effects of malathion on humoral immunity and macrophage function
TRANSCRIPT

FUNDAMENTAL AND APPLIED TOXICOLOGY 3 1 , 2 5 2 - 2 5 8 (1996)
ARTICLE NO. 0097
Effects of Malathion on Humoral Immunity and MacrophageFunction in Mast Cell-Deficient Mice1
KATHLEEN RODGERS,2 KAREN ST. AMAND, AND SHIQUAN XIONG
Livingston Research Center, University of Southern California, 1321 N Mission Road, Los Angeles, California 90033
Received October 27. 1995; accepted March 6, 1996
Effects of Malathion on Humoral Immunity and MacrophageFunction in Mast Cell-Deficient Mice. RODGERS, K., ST. AMAND,K., AND XIONG, S. (1996). Fundam. Appl. Toxicol. 31, 252-258.
Malathion, when administered at noncholinergic doses, was pre-viously shown to enhance the humoral immune response to a In-dependent antigen, sheep red blood cells (SRBC), and macrophagefunction. In addition, malathion was shown to cause mast celldegranulation. The hypothesis that mast cells contribute to theobserved alterations in humoral immunity and macrophage func-tion was determined by examination of the effects of acute admin-istration of malathion to mast cell-deficient mice on macrophagefunction and the generation of a humoral immune response toSRBC. Initial studies in two strains of mast cell-deficient mice(6-7 weeks old) indicated that oral administration of malathionreduced macrophage function in these mice, but enhanced macro-phage function in the wild-type strain. Because both strains re-acted in a similar fashion and the defect in the WBB6F1-W/WV
strain allowed reconstitution, further studies were conducted withthis strain. Exposure of either wild-type mice or mast cell-deficientmice with reconstituted with bone marrow-derived mast cells(BMMC) from the wild-type mice to malathion enhanced macro-phage function and the production of circulating IgM, but notIgG, antibodies to SRBC on Days 3 and 5 after immunization. Incontrast, administration of malathion to older mast cell-deficientmice suppressed the generation of IgM and IgG antibodies toSRBC on Days 3 and 5 after immunization, but did not affectmacrophage function. In summary, the results presented indicatethat the presence of mast cells was necessary for the increases inmacrophage function and humoral immunity observed after acuteoral administration of malathion to mice, e 19% Society of Toxicology
Malathion is an organophosphate pesticide which has a lowmammalian toxicity due to detoxification of malathion intonontoxic derivatives by carboxylesterases (Aldridge et ai,1979; Dauterman, 1971; Frawley et ai, 1957; March et ai,1956; Umetsu et ai, 1977. 1981). Because of this low toxicity.malathion is used in situations where large urban populationsand domesticated animals may be exposed (Weeks et ai, 1977).
1 Work was Supported by PHS ESO4337.2 To whom correspondence should be addressed.
Such situations include mosquito eradication, spraying of to-bacco plants, and aerial spraying for the eradication of fruitflies in citrus-producing states. One published study by Milbyand Epstein (1964) has suggested that malathion may causecontact dermatitis (immune hypersensitivity) in human popula-tions. In this study, 43% of persons exposed to technical mala-thion under an occlusive patch for 48 hr developed contactdermatitis upon reexposure. Due to the brevity of technicalinformation in this publication, the ability of malathion to causerash formation after the first exposure or during shorter expo-sures is not known. In addition, the study design did not distin-guish between a direct effect of malathion on the mast cell oran indirect effect through the generation of an immune responseto malathion.
Animal studies have shown that acute administration of non-cholinergic doses of purified malathion elevated the humoralimmune and proliferative response to mitogenic stimuli (Rod-gers et ai, 1986; Rodgers and Ellefson, 1990). The increasein proliferative responses to mitogen was shown to be due toalterations in the adherent cell population (Rodgers and Ellef-son, 1990). Further studies indicated that acute, oral administra-tion of malathion to mice or in vitro exposure of mouse orhuman white cells to metabolized malathion increased the res-piratory burst of leukocytes stimulated by phorbol esters (Rod-gers and Ellefson, 1990). More recently, it was shown thatadministration of very low doses of malathion increased macro-phage function as measured by respiratory burst function andphagocytosis (Rodgers and Ellefson, 1992). These studies alsoshowed that in vivo administration of malathion led to degranu-lation of peritoneal mast cells. Most recently, it was shownthat in vitro exposure of purified mast cells or a basophilictumor cell line to metabolites of malathion resulted in the re-lease of mast cell mediators (Xiong and Rodgers, submitted).This study confirms that malathion, particularly metabolizedmalathion, can cause the degranulation of basophilic cells.These data suggest that the observations made by Milby andEpstein (1964) may be the result of direct degranulation ofmast cells by malathion rather than through the generation ofa specific immune response.
Published reports indicate that mast cell mediators, whichinclude cytokines, lipid-derived mediators, histamine, hepa-
0272-0590/96 SI8.00Copyright © 1996 by the Society of ToxicologyAll rights of reproduction in any form reserved.
