endpoints and new targets for curing hepatitis...
TRANSCRIPT

Endpoints and New Targets for
Curing Hepatitis BProfessor Stephen Locarnini
WHO Regional Reference Laboratory for Hepatitis BVictorian Infectious Diseases Reference Laboratory,
Doherty InstituteMelbourne, Victoria 3000, AUSTRALIA
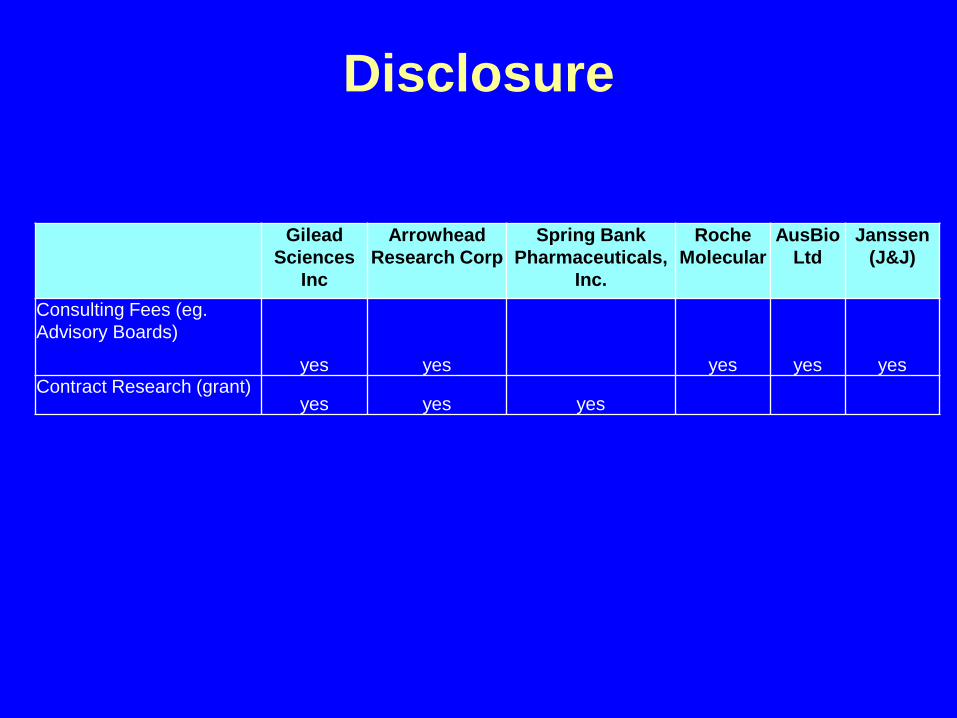
Disclosure
Gilead Sciences
Inc
Arrowhead Research Corp
Spring Bank Pharmaceuticals,
Inc.
Roche Molecular
AusBioLtd
Janssen (J&J)
Consulting Fees (eg. Advisory Boards)
yes yes yes yes yesContract Research (grant)
yes yes yes

Types of Chronic HBV Control and Cure
Peters, MG & Locarnini, S. 2016. Gastroenterol and Hepatol;13(6):348-356.
Inactive StateSustained, off drug:• No inflammation: normal ALT and liver biopsy• HBV DNA low or undetectable• HBsAg-positiveFunctional Cure (Clinical Resolution)Sustained, off drug:• No inflammation: normal ALT and liver biopsy• HBsAg loss• Anti-HBs gainComplete Cure (Virologic Cure)• All of the above plus• Loss of cccDNA in the liver

New Viral Targets
1. Entry and Pathways of Intrahepatic Spread (Re-Entry)
2. Cytosolic Transport of Nucleocapsids3. cccDNA Generation & Processing
(HBcAg and HBx)4. HBV Transcription5. HBV Nucleocapsid Assembly (HBcAg)6. HBsAg7. Putting It All Together

Electron Microscopy: HBV in Serum

Wang, J et al 2016. J Hepatol;65:700-710
1. Entry & Re-Entry and HBV
• HBV WT• HBV SPLICE VARIANTS• HBV FL RNA

