engineered probiotics for local tumor delivery of checkpoint … · tumor response and subsequently...
TRANSCRIPT

1
Engineered probiotics for local tumor delivery of checkpoint blockade nanobodies
Candice Gurbatri1, Courtney Coker1, Taylor E. Hinchliffe1, Ioana Lia1, Samuel Castro1, Piper M.
Treuting2, Nicholas Arpaia3,4, and Tal Danino1,4,5 *
1Department of Biomedical Engineering, Columbia University, New York, NY 10027, USA
2Department of Comparative Medicine, University of Washington, Seattle, WA, USA
3Department of Microbiology & Immunology, Vagelos College of Physicians and Surgeons of Co-
lumbia University, New York, NY 10032, USA
4Herbert Irving Comprehensive Cancer Center, Columbia University, New York, NY 10027, USA
5Data Science Institute, Columbia University, New York, NY 10027, USA
*Correspondence should be addressed to T.D. ([email protected])
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

2
ABSTRACT
Immunotherapies such as checkpoint inhibitors have revolutionized cancer therapy yet lead to a
multitude of immune-related adverse events, suggesting the need for more targeted delivery sys-
tems. Due to their preferential colonization of tumors and advances in engineering capabilities
from synthetic biology, microbes are a natural platform for the local delivery of cancer therapeu-
tics. Here, we present an engineered probiotic bacteria system for the controlled production and
release of novel immune checkpoint targeting nanobodies from within tumors. Specifically, we
engineered genetic lysis circuit variants to effectively release nanobodies and safely control bac-
teria populations. To maximize therapeutic efficacy of the system, we used computational model-
ing coupled with experimental validation of circuit dynamics and found that lower copy number
variants provide optimal nanobody release. Thus, we subsequently integrated the lysis circuit
operon into the genome of a probiotic E. coli Nissle 1917, and confirmed lysis dynamics in a
syngeneic mouse model using in vivo bioluminescent imaging. Expressing a nanobody against
PD-L1 in this strain demonstrated enhanced efficacy compared to a plasmid-based lysing variant,
and similar efficacy to a clinically relevant monoclonal antibody against PD-L1. Expanding upon
this therapeutic platform, we produced a nanobody against cytotoxic T-lymphocyte associated
protein -4 (CTLA-4), which reduced growth rate or completely cleared tumors when combined
with a probiotically-expressed PD-L1 nanobody in multiple syngeneic mouse models. Together,
these results demonstrate that our engineered probiotic system combines innovations in synthetic
biology and immunotherapy to improve upon the delivery of checkpoint inhibitors.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

3
SENTENCE SUMMARY
We designed a probiotic platform to locally deliver checkpoint blockade nanobodies to tumors
using a controlled lysing mechanism for therapeutic release.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

4
MAIN TEXT
INTRODUCTION
Cancers exploit checkpoint signaling pathways by expressing ligands such as programmed cell
death protein-ligand 1, PD-L1, which bind to the PD-1 receptor to inhibit the activation, expansion,
and function of T cells and support immune evasion (1-3). These inhibitory checkpoint blockade
mechanisms have prompted the exploration of blocking monoclonal antibodies (mAbs) as thera-
peutics against these molecules. While anti-PD-L1 mAbs have achieved some level of tumor re-
gression in ~30% of cancers like melanoma (4, 5), they can also result in immune-related adverse
effects (iRAEs), with up to 70% of patients experiencing a range of toxicity grades resulting in
fatigue, skin rashes, endocrine disorders, or hepatic toxicities (6-9). Furthermore, combination
therapies of anti-PD-L1/PD-1 mAbs and anti- cytotoxic T-lymphocyte associated protein-4 (CTLA-
4) mAbs are more efficacious than monotherapies, but cause higher grade toxicities that lead to
favoring of less efficacious monotherapies or eventual drug discontinuation (10, 11). Moreover,
iRAEs are often treated with steroids or other immunosuppressants that may comprise the anti-
tumor response and subsequently affect the efficacy of checkpoint blockade antibodies (9, 12).
Thus, there is a clear need for improved delivery of checkpoint blockade inhibitors to the tumor
site to minimize adverse effects and shift the treatment preference towards combination therapy
approaches.
Rapid development of genetic technologies has enabled the engineering of intelligent microbial
delivery systems for therapeutic applications. Specifically, synthetic biology has generated nu-
merous examples of genetic circuits controlling bacteria growth and gene expression (13-19),
allowing them to sense and respond to disease states of inflammation, infection, and cancer (20-
23). Particularly for cancer, a multitude of studies have shown that systemic administration of
bacteria results in their selective colonization of tumors, providing a unique opportunity for tumor
drug delivery. This occurs primarily due to reduced immune surveillance along with the ability of
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

5
bacteria to grow within the hypoxic and necrotic tumor core (24-27). At the same time, microbiome
research efforts have revealed the widespread prevalence of microbes within malignant tissue
that do not cause infections or other long-term detrimental health effects (28, 29). Since bacteria
are both inherently present and selectively grow in tumors, they provide a natural platform for the
development of programmable therapeutic delivery vehicles.
Harnessing the converging advancements in both immunotherapy and synthetic biology, we en-
gineered probiotic bacteria to locally and controllably release PD-L1 and CTLA-4 antagonists in
the form of blocking nanobodies. Specifically, we coupled immunotherapeutic expression with an
optimized lysing circuit mechanism, such that probiotic bacteria carrying the anti-PD-L1 nanobody
homes to the necrotic tumor core, grow to a critical density, and lyse effectively releasing the anti-
PD-L1 nanobody to block the PD-1/PD-L1 interaction between tumor and T cells (Fig. 1a).
RESULTS
Construction and characterization of a probiotically-expressed PD-L1 Nb
A single-domain antibody, or nanobody (Nb), blocking PD-L1 was chosen from the RCSB Protein
Data Bank as therapeutic cargo. Unlike antibodies with a molecular size of approximately 150
kDa, Nbs are only 15 kDa and lack an Fc region, which requires glycosylation by mammalian
cells, and can therefore be produced recombinantly in bacteria (30, 31). Nbs provide multiple
advantages, including their small size, which allows for increased diffusion within the tumor mi-
croenvironment and more rapid clearance from the bloodstream through glomerular filtration
thereby reducing systemic toxicity (32). While faster blood clearance may suggest shorter thera-
peutic impact, the bacterial platform allows for continuous and intratumoral Nb production to im-
prove upon this limitation. Therefore, the PD-L1 Nb sequence was cloned downstream of a strong
constitutive pTac promoter on a stabilized, high copy ColE1 origin of replication plasmid to allow
for maximal gene expression. A human influenza hemagglutinin (HA) protein tag was added to 3’
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

6
end of the Nb for in vitro visualization and an Axe/Txe stability mechanism was cloned into the
vector to prevent plasmid loss during bacterial replication (Fig. S1a) (33). The plasmid was trans-
formed into the probiotic strain, E. coli Nissle 1917, containing a genomically integrated
luxCDABE cassette for bacterial tracking in vivo (EcN-lux). E. coli Nissle 1917 was chosen as the
therapeutic vehicle because of its proven safety, as it is currently prescribed for oral administration
in humans, and for its ease in genetic manipulation (34).
Flow cytometry analysis (Fig. 1b) and immunofluorescence (Fig. S1b) were used to confirm PD-
L1 expression on CT26 cells, a colorectal cell line that has a modest antitumor response to PD-
L1 mAb (35, 36). To investigate the binding pattern of a previously uncharacterized PD-L1 Nb,
bacterial lysate containing the probiotically-produced PD-L1 Nb was incubated on a monolayer of
CT26 cells and an anti-HA mAb was used to probe for PD-L1 Nb binding. Imaging using fluores-
cence microscopy revealed a similar expression pattern to that of PD-L1 expression probed for
by an anti-PD-L1 mAb (Fig. S1b). To quantify binding kinetics of the PD-L1 Nb, multiple dilutions
of bacterial lysate containing the PD-L1 Nb were co-incubated on a monolayer of CT26 cells with
a constant concentration of fluorescently-tagged anti-PD-L1 mAbs known to bind to either the
10F.9G2 or MIH7 epitope. With a more diluted lysate concentration, an increase in the 10F.9G2
mAb fluorescence was observed (Fig. 1c, d). Notably, a very small fraction – approximately 5%--
of the total lysate produced was needed to achieve binding to 50% of PD-L1+ CT26 cells. How-
ever, no change in fluorescence of the MIH7 mAb was observed as a function of bacterial lysate
(Fig. 1c), suggesting that the PD-L1 Nb specifically binds to an epitope similar to that recognized
by 10F.9G2.
Optimization of lysis circuits for controllable therapeutic release
Since we observed dose-dependent binding of the Nb to PD-L1 on CT26 cells, we sought to
enhance therapeutic release by optimizing a synchronized lysis circuit (SLC), whereby a bacterial
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

