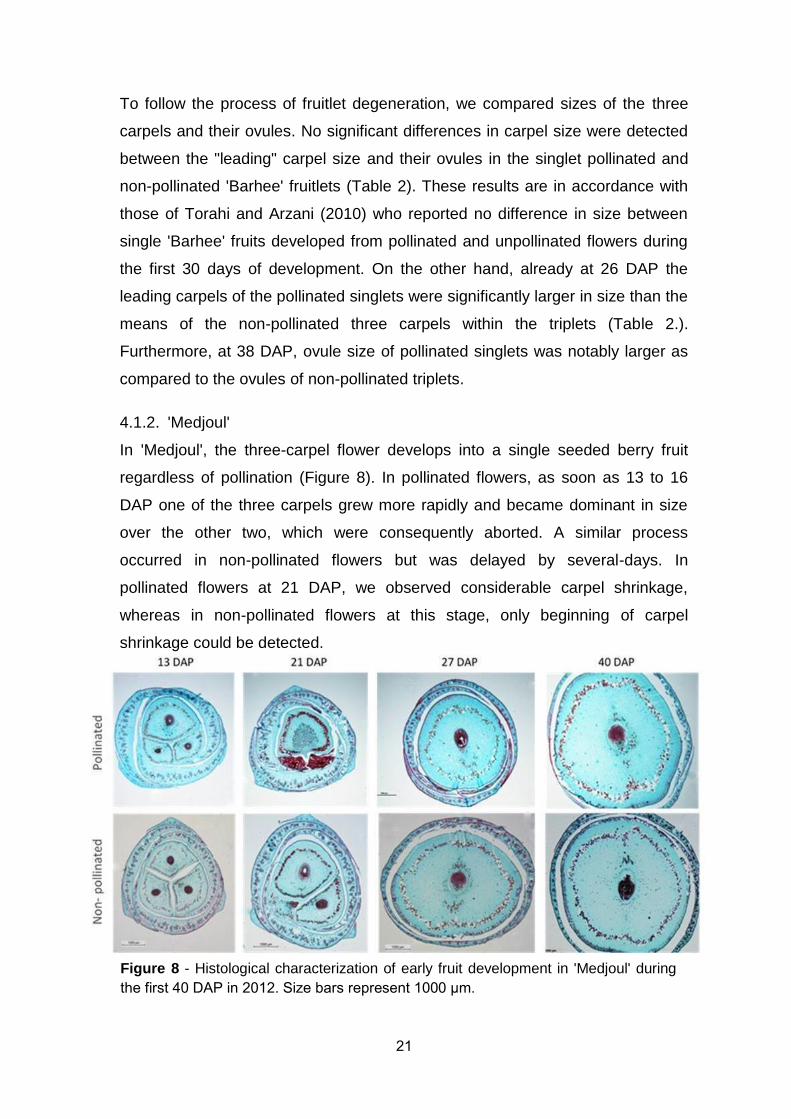
Figure 8 - Histological characterization of early fruit development in 'Medjoul' during
the first 41 DAP in 2112. Size bars represent 1111 µm.
To follow the process of fruitlet degeneration, we compared sizes of the three
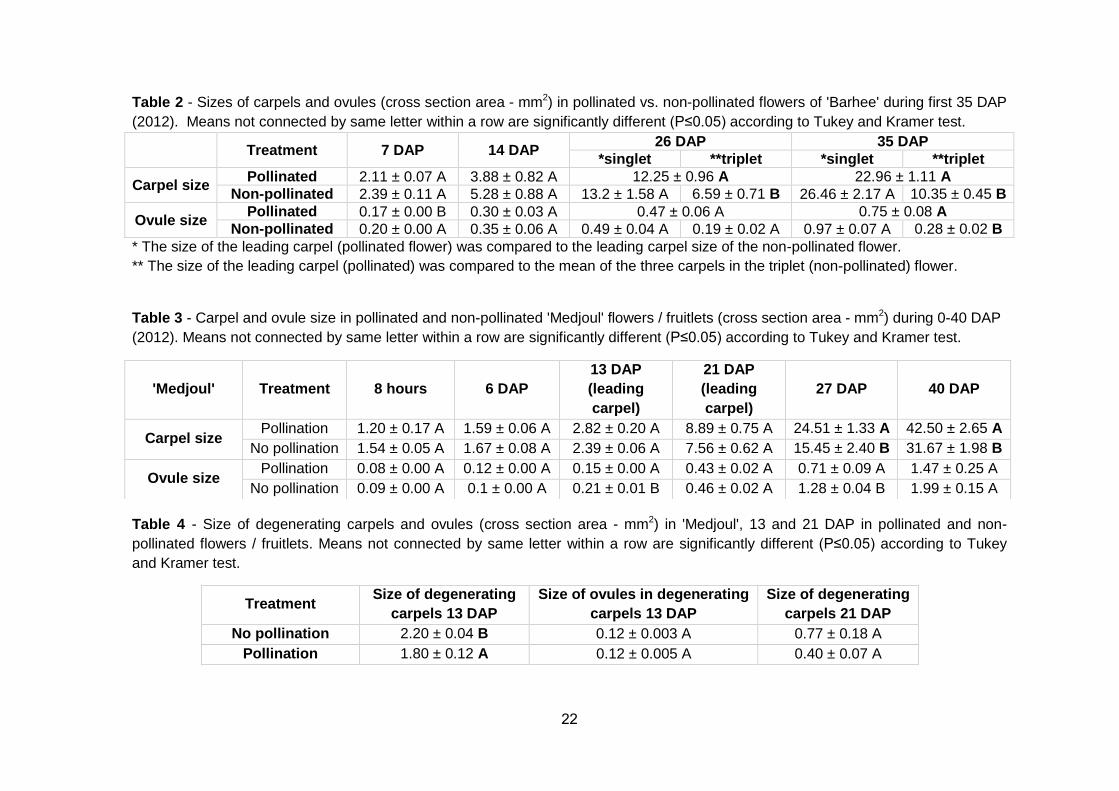
carpels and their ovules. No significant differences in carpel size were detected
between the "leading" carpel size and their ovules in the singlet pollinated and
non-pollinated 'Barhee' fruitlets (Table 2). These results are in accordance with
those of Torahi and Arzani (2010) who reported no difference in size between
single 'Barhee' fruits developed from pollinated and unpollinated flowers during
the first 30 days of development. On the other hand, already at 26 DAP the
leading carpels of the pollinated singlets were significantly larger in size than the
means of the non-pollinated three carpels within the triplets (Table 2.).
Furthermore, at 38 DAP, ovule size of pollinated singlets was notably larger as
compared to the ovules of non-pollinated triplets.
4.1.2. 'Medjoul'
In 'Medjoul', the three-carpel flower develops into a single seeded berry fruit
regardless of pollination (Figure 8). In pollinated flowers, as soon as 13 to 16
DAP one of the three carpels grew more rapidly and became dominant in size
over the other two, which were consequently aborted. A similar process
occurred in non-pollinated flowers but was delayed by several-days. In
pollinated flowers at 21 DAP, we observed considerable carpel shrinkage,
whereas in non-pollinated flowers at this stage, only beginning of carpel
shrinkage could be detected.
22
Table 2 - Sizes of carpels and ovules (cross section area - mm2) in pollinated vs. non-pollinated flowers of 'Barhee' during first 35 DAP
(2012). Means not connected by same letter within a row are significantly different (P≤0.05) according to Tukey and Kramer test.
Treatment 7 DAP 14 DAP 26 DAP 35 DAP
*singlet **triplet *singlet **triplet
Carpel size Pollinated 2.11 ± 0.07 A 3.88 ± 0.82 A 12.25 ± 0.96 A 22.96 ± 1.11 A
Non-pollinated 2.39 ± 0.11 A 5.28 ± 0.88 A 13.2 ± 1.58 A 6.59 ± 0.71 B 26.46 ± 2.17 A 10.35 ± 0.45 B
Ovule size Pollinated 0.17 ± 0.00 B 0.30 ± 0.03 A 0.47 ± 0.06 A 0.75 ± 0.08 A
Non-pollinated 0.20 ± 0.00 A 0.35 ± 0.06 A 0.49 ± 0.04 A 0.19 ± 0.02 A 0.97 ± 0.07 A 0.28 ± 0.02 B
* The size of the leading carpel (pollinated flower) was compared to the leading carpel size of the non-pollinated flower.
** The size of the leading carpel (pollinated) was compared to the mean of the three carpels in the triplet (non-pollinated) flower.
Table 3 - Carpel and ovule size in pollinated and non-pollinated 'Medjoul' flowers / fruitlets (cross section area - mm2) during 0-40 DAP
(2012). Means not connected by same letter within a row are significantly different (P≤0.05) according to Tukey and Kramer test.
Table 4 - Size of degenerating carpels and ovules (cross section area - mm2) in 'Medjoul', 13 and 21 DAP in pollinated and non-
pollinated flowers / fruitlets. Means not connected by same letter within a row are significantly different (P≤0.05) according to Tukey
and Kramer test.
'Medjoul' Treatment 8 hours 6 DAP
13 DAP
(leading
carpel)
21 DAP
(leading
carpel)
27 DAP 40 DAP
Carpel size Pollination 1.20 ± 0.17 A 1.59 ± 0.06 A 2.82 ± 0.20 A 8.89 ± 0.75 A 24.51 ± 1.33 A 42.50 ± 2.65 A
No pollination 1.54 ± 0.05 A 1.67 ± 0.08 A 2.39 ± 0.06 A 7.56 ± 0.62 A 15.45 ± 2.40 B 31.67 ± 1.98 B
Ovule size Pollination 0.08 ± 0.00 A 0.12 ± 0.00 A 0.15 ± 0.00 A 0.43 ± 0.02 A 0.71 ± 0.09 A 1.47 ± 0.25 A
No pollination 0.09 ± 0.00 A 0.1 ± 0.00 A 0.21 ± 0.01 B 0.46 ± 0.02 A 1.28 ± 0.04 B 1.99 ± 0.15 A
Treatment Size of degenerating
carpels 13 DAP
Size of ovules in degenerating
carpels 13 DAP
Size of degenerating
carpels 21 DAP
No pollination 2.20 ± 0.04 B 0.12 ± 0.003 A 0.77 ± 0.18 A
Pollination 1.80 ± 0.12 A 0.12 ± 0.005 A 0.40 ± 0.07 A
23
No significant differences in carpel size of 'Medjoul' flowers were observed in
the first 3 weeks following pollination. However, at 27 and 40 DAP, the
leading carpels of the pollinated flowers were significantly larger in size as
compared to those of the non-pollinated ones. Moreover, the ovules of the
leading carpels were larger in size in pollinated flowers, as compared to the
non-pollinated flowers, being significant at 13 and 27 DAP (Table 3).
Comparing sizes of ovules in degenerating carpels, we reported no statistical
difference among treatments. Nevertheless, at 13 DAP, the two degenerating
carpels were significantly larger in size in non-pollinated fruitlets, as
compared to those of the pollinated fruitlets, showing that the process of
carpel abortion starts earlier and occurs at a higher rate in pollinated fruitlets
(Table 4).
4.1.3. Comparison between 'Barhee' and 'Medjoul'
In pollinated 'Barhee' and 'Medjoul', first indications of programmed cell death
were reported 14 and 21 DAP respectively, through ovule deterioration and
carpel shrinkage. In pollinated 'Medjoul', carpel degeneration was faster as
compared to the non-pollinated 'Medjoul'.
'Barhee' and 'Medjoul' differ in their shedding of fruitlets. Fruit-drop was
compared at 3, 4, and 9 WAP (Table 5), showing great differences in
response to pollination. Highest fruit shedding was reported in non-pollinated
'Medjoul', differing from other treatments at all examination times.
Table 5 - Percentage of fruitlet-drop (abscission) in 'Barhee' versus 'Medjoul' 3, 4
and 9 WAP respectively grown in the field (March-April 2014). Means not connected
by same letter within a row are significantly different (P≤0.05) according to Tukey
and Kramer test.
63 DAP 28 DAP 21 DAP Treatment Cultivar
40.80 ± 3.02 BC 17.57 ± 3.57 B 6.5 ± 1.68 B Pollination 'Barhee'
51.03 ± 4.53 B 14.43 ± 2.87 B 3.72 ± 1.52 B No pollination
28.77 ± 3.28 C 21.19 ± 7.25 B 15.32 ± 2.85 B Pollination 'Medjoul'
90.04 ± 2.02 A 68.5 ± 5.51 A 40.82 ± 9.62 A No pollination

24
4.2. Effects of temperature regimes on pollination, fertilization
and fruit-set in date palm
4.2.1. Development of an in vitro assay for studying date palm fertilization
The in vitro fertilization assay was developed with the aim to study date
fertilization of isolated pollinated spikelets under controlled environmental
conditions: We tried to optimize survival conditions of spikelets, flowers and
organs incubated in different media and under different temperature regimes.
This section was performed in collaboration with David Birger, another
master student in the laboratory.
Optimization of growth media and cultivation conditions in vitro
In order to improve "vase life" of the flowers and spikelets, we calibrated the
in vitro assay. The vase life of spikelets was tested in five growth media:
Three different liquid media were tested: T.O.G.6, T.O.G.6 + 2% sucrose,
“Longlife” and solid agar media. Spikelets of 'Medjoul' were incubated at
three different temperature regimes: 20°C, 25°C and 30°C (Table 6).
In order to prevent fungal contamination, 0.2% Marpan fungicide was added
(dipped) to the media with spikelets which was then replaced every 7 days.
Without the use of Marpan, all the spikelets were contaminated after only two
days under all temperature treatments (data not shown).
In October 2013, we used Canary palm in order to calibrate in vitro assay
and extend spikelet vase-life. Both palms are close relatives and possess
similar reproductive mechanisms, but vary in their annual cycle and flowering
season. Date palm flowers during relatively short season in March-April,
while Canary palm flowers in October. Therefore, we used Canary palm to
complement our main research to obtain plant material out of the date
flowering season.

25
Table 6 - Effect of culture media and temperature on isolated spikelets viability and flower abscission of 'Medjoul'. Spikelet
sections were incubated at different temperatures, their viability and flower abscission was estimated at 1, 5 and 9 days
after setup (DAS) using a five-point scale, where 5 – is completely viable and 0 being dead / most contaminated. Means ±
standard errors are significantly different (P≤0.05) according to the Tukey-Kramer HSD test.
Temp. Culture media Overall vitality Abscission
1 DAS 5 DAS 9 DAS 1 DAS 5 DAS 9 DAS
20˚C TOG 5 ± 0 a 4.9 ± 0.1 a 5 ± 0 d 5 ± 0 a 4.9 ± 0.1 ab 5± 0 c
TOG+2% sucrose 5 ± 0 a 5 ± 0 a 5 ± 0 d 5 ± 0 a 0 ± 0 b 5 ± 0 c
LongLife 4.8 ± 0.3 a 4.3 ± 0.6 ab 3.5 ± 1.2 bcd 5 ± 0 a 4.9 ± 0.1 ab 3.6 ± 1.2 abc
DDW 4.8 ± 0.1 a 3.1 ± 0.6 abc 1 ± 1 ab 5 ± 0 a 4.8 ± 0.3 ab 1.1 ± 1.1 abc
MS Agar + 3% Sucrose 5 ± 0 a 4.1 ± 0.6 ab 1.8 ± 1.2 abc 5 ± 0 a 0 ± 0 b 2.3 ± 1.3 abc
25˚C TOG 5 ± 0 a 4.8 ± 0.1 a 1.5 ± 0.9 ab 5 ± 0 a 4.8 ± 0.1 ab 2.8 ± 1.0 abc
TOG+
2% sucrose
5 ± 0 a 5 ± 0 a 4.5 ± 0.3 cd 5 ± 0 a 4.5 ± 0 ab 4.6 ± 0.2 bc
LongLife 3.8 ± 0.4 b 2 ± 0.8bc 0 ± 0 a 5 ± 0 a 2.6 ± 1.0 a 0 ± 0 a
DDW 4.3 ± 0.1 ab 3.5 ± 0.5 abc 0 ± 0 a 4.9 ± 0.1 a 4.6 ± 0.1 ab 0 ± 0 a
MS Agar + 3% Sucrose 4.9 ± 0.1 a 3.5 ± 0.9 abc 0 ± 0 a 5 ± 0 a 3.8 ± 0.7 ab 2.3 ± 1.3 abc
30˚C TOG 5 ± 0 a 3.3 ± 0.5 abc 0 ± 0 a 5 ± 0 a 4.1 ± 1.5 ab 0.8 ± 0.8 ab
TOG+
2% sucrose
5 ± 0 a 4.1 ± 0.6 ab 0 ± 0 a 5 ± 0 a 4.1 ± 0.7 ab 2.0 ± 0.9 abc
LongLife 4.8 ± 0.3 a 3.5 ± 0.8 abc 0 ± 0 a 5 ± 0 a 4.1 ± 0.4 ab 0.8 ± 0.8 ab
DDW 4.8 ± 0.1 a 2.8 ± 0.3 abc 0 ± 0 a 5 ± 0 a 3.9 ± 0.4 ab 0 ± 0 a
MS Agar + 3% Sucrose 5 ± 0 a 1 ± 0 c 0 ± 0 a 5 ± 0 a 4.3 ± 0.3 ab 0 ± 0 a

26
Cut spikelets of Canary palm were treated with ethylene inhibitors, 1-
Methylcyclopropene (1-MCP) or silver thiosulfate (STS) aiming to hinder
senescence – the time-related deterioration of the physiological functions,
and support in vitro development of the spikelets. Both treatments extended
"vase life" of the flowers and spikelets (Figure 9). Effects of 1-MCP and STS
on different physiological parameters were tested in vitro and evaluated on a
five-point scale (e.g. 0 being most contaminated, 5 – no contamination)
(Figure 10).
Pollination greatly reduced viability, the most significant parameter of vase-
life in STS-treated spikelets as compared to 1-MCP-treated. Pollinated
spikelets treated with 1-MCP remained viable 13 DAP, i.e. they reached the
average grade of 3; while the STS-treated and control flowers have been
already dried at this time, reaching grade 3 already between 4-7 DAP. Grade
3 was used as indicator of spikelet half-life even though at this point it was
already late to use flowers for physiological studies due to pronounced
senescence. On the other hand, in non-pollinated flowers, viability of the STS
and 1-MCP treated flowers was insignificant.
Figure 9 - Effects of Ethylene inhibitors on spikelets "vase life" 10 days after
culturing. Left: First four samples represent non-pollinated STS-treated spikelets with
control; whereas additional four are pollinated STS-treated spikelets. Right: Non-
pollinated-1-MCP treated spikelet and non-pollinated control, and 1-MCP-treated
spikelets 1 and 2 DAP respectively, with the control.