HBx RNA Was Detected Very Early After HBV Infection
Beran, R et al 2017. EASL
*Mock infection. LHB, large HBV surface protein; d, day; h, hour; MHB, middle HBV surface protein; ORF, open reading frame; SHB, small HBV surface protein.
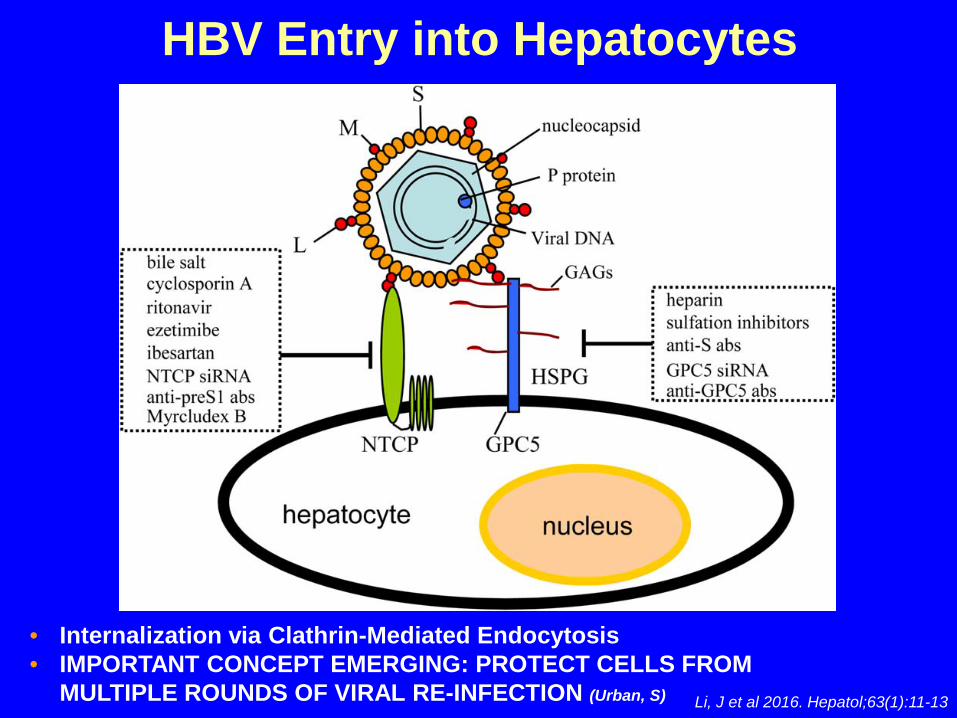
Li, J et al 2016. Hepatol;63(1):11-13
HBV Entry into Hepatocytes
• Internalization via Clathrin-Mediated Endocytosis• IMPORTANT CONCEPT EMERGING: PROTECT CELLS FROM
MULTIPLE ROUNDS OF VIRAL RE-INFECTION (Urban, S)

2. Cytosolic Transport of NucleocapsidsA. Primary infection
– Mature nucleocapsids transported via importin α/β whilst nucleoporin 153 delivers nucleocapsids to nuclear basket where they uncoat at nuclear pore
– Genomic RC DNA enters nucleoplasm with Pol covalently attached and is then converted to PF-RC DNA
– (?) HBV RNA species eg: HBx mRNA released and translated
B. Persistent infection– Intracellular conversion pathway provides low level recycling
of nucleocapsids- replication complexes from hepatocyte cytoplasm
– Multiple rounds of re-infection replenish key intermediates
TARGET: NUCLEOCAPSID DISASSEMBLY WITH CpAM COMPOUNDS: BLOCK cccDNA GENERATION

Key Step 2: Reverse Transcription
Host Enzymes
Covalently Closed Circular
(ccc) DNAHBV RC Genomic DNA
(-) (+)
HBV DNA Polymerase Host RNA
Polymerase II
HBV Reverse Transcriptase
HBV Minus (-) DNA
(-)
Pregenomic HBV (pg) RNA
AAAA
3. Key Step 1: Conversion of RC DNA into cccDNA
1. Removal of RT2. Removal of r3. Ligation of (-) DNA4. Completion of (+) DNA5. Removal of capped RNA6. Ligation of (+) DNA

Bock, T. et al 1994. Virus Genes;8:215
Bock, T. et al 2001. JMB;307:183
Newbold, J and Locarnini, S 1995. J. Virol;69:3350
activated nucleosomal spacing
High Replication Phenotype
Transcriptionally ActiveHigh Viraemia
repressive nucleosomal spacing
Low Replication Phenotype
Quiescent or activeMedium to Low Viraemia
HBV cccDNA is a Minichromosome
“OPEN”
“CLOSED”

