electrocortical changes observed in the n2 as an …
TRANSCRIPT

Northern Michigan University Northern Michigan University
NMU Commons NMU Commons
All NMU Master's Theses Student Works
8-2020
ELECTROCORTICAL CHANGES OBSERVED IN THE N2 AS AN ELECTROCORTICAL CHANGES OBSERVED IN THE N2 AS AN
OUTCOME OF ATTENTION BIAS MODIFICATION TRAINING OUTCOME OF ATTENTION BIAS MODIFICATION TRAINING
Hayley Gilbertson [email protected]
Follow this and additional works at: https://commons.nmu.edu/theses
Part of the Biological Psychology Commons, and the Cognitive Psychology Commons
Recommended Citation Recommended Citation Gilbertson, Hayley, "ELECTROCORTICAL CHANGES OBSERVED IN THE N2 AS AN OUTCOME OF ATTENTION BIAS MODIFICATION TRAINING" (2020). All NMU Master's Theses. 649. https://commons.nmu.edu/theses/649
This Open Access is brought to you for free and open access by the Student Works at NMU Commons. It has been accepted for inclusion in All NMU Master's Theses by an authorized administrator of NMU Commons. For more information, please contact [email protected],[email protected].

i
ELECTROCORTICAL CHANGES OBSERVED IN THE N2 AS AN OUTCOME OF ATTENTION BIAS MODIFICATION TRAINING
By
Hayley Elizabeth Gilbertson
THESIS
Submitted to Northern Michigan University
In partial fulfillment of the requirements For the degree of
MASTER OF SCIENCE IN PSYCHOLOGICAL SCIENCE
Office of Graduate Education and Research
October 2020

SIGNATURE APPROVAL FORM
ELECTROCORTICAL CHANGES OBSERVED IN THE N2 AS AN OUTCOME OFATTENTION BIAS MODIFICATION TRAINING
This thesis by Hayley Gilbertson is recommended for approval by the student’s Thesis Committee and Department Head in the Department of Psychological Science and by the Dean of Graduate Education and Research.
__________________________________________________________Committee Chair: Dr. Joshua M. Carlson Date
__________________________________________________________First Reader: Dr. Jon Barch Date
__________________________________________________________Second Reader: Dr. Christina Hartline Date
__________________________________________________________Department Head: Dr. Adam Prus Date
__________________________________________________________Dr. Lisa Schade Eckert DateDean of Graduate Education and Research
08/05/2020
08/04/2020
08/03/2020
08/05/2020
01/07/2021

i
ABSTRACT
ELECTROCORTICAL CHANGES OBSERVED IN THE N2 AS AN OUTCOME OF
ATTENTION BISA MODIFICATION TRAINING
By
Hayley Elizabeth Gilbertson
Anxiety disorders are currently one of the most predominant mental health conditions
worldwide. Increased anxiety is associated with elevated attentional focus to threat also known
as attentional bias to threat. Attention Bias Modification (ABM) is a type of computerized
training, attempting to reduce attentional focus for threatening stimuli and has been found to
successfully reduce symptoms of anxiety. Past studies have implemented ABM training as a
possible tool to modulate attention away from threat in attempt to decrease pathological anxiety.
The N2 is an event-related potential (ERP) detected in scalp EEG recordings that is associated
with conflict monitoring and complex cognitive functioning. Past studies have found that
anxious adults present increased N2’s during conflict processing, suggesting a relationship
between hyperactive conflict monitoring and anxious symptomology. This thesis measured the
effects of ABM on trait anxiety over a 6-week period. Each participant completed an attention
bias task, questionnaires for anxiety and emotion related traits, as well as an EEG session (i.e.,
flanker task) pre and post the 6-week ABM training period. ABM training was administered
through a modified dot-probe task; training attention away from threatening stimuli. The overall
purpose of this thesis was to establish the effect of ABM (compared to control) training on ERP
amplitudes and self-reported levels of anxiety. The results demonstrated a non-significant
correlation between pre-trait anxiety scores and pre- ΔN2. Additionally, there was no significant
effect of ABM on ΔN2 amplitudes or trait anxiety scores.

ii
Copyright by
Hayley Elizabeth Gilbertson
2020

iii
DEDICATION
I want to dedicate this thesis to my close friends and family who have supported me through long nights, lack of money, and busy days. I contribute a great amount of my success to these relationships/influences in my life. More specifically, I want to dedicate this thesis to my dad, Jeff Gilbertson, for shaping me into the individual I am today. Your selflessness and hard work is beyond inspiring. I would also like to thank my Mom, Vicki Vanderkolk for teaching me to always follow my dreams.
This thesis is also dedicated to all the individuals who have and/or are currently experiencing the negative effects associated with anxiety disorders. It is my hope that by identifying possible EEG-based neural biomarkers in the brain that it will increase the knowledge in order to establish efficacious treatment options and preventative measures in the future.

iv
ACKNOWLEDGEMENTS
The author firstly wants to thank her mentor and advisor, Dr. Joshua Carlson for all of the leadership, guidance, and instruction during this project and throughout her master’s program entirely. She also wants to thank the post-doctoral researcher of the CABIN lab, Dr. Lin Fang for all the assistance, management, and knowledge she provided her with throughout her master’s program. She also wants to thank her other two-committee members, Dr. Jon Barch and Dr. Christina Hartline for all of the great feedback and questions that influenced critical thinking into this project and related experiments. Additionally, she would like to thank the members of the CABIN lab for their assistance in data collection and recruitment of this project. Furthermore, the author would like to thank for the National Institute of Mental health for funding this project (as part of a larger research grant; R15MH1109051). Lastly, the author would like to thank Rourke Sylvain for his support and assistance throughout this endeavor. He has been an intellectual touchstone and has inspired her to delve deeper and question more. The format of this thesis follows the publication manual of the American Psychological Association (6th edition) and the Department of Psychological Science at Northern Michigan University.

v
TABLE OF CONTENTS
ABSTRACT. ......................................................................................................................... (i)
COPYRIGHT. ....................................................................................................................... (ii)
DEDICATION. ...................................................................................................................... (iii)
ACKNOWLEDGEMENTS ................................................................................................... (iv)
TABLE OF CONTENTS ....................................................................................................... (v)
LIST OF FIGURES ............................................................................................................... (vI)
INTRODUCTION ..................................................................................................................... 1
LITERATURE REVIEW ........................................................................................................... 4
Anxiety .......................................................................................................................... 4
Attention to Threat ....................................................................................................... 5
Anxiety and Attention Bias to Threat .......................................................................... 7
Attention Bias Modification ....................................................................................... 10
ABM-Positive-Search Training .................................................................................. 12
Error-Related Negativity (ERN) ................................................................................ 13
N2 ................................................................................................................................. 15
METHOD ................................................................................................................................ 17
Participants .................................................................................................................. 17
Procedure ..................................................................................................................... 17
Self-Reported Measures ............................................................................................. 18
State-Trait Anxiety Inventory .................................................................................... 18
Dot-Probe Task ........................................................................................................... 19
Flanker Task ............................................................................................................... 20

vi
Attention Bias Modification Training .........................................................................21 EEG Recording .......................................................................................................... 24
Data Processing ......................................................................................................... 24
Analytic Plan............................................................................................................... 25
RESULTS ................................................................................................................................ 26
N2 Manipulation Check ............................................................................................. 27
DISCUSSION .......................................................................................................................... 29
Effect of ABM Training on Self-Reported Anxiety and ΔN2 .................................... 32
Clinical Implications....................................................................................................34
Limitations and Future Directions ............................................................................. 35
Conclusions ..................................................................................................................37
REFERENCES ........................................................................................................................ 39
APPENDIX A ........................................................................................................................... 59
Figures .........................................................................................................................59
APPENDIX B……………………………………………………………………………………
IRB approval letter ...................................................................................................... 37

vii
LIST OF FIGURES
Figure 1. Example of the modified flanker task paradigm ........................................................ 59
Figure 2. 10-20 Electrode map highlighting FCz (electrode 4) ................................................. 59
Figure 3. Example of the dot-probe task during ABM training displaying a threatening (left) and neutral face (right) ............................................................................................................................... 59
Figure 4. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the control group (PRE data) . .................................................................................. 60
Figure 5. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the control group (POST data) .................................................................................. 60
Figure 6. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the experimental group (PRE data; i.e., pre 6-week ABM training) . ......................... 61
Figure 7. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the experimental group (POST data; i.e., post 6-week ABM training) . ..................... 61
Figure 8. Correlation between pre-training self-reported STAI – T score and pre-training ΔN2 amplitude (in μV) across all participants ......................................................................................................... 62
Figure 9. Correlation between pre-training self-reported STAI – T score and pre-training incompatible ΔN2 amplitude (in μV) across all participants ..................................................................................... 62
Figure 10. Evidence of the correlation (or lack thereof) between pre-training STAI-T score and pre-training ΔN2 amplitudes (in μV) in the experimental group...................................................... 63
Figure 11. Although not significant, this chart provides evidence that ABM may correlate with the ΔN2, suggesting it may be related to conflict monitoring. .................................................................. 63

1
Introduction
Anxiety is associated with increased attentional bias to threat and decreased executive
functioning, decreased cognitive control, and excessive error monitoring (Mogg & Bradley,
2016). This increases an individual’s susceptibility to external distractions, suggesting a
relationship between emotion and cognition in pathological anxiety.
Anxiety is characterized by both the irrational perception of threat and the duration and
intensity of fear experienced. Over 25 percent of individuals will be diagnosed with an anxiety
disorder during their lifetime (Kessler, Berglund, Demler Jin, Merikangas, & Walters, 2005).
The mapping and identification of neuro-correlates associated with anxious symptomatology
would allow for possible improved treatments and the establishment of preventative measures.
Current popular treatments for anxiety disorders consist of Cognitive Behavioral Therapy
(CBT) and Pharmacotherapy; both have been found to have a 50% success rate, while
pharmacotherapy can be generalized and susceptible to negative side effects such as drug
dependence (Bar-Haim, 2010; Hofmann, Asnaani, Vonk, Sawyer, & Fang, 2012). This can lead
to an increase in anxious symptoms and a prolonged recovery (Bishop, 2009). Attention bias
(AB) is the preferential tendency to attend to threat-related stimuli, in comparison to a neutral or
positive stimulus, and is a cardinal symptom of anxiety. Some research suggests an increased AB
to threat is a causal factor in the development and maintenance of anxiety (Beck & Clark, 1997;
Dennis & Chen, 2006; MacLeod, 2002). Researchers have shown a significant link between
anxiety disorders and hyperactive attention bias to threatening stimuli (Mogg & Bradley, 2016).
Recently, established Attention Bias Modification ABM treatments have ascertained the
ability to transfer participants' automatic attentional processing away from threat, towards non-
threatening stimuli, resulting in an improvement of the cognitive mechanisms associated with

2
inhibition and down regulation of attentional bias to threat (MacLeod et al, 2002; Mogg &
Bradley, 2006; Nelson et al, 2015).
Previous studies have examined the effectiveness of ABM using neuroimaging measures
such as MRI/fMRI (e.g., Aday & Carlson, 2017). This approach is costly and can be intrusive.
Electroencephalography (EEG), which measures electrocortical activity, and in particular interest
to this thesis, event-related potentials (ERP) could provide a more cost effective and relatively
non-invasive option, compared to fMRI. Two ERP’s previously associated with error processing
and conflict monitoring are the Error-Related Negativity (ERN) and the N2. The ERN is
associated with anxiety due to its role in worry, apprehension, and cognitive inhibition (Moser,
Moran, Schroder, Donnellan, & Yeung, 2013). The N2 is associated with conflict processing and
the demand for increased cognitive control (Dennis & Chen, 2009). Cognitive control and
inhibition are associated with the modulation of top-down and bottom-up processing. Top down
processing is goal-directed, involving effortful processing of one’s control of a task. Bottom-up
processing is stimulus driven involving unconscious automatic processing (Mogg & Bradley,
2016). Mogg and Bradley discovered that anxiety enhances stimulus-driven attention whereas
top-down cognitive mechanisms associated with inhibition become less effective (Mogg &
Bradley, 2016). The extent to which ABM modifies conflict monitoring (N2) processes is
unknown.
The purpose of this project is to identify and improve the understanding of electro-
cortical changes following ABM. Specifically, this thesis seeks to assess the extent to which
ABM reduces N2 ERP amplitudes, and self-reported anxiety.

3
Overall Research Question:
● Will ABM lead to a decrease in electrocortical measures of conflict processing in high-
anxious individuals measured through N2 amplitude?
Hypothesis 1: heightened levels of trait anxiety are negatively correlated with ΔN2 (ΔN2
= incongruent flanker N2 - congruent flanker N2) amplitudes. Such that, individuals with
heightened self-reported trait anxiety scores present larger negative ΔN2 amplitudes.
Hypothesis 2: 6-weeks of ABM training decreases ΔN2 amplitude and self-reported
anxiety levels.