252
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022

MALATHION IMMUNOTOXICITY 253
rin, proteases, and proteoglycans, can modulate macrophagefunction (Bonavida et al., 1990). In addition, macrophagescan ingest mast cell granules after degranulation (Baggioliniet al., 1982; Lindahl et al, 1979; Rodgers and Ellefson,1992). Further studies have shown that mast cells expresshistamine receptors and expression of these receptors is mod-ulated by macrophage differentiation (Feder et al, 1991;Gespach et al., 1986). Mouse macrophages have also beenshown to express heparin receptors. Studies by Yoffee andothers indicated that mast cell products and more specificallyheparin and other polyanions increased the production ofinflammatory mediators by and other activities of mononu-clear cells (Yoffee et al, 1985; Bleiberg et al, 1993; Schultzet ai, 1977; Jaques et al, 1979). Therefore, studies wereconducted to determine if the presence of mast cells wasrequired for the increases in immune activities observed afteradministration of noncholinergic doses of malathion.
MATERIALS AND METHODS
Animals and Mast Cell Reconstitution
Mice. Mast cell-deficient female mice, WBB6F1-W/WV(W/WV) orWCB6FI-SI/Sld, and their normal littermates, WBB6F1-+/+(+/+) orWCB6F1-+/+, respectively, were used in initial studies. Each mouse strainhas a separate genetic defect in the ability to generate mast cells, but asimilar phenotype (i.e., mast cell deficiency, Galh and Kitamura, 1987).The mice were purchased from the Jackson Laboratory (Bar Harbor, ME)and housed in the USC vivarium. Food and water were available ad libitumand the mice were housed on a 12:12 hr light:dark cycle. The malathionwas dissolved in corn oil and administered via oral gavage. For the majorityof the studies. WBB6F1-W/WV mice were utilized because the geneticdefect in this mouse strain leading to mast cell deficiency allows reconstitu-tion of the mice with bone marrow-derived mast cells (BMMC) from wild-type mice (Kitamura et al., 1978).
Preparation of bone marrow-derived mast cells. Bone marrow cellswere obtained from femurs of 10-week-old WBB6F1-+/+ (+/+) mice.WBB6FI-W/WV (W/Wv) mice were reconstituted with purified culturedBMMC from wild-type mice at age 90 days or more (Ramos et al., 1991,1992). BMMC were prepared according to a procedure previously described(Razin et ai. 1981; Frandji et al., 1993). Briefly, 5 X 10" bone marrowcells were suspended in 25 ml enriched culture medium: RPMI 1640 supple-mented with 10% heated-inactivated fetal calf serum (FCS), antibiotics [100//g/ml streptomycin, 100 U/ml penicillin, 250 /jg/ml amphotericin B], 25min Hepes, 4 ITIM L-glutamine, 0.1 mM nonessential amino acid, 1 minsodium pyruvate, and 50% WEHI-3-conditioned medium (WEHI-3-CM,source of IL-3) in 75-cm2 tissue culture flasks (Falcon) and incubated at37°C in humidified 5% CO2/95% air. The cell suspensions were centrifugedat 400g for 10 min and the cells were resuspended in fresh medium at 7-day intervals. Macrophages present within the cultures were removed byattachment to plastic surfaces of a flask. A sample of the nonadherent cellswas stained with toluidine blue (pH 3.5) to determine the percentage ofmetachromatic cells (Frandji et al., 1993).
WEHI-3-CM was produced by seeding WEHI-3 cells (1 X 106 cells/ml)into culture medium [Iscove's modified Dulbecco's medium with 0.05 mM2-mercaptoethanol, 4 mM L-glutamine, and 10% FCS] and incubating thesecultures at 37°C in 5% CO2 for 3 days. The cells were then transferred intomedium without serum for an additional 3 days. This serum-free mediumwas utilized as WEH1-CM above. The supernatant was centrifuged at 600gfor 20 minutes, filtered through a 0.2-/jm filter system, and stored at -70°Cuntil use. AH reagents were purchased from GIBCO (Grand Island, NY).
Mast cell reconstitution. After a 3-week culture with medium supple-mented with WEHI-CM, the cultures of bone marrow cells contained almostexclusively mast cells (BMMC). An aliquot, 5-10 X 106, of BMMC wasinjected intrapentoneally into each mouse undergoing reconstitution. Themast cell-deficient mice reconstituted with cultured BMMC were utilized3 weeks after injection (appropriately 16 to 17 weeks of age). The abilityto reconstitute with BMMC was assessed through microscopic examinationfor the presence of mast cells of the cells in peritoneal lavage fluid fromeach mouse stained with Alcian blue. In addition, a portion of the skinfrom one control mouse in each study was fixed in Carnoy's solution,stained with toluidine blue, and examined microscopically for the presenceof mast cells. Different groups examined in these studies were designatedas follows: (1) WAV" + corn oil; (2) W/Wv + malathion (600 mg/kg sameas below); (3) W/Wv + BMMC + corn oil; (4) WAV" + BMMC + mala-thion; (5) +/+ + corn oil; (6) +/+ + malathion.