HBcAg and HBx are key Components of the HBV Minichromosomes
Bock, T. et al 2001. JMB;307:183.;Belloni, L et al 2009. PNAS;106:19975-19979.;
Guo, YH et al 2011. Epigentics;6:720.;Levrero, M & Zucman-Rossi, J. 2016. J Hepatol; 64(1 Suppl):S84-S101
HBx blocks methylation complexHBcAg binds HBV DNA (CpG Island II)HBx knock-out result in transcriptional arrest of MC

HBx Contributes to Establish an Active cccDNA Chromatin State
Alarcon, V et al 2015. Sci Rep;6:1-11
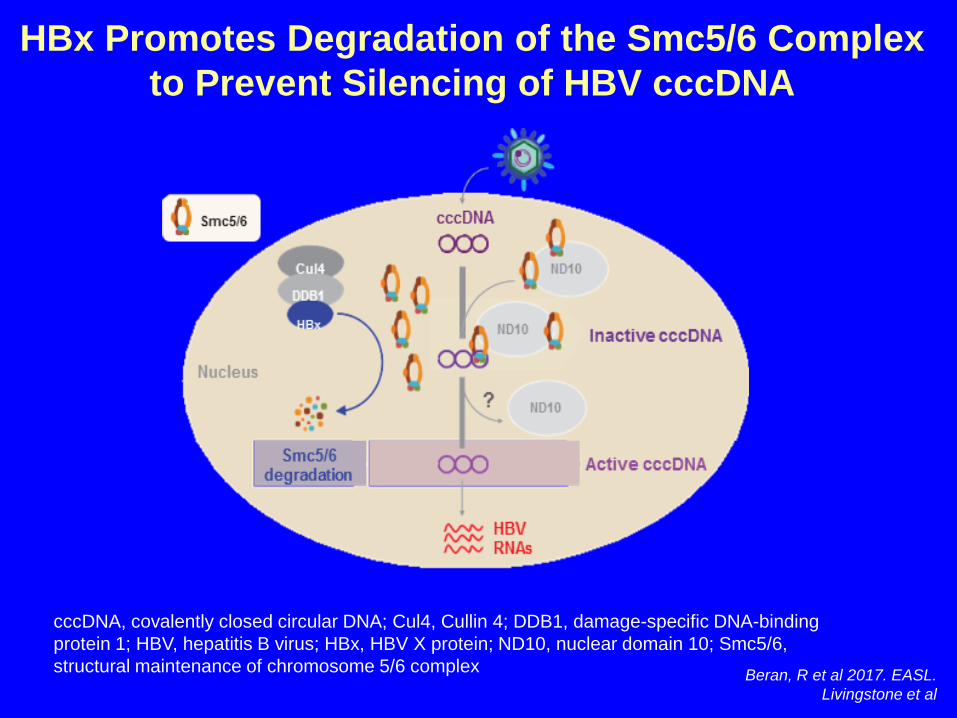
HBx Promotes Degradation of the Smc5/6 Complex to Prevent Silencing of HBV cccDNA
cccDNA, covalently closed circular DNA; Cul4, Cullin 4; DDB1, damage-specific DNA-binding protein 1; HBV, hepatitis B virus; HBx, HBV X protein; ND10, nuclear domain 10; Smc5/6, structural maintenance of chromosome 5/6 complex Beran, R et al 2017. EASL.
Livingstone et al

HBx Inhibitors: To Silence Minichromosome
• Crystal structure– CUL4A-DDB1 ubiquitin ligase complex
interacts with HBx
• Small molecule inhibitors– in development

HBcAg and Transcription: Preferential Binding of HBcAg to HBV Minichromosome
Bock, CT et al 2001. J Mol Biol;307:183
A B
• HBcAg binds to CpG Island II (Guo, YH et al 2011. Epigenetics;6:720)
• in the presence of HBcAg peak number of nucleosomes increased from 15 to 16, resulting in a 20bp decrease in nucelosomal spacing
• HBc enhances upstream NK-κβ binding thereby promoting activation of Enh II/pregenomic promoter (Kwon, JA & Rho HM. 2002. Biochem Cell Biol;80:445)

Interaction of APOBEC 3A/3B, HBV Core Protein (HBc) and cccDNA
Lucifora, J et al 2014. Science;343(6176):1221-8

Targeting HBV Nucleocapsids
Antimicrob Agents Chemother. 2002 Sep;46(9):3057-60. Yuen M-F, et al. AASLD 2015, San Francisco. #LB-10
Deres et al, Science 2003 Klumpp et al, PNAS 2015
#2 Heteroaryldihydropyrimidines
Destabilization of nucleocapsids
#1 Novel classes of capsid inhibitors based on the 3D structure of HBcNovira, Assembly Biosciences, Janssen, Roche, and othersPhase 1/2 studies with Novira compound (NVR3-778) completed
Lam A, et al. AASLD 2015, San Francisco. #33
[sulphonamide/sulfamoyl benzimide derivatives]

