endocrinology
TRANSCRIPT


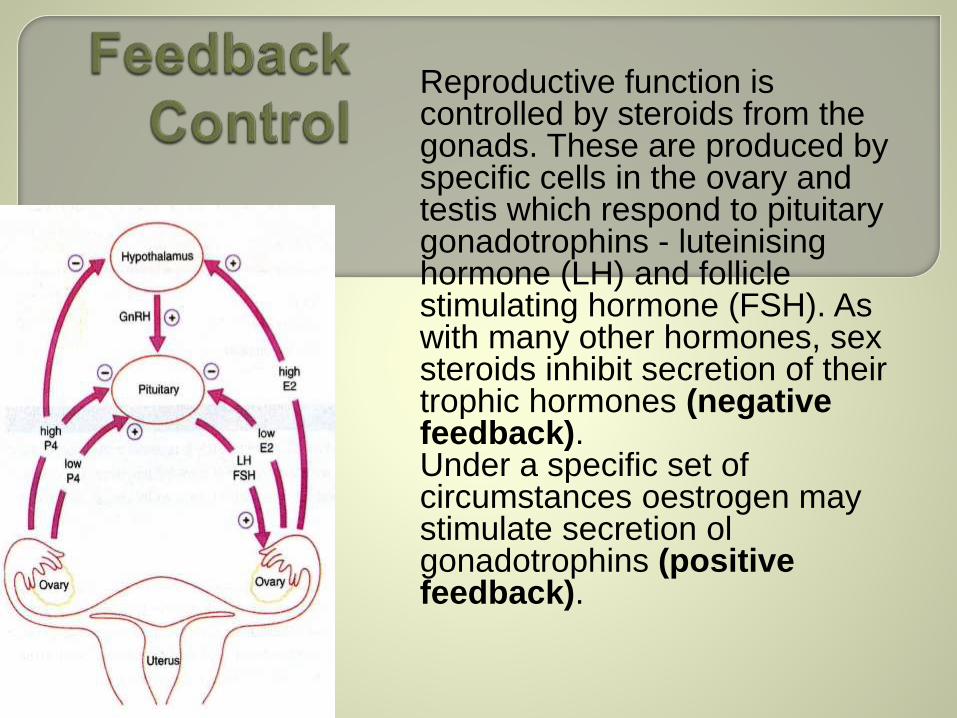
Reproductive function is controlled by steroids from the gonads. These are produced by specific cells in the ovary and testis which respond to pituitary gonadotrophins - luteinisinghormone (LH) and follicle stimulating hormone (FSH). As with many other hormones, sex steroids inhibit secretion of their trophic hormones (negative feedback). Under a specific set of circumstances oestrogen may stimulate secretion olgonadotrophins (positive feedback).

Protein hormones exert their effect on target cells by combining with cell surface receptors. Steroid receptors are located predominantly in the nucleus. The receptor is made up of one steroid binding unit and two non-binding subunits. The binding unit has one binding site for steroid and one for DNA; attachment of the latter to regulatory regions of nuclear DNA controls transcription of RNA and hence protein synthesis. Receptor molecules which are not attached to steroid cannot bind with DNA because the DNA binding site is capped with a specific protein known as heat-shock-protein 90 (HSP 90). The combination of steroid with receptor causesa conformational change, after which it can displace the HSP 90. All the nuclear steroid receptors are members of a single family of proteins. The receptors for thyroid hormone and retinoic acid are part of the same family. The binding units of these receptors are structurally related to some oncogenes. The biological activity is maintained only for as long as the receptor is occupied by the hormone. Thus steroids such as oestrogen, whose receptors have long half-lives, are present in lower concentrations than others, such as progesterone and cortisol, whose receptors have short half-lives.