27
Abscission, the natural detachment of plant organs (fruitlets), was
more pronounced in pollinated flowers as compared to non-pollination
control, and 1-MCP treatment reduced abscission more efficiently than STS
(Figure 10c and 10d). On the other hand, with regard to senescence, no
difference was observed in pollinated flowers versus the non-pollination
control.
Both anti-ethylene treatments extended "vase life" of the flowers and
spikelets, showing different effects on particular physiological parameters
(Figure 10). The flowers were healthier, less flowers had dropped 3-10 DAP
and lower fungal contamination was observed (data not shown). However,
overall, we can report that flowers treated with 1-MCP had vase-life of 13
days, whereas pollinated and non-pollinaed STS-treated spikelets were
deteriorated after 6 or 12 days respectively. Neither of these treatments was
sufficient to maintain in vitro system long enough and allow us to focus on
fruitlet development.
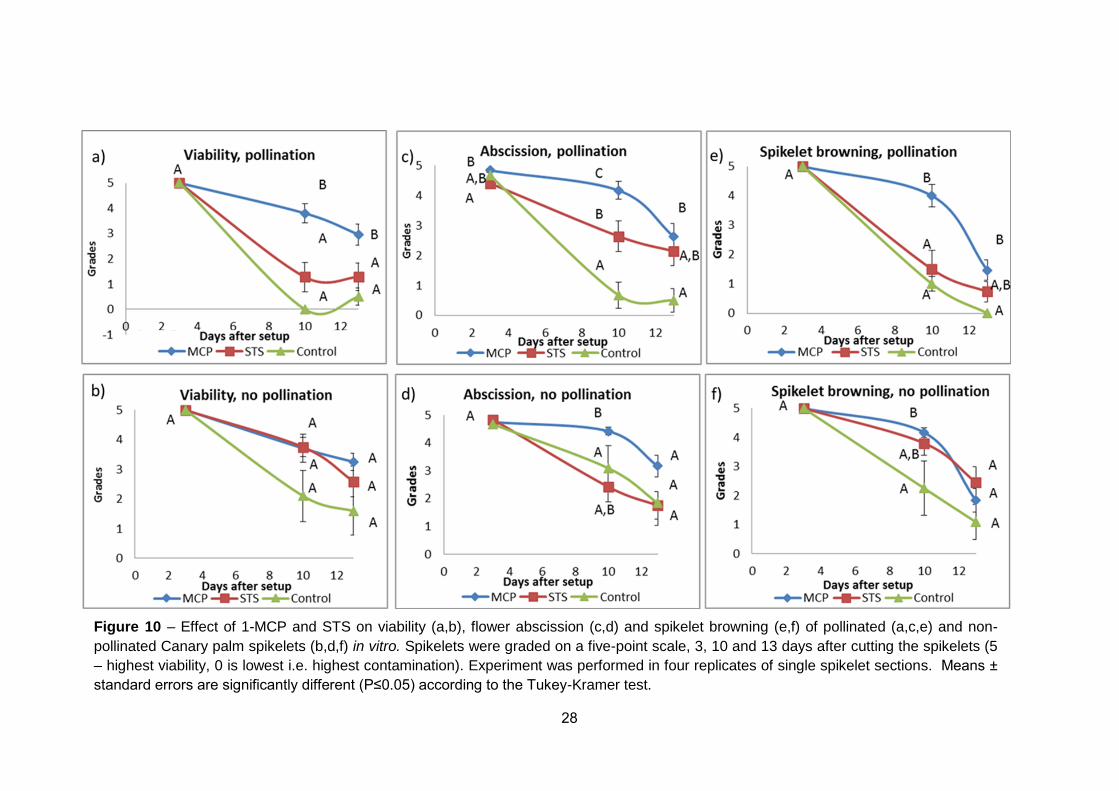
28
Figure 10 – Effect of 1-MCP and STS on viability (a,b), flower abscission (c,d) and spikelet browning (e,f) of pollinated (a,c,e) and non-
pollinated Canary palm spikelets (b,d,f) in vitro. Spikelets were graded on a five-point scale, 3, 10 and 13 days after cutting the spikelets (5
– highest viability, 0 is lowest i.e. highest contamination). Experiment was performed in four replicates of single spikelet sections. Means ±
standard errors are significantly different (P≤1.15) according to the Tukey-Kramer test.

29
In order to improve "vase life" of 'Medjoul' flowers in vitro, we tried to
culture larger sections of inflorescences instead of single spikelet sections.
These were pollinated and exposed to four alternating temperature regimes
(Figure 11).
Pollination had no significant effect on tested physiological parameters.
However, the temperature conditions greatly affected vase life of the flowers
(Figure 12), as well as their fungal contamination (data not shown).
Senescence and overall inflorescence deterioration were first reported only 6
days after setup at highest temperatures (34/28˚C). On the other hand,
deterioration was slowest in inflorescences exposed to lowest temperature
treatment (16/10˚C) which had longest vase life of approximately 15 days.
Figure 11 - Pollinated and non-pollinated inflorescences of 'Medjoul' exposed to
four temperature regimes 10 days after setup, in three biological replicates per
treatment.

31
Figure 12 – Effect of four temperature regimes on pollinated and non-pollinated
inflorescences of 'Medjoul' in vitro. Bunches were graded for carpel health, browning
and for fruitlet drop using three replicates per treatment, on a five-point scale (e.g. 5
– being most healthy, whereas 0 – the least). Bars represent standard errors.

31
Figure 13 - Pollen tube length of 'Medjoul' 9 hours after incubation at 4 constant
temperature regimes. Bars represent 51 µm.
4.2.2. Effect of temperature regimes on pollination and fertilization
Effects of temperature regimes on pollen germination and pollen tube
elongation in artificial media in vitro
Pollen grains were germinated in vitro at different constant temperatures, and
pollen tube length was measured after 3, 6 and 9 hours respectively (Figure
13).
Pollen germination in vitro was strongly influenced by temperature. At 15°C
pollen growth rate was slower and pollen tube elongation was retarded as
compared to those at 20-30°C (Figure 14). Eventually, the highest pollen tube
length was recorded at 25°C suggesting that this temperature may be optimal
for pollen germination and tube elongation of cultivar 'Medjoul'.
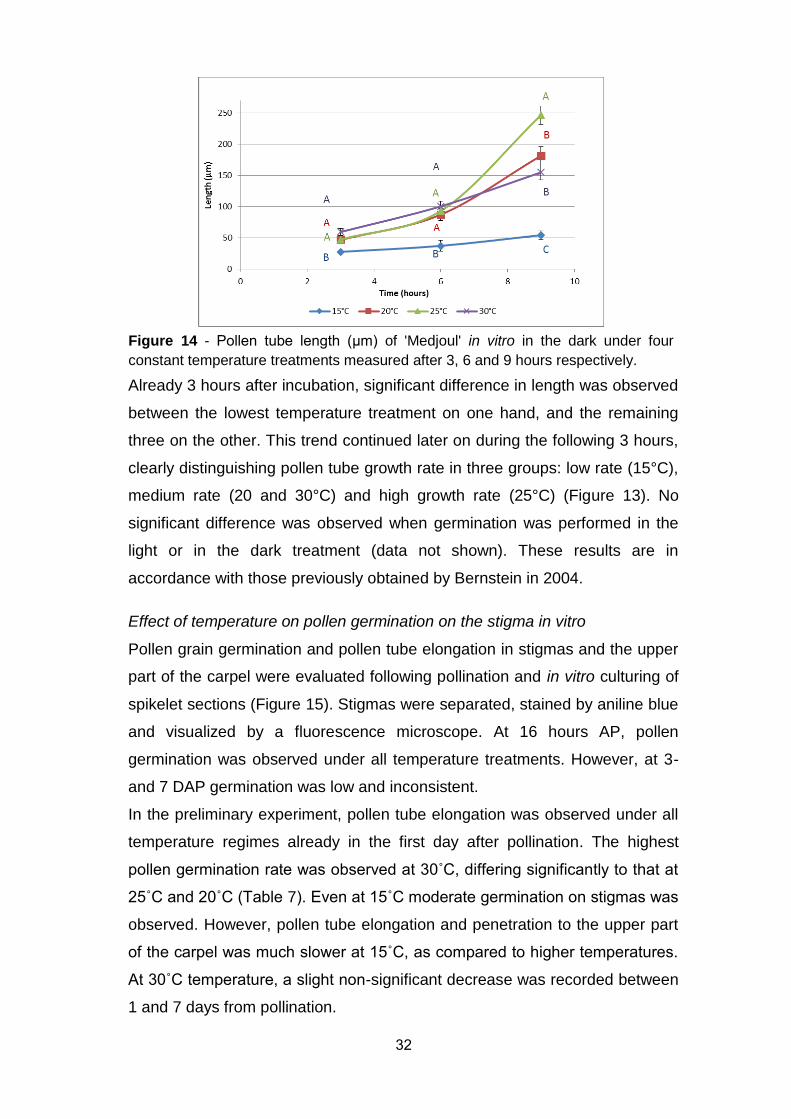
32
Already 3 hours after incubation, significant difference in length was observed
between the lowest temperature treatment on one hand, and the remaining
three on the other. This trend continued later on during the following 3 hours,
clearly distinguishing pollen tube growth rate in three groups: low rate (15°C),
medium rate (20 and 30°C) and high growth rate (25°C) (Figure 13). No
significant difference was observed when germination was performed in the
light or in the dark treatment (data not shown). These results are in
accordance with those previously obtained by Bernstein in 2004.
Effect of temperature on pollen germination on the stigma in vitro
Pollen grain germination and pollen tube elongation in stigmas and the upper
part of the carpel were evaluated following pollination and in vitro culturing of
spikelet sections (Figure 15). Stigmas were separated, stained by aniline blue
and visualized by a fluorescence microscope. At 16 hours AP, pollen
germination was observed under all temperature treatments. However, at 3-
and 7 DAP germination was low and inconsistent.
In the preliminary experiment, pollen tube elongation was observed under all
temperature regimes already in the first day after pollination. The highest
pollen germination rate was observed at 30˚C, differing significantly to that at
25˚C and 20˚C (Table 7). Even at 15˚C moderate germination on stigmas was
observed. However, pollen tube elongation and penetration to the upper part
of the carpel was much slower at 15˚C, as compared to higher temperatures.
At 30˚C temperature, a slight non-significant decrease was recorded between
1 and 7 days from pollination.
Figure 14 - Pollen tube length (µm) of 'Medjoul' in vitro in the dark under four
constant temperature treatments measured after 3, 6 and 9 hours respectively.

33
Figure 15 - Pollen germination on 'Medjoul' stigmas in vitro under 4 constant
temperatures visualized using fluorescence microscopy. Bar represents 50 µm.
Table 7 - Effects of four temperatures on pollen tube elongation in isolated 'Medjoul'
stigma sections in vitro. Pollen germination on the stigma was estimated on a five-
point scale, where 0 – no germination; 4 – high germination. Four flowers were used
per temperature treatment i.e. twelve stigmas. Statistical analysis included samples
from different temperature and DAPs. Means are significantly different (P≤0.05)
according to the Tukey-Kramer HSD test. *DAP- days after pollination.
Temperature,
day/night 16 hours AP 3 DAP 7 DAP
15˚C 1.95 ± 0.29 bcd 1.45 ± 0.29 c 1.62 ± 0.29 d
20˚C 1.37 ± 0.29 d 1.75 ± 0.29 bcd 1.68 ± 0.36 bcd
25˚C 1.5 ± 0.29 d 1.91 ± 0.29 bcd 1.35 ± 0.38 d
30˚C 3.41 ± 0.29 a 3.08 ± 0.29 ab 3.00 ± 0.29 abc
4.2.3. Effects of temperature regimes on fertilization and fruit set in "modular
phytotrons" - in vivo
Due to germination inconsistency in the in vitro assay, as well as the inability
to support "vase-life" of cut inflorescences long enough to study fruit
development, we attempted to induce controlled temperature on the bunches
using special units we called "modular phytotrons", and study pollen
germination and fertilization on bunch on the trees in the orchard in vivo
(Figure 2).