How Can cccDNA be Cleared? An Immunological Perspective
Fourel, I et al 1994. J Virol;68:8321.; Guo, JT et al 2000. J Virol;74:1495.Thimme, R et al 2003. J Virol;77:68.; Guidotti, L et al 1999. Science;284:825
CytolyticNon-Cytolytic

How Can cccDNA be Cleared? A Virological Perspective
• directly purge all of the pre-existing cccDNA [topoisomers] (Summers, J and Mason, WS. 2004. Proc Natl Acad Sci (USA);101(2):638-640)
OR• permanently silence cccDNA (condensed
chromatin) transcription (Locarnini, S and Newbold, J. 1997. J Antimicrob Chemoth;39:559-562)
• “cccDNA targeting” small molecule antivirals. eg: di-substituted sulfonamides(Cai, D et al 2012. Antimicrob Agents Chemoth;56:4277)

Direct cccDNA Destruction: Molecular Based Therapies
• cleave cccDNA moleculesVIAcccDNA sequence-specific endonuclease using
– zinc-finger nucleases (Hoeksema, KA & Tyrell, DL. 2010. Meth MolBiol;649:97-116)
– transcription activator-like effector nucleases (TALENS) (Chen, J et al 2014. Mol Ther;22:303-311)
– CRISPR/Cas9 technology (Seeger, C & Sohn, JA. 2014. Mol TherNucleic Acids;3:e216)

• clustered regularly interspaced short palindromic repeats (CRISPR)/Cas9 platform for efficient gene knockout
• can inhibit HBV replication up to eight fold• inhibition due to introduction of mutations and
deletions in cccDNA from recruited Cas9 repaired by non-homologous end joining (NHEJ)
• inhibition not affected when IFN-α used in combination
• independently confirmedKennedy, EM et al 2015. Virology;476:196.
Lin, S-R et al 2014. Mol Ther Nuc Acids;3:e186.

Modulating Transcription: Epigenetics• Structure of Chromatin
– altered by post-translational modification of histones– acetylation, phosphorylation, methylation and ubiquitylation
• Relaxed Chromatin: Histone Acetylase (HAT)
– transcription activation complex containing HATs– HATs acetylate lysine residues of the histone tails
• Compacted Chromatin: Histone Deacetylases (HDAC)
– transcription repression complex containing HDAC– HDACs deacetylate histone lysine tails
• Conclusion– acetylation status of HBV minichromosome (cccDNA-bound H3 & H4
histones) regulates HBV transcription/replication and is reflected in viral load
Activation of Gene Expression
Repression of Gene Expression
Pollicino, T. et al 2006. Gastroenterology;130:823
Haematologica. 2009;94(11):1618-22