4
Literature Review
Anxiety
Anxiety is associated with reduced emotional conflict adaptation (Larson, Clawson,
Clayson & Baldwin, 2013), impaired cognitive control (Sehlmeyer, Konrad, Zwitserlood, Arolt,
Falkenstein, & Beste, 2010), increased attention to threatening stimuli (Dennis & Chen, 2009),
avoidance behavior (Larson, Clawson, Clayson & Baldwin, 2013), and hyperactivity in specific
neural structures associated with emotional regulation and conflict processing (Mogg & Bradley,
2016). High-trait anxious individuals have an increased attention to threat-related stimuli and
diminished ability to transfer attention away from threat. This suggests a decrease in cognitive
control and difficulty orienting attention to goal-relevant stimuli (Eastwood, Smilek, & Merikle,
2003; Fox, Russo, Bowles, & Dutton, 2001; Mogg, Mathews, & Eysenck, 1992).
Research investigating emotional conflict processing has found that individuals
diagnosed with Generalized Anxiety Disorder (GAD) have an inability to implicitly regulate
emotional conflict (Etkin & Schatzberg, 2011). Furthermore, they found deficits in the neural
regions associated with implicit regulation during emotional conflict processing. That is, GAD
patients fail to show the typical increase in ventral cingulate activity and associated dampening
of amygdala activity during implicit emotion regulation (Etkin & Schatzberg, 2011). This
suggests a decrease in inhibitory control mechanisms and a deficit in attentional regulation.
The two most prevalent treatments for anxiety disorders include cognitive-behavioral
therapy (CBT) and pharmacotherapy; resultant in a 50% success rate for each approach in
clinical settings (Bar-Haim, 2010; Hofmann, Asnaani, Vonk, Sawyer, & Fang, 2012). The
importance of improving current treatments and the continuation of establishing innovative and

5
more successful options is imperative in psychology (and related fields). Moreover, developing
less invasive and more effective anxiety treatments for children is also of paramount importance
(Bar-Haim, 2010).
Trait anxiety is the increased tendency to be affected by environmental stressors (Fales et
al, 2008) while anxiety sensitivity represents the increased susceptibility to experience anxious
symptomology and the effects it has on behavior (McNally, 2002). Individuals with anxiety have
neurocognitive deficits in cognitive control and inhibition. Trait anxiety and anxiety sensitivity
have been linked to response inhibition in individuals who have anxiety. The concepts of anxiety
that trait anxiety and anxiety sensitivity represent are still unknown (Lilienfeld, 1996); although
it is assumed that the two work together, while trait anxiety is responsible for cognitive factors,
anxiety sensitivity is assumed to represent one’s proneness/probability to experience anxiety.
Recent research has focused on the relationship between cognitive differences, anxiety traits, and
electrophysiology for more insight into the psychopathology of psychiatric disorders related to
anxiety (Karch et al., 2008; Manly, Robertson, Galloway, & Hawkins, 1999; Roche, Garavan,
Foxe, & O’Mara, 2005).
Attention to Threat
Past research has provided evidence that AB to threat is increased in a subset of
individuals with anxiety disorders, linking this bias to the development and maintenance of such
disorders (Dennis & Chen, 2009). This bias can be measured through conflict interference of
cognitive control processing of attention (Fenske & Eastwood, 2003; Simpson et al., 2000;
Williams, Mathews, & MacLeod, 1996). When processing information, highly salient emotional
cues are prioritized over stimuli that are less salient. This process is crucial for survival, eliciting
avoidance of threat or signaling approach of a positive outcome. When an individual comes

6
across emotional stimuli, their perceptual and attentional systems prioritize the stimuli associated
with emotional mechanisms, ensuring that the individual is aware of this information primarily
(Sussman et al., 2016). Evidence of this was displayed in an experiment conducted by Ohman
and colleagues, measuring reaction time on stimulus detection, comparing threat-related words
and neutral/positive words. Results displayed that the participants responded more rapidly to the
words associated with threat compared to the neutral and positive words (Ohman et al., 2001).
Another example includes an experiment using a set of rapidly displayed images of threatening
and non-threatening stimuli, resulting in the participant’s ability to identify more of the
threatening images post presentation in comparison to neutral stimuli (Anderson, 2005).
Attention mechanisms allow individuals to selectively attend and respond to specific
stimuli (Bishop, 2009). Selective attention is the process of orienting attention towards one thing
while simultaneously disengaging focus towards all other external stimuli. This is regulated by
the interplay between “bottom-up” and “top-down” processing (Bishop, 2008). Top down
processing refers to goal-directed behaviors or processes, increasing one’s control of a task.
Bottom-up processing is stimulus driven, referring to unconscious automatic processing (Mogg
& Bradley, 2016). The resources for cognitive processing associated with attention and
emotional processes compete when increased threat is perceived. Top-down control is
diminished and attention towards emotionally salient stimuli is increased. (Easterbrook, 1959;
Hanoch & Vitouch, 2004; Leith & Baumeister, 1996; Meinhardt & Pekrun, 2003; Wood,
Mathews, & Dalgleish, 2001). Therefore, one’s AB is altered by the reciprocal influence
between top-down and bottom-up processing. In other words, an individual’s attention is
constantly queued in the environment through perceived novel/salient stimuli while
simultaneously competing for more focused and goal-driven cognitive ability (Bishop, 2008).

7
AB reflects the conflict interference on cognitive control processing of attention (Fenske
& Eastwood, 2003; Simpson et al., 2000; Williams et al., 1996). AB to threat refers to the
unconscious tendency to attend towards threatening stimuli in comparison to neutral/positive
stimuli. This is thought to be overrepresented in anxious populations, linking them to an
increased bias to threat. Individuals with low self-reported anxiety display low AB to threat and a
higher tolerance to threat-related situations (Bishop, 2008).
There are many examples of this dichotomy between anxious and non-anxious
individuals. Williams et al. (1996) investigated the effects of AB on task performance using an
emotional word Stroop task. Participants were presented one word at a time on a computer
screen. The words were split into two groups: emotionally salient and neutral and were displayed
in various colors. Participants were asked to respond to the color of the ink while ignoring the
word's implication. The participants that were classified as anxious had a slower response time
when responding to threat salient words (Williams et al., 1996). Another study, using a dot-probe
task, measured AB in anxious and non-anxious controls. The study found the anxious
participants were faster to detect the probe when behind threat-related trials compared to neutral
trials (Mogg & Bradley, 1998). These findings suggest that anxious individuals tend to exhibit a
heightened AB to threatening stimuli. Due to the emotional and cognitive factors linked to
anxiety, it makes sense that there is a relationship between anxiety and diminished performance
in situations encountering emotional or cognitive conflict (Mogg & Bradley, 2016).
Anxiety and Attention Bias to Threat
Individuals with anxious symptomology have enhanced attentional disruptions to threat-
related stimuli (Eastwood et al., 2003; Fox et al., 2001; Mogg et al., 1992). As mentioned
previously, AB to threat is seen in most individuals (regardless of anxious affiliation), although

8
the modulation of attention towards threatening stimuli is more persistent and impairing in
individuals diagnosed with an anxiety disorder (Mogg & Bradley, 2016). Individuals
experiencing anxiety often superimpose negative salience onto non-threatening stimuli
(Pergamin-High et al., 2015). Evidence of selective processing of threat in anxiety disorders
suggests a critical relationship between etiology and ailment of such disorders (Bar-Haim et al.,
2007).
Avoidance of threat is another potential factor for decreased task performance in anxious
populations. The degree to which anxious individuals efficiently recruit cognitive resources can
be completely inhibited when fear is present (Bishop et al., 2004; Compton, 2003; Compton et
al., 2007). Bishop and colleagues (2004) conducted a study using threat-related distractors (i.e.,
fearful faces) during a non-emotional decision making task in individuals experiencing anxiety.
Although task performance was unaffected, decreased neural activity was found in regions
linked to cognitive control, such as the left prefrontal cortex and dorsal Anterior Cingulate
Cortex (dACC; Bishop, et al., 2004). Researchers suggest this may reflect a neural mechanism
attempting to mitigate the degradation of task-performance when bottom-up emotional and
conflict mechanisms are enhanced (Gray, 2004).
The dot-probe task (sometimes also referred to as the visual-probe task) intrinsically
measures selective attention to threat in order to assess one’s AB (MacLeod, 1987,1988;
Thigpenet al., 2018). The typical experiment starts with a fixation point (center of the screen),
followed by the concurrent display of threatening and non-threatening cues on the left and right
side of the screen. After these stimuli disappear, a probe (i.e., dot, letter, arrow, etc.) replaces one
of the previously displayed cues (threatening vs non-threatening). The probes are administered
randomly and with equal probability between cues. Participants are asked to respond to the

9
location of the dot as quickly and as accurately as possible. AB is measured through reaction
time to each probe. AB to threat is reflected through a decreased response time to threat-
congruent trials, i.e. trials in which the probe replaces the threatening cue. An increased response
time in threat-incongruent compared to threat-congruent trials indicates that the participant’s
attention is unconsciously biased towards the side of the screen displaying the threatening
stimuli. This is often referred to as vigilance (Mogg & Bradley, 2016). Conversely, the
behavioral trait displayed when individuals react quicker to threat-incongruent trials compared to
threat-congruent trials is classified as avoidance (Mogg & Bradley, 2016). AB scores are
determined by the RT difference of threat-incongruent trials and threat-congruent trials (i.e.,
threat-incongruent trials minus threat-congruent trials). Positive scores represent vigilance while
negative scores reflect avoidance. An individual is said to have no attentional bias, if their
incongruent – congruent score is near zero.
Mounting evidence implicates the prefrontal cortex (PFC) and the extended amygdala
network as regions associated with AB mechanisms and targets of neuroplastic change following
ABM training (Aday & Carlson, 2017). Human lesion studies have demonstrated that the
amygdala is critical for the allocation of attentional resources to threatening stimuli. For
example, research involving an attentional blink paradigm showed that healthy observers
robustly perceived verbal stimuli of aversive content compared with stimuli of neutral content.
Conversely, patients with bilateral amygdala lesions showed no enhanced perception for such
aversive stimulus events (Anderson and Phelps, 2001). Furthermore, Vuileumier and colleagues
(2004) conducted event-related fMRI investigations assessing the amygdala’s influence on the
visual cortex when healthy, hippocampus lesion, and amygdala lesion patients were presented
emotional visual stimuli (i.e. fearful faces). The results demonstrated that, in contrast to the

10
patients with amygdala lesions, both healthy patients and patients with hippocampal lesions had
enhanced activation in the fusiform and occipital cortex for threatening compared to neutral
stimuli (Vuilleumier et al., 2004). Moreover, the nucleus basalis of Meynert (nbM), which
contain acetylcholine (Ach) neurons, are a major projection site of the amygdala and the nbM
sends projections diffusely throughout the cortex—including visual and other sensory cortices—
to facilitate perceptual processing (Himmelheber et al., 2000; Himmelheber et al., 2001; Peck &
Salzman, 2014; Sarter et al, 2003;).
In addition to the amygdala, numerous sub-regions of the PFC play an integral role in
attention orientation and modulation. For example, Wolf et al. used eye-tracking techniques to
investigate the effect the ventromedial PFC (vmPFC) has in orienting attention to fearful stimuli
(2014). The results concluded that patients with vmPFC lesions oriented their attention to eye
regions of fearful faces significantly less than those without vmPFC lesions. Prior human and
non-human primate investigations have shown that the vmPFC and the amygdala are densely and
reciprocally connected (Barbas, 2000; Ghashghaei & Barbas, 2002; Roy et al., 2009).
Furthermore, the attention orientation reflected by amygdala activity has been shown to be
correlated with activity in the ventral and dACC (Carlson et al., 2009). This amygdala-PFC
network may reflect the neural mechanism for allocating visual attention to emotionally and
socially salient stimuli.
Attention Bias Modification
As mentioned previously, anxious individuals have a heightened AB towards threatening
stimuli. This schema towards threatening stimuli has been found to play a large role in the
etiology and maintenance of such disorders (Beck & Clark, 1997; Eysenck, 1997; Rapee &
Heimberg, 1997; Williams et al., 1997).

11
Using neuroimaging and cognitive/behavioral methodologies, researchers are able to peer
into the neural mechanisms associated with such mental health disparities, establishing a
biological foundation, on which more efficacious treatments can be developed. Examples of this
include Gabrieli’s research demonstrating improved reading performance amongst dyslexic
children by administering an auditory phonological processing schema of print words (2009). In
addition, Fisher et al. found schizophrenia patients who underwent auditory training displayed
marked improvements in cognition, verbal working memory, and verbal learning memory as
well (2009). These examples display cognitive-behavioral treatments that improved psychiatric
disorders.
In light of the cognitive-behavioral therapies addressed above, recent investigations have
employed a modified dot-probe task, used to measure AB, in attempts to modulate a participant’s
AB towards or away from task relevant stimuli and has been classified as ABM (Mogg &
Bradley, 2016). ABM was first implemented in two studies, which evaluated the effect of a dot-
probe task on anxiety scores. The investigations demonstrated that participants who underwent
more than 6000 training trials, where the probe location consistently appeared behind the neutral
stimuli (opposed to threat), displayed not only lower AB scores but also a significant decrease in
trait-anxiety scores (reviewed in Mathews and MacLeod, 2002). These studies initiated the rising
interest in ABM research and its effects on anxiety symptoms (Bar-Haim, 2010).
Current investigations also attempt to understand the neural mechanisms of ABM
(Browning et al., 2010; Mansson et al., 2013; Mansson et al., 2016; Maslowsky et al., 2010;
Taylor et al., 2013). Similar to the regions discussed above (see AB section), the amygdala and
prefrontal cortices have been shown to reflect a scalar and reciprocal influence on the attentional
mechanisms, respectively (Away & Carlson, 2017; Taylor et al., 2013). Taylor et al.