34
Characterization of the modular phytotrons
Following pollination, "modular phytotrons" were installed on the trees around
individual pollinated inflorescences and sealed during the period of 14.03-
22.04.2013 (40 days) and 18.03-01.04.2014 (14 days). After removing the
units, further fruitlet development was followed under external weather
conditions. The setup and maintenance of the "modular phytotrons" was done
by Avi Sadowsky and the team of the Southern Arava R & D.
Temperature variation inside the "modular phytotrons" were recorded during
the season of 2013 (Figure 16). In general, the units induced different
temperature regimes. However, while the high temperature units operated
almost as planned, those of the medium and lower temperatures did not
(Figure16 B-C), Variation in the low temperature units was much higher as
compared to that in high temperature units (Figure 16 A-C), and during very
warm days could not keep the required temperatures. Still, the two units more
successful in reducing the temperatures had significantly lower temperatures
than the other treatments (Figure 16 D).
Effects of temperature regimes on pollen germination on stigma in vivo
Pollen germination on stigmas was reported under all temperature regimes
(Figure 17). However, in the cool treatment (20/8°C) pollen germination was
not detected at 16 hours after pollination and was also delayed at 3 DAP, as
compared with the medium (25/12°C), warm treatment (32/18°C) and the
non-controlled bunches (Figure 18). The highest germination and pollen tube
elongation was observed in the warm treatment, followed by the medium,
control and the cool treatment. At 7 DAP there was no difference in pollen
tube growth between the treatments, suggesting that even in cool
temperatures pollen grains can germinate on the stigma.
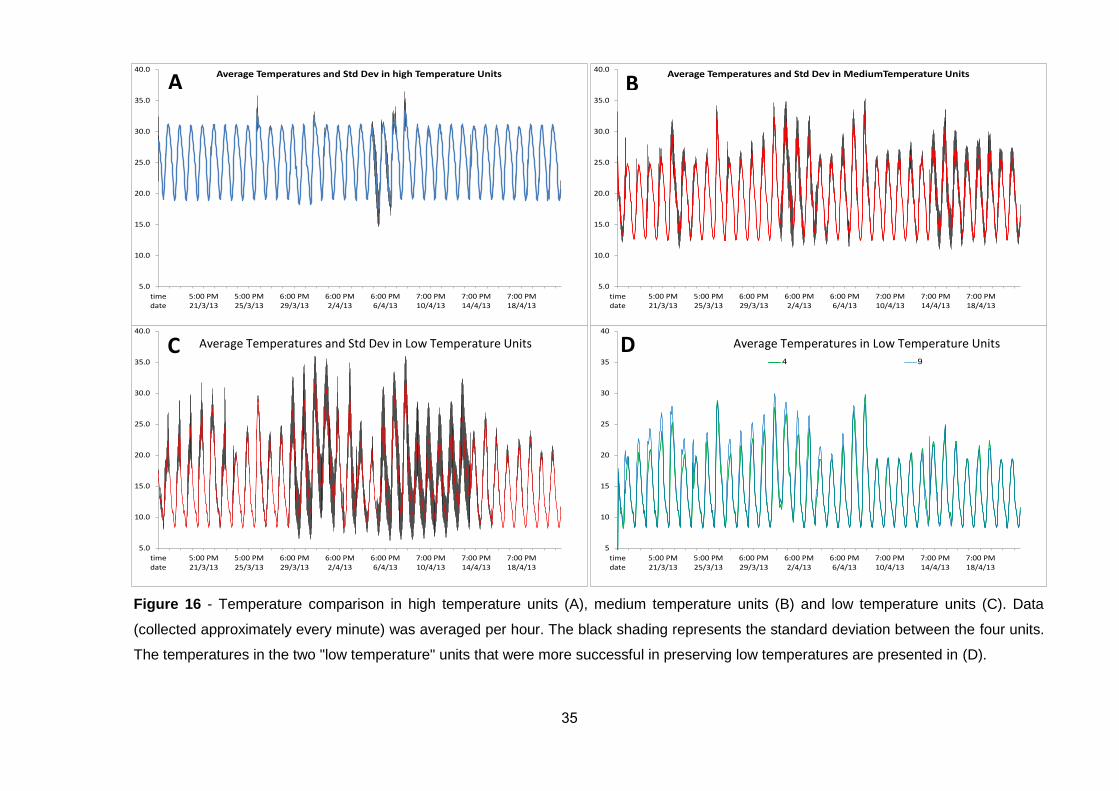
35
5.0
10.0
15.0
20.0
25.0
30.0
35.0
40.0
timedate
5:00 PM21/3/13
5:00 PM25/3/13
6:00 PM29/3/13
6:00 PM2/4/13
6:00 PM6/4/13
7:00 PM10/4/13
7:00 PM14/4/13
7:00 PM18/4/13
Average Temperatures and Std Dev in high Temperature Units
5.0
10.0
15.0
20.0
25.0
30.0
35.0
40.0
timedate
5:00 PM21/3/13
5:00 PM25/3/13
6:00 PM29/3/13
6:00 PM2/4/13
6:00 PM6/4/13
7:00 PM10/4/13
7:00 PM14/4/13
7:00 PM18/4/13
Average Temperatures and Std Dev in MediumTemperature Units
5.0
10.0
15.0
20.0
25.0
30.0
35.0
40.0
timedate
5:00 PM21/3/13
5:00 PM25/3/13
6:00 PM29/3/13
6:00 PM2/4/13
6:00 PM6/4/13
7:00 PM10/4/13
7:00 PM14/4/13
7:00 PM18/4/13
Average Average Temperatures and Std Dev in LowTemperature Units
5
10
15
20
25
30
35
40
timedate
5:00 PM21/3/13
5:00 PM25/3/13
6:00 PM29/3/13
6:00 PM2/4/13
6:00 PM6/4/13
7:00 PM10/4/13
7:00 PM14/4/13
7:00 PM18/4/13
4 9
A B
C D Average Temperatures in Low Temperature Units Average Temperatures and Std Dev in Low Temperature Units
Figure 16 - Temperature comparison in high temperature units (A), medium temperature units (B) and low temperature units (C). Data
(collected approximately every minute) was averaged per hour. The black shading represents the standard deviation between the four units.
The temperatures in the two "low temperature" units that were more successful in preserving low temperatures are presented in (D).
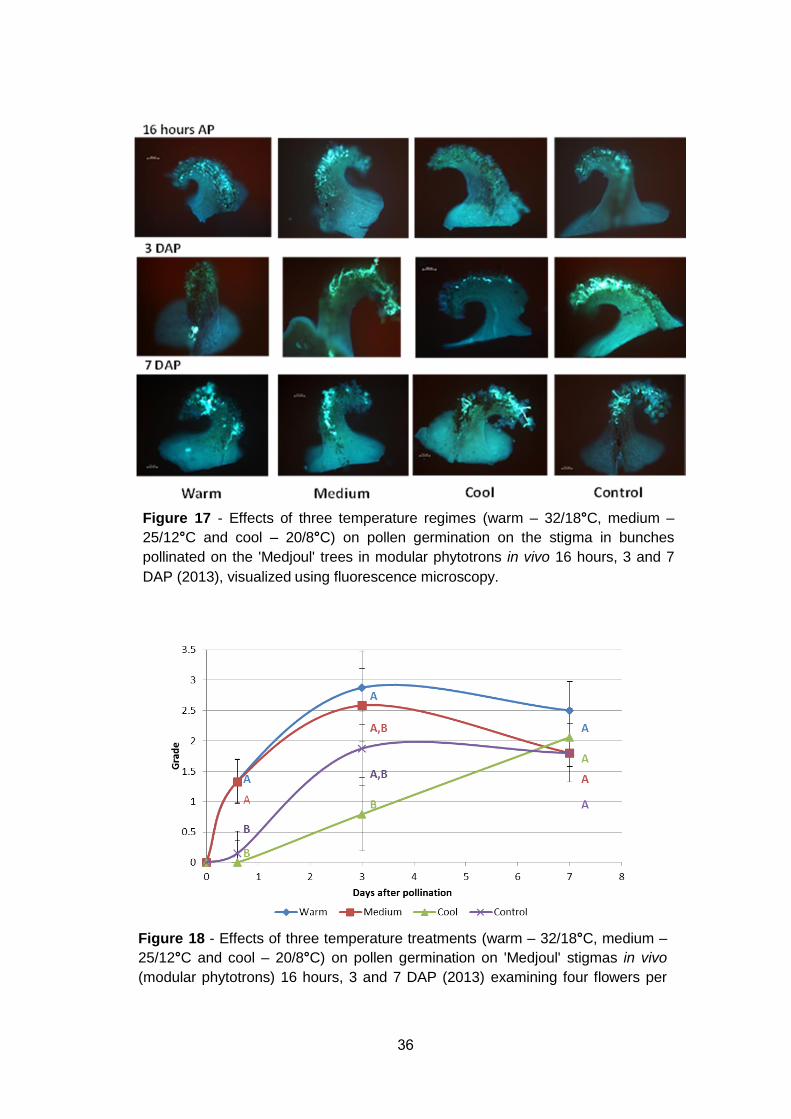
36
Figure 17 - Effects of three temperature regimes (warm – 32/18°C, medium –
25/12°C and cool – 20/8°C) on pollen germination on the stigma in bunches
pollinated on the 'Medjoul' trees in modular phytotrons in vivo 16 hours, 3 and 7
DAP (2013), visualized using fluorescence microscopy.
Figure 18 - Effects of three temperature treatments (warm – 32/18°C, medium –
25/12°C and cool – 20/8°C) on pollen germination on 'Medjoul' stigmas in vivo
(modular phytotrons) 16 hours, 3 and 7 DAP (2013) examining four flowers per
time point.

37
Effects of temperature regimes in vivo on fruitlet development of 'Medjoul'
The temperature treatments affected growth of fruitlets. Representative
fruitlets on spikelet section from each unit 5 WAP are presented in Figure 19
(2013). The fruitlets were much bigger under warmer condition (Table 8,
Table 9. Figure 20). Even though
variation from the required
temperature was observed in the units
(see above), the fruitlets in all four
repeats seems equally affected. The
fruitlets were smaller in the four cool
treatments and large in the four warm
treatments. Moreover, a distinctive
difference in colour development was
observed. However, temperature did
not affect the spikelet elongation and
the distances between the fruitlets on the spikelets did not vary significantly
(Table 8).
Table 8 – Variation in fruitlet weight of 'Medjoul' 5 WAP (2013) in
temperature controlled units. Means are significantly different (P≤0.05)
according to the Tukey-Kramer HSD test.
Treatment Units Fruitlet per cm spikelet
(means)
Single fruitlet weight (g) (means)
Fruitlet weight per cm spikelet (g)
(means)
Control 4 1.5 1.89 B 2.84 B
Warm 4 1.6 3.12 A 4.99 A
Medium 4 1.9 1.22 BC 2.20 BC
Cold 4 1.5 9.1 C 1.38 C
*Connecting Letters Report refers both to the single fruitlet weight and fruitlet weight
per cm spikelet.
Table 9 - Effect of different temperature treatments in vivo on fruitlet weight of
'Medjoul' 9 WAP in the season of 2013 and 10 WAP in 2014 respectively. Means
are significantly different (P≤0.05) according to the Tukey-Kramer HSD test.
Treatments Single fruitlet weight (g)
Means ± Std. Errors (2013)
Single fruitlet weight (g) Means
± Std. Errors (2014)
Control 2.98 ± 0.12 A 4.58 ± 0.13 A
Warm 3.76 ± 0.14 B 7.28 ± 0.34 B
Medium 1.78 ± 0.12 C 4.39 ± 0.10 A
Cool 1.48 ± 0.14 C 2.10 ± 0.08 C
Figure 19 'Medjoul' fruitlets 5 weeks
following pollination in temperature
controlled units (2013).
38
Figure 21 - Percentage of normal, parthenocarpic, aborted and non-developed
fruits of 'Medjoul', in response to pollination and growth under different
temperatures in vivo, 10 WAP in 2013. Fruitlets from middle parts of eight
spikelets taken from two temperature units of the same treatment were used for
evaluation. Means are significantly different (P≤1.15) according to the Tukey-
Kramer HSD test.
B B A A, B
B B A, B A
A, B A B A, B
A, B A, B B A
The percentage of normal, parthenocarpic, aborted and non-developed
fruitlets was measured ten weeks following pollination and incubation under
different temperatures in vivo (2013). Significant differences were found
between treatments in all tested parameters (Figure 21). Development of
normal fruits was enhanced in the warm treatment as compared to the
medium and cool.
The percentage of parthenocarpic fruits was significantly higher under cool
Figure 20 - Effect of three temperature treatments in vivo on 'Medjoul' fruitlet size
9 WAP (2014).

39
Figure 22 - Percentage of normal, parthenocarpic, aborted and non-
developed 'Medjoul' fruits, in response to pollination and growth under
different temperatures in vivo, 10 WAP in 2014. Fruitlets from middle
parts of eight spikelets taken from two temperature units of the same
treatment were used for evaluation. Means are significantly different
(P≤1.15) according to the Tukey-Kramer HSD test.
treatment in 2013, as compared to the warm treatment and the control
(Figure 21). In the following season (2014), the experiment was repeated,
and once again we confirmed that parthenocarpic fruit formation is
enhanced by the cool temperature treatment (Figure 22).
Histological characterization of effects of temperatures on fruitlet
development in vivo
Histological cross sections of pollinated 'Medjoul' dates showed that the three
temperature treatments in vivo affected the rate of early fruitlet development
(Figure 23). The rate of overall fruitlet development was highest in the warm
unit, followed by the medium and the cool unit. In addition, carpel abortion
was detected in the warm treatment, already 7 DAP, as opposed to the cool
unit, where abortion was still ongoing at 38 DAP. Both carpel size and ovule
size were greatly affected by the "modular phytotron" temperature treatments
(Table 10, 2013).
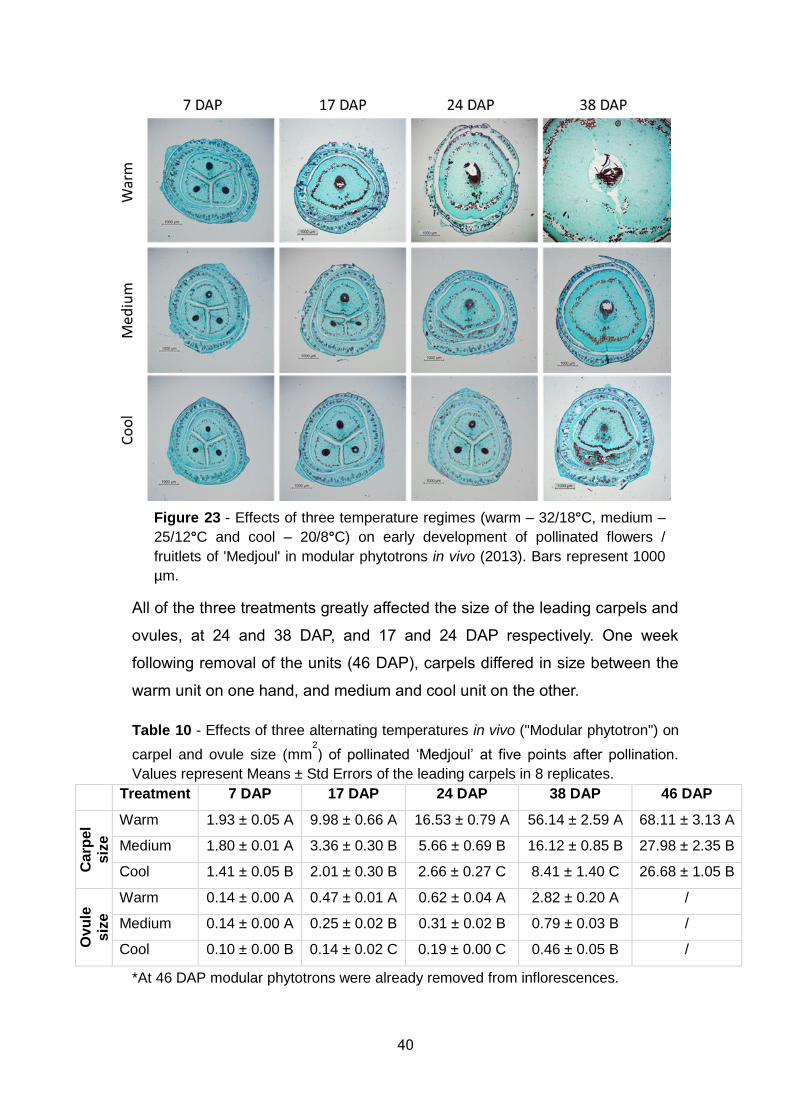
41
All of the three treatments greatly affected the size of the leading carpels and
ovules, at 24 and 38 DAP, and 17 and 24 DAP respectively. One week
following removal of the units (46 DAP), carpels differed in size between the
warm unit on one hand, and medium and cool unit on the other.
Table 10 - Effects of three alternating temperatures in vivo ("Modular phytotron") on
carpel and ovule size (mm2
) of pollinated ‘Medjoul’ at five points after pollination.
Values represent Means ± Std Errors of the leading carpels in 8 replicates.
Treatment 7 DAP 17 DAP 24 DAP 38 DAP 46 DAP
Ca
rpel
siz
e
Warm 1.93 ± 0.05 A 9.98 ± 0.66 A 16.53 ± 0.79 A 56.14 ± 2.59 A 68.11 ± 3.13 A
Medium 1.80 ± 0.01 A 3.36 ± 0.30 B 5.66 ± 0.69 B 16.12 ± 0.85 B 27.98 ± 2.35 B
Cool 1.41 ± 0.05 B 2.01 ± 0.30 B 2.66 ± 0.27 C 8.41 ± 1.40 C 26.68 ± 1.05 B
Ov
ule
siz
e
Warm 0.14 ± 0.00 A 0.47 ± 0.01 A 0.62 ± 0.04 A 2.82 ± 0.20 A /
Medium 0.14 ± 0.00 A 0.25 ± 0.02 B 0.31 ± 0.02 B 0.79 ± 0.03 B /
Cool 0.10 ± 0.00 B 0.14 ± 0.02 C 0.19 ± 0.00 C 0.46 ± 0.05 B /
*At 46 DAP modular phytotrons were already removed from inflorescences.
Figure 23 - Effects of three temperature regimes (warm – 32/18°C, medium –
25/12°C and cool – 20/8°C) on early development of pollinated flowers /
fruitlets of 'Medjoul' in modular phytotrons in vivo (2013). Bars represent 1000
µm.