Larger protein hormones, such as the trophic hormones, cannot enter the cell. They combine with a specific surface receptor which activates adenyl cyclase in the cell membrane. This catalyses production of a second messenger, cyclic adenosine 3' 5'-monophosphate (cyclic AMP), from ATP. Excessive build-up of this substance is prevented by another enzyme, phosphodiesterase. Cyclic AMP activates protein kinases which cause phosphorylation, and thereby activation of specific enzymes. A second and very similar system involves formation of inositoltriphosphate.
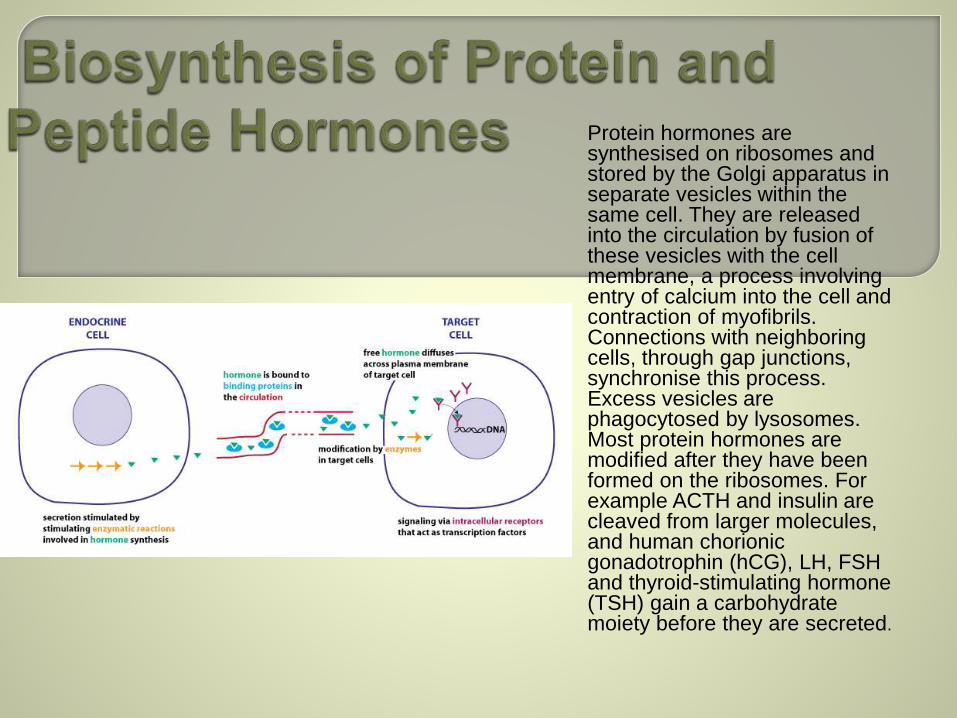
Protein hormones are synthesised on ribosomes and stored by the Golgi apparatus in separate vesicles within the same cell. They are released into the circulation by fusion of these vesicles with the cell membrane, a process involving entry of calcium into the cell and contraction of myofibrils. Connections with neighboring cells, through gap junctions, synchronise this process. Excess vesicles are phagocytosed by lysosomes. Most protein hormones are modified after they have been formed on the ribosomes. For example ACTH and insulin are cleaved from larger molecules, and human chorionic gonadotrophin (hCG), LH, FSH and thyroid-stimulating hormone (TSH) gain a carbohydrate moiety before they are secreted.

All steroid hormones (and thyronines) are carried in blood largely bound to proteins. It is only the ‘free` fraction that is able to move into cells and exert a biological effect. If there is an increase in binding proteins the free fraction is diminished; a compensatory increase in trophic hormone then occurs and restores the total amount of free hormone.