Macrophage Functional Assays
Macrophage respiratory burst function. The respiratory burst functionwas assessed by the measurement of hydrogen peroxide production afterstimulation with phorbol ester as previously described (Rodgers and Ellef-son, 1992; Rodgers, 1995). Briefly, murine peritoneal cells were harvested,the viability was assessed by trypan blue exclusion, and the cells wereresuspended at 2 X 106 cells/ml in phosphate-buffered saline (PBS, pH7.4). Aliquots, 100 y\, of cell suspension were added into wells of a 96-well microtiter plate (Falcon). After a 1-hr incubation, cells were washedthree times to remove nonadherent cells. Ninety microliters of PBS with2,7-dichlorofluoroscein diacetate [DcFD] (5 /XM final concentration; KodakChemicals, Rochester, NY) was added into each well and incubated for 15min. Ten microliters of PMA (100 ng/ml, Sigma, St. Louis, MO) was addedinto each well to stimulate hydrogen peroxide production. The increase influorescent product was measured after a 1-hr incubation at 37°C on aMillipore fluorescence measurement system (Cytofluor 2350) [excitation,505; emission, 530]. The intensity of the fluorescent product, 2,7-dichloro-fluoroscein, was compared to a standard (Polysciences, Warrington, PA).
Phagocytosis assay. Assessment of the phagocytic capability of peritonealmacrophages was conducted as previously described (Rodgers et ai, 1985;Rodqus and Ellefson, 1988). Yeast particles (Sigma) were hydrated in PBSand boiled for 1 hr to inactivate the particles. The inactivated particles werethen opsonized by mixing 2 X 109 yeast particles/ml with heat-inactivatedserum from WBB6F1--1-/+ mice. After 1 hr, the particles were washed threetimes with PBS to remove excess serum proteins. In order to adhere theperitoneal cells for the measurement of phagocytic capability, 2 x 106 mononu-clear cells in 1 ml were placed onto a glass coverslip (22 x 22 mm) that hadbeen placed in a 35-mm tissue culture dish. The cells were then allowed toadhere in a humidified environment of 5% CO2 in air at 37°C for 1 to 2 hr.At the end of this incubation, the coverslip was washed gently six times withPBS to remove nonadherent cells. After each rinse, one end of the coverslipwas blotted with an absorbent towel to remove the loosely associated liquid.An aliquot, 100 ^1, of opsonized yeast was then added to the adherent peritonealcells and incubated for 20 min longer. The coverslips were then washed threeto six times and placed cell side down on the microscope slide for assessment.The number of yeast ingested per macrophage was determined for 100 to 200cells per coverslip.
Humoral Immune Response
Immunization and sera collection. The mice were immunized to sheepred blood cells (SRBC, Colorado Serum, Denver, CO), 2 x 10s SRBC/mouse, via intraperitoneal injection. At Days 3, 5, and 7 after immunization,blood was harvested from the retroorbital space (Days 3 and 5) underMetofane anesthesia or by cardiac puncture immediately after terminationon Day 7. The blood was allowed to clot overnight at 4°C and centrifugedfor 15 min at 800g, and the serum harvested to assay for antibodies toSRBC. The sera were stored in cryogenic vials at -20°C until assayed.
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022

254 RODGERS, ST. AMAND. AND XIONG
ELISA plates. The ELISA plates were made 1 to 3 days prior to per-forming the assay. Fifty microliters of 0.1 mg/ml poly-D-lysine (Sigma)were added to each well of a U-bottom microtiter plate for ELISA (Dyna-tech, Chantilly, VA) and incubated for 30 min at room temperature. Theplates were then washed three times with distilled water. Plasma membranepreparations were generated by lysis of washed SRBC in distilled waterfollowed by somcation for 1 min and centrifuged at 10,000g for 10 min.The pellet from this centnfugation step was then diluted into coating buffer(carbonate buffer, pH 9.6) at an equivalent of 1 X I08 cells/ml. Fiftymicroliters of this SRBC plasma membrane preparation was added to eachwell followed by 25 /zl/well of 1.5% glutaraldehyde. The plates were thencovered with film wrap and incubated for 1 hr at room temperature. Theplates were nnsed three times with distilled water and the remaining bindingsites were blocked with 100 //I/well PBS, pH 7.2. with 0 5% bovine serumalbumin, 0.01% Tween, and 0.01% sodium azide (blocking buffer). Theplates were again covered with film wrap and incubated for either I hr at37°C or 2 hr at room temperature. The plates were then washed three timeswith PBS containing 0.01% Tween (PBS-Tween) and two times withdistilled water. The plates were stored at 4°C until use.
ELISA assay. The sera were diluted 1:100, 1:200, and 1:400 in dilutingbuffer (PBS, pH 7.2, containing 0.2% BSA). Each diluted serum samplewas aliquoted in tnplicate on two plates (one to measure anti-SRBC IgGlevels and the other to measure anti-SRBC IgM levels) at 50 /A per well.The plates were incubated at 37°C for 2 to 3 hr. After this incubation, theplates were washed three times with PBS-Tween. Second antibody (rabbitanti-mouse IgG or IgM, as appropnate, coupled with horseradish peroxi-dase, Sigma), diluted 1:500 in diluting buffer, was then added into eachwell. These plates were then incubated for 1 hr at room temperature andthe plates were washed five times with PBS-Tween and three times withdistilled water. Thereafter, 100 fi\ substrate, 2,2'-azino-bis(3-ethylbenzthia-zoline-6-sulfonic acid) [ABTS; 0.5 mg/ml in citrate phosphate buffer, pH4.0, Sigma] with 0 015% hydrogen peroxide (Sigma), was then added toeach well. After 30 min of incubation at room temperature, the reactionwas stopped by addition of 25 fi\ 0.33 M sodium fluoride. The absorbanceof each well was then measured on a Dynatech Microplale Reader MR650with a Apple IIC computer attached.