12
administered an attention modification program (AMP) to participants with high social anxiety,
and provided evidence of attenuated activation from pre- to post-AMP in the bilateral amygdala,
bilateral insula and subgenual anterior cingulate cortex (2013). Furthermore, participants
displayed enhanced activation in prefrontal cortices, such as the left anterior dorsal ACC, left
dorsolateral PFC, vmPFC and right ventrolateral PFC, following the AMP (Taylor et al., 2013).
This evidence suggests that decreased activation in the ACC and amygdala, as well as enhanced
activity in the PFC, following AMP may reflect overall improvements in the functioning of
attentional neural mechanisms during emotional processing. These findings add to the mounting
evidence implicating the ACC and the medial PFC as neural regions strongly associated with
emotional processing and ABM (Etkin et al., 2011).
ABM-Positive-Search Training
Although there is ample evidence regarding an anxious individual’s susceptibility
towards threat, anxiety disorders have been diagnosed without any perception to threat (Van
Bockstaele et al., 2014). ABM-positive-search training focuses on transferring one’s automatic
attention towards positive environmental cues. In this method, participants are instructed to
respond to positive stimuli only, inhibiting attention towards threat. This method could
potentially be administered to patients with anxiety disorders whose symptoms vary from those
associated with vigilance to threat (Dandeneau et al. 2007; De Voogd et al., 2014; Waters et al.,
2013).
Researchers discovered that using the same tools that measure AB could also be
applicable to manipulate one’s attention away from threat. As mentioned previously (Anxiety and
AB to threat), the dot-probe task is used to measure attentional bias to threat. The task is also
used to implement ABM training. The use of the dot-probe task in attention training involves an

13
increase in target probes associated with neutral/positive stimuli in attempt to guide the
participant’s attention away from the threatening stimuli. Numerous (>100) trials are required to
alter one’s pattern of attention, shaping an altered perception to threat. In one study, participants
who displayed high-trait anxiety participated in an attention training dot-probe task (consisting of
6000-7500 trials each), and displayed a significant decrease in trait-anxiety when pre and post
training were compared. In another study, patients diagnosed with General Anxiety Disorder
(GAD) completed dot-probe training comprised of 160 trials (8 sessions in four weeks). This
resulted in 58% of GAD patients no longer meeting the DSM-IV requirements for the disorder
(Bar-Haim, 2010).
Error-Related Negativity (ERN)
An acquisition process of EEG measurements, where an isolated electro-cortical
waveform is segmented into epochs, temporally bound to an event, and averaged across trials—
is used for analysis, and has become a popular technique deployed for cognitive and behavioral
investigations. These waveforms are known as event-related potentials (ERPs), first recorded in
1935 by Hallowell Davis et al. (1939), multiple facets of psychological and neurological research
deploy this technique in order to better understand the temporal dimensions of electro-cortical
activity emitted by the brain. In particular, research investigating performance monitoring and
error detection have provided substantial evidence of a specific ERP associated with these
behaviors classified as the error-related negativity (ERN; Riesel et al., 2013; Yeung et al., 2004).
The ERN is an ERP closely associated with anxiety due to its nature in apprehension/worry,
concept control, and cognitive inhibition. It is localized to the frontocentral electrode cites,
displayed approximately 50-100 ms after an erroneous response has been conducted (Zambrano-
Vazquez & Allen, 2014). ERN amplitude can vary across individuals depending on participants'

14
perceived response accuracy; however, latency is predominantly consistent (Bress et al., 2015).
Research suggests a strong correlation between the ERN and increased activity in the Anterior
Cingulate Cortex (ACC) due to its association with conflict, action monitoring, emotional
influences, and motivational factors (Dennis & Chen, 2009; Hajack et al., 2010). Accompanied
physiological changes include increased skin conductance, heart rate activity, and the startle
reflex (Bress et al., 2015).
In addition, as discussed above, research suggests that the ACC is involved in attention
bias to threat and is modulated by ABM training. With research providing evidence of ACC
modulation due to ABM training (Taylor et al. 2013), ABM may modulate additional functions
of the ACC, such as error monitoring as measured by the ERN. As mentioned previously,
disruptions in attentional processing to threat-related stimuli reflect the interference between
emotional and cognitive processing. Studies focusing on generalized anxiety disorder (GAD)
suggest that an individual’s inability to modulate attention and inhibit cognitive processes are
due to decreased ability to detect and respond to conflict (Etkin et al., 2010). This interference is
heightened in anxious compared to non-anxious individuals, due to their increased susceptibility
of emotional processing (Easterbrook, 1959; Hanoch & Vitouch, 2004; Leith & Baumeister,
1996; Meinhardt & Pekrun, 2003; Wood et al., 2001).
The ERN has been found to be noticeably enhanced in individuals with GAD and OCD
(Endrass et al., 2008; Gehring et al., 2000; Hajcak et al., 2008; Weinberg et al., 2010; Weinberg
et al., 2012). Family history of mental health (Carrasco et al., 2013; Riesel et al., 2011) and
inability to inhibit behavior at a young age (McDermott et al., 2009) are also linked to a
heightened ERN; both classified as common risk factors for anxiety. In two studies conducted by
Nelson, et al. (2015; Nelson, Jackson, Amir, & Hajcak, 2017), researchers examined the effect of

15
ABM on the neural processing of errors (i.e., ERN). Participants were administered single
session ABM training through a modified version of the spatial cueing task (Posner, 1980). Error
processing was measured through ERN amplitude, elicited through a flanker task. Participants
completed the experiment conversely (i.e. ABM before ERN was measured vs. ABM after ERN
was measured). The ERN was significantly decreased (i.e., less negative) in participants who
completed ABM prior to eliciting the ERN, suggesting that a single session of ABM
disengagement towards threatening stimuli may influence ERN amplitude through decreased
neural activity associated with anxiety (Nelson, et al., 2015). These findings suggest that ABM
may be successful in modulating attention away from threatening stimuli, signaling the ERN as a
possible mechanism of ABM-related changes.
N2
The N2 is stimulus-locked, scalp-recorded ERP displayed when individuals require
increased cognitive processing (Dennis & Chen, 2009). It is associated with the frontal midline
regions, displaying a negative amplitude occurring 200-350 ms after a stimulus was presented.
The mean amplitude is configured through the comparison of incongruent and congruent N2
amplitudes. The amplitude of incongruent trials are increased, compared to congruent trials;
when the task exhibits conflict (i.e. incongruent flanker display; < < > < <) or when the
incongruent stimuli is highly salient (i.e. it is associated with inhibitory control mechanisms;
Larson & Clayson, 2011). The N2 indicates the need for increased cognitive control, improving
one’s response to future conflict and increased inhibition to unrelated stimuli. The proposed
brain structure associated with N2 is the ACC responsible for conflict and action monitoring.
The amygdala and other limbic regions relay information to the ACC, further associating with
motivational and emotional factors. The ACC holds an important role in the ERN and N2, due to

16
its neural basis integrating emotion and cognition. Conflict-monitoring theory suggests that N2
amplitudes (mean amplitude from 300-350 ms) are larger when task-irrelevant stimuli is
presented, suggesting one must exhaust more attention when presented unrelated stimuli (Dennis
& Chen, 2009).
Taken together, research suggests that individuals with high levels of trait-anxiety have
an increased AB to threat. Evidence of selective processing of threat in such individuals proposes
there is a relationship between etiology and maintenance of anxiety disorders. Due to the high
concentration of individuals experiencing anxiety and the poor success rate of current treatments
(i.e., 50% success in CBT and pharmacotherapy), it is vital that new treatment options are
established for use in clinical settings. Top-down and bottom-up processing together determine
one’s selective attention towards external stimuli. Researchers have developed ABM in order to
transfer selective attention towards more goal-oriented behaviors, indirectly decreasing anxiety
through increased cognitive performance/decreased stimulus driven processing. The N2 is an
ERP associated with conflict processing and has shown to be elevated in anxiety. A decreased
N2 amplitude displayed in individuals suggests that ABM could influence cognitive processing,
possibly manipulating psychological/physiological components associated with trait anxiety.
Therefore, a decreased ΔN2 was expected following 6-week ABM training

17
Methods
Participants
Eighty-four men and women (M = 31) between the ages of 18-38 (M = 21.83, SD = 4.82)
were recruited from the Marquette Community. Participants were chosen randomly to be part of
the control (N = 32; i.e., completing six weeks of training, not implementing ABM training
towards or away from threat) or experimental group (N = 52 ; i.e., completing six weeks of
ABM-training away from threatening stimuli). Fifty-six of those recruited were not part of the
final results due to artifact rejection (N = 5), training attrition (N = 18), or quarantine in response
to the COVID19 pandemic (N = 33). The final analysis included 28 participants, with 14 in the
experimental group, and 14 in the control group. Participants were informed of the study through
various posters in the community and numerous Facebook posts from the Cognitive Affective
Behavioral and Integrative Neuroscience (CABIN) lab page. Data for this experiment were
collected as part of a larger research project (NIMH #R15MH1109051), following the same
recruitment method.
Procedure
Participants completed a pre-screening survey verifying eligible age range (18-42),
possession of a smartphone, normal (or corrected to) vision, current psychological treatment, any
past/current diagnosis of mental illness, history of head injury and lost consciousness, current
medications, anxiety elicited through small spaces, status of childbearing, and possible metal
placed inside or outside the body that could not be removed. Possible exclusion criteria included
recent head injuries (e. g., concussion), history of neurological disorders, current consumption of
psychoactive medication, and/or currently seeking mental health treatment through any form of

18
counseling or therapy. If participants met criteria based on the pre-screening survey, they were
contacted by a research assistant in the CABIN lab through the google voice text application.
Participants then completed one task and three self-report surveys to verify their
eligibility in the study: the dot probe task, the State and Trait Anxiety Inventory (STAI), the
Depression and Stress Scale (DASS), and the Cognitive Emotion Regulation Questionnaire
(CERQ). Participants were required to obtain an AB incongruent - congruent score of at least 7
ms and STAI-T score of 40 or higher to be included for participation in the study. If individuals
met inclusion criteria, they were asked to provide general background information such as age,
gender, dominant hand, and ethnicity. Next, they completed the screening (which measured
behavioral effects and AB score based on a dot-probe task) and the three previously mentioned
self-report surveys. If participants did not meet requirements of the in-person screening, they
were compensated $10 for their participation in the screening. If participants did meet inclusion
requirements for the study, they were compensated $65 following their post EEG session.
Self-Report Measures
State-Trait Anxiety Inventory.
The STAI was created to measure the prevalence of anxiety in individuals. This measure
has two subscales: State and Trait (Leal et al., 2017). State anxiety is prompted through
environmental factors; therefore, on the state scale participants were asked to respond how they
felt in that moment. Trait anxiety involves a character predisposition that increases susceptibility
to anxiety and is evaluated through factors that suggest an increase prone to anxiety (e.g, general
states of calmness, confidence, and security; (Guillen-Riquelme & Buela-Casal, 2011; Julian,
2011). Each category of anxiety involves 20 questions, assessed on a 4-point Likert-scale (1 =
almost never, 4 = almost always). Each subset score (state and trait anxiety) has a range of 20-80.