The median eminence of the hypothalamus is connected to the pituitary gland by the pituitary stalk. The hypothalamus stretches from the,optic chiasm in front to the mamillary bodies behind. The hypothalamus receives its arterial supply from the circle of Willis, and the supra-optic nucleus has the richest blood supply of any area in the brain. Capillaries draining the median eminence enter the portal system, which passes into the anterior pituitary.Many nuclear groups - hypothalamic centres - exist in the hypothalamus.The paraventricular and supra-optic centres synthesise posterior pituitary hormones and the ventromedial nuclei control satiety.
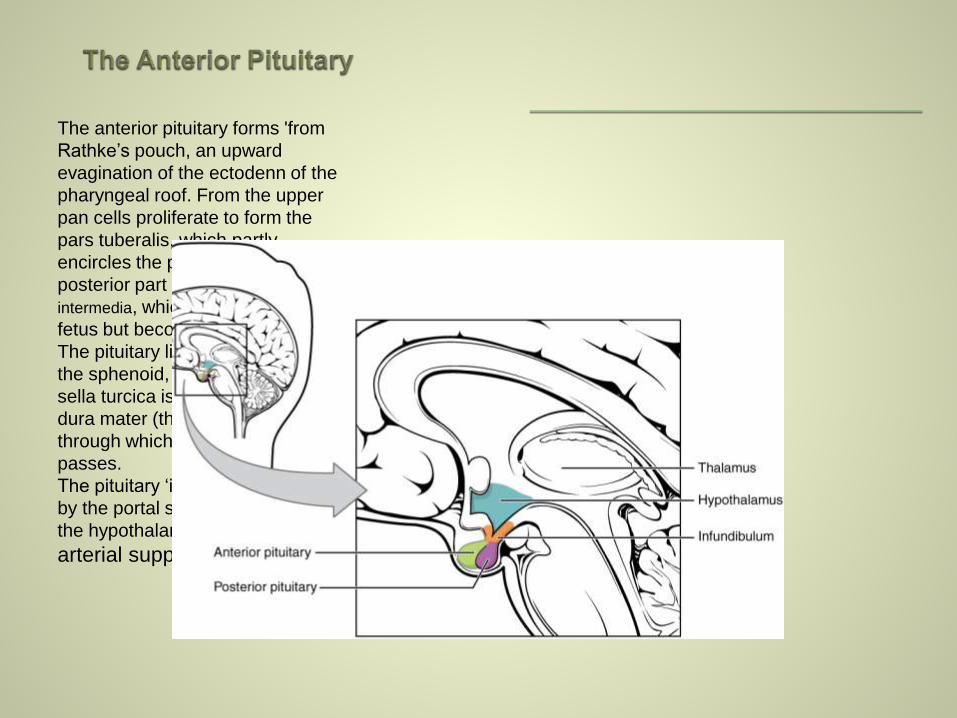
The anterior pituitary forms 'from
Rathke’s pouch, an upward
evagination of the ectodenn of the
pharyngeal roof. From the upper
pan cells proliferate to form the
pars tuberalis, which partly
encircles the pituitary stalk; the
posterior part forms the pars
intermedia, which is prominent in the
fetus but becomes atrophic at birth.
The pituitary lies in a depression in
the sphenoid, the sella turcica. The
sella turcica is covered by a layer of
dura mater (the diaphragma sellae)
through which the pituitary stalk
passes.
The pituitary ‘is supplied with blood
by the portal system originating in
the hypothalamus and by a direct
arterial supply.

Ovaries with developing oocytes are present in the female fetus from an early stage of development. By the end of the second trimester in utero, the number of oocytes has reached a maximum and they arrest at the first prophase step in meiotic division. No new oocytes are formed during the female lifetime.With the onset of menarche, the primordial follicles containing oocytes will activate and grow in a cyclical fashion, causing ovulation and subsequent menstruation in the event of non-fertilization. In the course of a normal menstrual cycle, the ovary will go through three phases:I Follicular phase2 Ovulation3 Luteal phase.

