energy storage in cells - vyuka-data.lf3.cuni.czvyuka-data.lf3.cuni.cz/cvse1m0001/energy storage in...
TRANSCRIPT

Energy storage in cells
Josef Fontana
EC - 58

Overview of the lecture
• Introduction to the storage substances of human body
– Overview of storage compounds in the body
• Glycogen metabolism
– Structure of glycogen
– Synthesis and degradation of glycogen
– Phosphorylation and dephosphorylation as a regulatory
mechanism of the glycogen metabolism
• Synthesis of fatty acids and TAG
– Differences between synthesis and degradation of fatty acids
– How works the fatty acid synthase
– Elongation and desaturation of fatty acids
– Synthesis of TAG

Introduction to the storage
substances of human body
Overview of storage
compounds in the body

Overview of storage compounds
in the body • TAG
• Glycogen
• No storage protein
• TAG are excelent for energy storage - 1g of fat
has 6 times more energy than 1g of
hydrated glycogen
• Complete oxidation of 1g of FA = 38 kJ
• Complete oxidation of 1g of saccharides or
proteins only 17 kJ

Overview of storage compounds
in the body • 70 kg man has:
• 1) 420 000 kJ in TAG
• 2) 10 000 kJ in proteins (muscle)
• 3) 2 500 kJ in glykogen
• 4) 170 kJ in glucose
• Glycogen and glucose are sufficient to
supply the body one day, TAG many
weeks

Glycogen metabolism
Structure of glycogen

Glycogen
• Animal saccharide storage
• In liver (100g), skeletal muscle
(500g) and in small quantities in each
cell
• 1) liver glycogen: to maintain glycemia
• 2) muscle glykogen: for internal
muscle use

Glycogen structure
• Branched
homopolymer
• Most residues bound
by α 1→4 bonds
• Branching: α 1→6
bond
• These branches are
extended by α 1→4
bond

Glycogen has two ends
• Only on the non-reducing ends can
take place reactions (lengthening or
shortening)
• Reducing end is the one with the
hemiacetal hydroxy group - bound to
tyrosine in glycogenin

Glycogen metabolism
Synthesis and degradation of
glycogen

Glycogenesis (glycogen synthesis)
• Cytosol
• Glucose phosphorylation to Glc-6-P:
glucokinase in liver and hexokinase in muscle
• Isomeration of Glc-6-P → Glc-1-P:
phosphoglucomutase
• Glc-1-P reacts with UTP → UDP-Glc (activated
Glc, bond on C1): Glc-1-P uridylyltransferase
• UDP-Glc is bound to the non-reducing end of
glycogen: glycogen synthase

Glycogen synthase
• Binds UDP-Glc to the non-reducing
end of glycogen
• UDP is released
• Chain of glucose molecules
lengthens, until it reaches a certain
length and branching occurs

Branching enzyme
• Removes oligosaccharide (6-7 Glc
residues) from growing chain and adds it to
a hydroxy group on the C6 in Glc
• Forms α 1→ 6 bond
• These branches are extended by glycogen
synthase
• Branching enzyme = amylo-(1,4 – 1,6)-
transglycosylase

Regulation of glycogen synthesis
• Glycogen synthase is regulated by
phoshorylation:
– phosphorylation inactivates
– dephosphorylation activates
• Insulin activates
• Glucagon and adrenaline inhibit

Glycogenolysis
• Cytosol
• 1) Phosphorolytic cleavage
(inorganic phosphate is used):
glycogen phosphorylase – Glc-1-P
(Cori ester)
• 2) Isomeration of Glc-1-P to Glc-6-P:
phosphoglukomutase

Cutting branches off
• Degradation of glycogen stops at the 4th Glc
before the branching point
• Glucanotransferase (glycosyltransferase)
transfers three glucose residues from the 4-
residue glycogen branch to the main chain
• Only one glucose molecule remains (α 1→6
bond) – cleaved by debranching enzyme
(amylo-α1→6-glucosidase)
• Linear glycogen chain – glycogen
phosphorylase