7
population lyses once a critical density is reached effectively releasing its therapeutic payload
(21). The SLC has been shown to aid in tumor-selective bacterial production, population limitation,
and therapeutic release, serving multiple purposes critical for translational efforts. However, one
drawback of the original SLC system is its reliance on plasmids. Since the quorum sensing genes
and more importantly the lysis gene are cloned onto plasmid vectors, they can be lost or mutated
during the bacterial growth cycle. To make this circuit more stable, we combined the original SLC
two-plasmid system into a single operon, one-plasmid system that we then genomically integrated
into EcN-lux. In this optimized system, the quorum sensing plux promoter drives transcription of
both quorum sensing genes, luxR and luxI, and the phage-derived lysis gene, φ X174E (Fig. 2a).
Use of only one promoter and one terminator allowed for this smaller module to be more easily
integrated and reduced repeat sequences that can lead to recombination.
While genomic integration of the SLC operon offers stability, it significantly reduces the copy num-
ber of quorum-sensing genes, prompting us to explore how this reduction would affect therapeutic
efficacy. Therefore, we computationally modeled the behavior of each circuit variant to further
understand the effect of copy number on lysis circuit dynamics. Using a system of ordinary differ-
ential equations, we modeled the dynamics of relevant variables in the lysing circuit: luxI, φ174E,
bacteria number, the small molecule used for bacterial communication AHL (N-Acyl homoserine
lactone), and therapeutic production. We first simulated the system to calculate two quantities:
number of bacteria required to reach quorum for the first lysis event and time required to reach
quorum (Fig. 2b, c). Here, we observed that decreasing copy number of the SLC circuit mono-
tonically increased the number of bacteria required to reach a quorum threshold before lysing.
However, time to lysis displayed non-monotonic behavior, where the single copy variant produced
a longer time to lysis than medium copy variants, but the high copy variant increased time to lysis
again. This non-monotonic behavior is due to the higher level of basal lysis when high copies of
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

8
the φ174E gene are present, resulting in slower growth, and thus longer time to reach quorum.
Further exploration of additional dynamic properties suggested that lower copy variants had a
higher mean oscillation amplitude and frequency (Fig. S2a, b).
The system’s behavior at lower copy number variants motivated our exploration into the dynamics
of therapeutic production as a direct function of population number over time, where we found
that lower copy variants produced more therapeutic in a given period of time and at a faster rate
compared to higher copy variants (Fig. 2d, Fig. S2c, d). Furthermore, the average rate of thera-
peutic production was time invariant, suggesting that this difference in therapeutic production rate
was primarily a function of copy number. Given the differences in therapeutic production observed
computationally, we sought to understand how our model could be used to predict bacterial dy-
namics in an in vivo setting. Previous studies have shown that bacterial growth kinetics are sig-
nificantly slower in vivo, with the doubling time of the bacteria increasing from ~30 minutes in vitro
to ~4 hours in vivo (26). Using our model, we simulated oscillation frequency as a function of
bacterial growth rate and copy number and found that lower copy number variants are more ro-
bust to a changing growth rate parameter (Fig. 2e). Together these findings suggest that the SLC-
int variant would result in maximal protein production across experimentally-relevant copy num-
bers.
To experimentally investigate system dynamics of the single operon SLC, we built a library of
plasmid variants covering a range of copy numbers for the quorum sensing genes, including a
single genomic integration of the operon into the φ80 site of the EcN-lux strain (SLC-int) (Fig. 2a).
Tracking of bacteria concentration over time in 96 well plates suggested that more copies of the
lysing circuit leads to lysis at a lower bacteria concentration with a rapid decay relationship relating
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

9
these two variables (Fig. 2f, g). Furthermore, the time it took each variant to reach quorum gen-
erally suggested that a lower copy number required more time to reach a critical density and the
relationship exhibited non-monotonic behavior, consistent with our simulations (Fig. 2g). To addi-
tionally visualize lysis events, the SLC-int variant was co-transformed with a constitutive plasmid
(ptac:GFP) and imaged over time using fluorescence microscopy (Movie S1). Moreover, multiple
oscillations for the SLC-int and SLC-P15A variants were observed by tracking the optical density
over time in a plate reader (Fig. S2e).
We next tested whether the SLC-int strain demonstrated lysis behavior and predictable dynamics
in vivo using a syngeneic CT26 hind flank tumor model (Fig. 3a). We monitored bacteria lumines-
cence from the integrated luxCDABE cassette in tumors over time using an In-vivo Imaging Sys-
tem (IVIS) and observed a critical density at approximately 40 hours and population decay to a
minimum level at 60 hours (Fig. 3b). Bacteria remaining then repopulated the system until the
measurements ceased at the 84 hour time point. When compared to the original SLC two-plasmid
system, the SLC-int system took twice as long to reach a critical density, similar to results obtained
from our simulations.
Determining the optimal release mechanism for the PD-L1 Nb
Considering the varying dynamics of the integrated and plasmid-based systems, both the SLC-
int and SLC-p15A systems were engineered to produce the PD-L1 Nb. By western blot we were
able to confirm production and release of the PD-L1 Nb into the supernatant and observed higher
protein levels from the lowest copy SLC-int variant compared to the non-lysing EcN-lux strain
transformed with the PD-L1 Nb plasmid at a 16 hour time point (Fig. S3a). We next explored the
therapeutic efficacy in vivo of both the SLC-int and SLC-p15A copy number variants producing
the PD-L1 Nb (SLC-int:PD-L1nb and SLC-p15A:PD-L1nb) in a CT26 hind flank syngeneic mouse
model. After 2 weeks, a moderate therapeutic difference was observed, with the SLC-int:PD-L1nb
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

10
variant resulting in slower relative tumor growth (Fig. 3c). As previously reported, some tumors
treated with an anti-PD-L1 monotherapy have a greater response than other tumors within the
same treatment group (35, 37). Therefore, we looked at individual mean trajectories and found
that for the SLC-int:PD-L1nb treated tumors, 2 of the 6 tumors grew much slower, with 1 tumor
trending towards complete regression (Fig. S3b). More interestingly, when therapeutic treatment
ceased at day 7 and tumors were left untreated for one week, the relative rate of tumor growth in
the SLC-p15A:PD-L1nb variant group increased dramatically in comparison to those in the SLC-
int:PD-L1nb group resulting in a significant difference in tumor growth rate (Fig.3d, S3c). This
difference in growth rate when treatment stopped is likely to be a result of a more stable SLC-int
bacterial system with less plasmid burden. Previous research suggests that increasing bacterial
plasmid burden may increase plasmid instability or lead to mutations, which could ultimately affect
the therapeutic efficacy of the strain (38). Together these findings led us to choose the SLC-int
strain as the release mechanism for the PD-L1 Nb therapeutic.
Comparison of the SLC-int: PD-L1nb system to anti- PD-L1 mAb
In the syngeneic hind flank mouse model of CT26, comparable therapeutic efficacy was observed
for both the 10F.9G2 mAb and the SLC-int:PD-L1nb at day 4 (Fig. 3e). Analysis of individual mean
tumor trajectories show that 1 out of 6 tumors in the 10F.9G2 mAb treated group showed minimal
relative tumor growth, whereas at least 2 of the 8 tumors in the SLC-int:PD-L1nb treated group
showed minimal growth or a reduction in tumor size (Fig. S4). Furthermore, the distribution plot
of the absolute tumor volumes for the 10F.9G2 mAb and the SLC-int:PD-L1nb were similar, with
the average absolute tumor volume for the SLC-int:PD-L1 Nb treated group being slightly smaller
than that of the 10F.9G2 mAb treated group (Fig. S4). While there was no striking difference in
tumor sizes between the two anti-PD-L1 treatment groups, histological analysis indicated a higher
dirty necrosis score in bacteria-treated tumors, suggesting that tumors treated with the SLC-
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

11
int:PD-L1nb had less viable tissue and more neutrophils present in their tumor microenvironment
compared to the anti-PD-L1 mAb group (Fig. 3f, Fig. S5).
We explored the versatility of a probiotically-produced PD-L1 Nb by comparing the therapeutic
effects of an EcN-lux:SLC-PD-L1nb and the 10F.9G2 mAb in a Balb/c-4T1 syngeneic hind flank
mouse model of triple negative breast cancer, where we observed severe toxicity and animal
death after only three anti- PD-L1 mAb doses (Fig.S6a, b). After the second dose of the anti-PD-
L1 mAb, the body condition of the mice visibly deteriorated with one-third of the mice losing more
than 10% of their weight and subsequently remained untreated for the rest of the trial. After the
third dose, the mice we had continued treating with the anti-PD-La mAb died and the remaining
mice, having never recovered body condition after the second dose, were euthanized (Fig. S6b).
In contrast, mice receiving the EcN-lux control and EcN-lux-SLC:PD-L1nb had no change in body
condition and appeared healthy throughout the trial. Toxicity to anti-PD-L1 monotherapy treatment
has been previously reported in this breast cancer model (39), but here we have demonstrated
that a probiotically-produced PD-L1 Nb mitigates this observed toxicity. Furthermore, our engi-
neered strain demonstrated moderate efficacy when compared to the WT EcN-lux control, sug-
gesting that our engineered probiotic delivery system maintained efficacy while reducing toxicity
(Fig. S6c).
Exploration of immunotherapeutic combination therapies
Due to its local delivery, bacterial therapy may be used to deliver combinations of therapeutics
without increasing iRAEs. Therefore, we explored combinations to enhance therapeutic efficacy
of our engineered SLC-int:PD-L1nb probiotic therapy in the CT26 model, where only moderate
efficacy of the PD-L1 monotherapy was observed consistent with previous results in literature (35,
36). The bacterially-expressed hemolysin exotoxin (hlyE) was chosen for combination with SLC-
int:PD-L1nb due to its ability to lyse mammalian cells and potentially release neoantigens that
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