Antiviral activity of GS-5801, a liver-targeted prodrug of a lysine demethylase-5 inhibitor, in an HBV primary human hepatocyte infection model
Gilmore S, et al. EASL 2017, Amsterdam. #SAT-160
AIM: To characterize the relationship between the PD effect (increase in H3K4 methylation) and antiviral activity of GS-5801 in primary human hepatocytes
HMT, histone methyltransferases; K4, lysine 4; KDM5i, lysine demethylase-5 inhibitor; Me, methyl.
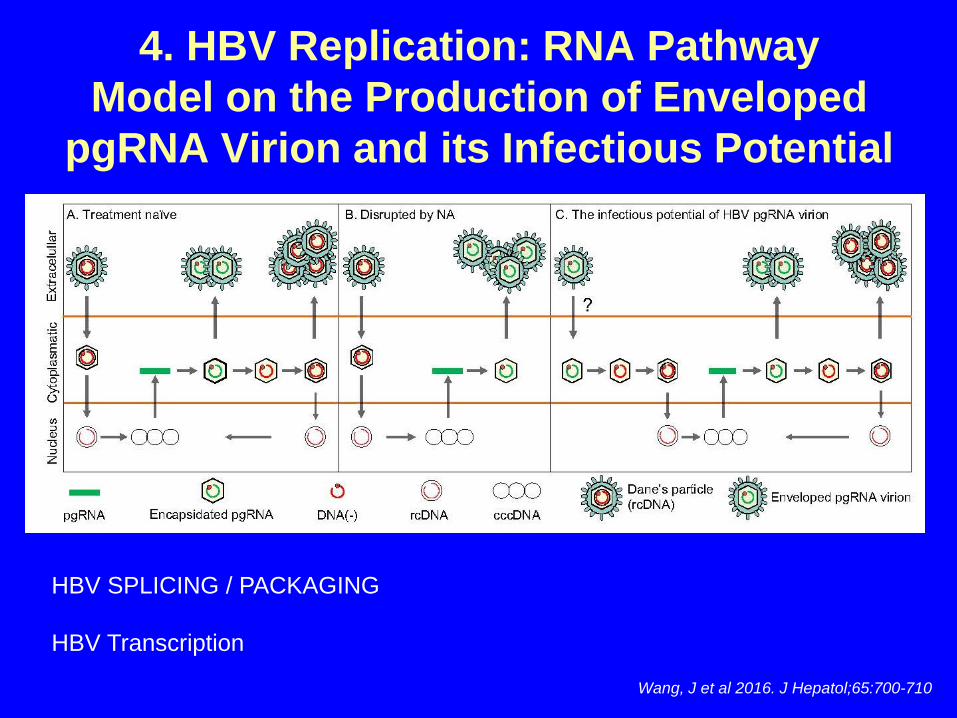
Wang, J et al 2016. J Hepatol;65:700-710
4. HBV Replication: RNA PathwayModel on the Production of Enveloped
pgRNA Virion and its Infectious Potential
HBV SPLICING / PACKAGING
HBV Transcription

The RNA Sensor RIG-I Dually Functions as an Innate Sensor and Direct Antiviral Factor
for Hepatitis B Virus
• RIG-I senses the HBV genotype A, B, and C for the induction of type I and III IFNs
• The 5’-ε region of HBV pgRNA is a key element for the RIG-I mediated recognition
• RIG-I counteracts the interaction of HBV polymerase with pgRNA to suppress viral replication
• Type III IFNs are predominantly induced in human hepatocytes during HBV infection
Sato et al., 2015, Cell Immunity 42, 123–132