19
Total scores for each are determined by the sum of anxiety symptoms, simultaneously
subtracting responses indicating the absence of symptoms. A score of 39 or higher indicates
clinically significant systems for state anxiety (Julian, 2011). Participants were selected for high
levels of anxiety (≥ 40 on the STAI-T) and heightened attentional bias to threat.
Dot-probe task.
A dot-probe task requires an incongruent-congruent difference score of greater than or
equal to seven ms. It measures intrinsically selective attention in two stimuli (See figure 3;
Thigpen et al., 2018). It is important for assessing and altering an individual’s AB. Participants
start with a fixation cue displayed in the center of the screen (1000ms). Then, there were two
stimuli displayed on either side of the screen (100ms). Following stimulus display, a dot was
displayed on either the same or the different side than the stimulus was displayed on and
remained until a response was made. Participants were asked to respond to the orientation of the
dot as quickly as possible, using their right hand; right index finger on the first button to respond
to dots displayed on the left side of the screen, right middle finger on the second button in
response to right-sided targets. AB was measured through response time of each trial; suggesting
a faster response indicates the participants’ unconscious attention was already biased towards
that side of the screen, with slower responses indicating fixation on the opposite side. Faster
reaction times to congruent threat stimuli predict a bias toward related stimuli (MacLeod, 1987-
1988; Mogg & Bradley, 2016).
Prior to beginning participation in the study, individuals were required to read and sign
the informed consent form, confirming they understood the procedure and agreed to volunteer in
data collection. Participants were notified in the consent form that they were volunteers and
allowed to leave at any time during the study. Once deemed eligible to participate, individuals

20
were asked to provide additional demographic information; full first, middle, and last name, date
of birth, and city/state of birth. The participants were measured for head circumference in cm in
order to anticipate the corresponding EEG cap size. In addition, the participants were scheduled
for their Pre-EEG session and the CABIN ABM application was installed on their smartphones.
Participants were then randomly placed in a control (completing ABM training sessions through
the modified version of the dot-probe task that are not modulating attention towards or away
from threatening stimuli) or an experimental group (using a modified version of the dot-probe
task to alter individuals attention away from threat).
During the EEG, participants were required to complete a seven block computer
administered flanker task (see Flanker task - above) and 8 minute resting state measure where
they were instructed to remain as relaxed as possible.
Flanker task.
A Flanker Task is the most common and effective measure used to elicit an ERN (Yeung,
Botvinivk, & Cohen, 2004). Participants were administered a modified flanker task using the
EPrime 3.0 presentation software. Participants were seated 59 cm from the computer screen.
Responses were recorded with a chronos box placed in front of the computer monitor. For each
trial (see Figure 1), participants were presented with a fixation point (i. e., a white plus sign in
the middle of the screen) for 1000ms. Following the fixation cue, the screen displayed five white
arrows centered on a black screen for 200 ms. Each trial could be classified as a congruent trial
(e.g., < < < < < or > > > > >) or an incongruent trial (e.g., < < > < < or > > < > >). All trial types
were administered with equal probability. Each trial had an inter-trial interval of 1000 to 1500
seconds where the participants were instructed to provide the exhibited direction of the center
arrow (left or right) using the corresponding buttons on the chronos box. The task included one

21
practice block (20 trials) and seven training blocks, consisting of 60 trials each, completing a
total of 420 trials. This experiment complied with a predefined protocol, which instructed
procedures for the collection of demographic information, fitting of the EEG cap, experiment
progression and recording, and subsequent cap cleaning (See Flanker protocol in APPENDIX).
Attention bias modification training.
The ABM task used was a slightly altered version of the ABM training dot-probe task
conducted by Aday and Carlson (2017). Participants completed training via their personal
smartphone on the “CABIN” cell phone application. To maintain consistency with the previous
paradigm, duration of the training remained six weeks and the same set of facial stimuli was
utilized as the original task. Sixty word stimuli, taken from the Affective Norms for English
words (ANEW), were added to the ABM task; they were rated by valence and arousal,
categorizing them as neutral or fearful stimuli. Neutral and fearful word pairs were matched by
length and frequency, for a total of 30 pairs (Bradley & Lang, 1999; Stevenson, Mikels, &
James, 2007).
The application was compatible with apple and android devices. Prior to beginning
training, participants were required to read through the training instructions and requirements.
They were then asked if they had any questions regarding training, the cell phone application, or
anything else related to the experiment. Training consisted of six weeks, with a requirement of
no more/no less than six training sessions to be completed per week. This resulted in a total of 36
sessions. To better control regulation of training, completion of one training session a day was
preferred for participants; performance of up to three sessions per day was accepted. Participants
were issued daily reminders to complete their sessions and warning messages if further action
was needed (e.g. participant missed a session). Participants were excluded from the experiment if

22
they were behind on their sessions for more than one week (as stated in the informed consent
document) and provided with $10 of compensation. The procedure of the ABM training is
explained in detail below:
Once the participant opens the “CABIN” application, a “Prepare for Trial” screen was
displayed; requiring the individual to set their phone to “Do Not Disturb” and ensure their level
of brightness is set to its highest setting. This screen also reminds participants to find a place
where they can complete the training distraction-free. Once the participants were prepared to
begin training, they were administered 10 PANAS items and asked to respond with how they
currently felt in the present moment using a Likert scale of 1 (“Not at all”) to 5 (“Extremely).”
The participants were administered a prompt with the following instructions: “Please try your
best to concentrate on the task. Your performance may be compared anonymously with other
participant’s performance at a later time.” Pressing the “next” button on the screen confirms the
participant has seen the instruction. Instructions were then provided on how to complete the
training, (i.e., “Focus your gaze on the cross. You will briefly see two stimuli. Tap the half of the
screen where the dot appears next as promptly as you can!”) Participants completed the training
session. Each training session consisted of 240 trials. Participants completed a total of 8,640
trials during the 6-week experiment. Feedback on response accuracy (correct vs. incorrect) and
response time (“Fast”, under 300 ms; “OK”, between 300 – 1000 ms; “Too Slow”; above 1000
ms) was given following each trial.
As mentioned previously, participants were assigned to two groups: the ABM treatment
group or the neutral control group. The cell phone application functioned identically in both
groups, although trial type differed between groups. Incongruent trials (i.e. trials where the dot
appeared behind the neutral stimuli) were administered to participants in the ABM treatment

23
group. This was done in order to modulate attention training away from threatening stimuli. The
control group was administered an equal amount of incongruent and congruent trial types in each
training session. Participants were able to track their progress and current accuracy during each
session. Progress was also monitored by participants in the CABIN lab to ensure sessions were
being completed appropriately. After the six weeks are completed, participants were scheduled
for their second EEG session and compensated $65 for their time spent on the study.
Following completion of the first EEG, participant’s application ID and pin were
implemented into the app., giving them access to begin training. They were given reminders to
complete their sessions on a daily basis and were sent warning messages when sessions were not
completed in the appropriate time. Participants were required to complete six test sessions via the
cell phone app each week, for six consecutive weeks. No more than three sessions were allowed
to be completed in one day, preferring the completion of one session each day, six days of the
week.
Since data was collected from a larger research project (NIMH R15MH1109051),
participants were required to get a pre and post fMRI, 10 minute resting state measure in addition
to the two EEG sessions and completion of the six week training in order to receive
compensation. Some participants in the treatment condition were offered increased compensation
to return for a third EEG, fMRI, and behavioral measure (i.e., dot-probe task, STAI, DASS, and
CERQ) session 6 weeks following post session data collection which was added two years after
beginning data collection. Participants received a total of $65 or $100; specific to whether they
returned for third session data collection (receiving $100 in comparison to $65 for completion of
the original experiment procedure).

24
EEG recording.
In this experiment, computer program setup was predefined in a protocol (See Flanker
Protocol in Appendix). Data were continuously recorded using a 64 channel Geodesic Sensor
Net (see figure 2; Electrical Geodesics Inc., Eugene, OR) with AgCl electrodes placed according
to the international 10-20 system. Signal acquisition was performed using Net Station 4.5.4
software (Electrical Geodesics Inc., Eugene, OR) and was digitized at a sampling rate of 500 Hz.
Electrode impedance was kept below 75 kilo-ohms.
Data processing.
EEG data was analyzed using EEGLAB V2019.1 (Delorme & Makeig, 2004) and
ERPLAB V8.0 (Lopez-Calderon, J., & Luck, S. J., 2014)., MatLab packages (2010, The
Mathworks Inc., Massachusetts, USA). Continuous EEG data set files were first iterated through
a preprocessing script. Each file was low-pass filtered at 30 Hz and high-pass filtered at 0.1 Hz.
Congruent and incongruent bins were time-locked to the stimulus presentation and epoched 200
ms prior to presentation and 800 ms post presentation and were then baseline corrected from -
200 ms to 0 ms. Files then underwent artifact detection where amplitude deflections of at least
140 μV at eye-blink electrodes were considered eye-blinks and amplitude deflections of 55 μV
or greater at eye-movement electrodes were considered horizontal eye-movements. Epochs
containing eye-blinks or eye movements were excluded from data analysis. Additionally, epochs
with more than 10 bad channels were discarded. Bad channels were replaced with interpolated
data using spherical splines from the remaining channels. Following interpolation, files were
subjected to an Independent Components Analysis (ICA) to identify and correct artifacts. The

25
ERP segments were then averaged for each participant so that each electrode had a single
waveform for each of the four conditions (pre-control, pre-training, post-control, post-training).
Following preprocessing, electrode averages amongst each subject were re-referenced
within groups and grand averaged. The data was then extracted and saved to an untraceable ID
number associated with each participant (Larson & Clayson, 2011).
Following data processing, the grand averaged frontocentral electrode waveforms of the
four groups were visually inspected in order to determine which electrode best displays the delta
N2. Statistical analysis utilized the amplitude extraction from 300 - 375 milliseconds of the
individual averages across groups.
Analytic Plan
Statistical analyses were performed using SPSS V21 (2012, IBM Statistics, New York,
USA).
Hypothesis 1: heightened levels of trait anxiety are negatively correlated with ΔN2 amplitudes.
To test the correlation between pre-ABM self-reported trait anxiety scores and ΔN2
amplitudes, a Pearson Correlation was used. A negative correlation between pre-ABM trait
anxiety scores and ΔN2 amplitudes would lend support towards hypothesis 1. It was
hypothesized that participants’ self-reported anxiety levels would be negatively correlated with
their ΔN2 amplitudes as measured by the STAI questionnaire and EEG respectively. That is,
participants who reported having higher anxiety displayed a larger negative ΔN2 (ΔN2 =
incongruent flanker N2 - congruent flanker N2). These results would provide evidence that
enhanced emotional processing mechanisms, present in anxious individuals, were interfering
with or disrupting the cognitive mechanisms that are engaged when monitoring conflict.

26
Hypothesis 2: 6-weeks of ABM training decreases self-reported anxiety levels and ΔN2
amplitudes.
To test for changes in self-reported trait anxiety following ABM training and changes in
ΔN2 following training, a 2 (ABM vs Control) × 2 (pre vs post) mixed ANOVA was performed
on anxiety and ΔN2 measures. It was hypothesized that there would be a significant interaction
for both ANOVAs demonstrating significant changes in ΔN2 amplitudes and self-reported
anxiety following training for the ABM group only. More specifically, post-session ΔN2
amplitudes, as measured with EEG, were expected to significantly decrease in the training group
compared to their pre-ABM training amplitudes. That is, the ΔN2 amplitudes of the training
participants is less negative following ABM training. Additionally, the self-reported anxiety
scores of the training participants is reduced compared to their pre ABM training scores.
Furthermore, the quantity of reduction between participants' ΔN2 amplitude and anxiety scores
were expected to be proportional across the training sample group. This suggests an intimate
relationship between AB, conflict monitoring, and anxiety.

27
RESULTS
N2 Manipulation Check
Following the statistical processing of the results, a N2 Manipulation Check was
implemented (i.e., test for amplitude differences between congruent and incongruent flanker
trials). Here, a statistical significance was documented for pre (Mdiff = 2.72µV, SD = 0.57), t (26)
= 4.781, p < 0.001 and post ΔN2 amplitudes Mdiff = 2.84 µV, SD = 0.49), t (26) = 5.828, p < .001.
Hypothesis 1: heightened levels of trait anxiety are negatively correlated with ΔN2
amplitudes
A Pearson correlation was used to test for a relationship between self-reported levels of
anxiety and ΔN2 amplitudes. There was not a statistically significant correlation between pre-
training STAI-T scores (M = 52.07, SD = 6.66) and pre training ΔN2 amplitudes (M = -2.72 µV,
SD = 3.01 µV), r (27) = 0.11, p = 0.59).
Hypothesis 2: 6-weeks of ABM training decreased anxiety levels and ΔN2
In order to test for the effect of ABM training on anxiety levels and ΔN2 amplitudes, a 2
(pre-training vs. post-training) × 2 (ABM vs. control) mixed factorial ANOVA was implemented
for both STAI-T scores and ΔN2 amplitudes. The between subject measure used was the type of
treatment group (control vs. ABM) with the repeated measures independent variable was time
(Pre and Post). The dependent variables tested were ΔN2 amplitude and STAI-T scores.
There were not significant main effects of time; pre-training (M = -2.72 µV, SD = 3.01
µV) and post-training (M = -2.84, SD = -2.58), F (1, 26) = 0.048, p =.828, 𝜂𝜂2 = 0.002, or group,

28
F (1, 26) = 0.095, p = 0.761, 𝜂𝜂2 = 0.004, on ΔN2 amplitudes. Additionally, there was no
significant interaction between the time × group on the ΔN2, F (1, 26) = 0.228, p =.637, 𝜂𝜂2 =
0.009 (Experimental Group pre: M = -2.71 µV, SD = .821µV; post: M = -2.57, SD = .698, and
Control Group pre: M = -2.74, SD = .821; post: M = -3.11, SD = .698)
For the STAI-T scores; there was not a statistically significant effect for time in self-
reported anxiety; pre-training (M = 52.07, SD = 6.67) and post-training scores (M = 52.19, SD =
8.2), F (1, 25) = 0.004, p = 0.952, 𝜂𝜂2 = 0.000. The tests of between-subjects group effect was
also not significant; (pre: M = 52.77, SD = 5.372; post: M = 52.00, SD = 7.714), F (1, 25) = .035,
p = .853, 𝜂𝜂2 = .001. Finally, the training × group interaction was not significant, F (1, 25) =
0.418, p = 0.524, 𝜂𝜂2 = 0.016.

