effects of neuropeptide y on single neuronal firing
TRANSCRIPT

1
EFFECTS OF NEUROPEPTIDE Y ON SINGLE NEURONAL FIRING
PATTERNS IN A GENETIC RAT MODEL OF ABSENCE EPILEPSY
Arun Gandrathi
Submitted in total fulfilment of the requirements of the degree of Doctor of Philosophy
August 2016
The Department of Medicine, Royal Melbourne Hospital The University of Melbourne

2

3
ABSTRACT
Absence seizures are most common type of generalized seizures where brain
goes into abnormal pathological rhythm characterized by bilateral and
synchronous 3Hz spike and wave discharges on EEG. Currently available
anti-epileptic drugs do not have adequate seizure control and have adverse
side effects, which increased the desire for development of novel therapeutic
options for treating absence seizures. Previous studies demonstrated that
neuropeptide Y (NPY) successfully suppresses absence seizures in genetic rat
model of absence epilepsy but the underlying neuronal mechanisms for
seizure suppression still remain unknown. Recent developments in
electrophysiology enabled the quantification of neuronal firing patterns in
vivo and many researchers have already demonstrated differential
characteristic firing patterns of neurons in thalamo cortical circuit in the brain
that plays a critical role in absence seizures. This study was designed to
investigate the effects of intracerebroventricular (ICV) and locally
administered NPY on neuronal firing patterns in thalamocortical structures,
which play a critical role in the generation of absence like seizures in GAERS
as well as effects on seizure induction threshold in the cortical S2 region.
Methods
All experiments were conducted on 8-12 weeks old male GAERS rats. This
involved performing in vivo electrophysiological recordings under neurolept
anaesthesia. In these experiments single neuronal firing activity was recorded
from different regions of thalamocortical circuit. Once stable
electrophysiological recordings are achieved, NPY was infused both ICV and
focally at the recording site and alterations in the neuronal firing patterns were
assessed before and after the drug intervention. In another experiment cortical
region S2 was stimulated by external electrical stimulation to induce absence

4
like seizure and effects of ICV administration of NPY on seizure triggering
threshold in the S2 region was assessed in GAERS.
Results
NPY suppressed absence seizures in GAERS by reducing total seizure length
and percentage of time spent in seizures under neurolept analgesia. In
addition, we also demonstrate that NPY mediated reductions in the seizure
levels (absence) were associated with increased firing frequency of the NRT
neurons interictally in these rats. ICV and focal administration of NPY
increased the mean firing frequency of the NRT neurons in interictal periods.
Control experiments were performed with saline injections .The saline
infusion data showed no change in the neuronal firing patterns of the NRT
cells after ICV and focal administered. Furthermore NPY decreased the
waveform correlation of local field potentials between NRT and cortical
regions. In addition to effect on neuronal firing activity, NPY significantly
increased the seizure induction threshold for triggering seizures by external
electrical stimulation in the S2 region of cortex in GAERS. Control
experiments with saline did not alter seizure-inducing threshold in GAERS.
Seizures were not triggered in response to similar external stimulations in
non- epileptic control rats.
Conclusions
In conclusion, NPY is an endogenous neuropeptide which suppresses absence
seizures. These results suggest that NPY mediates its anti-epileptic response
through alterations in firing patterns of NRT neurons. It also strengthens the
argument that S2 might be focus for origin of SWDs and implicates the NRT
as the key target structure of the thalamocortical circuit by which NPY
suppresses seizures. These findings are associative in nature and further
investigation using NPY receptor Y2 and Y5 subtype selective agents may
clarify cellular molecular reasons explaining the antiepileptic effects of NPY.

5
Finally, NPY associated mechanisms may provide novel therapeutic options
for absence seizures and other generalized epilepsy syndromes.

6
DECLARATION
This is to certify that:
The thesis comprises only my original work towards the PhD
Due acknowledgement has been made in the text to all other material used,
The thesis is fewer than 100,000 words in length, exclusive of tables, maps,
bibliographies and appendices.
Signed…………………………… Date………………………..

7
PUBLICATIONS/CONFERENCES Manuscripts arising from this thesis: Arun Gandrathi, Thomas Zheng, Patrick O’Brien, Idrish Ali, Terence J.
O’Brien and Christopher R French (2013) An in vivo technique for
investigating electrophysiological effects if centrally administered drugs on
single neurons and network behaviour. Published in Journal of Neuroscience
methods.
Zheng TW, O'Brien TJ, Morris MJ, Reid CA, Jovanovska V, O'Brien P, van
Raay L, Gandrathi AK, Pinault D (2012) Rhythmic neuronal activity in S2
somatosensory and insular cortices contribute to the initiation of absence-
related spike-and-wave discharges. Published in Epilepsia
Arun Gandrathi, Idrish Ali, Thomas Zheng, Chris French, Margaret J
Morris, Terence J O’Brien Suppression of absence seizures in the genetic rat
model by Neuropeptide Y is associated with effects in thalamocortical circuit.
(Ready for submission)

8
Conference proceedings:
Arun Gandrathi, Terence J O’Brien, Margaret Morris, Didier Pinault, Chris
French Suppression of absence seizures by neuropeptide Y is associated with
the increase in the interictal firing frequency of NRT neurons. Melbourne
Health Research Week, Melbourne, Australia (Poster).
Arun Gandrathi, Terence J O’Brien, Margaret Morris, Didier Pinault, Chris
French Suppression of absence seizures by neuropeptide Y is associated with
the increase in the interictal firing frequency of NRT neurons. Asian ocean
epilepsy congress, Melbourne, Australia (Poster).
Arun Gandrathi, Terence J O’Brien, Margaret Morris, Didier Pinault, Chris
French Suppression of absence seizures by neuropeptide Y is associated with
the increase in the interictal firing frequency of NRT neurons. Australian
neuroscience society, Auckland, New Zealand (oral presentation).

9
AWARDS
2011: Melbourne abroad travel scholarship
2008: Melbourne international fee remission scholarship
2008: Melbourne international research scholarship

10
Acknowledgment
It would have been never be possible to finish a research project like this
without the guidance, motivation encouragement and continuous support of
few people who I owe immense heartfelt gratitude.
I would like to express my deepest gratitude to my supervisors, Prof.
Margaret Morris, Prof. Terence O’Brien and Chris French for their excellent
guidance, caring, patience, and providing me with an excellent atmosphere for
doing research. I would like to express my special appreciation and thanks to
my supervisor Professor Margaret Morris for her continuous support and
care. Though she was in Sydney, she never lost contact and continuously
monitored my progress with frequent visits, and pouring her valuable inputs
and immense knowledge into my research. I consider myself luckiest to find
such a mentor.
I would like to express my sincere gratitude to my supervisor Professor
Terence J O’Brien for his support, patience, motivation, enthusiasm, immense
knowledge and for all that I have learned from him and his guidance and
support in all stages of my research. I could not have imagined a better
supervisor for my Ph.D study. I would like to thank you for encouraging my
research and for allowing me to grow personally and professionally during
this period.

11
Words fail me when I yearn to depict my profoundest feelings of gratitude my
other supervisor Dr. Chris French. It was my great fortune that I was under
supervision of Dr. Chris French. It would not be an exaggeration to say that
he spent more sleepless nights over my thesis than myself. He made me to
write and rewrite and he was bent upon pointing out my mistakes, so that I
can root them out. I express my gratitude from the bottom of my heart for his
constant support. From the bottom of my heart I know this would not be
enough, some gestures cannot be repayed.
Thanks to Dr. Nigel C. Jones, Dr. Kim Powell and Dr. Thomas Zheng for
their innovative suggestions, helping me to improve the quality of this work.
Thanks to Dr. Idrish Ali, Leena van Raay, and Patrick O.Brien for their
technical assistance.
A special thanks to all past and present members of the O’Brien Lab for
baring me very patiently and always encouraging me during my thesis and
providing me a great atmosphere to work, always felt like home. I will always
cherish your friendship and support.
I wish to thank the Department of Medicine that provided me with the facility
to conduct research as well as to the University of Melbourne to provide me
with financial assistance and scholarships including MIFRS, MIRS. I also

12
acknowledge animals used in this study for their noble sacrifice for benefit of
science.
I owe a debt of gratitude to Dr. Raju Yerra and his family. Without them I
may not have gathered enough courage to travel this far to pursue my PhD. I
would also like to thank for their love and support for all these years.
Personally, I would like to thank my parents Venkateshwarlu and Vijaya, my
brother Ranadheer, sister Priyanka krsihna, my uncle Dr. Srinivasulu, other
friends and family members for their unconditional love and support, who had
faith in made me what I am today.

13
List of Abbreviations
AP Action potential
APFP AP firing proportion per EEG spike
AMPA α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid
cDNA Complementary Deoxyribonucleic acid
CF Cycle frequency
CNS Central nervous system
CSF Cerebro spinal fluid
DAB 3,3 P-diaminobenzidine
DNA Deoxyribonucleic acid
EEG Electroencephalogram
EPSP excitatory post-synaptic potential
FC Febrile convulsions
FFT Fast Fourier transforms
GABA ɣ- Aminobutyric acid
GAERS Genetic absence epilepsy rats from Strasbourg
HCN Hyperpolarization gated cation
IC Insular cortex
ICV Intracerebro ventricular
IBF Intra burst frequency

14
ILAE International League against Epilepsy
IPSP Inhibitory post-synaptic potential
KA Kainic acid
LFP Local field potential
LTP Long term potentiation
MAP Mean number of AP’s per EEG spike
MAPB Mean number of AP’s per burst
MAPF Mean action potential firing frequency
MDS Mean duration of seizure
MRI Magnetic resonance imaging
mRNA Messenger Ribonucleic acid
MTLE Mesial temporal lobe epilepsy
MxAPB Maximum number of AP’s per burst
NEC Non-epileptic control
NMDA N-methyl D-aspartate
NPY Neuropeptide Y
NRT Nucleus of reticular thalamus
PABD Percentage of AP’s in burst
PB Percentage of burst with EEG spike
PBS Phosphate buffered saline

15
PFA Paraformaldehyde
PP Pancreatic peptide
PTS Proportion of recording time of seizures
PTZ Pentylenetetrazol
RNA Ribonucleic acid
SEM Standard error of mean
SI Synchronization index
SUDEP Sudden unexpected death due to epilepsy
SWDs Spike and wave discharges
TLE Temporal lobe epilepsy
TLS Total length of seizures
VB Ventrobasal
WAG/Rij Wistar albino glaxo kept in Rijswijk

16
TABLE OF CONTENTS
CHAPTER 1: LITERATURE REVIEW AND RATIONALE FOR THE
PRESENT RESEARCH
1.1 Introduction…………………………………………….. 1
1.1.1 Epilepsy………………………………………………... 1
1.1.1.1 Prevalence of epilepsy…………………………………. 1
1.1.1.2 Incidence of epilepsy………………………………….. 2
1.1.1.3 Aetiology of epilepsy…………………………………. 2
1.1.1.4 Mortality……………………………………………… 3
1.1.2 Classification of epilepsy……………………………… 3
1.1.2.1 Focal epilepsy………………………………………… 4
1.1.2.2 Generalised epilepsy…………………………………... 5
1.1.3 Prognosis of epilepsy …………………………………. 6
1.1.3 Table of relationship between type of epilepsy,
Prognosis and treatment………………………………. 7
1.2 Absence seizures………………………………………. 8
1.2.1 Classification of absence seizures………………….. … 8
1.2.2 Aetiology of absence seizures………………………...... 9
1.2.3 Age of onset...………………………………………….. 9
1.2.4 Morbidity and mortality………………………………... 10
1.2.5 Electroencephalography ………..……………………… 10
1.2.6 Thalamocortical circuits ad mechanism of absence
seizures………………………………………………... 11
1.2.6.1 Anatomy and physiology of thalamocortical circuit…... 11
1.2.7 Animal models of absence seizures…….……………… 16
1.2.7.1 The genetic absence epilepsy rats from Strasbourg
(GAERS)…………………….………………………… 18

17
1.2.8 Pharmacological treatment for absence epilepsies……… 20
1.3 Introduction to NPY and its receptors.………………...... 20
1.3.1 Discovery of NPY………………………………….….... 21
1.3.2 Primary structure of NPY……………………………..... 21
1.3.3 Cloning of NPY receptors……………………………..… 22
1.3.3.1 NPY Y1 receptor subtype……………………………..… 22
1.3.3.2 NPY Y2 receptor subtype……………………………….. 23
1.3.3.3 NPY Y3 receptor subtype……………………………….. 23
1.3.3.4 NPY Y4 receptor subtype……………………………….. 23
1.3.3.5 NPY Y5 receptor subtype……………………………...... 23
1.3.3.6 NPY Y6 receptor subtype………………………………. 24
1.3.4 Distribution of NPY in central and peripheral nervous
system……………………………………………….….. 24
1.3.5 Biological effects of NPY………………………………. 25
1.3.6 Effect of NPY of Seizures……………………………… 26
1.3.7 NPY and focal epilepsies………………………………. 26
1.3.7.1 NPY and focal epilepsies: in vivo studies………….. …. 26
1.3.7.2 NPY and focal epilepsies: in vitro studies……………… 27
1.3.7.3 NPY and focal epilepsies: human studies……,…………. 29
1.3.8 NPY and generalized epilepsy……………………….….. 30
1.4 Conclusion of literature review…………………………. 30
1.5 Rationale for this study…………………………..……… 31
1.6 Research questions…………………………………..…… 33
1.7 Hypotheses………………………………………………. 34
CHAPTER 2. MATERIALS AND METHODS
2.1 Study design…………………………………………….. 36
2.2 Animals………………………………………………........ 36

18
2.3 Anaesthesia and surgery…………………………………. 37
2.3.1 Anaesthesia………………………………………………...37
2.3.2 Penile vein catheterization……………………………........ 37
2.3.3 Tracheotomy………………………………………………. 38
2.3.4 EEG electrode implantation…………………..……………39
2.3.5 Electrophysiology………………………………………….40
2.3.6 ICV drug infusion…………………………………………. 43
2.3.7 Juxtacellular labelling…………………………………….. 44
2.3.8 Perfusion and Histology…………………………………... 45
2.4 Focal injection studies……………………………….......... 47
2.5 Cortical stimulation studies…………………………….…. 48
2.6 Data acquisition and analysis……………………………... 51
2.6.1 Autocorrelation analysis………………………………… ..55
2.6.2 Cross correlation analysis………………………………….56
2.6.3 Waveform correlation……………………………………...57
CHAPTER 3. EFFECTS OF ICV ADMINISTERED NPY ON EEG AND
NEURONAL FIRING PATTERNS IN THALAMOCORTICAL
CIRCUIT IN GENETIC RAT MODEL OF ABSENCE EPILEPSY.
3.1 Introduction………………………………………….........59
3.2 Materials and methods…………………………………….62

19
3.2.1 Anaesthesia …………………………………………….…....62
3.2.2 Surgery……………………………………………………….63
3.2.3 Electrophysiology………………………………………........63
3.2.4 ICV drug infusions……………………………………….….64
3.2.5 Juxtacellular labelling and histology………………………...64
3.3 Data acquisition and analysis…………………………….….65
3.4 Statistical analysis……………………………………….…..66
3.5 Results ………………………………………………........…66
3.5.1 Effect of NPY on Seizures…………………………………..66
3.5.2 EFFECT of NPY on neuronal firing patterns……………….68
3.5.3 Effect of NPY on rhythmicity and synchronization in
neuronal firing of NRT cells…………………………….......76
3.5.4 Effect of ICV administered NPY on the waveform correlation
between cortical EEG and local field potentials of NRT……78
3.6 Discussion………………………………………………….....80
CHAPTER 4. EFFECTS OF NPY FOCAL INFUSIONS ON SINGLE
NEURONAL FIRING PATTERNS IN RETICULAR THALAMILC
NUCLEUS CELLS IN A GENETIC RAT MODEL OF ABSENCE
EPILEPSY (GAERS).
4.1 Introduction……………………………………………….….88
4.2 Methods…………………………………………...……….…90
4.2.1 Anaesthesia and surgery……………………………..............90

20
4.2.2 EEG electrode implantation……………………………….....90
4.2.3 Electrophysiology………………………………………..…....91
4.2.4 Juxtacellular labelling and histology……………………….....93
4.2.5 Data acquisition and analysis……………………………..…..93
4.3 Results……………………………………………………..….94
4.4 Discussion………………………………………………….....96
CHAPTER 5. EFFECT OF ICV ADMINSTERED NPY ON SEIZURE
INDUCING THRESHOLD UPON EXTERNAL ELECTRICAL
STIMULATION IN S2 REGION IN GAERS.
5.1 Introduction………………………………………………..…100
5.2 Methods……………………………………………………....101
5.2.1 Anaesthesia and surgery……………………………….….….102
5.2.2 EEG electrode and ICV cannula implantation……………….102
5.2.3 Cortical stimulation and seizure induction…………………...102
5.2.4 Juxtacellular labelling and histology………………………....104
5.2.5 Data acquisition and analysis………………………….……..104
5.3 Results……………………………………………………..…104
5.4 Discussion………………………………………………..…..106
CHAPTER 6. GENERAL DISCUSSION AND CONCLUSION
6.1 Key findings of this study……………………………………111
6.1.1 Effect of NPY on seizures……………………………………112

21
6.1.2 Effect of NPY on interictal Neuronal firing patterns …….…..112
6.1.3 Effect of NPY on the waveform correlation between
cortical EEG and local field potentials of NRT ……….……..113
6.1.4 Effects of Focal administration of NPY on neuronal
firing pattern in NRT region …………………………………..113
6.1.5 Effects of NPY on Seizure induction threshold…………..…...113
6.2 Integration and interpretation of results…………….…...........114
6.3 Limitations of this study and experimental considerations…...119
6.3.1 Limitations in ICV NPY infusion study………………………119
6.3.2 Limitations in focal injection study…………………………...120
6.3.3 Limitations in cortical stimulations study…………………….120
6.4 Future research directions…………………………………….121
6.4.1 Investigating the particular receptor subtype with predominant
seizure suppression effect (ICV and focal administration)……121
6.4.2 Investigating the effect of NPY and its subtypes at network
level……………………………………………………………121
6.4.3 To investigate the effect of NPY and its subtypes on neuronal firing
patterns of freely moving rats…………………………….…...122
6.5 Other therapeutic options for absence epilepsy………..……..122
6.6 Final conclusion…………………………………….………...123
LIST OF REFERENCES……………………………………………..125

22
LIST OF FIGURES
Figure 1.1 Classification of epilepsy according to ILAE
Figure 1.2 Focal or partial seizures
Figure 1.3 Generalized seizures
Figure 1.4 3 Hz SWDs of human absence seizures
Figure 1.5 Thalamocortical circuit
Figure 1.6 Thalamocortical circuit and the thalamocortical interaction
Figure 1.7 Physiology of relay or tonic mode of firing in thalamocortical system
Figure 1.8 Physiology of oscillatory or burst firing mode in thalamocortical circuit
Figure 1.9 Transition from relay mode of firing to oscillatory mode along with EEG.
Figure 1.10 GAERS rats displaying 5-9Hz spike and wave discharges on EEG
Figure 1.11 Molecular structure of NPY and sequence homology between amino acid sequence of NPY, PPY and PP
Figure 2.1 Co-ordinates for the implantation of EEG electrodes, ICV cannula and co-ordinates for craniotomies.
Figure 2.2 Protocol for paired juxtacellular recordings with pictures of labelled neurons
Figure 2.3 Time line for Electrophysiological recordings and ICV NPY perfusions
Figure 2.4 Protocol for Juxtacellular labelling using neurobiotin
Figure 2.5 Experimental design for focal injection of NPY on recording neurons in vivo

23
Figure 2.6 Experimental design for cortical stimulations in S2 of somatosensory cortex
Figure 2.6.1 NRT neuron firing during non-seizing period
Figure 2.6.2 NRT neuron firing during seizure
Figure 2.7 Kanoeke’s program for assessing rhythmicity in the neuronal firing
Figure 2.8 Assessing synchronicity in the neuronal firing using cross correlation and synchronization index
Figure 2.9 Determining the correlation between field potentials of two different regions using waveform correlation in Spike 2.
Figure 3.1 Time line for paired juxtacellular recordings
Figure 3.2 Results of effect of NPY on Seizures
Figure 3.3 Results of effect of NPY on single neuronal firing patterns
Figure 3.3.1 Burst of action potentials during an EEG spike at a high resolution
Figure 3.3.2 Illustration of burst of action potentials during an EEG spike at a low- medium resolution
Figure 3.4.1 Interictal % burst of NRT cells
Figure 3.4.2 Interictal mean AP/Burst of NRT cells
Figure 3.4.3 Interictal maximum number of AP/Burst in NRT cells
Figure 3.4.4 Interictal intra burst frequency of the NRT cells
Figure 3.5.1 Interictal AP firing portions of NRT cells
Figure 3.5.2 Interictal AP/spike of NRT Cell
Figure 3.5.3 Ictal %burst firing/SPIKE of NRT cell
Figure 3.5.4 ICTAL intra burst frequency of NRT cell
Figure 3.6 Effect of NPY on rhythmicity in the neuronal firing

24
Figure 3.7 Effect of NPY on synchronicity of neuronal firing
Figure 3.8 Effect of NPY on waveform correlation
Figure 3.9 Possible mechanism of seizure suppression and increase in the interictal neuronal firing frequency of NRT neurons in GAERS
Figure 4.1 Experimental design for focal injection of NPY
Figure 4.2 Experimental time line for electrophysiology recordings in focal infusion studies
Figure 4.3 the results of focal NPY injections on the single neuronal firing patterns
Figure 4.4 Focal infusion studies showing the firing frequency of NRT neurons before and after focal administration of NPY
Figure 5.1 Three main structures of thalamocortical circuit involved in the generation of absence seizures (Experimental design for cortical stimulation protocol)
Figure 5.2 the results of ICV administration of NPY on the seizure inducing threshold in the S2 region (Effects of NPY of seizure inducing threshold in S2)
Figure 5.3 Showing the possible mechanism for the increase in seizure
induction threshold
Figure 6.1 Schematic diagram of neuron illustrating the locations of Y1, Y2
and Y5 receptors
Fig 6.2 The above figure illustrates the possible mechanism for seizure
suppression.

25
Chapter 1
Literature Review
1.1 Introduction:
1.1.1 EPILEPSY
Epilepsy is a neuro pathological disorder characterized by recurrent
seizures because of abnormal and irregular neuronal activity in the brain. A
seizure is a consequence of abnormal and irregular neuronal discharges in
the brain, it is also referred as a convulsion or fit. Seizure episodes are
characterized by impaired consciousness, loss or excess muscular activity,
or an abnormal sensation. The excessive neuronal discharges may be
restricted to small region of the brain (a lesion or focus) resulting in partial
seizures or focal seizures or initiate in small area (focus) and spread to the
whole brain resulting in generalized seizures. Seizures may occur during
any time of life and can occur intermittently or frequently. Some epilepsies
are limited to specific age groups; some patients suffer from epilepsy for a
limited time, and some for their whole lifetime. Epilepsies that develop
after an identifiable event or cause incident (e.g., asphyxia, head injury,
meningitis are referred to as symptomatic epilepsy or epilepsies developing
from unknown cause are referred as idiopathic epilepsy.
1.1.1.1 Prevalence of epilepsy
Epilepsy is a chronic condition with a high prevalence rate. The prevalence of
epilepsy is defined as the number of patient’s diagnosed with epilepsy at a
given point of time. When examining age-adjusted studies conducted in
developed and developing countries, the prevalence of epilepsy is more in
developing countries. The age adjusted prevalence was 41.0 per 1000 in

26
Nigeria (Olumide A.e · Oyediran A.B.O.d · Pearson C.A.d · Bolis C.L.f
1982)and 22.0 per 1000 in Ecuador (Cruz et al., 1985) much higher when
compared to the developed countries, prevalence in New York, United States
was 7.1 per 1000 (Haerer et al., 1986) and in prevalence in Europe ranged
from 3.7 to 3.3 in 1000 in studies conducted in Italy (Rocca et al., 2001,
Banerjee et al., 2009). Environmental and social differences play a
significant role in the development of epilepsy compared to racial
differences (Noronha et al., 2007). Generally, 5-10 people in each 1,000
around the world are identified with epilepsy. Two to five per cent of the
overall population has experienced epilepsy at least once in their lifetime. 40
to 50 million people are detected with epilepsy, and a significant section
(~30%) have treatment refractory epileptic situation (Wiebe, 2000).
1.1.1.2 Incidence of epilepsy
Incidence is defined as the rate at which new patients are diagnosed with a
disease within a given time in a given population. In epilepsy, the annual
incidence rate is generally calculated per 100,000 populations. The
estimated incidence rate of epilepsy is 40–70/ 100,000 in developed nations
and 100–190/100,000 in underdeveloped or developing nations; socially
and economically deprived individuals are at higher risk (Sander, 2003).
From surveys during the past decade, the epilepsy incidence rate is high in
young age groups (<15 years), and in older age groups (>70years)
(Olafsson et al., 2005, Banerjee et al., 2009)
1.1.1.3 Aetiology of epilepsy
Some significant changes were made to the terminology and classification of
epilepsy by ILAE after 2011. Novel techniques in neuroscience research have

27
led to development of rational system for classification of epilepsies based on
underlying mechanisms.
According to the ILAE the causative factors for epilepsy could be i) Genetic
reasons: Epilepsy is caused because of genetic defects e.g., genetic
generalised epilepsies and channelopathies ii) structural or metabolic
conditions: Epilepsy is the consequence of a structural or metabolic defect
such as brain lesions, malformations during the cortical development
(Barkovich et al., 2012) iii) unknown: there are many patients suffering with
epilepsy with unidentified aetiology (Berg and Scheffer, 2011) (Hauser,
1997).
1.1.1.4 Mortality
Mortality rate is generally elevated in patients with epilepsy. Mortality rate
in epilepsy is low in developing countries compared to underdeveloped or
developing countries. A recent study in China revealed that epileptic
patients had 3–4 times higher mortality rate than the normal population
(Ding et al., 2006). Epilepsy associated mortality rate is considerably
increased by 2-3 times in worlds population(Hitiris et al., 2007).
1.1.2 Classification of epilepsies
Many changes have been made to the classification of epilepsy and
terminology in recent times. Latest developments and advancements in
scientific techniques have improved the understanding of epilepsy. Currently
epilepsy is classified into three main types (Berg and Scheffer, 2011, Engel,
2011, Shorvon, 2011) (Commission on classification and terminology of the
International League against epilepsy, 2011) (Figure 1.1). The three types are
1) generalised epilepsy, where electrical activity is spread throughout the
brain (Figure 1.2) and, 2) focal epilepsy, where the electrical activity is

28
limited to particular areas of the brain, mostly frontal and temporal regions
(Figure 1.3). 3) Unknown, where seizures cannot be diagnosed either a
generalised or focal seizures are grouped as unknown such as epileptic
spasms or other seizures.
Figure 1.1 above is the current classification of epilepsy. Epilepsy is mainly
classified into two major types: Focal epilepsies and Generalised epilepsies,
these epilepsies are further classified to other syndrome types based on
pathophysiology of the syndrome (adopted from www.epilepsy.org.au).
1.1.2.1 Focal epilepsy
Focal (partial) epilepsies are mostly symptomatic in nature with an
identifiable cause. The focal point or focus may reside in frontal,
parietal, temporal, occipital lobes or in the motor cortex. These
seizures exhibit better responses to treatment compared to the
generalised symptomatic seizures.

29
Efficacious drugs in focal epilepsies are phenytoin and carbamazepine and
phenytoin. With the use of these anti-epileptic drugs the seizure become less
severe or less frequent. In the case of inadequate improvement with these
drugs, additional drugs such as valproate may be used. Maintaining a good
and healthy lifestyle is very important: sleep deprivation, emotional stress
and alcohol consumption may trigger these seizures. (Nashef et al., 2007).
The idiopathic focal epilepsies are rare and display good response (efficient
seizure reduction with drugs) to treatment (phenytoin or carbamazepine).
Figure 1.2 These are focal or partial seizures, in these seizure the abnormal electrical activity or discharges
start and confined to a localised region in the brain (adopted from www.epilepsy.org.au
accessed on 2/10/2013).
.
1.1.2.2 Generalized epilepsy
The majority of these epilepsies are idiopathic in nature. These epilepsies
usually show a good response to anti-epileptic drugs. The most common
generalised epilepsies are: absences (treatment: ethosuximide and
valproate), generalized tonic-clonic (treatment: carbamazepine, valproate or
phenobarbitone) and myoclonus (treatment: benzodiazepines and valproate).

30
Some of the generalized epilepsies are symptomatic. These may be caused
by asphyxia, neonatal brain damage or with inborn errors of metabolism.
Figure 1.3 These are generalised seizures where the seizure arises in both hemispheres of the brain
simultaneously (adopted from www.epilepsy.org.au accessed on 2/10/2013).
1.1.3: Prognosis of Epilepsy
Prognosis is used to predict the course or outcome of a medical condition or
disorder. For patients with epilepsy, prognosis means the probability of more
seizures after an unprovoked seizure or the chance of attaining seizure
freedom after recurrent seizures have been established (Sander, 1993). The
table below illustrates the relationship between the type of epilepsy,
medication available and the prognosis (Sander, 1993, Cockerell et al., 1997,
MacDonald, 2001).
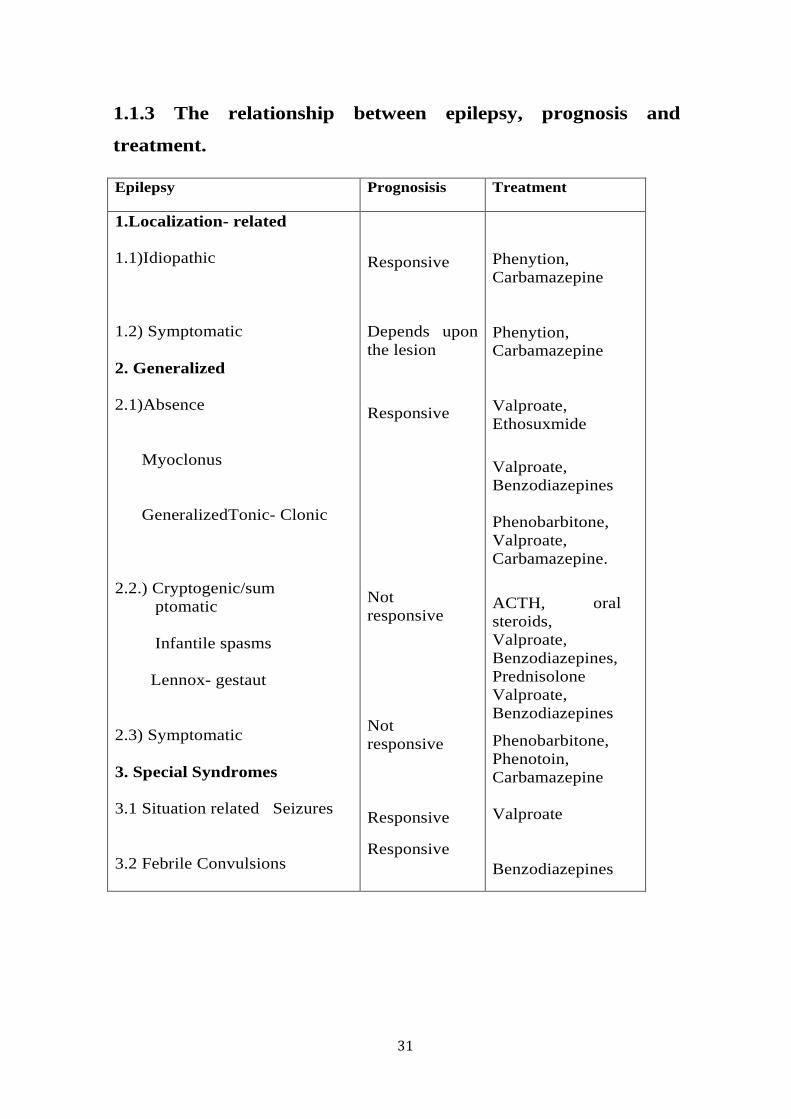
31
1.1.3 The relationship between epilepsy, prognosis and
treatment.
Epilepsy Prognosisis Treatment
1.Localization- related 1.1)Idiopathic 1.2) Symptomatic 2. Generalized 2.1)Absence Myoclonus GeneralizedTonic- Clonic 2.2.) Cryptogenic/sum ptomatic Infantile spasms Lennox- gestaut 2.3) Symptomatic 3. Special Syndromes 3.1 Situation related Seizures 3.2 Febrile Convulsions
Responsive
Depends upon the lesion
Responsive
Not responsive Not responsive Responsive
Responsive
Phenytion, Carbamazepine Phenytion, Carbamazepine Valproate, Ethosuxmide
Valproate, Benzodiazepines Phenobarbitone, Valproate, Carbamazepine.
ACTH, oral steroids, Valproate, Benzodiazepines, Prednisolone Valproate, Benzodiazepines
Phenobarbitone, Phenotoin, Carbamazepine Valproate Benzodiazepines

32
1.2 Absence seizures
Absence seizures are the most common type of generalized seizures
(Blumenfeld, 2003). These seizures were first designated by Poupart in
1705, and later were described by Tissot in 1770. Calmeil used the
term absence for the first time in (Temkin, 1994). Gibbs and his associates
described the association of signature 3-Hz spike and wave discharges
(SWDs) on electroencephalograms (EEG)(Gibbs FA, 1935). Some seizures
are recurrent and brief, limited to few seconds they are termed as
pyknoleptic seizures. Contrarily some seizures last for a few seconds to
minutes and could occur less frequently in a day, these are termed as non-
pyknoleptic absence seizures. Generally absence seizures onset at 2-13 year
of age peaking at 6 and females have higher incidence rate of absence than
males (Kramer et al., 1998).
1.2.1 Classification of absence seizures
The International League Against Epilepsy (ILAE) Commission has revised
the concepts, terminology, and approaches for classifying seizures and
epilepsy (1989).
According to ILAE, the classification of absence seizures is as follows:
• Absence seizures - Typical or atypical.
• Absence with special features - which includes myoclonic
absence and eyelid myoclonia
1.2.2 Aetiology of absence seizures
The underlying mechanism for generation and propagation of absence
seizures is complicated and not completely understood. In 1947, Jasper et al
electrically stimulated thalamic nucleus of cats at 3 Hz and triggered
bilaterally synchronous SWDs on EEG (Jasper HH, 1947). In 1953,

33
researchers were able to record SWDs from patient with absence seizures
by using depth electrodes in thalamus (Williams, 1953).
Gloor et al in 1977 revealed that 3Hz SWDs associated with absence
seizures were generated in the cortex in the feline penicillin model of
absence seizures. .
Pathological oscillatory rhythms during absence seizures are supposed to
initiate in thalamocortical circuit that involves gamma-aminobutyric acid
(GABA) mediated inhibition and glutamate mediated excitation.
1.2.3 Age of onset
Absence seizures appear in childhood and may continue to adulthood. The
cause for absence seizures may be genetic or a perinatal insult (Hamiwka
and Wirrell, 2009).
Five main syndromes are associated with absence epilepsy: Childhood
absence epilepsy, onset of this syndrome occurs at age 4-8 years, peaking at
the age of 6-7 years (P, 1985). Juvenile absence epilepsy, the onset of this
syndrome is around puberty and varies between pyknoleptic absences
which occur between 8.3 ± 4.5 years of age or non-pyknoleptic seizures
which occur between 14.8 ± 8.3 years (Wolf, 1979). Juvenile myoclonic
epilepsy, this syndrome has the age of onset from 8-26 years (Wolf, 1979,
Porter, 1993, Loiseau et al., 1995). As the absence seizure episodes are
brief, they are often unrecognised.
1.2.4 Morbidity and mortality
An absence seizure per se, does not directly result in deaths. However, if
the absence episodes occur while driving or operating dangerous machines,
patients may suffer from fatal injuries or death.