41
4.3. Expression analysis of genes involved in hormonal
regulation during early fruit development of cultivars
'Medjoul' and 'Barhee'
High-throughput gene expression analysis of pollinated and non-pollinated
flowers was performed in two date cultivars 'Medjoul' and 'Barhee', at four
stages of early fruit development. Using the recently published date palm
transcriptome data (Al-Mssallem, 2013), we selected ninety six candidate
genes/transcripts, associated with metabolism and signaling of main plant
hormones auxin, gibberellin, cytokinin, abscisic acid, and ethylene, as well as
with the processes of plant senescence and programmed cell death. Further
data mining and literature search were based on the published date
transcriptome, KEGG and NCBI databases. Basic local alignment search
tool (BLAST) was used to specify the identity and similarity of the sequences,
obtained from the date transcriptome and matching them with annotated
genes/proteins deposited in the NCBI database. With regard to different
paralogs from the same gene family, we included sequences with both high
and low relative expression, according to the transcriptome. All the selected
sequences were aligned in 5'-3' direction by checking open reading frames.
Specific primers were designed for the selected genes (Appendix). Four
constitutively expressed genes were used as references. The analysis was
performed with Microfluidic Dynamic Array (BioMark, Fluidigm).
The initial analysis, filtering and cleaning of the obtained data resulted in
elimination of the sequences of ethylene response transcription factor
(ERTF), gibberellic acid insensitive dwarf (GID2) and Bax inhibitor (BI), due
to very low expression. Moreover, two replicates of 'Medjoul' (stage 0, before
pollination, and stage of two WAP) were disregarded as their expression was
low in the reaction. Actin was selected as a reference gene, due to its highest
consistency throughout the developmental stages. Gene expression was
analyzed using the Fluidigm software, and the obtained heat map of
EvaGreen Ct values is presented in Figure 24. This map presents a large
overview of the selected candidate genes Ct values and relative expression
for all the selected transcripts.
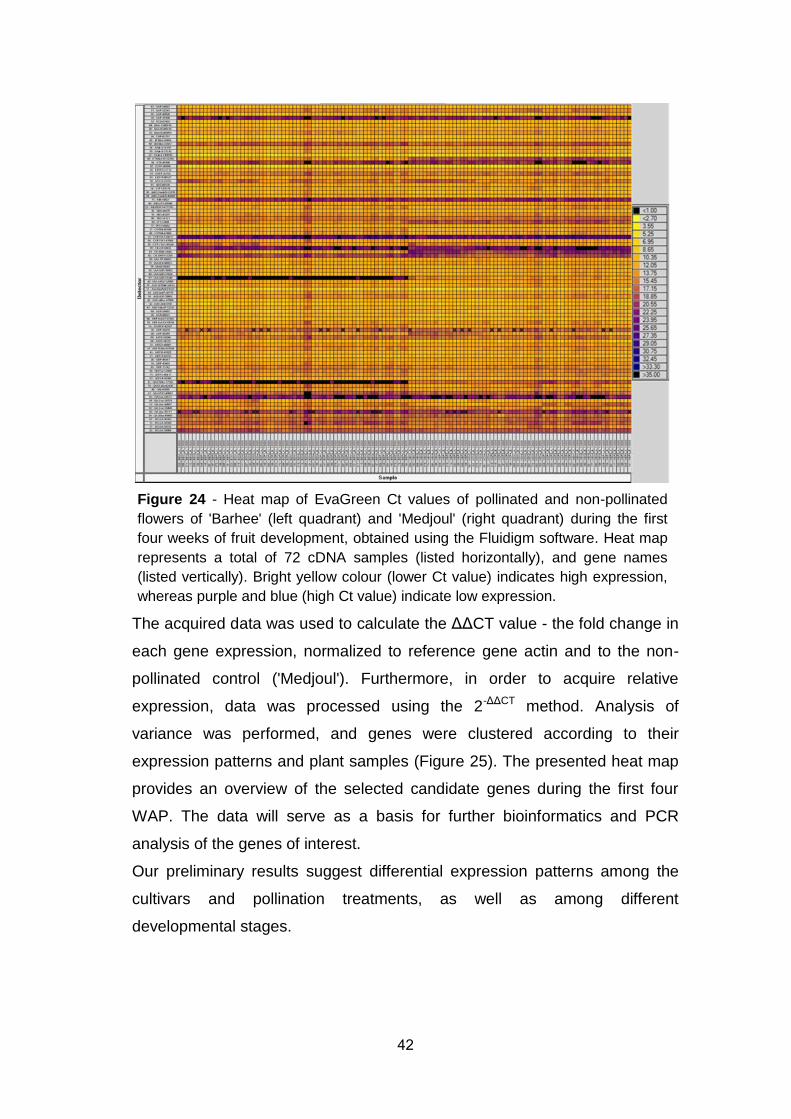
42
The acquired data was used to calculate the ΔΔCT value - the fold change in
each gene expression, normalized to reference gene actin and to the non-
pollinated control ('Medjoul'). Furthermore, in order to acquire relative
expression, data was processed using the 2-ΔΔCT method. Analysis of
variance was performed, and genes were clustered according to their
expression patterns and plant samples (Figure 25). The presented heat map
provides an overview of the selected candidate genes during the first four
WAP. The data will serve as a basis for further bioinformatics and PCR
analysis of the genes of interest.
Our preliminary results suggest differential expression patterns among the
cultivars and pollination treatments, as well as among different
developmental stages.
Figure 24 - Heat map of EvaGreen Ct values of pollinated and non-pollinated
flowers of 'Barhee' (left quadrant) and 'Medjoul' (right quadrant) during the first
four weeks of fruit development, obtained using the Fluidigm software. Heat map
represents a total of 72 cDNA samples (listed horizontally), and gene names
(listed vertically). Bright yellow colour (lower Ct value) indicates high expression,
whereas purple and blue (high Ct value) indicate low expression.
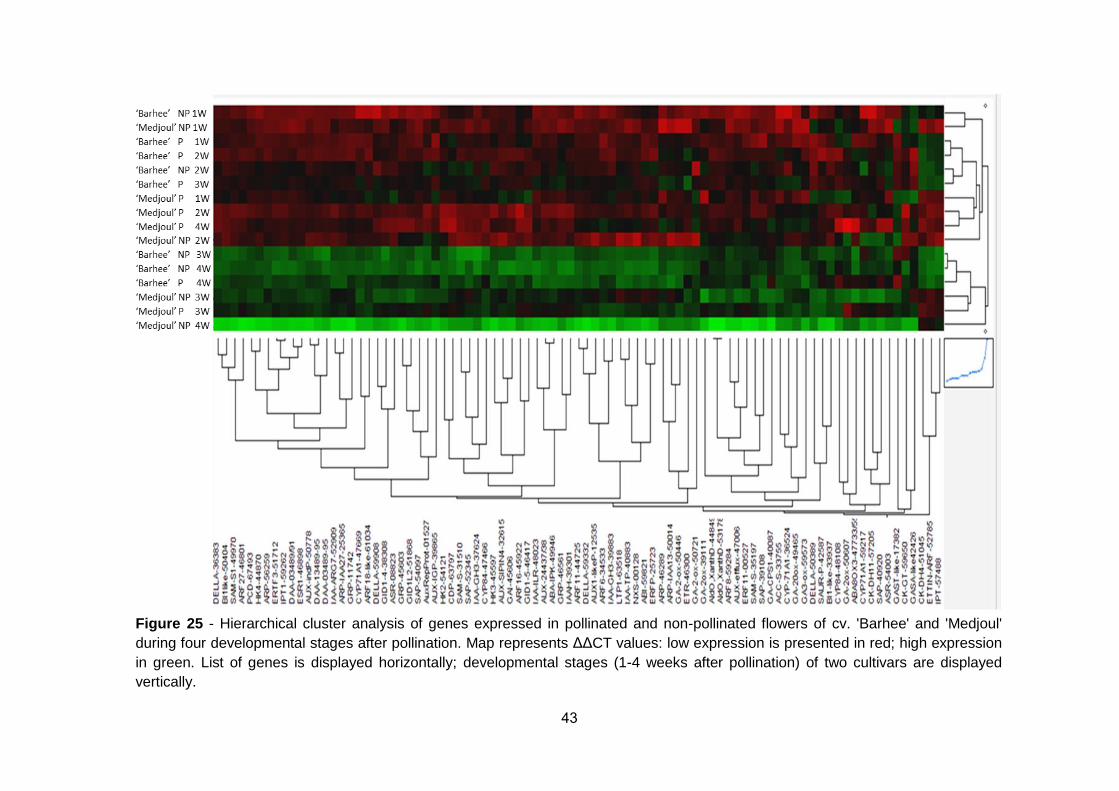
43
Figure 25 - Hierarchical cluster analysis of genes expressed in pollinated and non-pollinated flowers of cv. 'Barhee' and 'Medjoul'
during four developmental stages after pollination. Map represents ΔΔCT values: low expression is presented in red; high expression
in green. List of genes is displayed horizontally; developmental stages (1-4 weeks after pollination) of two cultivars are displayed
vertically.

44
Differential expression with respect to pollination. Four weeks after pollination
(4 WAP), abundance of transcripts of most of the selected categories was
recorded in non-pollinated flowers of 'Medjoul', as opposed to the pollinated
samples. At this time point, with respect to pollination, higher gene
expression was reported in 'Medjoul', in comparison to 'Barhee', e.g.,
transcript for GA-2 oxidase and abscisic acid stress ripening protein
homologue (ASR2). The examples of differential gene expression with
respect to pollination are presented in Table 11.
Table 11 – Differential expression of selected genes between non-pollinated and
pollinated flowers of 'Medjoul', four weeks after pollination.
Gene group Specific transcript Higher expression
Gib
be
relli
ns
DELLA proteins
In non-pollinated samples GA-2 oxidase
Gibberellin insensitive dwarf 1
GA repressed protein
Au
xin
s
auxin repressed protein – ARP
In non-pollinated samples
Auxin response factor - ARF (8;11;16;18-like;27)
auxin transport protein
auxin independent growth promoter
Cyto
kin
ins
CK DH 11
In non-pollinated samples
IPT9
CYP84A
CYP71A1
CK glucosyl-transferase 1
AB
A
ABA 8 hydroxylase In non-pollinated samples
ABA stress ripening protein
Differences in the expression of genes involved in AUX, GA and ABA
metabolism were recorded between pollinated and non-pollinated flowers of
'Medjoul' already at 1 WAP. Later, at four WAP in 'Medjoul', we observed a
trend of over-representation of GA transcripts in pollinated flowers,
associated with upregulation of GID1 (Figure 26 A). In 'Barhee', main
differences were observed in GA transcript levels, such as in DELLA
transcript.

45
Cultivar-specific gene expression was observed between 'Barhee' and
'Medjoul' already in the first WAP, (Figure 25, e.g. transcripts of IPT, ARF
and CK DH4). As an example, studied cultivars significantly varied in the
expression levels of gibberellin stimulated transcript like (GAST-like, Figure
26 B), IAA-amido synthetase (Figure 26 C) and cytokinin-related iso-
penthenyl transferase 9 transcript (Figure 26 D), showing variations both
regarding cultivars and pollination.
Expression dynamics during early fruit development. Our preliminary results
suggest that most of the selected genes showed the increasing trend
throughout the studied developmental stages, and expression was higher in
the fourth (latest) developmental stage as compared to the first three stages
in the two cultivars (e.g. ACC-synthase). In general, at four weeks after
anthesis, higher expression of most candidate genes was observed in non-
pollinated flowers as compared to pollinated ones (Figure 24). However,
some genes have high initial expression (CK GT) that declines at two and
three WAP, and show upregulation at four WAP in both cultivars, regardless
of pollination treatment.
Figure 26 Differential expressions of selected genes in 'Medjoul' and 'Barhee' during
four weeks of early fruit development. Pollinated and non-pollinated flowers were
analyzed in both cultivars. A) gibberellin insensitive dwarf 1 (GID1), B) gibberellin
associated transcript-like (GAST-like), C) IAA-amido synthetase and D) isopenthenyl
transferase 9 (IPT9).
-11
-8
-5
-2
1
4
0 1 2 3 4
Re
lati
ve e
xpre
ssio
n
Weeks after pollination
A AB
A
A A
ABC
BC BCC
CDCD
CDCD
D
A A
B
-3
-1
1
3
5
0 1 2 3 4
Re
lati
ve e
xpre
ssio
n
Weeks after pollination
AA
AB
AB
ABABAB
AB
AB
ABAB
ABAB
AB
AB
B
A
A
A
-11
-7
-3
1
5
9
0 1 2 3 4
Re
lati
ve e
xpre
ssio
n
Weeks after pollination
AA
AB
CDCDE
CDE CDEFDEF EF
EFF
AA
BC
A
CD
C
-2
0
2
4
6
8
10
0 1 2 3 4
Re
lati
ve e
xpre
ssio
n
Weeks after pollination
AB ABABCD
ABCDEBCDEF CDEFGDEFG
DEFG EFG
FG
FGFG
GG
AA
ABC
D

46
5. DISCUSSION
5.1. Morpho-anatomical traits of reproductive system and fruit
set in 'Medjoul' and 'Barhee'
The three-carpellate flower is the most common developmental pattern in
Arecaceae (Uhl and Moore, 1971). Similar to other common cultivars of date
palm, such as 'Deglet Noor' (Long, 1943), or 'Lulu' (Awad, 2010), at anthesis,
flowers of the two studied cultivars 'Medjoul' and 'Barhee' comprise three
equally developed, separated carpels, of which only one develops into a
single fruit. However, when pollination is prevented, 'Medjoul' and 'Barhee'
differed in number of carpels developing into fruit, as well as in rate of carpel
development, carpel size, and in fruit shedding ratio. Similar to the date palm,
examples of three-carpelate gynoecia were described in other palm species -
Corypha umbraculifera, Sabal palmetto (Rudall, 2011), as well as Elaeis
guineensis (Hartley, 1988). On the other hand, in the Coryphoid palm Thrinax
excels, gynoecium comprises a large single carpel, while four-carpelate
gynoecia exist in Latania palm. Furthermore, some palm species possess
larger multicarpelate gynoecia e.g. Orbignya speciosa, usually with 5-6
carpels, or Phytelephas macrocarpa (Tagua) comprising 7-10 carpels (Uhl
and Moore, 1971).
In general, fertilization patterns in dates resemble those of olive (Olea
europea), in which, only one ovule of the two-carpelate gynoecium bears
normal single-seeded fruit following carpel degeneration (King, 1938). Similar
to 'Medjoul' and 'Barhee', in most palms, the fruit is developing from a single
carpel, and the process of degeneration of two of the three additional carpels
is common. However, variation occurs, for instance, Phytelephas sp. usually
develops infructescences (the ensemble of multiple fruits derived from the
ovaries of an inflorescence), each containing numerous fruits (Bernal and
Galeano, 1993).
Effect of pollination and pollen quality on ovary and fruit development have
been reported in numerous plant species. For example, muskmelon ovaries
doubled in size within 48 h after pollination, similar processes were observed
in Brodiaea and carnation (O'Neill, 1997). In both studied palm cultivars

47
normal seed-bearing fruits develop upon successful pollination. In 'Barhee',
fruitlet formation was slightly faster. Following pollination, the morphological
traits of the developing fruits appear to be similar to those of other date
cultivars, such as 'Deglet Noor', 'Khadrawy' and 'Derry' (Haas and Bliss,
1935; Long, 1943; Reuveni, 1986;). On the other hand, two developmental
patterns were observed in parthenocarpic fruit formation. When pollination is
prevented, the non-pollinated flowers of 'Medjoul' form singlet partenocapric
fruits with high ratio of fruit shedding. In comparison, 'Barhee' produced
mainly triplets, and fruit shedding is relatively low.
Zaid and Al-Kaabi (2007) reported that ‘Barhee’ trees originated from tissue
culture were more susceptible to pollination failure and they produced more
abnormal fruits than ‘Medjool’. These in vitro generated somaclonal variants
of 'Barhee', 'Hallas' and other date palm cultivars, tend to form triplet
parthenocarpic fruitlets. Sometimes, multicarpelate fruit formation in dates
was observed (Al-Wasel, 2001; Awad, 2007, Cohen et al., 2004). The
additional carpels were probably developed by homeotic transformation from
the staminoid primordia. Triplet and multicarpelate parthenocarpic fruits are
also common in somaclonal variants obtained from in vitro propagation of oil
palm (Adam et a., 2005; Corley et al., 1986).
Using histological data, we defined several 'developmental checkpoints', i.e.,
critical stages of carpel and fruit development: (1) following pollination,
designation of a "leading carpel" forming the fruit, (2) degeneration vs.
continuous development of the two additional carpels, and (3)
(parthenocarpic) fruitlet shedding. These processes are similar in the
development of the normal, pollinated fruitlet, but differ between the cultivars
in developing parthenocarpic fruits.
For the first time, we provide evidence that, similar to 'Barhee' (Cohen et al.,
2004), pollen germination in ‘Medjoul’ occurrs on stigmas of all carpels,
suggesting that all three carpels are receptible and ready for fertilization
(Figure 17). These results suggest that the “decision making” checkpoint of
carpel abortion appears at post-fertilization stage and the first fertilized carpel
may inhibit the developments of two other carpels. For both cultivars, we
confirmed that processes of ovule degeneration and carpel reduction occur

48
earlier than we can record from the external morphological observations.
Under our experimental conditions, first histological evidence for ovule
degeneration in pollinaned 'Barhee' was recorded at 14 DAP, following by
pollinated 'Medjoul', in which similar developmental changes were observed
only after 21 DAP.
In another palm species Geonoma interrupta (Arecaceae) one of the three
carpels is completely developed and dominant already at anthesis, before
pollination, and fruit will be formed only from this carpel (pseudomonomerous
gynoecium) (Stauffer et al., 2002). The gynoecium development of G.
interrupta was divided into four stages. In stage II, the sterile carpels already
develop unequally, and in stage III the fertile carpel overtops the sterile ones
and is the most conspicuous organ of the gynoecium (Stauffer et al., 2002).
In contrary, in date palm, histological analysis as well as in situ expression of
flower development genes did not detect any differences in the three
developing carpels (Daher et al., 2010).
Pollination of 'Medjoul' flowers significantly accelerates carpel degeneration,
as compared to non-pollinated flowers (Table 4). Carpels of pollinated
flowers (only the one developing into fruit) were significantly larger in size, as
compared to the non-pollinated flowers, already at 27 DAP (Table 2 and 3).
We observed that in date palm, pollination provides a major cue for 'normal'
fruit development, acting as the switch for carpel degeneration and single
seed-bearing fruit development. When impaired, this process results in
parthenocarpic fruit development.
5.2. Temperature affects pollen germination, fertilization and
fruit-set processes
Temperature ranges and optima for pollen germination are known to vary
among species (Farlow et al., 1979; Kuo et al., 1981; Pearson, 1932). Pacini
et al. (1997) reported that pollen of the Meditteranean dwarf fan palm
Chamaerops humilis, remained viable for an exceptionally long time as
compared to Festuca arundinacea, Mercurialis annua, Acanthus mollis,
Cucurbita pepo and Spartium junceum. It was also suggested that pollen of