RESULTS
Macrophage Functional Assays
Initial studies were done on mice that were 6-8 weeksold to determine if the wild-type mice from two separatestrains that have mast cell-deficient counterparts (with de-fects in two separate sites for mast cell differentiation) hadan increase in their macrophage function following acute,oral administration of malathion. In addition, the effects oforal administration of malathion on the macrophage functionof these two strains of mast cell-deficient mice were deter-mined. The results from these studies are shown in Table 1.As can be seen, in both strains of mice, acute, oral adminis-tration of 300 mg/kg malathion to the wild-type mice ele-vated the production of hydrogen peroxide by peritonealleukocytes while the same treatment suppressed the produc-tion of hydrogen peroxide by the peritoneal leukocytes ofmice genetically altered to be deficient in mast cells (W/Wv and Sl/Sld). Because both strains were responsive tomalathion treatment and the genetic defect that leads to thephenotype of mast cell deficiency in the WAVV mice allowedreconstitution of mast cells by injection with bone marrow-
TABLE 1Effect of Malathion Administration on the Respiratory Burst
of Macrophages from Mast Cell-Deficient Mice
Production of hydrogen peroxide (nmol/106)
Groups Mean ± SEM
WB6-Wt corn oilWB6-Wt + malathionWAV" + corn oilWAV1 + malathionWC6-Wt + corn oilWC6-Wt + malathionSl/Sld + com oilSl/Sld + malathion
14.02 ± 1.2022.33 ± 2.1013.94 ± 0.946.14 ± 0.49
15.14 ± 1.6021.09 ± 2.1919.25 ± 1.9013.06 ± 1.31
Note. The effect of oral administration of malathion on macrophage respira-tory burst was examined. Mast cell-deficient (WAV or Sl/Sld) mice and wild-type (Wt) mice were treated with corn oil or malathion (300 mg/kg) via oralgavage and the respiratory burst function was determined 24 hr later. Eachdata point is the mean and standard error of six mice in two experiments, pwas less than 0.001 for all comparisons between com oil- and malathion-treated groups.
derived mast cells (BMMC) from the wild-type mice, allfurther studies were conducted using W/Wv mice.
In order to reconstitute the mice, older mice were used(90 days at the time of injection of BMMC). Attempts toreconstitute younger mice (6-8 weeks at the time of injec-tion) were not successful (data not shown). Because the micewere used for the experiments 20-21 days after injection,the mice used in these studies were 16-17 weeks of age attreatment rather than 6-8 weeks of age as above. The wild-type and mast cell-deficient controls were age matched tothe reconstituted group. In this study, acute, oral administra-tion of 600 mg/kg malathion did not suppress macrophagefunction in the mast cell-deficient mice, but did enhance thefunction of peritoneal macrophages in the wild-type and themast cell-deficient mice that had been reconstituted withBMMC (Fig. 1 and Tables 2 and 3). In Fig. 1, the effectsof malathion administration on the respiratory burst of peri-toneal macrophages after stimulation in vitro with phorbolester are shown. As stated above, malathion did not affectthe ability of macrophages from mast cell-deficient mice togenerate hydrogen peroxide, but did enhance the respiratoryburst function of wild-type and reconstituted mice. Thissame pattern of results was observed with regard to theeffects of malathion on the phagocytosis of opsonized yeastby peritoneal macrophages (Table 2 [percentage of macro-phages with no yeast ingested] and Table 3 [average numberof yeast ingested per macrophage]). However, the phagocyticcapability of the reconstituted mice was reduced comparedto wild-type controls (Table 3).