The initial stages of follicular development are independent of hormone stimulation. However, follicular development will fail at the preantralstage and follicular atresia will ensue if pituitary hormones LH and FSH are absent. FSH levels rise in the lirst days of the menstrual cycle, when oestrogen, progesterone and inhibin levels are low. This stimulates a cohort of small antralfollicles on the ovaries to grow. Within the follicles, there are two cell types which are involved in the processing of steroids, including oestrogen and progesterone. These are the theca and the granulosa cells, which respond to LH and FSH stimulation, respectively. LH stimulates production of androgens from cholesterol within theca cells. These androgens are converted into oestrogens by the process of aromatization in granulosa cells, under the influence of FSH. The roles of FSH and LH in follicular development are demonstrated by studies on women undergoing ovulation induction in whom endogenous gonadotrophin production has been suppressed. If pure FSH alone is used for ovulation induction, an ovulatory follicle can be produced, but oestrogen production is markedly reduced. Both FSH and LH are required to generate a normal cycle with adequate amounts of oestrogen.

The endocrine influences in ` menstruation are clear. However, the paracrine
mediators less so, Prostaglandin F2 alpha, endothelin-1 and platelet activating factor (PAP)
are vasoconstrictors which are produced within the endometrium and are thought likely to
be involved in vessel constriction, both initiating and controlling menstruation. They may be
balanced by the effect of vasodilator agents, such as prostaglandin EZ, prostacyclin
(PGI) and nitric oxide (NO), which are also produced by the endometrium. Recent
research has shown that progesterone withdrawal increases endometrial prostaglandin
(PG) synthesis and decreases PG metabolism. The COX-2 enzyme and chemokines are
involved in PG synthesis and this is likely to be the target of non-steroidal anti-
inflammatory agents used for the treatment of heavy and painful periods. Endometrial repair
involves both glandular and stromal regeneration and angiogenesis to reconstitute the
endometrial vasculature. VEGF and fibroblast growth factor (FGF) are found within the
endometrium and both are powerful angiogenic agents. Epidermal growth factor (EGF)
appears to be responsible for mediation of oestrogen-induced glandular and stromal
regeneration. Other growth factors, such as transforming growth factors (TGFS) and IGFS,
and the interleukins may also be important. Greater understanding of mediators of
menstruation is important in the search for medications to control heavy and painful periods.
Mefenamic acid is a PG synthetase inhibitor which is widely used as a treatment for heavy
menstrual bleeding. It is believed to act by increasing the ratio of the vasoconstrictor
PGF2ot to the vasodilator PGE2. Mefenamic acid reduces menstrual loss by a mean value
of 20-25 per cent in women with very heavy bleeding, and further more effective agents are
still being sought.

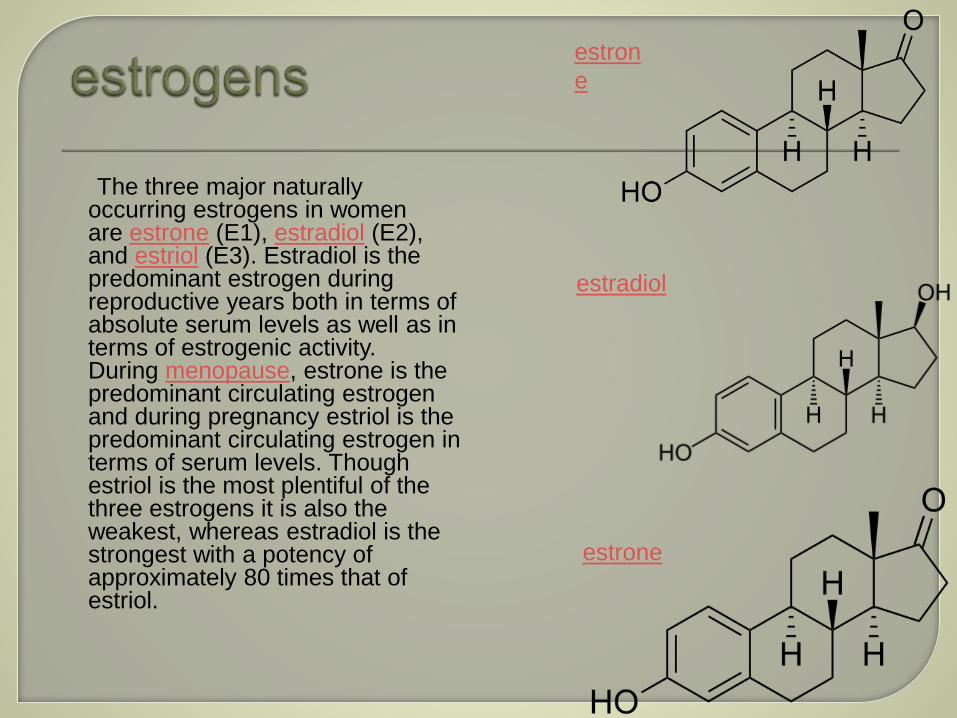
The three major naturally occurring estrogens in women are estrone (E1), estradiol (E2), and estriol (E3). Estradiol is the predominant estrogen during reproductive years both in terms of absolute serum levels as well as in terms of estrogenic activity. During menopause, estrone is the predominant circulating estrogen and during pregnancy estriol is the predominant circulating estrogen in terms of serum levels. Though estriol is the most plentiful of the three estrogens it is also the weakest, whereas estradiol is the strongest with a potency of approximately 80 times that of estriol.
The three major naturally occurring estrogens in women are estrone (E1), estradiol (E2), and estriol (E3). Estradiol is the predominant estrogen during reproductive years both in terms of absolute serum levels as well as in terms of estrogenic activity. During menopause, estrone is the predominant circulating estrogen and during pregnancy estriol is the predominant circulating estrogen in terms of serum levels. Though estriol is the most plentiful of the three estrogens it is also the weakest, whereas estradiol is the strongest with a potency of approximately 80 times that of estriol.
estrone
estron
e
estradiol