34
1.2.5 Electroencephalography
Electroencephalography is the only diagnostic test for absence seizures.
Generalized 3-Hz SWDs are presented during the seizures (Gibbs FA,
1935, Wolf, 1979, Lopes da Silva et al., 2003) (Figure 1.4). The onset and
termination of absence seizures are completely unexpected. During an
absence episode, mild clonic jerks, facial twitching and rhythmic eye blinks
may be seen and automatisms may be present during seizure progression
(Penry et al., 1975).
Figure 1.4 An image of human absence seizures displaying characteristic 3 Hz spike and wave
discharges. These absence seizures arise abruptly out of normal background and ceases after few
a seconds on EEG (adopted from (Lopes da Silva et al., 2003)).

35
1.2.6 Thalamocortical circuit and mechanisms of absence seizures
Many electrophysiological studies support SWDs are initiated in the
thalamocortical system that connects the thalamus and the cerebral cortex
(Danober et al., 1998, Panayitipoulos, 2005a, Benbadis SR, 2006).
1.2.6.1 Anatomy and Physiology thalamocortical circuit
The thalamocortical circuit consists of the cerebral cortex, VB thalamus,
nucleus of reticular thalamus (NRT) and their connecting neurons (Danober
et al., 1998) (Figure1.5). The thalamocortical circuit involves glutamatergic
(Fonnum et al., 1981b, Kaneko and Mizuno, 1988) axons projecting from
VB thalamus to layers III and IV of the cerebral cortex (Alloway et al.,
1993) and glutamatergic axon projections from layers V and VI of the
cerebral cortex to the relay nuclei of the thalamus (Bourassa et al., 1995).
The NRT consists of inhibitory GABAergic neurons that innervates the VB
and sends projections to its own neurons releasing GABA, a potent
inhibitory neurotransmitter forming a loop (Pinault et al., 1995, Cox et al.,
1996) (Figure 1.6). The NRT neurons receive projections from the
thalamocortical and corticothalamic circuit but do not project to the
cerebral cortex (Spreafico et al., 1991, Contreras et al., 1993, Bourassa et
al., 1995).
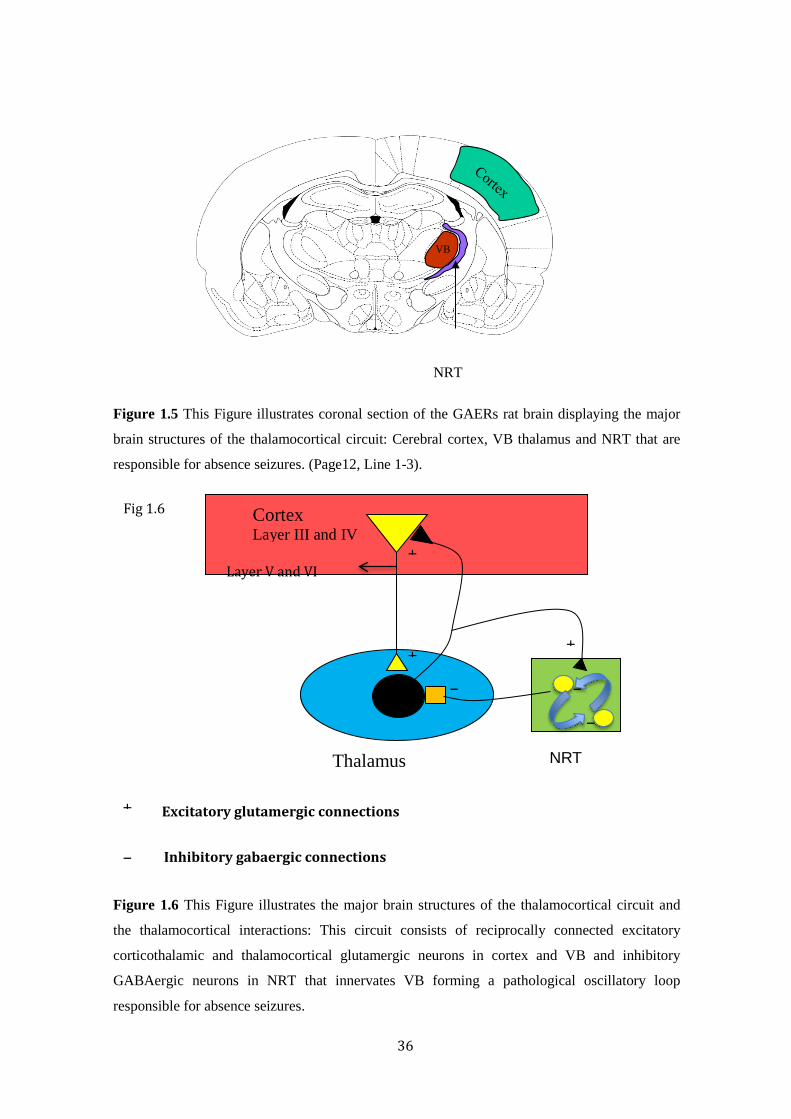
36
Figure 1.5 This Figure illustrates coronal section of the GAERs rat brain displaying the major
brain structures of the thalamocortical circuit: Cerebral cortex, VB thalamus and NRT that are
responsible for absence seizures. (Page12, Line 1-3).
Figure 1.6 This Figure illustrates the major brain structures of the thalamocortical circuit and
the thalamocortical interactions: This circuit consists of reciprocally connected excitatory
corticothalamic and thalamocortical glutamergic neurons in cortex and VB and inhibitory
GABAergic neurons in NRT that innervates VB forming a pathological oscillatory loop
responsible for absence seizures.
Thalamus
Cortex Layer III and IV
Layer V and VI
Excitatory glutamergic connections
Inhibitory gabaergic connections
NRT
+
+
−
+
−
−
+
−
VB
NRT
Fig 1.6

37
The view of generalized seizures is that, they arise simultaneously from
both hemispheres of the brain. Nevertheless, recent experiments in genetic
rat models of absence epilepsy, WAG/Rij and GAERS models have
proposed that a “cortical focus” within the cerebral cortex from which
seizures initiate and spreads over the cortex and subsequently to thalamic
regions. Meeren et al described that the focus was located in S1 region of
somatosensory cortex (Meeren et al., 2002). A recent study from our lab in
GAERS demonstrated that seizures were found to originate within the
somatosensory cortex (Zheng et al., 2012). The VB thalamus is the major
source of input to the cortex, transmitting sensory information. The cerebral
cortex receives its subcortical afferents from VB. The VB receives
inhibitory inputs from the NRT, which is located in the pathway that links
VB thalamus and cerebral cortex and receiving afferents from both
structures.
The thalamocortical and corticothalamic neurons are mostly glutamatergic
(Fonnum et al., 1981a). Most thalamocortical axons project to cortical
layers IV and II and information is transmitted to other cortical areas via
cortical interneurons (Jones, 1985). All sensory information, except smell,
is relayed to the cerebral cortex via the thalamus. In return, the pyramidal
neurons from cortical layers V and VI projects to VB thalamus (Jones,
1985, Bourassa and Deschenes, 1995). In addition, all VB thalamus
neurons receive GABAergic inhibitory projections from the NRT, which
is the major source of GABA in thalamus (Jones, 1985, Pinault et al., 1995,
Cox et al., 1996). NRT in return receives excitatory inputs from both
thalamocortical and corticothalamic axons.
Thalamocortical neurons exhibit two modes of firing patterns associated
with sleep/wake cycles: 1) relay mode or tonic mode, this occurs during

38
quite wakefulness and is associated with faithful transmission of sensory
information to the cerebral cortex where it is perceived and processed
(Llinas et al., 1998) (Figure 1.7). 2) Oscillatory mode or burst firing mode,
this occurs during sleep and is associated with filtering of sensory
information (Snead, 1995) (Figure 1.8).
During wakefulness the thalamus exhibits relay or tonic mode or firing with
fast sodium/potassium mediated action potentials (Figure 1.9). In relay
mode thalamic neurons are depolarized from resting membrane potential
levels positive to -55 mV. During sleep or drowsiness, the thalamus
switches its mode from relay to oscillatory or burst firing mode, where the
membrane potential is hyperpolarized to -60 mV. The de-inactivation of a
Ca2+ conductance causes an inward current through T-type Ca2+ channels
(Chemin et al., 2002) which may lead to high frequency burst of action
potentials (Steriade and Deschenes, 1984b) (Figure 1.9). This hyper-
polarization was believed to be mediated by the Gabaergic inhibitory
neurons in the NRT that provides rhythmic, synchronous inhibitory
postsynaptic potentials (IPSPs) to the VB thalamus. These oscillatory
bursts frequently occur during sleep, when the VB thalamus NRT neurons
are hyperpolarized. These conditions are suggested to be the functional
basis of sleep and drowsiness, where sensory information is not relayed to
the cortex, resulting in reduced responsiveness. These bursts are displayed
on EEG as spindles and delta waves during slow-wave sleep (Danober et
al., 1998). Spindles are frequent rhythmic oscillations of 7-14 Hz that occur
every 3-10 seconds slow-wave sleep (Steriade and Deschenes, 1984a).
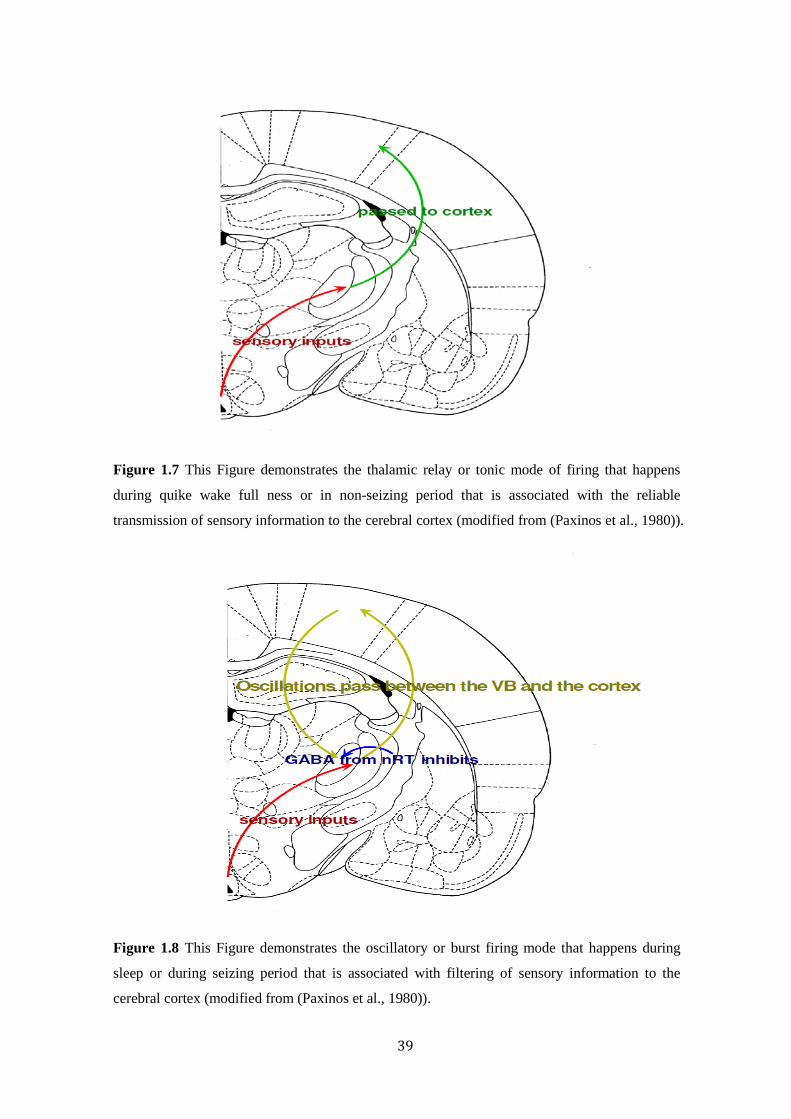
39
Figure 1.7 This Figure demonstrates the thalamic relay or tonic mode of firing that happens
during quike wake full ness or in non-seizing period that is associated with the reliable
transmission of sensory information to the cerebral cortex (modified from (Paxinos et al., 1980)).
Figure 1.8 This Figure demonstrates the oscillatory or burst firing mode that happens during
sleep or during seizing period that is associated with filtering of sensory information to the
cerebral cortex (modified from (Paxinos et al., 1980)).

40
Figure 1.9 This Figure demonstrates two modes of firing patterns, showing the transmission
between two modes of firing. Here in this picture thalamic neurons fires initially in a tonic mode
during interictal period, as the seizures start the firing mode switches to low frequency burst
firing mode during a seizure.
The above pathological change in thalamocortical mode of firing from relay
to oscillatory is the mechanism underlying the generation of absence seizures
(Snead, 1995, Kostopoulos, 2001). The initiation and triggering point of this
SWD’s is currently unknown but the possible explanation is that these SWD’s
may arise from disruption of delicate balance between inhibitory and
excitatory neurotransmitters acting on the Thalamocortical neurons (Slaght et
al., 2002). This disruption may result in the hyper-polarization and oscillatory
firing of thalamocortical neurons at inappropriate times.
1.2.7 Animal models of absence seizures
Absence epilepsy is a common neurological disorder that mainly occurs in
children and is not amendable to surgical intervention. It is ethically not
possible investigate the mechanism and pathophysiology of absence
seizures in humans. Therefore, many acquired and genetic models of
absence epilepsy have been developed to understand the cellular and
molecular mechanism underlying the generation and propagation of
Figure 1.9 5m
V
0.5 sec Tonic firing mode Oscillatory firing mode
0.5 sec
2mV
EEG
Neuronal firing activity

41
absence seizures. In some animal models seizure are induced by external
electrical stimulation and in some animal models, seizures are induced by
acute administration of a specific agent to an animal for example penicillin
epilepsy model (Fisher and Prince, 1977) low dose pentylenetetrazol (PTZ)
model (Marescaux et al., 1984) and the 4, 5, 6,7 tetrahydroxyisoxazolo
(4,5,c) pyridine 3-ol (THIP) model (Fariello and Golden, 1987). The
administration of these drugs induces SWDs accompanied by behavioral
arrest, twitching of vibrissae and facial myoclonus. Most of the these
models display similar pharmacological profile humans, seizures are
diminished on administration of ethosuximide and valproic acid and
worsened by carbamazepine (Marescaux et al., 1984). These
pharmacological models are play a crucial role examining the
neuropathological alterations related with absence epilepsy as well as in
screening the efficacy of antiepileptic drugs. The pharmacological models
exhibit similarity in seizure phenotype but they do not model the
underlying pathology of absence epilepsy to study the development of the
disease.
Unlike the chemical induced seizure models, genetic models, which exhibit
spontaneous recurrent seizures as witnessed in the human absence seizures.
Examples of commonly used mouse mutants include lethargic, stargazer
and totterer mice. However, most of these models are the result of single
gene mutation and are associated with other neurological abnormalities that
limit these models mostly to behavioral and physiological studies.
The spontaneous recurrence of SWDs in untreated rats lead to the
development and characterization of two genetic rat models of absence
epilepsy: 1) the GAERS, an inbred strain of Wistar rats developed in
France (Marescaux and Vergnes, 1995) the Wistar Albino Glaxo kept in
Rijswijk (WAG/Rij) (Coenen et al., 1992a). Both rat strains show similar

42
seizure characteristics, physiological and pharmacological profile to that of
the human condition. Studies using these models over the past 20 years
using multiple approaches have significantly improved the understanding of
underlying mechanism of absence seizures. In the current thesis we have
used the GAERS model for our investigations.
1.2.7.1 The Genetic Absence Epilepsy Rats from Strasbourg (GAERS)
GAERS is a genetic rat model of absence epilepsy originated in the Centre
of Neurochemistry in Strasbourg, France, where animals that displayed
SWDs were selectively inbred to produce a strain that exhibits absence type
seizure characterized by SWDs. A control strain, which does not exhibit
SWDs were inbred from the same colony and designated as non-epileptic
control rats (NEC).
Similar to human absence, SWDs in GAERS start and end abruptly on EEG
background accompanied by behavioral arrest, twitching of the vibrissae
and facial muscles. GAERS exhibit SWDs typically around 9Hz faster than
human absence seizures (approximately 3Hz) (Figure 1.10). However, this
difference in frequency is a species-dependent difference (Snead, 1995). In
GAERS SWDs last for 17 ± 10 seconds and occur 1.3 times per minute on
average. The SWD frequency of 9Hz in GAERS and can be interrupted by
strong and unexpected sensory stimulus. (Vergnes et al., 1991). GAERS
start to exhibit seizures in early adolescence (30-40 days post-natal) and are
fully expressed by the age of 3 months and persist for rest of their life.
GAERS exhibit similar pharmacological responsiveness to human absence
epilepsy.
As most characteristic features of absence epilepsy in GAERS are highly
similar to human absence epilepsy, GAERS are used as reference for
investigating the mechanisms underlying absence seizures.
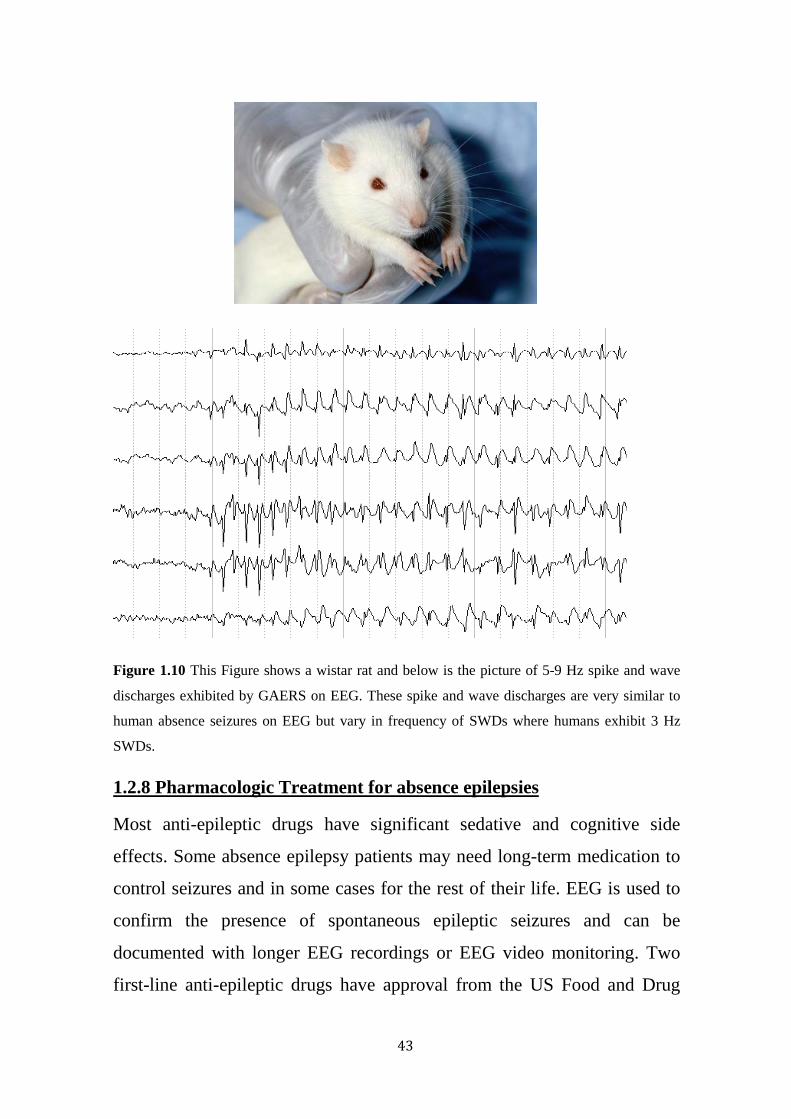
43
Figure 1.10 This Figure shows a wistar rat and below is the picture of 5-9 Hz spike and wave
discharges exhibited by GAERS on EEG. These spike and wave discharges are very similar to
human absence seizures on EEG but vary in frequency of SWDs where humans exhibit 3 Hz
SWDs.
1.2.8 Pharmacologic Treatment for absence epilepsies
Most anti-epileptic drugs have significant sedative and cognitive side
effects. Some absence epilepsy patients may need long-term medication to
control seizures and in some cases for the rest of their life. EEG is used to
confirm the presence of spontaneous epileptic seizures and can be
documented with longer EEG recordings or EEG video monitoring. Two
first-line anti-epileptic drugs have approval from the US Food and Drug

44
Administration (FDA) for treating patients with absence epilpesy: valproic
acid (Depakene, Depacon) and ethosuximide (Zarontin). Ethosuximide can
be used for absence seizures only but valproic acid has effectiveness for
absence seizures, generalized tonic-clonic seizures, and myoclonic seizures.
Some AEDs can aggravate seizures (Lerman, 1986), treatment with
carbamazepine (Snead and Hosey, 1985, Liu et al., 2006) and
oxcarbazepine (Vendrame et al., 2007) exacerbates the absence seizures
(Liu et al., 2006).
1.3 Introduction to NPY and its receptors:
Neuropeptide Y (NPY) is a naturally occurring 36 amino acid peptide, a
member of the pancreatic peptide family discovered from pig brain by
Tatemoto in 1982 (Tatemoto, 1982b). NPY is richly found in many brain
regions, such as the hypothalamus, amygdala, hippocampus, nucleus of the
solitary tract, locus ceruleus, nucleus accumbens and the cerebral cortex
(Allen et al., 1983, Chronwall et al., 1985, de Quidt and Emson, 1986).
NPY mediates many physiological responses such as feeding behaviour,
water consumption, learning and memory, locomotion, body temperature,
sexual behaviour, emotional behaviour, neuronal excitability,
cardiovascular homeostasis, hormone secretion, circadian rhythms
(Chronwall et al., 1985, Wahlestedt and Reis, 1993, Munglani et al., 1996,
Baraban et al., 1997, Hokfelt et al., 1998, Inui, 1999, Vezzani et al., 1999),
depression, anxiety (Kask et al., 2002, Berglund et al., 2003) and blood
pressure (Klapstein and Colmers, 1997).
NPY mediates its response by binding to G-protein coupled heptahelical
receptor subtypes, named Y1, Y2, Y4, Y5, and y6 (Stroud et al., 2005)

45
which inhibits adenylate cyclase and thus decrease intracellular calcium
levels (Michel et al., 1998, Berglund et al., 2003).
Receptor subtypes Y1, Y2 and Y5 are predominantly expressed in the
mammalian CNS. NPY has been implicated in anabolic activity through
circuits in the hypothalamus (Woods et al., 1998, Morris et al., 2007) and
seizure modulation in the thalamus(Sperk and Herzog, 1997, Bacci et al.,
2002, Xapelli et al., 2008)
1.3.1 Discovery of NPY
Many neuropeptides were discovered and identified basing on the biological
processes mediated by them. NPY was first identified by its C-terminal
tyrosine amide structure and is isolated from porcine brain extracts (Tatemoto
and Mutt, 1978, Tatemoto et al., 1982). This peptide is named as neuropeptide
Y as is contains numerous tyrosine residues (Y) in its structure (Tatemoto,
1982a, c).
1.3.2 Primary structure of NPY:
NPY is a 36 amino acid linear polypeptide. NPY shows a greater degree of
sequence homology between with PYY compared to PP (Figure 1.11). It was
then suggested that NPY, PYY and PP belong to an unrecognised peptide family
(Tatemoto, 1982a).
After that human NPY was isolated and the primary structure of NPY isolated
from human varied from the NPY isolated from porcine peptide only at one
location of the 36 residues (Corder et al., 1984) and NPY was subsequently
isolated from other animals such as birds, frogs, and others (Larhammar et al.,
1993). Dixon and co-workers first identified the cDNA encoding NPY.

46
Figure 1.11 This Figure illustrates the molecular structure of NPY and amino acid sequences of
porcine peptides PPY, NPY and pancreatic peptide PP. The blue structures with roman number I
to VII indicate 7 trans membrane type receptors and pink dot indicates the ion channels. The
asterisk at the end indicates the amidated C-terminus. Sequence homology between these
peptides is underlined (adopted from (Jasper HH, 1947)). (Page 22, Line 1-5)
1.3.3 Cloning of NPY specific receptor subtypes
Development in biotechnology techniques have led to the identification of
five NPY specific receptor subtypes, Y1, Y2, Y4, Y5, and Y6 (Michel et al.,
1998). These receptor subtypes exhibited only 30-50% sequence homologies to
each other. Moreover, each receptor subtype exhibits distinct tissue
localization and differential pharmacological profile.

47
1.3.3.1 NPY Y1 receptor subtype
The primary structure of the human NPY Y1 receptor was identified in 1992
(Herzog et al., 1992, Krause et al., 1992, Larhammar et al., 1992). It was the
first NPY specific receptor subtype to be cloned, it is a 384 amino acid protein. The
NPY Y1 receptor is widely distributed in mammalian CNS, heart, colon,
kidney, adrenal gland and placenta (Wharton et al., 1993).
1.3.3.2 NPY Y2 receptor subtype
The NPY Y2 receptor subtype is a 381 amino acid protein and exhibits
31% similarity with the Y1 receptor (Gerald et al., 1995, Rose et al., 1995,
Gehlert et al., 1996, Rimland JM, 1996). The NPY Y2 receptors are widely
expressed in human CNS, and are thought to mediate many physiological
responses including anti-epileptic actions. NPY Y2 receptor is widely
distributed in human CNS, heart, ileum and colon (Caberlotto et al., 1998b,
Ferrier et al., 2002, Jonsson-Rylander et al., 2003), the Y2 receptors are located
mostly on NPY-containing neurons (Caberlotto L, 2000).
1.3.3.3 NPY Y3 receptor subtype
There is no evidence for the existence of a NPY Y3 receptor (Michel et al.,
1998) and the reported Y3 receptor clone failed to confer NPY binding sites
(Rimland et al., 1991, Herzog et al., 1993, Jazin et al., 1993).
1.3.3.4 NPY Y4 receptor subtype
The NPY Y4 receptor is a 375 amino acid G-protein coupled receptor
(Lundell et al., 1995). The human NPY Y4 receptor exhibits 43% similarity
with the human NPY Y1 receptor (Bard et al., 1995, Lundell et al., 1995,
Yan et al., 1996). NPY Y4 receptors are localised in intestine, prostate gland,
and pancreas (Lundell et al., 1995), and are lightly expressed in the human

48
CNS (Parker and Herzog, 1999).
1.3.3.5 NPY Y5 receptor subtype
The NPY Y5 receptor is a 455 amino acid protein (Borowsky et al., 1998), this
receptor was originally cloned as a feeding receptor in hypothalamus. The Y5
receptor is broadly localised in the human CNS (Dumont et al., 1998) and is also
found in small intestine, colon, testis, spleen, and pancreas (Goumain et al.,
1998, Statnick et al., 1998).
1.3.3.6 NPY Y6 receptor subtype
The NPY Y6 receptor is a 290 amino acid protein present in chicken, cow, dog,
rabbit, mouse and human (Burkhoff et al., 1998). The NPY Y6 receptors are
functionally inactive in primates due to mutations occurred during evolution
(Matsumoto et al., 1996).
1.3.4 Distribution of NPY in central and peripheral nervous system
Bloom et al demonstrated that NPY is the most abundant neuropeptide and
is widely distributed in the brain (Adrian et al., 1983, Allen et al., 1983).
NPY is abundantly found in the paraventricular hypothalamic nucleus, hypo-
thalamic arcuate nucleus, paraventricular thalamic nucleus and suprachiasmatic
nucleus (Chronwall et al., 1985). Many in vitro studies have revealed the
coexistence of NPY with other neurotransmitters and neuropeptides
(Hokfelt et al., 1983). Selective receptor agonist studies revealed that NPY
Y1 and Y2 receptors are expressed independently in the human brain and
majority of the NPY receptors identified in the brain were Y2 receptors
(Aicher et al., 1991).
NPY like immune-reactivity in peripheral system was first identified in 1982
(Lundberg et al., 1982). NPY like immune-reactivity was identified in
sympathetic ganglia, heart atrium, spleen and blood vessels. NPY like

49
immune-reactivity was also found in gut and pancreas (Sundler et al., 1983).
NPY is mostly found in the sympathetic neurons (Mcdonald, 1988) thus seem
like an important peptide in sympathetic nervous system.
1.3.5 Biological effects of NPY
Several studies revealed that NPY mediates anti depressive, anti-stress,
anxiolytic and anti-nociceptive actions. NPY receptor subtype Y1 was
found to mediate anxiolytic-like action (Heilig et al., 1993, Wahlestedt and
Reis, 1993) and also found to mediate anxiogenic effect via the Y2 receptor
subtype (Nakajima et al., 1998). NPY transgenic mice exhibited anxiolytic
behaviours (Inui, 1999), additionally these transgenic rats with
hippocampal overexpression of NPY were unresponsive to restraint stress
and presented reduced spatial learning (Thorsell et al., 2000). NPY Y1
receptor-deficient mice were reported to develop hyperalgesia to pain, and
did not show any pharmacological analgesic effects of NPY (Naveilhan et
al., 1999). These reports suggest that NPY is involved in the mechanisms of
learning, stress, anxiety and nociception.
Low levels of NPY were found in patients who repeatedly attempted
suicide (Westrin et al., 1999). Alterations in the levels of NPY and NPY Y1
receptor mRNA were found in animal model of depression after treating
with anti-depressant drug (Caberlotto et al., 1998a) suggesting the role of
NPY in depression. These studies suggest the involvement of NPY in
depression.
1.3.6 Effect of NPY on Seizures
NPY is considered as an endogenous anti-convulsant. Studies have revealed
that NPY deficient mice showed less resistance to seizures induced by
GABA antagonist (Erickson et al., 1996). NPY deficient mice exhibited
uncontrollable limbic seizures induced by kainic acid which lead to 93 % of
mortality in mice, these deaths were prevented by ICV NPY infusion in