Humoral Immunity
Initial studies were conducted with mast cell-deficientmice to determine if the ability of the mice to respond to
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022
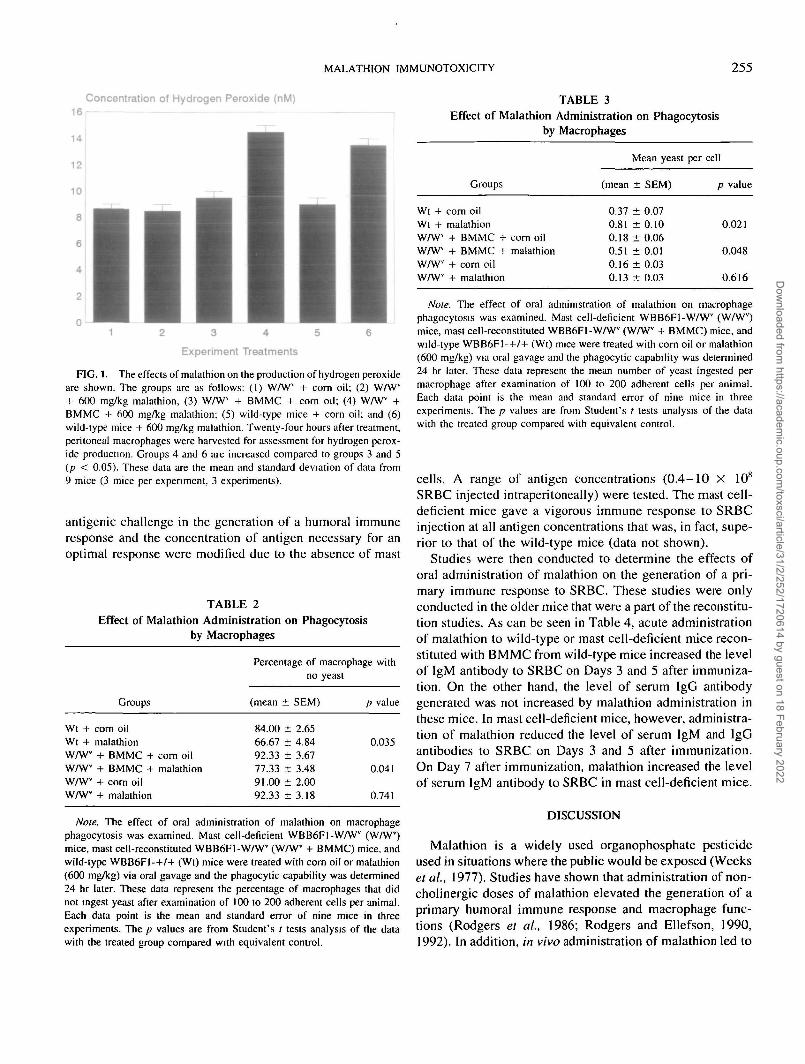
MALATHION IMMUNOTOXICITY 255
Concentration of Hydrogen Peroxide (nM) TABLE 3Effect of Malathion Administration on Phagocytosis
by Macrophages
Experiment Treatments
FIG. 1. The effects of malathion on the production of hydrogen peroxideare shown. The groups are as follows: (1) WAV" + corn oil; (2) W/Wv
+ 600 mg/kg malathion, (3) WAVV + BMMC + corn oil; (4) WAV" +BMMC + 600 mg/kg malathion; (5) wild-type mice + corn oil; and (6)wild-type mice + 600 mg/kg malathion. Twenty-four hours after treatment,peritoneal macrophages were harvested for assessment for hydrogen perox-ide production. Groups 4 and 6 are increased compared to groups 3 and 5(p < 0.05). These data are the mean and standard deviation of data from9 mice (3 mice per experiment, 3 experiments).
antigenic challenge in the generation of a humoral immuneresponse and the concentration of antigen necessary for anoptimal response were modified due to the absence of mast
TABLE 2Effect of Malathion Administration on Phagocytosis
by Macrophages
Wt +Wt +WAV"WAVV
WAVV
WAVV
Groups
corn oilmalathion+ BMMC + corn oil+ BMMC + malathion+ com oil+ malathion
Percentage of macrophage withno yeast
(mean ± SEM)
84.00 ± 2.6566.67 ± 4.8492.33 ± 3.6777.33 ± 3.4891.00 ± 2.0092.33 ± 3 . 1 8
p value
0.035
0.041
0.741
Note. The effect of oral administration of malathion on macrophagephagocytosis was examined. Mast cell-deficient WBB6FI-WAVV (WAVV)mice, mast cell-reconstituted WBB6F1-WAVV (WAVV + BMMC) mice, andwild-type W B B 6 F I - + / + (Wt) mice were treated with corn oil or malathion(600 mg/kg) via oral gavage and the phagocytic capability was determined24 hr later. These data represent the percentage of macrophages that didnot ingest yeast after examination of 100 to 200 adherent cells per animal.Each data point is the mean and standard error of nine mice in threeexperiments. The p values are from Student's / tests analysis of the datawith the treated group compared with equivalent control.
Wt +Wt +WAV1
W/WWAVV
WAVV
Groups
corn oilmalathion+ BMMC + corn oil+ BMMC + malathion+ corn oil+ malathion
Mean yeast
(mean :
0.37 d0.81 d0.18 i0.51 i0.16 i0.13 j
t SEM)
: 0.07t 0.10i 0.06: 0.01t 0.03t 0.03
per cell
p value
0.021
0.048
0.616
Note. The effect of oral administration of malathion on macrophagephagocytosis was examined. Mast cell-deficient WBB6F1-WAVV (WAV")mice, mast cell-reconstituted WBB6F1-WAVV (WAV" + BMMC) mice, andwild-type WBB6F1-+ /+ (Wt) mice were treated with corn oil or malathion(600 mg/kg) via oral gavage and the phagocytic capability was determined24 hr later. These data represent the mean number of yeast ingested permacrophage after examination of 100 to 200 adherent cells per animal.Each data point is the mean and standard error of nine mice in threeexperiments. The p values are from Student's I tests analysis of the datawith the treated group compared with equivalent control.
cells. A range of antigen concentrations (0.4-10 X 108
SRBC injected intraperitoneally) were tested. The mast cell-deficient mice gave a vigorous immune response to SRBCinjection at all antigen concentrations that was, in fact, supe-rior to that of the wild-type mice (data not shown).