Inarigivir (SB9200)• Small molecule nucleic acid
hybrid (SMNH)• Orally bioavailable prodrug• Active metabolite SB9000 • Actively transported into
hepatocytes via OATP1 • 30:1 liver to plasma ratio• Not metabolized, not
phosphorylated. • Biliary excretion of intact
molecule• No direct activity against DNA
polymerase
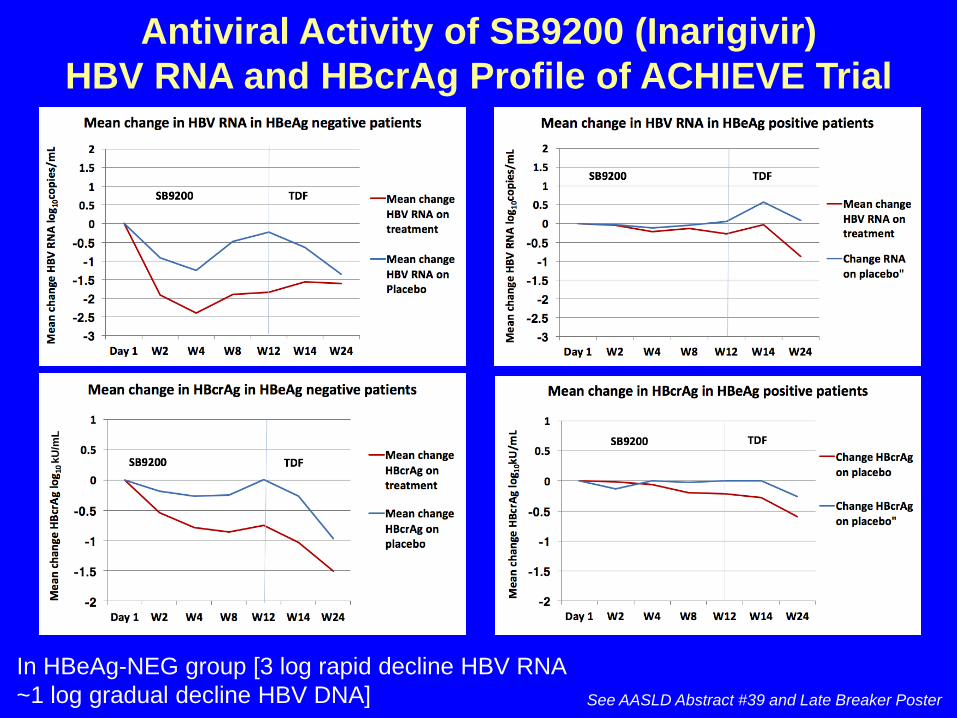
Antiviral Activity of SB9200 (Inarigivir) HBV RNA and HBcrAg Profile of ACHIEVE Trial
See AASLD Abstract #39 and Late Breaker Poster
In HBeAg-NEG group [3 log rapid decline HBV RNA ~1 log gradual decline HBV DNA]
kU/m
L
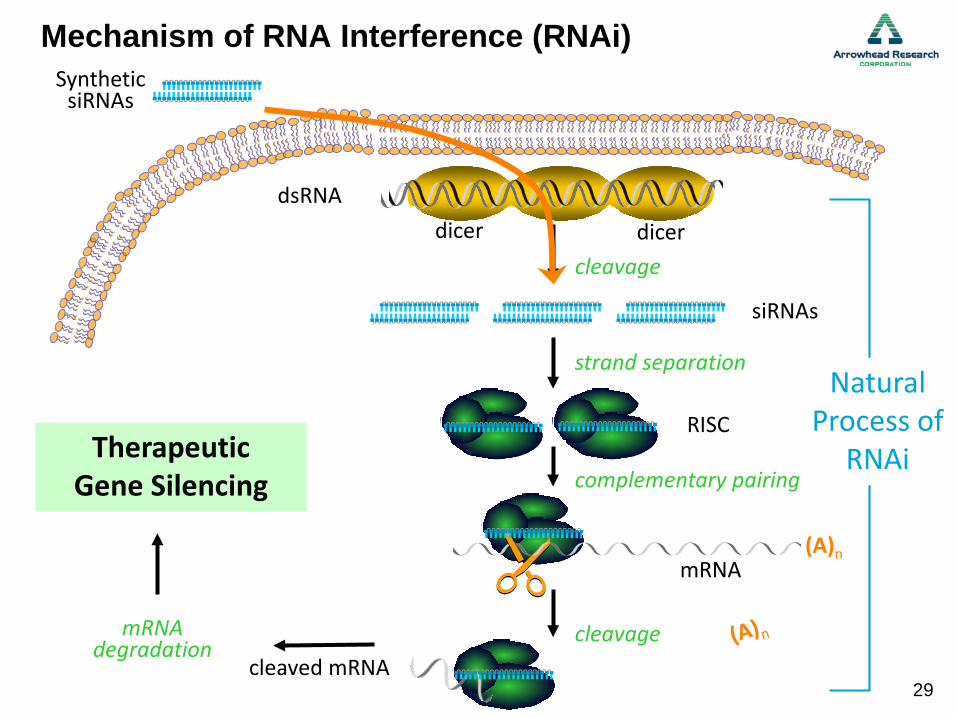
Mechanism of RNA Interference (RNAi)
Natural Process of
RNAi
cleaved mRNA
Selective GeneSilencing
mRNAdegradation
dicerdicerdsRNA
siRNAs
cleavage
RISC
strand separation
cleavage
Therapeutic Gene Silencing complementary pairing
mRNA(A)n
SyntheticsiRNAs
29

Yuen MF, et al. EASL 2017, Amsterdam. #PS-045
HBsAg reduction in HBeAg-positive patients
Smaller reductions in HBsAg in HBeAg– patients:Mean max –0.7 Log10; max observed –1.4 Log10
Multiple doses of ARC-520 resulted in additional reductions in all markers
Reduction in HBeAg+ patients greater than in HBeAg– patients
Difference reflects reductions in HBsAg from cccDNA in HBeAg+ patients vs. integrated DNA in HBeAg– patients
Immediate reductions in HBsAg in HBeAg+ patients:Mean max –2.2 Log10; max observed –3.1 Log10
7/8 patients reported at least one mild AE; none serious of that led to D/C
Prolonged RNAi therapy with ARC-520 in treatment-naive, HBeAg-positive and negative patients with chronic HBV results in significant reductions in HBsAg
HBsA
g (IU
/mL)
80
100
–1020 40 600
Week
1000
10000
100000
Single dose cohort 7
Extensioncohort 10
Patient 01-7981Patient 01-7982Patient 01-7985First dose of ext.Last dose of ext.