50
these mice (Baraban et al., 1997). In addition, transgenic rats with
overexpression of NPY displayed significant decrease in number of
seizures and length of seizures induced by kainic acid (Vezzani et al.,
2002). In in vitro models of epilepsy, infusion of NPY could successfully
inhibit epileptiform activity via Y2 receptor subtype (Klapstein and
Colmers, 1997) and Y5 receptor subtype potentially inhibited kainic acid
induced seizures (Woldbye et al., 1997). It was also shown that mice
lacking NPY Y5 receptor subtype were more susceptible to kainic acid
induced seizures (Marsh et al., 1999). In epileptic human hippocampus,
NPY released during profuse axonal sprouting and simultaneous up-
regulation of NPY Y2 receptors may instigate the inhibition of glutamate
release and consequently contribute to the seizure suppression mechanisms
of NPY (Furtinger et al., 2001).
1.3.7 NPY and focal epilepsy:
1.3.7.1 NPY and focal epilepsies: In-vivo
The anticonvulsive effects of NPY have been suggested in various animal
models of epilepsy. Changes in the immune-reactivity of NPY were
examined in rat brain after kainic acid induced limbic seizures. The NPY
levels consistently increased in the frontal cortex, hippocampus and
amygdala (Marksteiner et al., 1989). Increased NPY expression was
observed in neurons resistant to seizure-induced cell death (6-48
hours after i.p. Kainic acid). Using in situ hybridization histochemistry
NPY mRNA expression was assessed after kainic acid injection and
remarkably high hybridization signals were found in hippocampal after 8
months of kainic acid induced limbic seizures (Gruber et al., 1994). The
NPY Y1 receptor expression was assessed in hippocampus after kainic acid

51
induced seizures, the levels of NPY Y1 receptor decreased by 80% in
granule cells and concurrently increased by 75% in pyramidal neurons in
the CA2 region (Kofler et al., 1997). ICV administered NPY is a potent
inhibitor of kainic acid induced seizures, this study was the first to
determine the seizure suppression effect of NPY which is consistent with
the concept that NPY is an endogenous anticonvulsant (Woldbye et al.,
1997). Receptor autoradiography with the Y2 receptor ligand and in situ
hybridization of NPY Y2 receptor expression showed up-regulation of
presynaptic NPY Y2 receptors in schaffer collaterals after kainic acid-
induced recurrent seizures in the rat hippocampus. This up-regulation may
lead to presynaptic inhibition of glutamate release from hippocampus and
increased levels of NPY in mossy fibres may present endogenous seizure
suppression mechanism of NPY. (Schwarzer et al., 1998). A study on
expression of mRNAs for NPY and its subtypes in adult rats brains using in
situ hybridization in rapid kindling model demonstrated a cell and region-
specific mRNA expression for NPY and its receptor subtypes following
recurrent seizures (Kopp et al., 1999). By using auto-radiographic approach
it is shown that NPY Y2 and Y5 receptor binding receptors are reduced in
kindling and kainic acid models of epilepsy (Bregola et al., 2000). It has
been shown that seizure susceptibility in transgenic NPY knockout mice or
NPY deficit mice has a significantly high mortality rate than the control rats
after kainic acid injection (40 mg/kg i.p). In situ hybridization analyses in
these transgenic NPY knockout mice confirmed a decrease in prepro-NPY
gene expression (DePrato Primeaux et al., 2000). A constant increase of
NPY and elevation of hippocampal messenger RNA were found during a
spontaneous seizure in a Noda epileptic mutant rat (Jinde et al., 2002).
Overexpression of endogenous NPY in rat hippocampus is associated with
seizure inhibition and epileptogenesis in kindling and kainic acid models of

52
epilepsy (Vezzani et al., 2002). Results from all the above studies on
different models of epilepsies demonstrate the anti epileptic properties of
NPY but the underlying mechanism is still not completely understood and
this will require further investigations in the future.
1.3.7.2 NPY and focal epilepsies: In-vitro
Pharmacological studies of NPY demonstrated the critical role in regulation
of neuronal activity and neuro-modulatory effects of NPY. NPY inhibits
influx Ca2+ currents, this inhibition of presynaptic Ca2+ plays a critical role
in neurotransmitter release (Colmers and Bleakman, 1994, Wu and Saggau,
1997). The in vitro extracellular and whole cell patch-clamp recordings
from rat cortical slices demonstrated that NPY could significantly inhibit
epileptiform activity in in vitro models of epilepsy [0 Mg2+-, picrotoxin-, and
stimulus-train-induced bursting (STIB)] predominantly via NPY Y2 receptors
(Klapstein and Colmers, 1997). NPY inhibits synaptic excitation
of CA3 pyramidal cells of the rat hippocampus via presynaptic action of Y2
receptors (McQuiston and Colmers, 1996). A study showed that NPY
Y1 receptors in the hippocampus are involved in attenuating epileptic activity
in kainic acid model of limbic seizures(Gariboldi et al., 1998). On
examination of hippocampal function and responsiveness to kainic acid
induced seizures in Y5R-deficient (Y5R2y2) model, It was demonstrated that
Y5R2y2 were more prone to kainic acid seizures, suggesting significant role
of NPY Y5 receptor in mediating seizures (Marsh et al., 1999). Extracellular
or whole cell voltage-clamp recordings from CA3 regions in NPY Y5
receptor knock-out mice and matched wild type mice showed that, NPY and
NPY agonists caused a significant reduction in excitatory postsynaptic current
(EPSC) amplitudes concurrently suppressed-magnesium epileptiform burst
discharge in slices from wild type mice suggesting important role of NPY in
modulating excitatory synaptic transmission and inhibition of limbic seizure

53
activity in mouse hippocampus(Baraban, 2002). NPY Y2 receptors play a
prominent role in mediating NPY induced inhibition of glutamate release in
the hippocampus (Silva et al., 2001). It was shown that the antiepileptic
activity of NPY is mediated predominantly by the Y1 receptor subtype in the
frontal cortex and by Y2 and probably Y5 receptors in the hippocampal
CA3/CA1 areas, in rat cortical and hippocampal slices in Mg2+-free medium
(Bijak, 1999).
1.3.7.3 NPY and focal epilepsies: In humans
NPY is known to suppress hyper excitable activity in abnormal
pathological condition in rat brain. Intracellular recordings in hippocampal
slices from patients with hippocampal seizure onset showed NPY
containing cells throughout the hippocampus particularly in dentate
molecular layer suggesting role of NPY as a neuromodulator that may limit
hyper excitability in the human dentate gyrus (Patrylo et al., 1999). In
patients with Temporal lobe epilepsy (TLE), a significant increase in brain
derived neurotrophic factor (BDNF) was observed in the temporal
neocortex of patients. This increase was correlated significantly with levels
of NPY suggesting involvement of NPY in human epileptogenesis.
(Takahashi et al., 1999). Receptor binding and immune studies in
hippocampal specimens from patients with TLE showed increase in the
NPY mRNA, up regulation of Y2 receptors and down regulation of Y1
receptors (Furtinger et al., 2001). Patients with atypical febrile convulsions
(FC) showed lower NPY plasma levels compared to patients with typical
FC and controls suggesting patients with inadequate NPY inhibitory
activity are more susceptible to atypical FC (Lin et al., 2010).

54
1.3.8 NPY and generalised epilepsy:
Recent EEG studies with exogenous infusion of NPY in a genetic rat model
of absence epilepsy (GAERS) showed that NPY suppresses absence
seizures (Stroud et al., 2005). Another study showed NPY Y2 receptors
play a significant role in mediating seizure suppression effect compared to
Y1 and Y5 receptors in genetic rat models of absence epilepsy (Morris et
al., 2007). Focal injections studies of NPY showed that NPY suppresses
seizures when injected in cortex and showed a little but significant decrease
when injected in NRT but had contrasting effects when administered in
thalamus. Further, the NPY infusion in the S2 region of the somatosensory
cortex showed maximal effect of seizure suppression in GAERS (van Raay
et al., 2012).
1.4 Conclusion from literature review:
In summary, extensive literature suggests that, NPY is an endogenous
naturally occurring neuropeptide which activates three different receptor
subtypes NPY Y1, NPY Y2 and NPY Y5. NPY is widely distributed in the
human CNS and mediates many physiological responses. Our interest is it’s
anti-epileptic properties and ability to suppresses seizures in a various types
of epilepsy and the mechanism of seizure suppression. Numerous
experimental studies that have investigated this are summarized below.
In 1989, Marksteiner et al studied the changes in the immunoreactivity of
NPY in rat brain and found increased levels of NPY in cortex,
hippocampus and amygdala after seizures induced by kainic acid
(Marksteiner et al., 1989). Increased concentrations of NPY mRNA was
observed in different hippocampal regions after kainic acid induced limbic
seizures (Gruber et al., 1994). Infusion of NPY in in vitro models of

55
epilepsy, could successfully inhibit epileptiform activity (Klapstein and
Colmers, 1997). The first study to demonstrate an antiepileptic effect of
NPY was done by Woldbye et al, where NPY administered into the lateral
ventricle suppressed seizures induced by kainic acid (Woldbye et al., 1997).
An mRNA expression study in a rapid kindling model demonstrated a
neuronal and region specific mRNA expression for NPY after recurrent
seizures (Kopp et al., 1999). In 2001, it was shown that up regulation of
NPY receptors could be involved in the seizure suppression (Furtinger et
al., 2001). An NPY knockout study showed a significantly higher mortality
rate in NPY knockout mice than control mice, suggesting a critical role of
NPY in mediating neuronal excitability (DePrato Primeaux et al., 2000). A
consistent increase NPY expression was observed during a spontaneous
seizure in a Noda epileptic mutant rat (Jinde et al., 2002). Overexpression
of endogenous NPY in rat hippocampus is associated with seizure
inhibition and epileptogenesis in kindling and kainic acid models of
epilepsy (Vezzani et al., 2002). NPY was also shown to play an important
role modulating excitatory synaptic transmission and inhibition of limbic
seizure activity in mouse hippocampus (Baraban, 2002). Recent EEG
studies genetic rat model of absence epilepsy (GAERS) from our laboratory
showed that NPY suppresses absence seizures (Stroud et al., 2005, Morris
et al., 2007, van Raay et al., 2012).
The above studies strongly indicate that NPY plays a pivotal role in
suppression of seizures in various models of epilepsy. The current thesis
focuses more on investigating its mechanism of seizure suppression by
understanding the neuronal firing patterns in the thalamocortical circuit,
which is critical for development and propagation of absence seizures.

56
1.5 Rationale for this study
There is an increasing amount of in vivo and in vitro evidence on the seizure
suppression effects of NPY in various epilepsy models. However, the
underlying neuronal mechanisms for the seizure suppression effect are still
unknown. As discussed in section 1.2.6, the thalamocortical circuit plays a
pivotal role in generation and propagation of absence seizures. This circuit is
involved in the generation of signature spike and wave discharges and hyper-
synchronous rhythmic oscillatory activity in the brain during absence
seizures. Electrophysiological studies investigated the cellular firing patterns
of different neurons in the thalamic cortical circuit and their contribution in
mechanism of seizure initiation and spread of oscillatory thalamocortical
activity during a seizure. The mechanism underlying the generation of
absence seizures is the pathological and inappropriate change in the
thalamocortical mode from relay to oscillatory (Snead, 1995, Kostopoulos,
2001). No definitive cause for this change is currently known but it is highly
plausible that it could involve a lack of inhibition within the NRT causing
excessive release of GABA on thalamocortical neurons, resulting in a
disruption in the delicate balance between inhibitory and excitatory
neurotransmitters acting on the thalamocortical neurons (Slaght et al., 2002).
Recent studies in out laboratory have also demonstrated that ICV
administration of NPY clearly reduces the percentage-time spent in seizure
through reducing both seizure duration and number of seizures (Stroud et al.,
2005, Morris et al., 2007). Focal injections studies of NPY showed that NPY
suppresses seizures when injected in cortex and showed a small but
significant decrease when injected into the NRT(van Raay et al., 2012).
Nevertheless, the electrophysiological effects of NPY on neuronal firing
patterns and alteration in cellular patterns that underlie the mechanism of
seizure suppression have not yet been described.

57
In this thesis, we attempt to investigate the effects of NPY on neuronal
firing patterns to understand the mechanism underlying the seizure
suppression effect of NPY in vivo in genetic rat model of absence epilepsy
(GAERS). In this we have used in vivo electrophysiological approach to
investigate the alterations in neuronal firing patterns in different regions of
thalamocortical circuit, cortex, VB thalamus and NRT, which are basis for
absence seizures. Using GAERS as our rat model we had an advantage of
investigating the effects of NPY on spontaneously occurring seizures and as
it is an in vivo study there is an advantage of studying the neuronal firing
patterns during naturally occurring seizing and non-seizing periods and also
assess the alteration in the neuronal firing patterns induced by NPY that
may relate the seizure suppression. Thus, the data acquired in this study
will enable the investigation of mechanisms related to anti-epileptic effects
of NPY and also understand the underlying cellular mechanism of seizure
suppression.
1.6 Research questions
The present thesis focuses on investigating and understanding the
alterations in the neuronal firing properties of the thalamocortical circuit
neurons that support seizure suppression effect of NPY. The following
research questions will be investigated:-
1. What is the effect of ICV administration of NPY on spontaneously
occurring absence seizures in GAERS?
2. How does NPY affect the neuronal firing patterns in different
thalamocortical circuitry neurons when administered ICV?

58
3. What changes can occur to specific neuronal firing patterns that can
signify the alterations of neuronal activity at network level?
4. Does focally administered NPY show similar effects on neuronal
firing patterns on NRT neurons as it has when administered ICV?
5. Does NPY at amounts that suppress spontaneously occurring absence
seizures also show similar effects on electrically induced seizures in
GAERS?
1.7 Hypotheses
To address the above listed research questions the following hypotheses
will be tested:
1. ICV administration of NPY will induce anti-epileptic effects on
absence seizures in GAERS by decreasing the total and seizure
frequency on EEG.
2. To assess alternations in the neuronal firing patterns in the
thalamocortical circuit in vivo, electrophysiological recordings were
performed under neurolept anaesthesia, ICV NPY infusions were
also performed simultaneously along with EEG. NPY will induce
changes in the neuronal firing properties of different thalamocortical
circuitry neurons:
a. Cortical neurons (Layer IV and V)
b. VB thalamic neurons

59
c. NRT neurons
3. NPY administered ICV will induce changes in the specific neuronal
firing patterns that signify network activity:
a. Rhythmicity in the neuronal firing
b. Synchronicity of the neuronal firing
c. Correlation between field potentials of two different regions in
the thalamocortical circuit
4. NPY was injected focally at the recording site via micro-
iontophoretic process via multi-barrel electrode to investigate effect
of focal NPY administration on neuronal firing patterns. Focal
administration of NPY will induce similar effect on NRT neurons
further confirming the local effect of NPY on NRT.
5. Absence like seizures were induced in GAERS by external electrical
stimulation in somatosensory cortex and effect of ICV administration
of NPY is assessed on seizure inducing threshold of evoking
seizures. ICV administration of NPY will induce similar effects of
seizure suppression on induced seizures triggered by electrical
stimulation by increasing the seizure inducing threshold to evoke
seizures.
The understanding of the cellular and molecular mechanism related to
electrophysiological alterations induced by NPY infusion may contribute in
identifying the novel therapeutic target for seizure suppression in absence
seizures.

60
CHAPTER 2
MATERIALS AND METHODS
2.1 Study design:
All the experiments in this study involved in vivo electrophysiology. Three
different protocols of in vivo electrophysiology recordings have been
performed to understand NPY induced changes in neuronal firing patterns of
thalamocortical circuitry neurons in neurolept anaesthetised rats. The
protocols include single and paired juxta-cellular recordings with
simultaneous ICV drug administration and EEG recordings, juxta-cellular
recordings with simultaneous focal administration of NPY and cortical
stimulation to evoke spontaneous seizure activity combined with ICV drug
administration. The data collected from these experiments was analysed
before and after NPY administration and during ictal and interictal periods
and results are shown in the following chapters. Detailed methodology of
surgery, recordings and data analysis is explained in the current chapter.
2.2 Animals:
All experiments in this study were conducted on >12 weeks old male GAERS
and non-epileptic controls (NEC) rats. Rats were acquired from breeding
colonies at the animal house facility in department of Zoology, Royal
Melbourne Hospital, university of Melbourne and animal house facility in
Ludwig institute of Cancer research. All rats were bred and housed in
standard rat boxes and kept in a temperature-controlled animal room
maintained at 22-24°C with a 12hr light/dark cycle. All procedures performed
in this study were approved by the University of Melbourne Animal Ethics
Committee (0706287 & 1112079) and performed in accordance with the

61
guidelines published by the Australian National Health and Medical Research
Council (NHMRC).
2.3 Anaesthesia and surgery
2.3.1 Anaesthesia:
All experiments in this study were performed under two different kinds of
anaesthesia. Preparative surgeries for in vivo juxtacellular recordings were
performed under general anaesthesia using a combination of ketamine
(100mg/kg) and xylazine (10mg/kg) in 3:1 ratio at a dose of 0.1mg/100gm of
rat. Electrophysiological recordings were performed under “neurolept
anaesthesia” using a mixture of drugs d-tubocurarine (0.4 mg; Sigma
Aldrich), Fentanyl (0.375 μg; Mayne Pharma), haloperidol (50 μg; Janssen
Pharma) with glucose (25 mg; Sigma), which completely anaesthetises and
paralyses the animal but keeps it in a pseudo-awake state or
electrophysiogically active state which allows the generation of seizures
(Pinault et al., 2001).
Preparative surgeries included penile vein catheterization, tracheostomy, EEG
electrode implantations, ICV cannula implantation, craniotomy and duratomy.
2.3.2 Penile vein catheterization:
Dorsal penile vein catheterization is used to continuously infuse rat with
neurolept anaesthetic during the electrophysiological recordings. For this
procedure, the penis was retracted, and skin and connective tissue around the
penile vein was removed. The distal end of the penile vein was secured tightly
to stop the back flow of blood. A small incision made using a Vannas-scissors
and a polyethylene tube (0.8/.04mm) was inserted into the penile vein. The
position of the tubing was checked by with drawl of blood and the position of
the tubing was secured with a suture.

62
2.3.3 Tracheotomy:
Tracheotomy was performed to enable mechanical artificial ventilation to the
rats during the experiment. The artificial ventilation to the rat is facilitated
using small animal ventilator (SAR-830, CWE, USA). To perform
tracheostomy a small incision was made on the skin above the sternal notch
and trachea was isolated by separating the muscle layers. A small incision
was made between two rings of trachea and a 1 mm diameter plastic tube was
inserted and secured with suture. To prevent blockage of the airway, lung
secretions was aspirated with a tube connected to a 10ml syringe. The tracheal
tube was then attached to the ventilator for artificial ventilation of the rat (60
breaths/minute PH2O= 8-12cm). The body temperature of rat was maintained
at room temperature between 36.2-37.2°C using a thermo-regulated blanket
(Fine Science Tools., Inc, Germany) throughout the entire procedure. Needle
ECG electrodes were placed subcutaneously, bilaterally over lateral aspects of
thorax and signals from these electrodes were then conditioned and acquired
by using signal conditioner (Axon instruments, CyberAmp 380). The heart
rate of the animal was continuously monitored at (300-350 beats/minute) and
arterial PO2 and PCO2 were continuously monitored at all times by pulse-
oximeter and oxygenation was maintained >90%.
The experimental animal was then placed on a stereotaxic frame (David Kopf
Instruments, USA) and neurolept anaesthesia is initiated before the end of
general anaesthesia and was continuously administered at 0.5ml/hr using
infusion pump (KDS 220, Kd scientific Inc, Massachusetts, USA). The rate of
neurolept anaesthesia was regulated to maintain a predominantly
desynchronised EEG state during recordings. To record cortical EEG, three
electrodes were implanted on the rat skull.

63
2.3.4 EEG electrode and ICV cannula implantation:
For EEG electrodes and ICV cannula implantation, a midline incision was
made on the skull between the eyes and ears. The tissue across the scalp was
cleared and skull was exposed. Any bleeding was stopped using a diathermy
(Medtronic Solan, USA). Three holes were drilled on the skull for recording
electrodes without damaging the dura or brain (1.4 mm Easy Etch Engraver,
Arlec Australia, Blackburn North, Victoria, Australia); three extradural screw
electrodes were implanted in these burr holes to monitor EEG continuously.
One on the right hemisphere of the brain anterior to bregma for recording
cortical EEG, one hole on the left hemisphere of the brain anterior to lambda
which is used as a reference and the third burr hole posterior to lambda is
used as ground. The signal from these three electrodes was acquired and
displayed as one single channel. Another hole was drilled (-1.5mm relative to
bregma and +1mm relative to bregma) for implantation of ICV cannula
(Figure 2.1). ICV cannula (22G, 5mm cut guide cannula, Bioscientific PVT
LTD) was then lowered into the opening (dorsoventral -3.0-3.5mm relative to
dura). Intra-ventricular location of the cannula was confirmed by withdrawing
cerebrospinal fluid through the injecting line (22G injecting needle,
Bioscientific PVT LTD). The injecting line is removed from guide cannula
and a solid dummy line (28G, 5mm cut cannula internal, Bioscientific PVT
LTD) was then inserted into the ICV guide cannula to avoid blockage. The
electrodes and the ICV cannula was then secured in position using dental
acrylate.
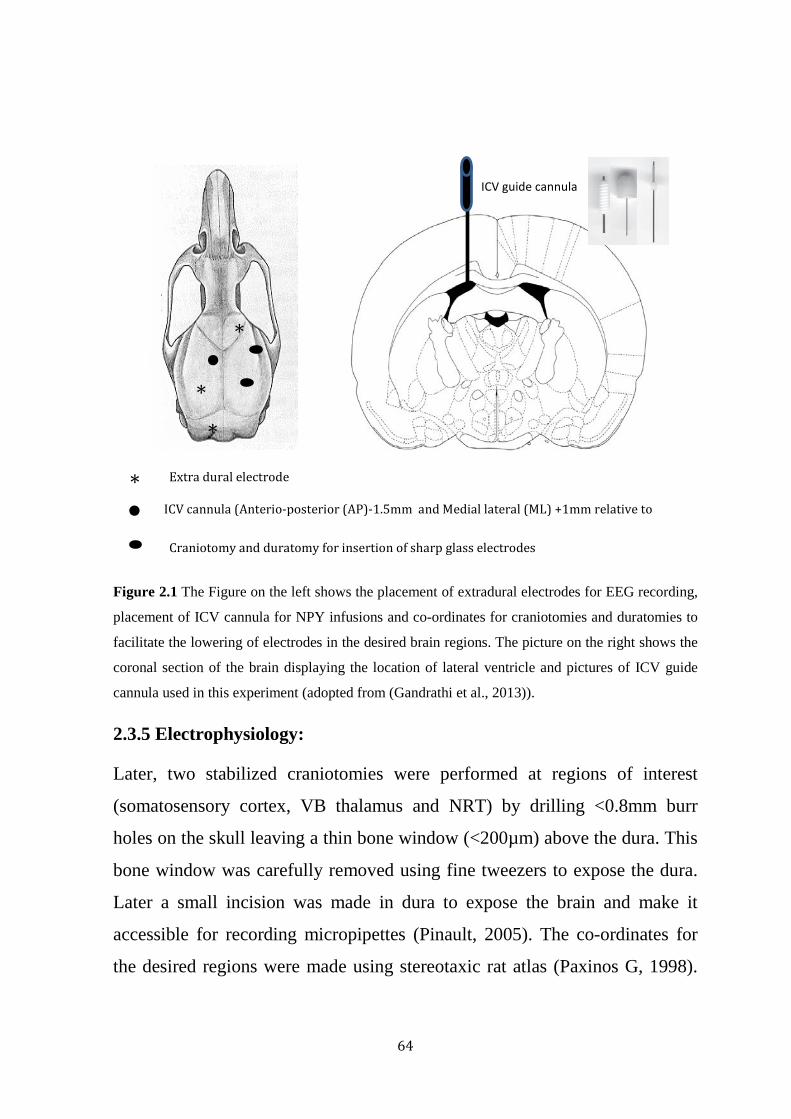
64
Figure 2.1 The Figure on the left shows the placement of extradural electrodes for EEG recording,
placement of ICV cannula for NPY infusions and co-ordinates for craniotomies and duratomies to
facilitate the lowering of electrodes in the desired brain regions. The picture on the right shows the
coronal section of the brain displaying the location of lateral ventricle and pictures of ICV guide
cannula used in this experiment (adopted from (Gandrathi et al., 2013)).
2.3.5 Electrophysiology:
Later, two stabilized craniotomies were performed at regions of interest
(somatosensory cortex, VB thalamus and NRT) by drilling <0.8mm burr
holes on the skull leaving a thin bone window (<200µm) above the dura. This
bone window was carefully removed using fine tweezers to expose the dura.
Later a small incision was made in dura to expose the brain and make it
accessible for recording micropipettes (Pinault, 2005). The co-ordinates for
the desired regions were made using stereotaxic rat atlas (Paxinos G, 1998).
*
*
*
*
ICV guide cannula
Extra dural electrode
ICV cannula (Anterio-posterior (AP)-1.5mm and Medial lateral (ML) +1mm relative to
Craniotomy and duratomy for insertion of sharp glass electrodes

65
Duratomies were covered with surgical sponge soaked in saline to avoid
desiccation.
Borosilicate glass capillaries of 1mm OD x 0.58mm ID (SDR technologies)
were pulled with pipette puller (P1000, Sutter instruments, USA) to a tip
diameter of ≈1 micron with resistance of 15-25 mega ohms typically recorded
in saline. These glass micropipettes were back filled with 1.5% N-(2
aminoethyl) biotinamide hydrochloride (Neurobiotin, Vector Laboratories,
USA) (50mg of neurobiotin in 3.3ml of 1M potassium acetate) and stored at
4oc one day prior to experiment. Glass electrodes were then lowered into the
brain through duratomies by using two motorized stereotaxic stepping
microdrivers, one for each electrode (Omnidrive by Neurostar, Germany).
These motorised microdrivers assisted slow movements of electrodes by 2µm
steps through the duratomies without causing significant damage to brain
tissue. After finding stable target cells from both electrodes, single neuronal
firing pattern were recorded (Figure 2.2) in the regions of interest
simultaneously with cortical EEG.
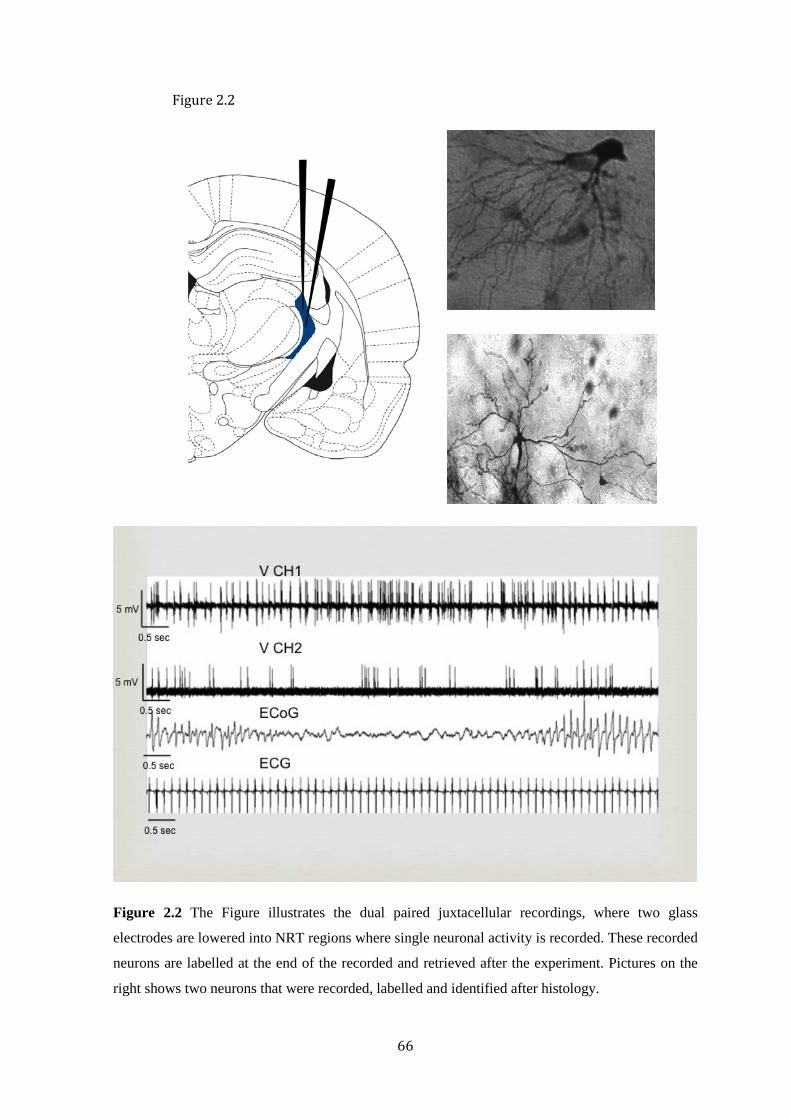
66
Figure 2.2 The Figure illustrates the dual paired juxtacellular recordings, where two glass
electrodes are lowered into NRT regions where single neuronal activity is recorded. These recorded
neurons are labelled at the end of the recorded and retrieved after the experiment. Pictures on the
right shows two neurons that were recorded, labelled and identified after histology.
Figure 2.2

67
2.3.6 ICV drug infusions
Before the start of electrophysiological recordings the dummy line was
replaced by an injecting line filled with 2µl of saline and 2µl drug separated
by air bubble. The injecting line was connected to a 10µl syringe enabling
manual administration of the drug ICV. Once stable electrophysiological
recordings were attained, baseline neuronal firing patterns were recorded for
15 minutes. 2µl saline was administered (as a control) manually followed by
another 15 minutes of recording. This was followed by administration of
1.5nmol NPY in 2µl of saline or saline as a control followed by another 30
minutes of recording (Figure 2.3). As used previously (Stroud et al., 2005,
Morris et al., 2007, van Raay et al., 2012) a 10µl glass Hamilton syringe is
connected to the injection needle using ling narrow bore line and a small air
bubble is introduced to enable tracking of the injection. The injecting line and
10 µl transparent glass micro-syringe is monitored thoroughly and a slow and
steady manual administration of saline and NPY is performed to ensure a
successful infusion. At the end of the recordings, the recorded cell was then
labelled with neurobiotin using micro iontophoresis process (Pinault, 1996).
At the end of the experiments, rats were then transcardially perfused with 4%
paraformaldehyde and the brains were removed for further histological
studies.