Studies were then conducted to determine the effects oforal administration of malathion on the generation of a pri-mary immune response to SRBC. These studies were onlyconducted in the older mice that were a part of the reconstitu-tion studies. As can be seen in Table 4, acute administrationof malathion to wild-type or mast cell-deficient mice recon-stituted with BMMC from wild-type mice increased the levelof IgM antibody to SRBC on Days 3 and 5 after immuniza-tion. On the other hand, the level of serum IgG antibodygenerated was not increased by malathion administration inthese mice. In mast cell-deficient mice, however, administra-tion of malathion reduced the level of serum IgM and IgGantibodies to SRBC on Days 3 and 5 after immunization.On Day 7 after immunization, malathion increased the levelof serum IgM antibody to SRBC in mast cell-deficient mice.
DISCUSSION
Malathion is a widely used organophosphate pesticideused in situations where the public would be exposed (Weekset at, 1977). Studies have shown that administration of non-cholinergic doses of malathion elevated the generation of aprimary humoral immune response and macrophage func-tions (Rodgers et al, 1986; Rodgers and Ellefson, 1990,1992). In addition, in vivo administration of malathion led to
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022

256 RODGERS. ST. AMAND, AND XIONG
TABLE 4Serum Antibody Titer to Sheep Red Blood Cells
Group
igMWt + corn oilWt + malathionWAV" + BMMC + corn oilWAVV + BMMC + malathionW/Wv + corn oilW/Wv + malathion
IgGWt + corn oilWt + malathionWAVV + BMMC + corn oilW/Wv + BMMC + malathionW/Wv + corn oilWAVV + malathion
Optical density,
3
0.324 ± 0.0300.400 ± 0.038*0.375 ± 0.0380.514 ± 0.051*0.551 ± 0.0560.235 ± 0.019*
0.320 ± 0.0400.391 ± 0.0430.318 ± 0.0370.400 ± 0.023*0.347 ± 0 0320.241 ± 0 021*
410 nM, mean ± SD Day
5
0.890 ± 0.0951.171 ± 0.106*0.966 ± 0.0481.265 ± 0.096*1.085 ±0.1020.903 ± 0.048*
0.657 ± 0.0600.770 ± 0.0800.652 ± 0.0690.832 ± 0.0960 843 ± 0.0890.625 ± 0.047*
postimmunization
7
0.831 + 0.0790.786 + 0.0690.850 ± 0.0950.741 ± 0.0820.910 ± 0.0931.137 ± 0.096*
0.606 + 0.0610.569 ± 0.0860.570 ± 0.0510.673 ± 0.0670.859 ± 0.0620.904 ± 0.093
Note. Wild-type WBB6F1-+/+ (Wt), mast cell-deficient WBB6FI- W/Wv (W/Wv), and reconstituted WAV1 (WAVV + BMMC) mice were treatedwith corn oil or 600 mg/kg malathion via oral gavage. The mice were immunized with SRBC and serum was harvested at various postimmunizationdays. The level of antibodies in the serum was assessed via ELISA. Experiments were performed three times with three mice per group in each experimentThese data are from one experiment and other experiments gave comparable results.
* p value =s 0.05 comparing corn oil and malathion group.
degranulation of peritoneal mast cells (Rodgers and Ellefson,1992). Published reports indicate that mast cell mediators,which include cytokines, lipid-derived mediators, histamine,heparin, proteases, and other enzymes, chemotactic factors,and proteoglycans, can modulate macrophage function andshow that macrophages can ingest mast cell granules (per-haps through a specific receptor on the macrophage mem-brane) after degranulation (Bonavida et ai, 1990; Klein etai, 1989; Baggiolini et ai, 1982; Lindahl el ai, 1979; Ishi-zaka and Ishizaka, 1984; Schwartz and Austen, 1984; Hen-derson and Kaliner, 1978; Dileepan et ai, 1989). Other stud-ies have shown that mast cell granules inhibit macrophagetumoricidal activity (perhaps through inhibition of bindingof lipopolysaccharide to its receptor) and nitric oxide produc-tion (Dileepan et ai, 1990, 1993).
Macrophages have also been shown to express receptorsfor mast cell mediators, including histamine and heparin(Bleiberg et ai, 1983; Feder et ai, 1991; Gespach et ai,1991). Through activation of these receptors, alterations inmacrophage activity have been observed. For example, hista-mine was shown to increase intracellular calcium in andinterleukin 1 production by macrophages (Ohno et ai, 1991;Okamato et ai, 1990). Studies by Yoffee and others indi-cated that mast cell products and more specifically heparinand other polyanions increased the production of inflamma-tory mediators by and other activities of mononuclear cells(Yoffee et ai, 1985; Bleiberg et ai, 1993; Schultz et ai,1977; Jaques, 1979). Therefore, studies were conducted todetermine if the presence of mast cells was required for the
expression of the alterations in immune effects observedafter administration of noncholinergic doses of malathion.