Regulation of glycogenolysis
• Glycogen phosphorylase is
activated phosphorylated
• Phosphorylase kinase
• Insulin inhibits
• Counter-regulatory hormones
activate

Synthesis of fatty acids and TAG
Differences between synthesis
and degradation of fatty acids

Differences between synthesis and
degradation of fatty acids
• FA synthesis in cytosol, degradation in
matrix
• Intermediates of FA synthesis are bound
to ACP (acyl carrier protein),
intermediates of degradation bound to
CoA
• Enzymes of FA synthesis form one big
multienzyme complex - Synthase of FA,
degradation enzymes are free in matrix

Differences between synthesis and
degradation of fatty acids
• FA chain is extended by 2 carbon
atoms from AcCoA – activated
substrate is malonyl~CoA
• Reducing cofactor for synthesis is
NADPH, oxidising cofactors for
degradation are FAD and NAD+

Differences between synthesis and
degradation of fatty acids
• FA synthesis (on FA synthase) ends
with palmitate (C16)
• Further chain elongation and
formation of unsaturated acids
catalyse other enzymes

Synthesis of fatty acids and TAG
Synthesis of malonyl~CoA

Synthesis of malonyl~CoA
• Substrate for FA synthesis: AcCoA
• Carboxylation to malonyl-CoA
AcCoA + ATP + HCO3- → malonyl~CoA + ADP
+ Pi + H+
• AcCoA carboxylase (biotin – vitamin H or B7)
• Regulatory enzyme
• CO2 removed during condensation with
growing FA

Synthesis of fatty acids and TAG
How works the fatty acid
synthase

Mammalian fatty acid synthase
• Homodimer of 2 identical subunits (260 kDa)
• Each subunit has three domains connected by
moving regions:
• 1) domain 1 – substrates entry and condensing unit -
both transferases and condensing enzyme - CE
• 2) domain 2 – reduction unit – includes ACP, β-
ketoacylreduktase, dehydratase and enoylreductase
• 3) domain 3 – thioesterase cleaves palmitate

Mammalian fatty acid synthase
• Places where are bound
intermediates on FA synthase:
• 1) thiol group of cysteine (CE)
• 2) thiol group of
phosphopantetheine (bound to
serine in ACP)

Steps of FA synthesis
1. Synthesis of malonyl-CoA: acetyl-CoA
carboxylase
2. Reaction AcCoA + CE: acetyltransacylase
3. Reaction malonyl-CoA + ACP:
malonyltransacylase
4. Condensation reaction: condensing enzyme
Acetyl-CE + malonyl-ACP → acetoacetyl-ACP +
CE + CO2

Steps of FA synthesis
5. First reduction: β-ketoacylreductase
Acetoacetyl-ACP + NADPH + H+ → D-3-
hydroxybutyryl-ACP + NADP+
6. Dehydration: 3-hydroxyacyldehydratase
D-3-Hydroxybutyryl-ACP → crotonyl-ACP + H2O
7. Second reduction: enoylreductase
Crotonyl-ACP + NADPH + H+ → butyryl-ACP +
NADP+