68
Figure 2.3 This is the timeline for the ICV NPY infusions that were performed during the
electrophysiological recordings under neurolept anaesthesia. After attaining stable recordings
conditions, baseline neuronal recordings were performed for 15 min followed by 15 minutes of
post saline and 30 minutes of post NPY recordings. The recorded neurons were than labelled
juxtacellularly and rats were perfused. Rat brain was extracted and stored for further histological
identification (adopted from (Gandrathi et al., 2013)).
.2.3.7 Juxtacellular labelling
At the end of recording, the recorded pairs of neurons were labelled using the
juxtacellular iontophoresis process described by Pinault to confirm their
location (Pinault, 1996). In this process, the recorded neurons were
juxtacellularly filled with 1.5 % neurobiotin iontophoretically by applying
positive currents of 0.5-8 nA through the micropipette for approximately 10
minutes (Figure 2.4). After labelling, the micropipettes were withdrawn
slowly, taking care not to damage the labelled cell. Following this, animals
were administered a lethal dose of phenobarbital (5mL/kg, i.p. injection;
Lethobarb, Virbac, Australia), perfused and brain was removed.
Electrophysiological recordings
Perfusion and Histology
Baseline Post-saline Post-NPY
Baseline recording
Injected Saline Injected NPY Juxtacellular labelling
15 min 15 min 30 min
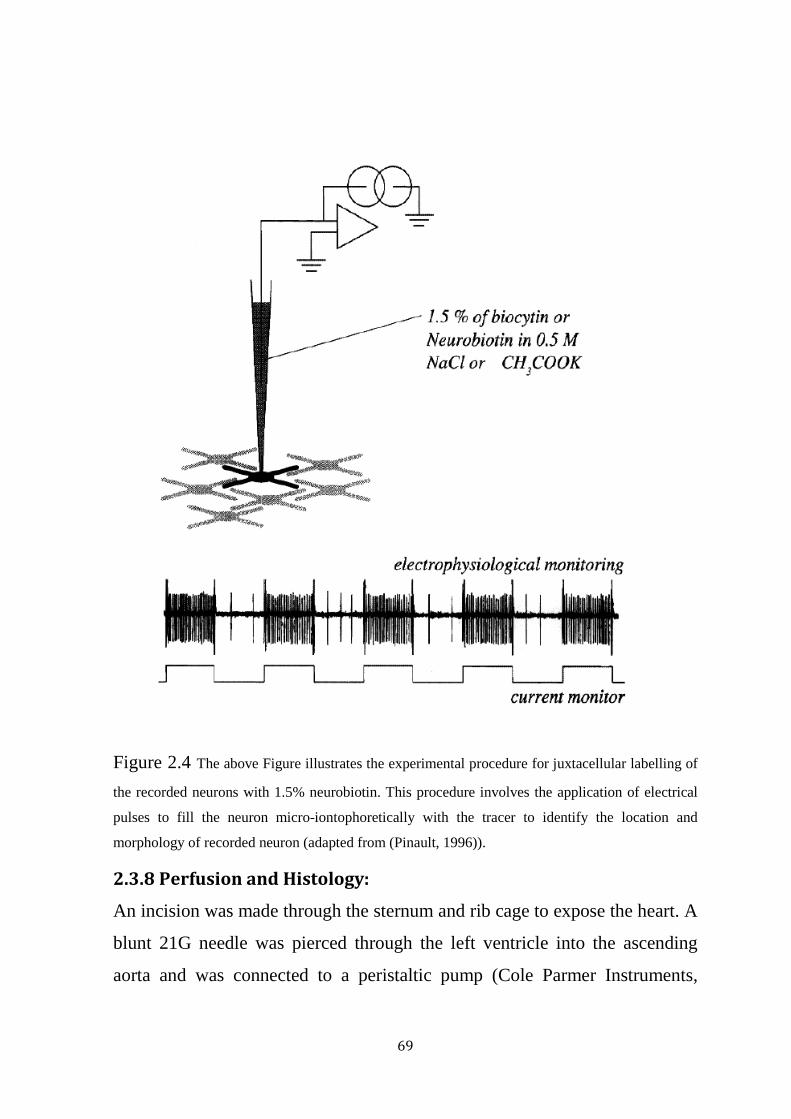
69
Figure 2.4 The above Figure illustrates the experimental procedure for juxtacellular labelling of
the recorded neurons with 1.5% neurobiotin. This procedure involves the application of electrical
pulses to fill the neuron micro-iontophoretically with the tracer to identify the location and
morphology of recorded neuron (adapted from (Pinault, 1996)).
2.3.8 Perfusion and Histology:
An incision was made through the sternum and rib cage to expose the heart. A
blunt 21G needle was pierced through the left ventricle into the ascending
aorta and was connected to a peristaltic pump (Cole Parmer Instruments,

70
Illinois, USA). Rats were then perfused transcardially with 300 ml 0.1 mM
phosphate buffered saline (PBS) to pump out the blood from the animal. This
is followed by 400 ml of 4 % paraformaldehydate (PFA) mixed with 0.25 %
glutaraldehyde. After perfusion, rats were decapitated and brains were
extracted and stored in 4% PFA at 4°C for histology.
For histological identification of the neurobiotin labelled neurons. The rat
brain was cut at the posterior end fixed to the vibratome with super glue.
Brain was cut into 100 μM axial sections using a vibrotome and placed in 24
well culturing plate in sequential order into each well. Brain slices were
washed thrice with 0.1M PBS (10 minutes between washes). Later, 400 μl of
1:100 avidin-biotin-peroxidase complex solution (Vectastain, ABC kit;
Vector Laboratories, USA) with 0.3% Triton was added to these slices and
Incubated overnight on a rotating stage. Next day, the brain slices were
washed and 400 μl of nickel intensified 3,3 P-diaminobenzidine
tetrahydrochloride activated by peroxidase (DAB; Vector Laboratories, USA)
DAB solution was added in each well to reveal the neurobiotin tracer. The
sections were than washed and mounted on gelatine-coated slides (6 sections
per slide) allowed to dry. Finally, sections were dehydrated using ethanol and
histolene and slides were cover slipped using DPX mounting media (BDH
chemicals, Australia). Later, the slides were observed under the microscope
for the localisation of any labelled cells.
Histological investigation in this thesis was only performed to identify and
confirm the location of the recording neuron. We have not performed any
investigations to morphologically identify and classify the subset of neurons
in a location, which would be more helpful but we rather confined our
investigation to assess and ensure the location of recorded neurons.

71
2.4 Focal injection studies:
For the focal injection study, a single craniotomy and duratomy was
performed at the right hemisphere of the brain (A/P: 3.6 mm, M/L: 3.6 mm).
A triple barrel borosilicate glass electrode was pulled to a diameter of ≈2
micron (15-30 MΩ). Only two barrels (out of three) were used in this
technique; one barrel was back filled with 1.5% neurobiotin and the other
barrel was filled with Saline/NPY for control and NPY investigations
respectively. The electrode was then slowly lowered to desired brain region to
locate a stable firing neuron. Once a stable neuronal recording is achieved, 15
mins of baseline recordings were performed followed by focal injection of
saline/NPY for control and NPY studies. NPY and saline were injected
focally using a microiontophoresis technique (Union-40 iontophoresis pump,
kation scientific). Post NPY/control recordings were performed for 15 mins.
In this technique neuroactive compounds are ejected by using small electrical
impulses (Curtis DR, 1964, D.R., 1964, Singh and Maibach, 1994). The
polarity and amount of current to be applied depends on the type of
chemicals. We have used 200nanoAmps of ejection current (+ve) for 10
minutes to deposit NPY (0.5nM) focally on the recording neuron and a
retaining current of 40 nanoAMPs was at all times of recordings (Ming and
Shao, 2000). Control (saline, 165mM NaCl) was ejected focally at
80nanoAMPs for 10 mins and retaining current of 30nanoAMPs was used at
all other times of recording (Haji and Takeda, 1993). All recordings were
performed simultaneously with EEG and recorded cells were again labelled
iontophoretically for later histological identification (Figure 2.5).

72
Figure 2.5 this is the experimental design for the focal injections of NPY. In this procedure
triple barrel glass capillaries were used, glass electrodes were lowered onto NRT region in
anaesthetized rats. One barrel filled with neurobiotin is used to record neuronal firing patterns,
another barrel filled with saline/NPY were used to inject microiontophoretically at the recording
site. Later the recorded neuron is labeled iontophoretically and the neurons are identified by
histology procedures.
Recording
Neuronal firing

73
2.5 Cortical stimulation studies:
For these experiments, single craniotomy and duratomy was performed (A/P
0.2mm, M/L 4.1). A borosilicate glass capillary was pulled to form a sharp tip
which was then broken gently with a paper to achieve a resistance of 2-5 mΩ.
This glass capillary was backfilled with 1.5% neurobiotin and positioned in
the somatosensory (S2) cortex with the stepping microdrive. The same
coordinates were used for all the experiments and seizures were induced by a
standard protocol to ensure minimal changes in the location of the electrode,
which could alter seizure-inducing threshold for these experiments. A
baseline recording was performed for 15 mins to assure stability. Baseline
seizure inducing threshold was assessed by applying an electrical stimulation
(7Hz) externally using and isolator stimulator. Current of 1-10milliAmps
were applied by increasing 1milliamp each step until the seizure was induced
and baseline seizure inducing threshold was recorded. 5 minutes after baseline
recordings saline was administered ICV and seizure inducing threshold was
assessed 15min, 1hour and 2 hours. Later NPY was administered ICV and the
same stimulation protocol was repeated and seizure inducing threshold
recorded. At the end of the experiment the whole region was labelled
iontophoretically for further histological identification (Figure 2.6). Location
of the electrodes and stimulation were confirmed by histological
investigations.

74
a) b)
c)
Figure 2.6 This is the experimental design for cortical stimulations: a) where ICV cannula is
implanted in lateral ventricle of left hemisphere, b) craniotomy and duratomy were performed in
the right hemisphere to facilitate electrode in to S2 region. Field potentials of the S2 region were
recorded along with cortical EEG. External electrical stimulation were given in S2 region to induce
absence like seizures, c) this illustrates the experimental timeline for the cortical stimulations
experiment. After attaining stable recording conditions baseline recordings were performed than
saline was injected. Seizures were triggered after 15 minutes of saline infusion and after 15
minutes, 1 hour and 2 hours of NPY infusion and seizure triggering threshold was recorded. The
Baseline
Inject Saline
15 min
Stimulation
15 min
Inject NPY
15 min
Stimulation Stimulation Stimulation
1 hour 2 hours Labeling
Experimental timeline
VB
NRT
Lateral ventricle
S2
7 Hz stimulation Seizure induced after stimulation

75
below picture is a snapshot of the recordings that displays the induction of seizure after stimulation
in GAERS.
2.6 Data acquisition and analysis
EEG signals were acquired from the rat brain, the recorded data was then
digitised at a sampling rate of 20 kHz (high-pass filtering) per channel
(Digidata 1440; Axon Instruments, USA) processed at band passes of 0.1-
600Hz for EEG (Clampex 10.0, Molecular Devices).
Neuronal signals recorded with glass electrodes were recorded with a
Neurodata dual channel recording amplifier (Cygnus technology Inc). The
data were processed with band passes of 0-6000 Hz (Cyberamp, Axon
Instruments, USA) for neuronal activity. The signals acquired were
conditioned using a programmable signal conditioner (Axon instruments,
CyberAmp 380).
Along with EEG and Neuronal recordings, field potentials were also recorded
for some experiments in this thesis. Unlike neuronal recordings and EEG,
field potentials is the activity of group of cells in a region. This is similar to
micro EEG and these are recorded by using blunt glass electrodes around 2-
5Mohm resistance. The field potentials are recorded and acquired similarly as
neuronal recordings using the same filters and programmable signal
conditioner.
To quantify single neuronal firing patterns, data was analysed using Clampfit
software (pClamp, Molecular Devices, California, USA) displaying multiple
channels of data, i.e. paired single cell recordings, 2 EEG traces and 1 ECG
trace. Spike events were semi-automatically detected using a threshold
detection protocol. A low-pass filtering of 20Hz was necessary and applied to
the single cell recording traces.

76
The recordings were arranged into standardized 15 minute time intervals –
baseline, following saline injection and following drug injection. These were
then subdivided into ictal, interictal and pre-ictal depending on the SWD’s
appearing on the recorded EEG. Periods with prominent SWDs on EEG are
defined as ictal episodes. Periods occurring between two ictal episodes or
periods without any SWDs are considered as interictal episodes. The pre-ictal
period is defined as the period just before an ictal episode, generally five
seconds before ictal period is considered as interictal period.
To assess neuronal firing properties, several firing parameters were derived
from single neural firing patterns. Different parameters were defined to
classify neural firing during ictal and interictal periods. Five parameters were
used for interictal firing properties: (1) Mean action potential (AP) firing
frequency (MAPF) –the average action potential firing frequency of the
neuron during the recorded period; (2) Percentage of APs in burst (PAPB) –
this gives us the proportional of action potential firing in bursts in a recording.
Bursts in the recording were identified as the group of action potentials with
interspike interval less than 6ms; (3) Mean number of APs per burst (MAPB)
– this parameter describes the average number of action potentials in burst in
a recording; (4) Maximum number of APs per burst (MxAPB) – describes the
maximum number of action potentials recorded in a burst; (5) Intraburst
frequency (IBF) – defined as frequency at which action potentials within the
burst. The same parameters were used to quantify the firing patterns of
thalamic and cortical neurons.
These interictal firing patterns are better described with an illustration of
interictal neuronal firing patterns in the below figure 2.6.1:
1) Mean action potential firing frequency (MAPF) for the whole stretch of
recording below is ≈ 11

77
2) Percentage of APs in burst (PAPB) ≈ 27%
3) Mean number of APs per burst (MAPB) ≈ 3
4) Maximum number of APs per burst (MxAPB) Maximum number of
APs per burst (MxAPB) ≈ 6
5) Intraburst frequency (IBF) is frequency measure within the burst
Ictal periods were parameterized by: (1) AP firing proportion per EEG spike
(APFP) – defined as the proportion of action potential firing per EEG spike;
(2) Percentage of burst with EEG spike (PB) describes the proportion of
action potential firing in burst within EEG spikes; (3) Mean number of AP’s
per EEG spike (MAP) gives the average number of action potentials firing per
EEG spike; and (4) Intraburst frequency gives the frequency of action
potentials firing in burst (IBF).
In the below exemplary figure (Fig 2.6.2) of an NRT neuron firing during
seizure, the ictal neuronal firing parameters are described as follows:
1) AP firing proportion per EEG spike (APFP) ≈ 1 as each EEG spike (in
SWDs) has an correlated action potential firing.
NRT cell
EEG
2 mV
1 sec
1 sec 1 mV
Fig 2.6.1 NRT neuron firing during non-seizing period

78
2) Percentage of burst with EEG spike (PB) is 100% as each EEG spike is
characterized by a burst of action potentials.
3) Mean number of APs per EEG spike (MAP) ≈ 7 i.e. an average number of
actions potentials per burst
4) Intraburst frequency is the frequency within the burst.
Four parameters were defined quantify drug effects on cortical EEG. These
were: (1) Total length of seizures (TLS) in the recording; (2) Mean duration
of seizure (MDS) – defined as the average duration of the seizure length; (3)
Cycle frequency (CF); and (4) Proportion of recording time of seizures (PTS).
Autocorrelation and cross-correlation analyses were performed to analyse the
rhythmicity of single neuronal firing, and synchronicity of firing between two
neurons, respectively. Rhythmicity is defined as the periodicity in neuronal
firing patterns of spontaneously firing neuronal activity. Lomb periodogram
of the autocorrelation function was used to access the periodicity of the
signals (Press et al., 1992). We defined synchronization as the presence of
simultaneous activity in the two signals. Wave form correlation was also
performed to assess the correlation between waveform signals (field
potentials) from different regions in the brain.
Fig 2.6.2 NRT neuron firing during seizure

79
2.6.1 Autocorrelation analysis:
The rhythmicity of the spontaneous neuronal (i.e significant periodicity
observed in the neuronal firing) was assessed using the method developed by
(Kaneoke and Vitek, 1996). In this method an autocorrelation function is
created by inverse Fourier transform of the power spectrum and significance
of the peaks in the autocorrelation function is determined by using the power
spectrum as described in (Press et al., 1992) for a frequency range from 0 to
50Hz (Figure 2.7). Using this method, multiple frequencies in the
spontaneous neuronal firing patterns can be assessed.
Juxtacellular neuronal firing data was divided into three different phases: ictal
period (seizing period), interictal period (non-seizing period) and pre-ictal
period (transition period from non-seizing period to seizing period) and
rhythmicity in neuronal firing was analysed before and after drug
administration.

80
Figure 2.7 Showing results from Kanoeke’s autocorrelation program which helps in detecting and
quantifying the rhythmicity in neuronal firing. These Figures here show an interspike interval
histogram, an autocorrelogram and spectrum of autocorrelogram which detects significant
rhythmicity in neuronal firing and also provides the frequency at which neuronal firing is
Rhythmic. The Figure below show an exemplary recording of a rhythmic and non-rhythmic
neuronal firing (adopted from (Gandrathi et al., 2013)).
2 m
V
2 m
V
0.1 sec
0.1 sec
Rhythmic firing
Non-rhythmic firing

81
2.6.2 Cross correlation analysis:
To assess the inter-dependence between two simultaneously firing neurons
cross correlation was used. Time stamps of the spontaneously firing neuronal
signals were collected and a cross-correlogram was constructed using a
custom program written in Matlab (1 milli second bin width and maximum of
500 lags). If two spike trains in the same recording are independent and
neuronal firing is randomly correlated with each other, the cross-correlogram
for that spike train will be flat. On the other hand, a correlated spike train and
a peak in the cross-correlogram would suggest that the neurons have a
common input or are synaptically connected.
We used the synchronization index (SI), developed by (Wiegner and
Wierzbicka, 1987) to quantify the height of the peak in the correlogram,
giving a measure of synchronicity between the two input spike trains. The SI
values range between 0.0 (not synchronous) to 1.0 (100% synchronous). To
access the statistical significance of SI, a critical value for the index is
calculated based on the notion of extra counts beyond ‘expected counts’,
which corrected for random fluctuations of the SI (Figure 2.8).
SI of ictal, interictal and pre-ictal phases were compared before and after and
ICV drug administration to examine the effect of the drug on synchronization
between two simultaneously firing neurons.

82
Figure 2.8 Cross correlation is the robust method to understand the synchronicity in the neuronal
firing and network behaviour. This Figure here shows cross correlograms constructed using
timestamps of the neuronal firing before and after ICV drug administration. Figures on the right
showing is a recording of typical NRT neurons firing recorded simultaneously with EEG during
ictal period (adopted from (Gandrathi et al., 2013)).
2.6.3 Waveform Correlation
Waveform correlation (spike 2, version 5.2.1) is used to understand the
correlation between waveforms recorded from two different regions in the
brain (Blumenfeld et al., 2007). The signals of interest were imported into
Spike 2 software (CED, UK) and processed to produce a cross correlogram.
The peak of the cross correlogram close to zero is calculated and the result of
waveform correlation is scaled from -1 (correlated but inverted), through 0
(uncorrelated), to +1 (full correlation).
Waveform correlations were assessed for cortical EEG and field potentials
recorded from NRT region during ictal, interictal and pre-ictal phases were
compared before and after and ICV drug administration to examine the effect
2 m
V
0 1
0 1
0 1
2 m
V
0 1
2 m
V
0 1
2 m
V
0 1
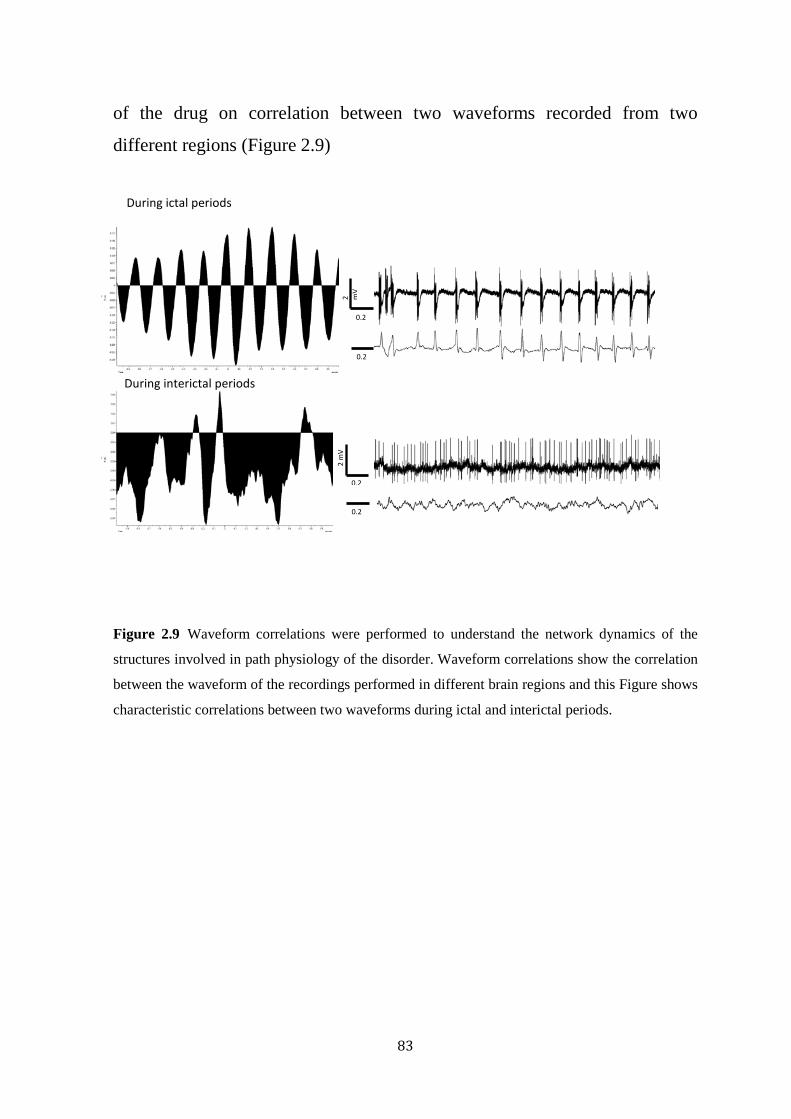
83
of the drug on correlation between two waveforms recorded from two
different regions (Figure 2.9)
Figure 2.9 Waveform correlations were performed to understand the network dynamics of the
structures involved in path physiology of the disorder. Waveform correlations show the correlation
between the waveform of the recordings performed in different brain regions and this Figure shows
characteristic correlations between two waveforms during ictal and interictal periods.
2 mV
0.2
2 m
V
0.2
0.2
0.2
During ictal periods
During interictal periods

84
CHAPTER 3
EFFECTS OF ICV ADMINISTERED NPY ON EEG AND
NEURONAL FIRING PATTERNS IN THALAMOCORTICAL
CIRCUIT IN GENETIC RAT MODEL OF ABSENCE
EPILEPSY.
3.1 Introduction
Absence (petit mal) seizures are a common seizure type in patients with
generalized epilepsy. Absence seizures are associated with symmetrical and
bilaterally synchronous 3Hz spike and wave discharges in the EEG (Gibbs
FA, 1935, Meeren et al., 2002). Absence episodes start and end abruptly on a
normal EEG background (Porter, 1993) with brief loss of consciousness
(Meeren et al., 2002). An absence seizure usually lasts 10-30 seconds and can
occur during quiet wakefulness with sudden arrest of ongoing behaviour.
Absence epilepsy generally occurs between the ages of 2-13, peaking at 6
years of age and females are twice more likely to have absence-like seizures
than males.
There are inadequate satisfactory therapeutic options available for patients
with absence epilepsy and many patients are unable to tolerate or are
unresponsive to currently available drugs: Ethosuximide, trimethadione,
valproate and benzodiazepines (Mattson et al., 1985, Micheletti et al., 1985,
Loiseau and Jallon, 1995, Liu et al., 2006). Valproate and ethosuximide cause
adverse effects in 40% and are ineffective in 20% of patients (Kwan and
Brodie, 2000, Murphy and Delanty, 2000) and benzodiazepines are highly
sedative. Drugs used effectively against focal epilepsies such as
carbamazepine are either ineffective or aggravating if used to treat absence
seizures(Liu et al., 2006). These facts show the necessity of developing new

85
agents for treating absence epilepsy. Neuropeptide associated mechanisms
may represent novel targets for seizure suppression, particularly neuropeptide
Y (NPY).
NPY is a 36 amino acid peptide discovered in 1982 in the pig brain by
Tatemoto (Tatemoto, 1982b). NPY is abundantly found in many brain regions
(Allen et al., 1983, Chronwall et al., 1985, de Quidt and Emson, 1986) and is
widely expressed in the human CNS (Morris, 1989).
NPY is a member of the pancreatic polypeptide (PP) family with six subunits
Y1, Y2, Y3, Y4, Y5, and y6. It acts by binding to six G-protein coupled
heptahelical receptor subtypes (Stroud et al., 2005), inhibiting adenylate
cyclase and thus decrease intracellular calcium levels (Michel et al., 1998,
Berglund et al., 2003). NPY plays a critical role in regulating the neuronal
excitability. In vivo studies in animal models of acquired focal epilepsy
revealed the evidence for involvement of NPY in seizure suppression
(Marksteiner et al., 1989, Gruber et al., 1994, Erickson et al., 1996, Kofler et
al., 1997, Woldbye et al., 1997, Kopp et al., 1999, DePrato Primeaux et al.,
2000, Vezzani et al., 2002) and the evidence for involvement of NPY is also
shown in in vitro studies (Greber et al., 1994b, Marsh et al., 1999, Baraban,
2002) and also in humans studies (Patrylo et al., 1999, Takahashi et al., 1999,
Furtinger et al., 2001).
The seizures in generalized epilepsies, especially in absence epilepsy, involve
particular neurophysiological circuitry that mediates hyper synchronous
oscillatory thalamocortical activity. The best documented example of this is
absence seizures. Recent studies have shown NPY reduces absence-like
seizures in genetic rat model of absence epilepsy (GAERS) (Stroud et al.,

86
2005, Morris et al., 2007) A recent study shows that NPY receptor subtype
Y2 plays a crucial role in mediating seizure suppression effect of NPY in
hippocampus (El Bahh et al., 2005). Additionally an intracerebro-vascular
injection study with NPY and subtypes (Y1, Y2 and Y5) on GAERS showed
that the Y2 subtype is more involved in anti-seizure activity compared to
other two subtypes compared to Y1 and Y5 (Morris et al., 2007).
Understanding the cellular and network dynamics in the brain during epileptic
activity and its changes with anti-epileptic drugs interference would help us
identify in greater detail the possible pathophysiology of these neurological
disorders. Many techniques have been developed in recent years to
understand the properties of cell, neuronal networks and changes occurring in
the brain. Juxtacellular recording (Pinault, 1996) is one such technique where,
sharp electrodes filled with neurobiotin were used to obtain extracellular
recordings and then labelling was performed by applying a train of current
pulses i.e. juxtacellular iontophoretic labelling. This technique permits high
resolution single cell electrophysiological recordings in anaesthetized animals
(Pinault, 1996, 2003, Pinault and O'Brien, 2005, Zheng et al., 2012). Pinault
has reported the properties and cellular mechanisms of thalamocortical cells
and reticular thalamic cells in GAERS (Pinault, 2003, Pinault and O'Brien,
2005). The staining technique assists in localisation and morphological
characterisation of recorded neurons (Pinault, 1996).
In the present study for this chapter we have used GAERS and NEC rats.
GAERS is a widely accepted genetic model for absence seizures, exhibiting
spontaneous and non-convulsive absence-like seizures. They display 5-9 Hz
SWDs on EEG and exhibit similar pathophysiological, behavioural,

87
pharmacological, aetiological characteristics to human absence seizures
(Danober et al., 1998).
From the previous studies we know that NPY suppresses seizures.
Understanding the underlying cellular and network changes involved in the
suppression of seizures will provide more knowledge on pathophysiology of
absence seizure and suggest mechanisms of action of NPY. In this study we
performed in vivo electrophysiological single and paired neuronal recordings
in thalamocortical circuit concurrently with ICV NPY infusions and assess
changes in the neuronal firing patterns that may contribute to the seizure
suppression effect of NPY.
3.2 Materials and Methods
The data showed in the present chapter involves EEG electrode implantation,
ICV cannula implantation and in vivo electrophysiology. Experiments were
conducted on 12-15 weeks old male GAERS (n=22) and NEC (n=6) rats.
Animals housing and all experimental procedures were performed in
accordance with the requirements of the Australian Code of Practice for the
Care and Use of Animals for Scientific Purposes.
3.2.1 Anaesthesia
Detailed descriptions of types of anaesthesia used in these experiments are
explained in methods (Chapter 2.3). Briefly, all surgical procedures were
performed under general anaesthesia (ketamine/xylazine in 3:1 ratio; 0.1ml
/100gm) and electrophysiological recordings were performed under neurolept
anaesthesia (Figure 3.1) (d-tubocurarine (2mg/1ml of water), Fentanyl
(0.2ml), Haloperidol (0.2ml) with glucose) which completely anaesthetises
and paralyses the animal it in a pseudo-awake state, allowing the rats to seize
spontaneously.

88
3.2.2 Surgery
The detailed description of surgical procedures involved in this study is given
in the methods (chapter 2.3). Briefly, the rats were anaesthetised with an
intraperitoneal injection of ketamine/xylazine (3:1). Once adequately
anaesthetised, penile vein catheterization and tracheotomy were performed
and rat was connected to mechanical ventilator for artificial ventilation (2.3.1,
2.3.2). The rats were then mounted on stereotaxic frame (David Kopf
instruments, Tujunga, CA, USA). Neurolept anaesthesia was initiated before
the end of general anaesthesia, and was administered continuously with micro
injection pump at 0.5ml/hr. An Active EEG electrode was epidurally
implanted on the remote cortex for cortical Electroencephalogram (EEG)
(2.3.3). For the ICV infusions, an ICV cannula was implanted on the left
hemisphere (A/P: 1mm, M/L: -1.5 mm and -3.0 to -3.5mm deep relative to
dura). Placement of cannula was confirmed by withdrawing cerebrospinal
fluid through the ICV injecting line (2.3.3) (Figure2.1).
Heart rate, body temperature and oxygen were continuously monitored and
maintained at physiological limits (PO2>90%, Heart rate 200-350
beats/minute, temperature-37OC) at all times during the experiment
3.2.3 Electrophysiology:
The surgeries and recording protocols have been described in methods
(chapter 2.4). Briefly, Two craniotomies and duratomies were performed on
the right hemisphere of the skull (A/P: 2.8 mm, M/L: 4.8 with -12.5 0o and
(A/P: 3.6 mm, M/L: 3.6 mm) for paired electrode recordings. Borosilicate
glass micropipettes filled with 1.5% were lowered into the desired brain
region using motorized stepping microdrive with 2µm steps and at relatively
low speed of 0.04mm/sec. Paired juxtacellular single neuronal recordings

89
were performed from target neuronal cells from thalamocortical regions
simultaneously with EEG and ECG (Figure 2.2).
3.2.4 ICV drug infusions:
The ICV drug infusion protocol is detailed in methods (chapter 2.5). Briefly,
juxtacellular single neuronal firing was recorded with the intervention of
saline and NPY at different time points. Baseline neuronal was performed for
the first 15 mins followed by ICV administration of 2µl saline. 15 minutes of
post-saline juxtacellular recordings were performed. This was followed by
ICV administration of 2µl of 1.5nmol NPY 30 minutes of post NPY recording
were performed (Figure 2.3). The recorded cell was then labelled and for
further histological investigations.
3.2.5 Juxtacellular labelling and Histology:
At the end of the ICV drug infusions, the recorded cells were
iontophoretically labelled for further histological investigations. The labelling
and histology procedures are described in methods (chapter 2.6, 2.7). Briefly,
the recorded cell was juxtacellularly filled with neurobiotin by applying
positive currents (0.5-8 nA). Rats were perfused and brain is removed. The
whole brain was sectioned into 100µm sections and stained with DAB
solution. The stained brain slices were mounted on glass slides, cover slipped
and left to dry overnight. The sections were then observed under bright field
inverted microscope (Olympus BX 51) under 20 x magnifications to discover
the labelled cells (Figure 2.2 and 2.4).

90
Figure 3.1 Figure illustrating the experimental time line for paired juxta cellular recordings.
Procedures under grey area are preparative surgeries performed under general anaesthesia (a
combination of Ketamine/xylazine), and electrophysiological recordings, ICV NPY infusions and
juxtacellular labelling performed under neurolept anaesthesia are shaded in black. At the end of the
experiment rats are perfused and brains are extracted for further histological identification.
3.3 Data acquisition and analysis:
The data acquisition and analysis is done using Clampex and Clampfit
software (pClamp, Molecular Devices). More information on data acquisition
and analysis is provided in the methods (chapter 2.10). Briefly, Data was
recorded at a sampling rate of 20,000 KHz/channel and processed at band
passes of 0.1-600Hz for EEG and 0-6000Hz for neuronal activity (Clampex
10.0, Molecular Devices). To quantify single neuronal firing patterns, data
was analysed using Clampfit software (pClamp, Molecular Devices,
California, USA).

91
The recordings were arranged into standardized 15 minute time intervals –
baseline, following saline injection and following drug injection. These were
then subdivided into ictal and interictal periods. To assess EEG and neuronal
firing patterns, several parameters have been determined. More detailed
information of these parameters is described in methods Chapter (2.10.1,
2.10.2, 2.10.3)
3.4 Statistical analysis:
Statistical analysis was performed using the Graph Pad Prism software
(GraphPad Software, San Diego California USA). One way ANOVA test was
performed to determine the significance levels of variables between the
treatments followed by planned analysis to compare significance between two
individual treatments. All data were expressed as mean ± standard error of
mean. P values less than 0.05 were considered to be significant.
3.5 Results:
3.5.1 Effect of NPY on seizures:
The percentage of recording time spent in seizures and total length of seizures
in the EEG recording were calculated before and after the ICV administration
of NPY. Highly significant decrease was found in Total length in seizures
(Baseline=131.41 ± 23.77 seconds, post NPY= 95.83 ± 20.48 seconds,
P=0.0001, n=11) and percentage of time spent in seizures (Baseline=15.91 ±
6.08, post NPY= 9.08 ± 4.12 seconds, P=0.0001, n=11) after ICV NPY
administration proving the anti-epileptic property of NPY (Figure3.2).
Other parameters were also assessed to determine NPY induced changes on
EEG such as mean length of seizures recorded and cycle frequency of
seizures. No significant change was observed in cycle frequency of seizures
(P=0.92, n=11) and Mean length of the seizures (P=0.10, n=11).