49
this palm, like that of other palms, is resistant to thermal (Al-Helal et al.,
1988) and water stresses (Bassani et al., 1994).
In our experiments, highest pollen germination on stigmas of 'Medjoul' in vitro
was observed at 30°C, whereas highest pollen tube elongation rate in a
solution in vitro was observed at 25°C, thus suggesting 25-30°C as optimal
temperature range (Figure 14 and 18). These results are in agreement with
Reuveni et al. (1986) and Bernstein (2004) that show fastest germination and
pollen tube elongation in styles at 25 and 28°C. In other species, optimum
temperatures for in vitro pollen germination varied from 28°C in cotton (Burke
et al 2004), to 23°C in peach (Weinbaum et al., 1984) and 22°C in
Arabidopsis (Boavida et al., 2007). In contrast, in almond, highest
germination was reported at 16°C (Weinbaum et al, 1984). Since date is
highly adaptive to high temperatures and can stand temperatures up to 50°C,
pollination and tube elongation require higher temperatures. Following to
pollination and during fruit ripening optimal reported temperature ranges from
21°C to 27°C (Zaid and De Wet, 2002b).
While optima of pollen germination lays between 25 and 30°C, general range
of suitable temperatures in orchard are much wider. Thus, even at 15˚C
moderate germination of ~ 50 % both in vitro and on date stigmas was
observed. However, pollen tube elongation and penetration to the upper part
of the carpel was much slower at 15˚C, as compared to higher temperatures,
making further germination into the carpel and ovule and fertilization
uncertain. Similarly, in avocado pollen tubes failed to reach the ovary at low
temperatures (17/12°C) (Sedgley, 1977). Under our experimental conditions,
in vitro assay of isolated spikelets was interrupted by early flower
senescence, and we could not follow fertilization and fruit setting. However,
the increase in parthenocarpic fruitlets in lower temperature in planta
experiments suggest reduced fertilization rate.
Although no morphological damage was observed within the first days after
pollen germination, further in vitro growth of pollen tubes on stigmas was
restricted and fertilization did not occur under any temperature regime. In
vitro cultured 'Medjuol' flowers wilt relatively fast either due to displacing from
the whole plant and shipment to the laboratory, or due to short "vase life" and

51
intense senescence and contamination processes. In a similar experiment,
performed with isolated spikelets of 'Barhee', pollen tube elongation was
detected within the carpel and up to the ovule within 3-7 days after pollination
(Cohen et al., 2004). Therefore we propose that the 'Medjoul' flowers under
in vitro conditions possess particularly short "vase life".
Beale and Johnson (2013) report that cross talk between the male and
female gametophyte is essential for early pollen tube growth and guidance in
several species, both monocots and dicots. We argue that under our
experimental conditions in vitro the interaction between the male and female
counterpart can be disturbed, in addition to pollen germination per se. The
reason for such “broken” interaction might be the hormonal imbalance in cut
female spikelets through overall senescence. Therefore, pollen tube
attractants secreted by the female cells, as well as the cross talk may had
been negatively affected.
Fast flower senescence might be also caused by ethylene emission,
hormonal imbalance or other factors. In spite of our efforts to extend in vitro
"vase life” of cut spikelets by using fungicide (Marpan) and anti-ethylene
compounds (STS, 1-MCP), flower senescence was faster than fruitlet
development, and even if pollination had been successful, we would not have
been able to follow the process of fertilization and fruitlet development.
In our in planta experiments pollen germination occurred under all
temperature regimes, but germination kinetics was clearly affected by
temperature range. High temperatures increased pollen germination and tube
growth rate in the stigma, reducing the time needed to reach the ovule at the
base of the carpel. Similar evidence of the effect of increasing temperature
on better pollen tube growth was reported in a range of herbaceous and
woody species (Hedhly, 2005). Relatively low temperatures (20/8°C) retarded
pollen germination, which was not detected at 16 hours after pollination, and
delayed at 3 DAP, as compared with higher temperatures. Only after 7 days,
pollen germination was observed at cool temperatures at the same
percentage as in other temperature treatments.
Even though we showed that pollen germination per se was not damaged
under cool temperatures, the major limiting factor for successful fertilization

51
can be stigma and ovule receptivity. For example, in sweet cherry, Hedhly et
al. (2003) reported that an increase in the average temperature of as little as
2.8°C was enough to negatively affect stigma receptivity. In addition, as high
temperatures accelerate and low temperatures retard pollen tube growth
rate, it would be expected that fertilization and, hence, fruit set would be
enhanced by high temperatures and reduced by low temperatures during
bloom (Sanzol, 2000). However, this is not always the case. Although high
temperatures during flowering accelerate pollen tube growth, they also
enhance maturation and early senescences and degeneration of the stigma
(Egea et al., 1991; Burgos et al., 1991) and ovule (Stosser and Anvari, 1982;
Postweiler et al., 1985; Cerovic and Ruzic, 1992), and, therefore, fertilization
will not succeed. In date palm, the length of the receptivity period of the
pistillate flowers varies between cultivars, and is usually up to 8 or 10 days
(Albert, 1930; Pereau- le Roy, 1958). ‘Medjoul’ flowers, however, have a
rather long flower receptability reaching up to 14 days (Bernstein, 2004).
Beyond these limits, fertilization fails and the percentage of parthenocarpic
fruits increases to 40 % (Zaid and De Wet, 2002). Moreover, in some
cultivars, such as 'Deglet Noor', female flowers do not become receptive for
possibly 7 days or more after the spathe cracks (Ream and Furr, 1969).
Taken together, fruit-set in dates is rather complex and sensitive to the
internal and external conditions. Therefore, as in many species, the main
question with respect to fruit-set is whether the equilibrium between pollen
tube growth rate and ovule receptivity/degeneration is reached at the given
temperature (Sanzol, 2000).
Since the temperature conditions in the "modular phytotoron" units were not
constant but changed in a sinusoidal pattern during the day (Figure 16), we
argue that in the lower temperature treatments, active pollen growth had
probably occurred mainly at the hours of highest temperature of the day. We
suggest that under lower temperatures a considerable fraction of the pollen
tubes did not reach the ovule, thus preventing efficient fertilization. The
increased occurrence of parthenocarpic fruits under lower temperatures
(20/8°C) implies that the reduced elongation rate of pollen tubes eliminated
efficient fertilization. It should be noted that in the experiment, large amount

52
of pollen was provided. Therefore, fertilization should lead to development of
a normal fruit, as fertilization of even a single of the three ovules is sufficient
for generation of a normal fruit. We speculate that if pollen was a limiting
factor, much larger variations in normal fruit set versus parthenocarpic fruit
would have been detected. In addition, our results regarding parthenocarpy
are in accordance with findings in mango, which, under low temperatures
(20/10°C), significantly increased the percentage of stenospermocarpic fruits
(Sukhvibul et al., 2000). This fact might also explain reduced fruit setting and
yields of dates occurring in commercial plantations under cooler conditions
during early spring (Y. Cohen, personal observations).
In planta experiments revealed differential effect of temperature regimes
during pollination and early fruit development on the levels of normal fruit
setting: higher temperatures significantly enhanced the formation of normal
seed-bearing fruits. At the same time, ratio of parthenocarpic fruits during
two consecutive seasons was significantly higher under cool temparatures,
whereas under normal conditions (non-induced conditions) or under
moderate temperatures, trees had much lower parthenocarpic fruit
percentage. This may suggest that even though conditions below 20˚C
reduce fertilization efficiency and favour parthenocarpic fruit development,
high temperatures during the middle of the day might compensate for the
lower extreme and prevent parthenocarpy. Similarly, in pear, parthenocarpic
fruit development is enhanced by frost damage (Lewis 1942, Modlibowska
1945). A tendency towards small parthenocarpic fruits in tomato was
observed under low (14 °C) temperature regimes (Adams, 2001; Nuez et al.,
1986; Preil, 1973; Preil and Reimann-Philipp, 1969). Alternatively,
parthenocarpy is enhanced by high temperatures in pommelo (Citrus
grandis) (Susanto, 1990), as well as in wheat (at 36/31˚C) (Tashiro et al.,
1990) and rice (39/34°C) (Tashiro et al., 1991). In tomato, however,
parthenocarpic fruits develop under optimal (28/22°C), as well as high
temperature treatments of 32/26°C (Sato, 2001). It seems that partenocarpy
is enhanced by suboptimal (too low or too high) temperatures and might also
be associated with additional environmental stresses.

53
Future research on parthenocarpic fruit development in date palm under
various temperature, including shifting and combination of several
temperature regimes (e.g., low temperature applied for pollen tube growth
and fertilization followed by high temperatures during early fruit setting) will
facilitate the optimization of temperature regime for each developmental
stage.
The histological analysis proved that the general developmental pattern of
carpel and fruit development was not altered under different temperature
treatments. However, overall rate of development and “decision making”
checkpoint of carpel degeneration were greatly affected, indicating that cell
division and elongation are promoted by higher temperatures. Furthermore,
temperature treatments in planta affected colour, size, and weight of the
fruitlets as well as overall rate of their development: fruitlets were much
bigger under warmer condition both 5 and 10 weeks following pollination,
whereas a distinct difference in colour was observed among the treatments 5
WAP. On the other hand, different treatments did not change the spikelet
growth, most likely because elongation of the spikelet within the spathe was
already terminated, and additional elongation occurs only at the very base of
the spikelet and fruit bunch.
Although in planta studying of whole trees in controlled temperature units is
beneficial, it is rather challenging, since it is not possible to place the entire
adult tree to standard rooms with controlled environments. Therefore,
alternative techniques are required. For example, in evaluating effects of
temperature regimes on pollination and stigma receptivity in peach, Hedhly et
al. (2005) covered trees with polyethylene cages with regulated temperature
regime. In addition, studying particular biological processes in large fruit trees
often relies on the use of isolated plant organs/parts in vitro (Hedhly, 2005;
Sukhvibul et al., 2000; Weinbaum et al., 1984). In the presented research,
two systems were developed for the evaluation of the fertilization process in
palms. In vitro assays on isolated inflorescences and in planta experiments in
"modular phytotrones" complemented each other. Each employed system
has specific advantages, as well as technical and biological limitations. The
in vitro approach facilitates a direct study of the inflorescences at constant

54
temperatures under artificial conditions. However, major limitations in this
approach are the short "vase life" of cut inflorescences, contamination,
absence of leaves and hence disturbance of hormonal and environmental
stimuli, as well as fast flower senescence, probably due to ethylene release
in cut inflorescences. Nevertheless, this approach allows focusing on pollen
germination, as well as on individual flowers and careful examination the
stigmas and carpels in vitro.
Our in planta study in controlled temperature units, actually presents a
beneficial “modular phytotron” approach. In this case, the inflorescence
remains the integral part of the whole tree, and its hormonal and nutritional
balance is intact. Moreover, this approach allowed us to study flowers and
fruitlets throughout development, much longer period than any in vitro assay
could allow for. However, in these experiments, environmental conditions
were modified only in the inflorescences, while temperature effects on the
other plant organs were not modified. We are aware of the limitations of this
approach: by enclosing inflorescences in environmental controlled units, the
tree trunk, leaves and root systems are being exposed to outdoor
temperatures and are not controlled. Hence, one of the drawbacks is the
disregard of hormonal and other environmental signals such as light and
humidity that affect the plant and can be transported from organ to organ.
5.3. Molecular analysis of genes involved in hormonal regulation
during early fruit development of cultivars 'Medjoul' and
'Barhee'
Plant hormones play a prominent role in fruit development of plants,
especially in reproductive traits and fruit setting (Nitsch 1970, Ozga et al.,
2003, De Jong et al., 2009, Perez-Amador et al., 2009). They are essential
for successful completion of each developmental stage and progression of
the developing fruit into the next stage. The abundance of certain hormones
at specific stages of fruit development indicates a possible role for these
hormones during that developmental stage (Srivastava and Handa, 2005). In
tomato, as in many other species, levels of AUX- and GA- genes are
upregulated in ovules after pollination, resulting in the activation of AUX and

55
GA response genes, which, in turn, will trigger fruit growth and development
by regulating cell division and cell expansion (De Jong et al., 2009). GAs are
effective in inducing parthenocarpy in apples (Hayashi et al., 1968), pears ,
stone fruits, including grape (Nagata, 1982), while AUX or CKs induce
parthenocarpy in figs, kiwifruit and strawberry (Kato et al., 2000). Moreover,
during fruit development of figs, following decline of endogenous levels of
CK, GA and AUX, fruit growth predominantly occurs through cell
enlargement; in addition, it has been demonstrated that ethylene promotes
expansion of cells (Crane, 1969, Srivastava, 2005). Taken together,
hormonal balance vary between different stages of fruit development, while
the same hormone can play different functions at different stages (Ozga and
Reinecke, 2003),
In the presented research, a large number of genes of date palm, associated
with hormonal regulation of the reproduction process, were analyzed, for the
first time, with respect to early stages of fruit development. The preformed
pioneer molecular analysis requires further bioinformatic analyses and PCR
validation. However, the initial results already provide an insight into some of
the regulatory hormonal processes occurring during early fruit development
in date palms.
As ripening of dates requires about 150 days, we used previously defined
seven developmental phases (Al-Mssallem et al., 2013), subsequently
merged into three stages (Zhang, 2012). We focused on early fruit
development from anthesis to 45 DAP and collected plant samples for
histological and molecular data in weekly intervals. It was shown that at early
stages up to 30 DAP young fruits are hard and green and are characterized
by high rate of cell multiplication. Later than 30 DAP, cell expansion
increases, and accumulation of starch begins in the fruit cells (Zhang, 2012).
Earliest stage of fruit development (0-15 DAP) is characterized by nine
groups of genes that are expressed higher than those of other fruiting stages.
These groups are involved in the molecular function of binding, catalytic
activity, structural molecule activity, nucleic acid binding, transcription factor
activity, transcription regulator activity, transporter activity, enzyme regulator
activity, electron carrier activity and antioxidant activity. In biological process

56
category, both cellular processes and metabolic processes such as
replication and repair, translation, and cell growth and death, etc. were
expressed at the highest level in the earliest stage (Zhang et al., 2012).
Similarly, we observed upregulation of genes associated with CK
biosynthesis in earliest stages of fruit set and development in both cultivars.
This fact may indicate high ratio of cell division, as shown by Zhang et al.
(2012) in the F1 stage of date fruit development, as well as by Janssen et al.
(2008) in apple. On the other hand, some genes coding for CK showed
cultivar-specific gene expression, suggesting that biosynthesis of this
hormone as well as the CK-related mitotic activity are regulated by a number
of genes. Moreover, timing of CK expression might vary between cultivars, as
observed in the histological results.
Assuming high cytokinin activity in early fruit development, and expecting
differential response to pollination of auxin and GA genes, we acquired and
validated data on numerous genes involved in metabolism and signaling of
these hormones during early fruit development. Indeed, the differential
expression of auxin-, GA- and cytokinin-related genes with respect to
pollination was found in both date palm cultivars. In addition, at 4 WAP
upregulation of GA transcripts through upregulation of GID1 was found in
pollinated flowers of 'Medjoul'. Similarly, the increase of GA levels in
response to pollination was reported in citrus (Ben-Cheikh et al. 1997) and
Arabidopsis (Gallego-Giraldo et al., 2014).
Our preliminary results suggest that main groups of growth regulators might
be involved in pollination, fertilization, fruit set and fruit development.
Pollination and fertilization may be regulated by the cross-talk between AUX
and GA. Similar to pea (Ozga and Reinecke, 2003) and tomato (De Jong et
al., 2009), increasing AUX levels promote biosynthesis of GA, The initial fruit
growth is associated with high mitotic activity, regulated by CK levels, while
the following stages are characterized by an increase in ET, which may be
involved in cell elongation and nutrient mobilization. Moreover, differential
expression of GA- and AUX- related genes in pollinated and non-pollinated
flowers might specify cultivar-specific developmental differences.