Structural Promote formation of
female secondary sex characteristics
Accelerate metabolism Increase fat stores Stimulate endometrial g
rowth Increase uterine growth Increase vaginal
lubrication Thicken the vaginal wall Maintenance of vessel
and skin Reduce bone
resorption, increase bone formation
Protein synthesis Increase hepatic
production of binding proteins
Coagulation
Increase circulating level
of factors 2, 7, 9, 10, plasminogen
Decrease antithrombin III
Increase platelet adhesiveness
Lipid
Increase HDL, triglyceride
Decrease LDL, fat deposition
Fluid balance
Salt (sodium) and water retention
Increase cortisol, SHBG
Gastrointestinal tract
Reduce bowel motility
Increase cholesterol in bile
Melanin
Increase pheomelanin,
reduce eumelanin
Cancer
Support hormone-sensitive breast
cancers (see section below)
Lung function
Promotes lung function by
supporting alveoli (in rodents but
probably in humans).
Uterus lining
Estrogen together with progesterone promotes
and maintains the uterus lining in preparation
for implantation of fertilized egg and
maintenance of uterus function during
gestation period, also
upregulates oxytocin receptor in myometrium
Ovulation
Surge in estrogen level induces the release
of luteinizing hormone, which then triggers
ovulation by releasing the egg from
the Graafian follicle in the ovary.
Sexual receptivity in estrus
Promotes sexual receptivity,[15] and
induces lordosis behavior.[16] In non-human
mammals, it also induces estrus (in heat) prior
to ovulation, which also induces lordosis
behavior. Female non-human mammals are
not sexually receptive without the estrogen
surge, i.e., they have no mating desire when
not inestrus.
Lordosis behavior
Regulates the stereotypical sexual receptivity
behavior; this lordosis behavior is estrogen-
dependent, which is regulated by
the ventromedial nucleus of thehypothalamus.
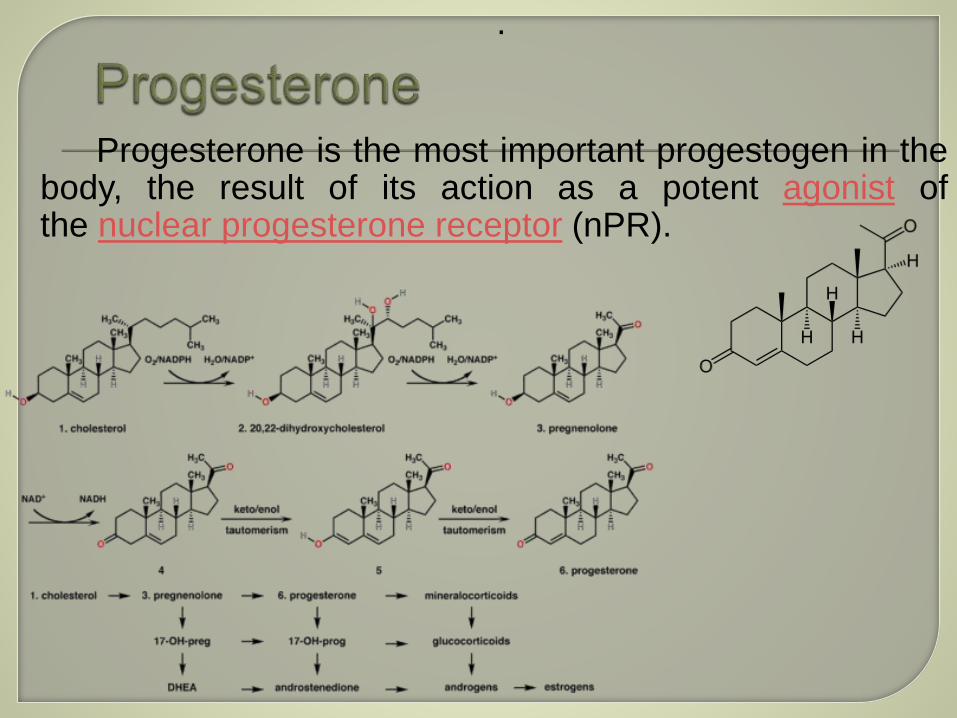
Progesterone is the most important progestogen in thebody, the result of its action as a potent agonist ofthe nuclear progesterone receptor (nPR).
.