2.4kb S
HBV Transcripts Differ Between HBeAg+ and HBeAg-Chimps PacBio Single Molecule Real-Time (SMRT) Sequencing
DR1
DR2 HBV Poly(A) signal
2.1kb S
HBeAg-(88A010)
HBeAg+(A2A004)
HBeAg-• Majority of S transcripts are
fused at the 3’ end to chimp sequence
• Fusion points typically between DR2 and DR1as expected if transcripts arose from integrated HBV dslDNA
HBeAg+• Most S transcripts terminate
near HBV poly(A) signal as expected
HBV-aligning
HBV non-aligning
S ORFs
S transcripts in HBeAg- chimps often lack target sites for ARC-520
ARC-520 siRNAs
Wooddell, C et al (2016)

Novel findings: Predominant liver HBV DNA differs in HBeAg neg and HBeAg pos chimps
Liver biopsy at initiation of ARC-520 treatment revealed:• Most HBV DNA in liver of HBeAg pos is cccDNA• 500-fold less cccDNA in HBeAg neg
– Only 5% of total HBV DNA in liver in HBeAg neg was cccDNA and total HBV DNA levels were not affected by NUCs
• HBV DNA profile in HBeAg neg chimps is consistent with a high proportion of integrated HBV DNA
• Direct antiviral effect on serum HBsAg, HBeAg, and HBcrAg levels which are substantial
• No rebound to baseline from nadir
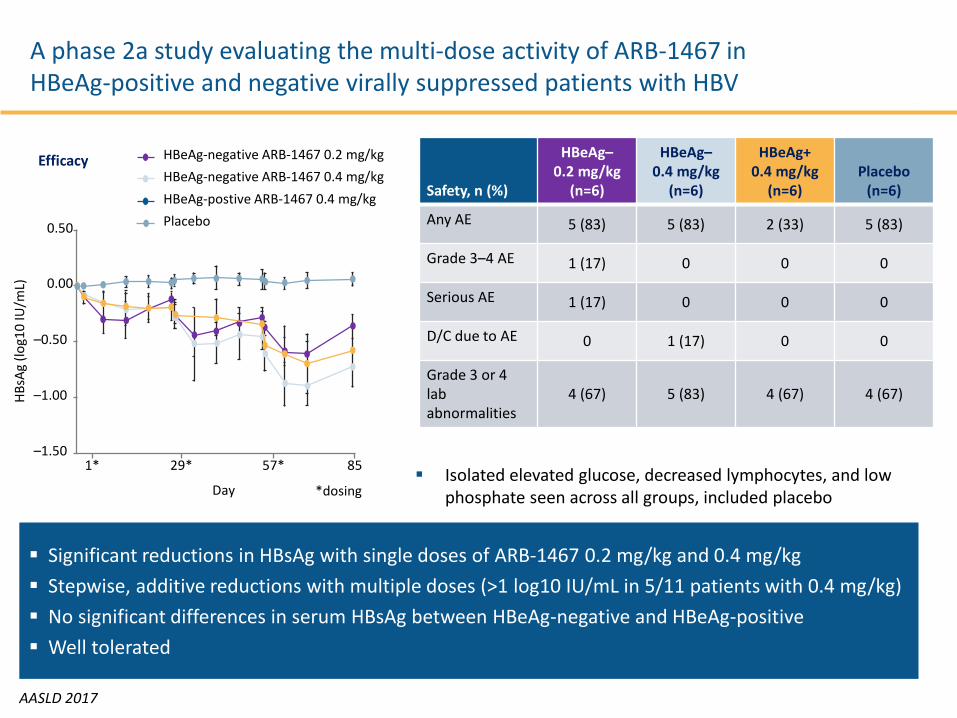
A phase 2a study evaluating the multi-dose activity of ARB-1467 in HBeAg-positive and negative virally suppressed patients with HBV
AASLD 2017
Significant reductions in HBsAg with single doses of ARB-1467 0.2 mg/kg and 0.4 mg/kg Stepwise, additive reductions with multiple doses (>1 log10 IU/mL in 5/11 patients with 0.4 mg/kg) No significant differences in serum HBsAg between HBeAg-negative and HBeAg-positive Well tolerated
Safety, n (%)
HBeAg–0.2 mg/kg
(n=6)
HBeAg–0.4 mg/kg
(n=6)
HBeAg+0.4 mg/kg
(n=6) Placebo
(n=6)
Any AE 5 (83) 5 (83) 2 (33) 5 (83)
Grade 3–4 AE 1 (17) 0 0 0
Serious AE 1 (17) 0 0 0
D/C due to AE 0 1 (17) 0 0
Grade 3 or 4 lab abnormalities
4 (67) 5 (83) 4 (67) 4 (67)
Isolated elevated glucose, decreased lymphocytes, and low phosphate seen across all groups, included placebo
0.50
0.00
–0.50
–1.00
–1.501* 29* 57* 85
Day
HBsA
g (lo
g10
IU/m
L)
HBeAg-negative ARB-1467 0.2 mg/kgHBeAg-negative ARB-1467 0.4 mg/kgHBeAg-postive ARB-1467 0.4 mg/kgPlacebo
Efficacy
*dosing