92
Figure 3.2 Cortical EEG of GAERS was analysed before and after administration of NPY. ICV
administration of NPY significantly suppresses seizures in GAERS. This above graphs illustrates
the seizure suppression effect of NPY by significantly decreasing the total length of seizures and
percentage of time spent in seizures and the below picture depicts the suppression of seizures after
n=11, p=0.0001
***
Tot
al le
ngth
of s
eizu
res i
n se
cond
s
% t
ime
in se
izur
es in
seco
nds
n=11, p=0.0001
After NPY administration:
1µV
1µ
V
0.8 sec
0.8 sec
Before NPY administration:

93
the administration of NPY. The below two graphs illustrates the results of the other two parameters
- EEG seizure duration and cycle frequency were not affected by ICV administration of NPY
fig 3.2.1
3.5.2 Effect of NPY on interictal Neuronal firing patterns
A significant increase was found in the mean firing frequency of the NRT
cells after administration of NPY (Baseline=12.32 ± 4.71, post NPY= 21.74
±8.09, P=0.008, n=12). A significant difference was observed in the
percentage difference of neuronal firing patterns between baseline and saline.
No significant change was observed in all other interictal parameters,
Percentage of burst firing (P=0.14, n=12), Mean number of action potentials
(P=0.46, n=12), Maximum number of action potentials (P=0.11, n=12),
interictal intra burst frequency (0.62, n=12) and ictal firing patterns. No
significant difference was observed between baseline and saline findings
contributing to the fact that this increase observed in the neuronal firing
frequency is not an artefact of ICV infusion procedure of saline injections but
an effect of NPY infusions.
None of the parameters of thalamic and cortical cells were found significant
during ictal and interictal periods. Though a trend was observed in some
parameters none of them were statistically significant (Figure 3.3).
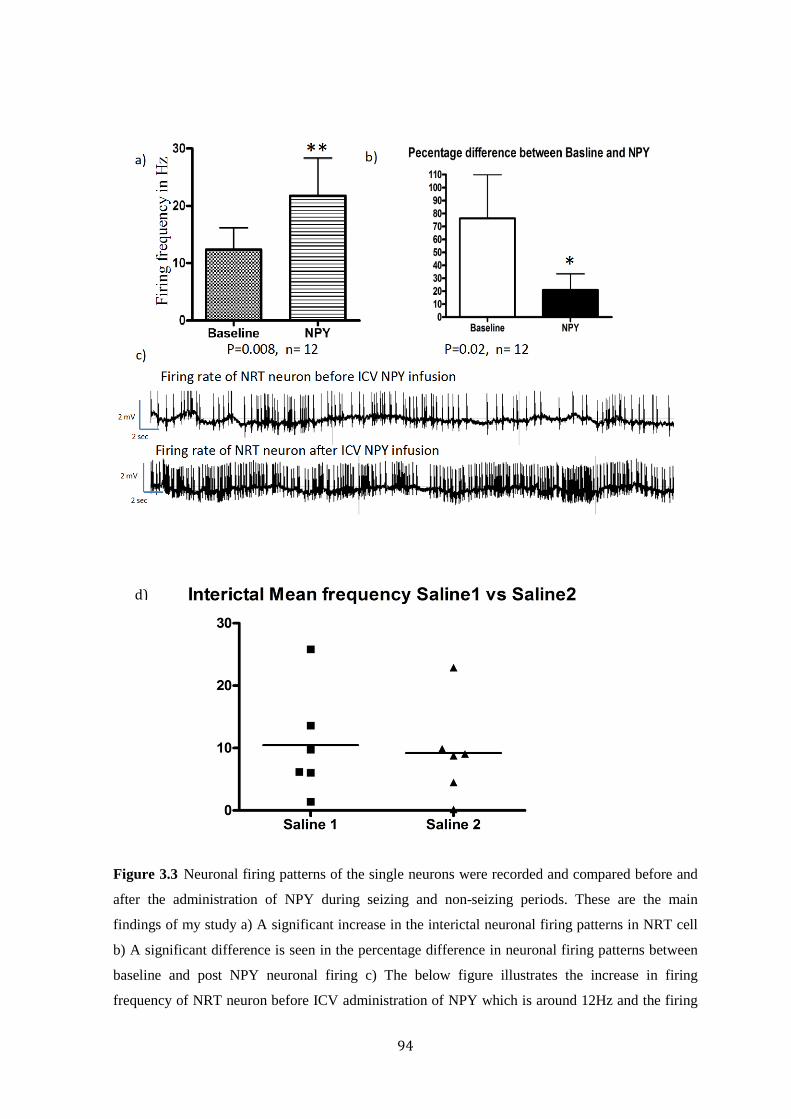
94
Figure 3.3 Neuronal firing patterns of the single neurons were recorded and compared before and
after the administration of NPY during seizing and non-seizing periods. These are the main
findings of my study a) A significant increase in the interictal neuronal firing patterns in NRT cell
b) A significant difference is seen in the percentage difference in neuronal firing patterns between
baseline and post NPY neuronal firing c) The below figure illustrates the increase in firing
frequency of NRT neuron before ICV administration of NPY which is around 12Hz and the firing
d)
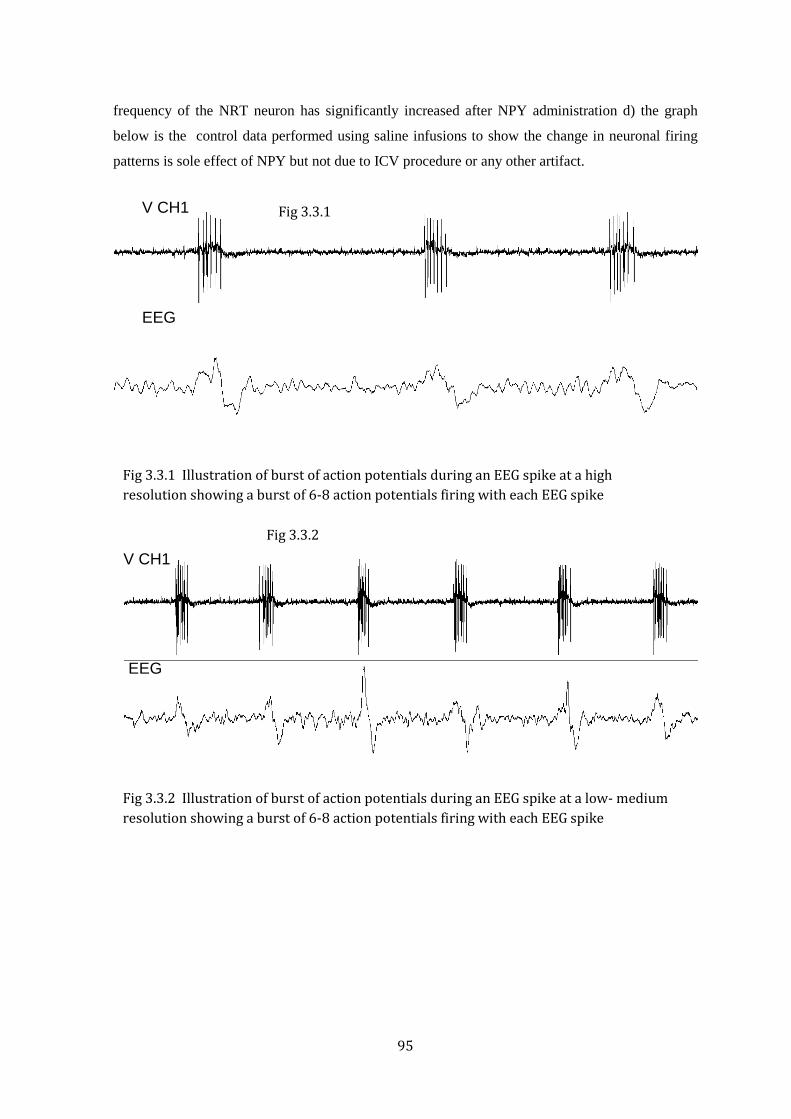
95
frequency of the NRT neuron has significantly increased after NPY administration d) the graph
below is the control data performed using saline infusions to show the change in neuronal firing
patterns is sole effect of NPY but not due to ICV procedure or any other artifact.
Fig 3.3.1 Illustration of burst of action potentials during an EEG spike at a high resolution showing a burst of 6-8 action potentials firing with each EEG spike
Fig 3.3.2 Illustration of burst of action potentials during an EEG spike at a low- medium resolution showing a burst of 6-8 action potentials firing with each EEG spike
V CH1
EEG
V CH1
EEG
Fig 3.3.1
Fig 3.3.2
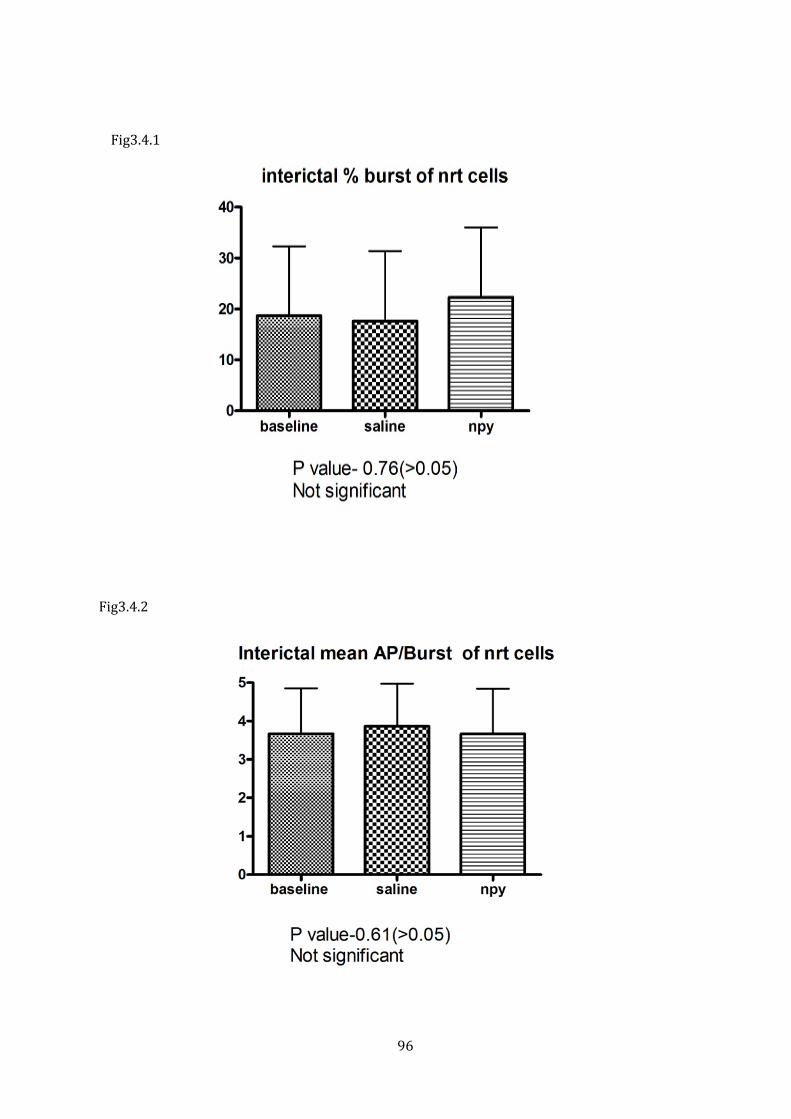
96
Fig3.4.2
Fig3.4.1
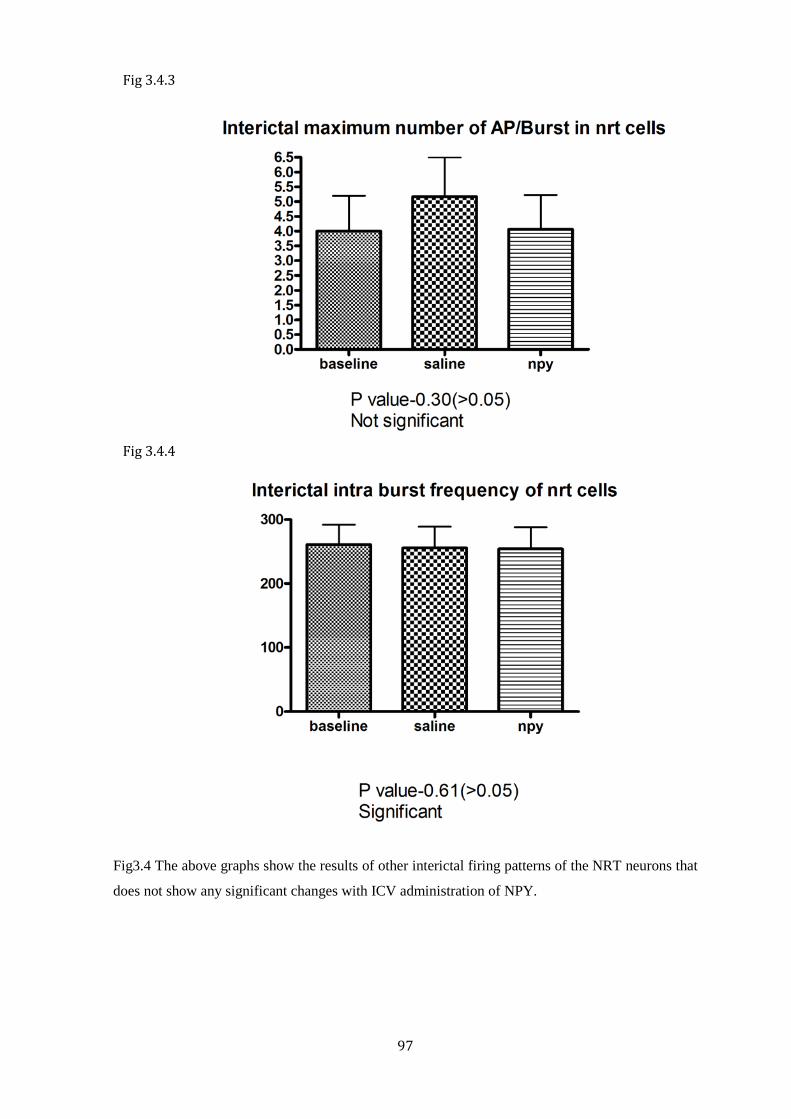
97
Fig3.4 The above graphs show the results of other interictal firing patterns of the NRT neurons that
does not show any significant changes with ICV administration of NPY.
Fig 3.4.3
Fig 3.4.4

98
P value – 0.05 (not significant)
P value – 0.05 (not significant)
Fig 3.5.1
Fig 3.5.2

99
Fig3.5 The above graphs show the results of other ictal firing patterns of the NRT neurons that does
not show any significant changes with ICV administration of NPY.
P value – 0.07 (not significant)
P value – 0.076 (not significant)
Fig 3.5.4
Fig 3.5.3

100
NRT interictal neuronal firing
patterns
ICV administered NPY induced
changes
MAPF Increased significantly
% Burst No significant difference
MAPB No significant difference
MxAPB No significant difference
IBF No significant difference
NRT ictal neuronal firing patterns ICV administered NPY induced
changes
APFP No significant difference
PB No significant difference
MAP No significant difference
IBF No significant difference

101
3.5.3 Effect of NPY on the Rhythmicity and synchronization in neuronal
firing of NRT cells
Rhythmicity of the neuronal firing was quantified applying spectral analysis
to the autocorrelogram, termed the Lomb periodogram. This method will
detect the multiple frequencies and also signifies the frequency at which
neuronal firing is rhythmic. In this study, the proportion of cells firing
significantly in rhythmic mode were identified and number of cells firing
rhythmically were compared before and after NPY administration in three
different phases (Ictal, pre-ictal, interictal). We have not assessed the changes
in the frequency before and after NPY infusions. No significant change was
observed in proportion of rhythmic cells before and after the ICV
administration of NPY during ictal, pre-ictal and interictal periods (Figure
3.6).
Figure 3.6 Data was recorded from 12 NRT cells and analysed. Kaneoke’s autocorrelation
program was used to detect rhythmicity in the spike train. a) This program uses Interspike intervals
of the spike train to plot a autocorrelogram and Lomb periodogram. This method enables to detect

102
the presence of multiple frequencies in the spike train and assess significances of frequencies
detected. b) Out of 12 nrt cells, proportion of rhythmically firing neurons showed a decreasing
trend after NPY infusion but results were not significant. c) Below is the illustration of neuronal
recording showing transition from non-rhythmic tonic firing in interictal period to rhythmic
neuronal firing.
To access the synchrony in the neuronal firing cross correlation analysis was
performed using the timestamps of spike events and synchronization index.
Time stamps of the spike events were used to plot cross correlograms and
synchronization index was calculated for each cross correlogram. To signify
the synchronicity in neuronal firing a critical index is also calculated using
peak values and off peak values in the cross correlogram. All the bin values in
the peaks present in correlogram are considered as peak values and values on
non-peak areas/troughs are classified as off peak values. Wiegner et al, has
given some clear calculations and formulas to determine these values and
calculate critical index (Wiegner and Wierzbicka, 1987). SI of the cross
correlogram for the neuronal recordings were compared before and after NPY
administration during interictal, pre-ictal and ictal phases of a seizure. No
significant change was observed in synchrony of the neuronal firing in NRT
cells before and after NPY in three different Phases (Figure 3.7).

103
Figure 3.7 As an approximation of the network behavior, we investigated the changes in the
synchronicity of neuronal firing. 8 pairs of NRT neurons were recorded and analysed, time stamps
of the recordings were used to plot cross correlograms and synchronization index was calculated
for each cross correlogram to signify the synchronicity in neuronal firing. SI showed a decreasing
trend during ictal period after NPY administration but was not significant. Here are exemplary
diagram of paired neuronal recording along with cortical EEG during seizure, here you can see two
neurons firing synchronously with each other along with the EEG spike.
3.5.4 Effect of ICV administered NPY on the waveform correlation
between cortical EEG and local field potentials of NRT
During a seizure cortex, thalamus and NRT are engaged in a highly rhythmic
oscillatory mode. To assess the effect NPY on correlation between networks,
we have performed cross correlation between local field potentials of two
different regions in thalamocortical circuit. As local field potentials signifies
the summation of neuronal activity of several cells in particular region,
performing correlation of field potentials will determine alterations in
correlation between networks. Waveform correlation analysis in Spike 2
Fig 3.7

104
software (Cambridge electronic design, Cambridge, England) measures the
similarity in waveforms with same sample interval. Cross correlation between
local field potentials of cortex and NRT was conducted and cross
correlograms were constructed. Observation of a peak near time displacement
zero suggested a correlated and synchronous signal (Blumenfeld et al., 2007).
A significant decrease in the correlation of field potentials was found between
cortex and NRT with ICV NPY drug administration (n=14, p=0.01) (Figure
3.8).
Fig.3.8
Figure 3.8 Waveform correlations were performed to understand the network dynamics of the
structures involved in path physiology of the disorder. Waveform correlations show the correlation
between the waveform of the recordings performed in different brain regions. Waveform
correlation between cortical EEG and field potential of NRT region decreased significantly after
NPY infusion. The Figure on the top right shows the wave form correlation between two regions

105
during ictal period. Below here is a visual example of correlation in field potentials of NRT neuron
and cortical EEG during ictal period.
3.6 Discussion:
Pathological oscillations within reciprocal connections of thalamocortical
circuits involving VB, NRT and somatosensory cortex, are responsible for
generation and initiation of absence seizures both in humans and rats. The
NRT is a critical structure in the thalamocortical circuit. The rhythmic hyper
synchronous oscillatory thalamocortical activity that characterizes absence
seizures is synchronized by the NRT (Steriade et al., 1986, Buzsaki et al.,
1990, Steriade et al., 1993, Contreras and Steriade, 1995, Snead et al., 1999,
Noebels et al., 2010). NRT is mainly composed of GABAergic neurons and
provides recurrent inhibitory synapses to the thalamic relay neurons (Kim et
al., 1997, Pinault et al., 2001, Pinault, 2003) and receives glutaminergic axon
collaterals from both thalamocortical and corticothalamic neurons (Ohara and
Lieberman, 1985, Pinault and Deschenes, 1998).
GABA is a major inhibitory neurotransmitter in vertebrate central nervous
system, blockade of GABA results in uncontrolled neuronal excitability in
neuronal circuits. GABA exhibits inhibitory actions via iontophoric GABA A
receptors, activation of these receptors triggers opening of a chloride ion-
selective channel and selectively conducts Cl¯ resulting increased chloride
conductance causing hyperpolarization in the neuron (Meldrum, 1989,
Treiman, 2001). Nevertheless, there are several reports of excitatory GABAA
receptors. The excitatory nature of GABAA is a result of increased intracellular
concentration of Cl¯ ions either during development of the nervous system or
in certain cell populations (Cherubini et al., 1991, Ben-Ari et al., 2007, Li and
Xu, 2008). The NRT neurons regulate the rhythmic oscillatory

106
thalamocortical activity via GABA-mediated hyperpolarization of VB
thalamus neurons, thereby de-inactivating low threshold T –type calcium
(Ca2+) channels, which results in high frequency action potential
bursts(Snead, 1995, Snead et al., 1999, Zhang and Jones, 2004) .
In the current chapter, we showed that ICV administration of NPY suppressed
absence seizures in a genetic rat model of absence epilepsy by exhibiting
reduction in total length of seizures and percentage of time spent in seizures
under neurolept analgesia. In addition, we also demonstrated that NPY
mediated reduction in the seizure levels (absence) i.e decrease in total length
of seizures and percentage of time spent in seizures were associated with
increased firing rate of the NRT neurons interictally in these rats. A study on
rat neocortex tissue showed NPY reduces evoked IPSP amplitude in
GABAergic interneurons which might lead to an increased firing rate.
Additionally, they also found an increase in IPSP amplitude in pyramidal
cells, this NPY mediated potentiation of inhibition might be derived from
increased excitability of interneurons by NPY (Bacci et al., 2002). Results of
the current studies indicate that the effects of NPY on neuronal firing patterns
were predominantly in NRT region in the thalamocortical circuit, as other
structures of the thalamocortical circuit did not have any significant change in
neuronal firing patterns, further confirming the critical role of NRT in
suppressing seizures. Previous studies have looked at the firing patterns of
NRT neurons in vivo in GAERS but this is the first study to look at the
changes in the neuronal firing patterns of the NRT neurons with application of
NPY. The neuronal recordings were only performed and assessed in
thalamocortical circuit, which is critical for generation of absence seizures.
Understanding the changes in this centrally based circuit responsible for
SWDs is more important to understand the underlying mechanism of absence

107
seizures. Given the complexity of the circuit and limitations of the method, it
is difficult to formulate a strong mechanistic hypothesis at this stage. To
briefly explore some possible mechanisms basing on the results of this
chapter: Increased NRT firing frequency following ICV administration of
NPY could be because of increased inhibition of GABA receptors with in the
NRT. This increase in firing frequency of NRT neurons may contribute to
seizure suppression by increased deposition of inhibitory GABA in thalamus
and causing decreased GABA mediated hyperpolarization of the
thalamocortical neurons there by maintaining the T type calcium channels
(Ca2+) in the inactivated state and inhibiting oscillatory thalamocortical
activity. During a seizure, the thalamocortical circuit goes into hyper-
synchronous state and we found a decrease in waveform correlation of field
potentials in cortex and NRT, this may also be due to increased firing rate of
NRT cells that might be disrupting the hyper-synchronous rhythm that
characterizes absence seizures. A previous study showed that enhanced
GABA activity in VB thalamus by injection of GABA transaminase inhibitor,
ɣ-vinyl GABA, aggravated seizures in GAERS(Liu et al., 1991) but this
technique is unlikely to be directly comparable with the increase in GABA-
ergic transmission proposed here. Another argument is that NPY may be
acting on specifically on interneurons present in cortex and NRT (Kuruba et
al., 2011). Further experiments with agents selective for NPY receptor
subtypes, and assessing changes in the firing of specific interneurons will
provide more information on the possible mechanisms of seizure suppression.

108
Fig. 3.9
Fig 3.9 the above figure illustrates the possible mechanism of seizure suppression and
increase in the interictal neuronal firing frequency of NRT neurons in GAERS.
Further, NPY injections led to a significant reduction in the correlation of
waveform activity between the NRT and cortical region suggesting an effect
on the overall thalamocortical network. As this occurs during the ictal period,
it may suggest a separate mechanism from the increased firing rate
interictally. We have analysed auto correlation and cross correlation with
custom made programs that provide a critical point or threshold to assess the
significance in autocorrelation and cross correlation. All statistical analysis
was performed by using widely accepted parameters for this kind of data.
Thalamus
Cortex
Layer IV and V
Excitatory glutamergic connections
Inhibitory gabaergic connections
NRT
+
+
−
+
−
−
+
−
Increased inhibition of GABA with in NRT
Increased NRT firing, which causes increased inhibitory GABA in thalamus resulting in decreased hyperpolarization in TC circuit
Possible mechanism of action for seizure suppression and concurrent increase in NRT neuronal firing

109
Further experiments with agents selective for NPY receptor subtypes, and
assessing changes in the firing of specific interneurons will provide more
information on the possible mechanisms of seizure suppression.
Previous studies from our laboratory have shown that ICV administration of
NPY has seizure suppressive effects (Stroud et al., 2005) that were mediated
through Y2 receptors (Morris et al., 2007, van Raay et al., 2012). The cellular
or molecular mechanism in suppression of seizures is poorly understood.
Reports from previous studies indicate that Y2 receptor is critical to mediate
the anti-epileptic actions of NPY (Colmers et al., 1991, Greber et al., 1994b,
Schwarzer et al., 1998, El Bahh et al., 2005). A study in rat brain slices
reported that NPY produced inhibitory effects on NRT and VB neurons
mostly via activation of GIRK channels coupled to NPY Y1 receptor (Sun et
al., 2001a). However, this study was performed in brain slices and on
stimulated spike responses. The neuronal and network dynamics are different
in live and intact brain compared to brain slices. Brain slices has limitations,
brain slices are not ideal to investigate neuronal excitability and pathological
oscillations of absence seizures, which arise from spontaneous hyper-
synchronous thalamocortical oscillations because of massive disconnections
involved in the preparation and lack of spontaneous firing in neurons. We
suspect that this is not the mechanism underlying our observation as increase
in K+ current would likely reduce spontaneous firing and the studies in vivo
strongly support predominant Y2 effect. Nevertheless, Sun et al demonstrated
that repetitive burst stimulation of the NRT induces NPY release that inhibits
the NRT by activating the Y1 coupled GIRK channels and t this endogenous,
oscillation induced NPY release acts to suppress the oscillations (Sun et al.,
2003). In the same paper it is shown that NPY Y2 receptor activation (by
application of NPY 3–36) decreased spontaneous and evoked GABA release

110
(Sun et al., 2001a, Sun et al., 2001b) in NRT. Overall, these studies on brain
slices may not be discordant with the current study, given the large
differences in preparation and even possibly complementary effects.
Unfortunately, NPY Y2 receptor antagonists were not available at that time to
investigate the effect of endogenous NPY Y2 activation on oscillations in
thalamic slices. The authors of these studies agree to the fact that NPY Y2
and NPY Y5 receptor subtypes may also be involved in the modulation of
neuronal excitability and GABA release (Sun et al., 2001a). The previous
results from our lab indicate that NPY mediates its anti-epileptic effects
primarily through Y2 receptor subtype (Morris et al., 2007, van Raay et al.,
2012). In vitro patch clamping with specific NPY receptor subtype agonists
and antagonists will be extremely useful to clarify these mechanisms.
NPY receptor subtypes Y1 and Y5 are primarily situated on somata and
dendrites of the neurons. These receptors act to inhibit their activity via
activation of G-protein-activated inwardly rectifying potassium channels
(GIRK). NPY Y2 receptors are predominantly located on nerve terminals and
act to inhibit GABA and glutamate release via inhibition of voltage-gated
calcium channels (Sun et al., 2001a, El Bahh et al., 2005). Possibly, NPY
might be acting post-synaptically via Y1 or Y5 receptor enhancing
GABAergic inhibition with in the NRT or cortex (decreasing the
hyperpolarizing output to VB), or pre-synaptically via Y2 receptor on the
NRT neurons projection on to VB, inhibiting GABA release and thus
reducing the hyperpolarization in these neurons that promote oscillatory
rhythmic activity during absence seizures (Stroud et al., 2005, Morris et al.,
2007).
In addition to anti-epileptic activity, NPY mediates other physiological
responses. NPY is anxiolytic, appetite stimulant, and shows cardiovascular

111
effects (Zini et al., 1984, Cabrele et al., 2000b). Nevertheless, the appetite
stimulating effects of NPY are primarily mediated by Y1 and Y5 receptors
(Gerald et al., 1996, Cabrele et al., 2000a, Chaffer and Morris, 2002).
Therefore, activating the NPY Y2 receptor could be a novel and effective
therapeutic option for treating absence seizures without inducing unwanted
side effects. Most importantly, NPY has a strong seizure suppressant effect at
very low concentrations than other drugs and difficult to compare as dose-
response study in not performed in this study. Other first line drugs for
absence seizures such as valproate and ethosuximide are effective but can
cause adverse effects in 40% of patients and are ineffective in 20% of patients
(Kwan and Brodie, 2000, Murphy and Delanty, 2000) and benzodiazepines
are highly sedative. ICV and focal administration studies of NPY Y2 agonists
and antagonists might provide more information on the mechanism of action
of NPY.
It would be more interesting to study effect of NPY at network level. One of
the limitations of this juxtacellular cellular recording is the sample size, we
could record from a maximum of two neurons at a time to interpret the
changes at network level. This may be the reason we do not have any result in
oscillation index and synchronization index to assess the changes at network
level. Multi electrode or Tetrode technology can be adopted to investigate and
understand network dynamics and properties of one region. Tetrode
technology is one of the novel techniques developed in neurophysiology, with
which one can record and analyse neuronal activity of more than two neurons
at a time.
In conclusion, the results of this study demonstrate for the first time that
exogenous NPY suppresses seizures concurrently increasing the mean firing
frequency of the NRT cells interictally in a model of genetic absence seizures.
A significant decrease was found in waveform correlation between NRT and

112
cortex suggesting the effect of NPY at network level. This decrease was
observed waveform correlation was observed during seizures. It is possible
that this may be attributed to the increased firing rate of NRT neurons
observed through disruption of hyper-synchronous rhythm seen in absence
seizures. Further investigations using focal injections are required to delineate
the site and mechanism action of NPY. Endogenous NPY release in the NRT
may play a critical role in decreasing the number of seizures and length in
GAERS. Moreover, NPY related mechanisms could be used to develop novel
anti-seizure therapies for absence seizures and other genetic generalized
epilepsy syndromes.

113
CHAPTER 4
EFFECTS OF NPY FOCAL INFUSIONS ON SINGLE
NEURONAL FIRING PATTERNS IN RETICULAR
THALAMIC NUCLUES CELLS IN A GENETIC RAT
MODEL OF ABSENCE EPILEPSY (GAERS).
4.1 Introduction
Rhythmic neuronal activity in the thalamocortical circuit vary from
moderately synchronous oscillations during sleep to hyper-synchronous
oscillations that characterize SWDs associated with absence epilepsy
(Huntsman et al., 1999). The NRT is considered to be a critical primarily
inhibitory component of the thalamocortical circuit important in generating
the normal thalamocortical rhythmicity of sleep wave and plays a critical role
in the maintenance and progression of abnormal rhythms associated with
absence seizures (Danober et al., 1998, Pinault and O'Brien, 2005). The NRT
consists of a large number of GABAergic cells (Houser et al., 1980, Pinault,
2004). The pathological rhythmic oscillatory activity during absence seizures
is synchronized by these NRT neurons, critically mediated by low threshold
calcium spikes generated in a rhythmic fashion (Steriade and Deschenes,
1984a).
Recent studies showed that NPY suppressed absence seizures in GAERS
model of absence epilepsy (Stroud et al., 2005, Morris et al., 2007), mainly
mediated through the NPY Y2 receptor subtype (Morris et al., 2007). Another
study from our lab investigated the site within the thalamocortical circuit that
effectively suppress seizures on EEG in GAERS upon NPY infusions. The
seizure suppression effect on seizures in freely moving GAERS were assessed
by focal microinjections of NPY into main regions of thalamocortical circuit:

114
the primary somatosensory cortex (S1), the secondary somatosensory cortex
(S2), the ventrobasal thalamus (VB) and the NRT. Focal infusions of NPY into
primary somatosensory cortex (S1), the secondary somatosensory cortex (S2)
resulted in a significant decrease of time spent in seizure over the 90-min EEG
recording in a dose dependent manner and a significant but small decrease in
average seizure duration with NPY (0.5nmol) administration into the NRT
(van Raay et al., 2012). The results of the previous chapter showing ICV
administration of NPY significantly suppresses seizures by a decrease in total
length of seizure, and percentage of time spent in seizures, complementing
these studies. Results of neuronal firing patterns showed increased firing
frequency of NRT neurons in the interictal period at the cellular level and also
decreasing the waveform correlation between cortex and NRT at the network
level confirming the critical role of the NRT in absence seizures. These
results were obtained using ICV administration of NPY. With the limitations
in the technique and complexity of the thalamocortical circuit, region specific
effect and mechanistic hypotheses for seizure suppression effect remain
tentative. A suggestive mechanism (increased firing rate) for seizure
suppression had been identified in the previous chapter, it was important to
try to assess the direct effect of NPY on single neurons to attempt to
differentiate direct effects on NRT neurons from possible indirect effects, for
example cortical inputs. It was hypothesized that focally administered NPY
would have a direct effect on NRT neuronal firing. Clarification of single
neuron effects would also contribute to an explanatory framework for the
potent anti-seizure activity of NPY.
To better understand the regional specific changes and to examine whether
the increase in neuronal firing frequency is a specifically NRT related
mechanism that supplements the mechanism of seizure suppression by NPY,
the effects of focal administered NPY on neuronal firing patterns of NRT

115
were studied. This is the first study where NPY is administered micro-
iontophoretically onto the NRT invivo to assess the changes in the neuronal
firing patterns upon direct administration of NPY.
4.2 Methods
The data presented in this chapter derives from single cell juxtacellular
electrophysiology recordings before and after the focal administration of
NPY. Focal administration of NPY was achieved with micro iontophoresis
and is complimented by the usage of multi-barrel electrodes. All the
experiments were conducted on male GAERS aged >12weeks.
4.2.1: Anaesthesia and surgery
For detailed information on animals, anaesthesia and preparative surgeries
like penile vein catheterization and tracheotomy please refer to sections 2.2,
2.3.1, 2.3.2, 2.3.3. Briefly, rats were anaesthetized with ketamine/xylazine
(3:1 ratio) at 0.1ml /100gm of rat. Penile vein catheterization and
tracheostomy were performed, and the rat was then placed in a stereotaxic
frame and connected to a mechanical ventilator for artificial ventilation.
Neurolept anaesthesia was initiated before the end of general anaesthesia
(Figure 3.1).
4.2.2 EEG electrode implantation
EEG electrodes were implanted to monitor EEG of the rat continuously
though out the experiment. For detailed description of EEG electrode
implantation please refer to section 2.3.4. Briefly, three active electrodes were
implanted on rat skull to record EEG (Figure 2.1). Continuous monitoring of
EEG will offer information about the physiological state of brain during the
experiment.