FA synthase works as a dimer
• Condensation
between
malonyl-ACP
(one subunit)
and acetyl-CE
(second
subunit)
• New acyl
remains on
ACP
CO2
KONDENZACE-
S
C
CH3
O
SH
CE ACP
SHS
C
CH2
CO
O
O
CEACP
SHSH
CE ACP
SHS
C
CH2
O
C
CH3
O
CEACP
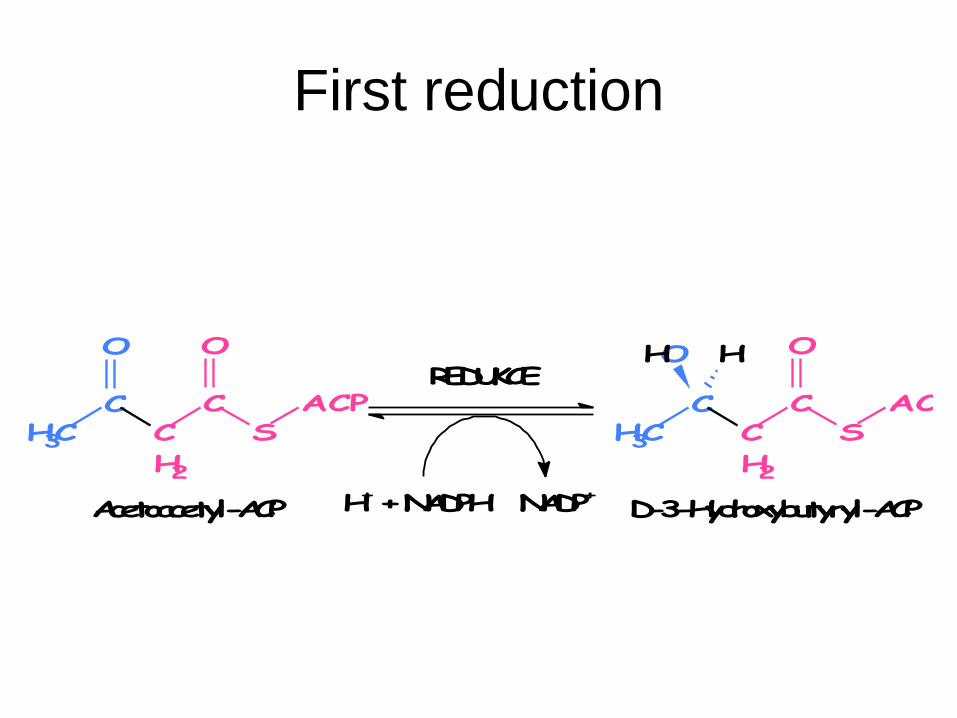
First reduction
D-3-Hydroxybutyryl-ACP
ACPS
CC
H2
O
CCH3
O
Acetoacetyl-ACP