12
could then be recognized by infiltrating T cells. Furthermore, previous studies have demonstrated
moderate efficacy of the hlyE as a monotherapy in mouse models of colorectal cancer (21). The
combination of SLC-int:PD-L1nb and EcN-lux producing hlyE (EcN-SLC-hlyE) significantly
slowed tumor growth when compared to the individual therapies (Fig. S7a). A distribution plot of
individual tumor volumes for each treatment group indicates that tumors with the combination
therapy were smaller than those treated with the monotherapies. (Fig. S7b). The overall effect of
the combination therapy is more apparent in the individual trajectories, where more mice receiving
the combination therapy demonstrated slowed tumor growth over time (Fig. S7c). Lastly, since
increased toxicity is observed when immunotherapeutics are used in combination, we tracked
body weight of the mice as a proxy for mouse health throughout the trial and saw no significant
changes in body weights between treatment groups at the end of the trial (Fig. S7d).
Combination of the SLC:PD-L1nb and EcN-SLC-hlyE therapeutics suggest that the PD-L1 Nb
therapeutic platform can be further maximized through combination therapy approaches. Clini-
cally, anti-CTLA-4 mAbs are frequently used in combination with anti-PD-L1 mAbs (40, 41).
Therefore, to make our platform more generalizable as a checkpoint blockade treatment option,
we engineered a microbial cocktail that could also block the interaction between CTLA-4 and its
associated ligands (42, 43). A Nb against CTLA-4 was chosen from the RCSB Protein Data Bank
and its sequence was cloned onto the same plasmid vector used to express the PD-L1 Nb (Fig.
S8a). In the CT26 mouse model, the SLC-int:CTLA-4nb monotherapy was moderately efficacious
and demonstrated similar efficacy to that of the anti-CTLA-4 mAb (Fig. S8b). While the therapeutic
response for tumors treated with SLC-int:CTLA-4nb was only modest, less toxicity was observed
in these mice compared to the anti-CTLA-4 mAb group, with the mice receiving EcN-lux only and
the SLC-int:CTLA-4nb increasing in body weight at a faster rate than those receiving the anti-
CTLA-4 mAb. Over the course of two weeks, the average weight of mice treated with the anti-
CTLA-4 mAb had increased by ~10%, while the average weight of the SLC-int:CTLA-4nb mice
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

13
had increased by ~30% (Fig. S8c). Additionally, treatment with the SLC-int:CTLA-4nb induced
the production of more IFNγ in the tumor microenvironment when compared to the anti- CTLA-4
mAb and the EcN-lux control (Fig. S8d). An increase in pro-inflammatory cytokines like IFNγ may
suggest an increase in T cell infiltration and proliferation, thereby providing further rationale for
combining the SLC-int:CTLA-4nb and SLC-int:PD-L1nb therapeutics.
To further explore efficacy in the CT26 model, we treated tumors with a combination of the check-
point blockade Nbs (Nb cocktail) against PD-L1 and CTLA-4 and observed a significant reduction
in the absolute tumor volume compared to the monotherapies (Fig. 4a). Furthermore, mouse body
weight was tracked throughout the trial and no significant weight changes were observed (Fig.
S9a). To explore the versatility of our combinatorial Nb platform to other models, we tested the
Nb cocktail in a syngeneic A20 lymphoma model, where previous literature reports therapeutic
efficacy when treated with combination therapies including checkpoint inhibitors (35). We ob-
served a striking efficacy of the Nb cocktail compared to individual SLC-int:CTLA-4nb and SLC-
int:PD-L1nb monotherapies, with 3 out of 5 tumors completely regressing and the remaining two
tumors showing significant size reduction (Fig. 4b). Moreover, no toxicity was observed and the
mice appeared to be healthy and maintain their body condition throughout the trial (Fig. S9b). To
further understand the underlying immune response of the observed therapeutic effect, we inter-
rogated the immunophenotype of extracted A20 tumors by flow cytometry and observed a de-
crease in the the population of regulatory T cells (Tregs, CD4+FOXP3+) in tumors treated with the
Nb cocktail (Fig. 4c). Furthermore, there was an increase in the proliferation of conventional CD4+
T cells (CD4+FOXP3–Ki67+) in the Nb cocktail-treated tumors, suggesting that the shift in favor of
responsive T cells and decrease in immunosuppressive Tregs may result in a more robust im-
mune response and mediate the therapeutic effect observed (Fig. 4d).
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

14
DISCUSSION
Here, we have demonstrated the first instance of PD-L1 and CTLA-4 antagonists being expressed
by bacteria, which allowed for local therapeutic production and minimized toxicity in multiple
syngeneic mouse models. Furthermore, we have characterized novel PD-L1 and CTLA-4 Nbs
that can be adapted into other biological circuits and we have optimized their therapeutic release
using a genomically-integrated lysing circuit, thereby reducing possible horizontal gene transfer
for translational studies. This circuit also serves as a biocontainment measure to confine the bac-
terial population to the tumor site thereby minimizing the risk of systemic iRAEs.
The SLC-int:Nb systems demonstrated comparable therapeutic efficacy to analogous clinically
relevant mAbs and combining the SLC-int:PD-L1 Nb with a bacterially-expressed hemolysin ex-
otoxin led to greater therapeutic efficacy than either monotherapy. Furthermore, the combined Nb
cocktail demonstrated tumor shrinkage in the CT26 model and resulted in complete tumor regres-
sion in the A20 model. We hypothesize that the synergistic effect of this combination therapy
might be a result of fewer immunosuppressive Tregs and an increase in the proliferation of CD4+
helper T cells. Moreover, SLC-int:CTLA-4nb led to more detectable IFNγ levels at the tumor site,
which could act cooperatively with the SLC-int:PD-L1nb by supporting increased infiltration of
immune cells into the tumor microenvironment. Additionally, the production of such cytokines is
specifically advantageous for CT26 tumors because colorectal cancers are considered “cold” due
to overproduction of prostaglandins that dampen the immune response (44). For this reason,
these tumors have only demonstrated modest responses to checkpoint blockade. Therefore, ad-
ministration of checkpoint blockade nanobodies in vehicles that might lead to more immune cell
recruitment could potentially enhance therapeutic responses which would be otherwise difficult to
attain for “cold” cancers. To understand the mechanism underlying the therapeutic effects of the
SLC-int:Nb system on these types of cancers, further investigation into the immune landscape
after treatment will be required.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

15
Cancer immunotherapies are often more effective in combination with other anticancer agents.
Therefore, future iterations of the SLC-int system will be programmed to produce a wide variety
of immunotherapeutics that can then be tested in combination with the SLC-int:Nb platform. Mi-
crobial-based therapeutic platforms are highly modular and optimal for the rapid production of
multiple drugs that can then be delivered as a cocktail. Furthermore, to make the system more
clinically relevant, other routes of therapeutic administration will be considered. With this in mind,
we developed the SLC-int:Nb system in the probiotic strain, E.coli Nissle 1917, which has been
shown to colonize liver metastases when delivered orally (45), thus offering a more translational
route of therapeutic delivery for more advanced metastatic disease.
All together, we have built a stable biological circuit integrated into a probiotic with therapeutics
analogous to the current treatment standard for optimization towards clinical translation. The SLC-
int:Nb system will help advance the cancer immunotherapy field by providing a delivery vehicle in
which combination therapies can be easily explored and toxicities minimized for a broader range
of cancer patients.
MATERIALS AND METHODS
Study Design
The goal of this study was to develop a probiotic platform that enables the controllable release
and effective delivery of checkpoint blockade Nbs within tumors. Due to the preferential growth of
microbes within the tumor core, we hypothesized that this delivery platform would reduce toxicities
and allow for the delivery of safer and more effective combination therapies. A lysing mechanism
was used to release the therapeutic and characterization of these therapeutics, including binding
kinetics was established in vitro with the CT26 cell line. We then sought to test our delivery system
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

16
in vivo, where all studies were performed on syngeneic hind flank mouse models of CT26, 4T1,
or A20 cancers. All mice were randomized prior to treatment and received injections of either
monotherapies (SLC-int:PD-L1nb, SLC-hlyE, SLC-int:CTLA-4nb) or combinations of therapies,
ensuring that the final concentration of bacteria injected was the same. Caliper measurements
were used to track tumor volume, and mouse weight was monitored as a proxy for mouse health.
Statistical analysis and sample sizes were determined from previous studies (21, 26, 45). Further
details on sample size and replications (technical or biological) are provided in the figure legends.
Plasmid and Strains
Our circuit strains were cultured in LB media with their respective antibiotics (SC101*variant:
100µg ml-1ampicillin, p15A variant: 100µg ml-1 spectinomycin, ColE1: 100µg ml-1 spectinomycin,
and all with 50µg ml-1 spectinomycin and kanamycin for strains also transformed with PD-L1 ther-
apeutic plasmid) with 0.2% glucose, in a 37oC shaking incubator. CT26 and 4T1 mammalian cells
were purchased from ATCC and cultured in RPMI supplemented with 10% fetal bovine serum
and 1% penicillin/streptomycin. A20 cells were purchased from ATCC and cultured in RPMI sup-
plemented with 10% fetal bovine serum and 1% penicillin/streptomycin and 0.01% 2-Mercap-
toethanol. Mammalian cells were grown inside a 37oC tissue culture incubator maintained at 5%
CO2. Plasmids were constructed using Gibson assembly cloning methods and integrated into
the φ80 site of E.coli Nissle 1917 using the CRIM protocol (46). The EcN-SLC-hlyE construct was
used in previous work from our group (21), while the PD-L1 Nb and CTLA-4 Nb sequence was
obtained from an the online RCSB protein data bank and synthesized.
Antibodies
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