29
Discussion
The main objective of this experiment was to assess the effects of ABM training on ERP
modulation and self-reported levels of anxiety in attempts to strengthen the connection between
electrocortical changes and anxiety levels following ABM. As documented in the results, there
was no significant correlation between pre-STAI-T scores and pre-ΔN2 amplitudes.
Additionally, there was no effect of training on STAI-T scores or ΔN2 amplitudes. This suggests
that there is no relationship between ABM training on anxiety reduction or ΔN2 modulation.
Prior to analyzing the ΔN2 amplitudes and the STAI-T scores, a N2 manipulation test was
conducted and provided evidence that there was a significant difference between incompatible
and compatible waveform amplitudes.
ΔN2 and Self-Reported Anxiety
The was a non-significant Pearson correlation between pre ΔN2 amplitudes and
participants self-reported anxiety. This does not support the hypothesis that there exists a
significant negative relationship between pre ΔN2 amplitudes and the STAI-T (i.e., an increase
in STAI-T associated with heightened negative N2 amplitudes). This is inconsistent with
previous research where the ΔN2 was found to be amplified in individuals experiencing greater
levels of anxiety (Dennis and Chen, 2008; Beste et al., 2010; Perez-Edgar et al., 2016; Baldwin
et al., 2013). An increased (more negative) ΔN2 has been correlated with heightened cognitive
processing; the amplitude is increased when the task displays conflict (incongruent trial during
flanker task, i.e., linked to inhibition; Larson & Clayson, 2011) or when a stimuli is highly
salient (Eldar and Bar-Haim, 2010). As mentioned previously, individuals with increased anxiety

30
have an AB to threatening stimuli, resulting in decreased cognitive control. When an individual’s
cognition is diminished, it inhibits their ability to orient to goal-relevant stimuli, facilitating
one’s inability to implicitly regulate emotions (Etkin & Schatzberg, 2011). Therefore, cognitive
control and inhibition are strongly correlated with conflict processing. Furthermore, because the
ΔN2 is associated with increased conflict processing, it was expected that a more negative ΔN2
amplitude correlates with pre-STAI-T scores.
The lack of significance toward the first hypothesis could be due to the small sample size
(N = 28) not providing enough power to detect a significant effect; a larger sample size may have
provided more evidence of a relationship between increased self-reported anxiety and the ΔN2.
Another reason could be due to the self-reported survey measuring for anxiety (STAI-T). Self-
reported surveys can sometimes provide inaccurate conclusions due to how the question is
perceived or the current state of the participant. Because inclusion criteria for participant
screening required both a heightened sense of anxiety(as measured with the STAI-T)—and the
dot-probe scores, it is less probable that the screening alone may have influenced the
participant’s perceptual framing, but may have had an effect on correlation performed above
(Tversky, A. and Kahneman,D., 1981).
A study conducted by Owens et al. (2015) looked into the effect of cognitive control
deficits modulated by task demands and emotional salience through a modified version of the
Erikson Flanker task (Eriksen & Eriksen, 1974). Participants were displayed five faces
simultaneously in the middle of the screen for each trial. The middle (third) face was the central
target and consisted of a neutral male or female face. The other four faces surrounding the
central target (two on each side) were faces displaying angry, happy, or neutral expressions; the
surrounding faces were of the opposite sex as the central target (i.e., if the central neutral face

38
was a female, the surrounding four faces were male). The participants were instructed to
determine the gender of the central target as quick and accurately as possible. Concurrent with
the flanker task, participants were presented with an auditory tone of low, mid, or high pitch
during each trial. This task was split into two conditions; low-load and high-load. During the
low-load condition, instructions were to say the word “tone.” In the high-load condition,
participants were instructed to report the pitch of the tone. In both conditions, participants were
to respond after the facial stimuli from the flanker task is displayed. In order to assess the effects
of self-reported anxiety and worry, participants were administered the STAI, the Worry Domains
Questionnaire (WDQ), & the Penn State Worry Questionnaire (PSWQ) prior to beginning the
experiment. Participants displayed larger ΔN2 amplitudes during the high-load condition
compared to the low-load condition, suggesting that increased need for cognitive resources is
linked to an increased N2. Additionally, trait worry was associated with increased ΔN2
amplitudes under the high-load condition. The use of the WDQ and the PSWQ in addition to the
STAI may provide more insight into the variability of individual levels of anxiety and worry.
Here, converse to the results provided above, a significant correlation between ΔN2 amplitudes
and self-reported anxiety--or in this case--trait worry, was documented. The increased cognitive
component may be a necessary variable to test N2 modulation.
As mentioned previously, the N2 is a negative ERP waveform associated with conflict
processing (i.e., increased conflict correlates with a more negative ΔN2 amplitude; Nieuwenhuis
et al., 2003;Yeung et al., 2004). When participants completed the flanker task (see figure 1), the
incongruent trials (< < > < <) elicited a more negative amplitude compared to congruent (< < < <
<) trials, reflecting the electro-cortical engagement of top-down control to decrease conflict.
(VanVeen & Carter, 2002; Yeung et al., 2004). Furthermore, N2 amplitudes are influenced by

39
the exposure to preceding trial condition. Such that, when an incongruent trial precedes an
incongruent trial, the N2 is diminished in comparison to a congruent trial preceding an
incongruent trial. The same is true for a congruent trial that is preceded by another congruent
trial (Clayson & Larson, 2011a; Clayson & Larson, 2011b). This is referred to as repetition
priming, and is understood to be the unconscious conditioning to specific patterns during
performance tasks (e.g., the flanker task). This phenomenon could be involved in the alteration
of data by diluting ERP amplitudes. For example, if a participant were to elicit an N2 in response
to two incongruent trials back to back, there is a probability that the N2 elicited during the
second trial becomes slightly diminished. A possible explanation could be that the neural
mechanisms involved in repetition priming recruit distributed neural resources that improve
cognitive functioning, in turn reducing conflict processing. If conclusive, this could lend
evidence to why the ΔN2 between conditions are relatively similar and could potentially
diminish the correlation between participant’s pre-STAI-T scores and the ΔN2.
Effect of ABM Training on Self-Reported Anxiety and ΔN2 Amplitude
There was no significance found between pre and post self-reported STAI-T or pre and
post ΔN2 amplitudes. Pre STAI-T scores for all participants had almost no change when
compared to post STAI-T scores. The experimental group displayed similar results between the
pre STAI-T scores and post STAI-T scores. Therefore, six weeks of ABM training did not have a
significant effect on decreasing trait anxiety, as determined by their STAI-T scores. One possible
reason is that although there is a large amount of evidence linking anxiety with AB, anxiety
disorders are diagnosed without any altered AB to threat. The ABM training is used to transfer
attention away from threatening stimuli in order to increase cognitive functioning and decrease
stimulus driven behaviors; this increases one’s ability to utilize goal-oriented behavior. If

40
participants were experiencing anxiety without a perception of threat, ABM would not be
sufficient to decrease their symptoms because the anxiety is not associated with increased
attention to threatening stimuli. Conversely, previous research has shown that there may be a
time lag between the conclusion of ABM training and both the electro-cortical and behavioral
modulation (Browning et al.2012). This may provide another reason that behavior and electro-
cortical activity were not shown to have been significantly modulated.
When comparing ΔN2 amplitudes, there was no significant relationship between pre and
post sessions. There was not much of a change between pre ΔN2 (M = -2.72 µV) and post ΔN2
(M = -2.84 µV) amplitude when comparing all participants (regardless of group), which was
expected due to only half (N = 14) of the participants receiving ABM treatment. Interestingly,
there was a slight increase (less negative) in the group receiving ABM when comparing pre ΔN2
amplitudes (M = -2.71 µV) and post ΔN2 amplitudes (M = -2.57 µV). Although there was no
significant change between these amplitudes, the data still displays a trend of ABM on ΔN2
amplitude, providing some evidence that ABM may be able to improve task performance during
increased cognitive conflict. This would need to be assessed in a larger sample in order to
properly examine the influence of ABM on the ΔN2. Further, the ΔN2 plays a large role in the
allocation of top-down cognitive control, impairing one’s conflict processing (i.e., when your
brain is competing for goal-driven and stimulus driven properties; vanVeen & Carter, 2002;
Yeung et al., 2004). The N2 measures conflict through the detection of novelty vs mismatch
stimuli when stimuli are attended to (Na¨a¨ ta¨nen & Gaillard, 1983; Na¨a¨ ta¨nen & Picton,
1986). Implementing additional variables to increase the cognitive load might improve or
establish a relationship between ABM and the ΔN2.

41
Other studies looking at the effect of ABM on the N2 have used No-go and flanker
paradigms. From this experiment, the results suggest that ABM does not decrease anxiety over a
six-week period, at least when using self-reported measures such as the STAI-T. In parallel with
other studies, this experiment suggests that ABM may strengthen cognitive functioning by
assisting in the reorientation of attention to goal-directed stimuli measured by the ΔN2 (Larson et
al., 2013). Although, the results were not significant, the present study did see a small effect of
ABM on post ΔN2 amplitude that could be replicated and expanded on to further assess its
reliability/validity.
Clinical Implications
The results from this study alone do not provide enough evidence to justify the usage of
the N2 as a target for treatment efficacy. When comparing the pre (M = -2.71) and post ΔN2 (M
= -2.57) averages, there was a slight increase (less negative) in the average post ΔN2 amplitude
in the experimental group. Although, the results were not significant, the results may suggest that
with more participants and further research, ABM could have a noteworthy effect on anxious
symptomology.
There was no effect of ABM on self-reported anxiety measures. The pre STAI-T scores
(M = 52.57) showed almost no difference compared to the post STAI-T scores (M = 52).
Although early ABM studies documented promising results for improving anxiety, (Mogg &
Bradley, 2016), these results do not support the justification for clinical application. As
mentioned previously, not all individuals diagnosed with anxiety experience an AB to threat
(Van Bockstaele et al, 2014). For example, a study conducted by Eldar et al (2012), assessed the
relationship between anxious children and ABM-threat avoidance. This excluded almost half of
the population of anxious children, due to the absence of pre-existing threat bias. This is an issue

42
because although this treatment may be beneficial for anxious children (& adults) who do have a
pre-existing bias to threat, it is simultaneously neglecting the many others experiencing the same
or similar mental illness without a bias to threat. Another approach of ABM is referred to as
ABM-positive search training. In contrast with the ABM-threat avoidance model, positive search
training directs your attention toward positive stimuli in comparison to orienting away from
negative (Dandeneau et al., 2007; De Voogd et al., 2014; Waters et al., 2013; Waters et al., 2015;
Waters, et al., 2016). This approach was designed to attempt to suit all individuals with anxiety
disorders, through the instruction to fixate on the positive images, while inhibiting background
stimuli (Dandeneau et al., 2007; De Voogd et al., 2014; Waters et al., 2013; Waters et al., 2015;
Waters, et al., 2016). Moreover, a recent paper published by Sylvain et al. (2020) investigated
the effects of a single session ABM to positive stimuli on the electro-cortical activity relating to a
reward processing ERP complex, P2-RewP-P3 (Sylvain et al. 2020). The investigation
demonstrated a significant effect on female P2 amplitudes following positive ABM training. If
more studies used the ABM positive search approach, researchers may find increased success of
ABM in the clinical setting.
Limitations & Future Directions
There are a few limitations to this study. As mentioned previously, the data from this
project were derived from a larger grant (#R15MH1109051). The grants original ERP of interest
was the ERN and the first 29 participants were not flagged for the N2 during the EEG
flanker/resting state measure. This provided a small sample (N = 28), diminishing the statistical
power. The study lost 18 participants due to attrition. If participants did not finish the
experiment, post-data ΔN2 amplitudes and behavioral measures did not exist, forcing the
removal of such participants from analysis. The present study lost an additional 33 participants

43
due to the emergence of the virus COVID-19, imposing an executive order to close labs and keep
the participants safe and healthy. Due to this, behavioral measures were altered to an online
format to continue collection of attainable data. Post EEG measures were not conducted due to
mandated quarantine and the closure of all school buildings/labs. This caused the exclusion of
many participants from analysis of this thesis. The small sample (N = 28) and within groups
(experimental = 14; control = 14), was a major limitation in this study.
Another limitation included the restriction of anxiety symptomology to the self-reported
STAI-T surveys. Self-reported measurements attempt to objectify subjective information and can
be influenced by a participant’s mental state. To improve or reduce inaccurate classification,
additional questionnaires, such as the Worry Domains Questionnaire (WDQ) and the Penn State
Worry Questionnaire (PSWQ) could be used.
An additional limitation of this study (as mentioned in the Effect of ABM training on self-
reported anxiety and ΔN2 amplitude section) included the possibility that the reliability/validity
of the dot-probe measure was suspect (Kappenman et al., 2014; Schmukle, 2005; Staugaard,
2009; Waechter et al., 2013). The dot probe paradigm was developed to quantify an individual’s
attention bias to threatening stimuli (please refer to the ABM section in the literature review for
more information on this measure). In an experiment conducted by MacNamara et al. (2013),
participants displayed a heightened attentional focus towards threatening stimuli in numerous
measures. Due to possible poor psychometric properties of the reaction time (RT) measure of the
dot-probe task (Schmukle, 2005; Staugaard, 2009), it is possible that some dot-probe measures
do not provide an accurate analysis (quantification) of the RT bias measure. For example, the
dot-probe task has been shown to have poor test-retest reliability (Schmukle, 2005; Staugaard,
2009), as well as reduced internal reliability (Kappenman et al., 2014; Schmukle, 2005;