In this report, it was shown that acute, oral administrationof noncholinergic doses of malathion to mast cell-deficientmice suppressed (young mice) or did not affect (older mice)the function of peritoneal macrophages as measured by theproduction of hydrogen peroxide and phagocytosis of opso-nized yeast. In addition, administration of malathion to mastcell-deficient mice (older) suppressed the level of serum IgMand IgG antibodies to SRBC on Days 3 and 5 after immuni-zation and increased the level of serum IgM antibodies toSRBC on Day 7 after immunization. These data show thatin the absence of mast cells oral administration of malathioncan suppress macrophage and humoral immune function.
On the other hand, reconstitution of the mast cell popula-tion of these mice with BMMC from wild-type mice restoredthe ability of oral exposure to malathion to increase macro-phage function (as measured by the production of hydrogenperoxide after stimulation and the ingestion of opsonizedyeast) and humoral immune function (as measured by thegeneration of a primary immune response to a T-dependentantigen) in a manner comparable to wild-type mice. Thesedata indicate that the genetic defect in the mast cell-deficientmice that led to this phenotype did not alter the ability ofthe mice to respond to malathion. Therefore, these studiessupport the hypothesis that mast cells are involved in theenhanced macrophage and humoral immune function ob-served after administration of malathion.
As stated, mast cells were shown to have a role in the
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022

MALATHION IMMUNOTOXICITY 257
increase in macrophage and humoral immune function ob-served after oral administration of malathion. These data,together with previous studies of the kinetics of macrophageactivation after malathion administration, are consistent withthe hypothesis that mast cells rapidly interact with macro-phages to modulate a variety of functional parameters includ-ing the generation of oxygen radicals and Fc receptor-medi-ated phagocytosis. Recent studies from one laboratory hasprovided compelling evidence that mast cell granules in-creased the production of hydrogen peroxide by alveolarmacrophage through an increase in the conversion of super-oxide anion to hydrogen peroxide. This was shown to bedue to the fact that superoxide dismutase was a componentof the mast cell granules and addition of this exogenousenzyme to the macrophages by ingestion of the granulesincreased hydrogen peroxide production (Dileepan et al.,1989; Dileepan and Stechschulte, 1986; Rock et al., 1990;Henderson and Kaliner, 1978). This may be one mechanismby which oral administration of malathion increased the pro-duction of hydrogen peroxide. On the other hand, studies byRock et al. (1990) showed that mast cell granules also con-tain peroxidase, an enzyme which metabolizes hydrogen per-oxide to water, that can actually decrease the amount ofhydrogen peroxide measured. Therefore, the results con-tained in this paper may be the result of (1) actual increasesin the respiratory burst system per se or (2) the addition ofexogenous enzymes to the system by the granules.
Studies have also shown that mast cells can respond toinducer T lymphocytes and that activated mast cells expressClass II major histocompatibility antigens. Based upon thesestudies, some have suggested that mast cells may act asantigen-presenting cells (Warbrick et al., 1995; Frandji etal., 1993; Nabel et al., 1981). In this study, the presence ofmast cells was necessary for the observed increase in thegeneration of a primary humoral immune response afteracute administration of malathion. It is therefore possiblethat the increase in this immune parameter is the result ofalterations in both macrophage and mast cell function aftermalathion exposure.
The data also showed that as the mast cell-deficient miceaged, oral administration of malathion no longer altered mac-rophage function. In contrast, the effect of malathion on themacrophage function of wild-type mice was not altered asthe mice aged (6 -8 weeks versus 16-17 weeks). There isno explanation for these differences at this time. However,the humoral immune response was inhibited in mice in whichmacrophage function was not altered; therefore, in the ab-sence of mast cells, the suppression in antibody formationis not always the result of alterations in macrophage function.
In summary, these data suggest that the presence of mastcells is necessary for the observed increases in macrophagefunction and humoral immune responses after malathion ex-posure. The mechanism by which malathion caused mast
cell degranulation and the mediator that is released by themast cell to affect these functional parameters are currentlyunknown. There are several mast cell mediators that arecapable of stimulating macrophage and humoral immunefunctions. Determination of the mediator involved is thesubject of ongoing studies.
REFERENCES
Aldridge. W. N., Miles, J. W., Mount, D. L., and Vershoyle. R. D. (1979).The toxicologic properties of impurities in malathion. Arch. Toxicol. 42,95-109.
Baggiolini, M., Horisberger. U.. and Martin, U. (1982). Phagocytosis ofmast cell granules by mononuclear phagocytes, neutrophils and eosmo-phils during anaphylaxis. Int. Arch. Allergy Appl Immunol. 67, 219.
Bleiberg, I., MacGregor, I., and Aronson, M. (1983). Heparin receptors onmouse macrophages. Thromb. Res. 29, 53.
Bonavida, B . Mencia-Huerta, J. M., and Braquet. P. (1990). Effects ofplatelet-activating factor on peripheral blood monocytes: Induction andpriming for TNF secretion. J. Lipid Mediators 2, S65-S76.
Dauterman, W. C. (1971). Biological and nonbiological modifications oforganophosphorus compounds Bull. WHO 44, 133-150.
Dileepan, K. N.. Simpson, K. M., and Stechschulte. D. J. (1989) Modulationof macrophage superoxide-induced cytochrome c reduction by mast cells.J. Lab. Clin. Med. 113, 577-585.