17
Antibodies used for in vivo experiments include: anti-mouse PD-L1 (BioXCell cat: BE0101and
anti-mouse CTLA-4 (BioXCell cat: BE0164). Pre-conjugated primary antibodies used for flow cy-
tometry and microscopy include: Pe/Cy7 anti-mouse CD274 (Biolegend cat: 124313), PE/Cy7 Rat
IgG2b (Biolegend cat: 400617), Pe anti-mouse CD274 (Biolegend, cat: 155403), Pe Rat IgG2a
isotype (BS biosciences cat: 557076), anti-HA-AF 647 (R&D System cat: IC6875R-025), and
mouse IgG1-AF 647 (R&D systems cat: IC200R). For western blot an anti-HA high affinity (Roche,
cat: 11867423001) and a rat HRP were used. Specific antibodies used for flow cytometry on tumor
tissue are listed in the corresponding section.
Immunofluorescence and Characterization
Collecting PD-L1 Nb protein
Non-lysing bacterial strains containing the PD-L1 Nb therapeutic plasmid were grown in a 50mL
LB culture with appropriate antibiotics to an optical density of 0.6 then centrifuged at 3000 rcf for
5 minutes. The bacterial pellet was resuspended in 5mL of RPMI media. Samples were frozen at
-80oC and thawed in a 30oC incubator 5 times and centrifuged at 3000 rcf for 5 minutes to remove
bacterial debris. 1mL of the resulting lysate was then filtered through a 0.2 micron filter.
Flow cytometry
For flow cytometry binding experiments, 1x106 CT26 cells were co-incubated in a 14mL round
bottom FACS compatible tube with a constant concentration of pre-conjugated PD-L1 Ab (1:800
for the 10F.9G2 clone and 1:200 for the MIH7 clone) and dilutions of the previously prepared
bacterial-lysate containing PD-L1 Nb (0.1%, 1%, 10%, 100% of full lysate) in a total working vol-
ume of 1mL for 2 hours at room temperature. Samples were spun and washed with ice cold PBS
and analyzed on an BD LSRII flow cytometer.
For flow cytometry on ex vivo tumor tissue, tumors were extracted for immunophenotyping
on day 8 following bacteria treatment on days 0, 4, and 7. Lymphocytes were isolated from tumor
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

18
tissue by mechanical homogenization and digestion with collagenase A (1mg/ml, Roche) and
DNAase I (0.5ug/mL, Roche) in isolation buffer (RPMI 1640 supplemented with 5% FBS, 1%L-
glutamine, 1%penicillin/streptomycin, and 10mM Hepes) for 1h at 37C. Cells were then filtered
through 100um cell strainers and washed in isolation buffer before staining. A Ghost Dye cell
viability stainin was used as a live/dead marker. Extracellular antibodies used include: anti-B220
(BD 1:400), anti-CD4 (Tonbo), anti-CD8 (eBioscience 1:400) and anti-NKp46(BD 1:200). Cells
were then fixed using FOXP3/transcription factor staining buffer set (Tonbo) as per the manufac-
turer’s protocol and then stained intracellularly. Intracellular antibodies used include: anti-
TCRβ(BD 1:400), anti-Ki67(Thermo 1:400) and anti-FOXP3(ebioscience 1:400). Samples were
analyzed using a BD LSR Fortessa cell analyzer.
Single-stains and isotype controls were used for compensation of the dual stain. FlowJo
was used for all data analysis.
Microscopy
For microscopy, 1x106 CT26 cells were stained on a poly-l-lysine coated glass flat bottom plate
with appropriate antibodies for detection of PD-L1 expression (10F.9G2 clone). In a separate
experiment, full bacterial lysate containing PD-L1 Nb was incubated on the monolayer of CT26
cells and probed with a pre-conjugated anti-HA antibody. A Nikon Eclipse Ti microscope was
used for all microscopy and FIJI was used for any post-image analysis.
Characterization of PD-L1 Nb Expression and Release
To determine protein expression and functionality of the HA tag, PD-L1 Nb protein was released
using the freeze/thaw method described above. A western blot was use for protein visualization
and was probed for 1hr with the primary anti-HA antibody (1:800) and 1hr with a rat HRP (1:4000)
at room temperature. Protein was detected using a chemiluminescent substrate.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

19
In vivo Studies
Tumor models
Animal experiments were performed on 6-8 week old female BALB/C mice from Taconic with
bilateral subcutaneous hind flank tumors from an implanted mouse colorectal cancer cell line
CT26, mouse lymphoma cell line A20, or a mouse breast cancer cell line, 4T1. Tumor cells were
prepared for implantation at a concentration of 5e7 cells/ml in RPMI without phenol red. Cells
were implanted at 100uL per flank, with each implant consisting of 5x106 cells. Tumors grew for
approximately 10 days or until the diameter was 4-10mm in diameter or volume was ~200mm3
for CT26 and 4T1 tumors and ~150mm3 for A20 tumors. All mice were randomized prior to treat-
ment.
Bacteria growth and therapeutic administration
Bacterial strains were grown overnight in LB media containing appropriate antibiotics and 0.2%
glucose for less than 12 hours. The overnight was subcultured at a 1:100 dilution in 50mL of fresh
media with antibiotics and glucose and grown until an OD of approximately 0.05 to prevent bac-
teria containing SLC operon from reaching quorum. Bacteria were spun down at 3000 rcf and
washed 3 times with sterile ice-cold PBS. Bacteria were delivered intratumorally at a concentra-
tion of 5e7 cells/ml in PBS with a total of 30-40uL injected per flank every 3-4 days (OD of 0.1 is
equivalent to 1e8 cells/mL). The PD-L1 mAb and the CTLA-4 mAb were injected intraperitoneally
at a concentration of 200ug/mouse and 100ug/mouse respectively.
Tumor and bacteria monitoring in vivo
CT26 and 4T1 tumor volumes were quantified using calipers to measure length, width, and height
of each tumor with volumes being reported as LxWxH. A20 tumors were measured as (LxW)/2.
Tumors were measured every 3 to 4 days and mouse weight was tracked to monitor overall
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

20
mouse health. All bacterial strains used were luminescent, which could be measured with the In
Vivo Imaging System. Living Image software was used for luminescent quantification.
Histology
Tumors were extracted, fixed in 10% formalin, and sent to the Histology and Imaging Core at the
University of Washington, where the tissue was processed and Haemotoxylin and Eosin stained.
The pathologist blinded to sample treatments manually scored the samples for dirty necrosis (0 =
not present, 1 = minimal, 2 = mild, 3 = moderate, and 4 = severe, abscess like)
Cytokine Analysis
Tumors were extracted from mice on day 14 following initial treatment. After one snap-freeze
cycle, tumors were mechanically lysed in complete lysis buffer (1x Protease Inhibitor cocktail,
Sigma Aldrich Cat: P27using a gentleMACS Dissociator.14-1BTL; EMD Millipore lysis buffer (Cat:
43-040) using a gentleMACS tissue dissociate. Once fully dissociated, the tumor homogenates
were centrifuged for 10min at 10,000g to clear debris. The lysate was spun for 5min at 5,000g.
Pre-cleared lysates were stored at -80C until analysis by Luminex Multiplex Assay. A Bradford
colorimetric assay was used to normalize protein concentrations and 35ug lysate in 25uL volume
was analyzed in the Luminex Multiplex Assay (EMD Millipore) according to the manufacturer’s
protocol.
Statistical Analysis
Statistical tests were calculated in GraphPad Prism 7.0. The details of the statistical tests are
indicated in the respective figure legends. Where data was assumed to be normally distributed,
values were compared using a one-way ANOVA for single variable or a two-way ANOVA for more
than one variable with a Bonferroni post-test applied for multiple comparisons. For categorical
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

21
data comparisons, data was assumed to be nonparametric and a Mann Whitney U Rank test was
used for single variable, two group comparisons.
SUPPLEMENTARY MATERIALS
Figure S1: Engineering and characterization of the ptac: PD-L1 Nb plasmid.
Figure S2: Computational modeling of the lysing variant dynamics.
Figure S3: Characterization of lysing circuit variants in a syngeneic colorectal model.
Figure S4: Individual tumor trajectories for comparison between probiotic expressing PD-L1nb
(SLC-int:PD-L1nb) and systemically delivered antibody in CT26 model.
Figure S5: Histological images and dirty necrosis scoring in CT26 mouse tumors.
Figure S6: Comparison between probiotically-produced PD-L1 Nb and systemically delivered an-
tibody in 4T1 model.
Figure S7: SLC-int:PD-l1nb and bacterially-expressed hemolysin exotoxin (hlyE) combination
therapy is more efficacious than individual therapies.
Figure S8: Engineered probiotic expressing CTLA-4 Nb.
Figure S9: Relative body weight for combination of PD-L1 and CTLA-4 probiotics.
Mathematical Model
Movie S1: Visualization of SLC-int lysis.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