44
Staugaard, 2009; Waechter et al., 2013). In regards to this thesis, it is possible that participant’s
AB scores were invalid due to the imprecision of the dot-probe measurement. Therefore,
implementing ABM treatment for participants that experience anxious symptomology, but do not
experience an enhanced attention bias to threat, would be clinically insufficient (Van Bockstaele
et al., 2014). Interestingly, Reutter and colleagues (2017) demonstrated significant internal
reliability when measuring attention bias to threatening images with the N2pc ERP in the dot-
probe task. Moreover, they provided evidence that there was a correlation between more
negative N2pc amplitudes and more severe symptoms of anxiety, as measured by the SPI and
STAI (Reutter et al., 2017). Future research incorporating the dot probe test may benefit by
looking at and comparing the N2pc amplitudes in substitution of the RT measures.
Furthermore, resent investigations documented evidence that a pooled FCz/Cz electrode
statistical analysis of N2 amplitudes provide a more complete picture (Iannaccone et al., 2015).
Here, the statistical analysis included the FCz electrode solely and pooling both electrode sites
might be a direction for future investigations.
Conclusions
This thesis sought to test the effect of ABM on trait anxiety and electro-cortical
modulation as quantified by the STAI-T questionnaire and the ΔN2, respectively. The study
investigated these effects by virtue of a multi-session mobile training application that featured
equally valenced stimuli for both control and training groups. The filtered inclusion of
participants provided a sample that had both high levels of trait anxiety and a preexisting
attention bias to threatening stimuli. Although the results did not lend significant support towards
the hypotheses, the investigation bore insight into future avenues of inquiry. Specifically, the
implementation of additional questionnaires such as the Worry Domains Questionnaire (WDQ)

45
and the Penn State Worry Questionnaire (PSWQ) could impart a more granular classification of
associated symptomologies to which an analytical comparison may furnish deeper insights.
Moreover, implementing a flanker paradigm that induces variability in cognitive load may
invoke differences between N2 activity elicited from congruent and incongruent trials that here
has been deemed negligible. In spite of the limitations addressed and the diminished expected
sample size, as a result of an ensuing global pandemic, the investigation demonstrated a subtle
decrease in the post-ΔN2 amplitude for the training group compared to their pre-ΔN2
amplitudes.

46
References
Aday, J., & Carlson, J. M. (2017). Structural MRI-based measures of neuroplasticity in an
extended amygdala network as a target for attention bias modification treatment
outcome. Medical hypotheses, 109(vol.), 6-16.
Anderson, A. K., & Phelps, E. A. (2001). Lesions of the human amygdala impair enhanced
perception of emotionally salient events. Nature, 411(6835), 305-309.
https://www.nature.com/nature/subscribe?gclid=EAIaIQobChM
In9vTuZmV6QIVDdbACh3H4ACpEAAYASAAEgJy5fD_BwE
Anderson, A. K. (2005). Affective influences on the attentional dynamics supporting awareness.
Journal of Experimental Psychology-General, 134(2), 258–
281. https://www.apa.org/pubs/journals/xge/
Bandelow, B., & Michaelis, S. (2015). Epidemiology of anxiety disorders in the 21st century.
Dialogues in Clinical Neuroscience, 17(3), 327.
Barbas, H. (2000). Connections underlying the synthesis of cognition, memory, and emotion in
primate prefrontal cortices. Brain Research Bulletin, 52(5), 319-330.
https://www.journals.elsevier.com/brain-research-bulletin
Bar-Haim, Y., Lamy, D., rin, L., Bakermans-Kranenburg, M.J., & van IJzendoorn, M.H.
(2007). Threat-related attentional bias in anxious and nonanxious individuals: A
Meta-analytic study. Psychological Bulletin, 133, 1–24.
https://www.apa.org/pubs/journals/bul/

47
Bar-Haim, Y. (2010). Research review: Attention bias modification (ABM)--A novel treatment
for anxiety disorders. Journal of Child Psychology and Psychiatry, 51(8), 859.
doi:10.1111/j.1469-7610.2010.02251.x
Beck, A. T., & Clark, D. A. (1997). An information processing model of anxiety: Automatic and
strategic processes. Behaviour Research and Therapy, 35(1), 49-58.
https://www.journals.elsevier.com/behaviour-research-and-therapy
Bishop, S. J., Duncan, J., Brett, M., & Lawrence, A.D. (2004). Prefrontal cortical function
and anxiety: Controlling attention to threat-related stimuli. Nature Neuroscience, 7,
184–188. https://www.nature.com/neuro/
Bishop, S. J. (2007). Neurocognitive mechanisms of anxiety: An integrative account.
Trends in Cognitive Sciences, 11(7), doi.org/10.1016/j
Bishop, S. J. (2009). Trait anxiety and impoverished prefrontal control of attention. Nature
Neuroscience, 12(1), 92-98. doi:10.1038/nn.2242
Bradley, M. M., & Lang, P. J. (1999). Fearfulness and affective evaluations of pictures.
Motivation and Emotion, 23(1), 1-13.
https://explorable.com/motivationandemotion
Bress, j., Meyer, A., & Hajack, G. (2015). Differentiating Anxiety and Depression in Children
and Adolescents: Evidence from Event-Related Brain Potentials, Journal of Clinical
Child & Adolescent Psychology, 44(2), 238-249, DOI:10.1080/15374416.2013.814544
Brosch, T., Pourtois, G., & Sander, D. (2010). The perception and categorization of emotional
stimuli: a review. Cognition & Emotion, 24(3), 377–400.
https://www.tandfonline.com/loi/pcem20

48
Browning, M., Holmes, E. A., & Harmer, C. J. (2010). The modification of attentional bias to
emotional information: A review of the techniques, mechanisms, and relevance to
emotional disorders. Cognitive, Affective, & Behavioral Neuroscience, 10(1), 8-
20.
Buhle, J. T., Silvers, J. A., Wager, T. D., Lopez, R., Onyemekwu, C.,Kober, H., & Ochsner, K.
N. (2014). Cognitive reappraisal of emotion: A meta-analysis of human
neuroimaging studies. Cerebral Cortex, 24(11), 2981e2990.
http://doi.org/10.1093/cercor/bht154.
Carlson, J. M., Reinke, K. S., & Habib, R. (2009). A left amygdala mediated network for rapid
orienting to masked fearful faces. Neuropsychologia, 47(5), 1386-1389.
https://www.journals.elsevier.com/neuropsychologia
Carrasco, M., Harbin, S. M., Nienhuis, J. K., Fitzgerald, K. D., Gehring, W. J., & Hanna, G. L.
(2013). Increased error‐related brain activity in youth with obsessive‐compulsive
disorder and unaffected siblings. Depression and Anxiety, 30(1), 39-
46. https://www.longdom.org/depression-and-anxiety.html
Cisler, J.M., & Koster, E.H. (2010). Mechanisms of attentional biases towards threat in anxiety
disorders: An integrative review. Clinical Psychology Review, 30( ), 203–216.
https://www.journals.elsevier.com/clinical-psychology-review
Compton, R. J. (2003). The interface between emotion and attention: A review of evidence from
psychology and neuroscience. Behavioral and Cognitive Neuroscience Reviews, 2(2), 115-
129. Craske, M. G., Prof, & Stein, M. B., Prof. (2016). Anxiety. The Lancet, 388(10063),
3048-3059. doi:10.1016/S0140-6736(16)30381-6
Dandeneau, S. D., Baldwin, M. W., Baccus, J. R., Sakellaropoulo, M., & Pruessner, J. C. (2007).
Cutting stress off at the pass: Reducing vigilance and responsiveness to social threat by

49
manipulating attention. Journal of personality and social psychology, 93(4), 651.
De Voogd, E. L., Wiers, R. W., Prins, P. J. M., & Salemink, E. (2014). Visual search attentional
bias modification reduced social phobia in adolescents. Journal of Behavior Therapy
and Experimental Psychiatry, 45(2), 252-259.
Dandeneau, S. D., Baldwin, M. W., Baccus, J. R., Sakellaropoulo, M., & Pruessner, J. C. (2007).
Cutting stress off at the pass: reducing vigilance and responsiveness to social threat
by manipulating attention. Journal of Personality and Social Psychology, 93(4),
651. https://www.apa.org/pubs/journals/psp/
Davis, H., Davis, P. A., Loomis, A. L., Harvey, E. N., & Hobart, G. (1939). Electrical reactions
of the human brain to auditory stimulation during sleep. Journal of Neurophysiology, 2,
500-514. https://journals.physiology.org/journal/jn
Davis, M., & Whalen, P. J. (2001). The amygdala: Vigilance and emotion. Molecular
Psychiatry, 6(1), 13e34. http://doi.org/10.1038/sj.mp.4000812.
Dennis, T. A., & Chen, C. (2009). Trait anxiety and conflict monitoring following threat:
An ERP study. Psychophysiology, 46(1), 122-131. doi:10.1111/j.1469-
8986.2008.00758.x
De Voogd, E. L., Wiers, R. W., Prins, P. J. M., & Salemink, E. (2014). Visual search attentional
bias modification reduced social phobia in adolescents. Journal of Behavior Therapy and
Experimental Psychiatry, 45(2), 252-259.
https://www.journals.elsevier.com/journal-of-behavior-therapy-and-
experimental- psychiatry

50
Easterbrook, J. A. (1959). The effect of emotion on cue utilization and the organization of
behavior. Psychological Review, 66, 183–201.
https://www.apa.org/pubs/journals/rev/
Eastwood, J. D., Smilek, D., & Merikle, P. M. (2003). Negative facial expression captures
attention and disrupts performance. Perception & Psychophysics, 65, 352–358.
https://publons.com/journal/49119/perception-and-psychophysics
Endrass, T., Klawohn, J., Schuster, F., & Kathmann, N. (2008). Overactive performance
monitoring in obsessive-compulsive disorder: ERP evidence from correct an
erroneous reactions. Neuropsychologia, 46(7), 1877-1887.
https://www.journals.elsevier.com/neuropsychologia
Etkin, A., Prater, K. E., Hoeft, F., Menon, V., & Schatzberg, A. F. (2010). Failure of anterior
cingulate activation and connectivity with the amygdala during implicit regulation of
emotional processing in generalized anxiety disorder. American Journal of Psychiatry,
167(5), 545-554. https://ajp.psychiatryonline.org/
Etkin, A., & Schatzberg, A. F. (2011). Common abnormalities and disorder-specific
compensation during implicit regulation of emotional processing in generalized
anxiety and major depressive disorders. American Journal of Psychiatry, 168, 968–
978. https://ajp.psychiatryonline.org/
Eysenck, M. W., & Derakshan, N. (1997). Cognitive biases for future negative events as a
function of trait anxiety and social desirability. Personality and Individual
Differences, 22(5), 597-605. https://www.journals.elsevier.com/personality-and-
individual-difference

51
Eysenck,M.W., Derakshan, N., Santos, R., & Calvo, M. G. (2007). Anxiety and cognitive
performance: Attentional control theory. Emotion, 7, 336–353.
https://www.apa.org/pubs/journals/emo/
Fenske, M. J., & Eastwood, J. D. (2003). Modulation of focused attention by faces expressing
emotion: Evidence from flanker tasks. Emotion, 3, 327–343.
https://www.apa.org/pubs/journals/emo/
Fisher, M., Holland, C., Merzenich, M. M., & Vinogradov, S. (2009). Using neuroplasticity-
based auditory training to improve verbal memory in schizophrenia. American
Journal of Psychiatry, 166(7), 805-811. https://ajp.psychiatryonline.org/
Fox, E., Russo, R., Bowles, R., & Dutton, K. (2001). Do threatening stimuli draw or hold visual
attention in subclinical anxiety? Journal of Experimental Psychology: General, 130,
681–700. https://www.apa.org/pubs/journals/xge/
Ghashghaei, H. T., & Barbas, H. (2002). Pathways for emotion: interactions of prefrontal and
anterior temporal pathways in the amygdala of the rhesus monkey. Neuroscience,
115(4), 1261-1279. https://www.jneurosci.org/
Gilmartin, M. R., Balderston, N. L., & Helmstetter, F. J. (2014). Prefrontal cortical
regulation of fear learning. Trends in Neurosciences, 37(8), 455-464.
doi:10.1016/j.tins.2014.05.004
Gray, M. (2004). Geodiversity: valuing and conserving abiotic nature. John Wiley & Sons:
Hoboken, New Jersey.
Hajack, G., MacNamara, A., & Olvet, D. M. (2010). Event-related potentials, emotion, and
emotion regulation: An integrative review. Developmental Neuropsychology, 35(2),
129- 155. doi:10.1080/87565640903526504