116
4.2.3 Electrophysiology
The basic surgery and recordings protocols for electrophysiology are
described in detail in section 2.4. For this particular study few changes have
been made to the basic protocol which is described in section 2.8. Briefly, a
double barrel borosilicate glass electrode was pulled to a diameter of ≈2
micron (15-30 MΩ). One barrel was back filled with 1.5% neurobiotin and
other barrel is filled with saline or NPY for control and NPY investigations
respectively. The electrode was then slowly lowered to NRT to locate a stable
firing neuron (Figure 4.1). Once a stable neuronal recording was achieved, 15
mins of baseline recordings were performed followed by focal injection of
saline/NPY for control and NPY studies. Post NPY/control recordings were
performed for 15 mins (Figure 4.2). NPY (0.5mM) and saline were injected
focally using a microiontophoresis technique (Union-40 iontophoresis pump,
kation scientific) (Hicks et al., 1993, Ouyang and Wang, 2000, van Raay et
al., 2012) (Figure 4.1). 200nA of cationic current was used to
microiontophorese NPY and a 40nA of anionic current was applied as a
holding current for NPY. These iontophoretic currents had a minimal effect
on the quality of recordings. The electrical interference between micro-
iontophoretic unit and the recording unit caused a decrease in the amplitude of
the spike by 20% and an increase in baseline noise levels in baseline but this
did not alter the stability of the recordings. Moreover we never lost a neuron
or recording because of this interference. All recordings were performed
simultaneously with EEG and recorded cells were again labelled
iontophoretically for later histological identification (Figure 4.1).

117
Figure 4.1 The above Figure illustrates the focal administration protocol: multiple barrel electrodes
were used in this procedure, here one barrel is filled with neurobiotin to record neuronal activity
and another barrel is filled with NPY/Saline and is infused iontophoretically on the recording
neuron. Alterations in the neuronal activity were assessed after the micro-iontophoretic
administration of NPY.
Figure 4.2 Timeline for the electrophysiological recordings performed under neurolept anaesthesia
for focal infusion experiments. In this procedure multi-barrel electrodes are lowered into brain
region and neuronal activity is recorded. In stable neuronal conditions, 15 minutes of baseline
recordings were performed, NPY is infused locally via micro-iontophoresis and 15 minutes of post
NPY recordings were performed.
Neurodata dual channel amplifier
Neuronal recording
Electrophysiological recordings
Perfusion and
Baseline recording Post NPY recording Microiontophoresis of NPY 15 min 15 min

118
4.2.4 Juxtacellular labelling and histology
The labelling histology protocols have been described in sections 2.6 and 2.7.
Briefly, at the end of the experiment, recorded neurons were labelled by
passing multiple depolarising currents through the neurobiotin-filled
recording electrode. The rat was then perfused via the left ventricle and the
brain was retained for histology. Histology was performed on the retained
brain sections to identify the cell location microscopically referring to
stereotaxic atlas (Paxinos G, 1998) (Figure 2.4).
4.2.5 Data acquisition and analysis:
Detailed information on data acquisition and analysis has been described in
section 2.10. In this chapter we have analysed the neuronal firing patterns of
NRT neurons interictally using the parameters described in the section 2.10.
Briefly, five parameters were used to quantify interictal firing patterns (1)
Mean AP firing frequency (MAPF), the average action potential firing
frequency, (2) Percentage of AP’s in burst (PAPB), the proportional of action
potential firing in bursts in a recording. Bursts were identified as a group of
action potentials with interspike interval less than 6ms (3) Mean number of
AP’s per burst (MAPB). (4) Maximum number of APs per burst (MxAPB).
(5) Intraburst frequency (IBF) is defined as the frequency at which action
potentials within the burst.
Relevant comparative t-test was used to assess significant differences in
neuronal firing patterns in response to individual treatments (saline/NPY). To
assess significance for this kind of data, a paired and non-parametric T-test
(Wilcoxon matched pair test) is performed to assess significance between two
individual treatments.

119
4.3 Results:
In this chapter, the changes in neuronal firing patterns of the NRT neurons
were assessed before and after focal administration of NPY. Similar to our
early findings with ICV NPY infusions, interictal mean firing frequency was
significantly increased after the focal administration of NPY (n=8, p=0.003)
(Figure 4.3). A significant decrease was also found in intra burst frequency
after focal NPY administration (n=8, p=0.01) as well as with saline control
experiments when compared with baseline intraburst frequency (n=9,
P=0.003) but this change was not significant when compared between saline
and NPY infusions. To assess the change in firing patterns between saline and
NPY infusion, a percentage difference of neuronal firing patterns between
treatment (NPY or saline) and baseline was calculated and compared between
saline and NPY focal infusions. A significant increase in the percentage
change of mean firing frequency was observed after NPY infusions (n=9,
p=0.0002) (Figure 4.3). No change was found in all other parameters. Control
saline experiments did not alter any firing patterns when compared with NPY.

120
Figure 4.3 This picture illustrates the results of focal NPY injections on the single neuronal firing
patterns. NPY was injected focally on the recorded neurons microiontophoretically and is found to
significantly increase the firing frequency of NRT cells interictally (n=9, P=0.003). The percentage
difference between treatment and baseline was calculated and compared to assess the change in
firing patterns between saline and NPY focal infusions. A significant increase in the percentage
change of mean firing frequency was found in NPY infusions (n=9, p=0.0002).
Firi
ng F
requ
ency
in H
z
n=9, P=0.003
**
n=9, P=0.0002
Perc
enta
ge ch
ange
in m
ean
firin
g fr
eque
ncy
***
2 m
V 2
mV
0.1
0.1
Firing rate before focal administration of NPY
Firing rate after focal administration of NPY

121
Figure 4.4 This is an example recording from focal infusion studies showing the firing frequency
of NRT neurons before and after focal administration of NPY, a strong increase in the neuronal
firing frequency after focal NPY administration.
NRT interictal neuronal firing
patterns
Focal NPY induced Changes
MAPF Increased significantly
% Burst No significant difference
MAPB No significant difference
MxAPB No significant difference
IBF No significant difference
Figure 4.5 The above table shows the results of all the interictal parameters of neuronal firing
patterns observed in this chapter.
4.4 Discussion:
The thalamocortical network involving somatosensory cortex, VB thalamus
(VB) and NRT are responsible for generation and progression of seizures
absence seizures (Danober et al., 1998). The effects of ICV administration of
NPY on cortical EEG, neuronal firing patterns of thalamocortical circuit
neurons, rhythmicity in the neuronal firing patterns of NRT neurons,
synchronicity of the firing in NRT neurons and waveform correlation between
cortex and NRT were described in the previous chapter. In the present chapter
effects of focally administered NPY on neuronal firing patterns of NRT
neurons have been investigated. Neuronal firing patterns between treatments
Fig 4.5

122
(saline/NPY) and baseline were compared. A significant increase in the
interictal neuronal firing frequency was found in NRT neurons after the focal
administration of NPY. We also found a significant decrease in intraburst
frequency of the interictal NRT neuronal firing after NRT firing. A similar
effect was found with saline infusions which were later found insignificant
when compared between saline and NPY. Control experiments with local
saline administration had no effect on neuronal firing patterns of NRT
neurons. All recordings were performed in the NRT region: the location and
morphology of these neurons were not compared to analyze the relative
change in effect for each neuron. The relative change in firing frequency after
NPY infusions could depend on the independent properties of the cells that
were recorded. In the present chapter we replicated the findings of the
previous chapter of an increase in the neuronal firing frequency of NRT after
ICV NPY administration. The mechanism of NPY effect via ICV application
in the previous chapter is further clarified by local application of NPY using
multiple barrel pipettes. Even though these two studies cannot be compared in
terms of doses and the method of drug infusions these observations strongly
suggest a direct effect of NPY on NRT neurons rather than an indirect effect
on other thalamocortical structures as could occur following ICV NPY
injection.
Possible mechanisms for increased NRT firing frequency following focal
administration of NPY could be because of increased inhibition of GABA
receptors with in the NRT. The increased firing rate of NRT cells may be
disrupting the hyper-synchronous rhythm that characterizes absence seizures.
Another argument is that NPY may be acting on specifically on interneurons
(Bacci et al., 2002, Kuruba et al., 2011). NPY receptor subtypes Y1, Y2, and
Y5 have all been demonstrated to be present in thalamocortical circuit
(Dumont et al., 1993, Sun et al., 2001a) and mediate NPY responses. The

123
anti-epileptic effect of NPY on the pathological oscillatory activity underlying
absence epilepsy is mainly mediated via the Y2 receptor (Morris et al., 2007).
In contrast a number of studies have reported the role of different NPY
receptor subtypes in suppression of other form of epilepsy (Reibel et al.,
2001). NPY specific receptor subtypes Y1 and Y5 primarily located in
postsynaptic position on somata and dendrites of neurons may act directly to
inhibit their activity by activation of G-protein-activated inwardly rectifying
potassium (GIRK) channels. On the other hand NPY Y2 receptors are located
at presynaptic positions on nerve terminals of neurons and act to inhibit
GABA and glutamate release via inhibition of voltage-gated calcium channels
(Sun et al., 2001a, El Bahh et al., 2005). Possibly, NPY might be acting post-
synaptically via Y1 or Y5 receptor enhancing GABAergic inhibition within
the NRT, or pre-synaptically via Y2 receptor on the NRT neurons projection
on to VB (Stroud et al., 2005, Morris et al., 2007). Further experiments with
NPY receptor subtype Y1, Y2 and Y5 agonists and antagonists in vivo will be
extremely useful in clarifying these mechanisms.
Although the effects of NPY on single neuronal firing properties could
plausibly account for seizure suppression, it is of great interest to see if we
could identify evidence of altered interaction between different populations of
neurons in this complex network. Because of the small sample size (n= 9)
autocorrelations and cross correlation were not performed as in the previous
chapter. For this kind of analysis a larger sample size is required to achieve
significant results. Multi electrode or tetrode technology is the best alternative
to record and analyse from higher population of neurons to understand the
effect of focally administered NPY at network level.
The results from this suggest NRT as the critical structure in seizure initiation
and progression and NPY acts on this structure for effective seizure
suppression. Further investigation using agonists at specific receptor subtypes

124
Y2 and Y5 on NRT could reveal more information on the molecular reasons
explaining the antiepileptic effects of NPY. The NRT could possibly become
a target region in the brain for anti epileptic drug delivery for effective seizure
suppression.
The advantage of the technique used in this chapter is that we can investigate
the effect of focally administered drugs on the single neuron firing patterns in
vivo. Conversely, there are some limitations too 1) The present study is
limited to a single treatment at a time, we could only deliver either control or
NPY at one time given that only one microiontophoresis unit was available.
2) Neuro-active compounds with high ejection current (<200nAmp) cannot be
used with this type of micro-iontophoretic pump. 3) Sample size is another
limitation of this study, with this current technique we can record and analyse
changes in neuronal firing patterns of a single neuron. 4) Quality of the
recordings is at considerable risk with electrical interphase between recording
and iontophoresis unit which decreases the amplitude of the spike train and
increases the noise levels in baseline. This problem is also accountable for
single treatment at a time in this study. This problem can be overcome by
finding relatively more stable firing neurons with higher amplitude (eg: 2mV)
and proper grounding of the equipment.
Overall, data from this chapter confirmed that interictal neuronal firing is
increased with focal administration of NPY in NRT neurons suggesting a
direct effect of NPY on NRT neurons.

125
CHAPTER 5
EFFECT OF ICV ADMINISTERED NPY ON SEIZURE
INDUCING THRESHOLD UPON EXTERNAL
ELECTRICAL STIMULATION IN S2 REGION IN GAERS.
5.1 Introduction
The thalamocortical circuit involving NRT, VB thalamus and cerebral cortex
is critical for generation and progression of absence seizures (Danober et al.,
1998) (Figure 1.4 and 1.6). The neuronal mechanism underlying the
generation and progression of SWDs associated with absence seizures has
been the basis for many in vitro and in vivo studies (McCormick and
Contreras, 2001, Pinault and O'Brien, 2005). This gave rise to several
hypotheses that are widely debated, with opposing views about whether the
cortex, the thalamus or both play a pivotal role in generation of absence
seizures (Pinault and O'Brien, 2005).
Genetic rat model of absence epilepsy from Strasbourg (GAERS)
(Vergnes et al., 1987) and Wistar Albino Glaxo/Rijswijk rats (WAG/Rij)
(Coenen et al., 1992b) are two genetic rat models of absence epilepsy that
exhibit spontaneous and generalized absence-like seizures characterized by
SWDs on the EEG (Figure 5a, b). Recent studies using these genetic models
demonstrated that absence related SWDs initiate from a specific region
“focus”, residing in the somatosensory cortex. In freely moving WAG/Rij rats
using multi-site field recordings, Meeren et al in 2002 demonstrated that
seizures were initiated in the S1 of somatosensory region and then generalized
to other regions of cortex and thalamus. (Meeren et al., 2002). Studies by
Pinault have revealed that layer VI neurons of S1 region play a critical role in
proliferation of SWD (Pinault, 2003, Pinault et al., 2006). Recently an in vitro

126
study in brain slices from thalamocortical region in GAERS proved that the
SWDs were initiated in the deeper cortical layers before scattering further to
other cortical and thalamus regions (Adams et al., 2011).
However, recent studies from our group have also demonstrated the
critical role of secondary somatosensory cortex (S2) in seizure initiation in
GAERS (van Raay et al., 2012, Zheng et al., 2012). From the results of
previous chapter and other studies from our lab we know that ICV
administered NPY suppresses spontaneous seizures in GAERS (Stroud et al.,
2005, Morris et al., 2007, van Raay et al., 2012). In addition to these results, it
is also demonstrated that absence related SWDs could be triggered by
external stimulation in somatosensory cortex in GAERS (Zheng et al., 2012).
It is also shown in WAG/Rij rats that SWDs like after discharges were
observed in somatosensory cortex (Luttjohann et al., 2011). In this chapter,
we assessed the seizure induction threshold (amount of stimulation required to
induce absence seizures) to trigger a seizure in GAERS in S2 region and the
changes in seizure induction threshold in response to ICV administered NPY.
By this approach we aim to investigate the effect of NPY on seizure initiation
and propagation in the thalamocortical circuit. In previous studies it has been
shown that NPY suppresses spontaneous seizures in GAERS; here, we
hypothesise that ICV NPY will affect externally induced seizures by altering
the seizure induction threshold
5.2 Methods
The data presented in this chapter is derived from cortical stimulations
performed in the S2 region. External cortical stimulations were performed to
induce seizure like activity and the inducting electrical threshold was
calculated and compared before and after ICV administration of NPY. All the
experiments were conducted on male GAERS aged >12weeks.

127
5.2.1: Anaesthesia and surgery
For detailed information on animals, anaesthesia and preparative surgeries
please refer to section 2.2, 2.3.1, 2.3.2, 2.3.3. Briefly, rats were anaesthetized
with ketamine/xylazine (3:1 ratio) at 0.1ml /100gm of rat. Penile vein
catheterization and tracheostomy were performed. The rat was then placed in
a stereotaxic frame and connected to a mechanical ventilator for artificial
ventilation. Neurolept anaesthesia was started before the general anaesthesia
was completed.
5.2.2 EEG electrode and ICV cannula implantation
EEG electrodes were implanted to monitor EEG of the rat continuously
throughout the experiment. For detailed description of EEG electrode
implantation please refer to section 2.3.4 (Figure 2.1). Briefly, three active
electrodes were implanted on rat skull to record EEG. Continuous monitoring
of EEG provided information about the physiological state of brain during the
experiment. An ICV cannula was also implanted for central administration of
NPY, detailed description of the procedure is provided in section 2.3.4.
Briefly, an ICV cannula was implanted on the left hemisphere (A/P: 1mm,
M/L: -1.5 mm and -3.0 to -3.5mm deep relative to dura). The position of
cannula was assured by withdrawal of cerebrospinal fluid through the ICV
injecting line.
5.2.3 Cortical stimulations and seizure induction
Absence-like seizures were induced by external electrical stimulation in the
cortical regions such as in motor cortex, insular cortex S1 and S2 region.
Seizure inducing threshold was recorded in S2 region before and after ICV
administration of NPY, detailed information about the procedure is provided
in methods (section 2.5). Briefly, for this experiment a single craniotomy and

128
duratomy was performed (A/P 0.2mm, M/L 4.1). Glass electrodes with a
resistance of 2-5 mΩ were backfilled with 1.5% neurobiotin and were used
for stimulation and recording from S2 region of somatosensory cortex (S2). In
this method, a 2-second monopolar stimulus train (1 millisec square pulse, 7
Hz) of 7-Hz electrical stimulation was given to S2 region in GAERS to
induce SWDs in GAERS rats (n=8) as described and published in Zheng et al
(Zheng et al., 2012). A baseline recording was performed for 15 mins,
followed by ICV saline infusions. After few minutes of saline infusion,
current of 1-10milliAmps were applied by increasing 1milliamp each step
until the seizure was induced and seizure induction threshold was recorded.
Thirty minutes after seizure induction NPY was administered ICV and
seizure-inducing threshold was assessed at 15min, 1hour and 2 hour time
intervals (Figure 5.1). At the end of the experiment the whole region was
labelled iontophoretically for further histological identification.
Figure 5.1 This Figure show the three main structures of thalamocortical circuit involved in the
generation of absence seizures. In this procedure, blunt glass electrodes were lowered into S2
region of the cortex and external electrical stimulation were applied to induce an absence like
VB
NRT
S2
Recording electrode
Stimulation

129
seizure. On right is an exemplary picture of cortical stimulation that induces seizures similar to
spontaneous absence seizures in GAERS.
5.2.4 Juxtacellular labelling and histology
The labelling histology protocols have been described in sections 2.6 and 2.7.
Briefly, at the end of the experiment, recorded neurons were labelled
iontophoretically, rats were perfused and brain was retained for histology
(Figure 2.4). Histology was performed on the retained brain sections to
identify the cell location microscopically referring to stereotaxic atlas(Paxinos
G, 1998).
5.2.5 Data acquisition and analysis:
Detailed information on data acquisition has been described in section 2.10.
Briefly, seizure induction threshold in S2 region is assessed before and after
the ICV administration of NPY. Non-parametric and repeated measure Anova
was performed to analyse the statistical difference between the treatments.
Relevant comparative t-test was also performed to assess significance
between two individual treatment time lines.
5.3 Results:
Taking advantage of method to induce seizures by external stimulation in
cortical regions from recent studies (Zheng et al., 2012). We have
successfully triggered absence like seizures by external electrical stimulation
in motor cortex, insular cortex, S1 and S2 region, S2 being more hyper-
excitable and epileptogenic, requiring less intensity of electrical stimulation to
induce seizure. The threshold for induction of absence like seizures in S2 was
measured. The seizure induction threshold in the somatosensory cortex was
assessed before and after the ICV administration of NPY. Seizure induction
threshold for seizure induction before NPY administration was 1.20 ± 0.51
mAmp (n= 8) which was significantly increased to 3.05 ± 0.68 mAmps 15

130
minutes after ICV NPY infusion (n=8, p=0.02). The seizure triggering
threshold showed an increasing trend after NPY infusion (2.00 ± 0.44 1 hour
after ICV NPY administration and 1.70 ± 0.50 2 hours after ICV NPY
administration, n=7) but this did not reach significance (Figure 5.2). No
difference was observed with control saline/saline treatment.
Figure 5.2 the above graph illustrates the results of ICV administration of NPY on the seizure
inducing threshold in the S2 region. ICV administration of NPY increased the seizure induction
threshold significantly 15 minutes after NPY infusion; an increasing trend was observed in seizure
induction threshold after 1 hour and 2 hours after NPY infusion but it did not reach significance.
n=8, P=0.01
*
P value – 0.25
Curr
ent i
n m
illiA
mps

131
Control experiments with saline did not show any significant changes to the seizure-inducing
threshold
5.4 Discussion:
A recent study from our lab demonstrated that SWDs could be induced by 7
Hz electrical stimuli in the cerebral cortex (Zheng et al., 2012). These
stimulations could reliably induce absence-like SWDs in S1, S2, or IC
(insular cortex) of somatosensory cortex in GAERS. However, the required
amplitude of electrical stimulation to induce seizure varied for each
experiment and also for the region of cortex where the seizure was triggered.
The seizure inducing threshold was significantly lower in S2 region (0.5 to 2
mAmps) than other cortical regions in GAERS, suggesting that the S2 cortical
region may have more pivotal role in generation of seizures than the S1region
(Zheng et al., 2012). External stimulations in S2 regions were performed
irrespective of the layers and the regions of stimulation were labelled
iontophoretically at the end of the recordings. The location of the recording
was later confirmed by histology. These evoked SWDs were similar in
morphology and frequency to typically occurring absence related spontaneous
SWDs. Zheng et al, has already described that these evoked seizures were
accompanied by similar behavioural characteristics such as immobility and
head nodding (Zheng et al., 2012). However, the evoked seizures can be
differentiated from spontaneous seizures as they longer than normal,
sometimes lasting for more than 30 seconds and also the evoked seizure starts
as soon as the stimulation ends as shown in figure 5.1. The seizure must be
induced at least 2 times with the same stimulation parameters to confirm that
the seizure is induced due to external stimulation.
It is significant that SWDs can only be induced when rats were quite awake
manifesting a desynchronised EEG. This state could be achieved by
anesthetizing the rat with neurolept anaesthesia, which keeps the rats in

132
pseudo-active state allowing this method to be implicated on anaesthetized
animals. In contrast, identical stimulations in the NEC rats could not induce
SWDs (Zheng et al., 2012).
Previous studies with ICV drug infusions decreased spontaneous absence
seizures after 90 min of drug infusion in a dose dependent manner (Stroud et
al., 2005, Morris et al., 2007) and also showed a similar prolonged effect on
spontaneous seizures by focal NPY administration (van Raay et al., 2012).
However, these prolonged effects were observed in spontaneous seizures
unlike in this study where we evoked seizures and used a standard dose of
1.5nmol. Given the fact that we could reliably induce absence like seizures
with external electrical stimulation and also that NPY suppresses naturally
occurring spontaneous seizures in GAERS, we investigated the seizure
suppression effect of NPY on induced seizures by evoking absence-like
seizures. We stimulated the S2 region of cortex to initiate seizures that
manifests similar hyper-resonant oscillatory thalamocortical activity allowing
us to investigate the alterations in this induced resonant activity in response to
NPY. In this chapter, we revealed that seizure induction threshold to trigger
SWDs was significantly increased after ICV NPY administration in GAERS.
As discussed in previous chapters, because of complexity of the network and
limitation of methodology a strong mechanistic hypothesis cannot be
formulated at this stage. In the previous chapter we demonstrated that the
seizure suppression effect of NPY is concomitant with increase in firing
frequency in NRT neurons interictally and a decrease in waveform correlation
between local field potentials of cortex and NRT.
From the above results, possible mechanism for escalation in the seizure
induction threshold might be enhanced GABAergic inhibition within NRT
instigating increased firing rate of NRT cells that might be disrupting the

133
hyper-synchronous rhythm and resonant oscillations in epileptic
thalamocortical circuit that characterizes absence seizures. This disruption in
NRT might restrict the generalization and spread of SWDs in the whole brain,
as the NRT is suggested to promote the propagation and synchronization of
SWDs (Kim et al., 1995, Cox et al., 1996). Another factor that facilitates the
thalamocortical circuit to express self-synchronising hyper excitable SWDs is
the ability of NRT and relay neurons to fire in phasic Ca2+ dependent burst of
action potentials mediated by T-type calcium channels. NPY would suppress
release of the inhibitory neurotransmitter GABA from NRT neurons
projecting to the thalamocortical neurons. This would lead to reduced GABA
facilitated hyperpolarization of the thalamocortical neurons, thus retaining the
Ca2+ T-channels in a de-inactivated state and preventing oscillatory hyper-
synchronous thalamocortical activity. Furthermore, previous studies have
confirmed that NPY receptors are located on the axon terminals of NRT
neurons projection to VB (Sun et al., 2001a), where they inhibit
neurotransmitter release by modulating the activity of voltage gated Ca2+
channels (Sun et al., 2001b). Another alternative possibility is that NPY may
be acting on specifically on interneurons present in cortex and NRT (Bacci et
al., 2002, Kuruba et al., 2011). Another study from our lab performed focal
injections of NPY in motor cortex, S1 and S2 cortical regions and found
greater seizure suppression effect in S2 suggesting this region is more critical
for seizure generation (van Raay et al., 2012). These results suggest that here
might be other cortical related mechanisms involved in the increase of seizure
induction threshold, which can be only clarified with focal NPY
administration experiments. Further in vivo ICV/focal NPY injection
experiments using specific receptor subtypes NPY Y1, Y2 and Y5 and more
specifically on interneurons involved in the thalamocortical circuit and similar
stimulation experiments with ICV/focal NPY administration in other regions

134
of the circuit might reveal more relevant information on underlying
mechanisms of NPY seizure suppression effect.
The results from this study suggest that NPY can not only suppress
spontaneous seizures, but also is effects against induced seizures, which
implies that NPY strongly suppresses seizures. An increase in seizure
induction threshold may be related to the increase in interictal NRT firing that
we found in previous chapters but another important consideration is that a
probable focus has been identified in the somatosensory cortex. Therefore it is
possible that ICV administered NPY also has effects more generally in the
thalamocortical circuit and perhaps has an interneuron specific effect. This
suppression could most possibly be mediated by affecting the initiation of
seizure or could be inhibiting these seizures to go into hyper synchronous
pathological oscillatory mode.

135
Fig5.3 Showing the possible mechanism for the increase in seizure induction threshold
The advantages of this study are that: it allows us to investigate the effects of
NPY on seizure induction in live animal models, it is already known that
electrical activity similar to SWDs has been observed in slice preparations but
an intact thalamocortical circuit is essential for expression of the seizure
(Gloor P, 1990, Vergnes and Marescaux, 1992) This method can be improved
with tetrode/multi-electrode technology to understand the dynamics of the
neuronal networks during pathological epileptic oscillations and their
response to antiepileptic drugs in different epileptic models. The only
drawback of this experiment is complexity in the methodology and restricted
to animals that are not freely moving.
Thalamus
Cortex
Layer IV and V
Excitatory glutamergic connections
Inhibitory gabaergic connections
NRT
+
+
−
+
−
−
+
−
Increased inhibition of GABA with in NRT
Increased NRT firing and this changes in the neuronal firing may possible disrupt the hyper-synchronous rhythm and resonant oscillations in thalamocortical circuit thus restricting the generalization SWD’s.
Possible mechanism of action for seizure suppression and concurrent increase in NRT neuronal firing
Fig 5.3

136
CHAPTER 6
General discussion and conclusion:
6.1 Key findings of this study:
Absence epilepsy is the most common type of generalized epilepsy,
characterized by symmetrical and bilaterally synchronous 3 Hz spike and
wave discharges (SWDs) in the electroencephalogram (EEG). The
thalmocortical circuit involving somatosensory cortex, VB thalamus and NRT
plays a critical role in generation of signature SWDs that characterizes
absence seizures. One third of the patients with absence seizures do not have
adequate seizure control with presently existing anti-epileptic drugs. This
increased the desire for better therapeutic agents to treat patients with absence
epilepsy. NPY related mechanisms may represent a novel therapeutic
approach for treating generalized epilepsies.
NPY is a naturally occurring neurotransmitter, found abundantly in many
brain regions and modulates many physiological responses such as sleep,
appetite, memory and stress etc. NPY more importantly is also shown to
mediate seizure suppression in different models of epilepsy. The underlying
neuronal mechanisms for the seizure suppression by NPY are still unknown.
The aim of the research conducted in this thesis was to investigate the effects
of NPY on single neuronal firing patterns of the thalamocortical circuitry
neurons to understand the underlying cellular mechanism of seizure
suppression in genetic model of absence seizure (GAERS) and the effect of
NPY on network dynamics. In this study we have investigated the effects of
NPY on EEG, single neuronal firing patterns and other properties that signify
the hyper-synchronous oscillation in the circuit.
For experiments in this research, genetic absence epilepsy rats from

137
Strasbourg (GAERS) were used. This is a widely accepted rat model for
absence epilepsy with similar pathophysiological and pharmacological
characteristics as human absence seizures. Juxtacellular electrophysiology
recordings were performed in rats in vivo under neurolept analgesia to
examine alterations in neuronal firing in the thalamocortical regions during
interictal and ictal periods that could be associated with seizure suppression
and alterations in the seizure threshold.
Electrophysiology recording under neurolept anaesthesia is well-
established technique and has been previously used in many studies to
understand the neural firing patterns of several types of neurons. In
current thesis, typical absence related SWDs are more related to a state
of awakeness. In GAERS rats, the characteristic SWDs and 5-9 Hz
oscillations arise from a relatively desynchronized EEG activity(Pinault et
al., 2006). Neurolept anaesthesia maintains the rats in the similar state of
immobile wakeful ness or drowsiness (Pinault et al., 2001, Pinault et al.,
2006). Such experimental conditions are suitable for recording SWDS at
similar frequency to those of freely moving rats. It has also been shown
that cellular and network mechanisms underling 5-9 Hz oscillations
during neurolept anaesthesia are similar to those in non anesthetised
intact brain(Pinault et al., 2006).
The key findings from the current project were:
6.1.1. Effect of NPY on seizures:
These findings of this chapter indicate that we have successfully are
replicated our previous studies findings demonstrating seizure suppression
after NPY infusion both ICV and focally. In the current study, NPY had a
strong seizure suppressant effect. A significant reduction was found in the

138
total length of seizures and percentage of time spent in seizures after ICV
NPY administration.
6.1.2. Effect of NPY on interictal Neuronal firing patterns
This is a novel and key finding of the current study. ICV administration of
NPY led to a significant increase in the neuronal action potential firing
frequency in the NRT region concurrently supressing seizures. This may
account for the possible mechanism of seizure suppression and also suggest
that NRT could be the critical structure in thalamocortical circuit involved in
the absence seizures.
6.1.3. Effect of NPY on the waveform correlation between cortical EEG and
local field potentials of NRT.
This is another novel finding from this study. ICV administered NPY
decreased the waveform correlation between cortical EEG and local field
potential recorded from the NRT region. These results compliment the
precious results suggesting that the decrease in correlation could be because
of changes in neuronal firing patterns demonstrated in NRT. These data also
suggest that, NRT could be critical structure involved in seizures and that
NPY could be acting more at a network level.
6.1.4. Effects of Focal administration of NPY on neuronal firing pattern in
NRT region:
This is again a new key finding. Significant changes in the neuronal firing
patterns in NRT were observed after ICV NPY infusions. We found similar
results after focal administration of NPY, whereby interictal mean firing
frequency was significantly increased after NRT administration of NPY. The
difference in percentage of neuronal firing frequency between NPY and
baseline group was assessed. A significant increase in percentage change of

139
mean firing frequency was found after NPY infusions. These results again
suggest that the NRT could be the target region in the thalamocortical circuit,
where NPY acts to suppress seizures.
6.1.5. Effects of NPY on Seizure induction threshold:
Previous studies showed that NPY suppresses spontaneous absence seizures
in GAERS. This study showed that ICV administered NPY suppresses
seizures evoked by external stimulation. Most importantly we investigated the
changes in the threshold to induce a seizure following a cortical stimulation.
The seizure inducing threshold doubled after ICV administration of NPY. The
maximal effect was seen during the initial recording period, i.e. 15 minutes
after NPY infusion as compared to the observations made during later time
points. NPY injected ICV may have effects throughout the thalamocortical
circuit and perhaps has an interneuron specific effect.
6.2 Integration and interpretation of results:
A large number of studies have described the effects of NPY on seizure
suppression Previous studies on focal seizures suggest that NPY is a potent
anti-convulsant, when considering the effect of endogenous NPY deficiency
(Erickson et al., 1996, DePrato Primeaux et al., 2000, Richichi et al., 2004,
Woldbye et al., 2005) and NPY administration on seizures (Colmers and
Bleakman, 1994, Greber et al., 1994a, Woldbye et al., 1996, Klapstein and
Colmers, 1997, Woldbye et al., 1997, Woldbye, 1998, Vezzani et al., 2000,
Reibel et al., 2001, Silva et al., 2001, El Bahh et al., 2002, Husum et al., 2002,
Reibel et al., 2003, Woldbye and Kokaia, 2004). Studies from our lab
suggested the anti-epileptic role of NPY in generalized seizures by

140
suppressing absence seizures in a genetic rat model of absence epilepsy
(Stroud et al., 2005, Morris et al., 2007, van Raay et al., 2012).
Despite a large number of studies suggesting a role of NPY on seizure
suppression the mechanism underlying the seizure suppression is still highly
debated. In the present research, we were able to show similar seizure
suppression effects of NPY administration in GAERS, when they were
anesthetized under a neurolept cocktail. We also demonstrated that NPY
mediated seizure suppression was associated with an increased interictal
firing frequency of neurons in the NRT in these rats. This escalation in NRT
firing frequency following ICV NPY infusion could be possibly due to an
increased inhibition of GABA receptors within the NRT and contribute to
seizure suppression by increased deposition of inhibitory GABA in thalamus
decreasing GABA mediated hyperpolarization of the thalamocortical neurons.
Given the complexity of the network and limitations of the NPY infusion
method, there could be many other mechanisms associated with the increase
in firing frequency and seizure suppression. To get away from these
limitations, we used NPY focal injections to study the effects on neuronal
firing patterns of NRT, which showed similar results to ICV administration of
NPY by increasing the interictal firing frequency of NRT neurons. This
confirmed the direct effect of NPY on NRT and not a network-dependent
effect.
During a seizure, structures involved in thalamocortical circuit engage
themselves in a hyper-synchronous oscillatory rhythmic state. In this thesis,
we have also demonstrated a decrease in waveform correlation in local field
potentials between the cortex and NRT during seizures after ICV NPY
administration. Increase in seizure induction threshold may be related to the
increase in interictal NRT firing and these changes in the neuronal firing

141
frequency or increased firing rate of NRT cells might be disrupting the hyper-
synchronous rhythm that characterizes absence seizures. These results
suggested an effect of NPY on overall thalamocortical circuit, possibly acting
on interneurons present in the network but the results from focal injection of
neuronal firing patterns in NRT further confirmed the direct effect of NPY on
the NRT.
Zheng et al from our lab recently demonstrated that absence related SWD’s
were first identified in S2 region before spreading to other regions of the brain
(Zheng et al., 2012), it was also demonstrated that absence like SWDs could
be induced in cortical regions by 7 Hz electrical stimuli in the cortical regions
with a lower intensity in S2, suggesting its critical role in generation of
seizures compared to other cortical regions IC and S1 (Zheng et al., 2012). In
this thesis, we have adapted the same technique to induce seizures in S2
region, which is considered as focus for seizure generation and investigated
the effects of NPY on hyper resonance of the thalamocortical circuit. In this
thesis, we showed an increase in the seizure induction threshold to induce a
SWD after ICV NPY administration. As discussed in previous chapters,
because of complexity of the network and limitation in methodology a strong
mechanistic hypothesis cannot be formulated at this stage. Basing on the
results from previous chapters in this thesis, mechanism for escalation in the
seizure induction threshold might be enhanced GABAergic inhibition within
NRT instigating increased firing rate of NRT cells that might be disrupting
the hyper-synchronous resonant oscillations that signifies absence seizures, as
NRT is postulated to promote the spread and synchronization of SWDs (Kim
et al., 1995, Cox et al., 1996), this disruption in NRT might restrict the
generalization and spread of SWDs in the whole brain.