REDUKCE
H+ + NADPH NADP+
ACPS
CC
H2
O
CCH3
OH H
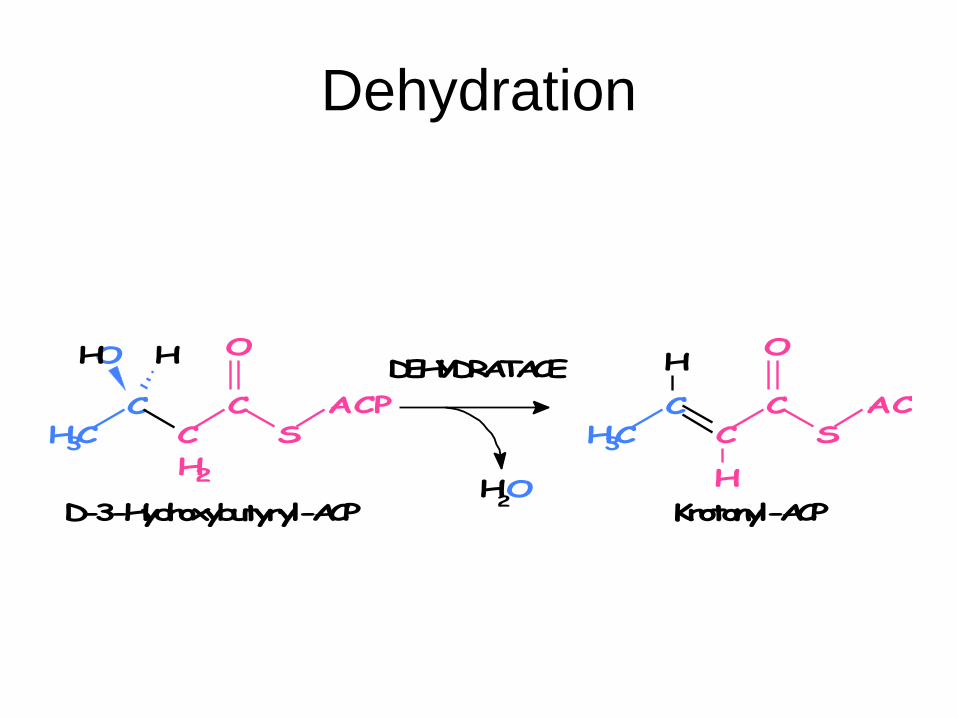
Dehydration
H2O
ACPS
CC
O
CCH3
H
H
Krotonyl-ACP
DEHYDRATACE
D-3-Hydroxybutyryl-ACP
ACPS
CC
H2
O
CCH3
OH H
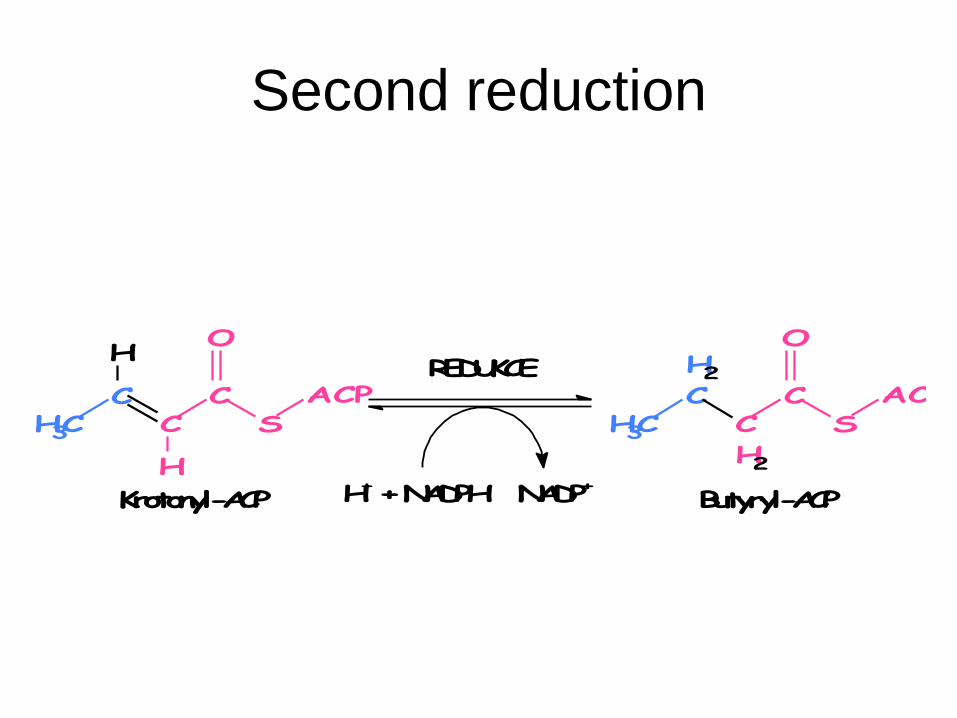
Second reduction
REDUKCE
H+ + NADPH NADP+
ACPS
CC
O
CCH3
H
H
Krotonyl-ACP
ACPS
CC
H2
O
C
H2
CH3
Butyryl-ACP