52
Hakamata, Y., Lissek, S., Bar-Haim, Y., Britton, J. C., Fox, N. A., Leibenluft, E., Pine, D. S.
(2010). Attention bias modification treatment: A meta-analysis toward the
establishment of novel treatment for anxiety. Biological Psychiatry, 68(11), 982-
990. doi:10.1016/j.biopsych.2010.07.021
Hajcak, G., Franklin, M. E., Foa, E. B., & Simons, R. F. (2008). Increased error-related brain
activity in pediatric obsessive-compulsive disorder before and after treatment.
American Journal of Psychiatry, 165(1), 116-
123. https://ajp.psychiatryonline.org/
Hanoch, Y., & Vitouch, O. (2004). When less is more: Information, emotional arousal and the
ecological reframing of the Yerkes-Dodson law. Theory & Psychology, 14, 427–452/
https://journals.sagepub.com/home/tap
Heeren, A., De Raedt, R., Koster, E.H.W., & Philippot, P. (2013). The (neuro) cognitive
mechanisms behind attention bias modification in anxiety: Proposals based on
theoretical accounts of attentional bias. Frontiers in Human Neuroscience, 7.
https://www.frontiersin.org/journals/human-neuroscience
Himmelheber, A. M., Sarter, M., & Bruno, J. P. (2000). Increases in cortical acetylcholine
release during sustained attention performance in rats. Cognitive Brain Research,
9(3), 313-325. https://www.sciencedirect.com/journal/cognitive-brain-research
Himmelheber, A. M., Sarter, M., & Bruno, J. P. (2001). The effects of manipulations of
attentional demand on cortical acetylcholine release. Cognitive Brain Research,
12(3), 353-370. https://www.sciencedirect.com/journal/cognitive-brain-research

53
Iannaccone, R., Hauser, T. U., Staempfli, P., Walitza, S., Brandeis, D., & Brem, S. (2015).
Conflict monitoring and error processing: new insights from simultaneous EEG–
fMRI. Neuroimage, 105, 395-407.
Kessler, R. C., Berglund, P., Demler, O., Jin, R., Merikangas, K. R., & Walters, E. E. (2005).
Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National
Comorbidity Survey Replication. Archives of General Psychiatry, 62(6), 593-602.
Kolb, B., Mychasiuk, R., Muhammad, A., Li, Y., Frost, D. O., & Gibb, R. (2012). Experience
and the developing prefrontal cortex. Proceedings of the National Academy of
Sciences, 109(Supplement 2), 17186-17193. doi:10.1073/pnas.1121251109
Larson, M. J., & Clayson, P. E. (2011). The relationship between cognitive performance
and electrophysiological indices of performance monitoring. Cognitive,
Affective, & Behavioral Neuroscience, 11(2), 159-171. doi:10.3758/s13415-
010-0018-6
Leal, P. C., Goes, T. C., da Silva, Luiz Carlos Ferreira, & Teixeira-Silva, F. (2017). Trait vs.
State anxiety in different threatening situations. Trends in Psychiatry and
Psychotherapy, 39(3), 147-157. doi:10.1590/2237-6089-2016-0044
LeDoux, J.E. (2000). Emotion circuits in the brain. Annual Review of Neuroscience, 23,
155–184. https://www.annualreviews.org/journal/neuro
LeDoux, J. (2012). Rethinking the emotional brain. Neuron, 73(4), 653e676. http://
dx.doi.org/10.1016/j.neuron.2012.02.004.
Leith, K. P., & Baumeister, R. F. (1996). Why do bad moods increase self-defeating behavior?
Emotion, risk tasking, and self-regulation. Journal of Personality and
Social Psychology, 71, 1250–1267.
https://www.apa.org/pubs/journals/psp/

54
MacLeod, C., & Mathews, A. (1988). Anxiety and the allocation of attention to threat. The
Quarterly Journal of Experimental Psychology, 40(4), 653-670.
https://journals.sagepub.om/home/qjp
MacLeod, C., Rutherford, E., Campbell, L., Ebsworthy, G., & Holker, L. (2002). Selective
attention and emotional vulnerability: assessing the causal basis of their
association through the experimental manipulation of attentional bias. Journal of
Abnormal Psychology, 111(1), 107. https://www.apa.org/pubs/journals/abn/
MacNamara, A., Post, D., Kennedy, A. E., Rabinak, C. A., & Phan, K. L. (2013). Electrocortical
processing of social signals of threat in combat-related post-traumatic stress disorder.
Biological Psychology, 94(2), 441-449.
Månsson, K. N., Carlbring, P., Frick, A., Engman, J., Olsson, C. J., Bodlund, O., & Andersson,
G. (2013). Altered neural correlates of affective processing after internet-
delivered cognitive behavior therapy for social anxiety disorder. Psychiatry
Research: Neuroimaging, 214(3), 229-237.
https://www.journals.elsevier.com/psychiatry-research-neuroimaging
Månsson, K. N., Salami, A., Frick, A., Carlbring, P., Andersson, G., Furmark, T., & Boraxbekk,
C. J. (2016). Neuroplasticity in response to cognitive behavior therapy for social
anxiety disorder. Translational Psychiatry, 6(2), e727-e727.
https://www.nature.com/tp/
Maslowsky, J., Mogg, K., Bradley, B. P., McClure-Tone, E., Ernst, M., Pine, D. S., & Monk, C.
S. (2010). A preliminary investigation of neural correlates of treatment in
adolescents with generalized anxiety disorder. Journal of Child and Adolescent
Psychopharmacology, 20(2), 105-111.

55
https://home.liebertpub.com/publications/journal-of-child-and-
adolescent- psychopharmacology/29
Mathews, A., & Macleod, C. (1994). Cognitive approaches to emotion and emotional
disorders. Annual Review of Psychology, 45, 25–50.
https://www.annualreviews.org/journal/psych
Mathews, A., & MacLeod, C. (2002). Induced processing biases have causal effects on anxiety.
Cognition & Emotion, 16, 331–354.
https://www.tandfonline.com/em20
McDermott JM, Perez-Edgar K, Henderson HA, Chronis-Tuscano A, Pine DS, Fox NA (2009).
A history of childhood behavioral inhibition and enhanced response
monitoring in adolescence are linked to clinical anxiety. Biol Psychiatry 65,
445–448. biologicalpsychiatryjournal.com
Meinhardt, J., & Pekrun, R. (2003). Attentional resource allocation to emotional events: An ERP
study. Cognition & Emotion, 17, 477–500.
https://www.tandfonline.com/loi/pcem20
Meyer, A., Weinberg, A., Klein, D. N., & Hajack, G. (2012). The development of the
error-related negativity (ERN) and its relationship with anxiety: Evidence from
8 to 13 year-olds. Developmental Cognitive Neuroscience, 2(1), 152-161.
doi:10.1016/j.dcn.2011.09.005
Mogg, K., Mathews, A., & Eysenck, M. (1992). Attentional bias to threat in clinical anxiety
states. Cognition & Emotion, 6, 149–159.
https://www.tandfonline.com/loi/pcem20

56
Mogg, K., & Bradley, B. P. (2016). Anxiety and attention to threat: Cognitive mechanisms and
treatment with attention bias modification. Behaviour Research and Therapy, 87, 76-
108. doi:10.1016/j.brat.2016.08.001
Mogg, K., Millar, N., & Bradley, B. P. (2000). Biases in eye movements to threatening facial
expressions in generalized anxiety disorder and depressive disorder. Journal of
Abnormal Psychology, 109(4), 695-704. doi:10.1037/0021-843X.109.4.695
Monk, C. S., Nelson, E. E., McClure, E. B., Mogg, K., Bradley, B. P., Leibenluft, E., et al.
(2006). Ventrolateral prefrontal cortex activation and attentional bias in
response to angry faces in adolescents with generalized anxiety disorder.
American Journal of Psychiatry, 163, 1091–1097.
https://ajp.psychiatryonline.org/
Monk, C. S., Telzer, E. H., Mogg, K., Bradley, B. P., Mai, X., Louro, H. M. C., et al. (2008).
Amygdala and ventrolateral prefrontal cortex activation to masked
angry faces in children and adolescents with generalized anxiety
disorder. Archives of General Psychiatry, 65(5), 568.
http://doi.org/10.1001/archpsyc.65.5.568.
Moser, J. S., Moran, T. P., Schroder, H. S., Donnellan, M. B., & Yeung, N. (2013). On the
relationship between anxiety and error monitoring: A meta-analysis and
conceptual framework. Frontiers in Human Neuroscience, 7, 466.
doi:10.3389/fnhum.2013.00466
Näätänen, R., & Gaillard, A. W. K. (1983). 5 The orienting reflex and the N2 deflection of the
event-related potential (ERP). In Advances in psychology (Vol. 10, pp. 119-141).
North- Holland.
Näätänen, R., & Picton, T. W. (1986). N2 and automatic versus controlled processes.

57
Nelson BD, Jackson F, Amir N, Hajack G. (2004). Single-session attention bias
modification and error-related brain activity. Cognitive, Affective, &
Behavioral Neuroscience.
(15)776-786. https://www.springer.com/journal/13415
New, J., Cosmides, L., & Tooby, J. (2007). Category-specific attention for animals reflects
ancestral priorities, not expertise. Proceedings of the National Academy of
Sciences, 104(42), 16598–16603. https://www.pnas.org/
Nofmann, S. G., Asnaani, A., Vonk, I. J., Sawyer, A. T., & Fang, A. (2012). The efficacy of
cognitive behavioral therapy: A review of meta-analyses. Cognitive Therapy and
Research, 36(5), 427-440.
Ohman, A., Flykt, A., & Esteves, F. (2001). Emotion drives attention: detecting the snake in the
grass. Journal of Experimental Psychology-General, 130(3), 466–478.
Olatunji, B. O., Cisler, J. M., & Deacon, B. J. (2010). Efficacy of cognitive behavioral therapy
for anxiety disorders: a review of meta-analytic findings. Psychiatric Clinics, 33(3),
557- 577.
Ouimet, A.J., Gawronski, B., & Dozois, D.J.A. (2009). Cognitive vulnerability to anxiety: A
review and an integrative model. Clinical Psychology Review, 29, 459–470.
https://www.journals.elsevier.com/clinical-psychology-review
Peck, C. J., & Salzman, C. D. (2014). The amygdala and basal forebrain as a pathway for
motivationally guided attention. Journal of Neuroscience, 34(41), 13757-
13767. https://www.jneurosci.org/
Pessoa, L. (2009). How do emotion and motivation direct executive control? Trends in Cognitive
Sciences, 13(4), 160e166. http://doi.org/10.1016/j.tics.2009.01.006.

58
Pessoa, L., & Adolphs, R. (2010). Emotion processing and the amygdala: From a ‘lowroad’ to ‘many
roads’ of evaluating biological significance. Nature Reviews
Neuroscience, 11(11), 773e783. http://doi.org/10.1038/nrn2920.
Pine, D. S., Helfinstein, S. M., Bar-Haim, Y., Nelson, E., & Fox, N. A. (2009). Challenges in
developing novel treatments for childhood disorders: Lessons from research on
anxiety. Neuropsychopharmacology, 34, 213–228. https://www.nature.com/npp/
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology,
32(1), 3-25. https://journals.sagepub.om/home/qjp
Quirk, G.J., & Mueller, D. (2008). Neural mechanisms of extinction learning and retrieval.
Neuropsychopharmacology, 33.
https://www.nature.com/npp/
Rapee, R. M., & Heimberg, R. G. (1997). A cognitive-behavioral model of anxiety in social
phobia. Behaviour Research and Therapy, 35(8), 741-756.
https://www.journals.elsevier.com/behaviour-research-and-therapy
Reutter, M., Hewig, J., Wieser, M. J., & Osinsky, R. (2017). The N2pc component reliably
captures attentional bias in social anxiety. Psychophysiology, 54(4), 519-527.
Riesel, A., Endrass, T., Kaufmann, C., & Kathmann, N. (2011). Overactive error-related brain
activity as a candidate endophenotype for obsessive-compulsive disorder: evidence
unaffected first-degree relatives. American Journal of Psychiatry, 168(3), 317-
324. https://ajp.psychiatryonline.org/
Riesel, A., Weinberg, A., Endrass, T., Meyer, A., & Hajack, G. (2013). The ERN is the ERN is
the ERN? Convergent validity of error-related brain activity across different tasks.
Biological Psychology, 93(3), 377–385. https://doi.org/10.1016/j.biopsycho.2013.04.007