142
However, the cellular and molecular mechanism underlying NPY related
suppression of seizures is still unknown and the specific receptor subtype
mediating the seizure suppression effect is highly debated. Reports showed
that NPY receptor subtypes Y1 (Sun et al., 2001a, Sun et al., 2001b, Sun et
al., 2003), NPY Y2 (Greber et al., 1994b, McQuiston and Colmers, 1996,
Schwarzer et al., 1998, Furtinger et al., 2001, Morris et al., 2007, van Raay et
al., 2012) and NPY Y5 (Woldbye et al., 1997, Marsh et al., 1998, Morris et
al., 2007) may be mediating seizure suppression effect by NPY in different
models and types of epilepsies. Previous studies from our lab in freely
moving animals indicate that NPY primarily mediates its anti-epileptic effects
via Y2 receptor subtype (Morris et al., 2007, van Raay et al., 2012). Further
research with NPY specific receptor subtypes especially Y2 will clarify the
possible mechanisms underlying the seizure suppression effect of NPY.

143
Figure 6.1 Above is the schematic diagram of neuron illustrating the locations of Y1, Y2 and Y5
receptors for better understanding of the possible mechanism.
Y1 receptors predominantly located post-synaptically on somata and dendrites of the gabaergic NPY neurons and glutamergic neurons in cortex and VB Y5 receptors predominantly located post synaptically
Y5 receptors are located pre-synaptically
Fig 6.1

144
Fig6.2 The above figure illustrates the possible mechanism for seizure suppression,
increase in the neuronal firing patterns in response to NPY administration ICV and focally
and also possible mechanism for increase in seizure induction threshold and decrease in
waveform correlation
Thalamus
Cortex
Layer IV and
Excitatory glutamergic connections
Inhibitory gabaergic connections
NRT
+
+
−
+
−
−
+
−
Increased inhibition of GABA with in NRT
Increased NRT firing may disrupt the hyper-synchronous rhythm and resonant oscillations in thalamocortical circuit thus restricting the generalization
Possible mechanism of action for seizure suppression and concurrent increase in NRT neuronal firing
Increased NRT firing, which causes increased inhibitory GABA in thalamus resulting in decreased hyperpolarization in TC circuit
Another possibility is that NPY could be primarily acting on interneurons present in the circuit but focal injection studies suggest that NRT could be the key structure involved in seizure suppression effect of NPY
Fig 6.2

145
6.3 Limitations of this study and experimental considerations:
Longitudinal studies in humans have shown an association of adverse prenatal
and postnatal experiences on the development of neurological disorders later
in life (Lupien et al., 2009). The mechanisms related to these effects are
difficult to study in humans. Furthermore, some studies have also argued an
opposite cause-and-effect interpretation of this association, i.e. that children
with pre-existing brain abnormalities are at higher risk of abuse. Therefore,
studies in animal models including rodents and primates have provided a rich
framework for hypothesizing the changes in neurodevelopment and
behavioural outcomes, and the related mechanisms.
6.3.1 Limitations in ICV NPY infusion study:
Despite having a rat model, these experiments cannot be carried out in freely
moving rats because of the experimental complications and surgeries. The
alterations in neuronal firing patterns induced by ICV and focally
administered NPY have been observed in rats under neurolept analgesia.
Neurolept anaesthesia is a mixture of drugs that can set the brain in a state
that electrophysiologically corresponds to immobile wakefulness, altering
between desynchronized and synchronized states that allow generation of
seizures similar to spontaneous epileptic seizures (Pinault et al., 2001).
However, neurolept analgesia itself is known to promote certain pathological
thalamic oscillations associated with absence-like seizures (Warter et al.,
1988, Inoue et al., 1994). All the recordings were conducted in rats during
predominantly desynchronized EEG conditions by titrating the levels of
analgesia as well as by monitoring and controlling critical physiological

146
parameters.
Another methodological consideration in ICV infusion studies is maintaining
the stability of the paired neuronal recording for longer periods is always an
issue. We have previously reviewed this concern in methods section 2.4.2 in
detail to achieve paired stable neuronal recordings.
6.3.2 Limitations in focal injection study:
Yet again this study is also restricted to anaesthetized animals. Maintaining
the quality and stability of the recording is always an issue with triple barrel
electrodes, which is addressed in the methods section 2.4 and 4.2.3.
6.3.3 Limitations in cortical stimulations study:
The cortical stimulations study has an experimental complication where
seizures can be triggered only when rats were in a state of quiet wakefulness
accompanied by desynchronized EEG. This state could be achieved by
anesthetizing the rat with neurolept anesthesia, which keeps the rats in
pseudo-active state allowing this method to be carried out on anaesthetized
animals (Zheng et al., 2012).
All the experiments in these studies were performed under neurolept
anaesthesia. Therefore all comparison was made between observations in rats
under neurolept analgesia.
NPY is believed to suppress absence seizures predominantly by two specific
receptor subtypes Y2 and Y5 (Morris et al., 2007, van Raay et al., 2012).
Further ICV and focal infusion experiments in GAERS rats are required to
investigate and understand the underlying mechanisms of seizure suppression
at molecular level. Recordings from chronically-implanted tetrodes in freely
moving conditions is one possibility that has further advantage, as it is

147
possible to sample a large population of neurons from a particular region,
which was not possible in the experimental set up of the current study.
6.4 Future research directions:
6.4.1 Investigating the particular receptor subtype with predominant
seizure suppression effect (ICV and focal administration)
The current study provided important information on changes in the neuronal
firing patterns in NRT associated with the seizure suppression and also about
the possible mechanisms of seizure suppression. It is important to investigate
the most possible mechanism and find a particular receptor subtype with
predominant seizure suppressing effect. Investigating the effects of ICV
administered Y2 and Y5 receptor agonists will provide extended knowledge
on the specific receptor subtype principally suppressing seizures and its
mechanism of action.
6.4.2 Investigating the effect of NPY and its subtypes at network level
This current study is limited to paired neuronal recordings. Much more
knowledge could be gained if we could sample multiple neurons from a
region at a time. Neuronal recordings with tetrodes will overcome the
disadvantage of limited sample size as in the current study, since a large
population of neurons can be recorded simultaneously and can be sorted and
analysed individually. Tetrodes can be used to record from multiple units
(max 4 units per tetrode) and more than one tetrode can be used in
experiments. Thus using the novel tetrode system recordings can be made
from 32-128 units at a time in one experiment, from different regions of the
brain. By combining ICV infusion technique with Tetrode recording
techniques we can analyze changes in the neuronal properties in a large
sample size and from different regions at a time. In addition to these

148
advantages, it will also facilitate recordings in freely moving rats. This
combined technique would also enable assessment of correlations between
two different regions a t a time using large sample sizes and help us
understand the network dynamics and also the mechanism of seizure
suppression at a network level. In addition to these advantages, it will also
facilitate recordings in freely moving rats.
6.4.3 To investigate the effect of NPY and its subtypes on neuronal firing
patterns of freely moving rats
This study provided insight on possible mechanisms related to seizure
suppression and alteration in the neuronal firing patterns in thalamocortical
circuit neurons during ictal and interictal periods in controlled conditions
under neurolept analgesia. As discussed previously, it cannot be certain that
the observed findings are not specific to the experimental conditions. Hence
future experiments with a similar study design should be conducted in freely
moving conditions. Neuronal recordings with tetrodes chronically implanted
in freely moving rats have proved useful in understanding hippocampal cell
firing during interictal (Zhou et al., 2007), ictal (Bower and Buckmaster,
2008, Cymerblit-Sabba and Schiller, 2010)(Bower and Buckmaster, 2008,
Cymerblit-Sabba and Schiller, 2010, 2012) and postictal states in rodent
models. Thus, similar experiments in GAERS could assist in overcoming the
limitations of the current study. In addition to these advantages, recordings in
freely moving rats would also allow precise monitoring of the
electrophysiological alterations in response to NPY and it receptor subtypes.
6.5 Other new therapeutic options:
Until now, ethosuximide and valproate remain the principal anti-absence
drugs. However, these drugs do not show adequate seizure control in some

149
patients and are not well tolerated in some patients as they cause unwanted
side effects (Panayiotopoulos, 2001). Thus recent research is focused on
developing drugs that are more effective and show minimal side effects.
Several experimental drugs are under development for treating absence
seizures and have also shown efficacy in animal absence epilepsy models:
1) Brivaracetam (BRV), a novel synaptic vesicle glycoprotein 2A ligand
(SV2A) demonstrated anti-absence activity in GAERS (Bialer et al., 2013).
2) Rapamycin is an mTOR (mammalian target of rapamycin) inhibitor, that
predominantly exhibits modulatory activity on cerebral inflammation has
displayed anti-epileptic effects in WAG/Rij rats. In addition to antiepileptic
effects it also showed sustained anti-epileptogenic effect (Russo et al., 2013).
3) The GABA B receptor antagonists are also found to exhibit effective anti-
epileptic properties in animal models of absence seizures (Han et al., 2010).
4) Metabotrophic glutamate (mGlu) receptors, which are located at the
synapse of thalamocortical neurons and underlie development of absence
related SWDs are presently undergoing clinical evaluation after exhibiting
encouraging results in animal models of absence epilepsy (Ngomba et al.,
2011).
5) Two new compounds, Z941 and Z944 that specifically block T-type calcium
channels displayed significant and promising anti-absence activity in GAERS
(Tringham et al., 2012).
6.6 Final conclusion:
The findings in the current study provide electrophysiological insight into the
possible mechanisms underlying the seizure suppression effect of NPY in
GAERS. It also strengthens the argument that the S2 region might be focus

150
for origin of SWDs and NRT as the key target structure of thalamocortical
circuit by which NPY suppresses seizures. These findings are associative in
nature and further studies are required to ascertain if alterations in neuronal
firing patterns are induced by specific NPY receptor subtypes to understand
more about the seizure suppressive effects at cellular and network level. The
clinical relevance of this study is that it potentially uncovers the possibility of
ICV and focal drug delivery systems, administering NPY or other anti-
epileptic neuro-active agents via mechanical or viral infusion methods to
control seizures in patients with generalized epilepsies.

151
REFERENCES
Factors effecting epilpesy. (1989) Proposal for revised classification of epilepsies and epileptic syndromes. Commission
on Classification and Terminology of the International League Against Epilepsy. Epilepsia 30:389-399.
Adams BE, Kyi M, Reid CA, Myers DE, Xu S, Williams DA, O'Brien TJ (2011) Seizure-like thalamocortical rhythms initiate in the deep layers of the cortex in a co-culture model. Exp Neurol 227:203-209.
Adrian TE, Allen JM, Bloom SR, Ghatei MA, Rossor MN, Roberts GW, Crow TJ, Tatemoto K, Polak JM (1983) Neuropeptide Y distribution in human brain. Nature 306:584-586.
Aicher SA, Springston M, Berger SB, Reis DJ, Wahlestedt C (1991) Receptor-selective analogs demonstrate NPY/PYY receptor heterogeneity in rat brain. Neurosci Lett 130:32-36.
Allen YS, Adrian TE, Allen JM, Tatemoto K, Crow TJ, Bloom SR, Polak JM (1983) Neuropeptide Y distribution in the rat brain. Science 221:877-879.
Alloway KD, Johnson MJ, Wallace MB (1993) Thalamocortical interactions in the somatosensory system: interpretations of latency and cross-correlation analyses. Journal of neurophysiology 70:892-908.
Bacci A, Huguenard JR, Prince DA (2002) Differential modulation of synaptic transmission by neuropeptide Y in rat neocortical neurons. Proceedings of the National Academy of Sciences of the United States of America 99:17125-17130.
Banerjee PN, Filippi D, Allen Hauser W (2009) The descriptive epidemiology of epilepsy-a review. Epilepsy research 85:31-45.
Baraban SC (2002) Antiepileptic actions of neuropeptide Y in the mouse hippocampus require Y5 receptors. Epilepsia 43 Suppl 5:9-13.
Baraban SC, Hollopeter G, Erickson JC, Schwartzkroin PA, Palmiter RD (1997) Knock-out mice reveal a critical antiepileptic role for neuropeptide Y. J Neurosci 17:8927-8936.
Bard JA, Walker MW, Branchek TA, Weinshank RL (1995) Cloning and functional expression of a human Y4 subtype receptor for pancreatic polypeptide, neuropeptide Y, and peptide YY. The Journal of biological chemistry 270:26762-26765.
Barkovich AJ, Guerrini R, Kuzniecky RI, Jackson GD, Dobyns WB (2012) A developmental and genetic classification for malformations of cortical development: update 2012. Brain : a journal of neurology 135:1348-1369.
Ben-Ari Y, Gaiarsa JL, Tyzio R, Khazipov R (2007) GABA: A pioneer transmitter that excites immature neurons and generates primitive oscillations. Physiol Rev 87:1215-1284.
Benbadis SR BS (2006) Absence seizures. Philadelphia: Lippincott, Williams and Wilkins. Berg AT, Scheffer IE (2011) New concepts in classification of the epilepsies: entering the 21st
century. Epilepsia 52:1058-1062. Berglund MM, Hipskind PA, Gehlert DR (2003) Recent developments in our understanding of
the physiological role of PP-fold peptide receptor subtypes. Experimental Biology and Medicine 228:217-244.
Bialer M, Johannessen SI, Levy RH, Perucca E, Tomson T, White HS (2013) Progress report on new antiepileptic drugs: a summary of the Eleventh Eilat Conference (EILAT XI). Epilepsy Res 103:2-30.
Bijak M (1999) Neuropeptide Y suppresses epileptiform activity in rat frontal cortex and hippocampus in vitro via different NPY receptor subtypes. Neurosci Lett 268:115-118.
Blumenfeld H (2003) From molecules to networks: cortical/subcortical interactions in the pathophysiology of idiopathic generalized epilepsy. Epilepsia 44 Suppl 2:7-15.
Blumenfeld H, Rivera M, Vasquez JG, Shah A, Ismail D, Enev M, Zaveri HP (2007) Neocortical and thalamic spread of amygdala kindled seizures. Epilepsia 48:254-262.

152
Borowsky B, Walker MW, Bard J, Weinshank RL, Laz TM, Vaysse P, Branchek TA, Gerald C (1998) Molecular biology and pharmacology of multiple NPY Y5 receptor species homologs. Regul Pept 75-76:45-53.
Bourassa J, Deschenes M (1995) Corticothalamic projections from the primary visual cortex in rats: a single fiber study using biocytin as an anterograde tracer. Neuroscience 66:253-263.
Bourassa J, Pinault D, Deschenes M (1995) Corticothalamic projections from the cortical barrel field to the somatosensory thalamus in rats: a single-fibre study using biocytin as an anterograde tracer. The European journal of neuroscience 7:19-30.
Bower MR, Buckmaster PS (2008) Changes in granule cell firing rates precede locally recorded spontaneous seizures by minutes in an animal model of temporal lobe epilepsy. J Neurophysiol 99:2431-2442.
Bregola G, Dumont Y, Fournier A, Zucchini S, Quirion R, Simonato M (2000) Decreased levels of neuropeptide Y-5 receptor binding sites in two experimental models of epilepsy. Neuroscience 98:697-703.
Burkhoff A, Linemeyer DL, Salon JA (1998) Distribution of a novel hypothalamic neuropeptide Y receptor gene and it's absence in rat. Brain Res Mol Brain Res 53:311-316.
Buzsaki G, Smith A, Berger S, Fisher LJ, Gage FH (1990) Petit mal epilepsy and parkinsonian tremor: hypothesis of a common pacemaker. Neuroscience 36:1-14.
Caberlotto L FK, Hurd YL. (2000) Characterization of NPY mRNA-expressing cells in the human brain: co-localization with Y2 but not Y1 mRNA in the cerebral cortex, hippocampus, amygdala, and striatum. Journal of Chemical Neuroanatomy.
Caberlotto L, Fuxe K, Overstreet DH, Gerrard P, Hurd YL (1998a) Alterations in neuropeptide Y and Y1 receptor mRNA expression in brains from an animal model of depression: region specific adaptation after fluoxetine treatment. Brain Res Mol Brain Res 59:58-65.
Caberlotto L, Fuxe K, Rimland JM, Sedvall G, Hurd YL (1998b) Regional distribution of neuropeptide Y Y2 receptor messenger RNA in the human post mortem brain. Neuroscience 86:167-178.
Cabrele C, Langer M, Bader R, Wieland HA, Doods HN, Zerbe O, Beck-Sickinger AG (2000a) The first selective agonist for the neuropeptide YY5 receptor increases food intake in rats. Journal of Biological Chemistry 275:36043-36048.
Cabrele C, Langer M, Bader R, Wieland HA, Doods HN, Zerbe O, Beck-Sickinger AG (2000b) The first selective agonist for the neuropeptide YY5 receptor increases food intake in rats. J Biol Chem 275:36043-36048.
Chaffer CL, Morris MJ (2002) The feeding response to melanin-concentrating hormone is attenuated by antagonism of the NPYY1-receptor in the rat. Endocrinology 143:191-197.
Chemin J, Nargeot J, Lory P (2002) Neuronal T-type alpha 1H calcium channels induce neuritogenesis and expression of high-voltage-activated calcium channels in the NG108-15 cell line. J Neurosci 22:6856-6862.
Cherubini E, Gaiarsa JL, Ben-Ari Y (1991) GABA: an excitatory transmitter in early postnatal life. Trends Neurosci 14:515-519.
Chronwall BM, DiMaggio DA, Massari VJ, Pickel VM, Ruggiero DA, O'Donohue TL (1985) The anatomy of neuropeptide-Y-containing neurons in rat brain. Neuroscience 15:1159-1181.
Cockerell OC, Johnson AL, Sander JW, Shorvon SD (1997) Prognosis of epilepsy: a review and further analysis of the first nine years of the British National General Practice Study of Epilepsy, a prospective population-based study. Epilepsia 38:31-46.
Coenen AM, Drinkenburg WH, Inoue M, van Luijtelaar EL (1992a) Genetic models of absence epilepsy, with emphasis on the WAG/Rij strain of rats. Epilepsy Res 12:75-86.

153
Coenen AML, Drinkenburg WHIM, Inoue M, Vanluijtelaar ELJM (1992b) Genetic Models of Absence Epilepsy, with Emphasis on the Wag Rij Strain of Rats. Epilepsy Research 12:75-86.
Colmers WF, Bleakman D (1994) Effects of neuropeptide Y on the electrical properties of neurons. Trends Neurosci 17:373-379.
Colmers WF, Klapstein GJ, Fournier A, St-Pierre S, Treherne KA (1991) Presynaptic inhibition by neuropeptide Y in rat hippocampal slice in vitro is mediated by a Y2 receptor. Br J Pharmacol 102:41-44.
Contreras D, Curro Dossi R, Steriade M (1993) Electrophysiological properties of cat reticular thalamic neurones in vivo. The Journal of physiology 470:273-294.
Contreras D, Steriade M (1995) Cellular basis of EEG slow rhythms: a study of dynamic corticothalamic relationships. J Neurosci 15:604-622.
Corder R, Emson PC, Lowry PJ (1984) Purification and characterization of human neuropeptide Y from adrenal-medullary phaeochromocytoma tissue. Biochem J 219:699-706.
Cox CL, Huguenard JR, Prince DA (1996) Heterogeneous axonal arborizations of rat thalamic reticular neurons in the ventrobasal nucleus. J Comp Neurol 366:416-430.
Cruz ME, Schoenberg BS, Ruales J, Barberis P, Proano J, Bossano F, Sevilla F, Bolis CL (1985) Pilot study to detect neurologic disease in Ecuador among a population with a high prevalence of endemic goiter. Neuroepidemiology 4:108-116.
Curtis DR NW (1964) Physical Techniques in Biological Research. New York: Academic Press V:144-192.
Cymerblit-Sabba A, Schiller Y (2010) Network dynamics during development of pharmacologically induced epileptic seizures in rats in vivo. J Neurosci 30:1619-1630.
D.R. C (1964) Microiontophoresis. In : Physical techniques of biological research. New york- London.
Danober L, Deransart C, Depaulis A, Vergnes M, Marescaux C (1998) Pathophysiological mechanisms of genetic absence epilepsy in the rat. Prog Neurobiol 55:27-57.
de Quidt ME, Emson PC (1986) Distribution of neuropeptide Y-like immunoreactivity in the rat central nervous system--I. Radioimmunoassay and chromatographic characterisation. Neuroscience 18:527-543.
DePrato Primeaux S, Holmes PV, Martin RJ, Dean RG, Edwards GL (2000) Experimentally induced attenuation of neuropeptide-Y gene expression in transgenic mice increases mortality rate following seizures. Neurosci Lett 287:61-64.
Ding D, Wang W, Wu J, Ma G, Dai X, Yang B, Wang T, Yuan C, Hong Z, de Boer HM, Prilipko L, Sander JW (2006) Premature mortality in people with epilepsy in rural China: a prospective study. Lancet Neurol 5:823-827.
Dumont Y, Fournier A, Quirion R (1998) Expression and characterization of the neuropeptide Y Y5 receptor subtype in the rat brain. J Neurosci 18:5565-5574.
Dumont Y, Fournier A, St-Pierre S, Quirion R (1993) Comparative characterization and autoradiographic distribution of neuropeptide Y receptor subtypes in the rat brain. J Neurosci 13:73-86.
El Bahh B, Balosso S, Hamilton T, Herzog H, Beck-Sickinger AG, Sperk G, Gehlert DR, Vezzani A, Colmers WF (2005) The anti-epileptic actions of neuropeptide Y in the hippocampus are mediated by Y and not Y receptors. Eur J Neurosci 22:1417-1430.
El Bahh B, Cao JQ, Beck-Sickinger AG, Colmers WF (2002) Blockade of neuropeptide Y(2) receptors and suppression of NPY's anti-epileptic actions in the rat hippocampal slice by BIIE0246. British journal of pharmacology 136:502-509.
Engel J, Jr. (2011) The etiologic classification of epilepsy. Epilepsia 52:1195-1197; discussion 1205-1199.
Erickson JC, Clegg KE, Palmiter RD (1996) Sensitivity to leptin and susceptibility to seizures of mice lacking neuropeptide Y. Nature 381:415-421.

154
Fariello RG, Golden GT (1987) The THIP-induced model of bilateral synchronous spike and wave in rodents. Neuropharmacology 26:161-165.
Ferrier L, Segain JP, Bonnet C, Cherbut C, Lehur PA, Jarry A, Galmiche JP, Blottiere HM (2002) Functional mapping of NPY/PYY receptors in rat and human gastro-intestinal tract. Peptides 23:1765-1771.
Fisher RS, Prince DA (1977) Spike-wave rhythms in cat cortex induced by parenteral penicillin. I. Electroencephalographic features. Electroencephalography and clinical neurophysiology 42:608-624.
Fonnum F, Storm-Mathisen J, Divac I (1981a) Biochemical evidence for glutamate as neurotransmitter in corticostriatal and corticothalamic fibres in rat brain. Neuroscience 6:863-873.
Fonnum F, Stormmathisen J, Divac I (1981b) Biochemical-Evidence for Glutamate as Neurotransmitter in Corticostriatal and Corticothalamic Fibers in Rat-Brain. Neuroscience 6:863-873.
Furtinger S, Pirker S, Czech T, Baumgartner C, Ransmayr G, Sperk G (2001) Plasticity of Y1 and Y2 receptors and neuropeptide Y fibers in patients with temporal lobe epilepsy. J Neurosci 21:5804-5812.
Gandrathi A, Zheng T, O'Brien P, Ali I, O'Brien TJ, French CR (2013) An in vivo technique for investigating electrophysiological effects of centrally administered drugs on single neurons and network behaviour. J Neurosci Methods 219:197-204.
Gariboldi M, Conti M, Cavaleri D, Samanin R, Vezzani A (1998) Anticonvulsant properties of BIBP3226, a non-peptide selective antagonist at neuropeptide Y Y-1 receptors. European Journal of Neuroscience 10:757-759.
Gehlert DR, Beavers LS, Johnson D, Gackenheimer SL, Schober DA, Gadski RA (1996) Expression cloning of a human brain neuropeptide Y Y2 receptor. Mol Pharmacol 49:224-228.
Gerald C, Walker MW, Criscione L, Gustafson EL, Batzl-Hartmann C, Smith KE, Vaysse P, Durkin MM, Laz TM, Linemeyer DL, Schaffhauser AO, Whitebread S, Hofbauer KG, Taber RI, Branchek TA, Weinshank RL (1996) A receptor subtype involved in neuropeptide-Y-induced food intake. Nature 382:168-171.
Gerald C, Walker MW, Vaysse PJ, He C, Branchek TA, Weinshank RL (1995) Expression cloning and pharmacological characterization of a human hippocampal neuropeptide Y/peptide YY Y2 receptor subtype. The Journal of biological chemistry 270:26758-26761.
Gibbs FA DH, Lennox WG (1935) The EEG in epilepsy and in conditon of impaired consciousness. 1134-1148.
Gloor P AM, Kostopoulos G (1990) Thalamocortical Relationships in Generalized Epilepsy with Bilaterally Synchronous Spike-and-Wave Discharge. In: Neurobiological Approaches, vol. III, pp 119-212.
Goumain M, Voisin T, Lorinet AM, Laburthe M (1998) Identification and distribution of mRNA encoding the Y1, Y2, Y4, and Y5 receptors for peptides of the PP-fold family in the rat intestine and colon. Biochem Biophys Res Commun 247:52-56.
Greber S, Schwarzer C, Sperk G (1994a) Neuropeptide Y inhibits potassium-stimulated glutamate release through Y2 receptors in rat hippocampal slices in vitro. British journal of pharmacology 113:737-740.
Greber S, Schwarzer C, Sperk G (1994b) Neuropeptide-Y Inhibits Potassium-Stimulated Glutamate Release through Y-2 Receptors in Rat Hippocampal Slices in-Vitro. British journal of pharmacology 113:737-740.
Gruber B, Greber S, Rupp E, Sperk G (1994) Differential NPY mRNA expression in granule cells and interneurons of the rat dentate gyrus after kainic acid injection. Hippocampus 4:474-482.
Haerer AF, Anderson DW, Schoenberg BS (1986) Prevalence and clinical features of epilepsy in a biracial United States population. Epilepsia 27:66-75.