Process continues
• Change of subunits after one rotation
• Palmitate (C16) is an end product
• Thioesterase cleaves palmitate
from ACP - hydrolysis of the thioester
bond with phosphopantetheine

Palmitate synthesis requires
• 8 AcCoA, 14 NADPH a 7 ATP
• AcCoA produced in matrix – inner
mitochondrial membrane is
impermeable – transport via citrate
• 8 NADPH from the citrate transport
to cytosol and remaining 6 NADPH
in pentose cycle
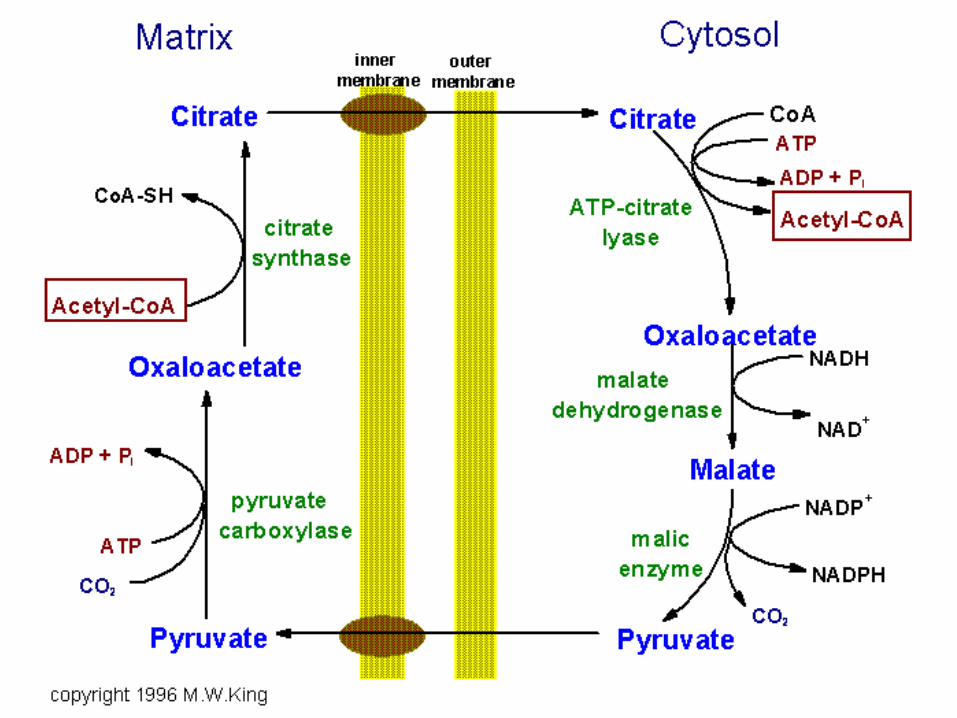

Citrate as AcCoA bearer
• High level of citrate in matrix – transport
to cytosol – cleavage by ATP-citrate
lyase:
Citrate + ATP + HSCoA + H2O → AcCoA
+ ADP + Pi + OAA
• AcCoA and OAA have different fate in
cytosol

OAA returns to matrix
• Inner mitochondrial membrane is
impermeable to OAA
• Reduction of OAA to malate by cytosolic
malate dehydrogenase:
OAA + NADH + H+ → malate + NAD+
• Oxidative decarboxylation of malate by
NADP+-malate enzyme (malic enzyme):
Malate + NADP+ → Pyr + CO2 + NADPH

OAA returns to matrix
• Pyruvate transport to matrix –
carboxylation by pyruvate carboxylase:
Pyr + CO2 + ATP + H2O → OAA + ADP
+ Pi + 2 H+
• Summary equation:
NADP+ + NADH + ATP + H2O →
NADPH + NAD+ + ADP + Pi + H+

Regulation of FA synthesis
• Enough substrates (saccharides/AA) and energy
• AcCoA-carboxylase:
• 1) insulin activates
• 2) glucagon and epinephrine inhibit
• 3) citrate activates
• 4) inhibition by palmitoyl-CoA – feedback
inhibition
• 5) AMP inhibits

Synthesis of fatty acids and TAG
Elongation and desaturation of
fatty acids

Synthesis of other fatty acids
• Chain elongation – elongases
• Synthesis of unsaturated FA –
desaturation – desaturases
• ER membrane

Desaturation
• Mammals lack enzymes catalyzing
formation of the double bond further than
on C9
• New double bonds are always formed
between the existing double bond and a
carboxyl group
• Mammals can not synthesize linoleic (18 : 2
cis D9, D12) and linolenic (18 : 3 cis D9, D12,
D15) acid – both eare essential

Synthesis of fatty acids and TAG
Synthesis of TAG

Synthesis of TAG