59
Roy A. K., Shehzad Z., Margulies D. S., Kelly A. M. C., Uddin L. Q., & Gotimer, K.(2009).
Functional connectivity of the human amygdala using resting state fMRI.
Neuroimage, 45, 614–626. https://www.journals.elsevier.com/neuroimage
Sarter, M., Bruno, J. P., & Givens, B. (2003). Attentional functions of cortical cholinergic inputs:
what does it mean for learning and memory? Neurobiology of Learning and Memory,
80(3), 245-256.
https://www.journals.elsevier.com/neurobiology-of-
learning-and-memory
Seligman, M. E. (1971). Phobias and preparedness. Behavior Therapy, 2(3), 307–320. Retrieved
from https://effectivechildtherapy.org/therapies/what-is-behavior-therapy/
Sergerie, K., Chochol, C., & Armony, J. L. (2008). The role of the amygdala in
emotional processing: A quantitative meta-analysis of functional
neuroimaging studies. Neuroscience & Biobehavioral Reviews, 32(4),
811e830. http://doi.org/10. 1016/j.neubiorev.2007.12.002.
Simpson, J. R.,O¨ ngu¨ r, D., Akbudak, E., Conturo, T. E., Ollinger, J. M., Snyder, A. Z., et al.
(2000). The emotional modulation of cognitive processing: An fMRI study. Journal
of Cognitive Neuroscience, 12, 157–170.
https://www.mitpressjournals.org/journal/jocn
Sotres-Bayon, F., Cain, C. K., & LeDoux, J. E. (2006). Brain mechanisms of fear
extinction: Historical perspectives on the contribution of prefrontal
cortex. Biological Psychiatry, 60(4), 329e336.
http://dx.doi.org/10.1016/ j.biopsych.2005.10.012.
Shin, L. M., & Liberzon, I. (2010). The neurocircuitry of fear, stress, and anxiety

60
disorders. Neuropsychopharmacology, 35(1), 169e191.
http://dx.doi.org/10.1038/ npp.2009.83.
Stevenson, R. A., Mikels, J. A., & James, T. W. (2007). Characterization of the affective norms
for English words by discrete emotional categories. Behavior Research Methods,
39(4), 1020-1024. https://www.springer.com/journal/13428
Sylvain, R., Gilbertson, H., & Carlson, J. M. (2020). Single session positive attention bias
modification training enhances reward-related electrocortical responses in
females. International Journal of Psychophysiology, 156, 10-17.
https://doi.org/10.1016/j.ijpsycho.2020.07.002
Taylor, J. M., & Whalen, P. J. (2015). Neuroimaging and anxiety: The neural substrates
of pathological and non-pathological anxiety. Current Psychiatry Reports,
17(6), 49. http://dx.doi.org/10.1007/s11920-015-0586-9.
Thigpen, N. N., Gruss, L. F., Garcia, S., Herring, D. R., & Keil, A. (2018). What does the
dot‐probe task measure? A reverse correlation analysis of electrocortical
activity.
Psychophysiology, 55(6), e13058-n/a. doi:10.1111/psyp.13058
Thomas, S. J., Johnstone, S. J., & Gonsalvez, C. J. (2007). Event-related potentials during an
emotional stroop task. International Journal of Psychophysiology, 63(3), 221-
231. doi:10.1016/j.ijpsycho.2006.10.002
Tversky, A., & Kahneman, D. (1981). The framing of decisions and the psychology of choice.
Science, 211(4481), 453-458.
Van Bockstaele, B., Verschuere, B., Tibboel, H., De Houwer, J., Crombez, G., & Koster, E. H.
(2014). A review of current evidence for the causal impact of attentional bias on
fear and

61
anxiety. Psychological Bulletin,
Van Veen, V., & Carter, C. S. (2002). The anterior cingulate as a conflict monitor: FMRI and
ERP studies. Physiology & Behavior, 77(4), 477-482.
doi:10.1016/S0031-9384(02)00930-7
Vuilleumier, P., Richardson, M. P., Armony, J. L., Driver, J., & Dolan, R. J. (2004). Distant
influences of amygdala lesion on visual cortical activation during emotional face
processing. Nature Neuroscience, 7(11), 1271-1278.
https://www.nature.com/neuro/
Vuilleumier, P., & Driver, J. (2007). Modulation of visual processing by attention and
emotion: Windows on causal interactions between human brain regions.
Philosophical Transactions of the Royal Society B: Biological Sciences,
362(1481), 837e855. http://doi.org/10.1098/rstb.2007.2092
Waters, A. M., Pittaway, M., Mogg, K., Bradley, B. P., & Pine, D. S. (2013). Attention training
towards positive stimuli in clinically anxious children. Developmental Cognitive
Neuroscience, 4, 77-84.
https://www.journals.elsevier.com/developmental-cognitive-neuroscience
Waters, A. M., Zimmer-Gembeck, M. J., Craske, M. G., Pine, D. S., Bradley, B. P., & Mogg,
(2015). Look for good and never give up: A novel attention training treatment
for childhood anxiety disorders. Behaviour research and therapy, 73, 111-123.
Waters, A. M., Zimmer-Gembeck, M. J., Craske, M. G., Pine, D. S., Bradley, B. P., & Mogg, K.
(2016). A preliminary evaluation of a home-based, computer-delivered attention

62
training treatment for anxious children living in regional communities.
Journal of Experimental Psychopathology, 7(3), 511-527.
Whitfield-Gabrieli, S., Thermenos, H. W., Milanovic, S., Tsuang, M. T., Faraone, S. V.,
McCarley, R. W., & Wojcik, J. (2009). Hyperactivity and hyperconnectivity
of the default network in schizophrenia and in first-degree relatives of
persons with schizophrenia. Proceedings of the National Academy of
Sciences, 106(4), 1279-1284. https://www.pnas.org/
Williams, J. M. G., Mathews, A., &MacLeod, C. (1996). The emotional Stroop task and
psychopathology. Psychological Bulletin, 120, 3–24.
https://www.apa.org/pubs//bul/
Williams, J. M., Watts, F. N., MacLeod, C., & Mathews, A. (1988). Cognitive psychology and
emotional disorders. Chichester, U.K.: John Wiley & Sons.
Wood, J., Mathews, A., & Dalgleish, T. (2001). Anxiety and cognitive inhibition. Emotion, 1,
166–181. https://www.apa.org/pubs/journals/emo/
Weinberg, A., Olvet, D. M., & Hajcak, G. (2010). Increased error-related brain activity in
generalized anxiety disorder. Biological Psychology, 85(3), 472-480.
https://www.simplypsychology.org/biological-psychology.html
Weinberg, A., Klein, D. N., & Hajcak, G. (2012). Increased error-related brain activity
distinguishes generalized anxiety disorder with and without comorbid major
depressive disorder. Journal of Abnormal Psychology, 121(4), 885.
https://www.apa.org/pubs/journals/abn/
Weumier, P. (2005). How brains beware: Neural mechanisms of emotional
attention. Trends in Cognitive Science, 9, 585e594.
http://dx.doi.org/10.1016

63
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). The neural basis of error detection Conflict
monitoring and the error-related negativity. Psychological Review, 111(4), 931-
959. doi:10.1037/0033-295X.111.4.931
Zambrano-Vazquez, L., & Allen, J. J. B. (2014). Differential contributions of worry, anxiety, and
obsessive-compulsive symptoms to ERN amplitudes in response monitoring and
reinforcement learning tasks. Neuropsychological, 61, 197-209
doi:10.1016/j.neuropsychologia.2014.06.023

64
Appendix A
Figure 1. Example of the modified flanker task paradigm.
Figure 2. 10-20 Electrode map highlighting FCz (electrode 4)
Negative
Neutral
1000 ms 100 ms Until Response 1000 msse
Figure 3: Example of the dot-probe task during ABM training displaying a threatening (left) and neutral face (right).
+
+ +
+
+
+
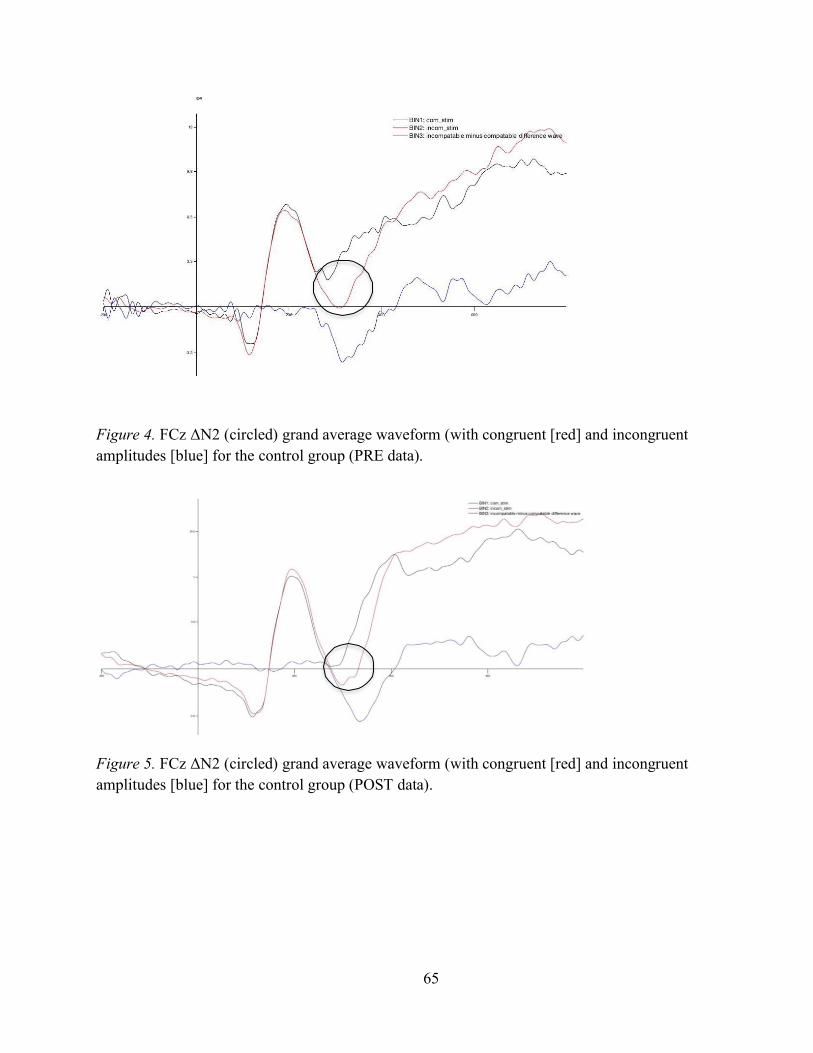
65
Figure 4. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the control group (PRE data).
Figure 5. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the control group (POST data).

66
Figure 6. Figure 4. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the experimental group (PRE data; i.e., pre 6-week ABM training).
Figure 7. FCz ΔN2 (circled) grand average waveform (with congruent [red] and incongruent amplitudes [blue] for the experimental group (POST data; i.e., post 6-week ABM training).

67
Figure 8. Correlation between pre-training self-reported STAI – T score and pre-training ΔN2 amplitude (in µV) across all participants.
Figure 9. Correlation between pre-training self-reported STAI – T score and pre-training incompatible ΔN2 amplitude (in µV) across all participants.
Correlation between pre-STAI scores and Pre ΔN2 amplitudes 80
60
40
20
0 STAI-T scores
Differences in STAI-T scores comparing pre vs post ABM 60
55 50 45 40 35 30 25 20 15 10
5 0
52.57142857 52
Pre ABM
Post ABM
1 Pre vs. Post ABM training
Pre
ΔN2
Inco
mpa
tible
(a
mpl
itude
s in
µV)
Self-
repo
rted
STA
I-T sc
ores
Figure 10. Evidence of the correlation (or lack thereof) between pre-training STAI-T score and pre-training ΔN2 amplitudes (in µV) in the experimental group.

68
Figure 11. Although not significant, this chart provides evidence that ABM may correlate with the ΔN2, suggesting it may be related to conflict monitoring.
Pre ΔN2 amplitudes pre vs. post ABM training
-2.8
-2.7
-2.6 Pre-ABM
Post-ABM
-2.5
-2.4 Session (pre vs post)
ΔN2
ampl
itude
s in
µV

NORTHERN MICHIGAN
UNIVERSITY
OFFICE OF GRADUATE EDUCATION AND RESEARCH
1401 Presque Isle Avenue Marquette, "11 49855-5301
906-227-2300906-227-2315
www.nmu.edu/graduatestudies MEMORANDUM
TO:
DATE:
FROM:
RE:
Joshua Carlson Lin Fang Department of Psychological Science
Taylor Susa Hayley Gilbertson Katie Elwell Haley Gaboury Justine Nelson Jacquie Cammarata Hannah Kaull Natalie Strand Nathan Schmitz Mason Steinhauer Madeline Voltz Diego Ulloa Abigail Schaedig Abbey Zigarac Analise Osgood
Sep tern bee 5, 2019 \ cjJ Lisa Schade Eckert 'o. Dean of Graduate Education and Research
Modification to HS 13-555 Original !RB Approval Date: 11/12/2013 Modification Approval Date: 9/5/2019 "Characteristics of Attention Bias Modification - New Title: "R 15MH 1109051: Neuroplasticity in an Extended Amygdala Network as a Target Mechanism for Attention Bias Modification Outcome"
Your modification for the project "Characteristics of Attention Bias Modification - New Title: "RI 5MH 1109051: Neuroplasticity in an Extended Amygdala Network as a Target Mechanism for Attention Bias Modification Outcome" has been approved by the Northern Michigan University Institutional Review Board. Please include your proposal number (HS 13-555) on all research materials and on any correspondence regarding this project.
Any additional personnel changes or revisions to your approved research plan must be approved by the IRB prior to implementation. Unless specified otherwise, all previous requirements included in your original approval notice remain in effect.
If you have any questions, please contact the IRB at [email protected].
Appendix B
69