57
Future research can clarify differences in hormonal regulation of the
reproductive processes in date palm. Morpho-physiological, combined with
the transcriptome analysis, PCR validation of the selected genes involved in
hormonal regulation, and biochemical analysis of plant hormones, will
increase our understanding of the complex process of fruit development in
dates. Taken together, this new information will be used for the optimization
of fruit production of this interesting and useful crop.
The presented research provides characterization of pollination, fertilization
and early fruit development in date palm. In-depth histological analysis
complemented with study on effects of temperature regime on reproductive
process increased our understanding of fruit developmental physiology and
its sensitivity to stress. Cultivar differences with respect to their reaction to
different environmental conditions may serve as basis for improving protocols
for pollination, fruit set and thinning.

58
References
Abd Alaal, A. F., Al Salih, K. K., Shabana, H., and Al Salihy, G. J. (1983). Production
of seedless dates by application of growth regulators. Proceedings of the first
symposium on the date palm in Saudi Arabia, 276-282.
Adam, H., Jouannic, S., Escoute, J., Duval, Y., Verdeil, J.-L. and Tregear, J.W.
(2005). Reproductive developmental complexity in the African oil palm (Elaeis
guineensis, Arecaceae). American Journal of Botany, 92:1836-1852.
Adams, S.R., Cockshull, K.E., Cave, C.R.J. (2001). Effect of temperature on the
growth and development of tomato fruits. Annals of Botany, 88, 869–877.
Al-Dous, E.K. (2011). De novo genome sequencing and comparative genomics of
date palm (Phoenix dactlyfera). Nature Biotechnology 29 (6): 521-7.
Al-Helal, A.A., Basalah, M.O., and Mohammed, S. (1988). Effect of storage and
temperature on pollen germination and rate of pollen tube elongation of date
palm (Phoenix dactilifera L.). Phyton 48: 119-122.
Al-Kaabi, H.H., Zaid, A. and Ainsworth, C. (2007). Plant-off-types in tissue culture-
derived date palm (Phoenix dactylifera L.) plants. Acta Horticulturae 736: 267-
281.
Al-Mssallem, I.S. (2013). Genome sequence of date palm Phoenix dactylifera L.
Nature Communications 4:2274.
Al-Wasel, A.S. (2001). Field performance of somaclonal variants of tissue culture-
derived date palm (Phoenix dactylifera L.). Plant Tissue Culture, 11:97-105.
Awad, M.A. (2007). Fruit set failure in tissue culture-derived date palm trees
(Phoenix dactylifera L.) cv. 'Nabt Saif' as affected by pollinator type and
pollination density. Acta Horticulturae 736, 441-448.
Awad, M. A. (2010). Pollination of date palm (Phoenix dactylifera L.) with pollen
grains water suspension. Acta Horticulturae 882, 337-344.
Barrow, S.C. (1998). A monograph of Phoenix L. (Palmae: Coryphoideae). Kew Bull.
53:513-575.
Bassani, M., Pacini, E., and Franchi, G.G. (1994). Humidity stress responses in
pollen of anemophilous and entomophilous species. Grana 33: 146-150.
Beale, K. M. and Johnson M. A. (2013). Speed dating, rejection, and finding the
perfect mate: advice from flowering plants. Current Opinion in Plant Biology 16:1-
8.
Ben-Cheikh, W., Perez-Botella, J., Tadeo, F. R., Talon, M. and Primo-Millo, E.
(1997). Pollination increases gibberellin levels in developing ovaries of seeded
varieties of citrus. Plant Physiology 114, 557–564.
Bernal, R. G. and Galeano, G. (1993). Tagua. In: Clay J. W. and Clement C. R. (ed.)
Selected species and strategies to enhance income generation from Amazonian
forests. FO: Misc/93/6 Working paper. Food and Agriculture Organization of the
United nations, Rome, May 1993.

59
Bernestein, Z. (2004). The Date Palm. Israeli Fruit Board (In Hebrew), Tel Aviv.
Boavida, L. C. and McCormick, S. (2007). Temperature as determinant factor for
increased and reproducible in vitro pollen germination in Arabidopsis thaliana.
The Plant Journal 52:570-582.
Bourgis, F., Kilaru, A., Cao, X., Ngando-Ebongue, G., Drira, N., Ohlrogge, J.B. and
Arondel, V., (2011). Comparative transcriptome and metabolite analysis of oil
palm and date palm mesocarp that differ dramatically in carbon partitioning.
10.1073/pnas.1106502108.
Burgos, L., Egea, J., and Dicenta, F., (1991). Effective pollination period in apricot
(Prunus armeniaca L.) cultivars. Annals of Applied Biology, 119, 533-539.
Burke, J. J., Velten, J. and Oliver, M. J. (2004). In vitro analysis of cotton pollen
germination. Agronomy journal 96 (2): 359-368.
Carimi, F., Zottini, M., Formentin, E., Terzi, M. and Lo Schiavo, F. (2003).
Cytokinins: new apoptotic inducers in plants, Planta 216:413-421.
Cerovic, R., and Ruzic R., (1992). Senescence of ovules at different temperatures
and their effect on the behavior of pollen tubes in sour cherry. Scientia
Horticulturae, 51, 321-327.
Chao, C. T. and Krueger, R. R. (2007). The Date Palm (Phoenix dactylifera L.):
Overview of Biology, Uses and Cultivation. Horticultural Science 42 (5), 1077-
1082.
Cohen, Y., Korchinsky, R. and Tripler, E. (2004). Flower abnormalities cause
abnormal fruit setting in tissue culture-propagated date palm (Phoenix dactylifera
L.). Journal of Horticultural Science and Biotechnology 79 (6),1007-1013.
Cohen, Y. and Glasner, B., (2014). Date palm status and perspective in Israel. In:
J.M. Al-Khayri, Date palm genetic resources, cultivar assessment, cultivation
practices and novel products. pp.1-35, Springer Science+Bussines Media. In
Press.
Corley, R.H.V., Lee, C.H., Law, I.M. and Wong, C.Y. (1986). Abnormal flower
development in oil palm clones. Planter, 62 (723): 233-240.
Crane, J. C. (1969). The role of hormones in fruit set and development. HortScience
4 (2): 108-111.
Daher, A., Adam, H., Chabrillange, N., Collin, M., Mohamed, N., Tregear, J.W. and
Aberlenc-Bertossi, F. (2010). Cell cycle arrest characterizes the transition from a
bisexual floral bud to a unisexual flower in Phoenix dactylifera. Annals of Botany
106:255.
De Jong, M., Mariani, C. and Vriezen, W. H. (2009). The role of auxin and
gibberellin in tomato fruit set. Journal of Experimental Botany 60 (5):1523-1532.
Dorcey, E., Urbez C., Blazquez, M.A., Carbonell, J. and Perez-Amador, M.A. (2009).
Fertilization-dependent auxin response in ovules triggers fruit development
through the modulation of gibberellin metabolism in Arabidopsis. The Plant
Journal, 58(2):318-332.

61
Dransfield, J., Uhl, N.W., Asmussen, C.B., Baker, W.J., Harley, M.M. and Lewis,
C.E. (2008). Genera palmarum: the evolution and classification of palms. Kew
Publishing, Kew, UK.
Eckardt, N.A. (2007). GA perception and signal transduction: molecular interactions
of the GA receptor GID1 with GA and the DELLA protein SLR1 in rice. Plant Cell,
19:2095-2097.
Egea, J., and Burgos, L. (1992). Effective pollination period as related to stigma
receptivity in apricot. Scientia Horticulturae 52, 77-83.
Fang, Y., Wu, H., Zhang, T., Yang, M., Yin, Y., Pan, L., Yu, X., Zhang, X., Hu, S., Al-
Mssallem, I.S. and Yu, J. (2012). A complete sequence and transcriptomic
analyses of Date Palm (Phoenix dactylifera L.) mitochondrial genome. PloS ONE
7(5): e37164.
Farlow, P. J., Dyth, D.E. and Kruger, N. S. (1979). Effect of temperature on seed set
and in vitro pollen germination in French beans (Phaseolus vulgaris), Australian
Journal of Experimental Agriculture and Animan Husbandry. 19: 725-731.
Frank, M. and Schmulling, T. (1999). Cytokinin cycles cells. Trends in Plant Science,
4:243-244.
Fuentes, S., Ljung, K., Sorefan, K., Alvey, E., Harberd, N. P. and Østergaard, L.
(2012). Fruit Growth in Arabidopsis Occurs via DELLA-Dependent and DELLA-
Independent Gibberellin Responses. The Plant Cell, 24 (10): 3982-3996.
Gallego-Giraldo, C., Hu, J., Urbez, C., Dolores Gomez, M., Sun, T. and Perez-
Amador, M. A. (2014). Role of the gibberellin receptors GID during fruit-set in
Arabidopsis. The Plant J79 (6): 1020-1032.
Gil, G. F., Martin, G. C. and Griggs, W. H. (1972). Fruit set and development in pear:
Extractable endogenous hormones in parthenocarpic and seeded fruit. Journal of
the American Society for Horticultural Science 97, 731-735.
Gillaspy, G., Ben-David, H. and Gruissem, W. (1993). Fruits: A developmental
perspective. Plant Cell 5 (10): 1439–1451.
Goetz, M., Hooper, L. C., Johnson, S. D., Carlyle, J., Rodrigues, M., Vivian-Smith,
A., and Koltunow, A. M. (2007). Expression of Aberrant forms of AUXIN
RESPONSE FACTOR8 stimulates parthenocarpy in Arabidopsis and tomato,
Plant Physiology 145: 351-366.
Gustafson, F. (1939b). The cause of natural parthenocarpy. American Journal of
Botany 26, 135-138.
Hardtke, C.S., Ckurshumova, W., Vidaurre, D.P., Singh, S.A., Stamatiou, G., Tiwari,
S.B., Hagen, G., Guilfoyle, T.J., and Berleth, T. (2004). Overlapping and non-
redundant functions of the Arabidopsis auxin response factors MONOPTEROS
and NONPHOTOTROPIC HYPOCOTYL 4. Development 131, 1089–1100.
Hartley, C. W. S. (1988). The oil palm. London: Longman.

61
Hass, A.R.C. and Bliss, D.E. (1935). Growth and composition of Deglet Noor dates
in relation to water injury. Journal of Agricultural Science published by the
California Agricultural Experiment station, Hilgardia 9 (6): 295-344.
Hayashi, F., Naito, R., Bukovac, M. J. and Sell, H. M. (1968). Occurrence of
Gibberellin GA3 in Parthenocarpic Apple Fruit. Plant Physiology 43:448-450.
Hedhly, A., Hormaza, J.I., and Herrero, M., (2003). The effect of temperature on
stigmatic receptivity in sweet cherry (Prunus avium L.). Plant Cell and
Environment, 26, 1673–1680.
Hedhly, A., Hormaza, J. I. and Herrero, M. (2005). The effect of temperature on
pollen germination, pollen tube growth and stigmatic receptivity in peach (Prunus
persica L. Batsch.). Plant Biology 7, 476-483.
Hedhly A., (2011). Sensitivity of flowering plant gametophytes to temperature
fluctuations. Environemntal and Experimental Botany, 74, 9-16.
http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor. Accessed
on January 8th, 2015.
Jain, S. M., Al-Khayri, J. M. and Johnson, D. V. (eds), (2011). Date palm
biotechnology. Springer Science+Business Media B.V. London, UK, pp. 320-327.
Janssen, B. J., Thodey, K., Schaffer, R. J., Alba, R., Balakrishnan, L., Bishop, R.,
Bowen, J. H., Crowhurst, R. N., Gleave, A. P., Ledger, S., McArtney, S.,
Pichler, F.B., Snowden, K.C. and Ward, S. (2008). Global gene expression
analysis of apple fruit development from the floral bud to ripe fruit. BMC Plant
Biology 8:16.
Jong, M., Mariani, C. and Vriezen, W. (2009). The role of auxin and gibberellin in
tomato fruit set, Journal of Experimental Botany, doi:10.1093/jxb/erp094.
Kato, K., Ohara, H., Takahashi, E. and Matsui H. (2000). Endogenous gibberellin-
induced parthenocarpy in grape berries. Acta Horticulturae 514: 69-74.
King, J.R., (1938). Morphological development of the fruit of the olive, in Hilgardia,
Journal of Agricultural Science 11 (8) 437-458.
Koshioka, M., Nishijima, T., Yamazaki, H., Nonaka, M. and Mander, L.N. (1994).
Analysis of gibberellins in growing fruits of Lycopersicon esculentum after
pollination or treatment with 4-chlorophenoxyacetic acid. Journal of Horticultural
Science 69:171–179.
Kuo, C. G., Peng J. S. and Tsay J. S. (1981). Effect of high temperature on pollen
grain germination, pollen tube growth and seed yield of Chinese cabbage.
HortScience 16: 67-68.
Lewis, D. (1942). The physiology of incompatibility in plants. I. The effect of
temperature. Proceedings of the Royal Society of London, Series B 131, 13-26.
Long, E. M. (1943). Developmental anatomy of the fruit of the 'Deglet Noor' Date.
Botanical Gazette, 104 (3): 426-436.

62
Luza, J. G., Polito, V. S. and Weinbaum, S. A. (1987). Pollen germination and tube
growth in two walnut (Juglans) species, American Journal of Botany 74 (12):
1989-1903.
Mapelli, S., Torti, G. and Soressi M.B.G. (1979). Effects of GA3 on flowering and
fruit-set in a mutant of tomato. Horticultural Science 14, 736-737.
Mapelii, S. and Lombardi, L. (1982). A comparative auxin and cytokinin study in
normal and to-2 mutant tomato plants. Plant and Cell Physiology 23, 751-757.
Marti, C., Orzaez, D., Ellul, P., Moreno, V., Carbonell, J. and Granell, A. (2007).
Silencing of DELLA induces facultative parthenocarpy in tomato fruits, The Plant
Journal 52, 865-876.
Modlibowska, I. (1945). Pollen tube growth and embryo sac development in apples
and pears. Journal of Pomology, 21, 57-89.
Molesini, B., Pandolfini, T, Rotino, G.L., Dani, V., and Spena, A. (2008), Aucsia
Gene Silencing Causes Parthenocarpic Fruit Development in Tomato. Plant
Physiology 149 (1):534-548.
Nagata, K., Kurihara, A. (1982). The varietal difference in the response of grape
cultivars to gibberellin application. Bulletin of fruit tree research station E, 4: 7-19.
Nitsch, J. P. (1952). Plant hormones in the development of fruits. The quarterly
Review of Biology 27 (1): 33-57.
Nixon, R. W., and Carpenter, J. B. (1978). Growing dates in the United States.
USDA Inform. Bull. 207. Washington, D.C., USA. pp. 3-5.
Nuez, F., Cuartero, J., Ferrando, C., Catala, M.S., and Costa, J. (1988). Genetic
model for the inheritance of the parthenocarpy in the tomato line '75/59'. Anales
de la Estacion Experimental de Aula Dei, 19 (1-2): 7-11.
O'Neill, S.D. (1997). Pollination regulation of flower development Annual Review of
Plant Physiology and Plant Molecular Biology 48:547-74.
Ozga, J. A. and Reinecke, D. M. (2003). Hormonal interaction in fruit development,
Journal of Plant Growth Regulation 22:73-81.
Pacini, E., Franchi, G. G., Lisci, M and Nepi, M. (1997). Pollen viability related to
type of pollination in six angiosperm species. Annals of Botany 80 (1): 83-97.
Pandolfini, T., Molesini, B., Rotino, G.L., Dani, V. and Spena A. (2009). Aucsia gene
silencing causes parthencarpic fruit development in tomato. Plant Physiology 149
(1): 534-548.
Pearson, O. H. (1932). Breeding plants of the cabbage group. California Agricutlural
Experiment Station Bulletin 532.
Postweiler, K., Stoesser, R., and Anvari S. F., (1985). The effect of different
temperatures on the viability of ovules in cherries. Scientia Horticulturae 25, 235-
239.
Preil, W., and Reimann-Philipp, R. (1969). Untersuchungen über die Einflüsse
verschiedener Umweltfaktoren auf die Funktionsfähigkeit der Pollen von Tomaten

63
(Lycopersicon esculentum Mill.) insbesondere solcher mit erblicher neigung zur
Parthenocarpie. Angewandte Botanik, 43: 175-193.
Preil, W. (1973). Zur Parthenocarpie bei Tomaten in Abhängigkeit vom
Temperaturverlauf. Angewandte Botanik 47, 135-140.
Reuveni, O. (1986). Date, CRC Handbook of Fruit Set and Development. Boca
Raton, FL: CRC Press,119-122.
Reuveni, O., Abu, S. and Golobovitz, S. (1986). Date palm pollen germination and
tube elongation on pistillate flowers cultured at different temperatures. Acta
Horticulturae 175:91-95.
Rezazadeh, R., Hassanzadeh, H., Hosseini, Y., Karami, Y. and Williams, R. (2013).
Influence of pollen source on fruit production of date palm (Phoenix dactylifera L.)
cv. 'Barhee' in humid coastal regions of southern Iran, Scientia Horticulturae
160:182-188.
Richards, D. E., King, K. E., Ait-ali, T. and Harberd, N. P. (2001). How gibberellin
regulates plant growth and development: a molecular genetic analysis of
gibberellin signaling. Annual Review of Plant Physioliogy and Plant Molecular
Bioliogy 52, 67-88.
Rudall, P. J., Ryder, R. A. and Baker, W. J. (2011). Comparative gynoecium
structure and multiple origins of apocarpy in coryphoid palms (Arecaceae).
International Journal of Plant Science 172 (5):674-690.
Ruzin, S. E., (1999). Plant microtechniques and microscopy. Oxford University
Press. pp. 57-116.
Saidi, M. N., Ladouce, N., Hadhri, R., Grima-Pettenati, J., Drira, N. and Gargouri-
Bouzid, R. (2010). Identification and characterization of differentially expressed
ESTs in date palm leaves affected by brittle leaf disease, Plant Science 179:325-
332.
Sanzol, J. and Herrero, M. (2001). The "effective pollination period" in fruit trees.
Scientia Horticultuae 90, 1-17.
Sato, S., Peet, M. M. and Gardner, R. G. (2000). Formation of parthenocarpic fruit,
undeveloped flowers and aborted flowers in tomato under moderately elevated
temperatures. Scientia Horticultuae 90: 243-254.
Sedgley, M. (1977). The effect of temperature on floral behavior pollen tube growth
and fruit set in the avocado. Journal of Horticultural Science 52: 135-141.
Sedgley, M. and Shuraki, Y. D. (1996). Fruit development of Pistacia vera
(Anacardiaceae) in relation to embryo abortion and abnormalities at maturity.
Australian Journal of Botany 44: 35-45.
Serrani, J.C., Fos, M., Atares, A. and Garcia-Martınez, J. L. (2007). Effect of
gibberellin and auxin on parthenocarpic fruit growth induction in the cv. Micro-tom
of tomato. Journal of Plant Growth Regulation 26:211–221.