5. Targeting the HBV Nucleocapsid

Heteroaryldihydropyrimidine compound GLS4 regulates both assembly and disassembly of HBV
capsids to inhibit cccDNA formation
Wang J, et al. AASLD 2017, Washington DC. #937
Different classes of capsid assembly modulators
Heteroaryldipyrimidine derivatives(HAP)
Phenylpropenamide derivatives(AT series)
Compounds in evaluationBAY41-4109
HAP-12AT-130
NVR3-778JNJ-379
ABI-H0731ABI-H0808
GLS4JHSHAP_R01 SBA_R01AB-423

Multiple-dose study of GLS4JHS, interfering with the assembly of HBV core particles, in patients infected with HBV
Improved antiviral potency with RTV boosting but no effect on cccDNA markersDing Y, et al. AASLD 2017, Washington DC. #920
Changes in therapeutic effect factors at different time points after treatment
Compared with values at baseline
1
10 40
–3
–5
–1
0
Mea
n H
BV D
NA
chan
ge fr
om b
aslin
e (L
og10
IU/m
L)
0 20 30
–2
–4
1 400
3M
ean
HBe
Ag le
vel
(Log
10IU
/mL)
0 14 29
2
1
4
33
1 40
Time (day)
3
5
Mea
n H
BsAg
leve
l (L
og10
IU/m
L)
0 14 29
4
33 7 40
Time (day)
–40
40
Mea
n AL
T ch
ange
from
bas
elin
e (IU
/mL)
3 14 21
0
–20
60
33
20
29
GLS4JHS/r 240/100 mg QDETV 0.5 mg QD GLS4JHS/r 120/100 mg QD
GLS4JHS/r combination is generally well-tolerated
PK profiles and efficacy support QD dosing
In chronic HBV patients, GLS4JHS/r combination was demonstrated potent and rapid HBV DNA and HBeAg reduction
These findings support further clinical investigation of GLS4JHS/r combination for the treatment of chronic HBV infection
Cohort
Decline from baseline after 28 days
HBV DNA (log
IU/mL)
HBsAg (log
IU/mL)HBeAg
(PE IU/mL)ETV –3.5 –0.33 –0.43
GLS4JHS/r 120/100 –1.42 –0.06 –0.25
GLS4JHS/r 240/100 –2.13 –0.14 –0.30

6. HBsAg Targeting Strategies
• HBsAg clearance an endpoint of therapy• Decline in HBsAg levels may restore the
antiviral activity of exhausted T cells• Several strategies in evaluation
-RNA interference (SiRNA): « gene silencing »
-Nucleic acid polymers (NAPs): HBsAg release
-HBs antibodies

Immune Regulation by HBsAg
• HBsAg secreted in vast excess over virions (>103 fold)
• Circulate in blood 100-400 µg/ml(1% of total serum protein)
• Unique conformational structure (8 cysteines and 8 prolines )
• Associated with increased risk of HCC (Yuen, MF. et al 2008. Gastro;135:1192–1199)
• Plays a key role in HBV persistence
• Suppress both innate (TLR-2, TLR-9 and IFN-α) as well as adaptive (mDC) responses to infection
Wang, S et al 2013. J Immunol;190:5142.; Xu, Y et al 2009. Mol Immunol;46:2640.; Op den
Brouw, ML et al 2009. Immunol;126:280.

7. Putting It All TogetherPotent
NA
cccDNA Inhibitor
HBV Antigen Inhibitor
Immune Activator
⊕
⊕
⊕/
agent to prevent viral spread and cccDNA re-amplification
safe and selective agent to reduce or silence cccDNA
agent(s) to block/inhibit the HBV life-cycle [entry, cell-spread, capsid assembly, HBx, HBeAg, HBsAg]
agent(s) to activate specific antiviral immune responses or relieve repression/exhaustion of the system
• The medium-term aim for the field is to achieve “functional cure” − HBsAg seroconversion off treatment