Progesterone is sometimes called the "hormone of pregnancy", and it has many roles relating to the
development of the fetus:
Progesterone converts the endometrium to its secretory stage to prepare the uterus for implantation. At the
same time progesterone affects the vaginal epithelium and cervical mucus, making it thick and impenetrable
to sperm. If pregnancy does not occur, progesterone levels will decrease, leading, in the human, to menstruation.
Normal menstrual bleeding is progesterone-withdrawal bleeding. If ovulation does not occur and the corpus
luteum does not develop, levels of progesterone may be low, leading to anovulatory dysfunctional uterine
bleeding.
During implantation and gestation, progesterone appears to decrease the maternal immune response to
allow for the acceptance of the pregnancy.
Progesterone decreases contractility of the uterine smooth muscle.In addition progesterone
inhibits lactation during pregnancy. The fall in progesterone levels following delivery is one of the triggers for milk
production.It belongs to an important group of endogenous steroids called neurosteroids. It can be synthesized
within the central nervous system
Progesterone increases core temperature (thermogenic function) during ovulation.
Progesterone reduces spasm and relaxes smooth muscle. Bronchi are widened and mucus regulated.
(PRs are widely present in submucosal tissue.)
Progesterone acts as an antiinflammatory agent and regulates the immune response.
Progesterone reduces gall-bladder activity.
Progesterone normalizes blood clotting and vascular tone, zinc and copper levels, cell oxygen levels, and
use of fat stores for energy.
Progesterone appears to prevent endometrial cancer (involving the uterine lining) by regulating the effects of
estrogen.
Progesterone plays an important role in the signaling of insulin release and pancreatic function, and may
affect the susceptibility to diabetes or gestational diabetes
The use of progesterone and its analogues have many medical applications, both to address acute
situations and to address the long-term decline of natural progesterone levels. Because of the poor bioavailability
of progesterone when taken orally, many synthetic progestins have been designed with improved oral
bioavailability and have been used long before progesterone formulations became available.