64
Shaheen, M. A., Nasr, T. A. and Bacha, M. A. (1988). Effect of some plant growth
regulators on induction of seedless fruits in some date palm cultivars. Journal of
the College of Agriculture, King Saud University 10 (1): 129-138.
Spurgeon, S. L., Jones, C. J. and Ramakrishnan, R. (2008). High throughput gene
expression measurement with real time PCR in a microfluidic dynamic array.
PlosOne 3 (2) e1662.
Srivastava, A. and Handa, A. K. (2005). Hormonal regulation of tomato fruit
development: A molecular perspective. Journal of Plant Growth Regulation 24:
67-82.
Stauffer, F. W., Rutishauser R. and Endress, P. K (2002). Morphology and
development of the female flowers in Geonoma interrupta (Arecaceae). American
Journal of Botany 89 (2): 220-229.
Stoesser, R., and Anvari, S. F., (1982). On the senescence of ovules in cherries.
Scientia Horticulturae, 16, 29-38.
Sukhvibul, N., Hetherington, S. E., Whiley, A. W., Smith, M. K. and Vithanage, V.
(2000). Effect of temperature on pollen germination, pollen tube growth and seed
development in mango (Mangifera indica L.), Acta Horticulturae 509: 609-616.
Sundberg, E., and Østergaard L. (2009). Distinct and dynamic auxin activities during
reproductive development. Cold Spring Harbor Perspectives in Biology
Susanto, S. and Nakajima, Y. (1990). Effect of winter heating on flowering time,
fruiting and fruit development in pummelo grown in a plastic house. Journal of
Japanese Society for Horticultural Science, 59 (2): 245-253.
Swain S. M., and Koltunow A.M. (2006). Auxin and fruit initiation. In L. Taiz, E.
Zeiger, (eds), Plant Physiology, 4. Sinauer Associates, Inc., Sunderland, MA.
Tashiro, T. and Wardlaw I.F. (1990) The response to high temperature shock and
humidity changes prior to and during the early stages of grain development in
wheat. Australian Journal of Plant Physiology, 17(5): 551 – 561.
Tatematsu, K., Kumagai, S., Muto, H., Sato, A., Watahiki, M.K., Harper, R.M.,
Liscum, E., and Yamamoto, K.T. (2004). MASSUGU2 encodes Aux/IAA19, an
auxin-regulated protein that functions together with the transcriptional activator
NPH4/ARF7 to regulate differential growth responses of hypocotyl and formation
of lateral roots in Arabidopsis thaliana. Plant Cell 16, 379–393.
Torahi, A. and Arzani, K. (2010). Date palm (Phoenix dactylifera L.) fruit growth
pattern. Acta Horticultuae 864: 201-206.
Tyler, L., Thomas, S. G., Hu, J., Dill, A., Alonso, J. M., Ecker, J. R., and Sun, T.
(2004). DELLA proteins and gibberellin-regulated seed germination and floral
development in Arabidopsis, Plant Physiology 135:1008-1019.
Uhl, N. W. and Moore, H. E. (1971). The palm gynoecium. American Journal of
Botany, 58 (10): 945-992.

65
Ulmasov, T., Hagen, G., and Guilfoyle, T.J. (1999b). Activation and repression of
transcription by auxin response factors. Proceedings of the National Academy of
Sciences, USA 96, 5844–5849.
Van Huizen, R., Ozga, J.A. and Reinecke, D.M. (1997). Seed and hormonal
regulation of gibberellin 20-oxidase expression in pea pericarp. Plant Physiology,
115, 123–128.
Vivian-Smith, A., Luo, M., Chaudhury, A. and Koltunow, A. (2001). Fruit
development is actively restricted in the absence of fertilization in Arabidopsis.
Development, 128 (12): 2321-31.
Wang, H., Jones, B., Li, Z., Frasse, P., Delalande, C., Regad, F., Chaabouni, S.,
Latche, A., Pech, J.-C., and Bouzayen, M. (2005). The tomato Aux/IAA
transcription factor IAA9 is involved in fruit development and leaf morphogenesis.
Plant Cell 17, 2676–2692
Weinbaum, S. A., Parfitt, D. E. and Polito, V. S. (1984). Differential cold sensitivity of
pollen grain germination in two Prunus species, Euphytica 33: 419-426.
White, P. J. (2002). Recent advances in fruit development and ripening: an
overview. Journal of Experimental Botany, 53 (377):1995-2000.
Wrigley, G. (1995). Date palm, Phoenix dactylifera. In: Smart J. and Simmonds N.
W. (eds.), Evolution of crop plants, 2nd ed. Longman, London: pp. 399-403.
Yang, M., Zhang, X., Liu, G., Yin, Y., Chen, K., Yun, Q., Zhao, D., Al-Mssallem, I. S.
and Yu, J. (2012). The complete chloroplast genome sequence of date palm
(Phoenix dactylifera L.). PloS One 15:5 (9):e12762.
Yin, Y., Zhang X., Fang, Y., Pan, L., Sun, G., Xin, C., Ba Abdullah M.M., Yu, X., Hu,
S., Al-Mssallem, I.S., Yu, J. (2012). High-throughput sequencing-based gene
profiling on multi-staged fruit development of date palm (Phoenix dactylifera L.)
Plant Molecular Biology, 78:617-626.
Zaid, A. and De Wet, P. F. (2002). Date Palm Cultivation, Food and Agriculture
Organization, Plant Production and protection Paper no. 156,
http://www.fao.org/docrep/006/y4360e/y4360e00.htm (accessed October 5th,
2013).
Zaid, A. and De Wet, P. F. (2002a). Botanical and systematic description of the date
palm. In: A. Zaid (ed.), Date Palm Cultivation, Vol. 156. FAO, Rome. p. 29-44.
Zaid, A. and De Wet, P. F. (2002b). Climatic requirements of date palm. In: A. Zaid
(ed.), Date Palm Cultivation. FAO, Rome. p. 58-73.
Zaid, A. and Al-Kaabi, H. (2003). Plant-off types in tissue culture-derived date palm
(Phoenix dactylifera L.). Emirates Journal of Agricultural Science 15:17-35.
Zaid, A., and Al-Kaabi, H. (2007). Morphological abnormalities in tissue culture-
derived date palm (Phoenix dactylifera L.). Acta Horticultureae 736, 329-335.
Zhang, G., Pan, L., Yin, Y., Liu, W., Huang, D., Zhang, T., Wang, L., Xin, C., Lin, Q.,
Sun, G., Ba Abdullah, M., Zhang, X., Hu, S., Al-Msallem, I.S. and Yu, J. (2012).

66
Large-scale collection and annotation of gene models for date palm (Phoenix
dactylifera L.). Plant Molecular Biology, 79(6):521-536.
Zhang, X.S., O’Neill, S.D. (1993). Ovary and gametophyte development are
coordinately regulated by auxin and ethylene following pollination. Plant Cell
5:403–418.
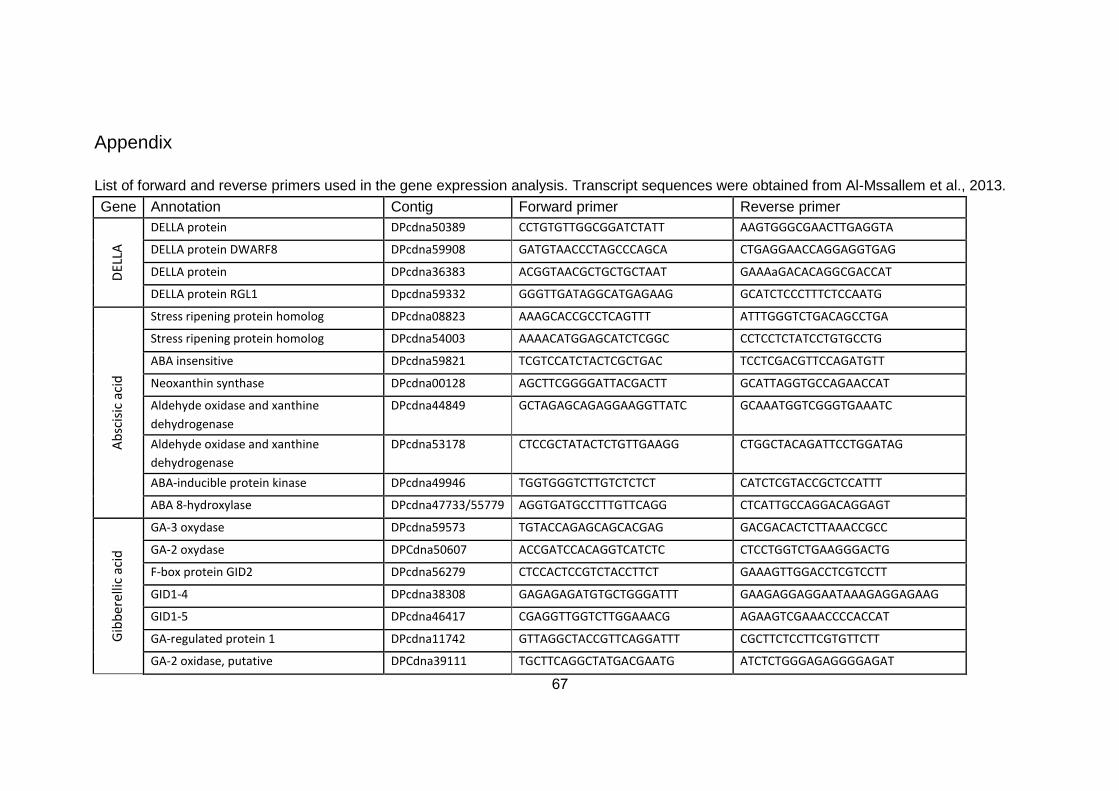
67
Appendix
List of forward and reverse primers used in the gene expression analysis. Transcript sequences were obtained from Al-Mssallem et al., 2013.
Gene Annotation Contig Forward primer Reverse primer
DEL
LA
DELLA protein DPcdna50389 CCTGTGTTGGCGGATCTATT AAGTGGGCGAACTTGAGGTA
DELLA protein DWARF8 DPcdna59908 GATGTAACCCTAGCCCAGCA CTGAGGAACCAGGAGGTGAG
DELLA protein DPcdna36383 ACGGTAACGCTGCTGCTAAT GAAAaGACACAGGCGACCAT
DELLA protein RGL1 Dpcdna59332 GGGTTGATAGGCATGAGAAG GCATCTCCCTTTCTCCAATG
Ab
scis
ic a
cid
Stress ripening protein homolog DPcdna08823 AAAGCACCGCCTCAGTTT ATTTGGGTCTGACAGCCTGA
Stress ripening protein homolog DPcdna54003 AAAACATGGAGCATCTCGGC CCTCCTCTATCCTGTGCCTG
ABA insensitive DPcdna59821 TCGTCCATCTACTCGCTGAC TCCTCGACGTTCCAGATGTT
Neoxanthin synthase DPcdna00128 AGCTTCGGGGATTACGACTT GCATTAGGTGCCAGAACCAT
Aldehyde oxidase and xanthine
dehydrogenase
DPcdna44849 GCTAGAGCAGAGGAAGGTTATC GCAAATGGTCGGGTGAAATC
Aldehyde oxidase and xanthine
dehydrogenase
DPcdna53178 CTCCGCTATACTCTGTTGAAGG CTGGCTACAGATTCCTGGATAG
ABA-inducible protein kinase DPcdna49946 TGGTGGGTCTTGTCTCTCT CATCTCGTACCGCTCCATTT
ABA 8-hydroxylase DPcdna47733/55779 AGGTGATGCCTTTGTTCAGG CTCATTGCCAGGACAGGAGT
Gib
ber
ellic
aci
d
GA-3 oxydase DPcdna59573 TGTACCAGAGCAGCACGAG GACGACACTCTTAAACCGCC
GA-2 oxydase DPCdna50607 ACCGATCCACAGGTCATCTC CTCCTGGTCTGAAGGGACTG
F-box protein GID2 DPcdna56279 CTCCACTCCGTCTACCTTCT GAAAGTTGGACCTCGTCCTT
GID1-4 DPcdna38308 GAGAGAGATGTGCTGGGATTT GAAGAGGAGGAATAAAGAGGAGAAG
GID1-5 DPcdna46417 CGAGGTTGGTCTTGGAAACG AGAAGTCGAAACCCCACCAT
GA-regulated protein 1 DPcdna11742 GTTAGGCTACCGTTCAGGATTT CGCTTCTCCTTCGTGTTCTT
GA-2 oxidase, putative DPCdna39111 TGCTTCAGGCTATGACGAATG ATCTCTGGGAGAGGGGAGAT

68
Gene Annotation Contig Forward primer Reverse primer
Gib
ber
ellic
aci
d
Chitin inducible GA-responsive protein DPcdna45603 CCCAGTACAGGCTCTTGGAT CTGCTTACATCGTCGTCAGC
Putative chitin-inducible GA-responsive
protein
DPcdna46561 AGAGTTGCCCGTTCAGATGA GGCTTTTATGAGGAGCTGGC
GA 20 oxidase DPcdna49465 GCTTCAACCACTACCCTCCA ACCAACATCTTGAGCCAGGA
GID1L2 DPcdna51868 ATGCGTTCCACTCCAAGAA CCAGCATCCACAATTCCAAAG
GA 2 oxidase DPcdna50446 ACTGTTGCCTGACTGGATTT ACTCCCATCCCTCAGAGATATT
GA 2 oxidase DPcdna50721 ACCTTCAGCAAGCTCCTTAC CTTAGCCCTTCAGCCATCAA
GA insensitive protein DPcdna45606 CAGGAATCCGTACACACATACA CAGAGCCCTATTGGTGGAAG
Ent-copalyl diphosphate synthase 1 DPcdna40087 AGCAAGCCAAGGATTTCTCA GTACagCCTCGCCTCTATGC
GASA-like protein DPcdna42426/25 GCTTCACTCACGGCAGAA CATGGGTCTTCCAATCGGTATAG
GAST-like protein DPcdna17382 CTCTTAGTCCCAACCTGTGTATC GAACCGTGCTGAGCTTCTAA
Au
xin
s
IAA-amido synthetase DPcdna37624/25 CAAGCAGGACAACAGCAATTT GTGGTGAGGGCAATCTCATAC
IAA-amido synthetase DPcdna39883 CTGGAGCAACCTGATACCTAATC CCAGCATAGTGCCTTAACTTCT
IAA-induced protein ARG7 DPcdna52909 CTGAGCCTGCCATTGTTTAAG TCTCGCAAGGGATTGTGATAG
ARP IAA 27 DPcdna25365 TGTGCCTGCCATGTTATTCC ACACAGGAAACAGGGACCAA
IAA hydrolase DPcdna39301 CAGTGCTACTGTGGACTTTCTT CATTTCCTCCGCAACCTTCT
IAA type protein DPcdna40883 CAAAGTGAATGGGAGCTGGA CATGACACAGGACTTGAGGAAA
IAA-amino acid hydrolase ILR1 DPcdna48023 CCTCATGGGCGTCAAGTATAG GAGTGCTACAAAGGGTGGAA
Auxin-responsive protein IAA13 DPcdna50014 GAAGAGCAACAGCAACGACA ACGGATGAGCATTGAGATCC
Auxin-repressed protein DPcdna01527 AGCAACCTAGCCACCAAGAA CCTCGGCACACaAAAAGTCT
Putative AUX1-like permease DPcdna12535/36 GCCCATGAGAAGCTGTACTATC CACCAAGCCTAGTAGCTCAAA
SAUR family protein DPcdna42587 TAGGTGGAGcCCTAGCtTTG AAATCGCCCTCCTCCTTCTA
IAA-amido synthetase DPcdna33546 cACAGCTaCaGCTCCTcataC TTCTCGGCATCTTTCTCGTT