155
Haji A, Takeda R (1993) Microiontophoresis of baclofen on membrane potential and input resistance in bulbar respiratory neurons in the cat. Brain Res 622:294-298.
Hamiwka LD, Wirrell EC (2009) Comorbidities in Pediatric Epilepsy: Beyond "Just" Treating the Seizures. Journal of child neurology 24:734-742.
Han HA, Cortez MA, Snead OC (2010) GABA(B) receptors in absence epilepsy. Epilepsia 51:24-24.
Hauser WA (1997) Sudden unexpected death in patients with epilepsy: issues for further study. Epilepsia 38:S26-29.
Heilig M, McLeod S, Brot M, Heinrichs SC, Menzaghi F, Koob GF, Britton KT (1993) Anxiolytic-like action of neuropeptide Y: mediation by Y1 receptors in amygdala, and dissociation from food intake effects. Neuropsychopharmacology 8:357-363.
Herzog H, Hort YJ, Ball HJ, Hayes G, Shine J, Selbie LA (1992) Cloned human neuropeptide Y receptor couples to two different second messenger systems. Proceedings of the National Academy of Sciences of the United States of America 89:5794-5798.
Herzog H, Hort YJ, Shine J, Selbie LA (1993) Molecular cloning, characterization, and localization of the human homolog to the reported bovine NPY Y3 receptor: lack of NPY binding and activation. DNA Cell Biol 12:465-471.
Hicks TP, Albus K, Kaneko T, Baumfalk U (1993) Examination of the effects of cholecystokinin 26-33 and neuropeptide Y on responses of visual cortical neurons of the cat. Neuroscience 52:263-279.
Hitiris N, Mohanraj R, Norrie J, Brodie MJ (2007) Mortality in epilepsy. Epilepsy Behav 10:363-376.
Hokfelt T, Broberger C, Zhang X, Diez M, Kopp J, Xu ZQ, Landry M, Bao L, Schalling M, Koistinaho J, DeArmond SJ, Prusiner S, Gong J, Walsh JH (1998) Neuropeptide Y: some viewpoints on a multifaceted peptide in the normal and diseased nervous system. Brain Research Reviews 26:154-166.
Hokfelt T, Lundberg JM, Lagercrantz H, Tatemoto K, Mutt V, Lindberg J, Terenius L, Everitt BJ, Fuxe K, Agnati L, Goldstein M (1983) Occurrence of neuropeptide Y (NPY)-like immunoreactivity in catecholamine neurons in the human medulla oblongata. Neurosci Lett 36:217-222.
Houser CR, Vaughn JE, Barber RP, Roberts E (1980) GABA neurons are the major cell type of the nucleus reticularis thalami. Brain research 200:341-354.
Huntsman MM, Porcello DM, Homanics GE, DeLorey TM, Huguenard JR (1999) Reciprocal inhibitory connections and network synchrony in the mammalian thalamus. Science 283:541-543.
Husum H, Gruber SH, Bolwig TG, Mathe AA (2002) Extracellular levels of NPY in the dorsal hippocampus of freely moving rats are markedly elevated following a single electroconvulsive stimulation, irrespective of anticonvulsive Y1 receptor blockade. Neuropeptides 36:363-369.
Inoue M, Ates N, Vossen JM, Coenen AM (1994) Effects of the neuroleptanalgesic fentanyl-fluanisone (Hypnorm) on spike-wave discharges in epileptic rats. Pharmacol Biochem Behav 48:547-551.
Inui A (1999) Neuropeptide Y feeding receptors: are multiple subtypes involved? Trends in Pharmacological Sciences 20:43-46.
Jasper HH D-FJ (1947) Experimental studies on the functional anatomy of petit mal epilepsy. 272-298.
Jazin EE, Yoo HY, Blomqvist AG, Yee F, Weng GH, Walker MW, Salon J, Larhammar D, Wahlestedt C (1993) A Proposed Bovine Neuropeptide-Y (Npy) Receptor Cdna Clone, or Its Human Homolog, Confers Neither Npy Binding-Sites nor Npy Responsiveness on Transfected Cells. Regulatory Peptides 47:247-258.
Jinde S, Masui A, Morinobu S, Noda A, Kato N (2002) Differential changes in messenger RNA expressions and binding sites of neuropeptide Y Y1, Y2 and Y5 receptors in the

156
hippocampus of an epileptic mutant rat: Noda epileptic rat. Neuroscience 115:1035-1045.
Jones EG (1985) The Thalamus. Jonsson-Rylander AC, Nordlander M, Svindland A, Ilebekk A (2003) Distribution of
neuropeptide Y Y1 and Y2 receptors in the postmortem human heart. Peptides 24:255-262.
Kaneko T, Mizuno N (1988) Immunohistochemical Study of Glutaminase-Containing Neurons in the Cerebral-Cortex and Thalamus of the Rat. Journal of Comparative Neurology 267:590-602.
Kaneoke Y, Vitek JL (1996) Burst and oscillation as disparate neuronal properties. J Neurosci Methods 68:211-223.
Kask A, Harro J, von Horsten S, Redrobe JP, Dumont Y, Quirion R (2002) The neurocircuitry and receptor subtypes mediating anxiolytic-like effects of neuropeptide Y. Neuroscience and biobehavioral reviews 26:259-283.
Kim U, Bal T, McCormick DA (1995) Spindle waves are propagating synchronized oscillations in the ferret LGNd in vitro. J Neurophysiol 74:1301-1323.
Kim U, Sanchez-Vives MV, McCormick DA (1997) Functional dynamics of GABAergic inhibition in the thalamus. Science 278:130-134.
Klapstein GJ, Colmers WF (1997) Neuropeptide Y suppresses epileptiform activity in rat hippocampus in vitro. J Neurophysiol 78:1651-1661.
Kofler N, Kirchmair E, Schwarzer C, Sperk G (1997) Altered expression of NPY-Y1 receptors in kainic acid induced epilepsy in rats. Neurosci Lett 230:129-132.
Kopp J, Nanobashvili A, Kokaia Z, Lindvall O, Hokfelt T (1999) Differential regulation of mRNAs for neuropeptide Y and its receptor subtypes in widespread areas of the rat limbic system during kindling epileptogenesis. Brain Res Mol Brain Res 72:17-29.
Kostopoulos GK (2001) Involvement of the thalamocortical system in epileptic loss of consciousness. Epilepsia 42 Suppl 3:13-19.
Kramer U, Nevo Y, Neufeld MY, Fatal A, Leitner Y, Harel S (1998) Epidemiology of epilepsy in childhood: a cohort of 440 consecutive patients. Pediatr Neurol 18:46-50.
Krause J, Eva C, Seeburg PH, Sprengel R (1992) Neuropeptide Y1 subtype pharmacology of a recombinantly expressed neuropeptide receptor. Mol Pharmacol 41:817-821.
Kuruba R, Hattiangady B, Parihar VK, Shuai B, Shetty AK (2011) Differential susceptibility of interneurons expressing neuropeptide Y or parvalbumin in the aged hippocampus to acute seizure activity. PLoS One 6:e24493.
Kwan P, Brodie MJ (2000) Early identification of refractory epilepsy. The New England journal of medicine 342:314-319.
Larhammar D, Blomqvist AG, Soderberg C (1993) Evolution of neuropeptide Y and its related peptides. Comp Biochem Physiol C 106:743-752.
Larhammar D, Blomqvist AG, Yee F, Jazin E, Yoo H, Wahlested C (1992) Cloning and functional expression of a human neuropeptide Y/peptide YY receptor of the Y1 type. The Journal of biological chemistry 267:10935-10938.
Lerman P (1986) Seizures induced or aggravated by anticonvulsants. Epilepsia 27:706-710. Li K, Xu E (2008) The role and the mechanism of gamma-aminobutyric acid during central
nervous system development. Neurosci Bull 24:195-200. Lin LC, Lee WT, Chen IJ, Yang RC (2010) Lower plasma neuropeptide Y level in patients with
atypical febrile convulsions. Kaohsiung J Med Sci 26:8-12. Liu L, Zheng T, Morris MJ, Wallengren C, Clarke AL, Reid CA, Petrou S, O'Brien TJ (2006) The
mechanism of carbamazepine aggravation of absence seizures. J Pharmacol Exp Ther 319:790-798.
Liu Z, Vergnes M, Depaulis A, Marescaux C (1991) Evidence for a critical role of GABAergic transmission within the thalamus in the genesis and control of absence seizures in the rat. Brain Res 545:1-7.

157
Llinas R, Ribary U, Contreras D, Pedroarena C (1998) The neuronal basis for consciousness. Philos Trans R Soc Lond B Biol Sci 353:1841-1849.
Loiseau P, Duche B, Pedespan JM (1995) Absence epilepsies. Epilepsia 36:1182-1186. Loiseau P, Jallon P (1995) [Drug-resistant epilepsy in adults]. Rev Neurol (Paris) 151:295-
306. Lopes da Silva FH, Blanes W, Kalitzin SN, Parra J, Suffczynski P, Velis DN (2003) Dynamical
diseases of brain systems: different routes to epileptic seizures. IEEE Trans Biomed Eng 50:540-548.
Lundberg JM, Terenius L, Hokfelt T, Martling CR, Tatemoto K, Mutt V, Polak J, Bloom S, Goldstein M (1982) Neuropeptide Y (NPY)-like immunoreactivity in peripheral noradrenergic neurons and effects of NPY on sympathetic function. Acta Physiologica Scandinavica 116:477-480.
Lundell I, Blomqvist AG, Berglund MM, Schober DA, Johnson D, Statnick MA, Gadski RA, Gehlert DR, Larhammar D (1995) Cloning of a human receptor of the NPY receptor family with high affinity for pancreatic polypeptide and peptide YY. The Journal of biological chemistry 270:29123-29128.
Lupien SJ, McEwen BS, Gunnar MR, Heim C (2009) Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat Rev Neurosci 10:434-445.
Luttjohann A, Zhang S, de Peijper R, van Luijtelaar G (2011) Electrical stimulation of the epileptic focus in absence epileptic WAG/Rij rats: assessment of local and network excitability. Neuroscience 188:125-134.
MacDonald B (2001) The prognosis of epilepsy. Seizure 10:347-358. Marescaux C, Micheletti G, Vergnes M, Depaulis A, Rumbach L, Warter JM (1984) A model of
chronic spontaneous petit mal-like seizures in the rat: comparison with pentylenetetrazol-induced seizures. Epilepsia 25:326-331.
Marescaux C, Vergnes M (1995) Genetic Absence Epilepsy in Rats from Strasbourg (GAERS). Italian journal of neurological sciences 16:113-118.
Marksteiner J, Sperk G, Maas D (1989) Differential increases in brain levels of neuropeptide Y and vasoactive intestinal polypeptide after kainic acid-induced seizures in the rat. Naunyn-Schmiedeberg's archives of pharmacology 339:173-177.
Marsh DJ, Baraban SC, Hollopeter G, Palmiter RD (1999) Role of the Y5 neuropeptide Y receptor in limbic seizures. Proceedings of the National Academy of Sciences of the United States of America 96:13518-13523.
Marsh DJ, Hollopeter G, Kafer KE, Palmiter RD (1998) Role of the Y5 neuropeptide Y receptor in feeding and obesity. Nat Med 4:718-721.
Matsumoto M, Nomura T, Momose K, Ikeda Y, Kondou Y, Akiho H, Togami J, Kimura Y, Okada M, Yamaguchi T (1996) Inactivation of a novel neuropeptide Y/peptide YY receptor gene in primate species. The Journal of biological chemistry 271:27217-27220.
Mattson RH, Cramer JA, Collins JF, Smith DB, Delgado-Escueta AV, Browne TR, Williamson PD, Treiman DM, McNamara JO, McCutchen CB, et al. (1985) Comparison of carbamazepine, phenobarbital, phenytoin, and primidone in partial and secondarily generalized tonic-clonic seizures. The New England journal of medicine 313:145-151.
McCormick DA, Contreras D (2001) On the cellular and network bases of epileptic seizures. Annu Rev Physiol 63:815-846.
Mcdonald JK (1988) Npy and Related Substances. Critical Reviews in Neurobiology 4:97-135. McQuiston AR, Colmers WF (1996) Neuropeptide Y2 receptors inhibit the frequency of
spontaneous but not miniature EPSCs in CA3 pyramidal cells of rat hippocampus. J Neurophysiol 76:3159-3168.
Meeren HK, Pijn JP, Van Luijtelaar EL, Coenen AM, Lopes da Silva FH (2002) Cortical focus drives widespread corticothalamic networks during spontaneous absence seizures in rats. J Neurosci 22:1480-1495.
Meldrum BS (1989) GABAergic mechanisms in the pathogenesis and treatment of epilepsy. Br J Clin Pharmacol 27 Suppl 1:3S-11S.

158
Michel MC, Beck-Sickinger A, Cox H, Doods HN, Herzog H, Larhammar D, Quirion R, Schwartz T, Westfall T (1998) XVI. International Union of Pharmacology recommendations for the nomenclature of neuropeptide Y, peptide YY, and pancreatic polypeptide receptors. Pharmacol Rev 50:143-150.
Micheletti G, Vergnes M, Marescaux C, Reis J, Depaulis A, Rumbach L, Warter JM (1985) Antiepileptic drug evaluation in a new animal model: spontaneous petit mal epilepsy in the rat. Arzneimittel-Forschung 35:483-485.
Ming OY, Shao W (2000) Dexamethasone attenuates the depressor response induced by neuropeptide Y microinjected into the nucleus tractus solitarius in rats. Brit J Pharmacol 129:865-870.
Morris BJ (1989) Neuronal localisation of neuropeptide Y gene expression in rat brain. J Comp Neurol 290:358-368.
Morris MJ, Gannan E, Stroud LM, Beck-Sickinger AG, O'Brien TJ (2007) Neuropeptide Y suppresses absence seizures in a genetic rat model primarily through effects on Y receptors. Eur J Neurosci 25:1136-1143.
Munglani R, Hudspith MJ, Hunt SP (1996) The therapeutic potential of neuropeptide Y. Analgesic, anxiolytic and antihypertensive. Drugs 52:371-389.
Murphy K, Delanty N (2000) Primary Generalized Epilepsies. Curr Treat Options Neurol 2:527-542.
Nakajima M, Inui A, Asakawa A, Momose K, Ueno N, Teranishi A, Baba S, Kasuga M (1998) Neuropeptide Y produces anxiety via Y2-type receptors. Peptides 19:359-363.
Nashef L, Hindocha N, Makoff A (2007) Risk factors in sudden death in epilepsy (SUDEP): the quest for mechanisms. Epilepsia 48:859-871.
Naveilhan P, Hassani H, Canals JM, Ekstrand AJ, Larefalk A, Chhajlani V, Arenas E, Gedda K, Svensson L, Thoren P, Ernfors P (1999) Normal feeding behavior, body weight and leptin response require the neuropeptide Y Y2 receptor. Nat Med 5:1188-1193.
Ngomba RT, Santolini I, Salt TE, Ferraguti F, Battaglia G, Nicoletti F, van Luijtelaar G (2011) Metabotropic glutamate receptors in the thalamocortical network: strategic targets for the treatment of absence epilepsy. Epilepsia 52:1211-1222.
Noebels JL, Avoli M, Rogawski M, Olsen R, Delgado-Escueta AV (2010) "Jasper's Basic Mechanisms of the Epilepsies" Workshop. Epilepsia 51 Suppl 5:1-5.
Noronha AL, Borges MA, Marques LH, Zanetta DM, Fernandes PT, de Boer H, Espindola J, Miranda CT, Prilipko L, Bell GS, Sander JW, Li LM (2007) Prevalence and pattern of epilepsy treatment in different socioeconomic classes in Brazil. Epilepsia 48:880-885.
Ohara PT, Lieberman AR (1985) The thalamic reticular nucleus of the adult rat: experimental anatomical studies. J Neurocytol 14:365-411.
Olafsson E, Ludvigsson P, Gudmundsson G, Hesdorffer D, Kjartansson O, Hauser WA (2005) Incidence of unprovoked seizures and epilepsy in Iceland and assessment of the epilepsy syndrome classification: a prospective study. Lancet Neurol 4:627-634.
Olumide A.e · Oyediran A.B.O.d · Pearson C.A.d · Bolis C.L.f OBOaSBSbNVAcAAaKOdAAdBOa (1982) Research Protocol for Measuring the Prevalence of Neurologic Disorders in Developing Countries
Results of a Pilot Study in Nigeria. Neuroepidemiology 1:143-153. Ouyang M, Wang S (2000) Dexamethasone attenuates the depressor response induced by
neuropeptide Y microinjected into the nucleus tractus solitarius in rats. Br J Pharmacol 129:865-870.
P L (1985) Childhood absence epilepsy. 106-120. Panayiotopoulos CP (2001) Treatment of typical absence seizures and related epileptic
syndromes. Paediatr Drugs 3:379-403. Panayitipoulos C (2005a) Typical absence seizures. Parker RM, Herzog H (1999) Regional distribution of Y-receptor subtype mRNAs in rat brain.
Eur J Neurosci 11:1431-1448.

159
Patrylo PR, van den Pol AN, Spencer DD, Williamson A (1999) NPY inhibits glutamatergic excitation in the epileptic human dentate gyrus. J Neurophysiol 82:478-483.
Paxinos G, Watson CR, Emson PC (1980) AChE-stained horizontal sections of the rat brain in stereotaxic coordinates. J Neurosci Methods 3:129-149.
Paxinos G WC (1998) The rat brain in stereotaxic coordinates. San Diego: CA: Academic press. Penry JK, Porter RJ, Dreifuss RE (1975) Simultaneous recording of absence seizures with
video tape and electroencephalography. A study of 374 seizures in 48 patients. Brain : a journal of neurology 98:427-440.
Pinault D (1996) A novel single-cell staining procedure performed in vivo under electrophysiological control: morpho-functional features of juxtacellularly labeled thalamic cells and other central neurons with biocytin or Neurobiotin. J Neurosci Methods 65:113-136.
Pinault D (2003) Cellular interactions in the rat somatosensory thalamocortical system during normal and epileptic 5-9 Hz oscillations. The Journal of physiology 552:881-905.
Pinault D (2004) The thalamic reticular nucleus: structure, function and concept. Brain Res Brain Res Rev 46:1-31.
Pinault D (2005) A new stabilizing craniotomy-duratomy technique for single-cell anatomo-electrophysiological exploration of living intact brain networks. J Neurosci Methods 141:231-242.
Pinault D, Bourassa J, Deschenes M (1995) The axonal arborization of single thalamic reticular neurons in the somatosensory thalamus of the rat. Eur J Neurosci 7:31-40.
Pinault D, Deschenes M (1998) Anatomical evidence for a mechanism of lateral inhibition in the rat thalamus. Eur J Neurosci 10:3462-3469.
Pinault D, O'Brien TJ (2005) Cellular and network mechanisms of genetically-determined absence seizures. Thalamus Relat Syst 3:181-203.
Pinault D, Slezia A, Acsady L (2006) Corticothalamic 5-9 Hz oscillations are more pro-epileptogenic than sleep spindles in rats. The Journal of physiology 574:209-227.
Pinault D, Vergnes M, Marescaux C (2001) Medium-voltage 5-9-Hz oscillations give rise to spike-and-wave discharges in a genetic model of absence epilepsy: in vivo dual extracellular recording of thalamic relay and reticular neurons. Neuroscience 105:181-201.
Porter RJ (1993) The absence epilepsies. Epilepsia 34 Suppl 3:S42-48. Press WH, Teukolsky SA, Vetterling WT, Flannery BP (1992) numerical recipies in C: The art
of scientific computing: cambridge university press. Reibel S, Benmaamar R, Le BT, Larmet Y, Kalra SP, Marescaux C, Depaulis A (2003)
Neuropeptide Y delays hippocampal kindling in the rat. Hippocampus 13:557-560. Reibel S, Nadi S, Benmaamar R, Larmet Y, Carnahan J, Marescaux C, Depaulis A (2001)
Neuropeptide Y and epilepsy: varying effects according to seizure type and receptor activation. Peptides 22:529-539.
Richichi C, Lin EJ, Stefanin D, Colella D, Ravizza T, Grignaschi G, Veglianese P, Sperk G, During MJ, Vezzani A (2004) Anticonvulsant and antiepileptogenic effects mediated by adeno-associated virus vector neuropeptide Y expression in the rat hippocampus. J Neurosci 24:3051-3059.
Rimland J, Xin W, Sweetnam P, Saijoh K, Nestler EJ, Duman RS (1991) Sequence and expression of a neuropeptide Y receptor cDNA. Mol Pharmacol 40:869-875.
Rimland JM SE, Humbert Y, Ratti E, Trist DG, North RA. (1996) Coexpression with potassium channel subunits used to clone the Y2 receptor for neuropeptide Y. Molecular Pharmacology.
Rocca WA, Savettieri G, Anderson DW, Meneghini F, Grigoletto F, Morgante L, Reggio A, Salemi G, Patti F, Di Perri R, Sicilian Neuro-Epidemiologic Study G (2001) Door-to-door prevalence survey of epilepsy in three Sicilian municipalities. Neuroepidemiology 20:237-241.

160
Rose PM, Fernandes P, Lynch JS, Frazier ST, Fisher SM, Kodukula K, Kienzle B, Seethala R (1995) Cloning and functional expression of a cDNA encoding a human type 2 neuropeptide Y receptor. The Journal of biological chemistry 270:29038.
Russo E, Citraro R, Donato G, Camastra C, Iuliano R, Cuzzocrea S, Constanti A, De Sarro G (2013) mTOR inhibition modulates epileptogenesis, seizures and depressive behavior in a genetic rat model of absence epilepsy. Neuropharmacology 69:25-36.
Sander JW (1993) Some aspects of prognosis in the epilepsies: a review. Epilepsia 34:1007-1016.
Sander JW (2003) The epidemiology of epilepsy revisited. Curr Opin Neurol 16:165-170. Schwarzer C, Kofler N, Sperk G (1998) Up-regulation of neuropeptide Y-Y2 receptors in an
animal model of temporal lobe epilepsy. Mol Pharmacol 53:6-13. Shorvon SD (2011) The etiologic classification of epilepsy. Epilepsia 52:1052-1057. Silva AP, Carvalho AP, Carvalho CM, Malva JO (2001) Modulation of intracellular calcium
changes and glutamate release by neuropeptide Y1 and Y2 receptors in the rat hippocampus: differential effects in CA1, CA3 and dentate gyrus. Journal of Neurochemistry 79:286-296.
Singh P, Maibach HI (1994) Iontophoresis in drug delivery: basic principles and applications. Crit Rev Ther Drug Carrier Syst 11:161-213.
Slaght SJ, Leresche N, Deniau JM, Crunelli V, Charpier S (2002) Activity of thalamic reticular neurons during spontaneous genetically determined spike and wave discharges. J Neurosci 22:2323-2334.
Snead OC, 3rd (1995) Basic mechanisms of generalized absence seizures. Annals of neurology 37:146-157.
Snead OC, 3rd, Depaulis A, Vergnes M, Marescaux C (1999) Absence epilepsy: advances in experimental animal models. Adv Neurol 79:253-278.
Snead OC, 3rd, Hosey LC (1985) Exacerbation of seizures in children by carbamazepine. The New England journal of medicine 313:916-921.
Sperk G, Herzog H (1997) Anticonvulsant action of neuropeptide Y. Neuropeptide Y may act as an endogenous anticonvulsant through Y5 receptors suggesting a new target for antiepileptic drugs. Nat Med 3:728-729.
Spreafico R, Battaglia G, Frassoni C (1991) The reticular thalamic nucleus (RTN) of the rat: cytoarchitectural, Golgi, immunocytochemical, and horseradish peroxidase study. J Comp Neurol 304:478-490.
Statnick MA, Schober DA, Gackenheimer S, Johnson D, Beavers L, Mayne NG, Burnett JP, Gadski R, Gehlert DR (1998) Characterization of the neuropeptide Y5 receptor in the human hypothalamus: a lack of correlation between Y5 mRNA levels and binding sites. Brain research 810:16-26.
Steriade M, Deschenes M (1984a) The thalamus as a neuronal oscillator. Brain research 320:1-63.
Steriade M, Deschenes M (1984b) The Thalamus as a Neuronal Oscillator. Brain Res Rev 8:1-63.
Steriade M, Domich L, Oakson G (1986) Reticularis thalami neurons revisited: activity changes during shifts in states of vigilance. J Neurosci 6:68-81.
Steriade M, McCormick DA, Sejnowski TJ (1993) Thalamocortical oscillations in the sleeping and aroused brain. Science 262:679-685.
Stroud LM, O'Brien TJ, Jupp B, Wallengren C, Morris MJ (2005) Neuropeptide Y suppresses absence seizures in a genetic rat model. Brain research 1033:151-156.
Sun QQ, Akk G, Huguenard JR, Prince DA (2001a) Differential regulation of GABA release and neuronal excitability mediated by neuropeptide Y1 and Y2 receptors in rat thalamic neurons. J Physiol 531:81-94.
Sun QQ, Baraban SC, Prince DA, Huguenard JR (2003) Target-specific neuropeptide Y-ergic synaptic inhibition and its network consequences within the mammalian thalamus. J Neurosci 23:9639-9649.

161
Sun QQ, Huguenard JR, Prince DA (2001b) Neuropeptide Y receptors differentially modulate G-protein-activated inwardly rectifying K+ channels and high-voltage-activated Ca2+ channels in rat thalamic neurons. J Physiol 531:67-79.
Sundler F, Moghimzadeh E, Hakanson R, Ekelund M, Emson P (1983) Nerve fibers in the gut and pancreas of the rat displaying neuropeptide-Y immunoreactivity. Intrinsic and extrinsic origin. Cell and tissue research 230:487-493.
Takahashi M, Hayashi S, Kakita A, Wakabayashi K, Fukuda M, Kameyama S, Tanaka R, Takahashi H, Nawa H (1999) Patients with temporal lobe epilepsy show an increase in brain-derived neurotrophic factor protein and its correlation with neuropeptide Y. Brain research 818:579-582.
Tatemoto K (1982a) Isolation and Characterization of Peptide Yy (Pyy), a Candidate Gut Hormone That Inhibits Pancreatic Exocrine Secretion. Proceedings of the National Academy of Sciences of the United States of America-Biological Sciences 79:2514-2518.
Tatemoto K (1982b) Neuropeptide Y: complete amino acid sequence of the brain peptide. Proceedings of the National Academy of Sciences of the United States of America 79:5485-5489.
Tatemoto K (1982c) Neuropeptide-Y - Complete Amino-Acid-Sequence of the Brain Peptide. Proceedings of the National Academy of Sciences of the United States of America-Biological Sciences 79:5485-5489.
Tatemoto K, Carlquist M, Mutt V (1982) Neuropeptide-Y - a Novel Brain Peptide with Structural Similarities to Peptide-Yy and Pancreatic-Polypeptide. Nature 296:659-660.
Tatemoto K, Mutt V (1978) Chemical determination of polypeptide hormones. Proceedings of the National Academy of Sciences of the United States of America 75:4115-4119.
Temkin O (1994) The Falling Sickness: A History of Epilepsy from the Greeks to the Beginnings of Modern Neurology. The New England journal of medicine.
Thorsell A, Michalkiewicz M, Dumont Y, Quirion R, Caberlotto L, Rimondini R, Mathe AA, Heilig M (2000) Behavioral insensitivity to restraint stress, absent fear suppression of behavior and impaired spatial learning in transgenic rats with hippocampal neuropeptide Y overexpression. Proceedings of the National Academy of Sciences of the United States of America 97:12852-12857.
Treiman DM (2001) GABAergic mechanisms in epilepsy. Epilepsia 42 Suppl 3:8-12. Tringham E, Powell KL, Cain SM, Kuplast K, Mezeyova J, Weerapura M, Eduljee C, Jiang XP,
Smith P, Morrison JL, Jones NC, Braine E, Rind G, Fee-Maki M, Parker D, Pajouhesh H, Parmar M, O'Brien TJ, Snutch TP (2012) T-Type Calcium Channel Blockers That Attenuate Thalamic Burst Firing and Suppress Absence Seizures. Sci Transl Med 4.
van Raay L, Jovanovska V, Morris MJ, O'Brien TJ (2012) Focal administration of neuropeptide Y into the S2 somatosensory cortex maximally suppresses absence seizures in a genetic rat model. Epilepsia 53:477-484.
Vendrame M, Khurana DS, Cruz M, Melvin J, Valencia I, Legido A, Kothare SV (2007) Aggravation of seizures and/or EEG features in children treated with oxcarbazepine monotherapy. Epilepsia 48:2116-2120.
Vergnes M, Marescaux C (1992) Cortical and thalamic lesions in rats with genetic absence epilepsy. J Neural Transm Suppl 35:71-83.
Vergnes M, Marescaux C, Boehrer A, Depaulis A (1991) Are rats with genetic absence epilepsy behaviorally impaired? Epilepsy research 9:97-104.
Vergnes M, Marescaux C, Depaulis A, Micheletti G, Warter JM (1987) Spontaneous spike and wave discharges in thalamus and cortex in a rat model of genetic petit mal-like seizures. Exp Neurol 96:127-136.
Vezzani A, Michalkiewicz M, Michalkiewicz T, Moneta D, Ravizza T, Richichi C, Aliprandi M, Mule F, Pirona L, Gobbi M, Schwarzer C, Sperk G (2002) Seizure susceptibility and

162
epileptogenesis are decreased in transgenic rats overexpressing neuropeptide Y. Neuroscience 110:237-243.
Vezzani A, Rizzi M, Conti M, Samanin R (2000) Modulatory role of neuropeptides in seizures induced in rats by stimulation of glutamate receptors. J Nutr 130:1046S-1048S.
Vezzani A, Sperk G, Colmers WF (1999) Neuropeptide Y: emerging evidence for a functional role in seizure modulation. Trends Neurosci 22:25-30.
Wahlestedt C, Reis DJ (1993) Neuropeptide-Y-Related Peptides and Their Receptors - Are the Receptors Potential Therapeutic Drug Targets. Annual Review of Pharmacology and Toxicology 33:309-352.
Warter JM, Vergnes M, Depaulis A, Tranchant C, Rumbach L, Micheletti G, Marescaux C (1988) Effects of drugs affecting dopaminergic neurotransmission in rats with spontaneous petit mal-like seizures. Neuropharmacology 27:269-274.
Westrin A, Ekman R, Traskman-Bendz L (1999) Alterations of corticotropin releasing hormone (CRH) and neuropeptide Y (NPY) plasma levels in mood disorder patients with a recent suicide attempt. Eur Neuropsychopharmacol 9:205-211.
Wharton J, Gordon L, Byrne J, Herzog H, Selbie LA, Moore K, Sullivan MH, Elder MG, Moscoso G, Taylor KM, et al. (1993) Expression of the human neuropeptide tyrosine Y1 receptor. Proceedings of the National Academy of Sciences of the United States of America 90:687-691.
Wiebe S (2000) Epilepsy: A comprehensive textbook on CD-ROM. BMJ 320:810A. Wiegner AW, Wierzbicka MM (1987) A method for assessing significance of peaks in cross-
correlation histograms. Journal of neuroscience methods 22:125-131. Williams D (1953) A study of thalamic and cortical rhythms in petit mal. Brain : a journal of
neurology 76:50-69. Woldbye DP (1998) Antiepileptic effects of NPY on pentylenetetrazole seizures. Regul Pept
75-76:279-282. Woldbye DP, Kokaia M (2004) Neuropeptide Y and seizures: effects of exogenously applied
ligands. Neuropeptides 38:253-260. Woldbye DP, Larsen PJ, Mikkelsen JD, Klemp K, Madsen TM, Bolwig TG (1997) Powerful
inhibition of kainic acid seizures by neuropeptide Y via Y5-like receptors. Nat Med 3:761-764.
Woldbye DP, Madsen TM, Larsen PJ, Mikkelsen JD, Bolwig TG (1996) Neuropeptide Y inhibits hippocampal seizures and wet dog shakes. Brain research 737:162-168.
Woldbye DP, Nanobashvili A, Sorensen AT, Husum H, Bolwig TG, Sorensen G, Ernfors P, Kokaia M (2005) Differential suppression of seizures via Y2 and Y5 neuropeptide Y receptors. Neurobiol Dis 20:760-772.
Wolf P (1979) [Nomenclature and classification of epileptic seizures and syndromes (author's transl)]. Der Nervenarzt 50:547-554.
Woods SC, Seeley RJ, Porte D, Schwartz MW (1998) Signals that regulate food intake and energy homeostasis. Science 280:1378-1383.
Wu LG, Saggau P (1997) Presynaptic inhibition of elicited neurotransmitter release. Trends Neurosci 20:204-212.
Xapelli S, Bernardino L, Ferreira R, Grade S, Silva AP, Salgado JR, Cavadas C, Grouzmann E, Poulsen FR, Jakobsen B, Oliveira CR, Zimmer J, Malva JO (2008) Interaction between neuropeptide Y (NPY) and brain-derived neurotrophic factor in NPY-mediated neuroprotection against excitotoxicity: a role for microglia. European Journal of Neuroscience 27:2089-2102.
Yan H, Yang J, Marasco J, Yamaguchi K, Brenner S, Collins F, Karbon W (1996) Cloning and functional expression of cDNAs encoding human and rat pancreatic polypeptide receptors. Proceedings of the National Academy of Sciences of the United States of America 93:4661-4665.
Zhang L, Jones EG (2004) Corticothalamic inhibition in the thalamic reticular nucleus. J Neurophysiol 91:759-766.

163
Zheng TW, O'Brien TJ, Morris MJ, Reid CA, Jovanovska V, O'Brien P, van Raay L, Gandrathi AK, Pinault D (2012) Rhythmic neuronal activity in S2 somatosensory and insular cortices contribute to the initiation of absence-related spike-and-wave discharges. Epilepsia 53:1948-1958.
Zhou JL, Shatskikh TN, Liu X, Holmes GL (2007) Impaired single cell firing and long-term potentiation parallels memory impairment following recurrent seizures. Eur J Neurosci 25:3667-3677.
Zini I, Pich EM, Fuxe K, Lenzi PL, Agnati LF, Harfstrand A, Mutt V, Tatemoto K, Moscara M (1984) Actions of Centrally Administered Neuropeptide-Y on Eeg Activity in Different Rat Strains and in Different Phases of Their Circadian Cycle. Acta Physiol Scand 122:71-77.

Minerva Access is the Institutional Repository of The University of Melbourne
Author/s:
GANDRATHI, ARUN
Title:
Effects of neuropeptide y on single neuronal firing patterns in a genetic rat model of absence
epilepsy
Date:
2016
Persistent Link:
http://hdl.handle.net/11343/113597
File Description:
Effects of neuropeptide y on single neuronal firing patterns in a genetic rat model of absence
epilepsy