22
REFERENCES
1. Baumeister SH, Freeman GJ, Dranoff G, & Sharpe AH (2016) Coinhibitory pathways in
immunotherapy for cancer. Annual review of immunology 34:539-573.
2. Hude I, Sasse S, Engert A, & Bröckelmann PJ (2017) The emerging role of immune
checkpoint inhibition in malignant lymphoma. Haematologica 102(1):30-42.
3. Parcesepe P, Giordano G, Laudanna C, Febbraro A, & Pancione M (2016) Cancer-
associated immune resistance and evasion of immune surveillance in colorectal cancer.
Gastroenterology research and practice 2016.
4. Zhang T, Xie J, Arai S, Wang L, Shi X, Shi N, Ma F, Chen S, Huang L, & Yang L (2016)
The efficacy and safety of anti-PD-1/PD-L1 antibodies for treatment of advanced or
refractory cancers: a meta-analysis. Oncotarget 7(45):73068.
5. D’Angelo SP, Larkin J, Sosman JA, Lebbé C, Brady B, Neyns B, Schmidt H, Hassel JC,
Hodi FS, & Lorigan P (2017) Efficacy and safety of nivolumab alone or in combination with
ipilimumab in patients with mucosal melanoma: a pooled analysis. Journal of Clinical
Oncology 35(2):226.
6. Naidoo J, Page D, Li B, Connell L, Schindler K, Lacouture M, Postow M, & Wolchok J
(2015) Toxicities of the anti-PD-1 and anti-PD-L1 immune checkpoint antibodies. Annals
of Oncology 26(12):2375-2391.
7. Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC, McDermott DF, Powderly
JD, Carvajal RD, Sosman JA, & Atkins MB (2012) Safety, activity, and immune correlates
of anti–PD-1 antibody in cancer. New England Journal of Medicine 366(26):2443-2454.
8. Brahmer JR, Tykodi SS, Chow LQ, Hwu W-J, Topalian SL, Hwu P, Drake CG, Camacho
LH, Kauh J, & Odunsi K (2012) Safety and activity of anti–PD-L1 antibody in patients with
advanced cancer. New England Journal of Medicine 366(26):2455-2465.
9. Michot J, Bigenwald C, Champiat S, Collins M, Carbonnel F, Postel-Vinay S, Berdelou A,
Varga A, Bahleda R, & Hollebecque A (2016) Immune-related adverse events with
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

23
immune checkpoint blockade: a comprehensive review. European Journal of Cancer
54:139-148.
10. Chae YK, Arya A, Iams W, Cruz MR, Chandra S, Choi J, & Giles F (2018) Current
landscape and future of dual anti-CTLA4 and PD-1/PD-L1 blockade immunotherapy in
cancer; lessons learned from clinical trials with melanoma and non-small cell lung cancer
(NSCLC). Journal for immunotherapy of cancer 6(1):39.
11. Du X, Liu M, Su J, Zhang P, Tang F, Ye P, Devenport M, Wang X, Zhang Y, & Liu Y (2018)
Uncoupling therapeutic from immunotherapy-related adverse effects for safer and
effective anti-CTLA-4 antibodies in CTLA4 humanized mice. Cell research 28(4):433.
12. Arbour KC, Mezquita L, Long N, Rizvi H, Auclin E, Ni A, Martínez-Bernal G, Ferrara R, Lai
WV, & Hendriks LE (2018) Impact of Baseline Steroids on Efficacy of Programmed Cell
Death-1 and Programmed Death-Ligand 1 Blockade in Patients With Non–Small-Cell
Lung Cancer. Journal of Clinical Oncology 36(28):2872-2878.
13. Gardner TS, Cantor CR, & Collins JJ (2000) Construction of a genetic toggle switch in
Escherichia coli. Nature 403(6767):339.
14. Elowitz MB & Leibler S (2000) A synthetic oscillatory network of transcriptional regulators.
Nature 403(6767):335.
15. Stricker J, Cookson S, Bennett MR, Mather WH, Tsimring LS, & Hasty J (2008) A fast,
robust and tunable synthetic gene oscillator. Nature 456(7221):516.
16. Danino T, Mondragón-Palomino O, Tsimring L, & Hasty J (2010) A synchronized quorum
of genetic clocks. Nature 463(7279):326.
17. Prindle A, Samayoa P, Razinkov I, Danino T, Tsimring LS, & Hasty J (2012) A sensing
array of radically coupled genetic ‘biopixels’. Nature 481(7379):39.
18. Tabor JJ, Salis HM, Simpson ZB, Chevalier AA, Levskaya A, Marcotte EM, Voigt CA, &
Ellington AD (2009) A synthetic genetic edge detection program. Cell 137(7):1272-1281.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

24
19. Liu C, Fu X, Liu L, Ren X, Chau CK, Li S, Xiang L, Zeng H, Chen G, & Tang L-H (2011)
Sequential establishment of stripe patterns in an expanding cell population. Science
334(6053):238-241.
20. Patyar S, Joshi R, Byrav DP, Prakash A, Medhi B, & Das B (2010) Bacteria in cancer
therapy: a novel experimental strategy. Journal of biomedical science 17(1):21.
21. Din MO, Danino T, Prindle A, Skalak M, Selimkhanov J, Allen K, Julio E, Atolia E, Tsimring
LS, Bhatia SN, & Hasty J (2016) Synchronized cycles of bacterial lysis for in vivo delivery.
Nature 536(7614):81-85.
22. Zheng JH, Nguyen VH, Jiang S-N, Park S-H, Tan W, Hong SH, Shin MG, Chung I-J, Hong
Y, & Bom H-S (2017) Two-step enhanced cancer immunotherapy with engineered
Salmonella typhimurium secreting heterologous flagellin. Science translational medicine
9(376):eaak9537.
23. Anderson JC, Clarke EJ, Arkin AP, & Voigt CA (2006) Environmentally controlled invasion
of cancer cells by engineered bacteria. Journal of molecular biology 355(4):619-627.
24. Leschner S, Westphal K, Dietrich N, Viegas N, Jablonska J, Lyszkiewicz M, Lienenklaus
S, Falk W, Gekara N, & Loessner H (2009) Tumor invasion of Salmonella enterica serovar
Typhimurium is accompanied by strong hemorrhage promoted by TNF-α. PloS one
4(8):e6692.
25. Forbes NS (2010) Engineering the perfect (bacterial) cancer therapy. Nature Reviews
Cancer 10(11):785-794.
26. Danino T, Lo J, Prindle A, Hasty J, & Bhatia SN (2012) In vivo gene expression dynamics
of tumor-targeted bacteria. ACS synthetic biology 1(10):465-470.
27. Zhao M, Yang M, Li X-M, Jiang P, Baranov E, Li S, Xu M, Penman S, & Hoffman RM
(2005) Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing
Salmonella typhimurium. Proceedings of the National Academy of Sciences of the United
States of America 102(3):755-760.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

25
28. Berg RD (1996) The indigenous gastrointestinal microflora. Trends in microbiology
4(11):430-435.
29. Savage D (1987) Microorganisms associated with epithelial surfaces and stability of the
indigenous gastrointestinal microflora. Molecular Nutrition & Food Research 31(5-6):383-
395.
30. Bannas P, Hambach J, & Koch-Nolte F (2017) Nanobodies and nanobody-based human
heavy chain antibodies as antitumor therapeutics. Frontiers in immunology 8:1603.
31. Lee YJ & Jeong KJ (2015) Challenges to production of antibodies in bacteria and yeast.
Journal of bioscience and bioengineering 120(5):483-490.
32. Hu Y, Liu C, & Muyldermans S (2017) Nanobody-based delivery systems for diagnosis
and targeted tumor therapy. Frontiers in immunology 8:1442.
33. Fedorec AJ, Ozdemir T, Doshi A, Rosa L, Velazquez O, Danino T, & Barnes CP (2018)
Two new plasmid post-segregational killing mechanisms for the implementation of
synthetic gene networks in E. coli. bioRxiv:350744.
34. Secher T, Kassem S, Benamar M, Bernard I, Boury M, Barreau F, Oswald E, & Saoudi A
(2017) Oral administration of the probiotic strain Escherichia coli Nissle 1917 reduces
susceptibility to neuroinflammation and repairs experimental autoimmune
encephalomyelitis-induced intestinal barrier dysfunction. Frontiers in immunology 8:1096.
35. Sagiv-Barfi I, Kohrt HE, Czerwinski DK, Ng PP, Chang BY, & Levy R (2015) Therapeutic
antitumor immunity by checkpoint blockade is enhanced by ibrutinib, an inhibitor of both
BTK and ITK. Proceedings of the National Academy of Sciences:201500712.
36. Kim K, Skora A, Li Z, Tam A, Diaz L, Papadopolous N, Blosser L, Kinzler K, Vogelstein B,
& Zhou S (2014) Eradication of metastatic mouse cancers resistant to immune checkpoint
blockade by suppression of myeloid-derived cells. Journal for immunotherapy of cancer
2(3):P267.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

26
37. Rios-Doria J, Durham N, Wetzel L, Rothstein R, Chesebrough J, Holoweckyj N, Zhao W,
Leow CC, & Hollingsworth R (2015) Doxil synergizes with cancer immunotherapies to
enhance antitumor responses in syngeneic mouse models. Neoplasia 17(8):661-670.
38. Fedorec AJ (2014) Mechanisms for Plasmid Maintenance. CoMPLEX, University College
London.
39. Mall C, Sckisel GD, Proia DA, Mirsoian A, Grossenbacher SK, Pai C-CS, Chen M,
Monjazeb AM, Kelly K, & Blazar BR (2016) Repeated PD-1/PD-L1 monoclonal antibody
administration induces fatal xenogeneic hypersensitivity reactions in a murine model of
breast cancer. Oncoimmunology 5(2):e1075114.
40. Lussier DM, Johnson JL, Hingorani P, & Blattman JN (2015) Combination immunotherapy
with α-CTLA-4 and α-PD-L1 antibody blockade prevents immune escape and leads to
complete control of metastatic osteosarcoma. Journal for immunotherapy of cancer
3(1):21.
41. Ott PA, Hodi FS, & Robert C (2013) CTLA-4 and PD-1/PD-L1 blockade: new
immunotherapeutic modalities with durable clinical benefit in melanoma patients. (AACR).
42. Contardi E, Palmisano GL, Tazzari PL, Martelli AM, Fala F, Fabbi M, Kato T, Lucarelli E,
Donati D, & Polito L (2005) CTLA-4 is constitutively expressed on tumor cells and can
trigger apoptosis upon ligand interaction. International journal of cancer 117(4):538-550.
43. Leach DR, Krummel MF, & Allison JP (1996) Enhancement of antitumor immunity by
CTLA-4 blockade. Science 271(5256):1734-1736.
44. Sandage BW, Talley JJ, Martinez EJ, Franklin MR, Meade MA, Saims D, Thayer M, Wise
S, Draper D, & Leopold W (2017) Abstract 2619: Combination of ECP1014 and anti-PD-
L1 reduces tumor growth in the CT26 murine colon carcinoma model of a cold tumor.
Cancer Research 77(13 Supplement):2619-2619.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