69
Gene Annotation Contig Forward primer Reverse primer
Au
xin
s
Auxin efflux facilitator SlPIN4 DPcdna32615/13 CGTGTTCGCCAAGGAGTATAA CCATCGTAATCGGAAGAGCTATC
Auxin-independent growth promoter DPcdna39865 TGAGGTATATGGCGGAGAGG CATGCGTGATGAAAATCCTG
Auxin-regulated protein DPcdna46289 GTCTGGGAAGAGCTTTGTGC CAGATCAGGATTGGGGCTTA
Auxin efflux carrier protein-like DPcdna47006 CTCTTGTGGATGGTGGAGTAAG CAAGTCCTCGTGTCGGTTT
Putative ARF 1 DPcdna24437/38 CCACCAGAAGGAAGCGATAAG CGTGGATCAGTGGGTCATTT
ARP putative DPcdna56259 GTCGTTTtAggggtggtgaa agcagtatcccagcttccaa
AUX-induced protein DPcdna50778 GTCACAGATTCACCGTGTGG GAAACCCAACCTGGATACCTC
Auxin-induced lipid transfer protein DPcdna63518 GCAATCCTGTTCTCGTCTCC ATGTAGCCGCTCAAGTTTGG
Putative auxin response factor 6 DPcdna34533 ACCGTGGATACAgCCAAGAC AGCTTTGAGGTTGCTGGAAA
Putative auxin response factor 8 DPcdna59284 CTCTCTATTGCGGTGCCTAAT CAGAAGACCCATCCAGGTAAC
Auxin response factor 11 DPcdna44725 GCGACCAAGATTAGAAACAACAC GCAGCATCCAGTCCTCATTAT
Auxin response factor 27 DPcdna46801 ACGGTCTCTACGAGTCATTCT CAGGCGAGAGTTCTTGGTTATAC
ARF16 DPCdna45922 ATGGGAGGCTTGCTGATATG GAACGGTTCATCTCCAGTTTG
Auxin response factor Dpcdna61034 CAGCCAAGGTAGCATCCATT AgCATCAGGGGACTGGTCTA
ETTIN-like ARF (3) DPcdna52785 TGTCAGTATAAGGTCAGACGAAATG GGTAGTTTGGCAGCTACTCTTATC
ACTIN CACTGCGGAACGGGAAAT GGATGGCTGGAAGAGGAC
F-BOX TGGCTGCTGTAGTTGTAGGATG CACCACCACCTGTTGATTTG
ELONGATION FACTOR CCAAGTGTGAAAGCAAGCAA TACTTCGCAGGCTGATTGTG
G6PDH ACAATCCGAGTCCACCCAC TTGCCTCCATCTGTTTACCC
Cyt
oki
nin
s
Cytokinin-O-glucosyltransferase 1 DPcdna59650 CCCTcCGAcTcCCATAAAAT GGTCCAGTGGAAGTGGATGT
Cytokinin dehydrogenase 4 DPcdna51045 GTGTTCGTAGCGGATGTTCT GCCTCTCTTCTCCTTCTCATTT
CYP84A33 DPcdna47466 CTGTGTCTCAAAGGAAGAGCA CCGCAGAGAAAGTACGAAGAG
CYP71AP4 DPcdna47669 CATTCGGCATGGCTAGTgTC CAACAGCAACCAGTTCAGCT
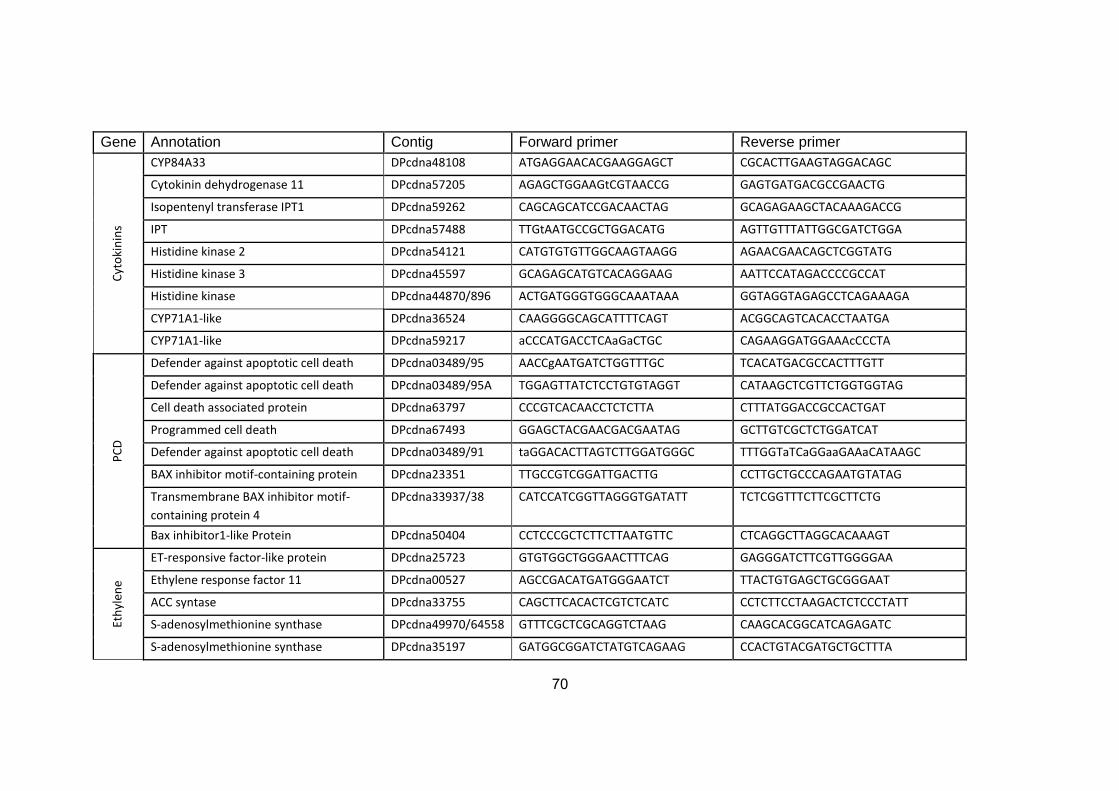
71
Gene Annotation Contig Forward primer Reverse primer
Cyt
oki
nin
s
CYP84A33 DPcdna48108 ATGAGGAACACGAAGGAGCT CGCACTTGAAGTAGGACAGC
Cytokinin dehydrogenase 11 DPcdna57205 AGAGCTGGAAGtCGTAACCG GAGTGATGACGCCGAACTG
Isopentenyl transferase IPT1 DPcdna59262 CAGCAGCATCCGACAACTAG GCAGAGAAGCTACAAAGACCG
IPT DPcdna57488 TTGtAATGCCGCTGGACATG AGTTGTTTATTGGCGATCTGGA
Histidine kinase 2 DPcdna54121 CATGTGTGTTGGCAAGTAAGG AGAACGAACAGCTCGGTATG
Histidine kinase 3 DPcdna45597 GCAGAGCATGTCACAGGAAG AATTCCATAGACCCCGCCAT
Histidine kinase DPcdna44870/896 ACTGATGGGTGGGCAAATAAA GGTAGGTAGAGCCTCAGAAAGA
CYP71A1-like DPcdna36524 CAAGGGGCAGCATTTTCAGT ACGGCAGTCACACCTAATGA
CYP71A1-like DPcdna59217 aCCCATGACCTCAaGaCTGC CAGAAGGATGGAAAcCCCTA
PC
D
Defender against apoptotic cell death DPcdna03489/95 AACCgAATGATCTGGTTTGC TCACATGACGCCACTTTGTT
Defender against apoptotic cell death DPcdna03489/95A TGGAGTTATCTCCTGTGTAGGT CATAAGCTCGTTCTGGTGGTAG
Cell death associated protein DPcdna63797 CCCGTCACAACCTCTCTTA CTTTATGGACCGCCACTGAT
Programmed cell death DPcdna67493 GGAGCTACGAACGACGAATAG GCTTGTCGCTCTGGATCAT
Defender against apoptotic cell death DPcdna03489/91 taGGACACTTAGTCTTGGATGGGC TTTGGTaTCaGGaaGAAaCATAAGC
BAX inhibitor motif-containing protein DPcdna23351 TTGCCGTCGGATTGACTTG CCTTGCTGCCCAGAATGTATAG
Transmembrane BAX inhibitor motif-
containing protein 4
DPcdna33937/38 CATCCATCGGTTAGGGTGATATT TCTCGGTTTCTTCGCTTCTG
Bax inhibitor1-like Protein DPcdna50404 CCTCCCGCTCTTCTTAATGTTC CTCAGGCTTAGGCACAAAGT
Eth
ylen
e
ET-responsive factor-like protein DPcdna25723 GTGTGGCTGGGAACTTTCAG GAGGGATCTTCGTTGGGGAA
Ethylene response factor 11 DPcdna00527 AGCCGACATGATGGGAATCT TTACTGTGAGCTGCGGGAAT
ACC syntase DPcdna33755 CAGCTTCACACTCGTCTCATC CCTCTTCCTAAGACTCTCCCTATT
S-adenosylmethionine synthase DPcdna49970/64558 GTTTCGCTCGCAGGTCTAAG CAAGCACGGCATCAGAGATC
S-adenosylmethionine synthase DPcdna35197 GATGGCGGATCTATGTCAGAAG CCACTGTACGATGCTGCTTTA

71
Gene Annotation Contig Forward primer Reverse primer
Eth
ylen
e
Ethylene-responsive transcription factor DPcdna43214 ACCTTAGCCTGGACCTCAAC CGAACCCTCCCAGCATCAAT
S-adenosylmethionine synthase DPcdna31510 GAGGTGCGGAAGAATGGAAC CTGGGTGGAGATGAGGACAG
Ethylene-responsive transcription factor
3
DPcdna51712 AAGCAATCAAATCGGCGTgT GGTTCCCTCCCTCTCCTTTG
Ethylene signal transcription factor DPcdna46898 ACTTTGGATGGGAACGTGAG CCTTCAGTGATGGTGGGAATAG
ET receptor DPcdna46300 CCAAGCACCATCTACCcACT GCATTGAGTCCCAGAGGAAA
Sen
esc
ence
Senescence-associated protein DPcdna54097 CGATACTGGCAGGAGGGATA CATGAGCGAGATCACCATGA
Putative senescence-associated protein DPcdna39108 GTTGAGCGTATGACTAGGAGATG TGGCGATTGAGTGATGAAGA
Senescence-associated protein DPcdna52345 GGGATAAACCTCCTGGCTTC GATTGGCAGTCATAGCAGAG
Senescence-associated protein 6 DPcdna40920 GGACTGTAGTGATCTCTTCTCTTG GTCTCCCTGATTGACTCCTTTG

תפתחות הפרי. בהשוואת מצביעות על כך שבתמר להאבקה יש חשיבות בהפעלת דפוס ה
ונשירה נשירת הפירות בין הזנים נמצאה נשירת פרי רבה בתפרחות 'מג'הול' שלא הואבקו
.מעטה יחסית בזן 'ברהי'
שלא הואבקו משני הזנים בכאלה ביצענו אנליזת ביטוי רחבת היקף בפרחים שהואבקו ו
גנים המעורבים . תשעים ושישה Microfluidic Dynamic Array (Fluidigm)במערכת
ואתילן) נבחרו על סמך מידע ABAן, יבבקרה הורמונאלית (לאוקסין, גיברלין, ציטוקינינ
מטרנסקריפטום פרי התמר שפורסם לאחרונה. רמות הביטוי היחסיות של גנים אלו נבחנו
בחמישה שלבי התפתחות מוקדמים של הפרי בשני הזנים. תוצאות ראשוניות מצביעות על
שונים בין זנים, טיפולי האבקה ושלבים התפתחותיים. דפוסי ביטוי

תקציר
במטע התמרים המודרני האבקה ודילול הפירות הינם תהליכים מחושבים. הפריה של
מרבית הפרחים תביא להתפתחות פירות קטנים, אשר תחייב דילול פרי ידני בהיקף רחב
פרחים לא האבקה לא יעילה תביא ליבול נמוך. בתנאים אלה עלולים ובעלות גבוהה.
בודדים או משולשים, להם אין כל ערך מסחרי. ,פרתנוקרפייםפירות ללהתפתח מופרים
למרות שהפרח הנקבי מכיל שלוש שלחות, רק אחת מהן מתפתחת לפרי ושתי האחרות
. בנוסף, ליעילות ההאבקה, תנאי הסביבה והרקע הגנטי (זנים שונים) השפעה על מתנוונות
החנטה והתפתחות הפרי. ,התהליכים ההתפתחותיים של ההפריה
מטרת העבודה היא אפיון מקיף של ההפריה, החנטה והתפתחות הפרי המוקדמת בתנאי
:הםהתמקדנו בעבודה הנושאים הספציפיים בהם סביבה שונים.
ני תמר, 'ברהי' ו'מג'הול'.זפיזיולוגי של תהליכי ההפריה והחנטה בשני -איפיון מורפולוגי .1
המוקדמת.לימוד השפעות הטמפרטורה על ההפריה והתפתחות הפרי .2
בחינה של דפוסי הביטוי של גנים המעורבים בבקרה ההורמונלית של התפתחות הפרי .3
ושל התנוונות שתיים מהשחלות.
. לבחינת אותו קשה להכניס לחדרי גידול ולבחון בתנאים מבוקרים התמר הינו עץ גדול מאוד
וניות. השפעות הסביבה על ביולוגיית הרבייה של התמר השתמשנו במספר גישות ניסי
במטע. הצלחנו רק באופן חלקי לכייל in plantaלבין ניסיונות in vitroשילבנו בן ניסיונות
, משום ש"חיי המדף" של in vitroפרוטוקול להאבקה של מקטעי תפרחות תמר במצע נוזלי
הפרחים המנותקים היו קצרים מאוד ותהליכי הזדקנות של הפרח והתפרחת חלו תוך מספר
עיים.ימים עד שבו
שמקיפים את התפרחות המואבקות בעצים שלמים , "פיטוטרונים מודולריים", תאים ייחודיים
על in plantaבמטע ומאפשרים השראה של משטרי סביבה מבוקרים בסביבת האשכול
התנוונות שתיים , . תהליכי צמיחת הנחשון, ההפריה, החטנהפותחו בפרויקט העץ במטע
פרתנוקרפיים אופיינו באנליזות מקרוסקופיות מהשחלות, וכן התפתחות פירות
ומיקרוסקופיות. הצגנו הבדלים משמעותיים בביולוגית הרבייה של שני הזנים, בהתפתחות
פירות פרתנוקרפיים, בנשירת חנטים ובבקרה שונה של התהליכים ההתפתחותיים.
ונים, טמפרטורות נמוכות יחסית שהושרו במהלך ההפריה הורידו את קצב צמיחת הנחש
העלו את רמת הפירות הפרתנוקרפים שנוצריים והאטו את קצב התפתחות הפרי הנורמאלי.
באמצעות אנליזה היסטולוגית הגדרנו "צמתים התפתחותיים" ושלבים התפתחות הפרי
המוקדמת בשני הזנים, ואישרנו שתהליכי ההתנוונות של ביציות ושחלות מתרחשים מוקדם
גית. בתנאי הניסוי, התנוונות ביציות זוהתה לראשונה ב'ברהי' יותר משניתן לזהות מורפולו
רחים מואבקים היו השחלות בפיום, והחלה כשבוע מאוחר יותר ב'מג'הול'. 14-לאחר כ
יום בשני הזנים. תוצאות אלה 27גדולות יותר לעומת פרחים שלא הואבקו כבר אחרי

עבודה זו נעשתה בהדרכתם של
דר' יובל כהן, מינהל המחקר החקלאי, מכון וולקני
וולקניפרופ' רינה קמנצקי, מינהל המחקר החקלאי, מכון

השפעות תנאי הסביבה על תהליכי ההפריה והחנטה
(.Phoenix dactylifera L)בתמרים
עבודת גמר
ע"ש רוברט ה. סמית, מוגשת לפקולטה למדעי החקלאות, המזון והסביבה
האוניברסיטה העברית בירושלים
לשם קבלת תואר "מוסמך למדעי החקלאות"
מאת
פיליפ סלבקוביץ'
5102ינואר טבת תשע"ה