27
45. Danino T, Prindle A, Kwong GA, Skalak M, Li H, Allen K, Hasty J, & Bhatia SN (2015)
Programmable probiotics for detection of cancer in urine. Science translational medicine
7(289):289ra284-289ra284.
46. Haldimann A & Wanner BL (2001) Conditional-replication, integration, excision, and
retrieval plasmid-host systems for gene structure-function studies of bacteria. Journal of
bacteriology 183(21):6384-6393.
ACKNOWLEDGEMENTS
This work was supported in part by the NIH Pathway to Independence Award (R00CA197649-
02), DoD Idea Development Award (LC160314), DoD Era of Hope Scholar Award (BC160541),
and the National Science Foundation Graduate Research Fellowship under Grant No. (1644869).
Research reported in this publication performed in the CCTI Flow Cytometry Core, was supported
in part by the Office of the Director, National Institutes of Health under awards S10RR027050. Ad-
ditionally, these studies used the resources of the Cancer Center Flow Core Facility funded in
part through Center Grant P30CA013696. The content is solely the responsibility of the authors
and does not necessarily represent the official views of the National Institutes of Health. We would
like to thank Omar Din for assistance with the computational modeling and in vitro lysing circuit
variant dynamics. We would also like to thank members of the Danino lab for critical review of the
manuscript.
AUTHOR CONTRIBUTIONS
C.G. and T.D. conceived and designed the therapeutic platform. C.G. built and characterized the
library of SLC variants, optimized the computational modeling platform, and performed experi-
ments in vitro to test bacterial strains and characterize the immunotherapeutic. C.G., C.C., T.H.,
I.L, and S.C. designed and performed in vivo experiments. C.G and N.A designed and analyzed
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

28
the immune characterization assays. P.T performed histology.C.G. and T.D. analyzed data and
wrote the manuscript with input from all of the other authors.
COMPETING INTERESTS STATEMENT
C.G., N.A. and T.D. have filed a provisional patent application with the US Patent and Trademark
Office (US Patent Application No. 62/747,826) related to this work.
DATA AVAILABILITY
The data that support the findings of this study are available within the paper and its supplemen-
tary information files. Additional data are available from the authors upon reasonable request.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

29
FIGURE LEGENDS
Figure 1: Design of a probiotic cancer therapy system for release of a functional anti-PD-
L1 blocking nanobody. (a) Bacteria engineered to controllably release a constitutively produced
PD-L1 blocking nanobody (Nb), which binds to PD-L1 on the tumor cell surface and blocks its
interaction with PD-1 expressed on T cells. (b) Flow cytometry analysis of PD-L1 expression on
CT26 cells (pink peak: isotype control; blue peak: stained with an anti-PD-L1 mAb, 10F.9G2
clone). (c) Binding curve of PD-L1 Nb to the 10F.9G2 and MIH7 PD-L1 epitopes on CT26 cells.
(d) Flow cytometry plots of CT26 populations, where CT26 cells were co-incubated with a con-
stant concentration of PE/Cy7-conjugated 10F.9G2 mAb and varying concentrations of bacterial
lysates containing the PD-L1 Nb.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

30
Figure 2: Characterization of lysing variant dynamics. (a) Circuit diagram of the single operon
SLC circuit in which plux drives the transcription of luxR, luxI and φ 174E genes under a single
promoter. The circuit was cloned onto three plasmids of different copy numbers: SC101* (3-4
copies, low), p15A (15-20 copies, medium), and ColE1 (70-100 copies, high) and integrated once
into the φ80 site of the EcN-lux genome. (b) Computational simulation of one lysis event for the
copy number variants over time (green: single integrant, red: low copy, blue: medium copy, black:
high copy). (c) Computational simulation of the number of bacteria needed to reach quorum as a
function of copy number (left y axis: black) and time to first lysis event (right y axis: gray). (d)
Simulated heatmap of the PD-L1 Nb protein produced (z-axis) as a function of both copy number
(y-axis) and time (x-axis). (e) Simulated heatmap of the oscillation frequency (z-axis) as a function
of copy number (y-axis) and growth rate (x-axis). (f) Bacterial population dynamics in batch culture
of the four lysing variants. (g) Experimental data of the number of bacteria needed to reach
quorum as a function of copy number and time to first lysis event. Error bars represent S.E.M of
repeated experiments
a
Time (au)
100
50
00 2000
Copy Number
Time to lysis (au)
0.5
0.25
0 0
250
500
# ba
cter
ia (a
u)Tim
e to lysis (min)
Copy Number
0 70
# ba
cter
ia (a
u) SLC-intSC101*P15AColE1
b
AHL
φ X174E
LuxR-AHL
luxI Tp15A
mediumcopy
SC101*
lowcopy
single copy
ColE1
high copy
LYSIS
d f
Copy
#
c e g
0
2
4
6
100 200 400 0 120 240 360 480
0.0
0.2
0.4
Time (min)
OD
600
OD
600
0 700
3
6
150
275
400
SLC-intSC101*P15AColE1
Growth Rate
100
50
010-3
Copy
#
10-2
log10[PD-L1 Nb]
Oscillation Frequency (au)
1000
5x10-3
×10-3
0
1
2
3
-3
-1012
4
-2
3
Time (au)
35
Figure 2, Gurbatri et al.
35
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint
31
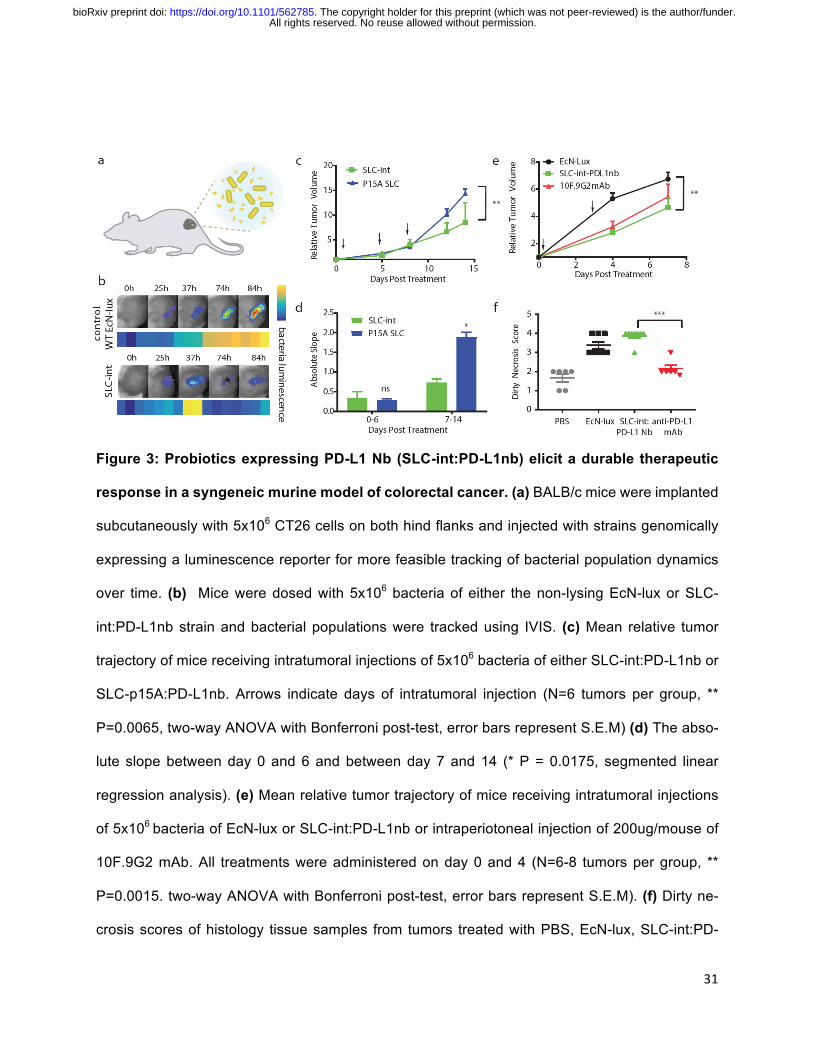
Figure 3: Probiotics expressing PD-L1 Nb (SLC-int:PD-L1nb) elicit a durable therapeutic
response in a syngeneic murine model of colorectal cancer. (a) BALB/c mice were implanted
subcutaneously with 5x106 CT26 cells on both hind flanks and injected with strains genomically
expressing a luminescence reporter for more feasible tracking of bacterial population dynamics
over time. (b) Mice were dosed with 5x106 bacteria of either the non-lysing EcN-lux or SLC-
int:PD-L1nb strain and bacterial populations were tracked using IVIS. (c) Mean relative tumor
trajectory of mice receiving intratumoral injections of 5x106 bacteria of either SLC-int:PD-L1nb or
SLC-p15A:PD-L1nb. Arrows indicate days of intratumoral injection (N=6 tumors per group, **
P=0.0065, two-way ANOVA with Bonferroni post-test, error bars represent S.E.M) (d) The abso-
lute slope between day 0 and 6 and between day 7 and 14 (* P = 0.0175, segmented linear
regression analysis). (e) Mean relative tumor trajectory of mice receiving intratumoral injections
of 5x106 bacteria of EcN-lux or SLC-int:PD-L1nb or intraperiotoneal injection of 200ug/mouse of
10F.9G2 mAb. All treatments were administered on day 0 and 4 (N=6-8 tumors per group, **
P=0.0015. two-way ANOVA with Bonferroni post-test, error bars represent S.E.M). (f) Dirty ne-
crosis scores of histology tissue samples from tumors treated with PBS, EcN-lux, SLC-int:PD-
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

32
L1nb, and anti-PD-L1 mAb (N=6-8 scores per group of biological replicates, ***P=0.007 Mann
Whitney Test ordinal non parametric between SLC-int:PD-L1nb and anti-PD-L1 mAb, error bars
represent S.E.M).
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

33
Figure 4: Combination therapy of SLC-int:probiotics producing anti-PD-L1 or anti-CTLA-4
Nbs enhances therapeutic response characterized by a decrease in Treg cells and an
increase in proliferative conventional CD4+ cells. (a) BALB/c mice were implanted
subcutaneously with 5x106 CT26 cells on both hind flanks. When tumors reached ~200mm3, mice
received intratumoral injections every 3-4 days of either EcN-lux, SLC-int:CTLA-4nb, SLC-int:PD-
L1nb, or an equal parts Nb cocktail. Mean absolute tumor trajectory and individual trajectories of
mice (N=6-8 tumors per group, , **** P<0.0001, two way-ANOVA with Bonferroni post-test, error
bars represent S.E.M). (b) BALB/c mice were implanted subcutaneously with 5x106 A20 cells on
both hind flanks. When tumors reached ~150mm3 mice received intratumoral injections every 3-
4 days of either EcN-lux, SLC-int:CTLA-4nb, SLC-int:PD-L1nb, or an equal parts Nb cocktail.
Mean absolute tumor trajectory and individual trajectories of mice (N=4-6, *** P = 0.002, two-way
ANOVA, Bonferroni post-test). (c–d) Tumor-infiltrating lymphocytes were isolated on day 8 after
initial treatment and analyzed by flow cytometry for frequencies of (c) CD4+FOXP3+ regulatory T
cells (N=3, unpaired t test between PBS and Nb cocktail, ** P=0.0013) and (d) proliferating
a
0 5 10 15
200
400
600
800
Days Post Treatment
Abso
lute
Tum
orVo
lum
e(m
m3 )
EcN-lux
SLC-int PDL1 NbSLC-int CTLA4 Nb
Nb Cocktail
Abso
lute
Tum
orVo
lum
e(m
m3)CT26 model
0 5 10 150
500
1000
1500
2000
2500b
Abso
lute
Tum
orVo
lum
e(m
m3 )
Days Post Treatment
EcN-lux
SLC-int PDL1 NbSLC-int CTLA4 Nb
Nb Cocktail
A20 modelAb
solu
teTu
mor
Volu
me
(mm
3)
0 5 10 150
500
1000
Days Post Treatment
0 5 10 15 0 5 10 15 0 5 10 15
0
1000
2000
3000
Days Post Treatment
CT26 model
A20 model
PBS0
20
40
60
Freq
uenc
yof
CD4+
FOXP
3+ i
nTu
mor
**
0
5
10
15
20
Freq
uenc
yof
CD
4+ F
OXP
3-Ki
67+
inTu
mor
EcN-lu
x
CTLAA-4 N
b
PD-L1 N
b
Nb Cocktail
0 5 10 15 0 5 10 15 0 5 10 15 0 5 10 15
PBS
EcN-lu
x
CTLAA-4 N
b
PD-L1 N
b
Nb Cocktail
c
d
Figure 4, Gurbatri et al.
****
**
***
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

34
CD4+FOXP3–Ki67+ conventional T cells (N=2-3, unpaired t test between PBS and Nb cocktail, **
P = 0.0013). All error bars represent S.E.M.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

35
SUPPLEMENTARY MATERIALS
Figure S1: Engineering and characterization of the ptac: PD-L1 Nb plasmid. (a) Plasmid map
for the constitutive expression of the PD-L1 Nb on a high copy plasmid. (b) Immunofluorescence
microscopy image of CT26 cells (purple: PD-L1, blue: nuclei). Left panel shows CT26 cells stained
for PD-L1 expression with a flourescently-tagged 10F.9G2 mAb and its respective isotype. Right
panel shows CT26 incubated with bacterial lysate containing PD-L1 Nb where the PD-L1 Nb is
probed for using an anti-HA mAb.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint
36
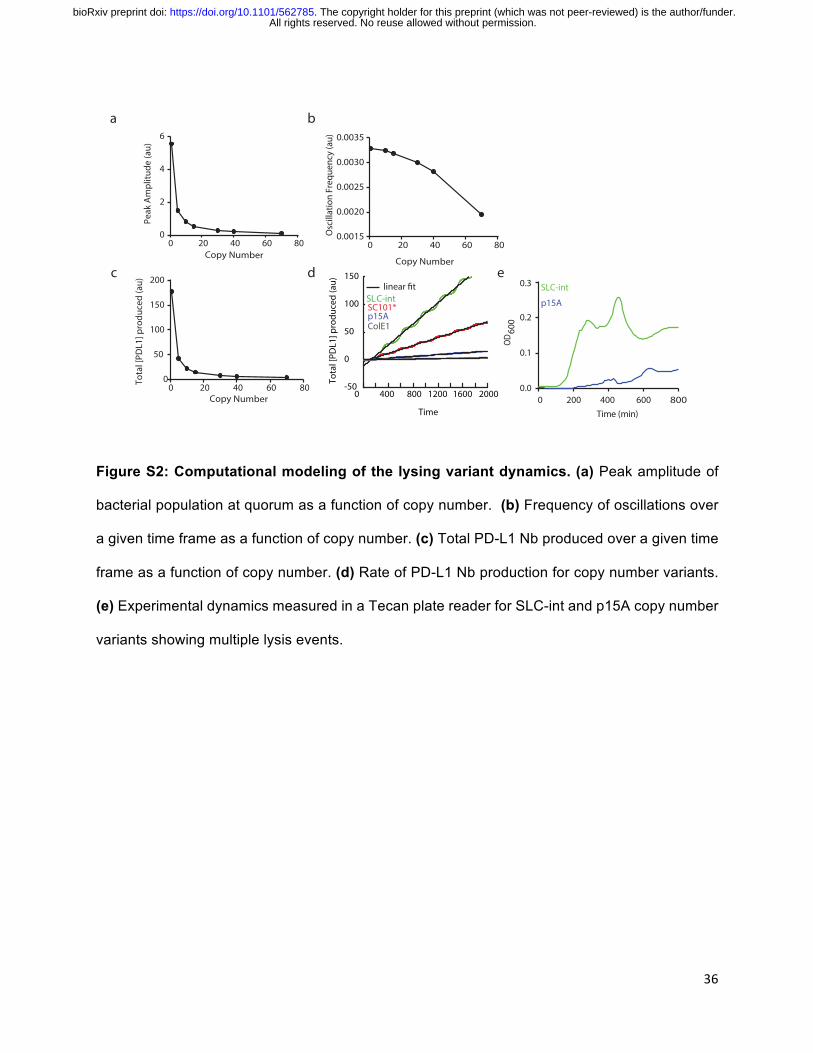
Figure S2: Computational modeling of the lysing variant dynamics. (a) Peak amplitude of
bacterial population at quorum as a function of copy number. (b) Frequency of oscillations over
a given time frame as a function of copy number. (c) Total PD-L1 Nb produced over a given time
frame as a function of copy number. (d) Rate of PD-L1 Nb production for copy number variants.
(e) Experimental dynamics measured in a Tecan plate reader for SLC-int and p15A copy number
variants showing multiple lysis events.
a
Peak
Am
plitu
de (a
u)
b
c
0 20 40 60 800
2
4
6
0 20 40 60 800
50
100
150
200
Tota
l [PD
L1] p
rodu
ced
(au)
e
0 200 400 600 8000.0
0.1
0.2
0.3
Time (min)
OD60
0
SLC-int
p15A
Copy Number
Copy Number
d
0 20 40 60 800.0015
0.0020
0.0025
0.0030
0.0035
Osc
illat
ion
Freq
uenc
y (a
u)
Copy Number
Time
0 400 800 1200 1600 2000
Tota
l [PD
L1] p
rodu
ced
(au)
-50
0
50
100
150
SLC-intSC101*p15AColE1
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

37
Figure S3: Characterization of lysing circuit variants in a syngeneic colorectal model. (a)
Western blot showing PD-L1 Nb protein levels collected from the supernatant of EcN-lux, EcN-
lux:PD-L1nb (non-lysing), and SLC-int:PD-L1nb strains. (b) Individual trajectories of mean tumor
volume for CT26 tumor bearing BALB/c mice receiving injections of the SLC-int:PD-L1nb or SLC-
p15A: PD-L1nb variant on day 0, 4 and 7 post treatment. (c) Segmented linear regression (dotted
line) analysis of the relative tumor volume from 0-6 and 7-14 days. For days 0-6, linear regression
fit R2 = 0.85 and 0.99 for SLC-int:PD-L1nb and SLC-p15A:PD-L1nb groups respectively. For days
7-14, linear regression fit R2 = 0.99 and 0.99 for SLC-int:PD-L1nb and SLC-p15A:PD-L1nb groups
respectively. N=6 per group, *P = 0.0175, error bars represent S.E.M.
a
EcN-lux Nonlysing SLC-int
0
5
10
15
20
Days Post Treatment
Rela
tive
Tum
orVo
lum
e
b
0 5 10 15 0 5 10 150
5
10
15
20
Rela
tive
Tum
orVo
lum
e
ns
*
c
Days Post Treatment
SLC-int: PD-L1nbSLC-p15A: PD-L1nb
SLC-int: PD-L1nbSLC-p15A: PD-L1nb
~15kDa
IB:H
A
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

38
Figure S4: Individual tumor trajectories for comparison between probiotic expressing PD-
L1nb (SLC-int:PD-L1nb) and systemically delivered antibody in CT26 model. (a) Plot show-
ing distribution of tumor sizes in CT26 model. (b) Individual relative tumor volumes for CT26 tumor
bearing BALB/c mice injected with EcN-lux, SLC-int:PD-L1 Nb, and 10F.9G2 mAb. This experi-
ment corresponds to Figure 3e in the text.
0
500
1000
1500
0 4 7Abso
lute
Tum
or V
olum
e (m
m3 )
Days Post Treatment
aEcN-l uxSLC-int: PD-L1nbanti-PD-L1 mAb
CT26 model
2
4
6
8
10
Rela
tive
Tum
orVo
lum
e
Days Post Treatment
0 4 6 82 0 4 6 82 0 4 6 82
b
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

39
Figure S5: Histological images and dirty necrosis scoring in CT26 mouse tumors. Main
panels are 40x, inserts are 600x of the approximate areas of boxed regions in low magnification
images. VT is viable tumor; Circles in insets are bacterial colonies; arrows degenerate neutrophils
(i.e. dirty necrosis). (A) PBS, (B) 10F.9G2 mAb, (C) EcN-lux, (D) SLC-int:PD-L1-nb. This exper-
iment corresponds to Figure 3e in the text.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

40
Figure S6: Comparison between probiotically-produced PD-L1 Nb and systemically deliv-
ered antibody in 4T1 model. BALB/c mice were implanted subcutaneously with 5x106 4T1 cells
on both hind flanks. When tumors reached ~200mm3, mice received intratumoral injections of
5x106 bacteria every 3-4 days of either EcN-lux, SLC-PD-L1nb, or 200ug/mouse of 10F.9G2 mAb
(a) Survival of mice following 5x106 bacteria dosed on days 0, 5, and 8 days post treatment, and
systemically delivery mAb dosed on days 0, 5, 8 and 12. Severe toxicities in weight and body
conditions of antibody treated mice were observed, leading to death or required euthanasia of all
mice within two weeks. (b) Mean relative body weight. (c) Distribution of tumor sizes. N=6-8 per
group, *P = 0.0199, two-way ANOVA with Bonferroni post-test, error bars represent S.E.M.
0 5 10 150.7
0.8
0.9
1.0
1.1
Days Post Treatment
Rela
tive
Body
Wei
ght
0 5 10 150
50
100
Days Post Treatment
Perc
ents
urvi
val
0
200
400
600
Days Post Treatment0 5 8 12Ab
solu
te T
umor
Vol
ume
(mm
3 )
a b c 4T1 model*EcN-lux
SLC-PDL1nb
EcN-luxSLC-PD-L1nbanti-PD-L1 mAb
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

41
Figure S7: SLC-int:PD-l1nb and bacterially-expressed hemolysin exotoxin (hlyE) combina-
tion therapy is more efficacious than individual therapies. BALB/c mice were implanted sub-
cutaneously with 5x106 CT26 cells on both hind flanks. When tumors reached ~200mm3, mice
received intratumoral injections of 5x106 bacteria every 3-4 days of either EcN-lux, EcN-SLC-
hlyE, SLC-int:PD-L1nb, or an equal parts cocktail of EcN-SLC-hlyE and SLC-int:PD-L1nb strains.
(a) Mean relative tumor trajectory of mice. (b) Distribution of tumor sizes. (c) Individual trajectories
of relative tumor volumes for each treatment group. N=6-9 tumors per group, *** P=0.0004, two-
way ANOVA with Bonferroni post-test, error bars represent S.E.M. (d) Mean relative body weight.
0 5 10 151
2
3
4
Days Post Treatment
Rela
tive
Tum
orVo
lum
e
***
SLC-hlyESLC-int:PD-L1nb
EcN-lux
PDL1 Nb + hlyE
a
0
200
400
600
0 4 7 11Days Post Treatment
b
Abso
lute
Tum
or V
olum
e (m
m3 )
0 5 10 150.9
1.0
1.1
1.2
1.3
Rela
tive
Body
Wei
ght
Days Post Treatment
d
1
2
3
4
5
Rela
tive
Tum
orVo
lum
e
0 5 10 15 0 5 10 15Days Post Treatment
c
0 5 10 150 5 10 15
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

42
Figure S8: Engineered probiotic expressing CTLA-4 Nb. (a) Plasmid map for the constitutive
expression of the CTLA-4 Nb on a high copy plasmid. (b) BALB/c mice were implanted subcuta-
neously with 5x106 CT26 cells on both hind flanks. When tumors reached ~200mm3, mice re-
ceived intratumoral injections every 3-4 days of 5x106 bacteria of either EcN-lux, SLC-int:CTLA-
4nb or 100ug/mouse intraperitoneal injections of a 9D9 mAb. Graph shows mean relative tumor
trajectories. (c) Mean relative body weight. N=6-7 per group, *P=0.0407, two-way ANOVA with
Bonferroni post-test, error bars represent S.E.M. (d) Cytokine levels of tumor lysates measured
by Luminex Multiplex Assay. N=3 biological replicates per group, *P=0.0224, one-way ANOVA
with Bonferroni post-test, error bars represent S.E.M.
b
0 5 10 150.9
1.0
1.1
1.2
1.3
1.4
Days Post Treatment
Rela
tive
Body
Wei
ght
0 5 10 151
2
3
4
5
6
Days Post Treatment
Rela
tive
Tum
oroV
lum
e EcN-lux9D9 Ab *
d
Axe/Txe
HA tag
KanR
pBR3222anti-CTLA4
a
c
ExN
-Lux
SLC-
int-
CTLA
4-N
b
9D9
Ab
*
ptac:CTLA-4 Nb-HASLC-int:CTLA-4nb
0
10
20
30
IFN
-Y(p
g/m
l)
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint
43
Figure S9: Relative body weight for combination of PD-L1 and CTLA-4 probiotics. Mean
relative body weight in a syngeneic (a) CT26 and (b) A20 mouse model. This experiment corre-
sponds to Figure 4 in the text.
0 5 10 150.9
1.0
1.1
1.2
1.3
1.4
Days Post Treatment
Rela
tive
Body
Wei
ght
a
0 5 10 150.7
0.8
0.9
1.0
1.1
Rela
tive
Body
Wei
ght
Days Post Treatment
bCT26 model A20 modelEcN-lux
SLC-int: PD-L1nbSLC-int: CTLA-4nb
Nb Cocktail
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

44
Mathematical Model
We built upon the ordinary differential equation model explored in the original sycnhrionized lysis
circuit system (19). The following set of ordinary differential equations were derived to describe
the bacterial population number (N, equation[1]), total extracellular AHL (H, equation[2]),
intracellular concentrations of the lysis protein E (L, equation[3]), intracellular concentrations of
the LuxI protein (I, equation[4]), and intracellular concentrations of the PD-L1 Nb protein (P,
equation[5]). In this system, bacteria grow logistically at a rate of µN to a maximum capacity, N0
and lyse once at quorum. AHL is produced at a rate proportional to bacterial population and is
cleared out at a rate of µ. The lysis protein is produced at a rate proportional to copy number and
has a basal degradation (γL) and is also diluted as the bacterial population grows (µg). The luxI
protein has similar dynamics, but is further degraded by ClpXP machinery (γc). The therapeutic
protein production is proportional to bacteria number and has basal degradation (γp) and is diluted
as the bacterial population grows (µg). Internal production of LuxI and lysis protein is described
by Plux (equation[6]) and the rate of cell degradation due to lysis is described by the hill function,
γN (equation[7]).
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

45
Model Parameters Values:
To explore the dynamics of our library of lysing variants, we iterated through a large range of copy
numbers, CI and CL ranging from 1 to 100. Growth rate (µN). was experimentally-derived and the
doubling time of ~30 minutes for SLC-int:PD-L1nb was used to calculate a growth rate of
0.023min-1 for in vitro plots (Fig. 2b, c, Fig. S2a-d) using the doubling time formula. Based on
previous literature (24), the doubling time of bacteria increases to ~4 hours, which yielded a
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint

46
growth rate of 0.00289min-1 (Fig. 2d, e). Since growth rate and dilution due to cell growth was
assumed to be equal µg was set equal to µN. Other parameter values include: α0 = 0.1, αH = 50,
h0=5, b=20, µg = 1.5, L0 = 8, n=2, m=4. γL = 1.5, K =0.05, γI = 5, γC = 12, N0=10.
Movie S1: Visualization of SLC-int lysis. The EcN-lux:SLC-int strain was transformed with a
ptac:sfGFP plasmid. Bacteria were seeded onto a 96 well glass bottom plate and timelapse was
used to capture GFP fluorescence change over time at 10X magnification.
All rights reserved. No reuse allowed without permission. The copyright holder for this preprint (which was not peer-reviewed) is the author/funder.. https://doi.org/10.1101/562785doi: bioRxiv preprint