Dileepan, K. N., Lei. M. G., Morrison, D. C . and Stechschultze. D. J.(1990). Inhibition of macrophage-mediated tumor cell killing by mastcell granules: Evidence for modulation of lipopolysaccharide binding toreceptor. J. Leukocyte Biol. 19, 48.
Dileepan, K. N., Lorsbach, R. B , and Stechschultze, D. J. (1993). Mastcell granules inhibit macrophage-mediated lysis of mastocytoma cells(P815) and nitric oxide production. /. Leukocyte Biol. 53, 446-453.
Dileepan, K. N., and Stechschultze, D J. (1986). Influence of mast cellson macrophage function. Biochem. Soc. Trans. 14, 913-914.
Feder. D.. Bueno, M. A., Krybus, J , Novak, A.. Martins, M. C , Andrade-Junior, A. F., and Cabral, M. (1991). Effects of H2 blockers on thespreading of macrophages in vitro. Rev. Paul. Med. 109, 213-216.
Frandji P., Oskeritzian C , Cacaraci F., Lapeyre J., Peronet R., David B.,Guillet J. G., and Mecheri S. (1993). Antigen-dependent stimulation bybone marrow-derived mast cells of MHC class II-restricted T cell hybrid-oma. J. Immunol. 151, 6318-6328.
Frawley, J. P.. Fuyat, H. N., Hagan, E. C , Blake, J. R., and Fitzhugh,O. G. (1957). Marked potentiation in mammalian toxicity from simultane-ous administration of two anticholinesterase compounds. J. Pharmacol.121,96-104.
Galli, S. J., and Kitamura, Y. (1987). Genetically mast cell-deficient W/Wv and Sl/Sld mice' Their value for the analysis of the roles of mastcells in biologic responses in vivo. Am. J. Pathol. 127, 191 — 198.
Gespach, C , Courillon-Mallet, A.. Launay, J. M., Cost. H.. and Abita,J. J. P. (1986). Histamine receptor activity and histamine metabolism inhuman U937 monocyte-like cells and human peripheral blood monocytes.Agents Actions 18, 124.
Henderson, W. R., and Kaliner, M. (1978). Immunologic and nonimmuno-logic generation of superoxide from mast cells and basophils. J. Clin.Invest. 61, 187-196.
Ishizaka, T. R., and Ishizaka, K. (1984). Activation of mast cells for media-tor release through IgE receptors. Prog. Allergy 34, 188-235.
Jaques, L. B. (1979). Heparin: An old drug with a new paradigm. Science206, 528.
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022

258 RODGERS, ST. AMAND, AND XIONG
Kitamura, Y., Go, S., and Hatanaka, K. (1978). Decrease of mast cells inW/Wv mice and their increase in bone marrow transplantation. Blood
52, 447-452.
Klein, L. M , Lavker, R. M , Matis, W. L., and Murphy, G. F. (1989).Degranulation of human mast cells induces an endothelial antigen central
to leukocyte adhesion. Proc. Natl. Acad. Sci. USA 86, 8972-8976.
Lindahl, U., Pertoft, H., and Selgelid, R. (1979). Uptake and degradation
of mast cell granules by peritoneal macrophages. Biochem. J. 182, 189.
March, R. B., Fukuto, T. R., Metcalf, R. L., and Maxon, M. G. (1956). Fateof P32-labelled malathion in the laying hen, white mouse and American
cockroach. J. Econ. Entomol. 49, 185-195.
Milby, T. B., and Epstein, W. L. (1964). Allergic contact hypersensitivity
to malathion. Arch. Environ. Health 9, 434-437.
Nabel, G., Galli, S. J., Dvorak, M., Dvorak, H. F., and Cantor, H. (1981).Inducer T lymphocytes synthesize a factor that stimulates proliferation
of cloned mast cells. Nature 291, 332-334.
Ohno, S., Shirai, A., Ueda, U., Igarashi, T , Ishigatsubo, Y., Tani, K.,Okubo, T., Hikawa, N., Kawakami, T., and Takenaka, T. (1991). Increasein intracellular calcium induced by stimulating histamine HI receptorsin macrophage-like P388D1 cells. Biochem. Biophys. Res. Commun. 181,
1156-1163.
Okamoto, H., Oh, C , and Nakano, K. (1990). Possible involvement ofadenosine 3':5'-cyclic monophosphate and extracellular calcium ions inhistamine stimulation of interleukin 1 release from macrophage line
P388D1 cells. Immunology Id, 186-190.
Ramos, B. F., Zhang Y., Qureshi R., and Jakschik B. A. (1991). Mast cellsare critical for the production of leukotrienes responsible for neutrophilrecruitment in immune complex-induced peritonitis in mice. J. Immunol.
147, 1636-1641.
Ramos, B. F., Zhang, Y., Angkachatchai, Z., and Jakschik, B. A., (1992).Mast cell mediators regulate vascular permeability changes in Arthus
reaction. J. Pharmacol. Exp. Ther. 262, 2559-2565.
Razin, E., Cordon-Cardo, C , and Good, R. A. (1981). Growth of a purepopulation of mouse mast cells in vitro with conditioned medium derivedfrom concanavalin A-stimulated splenocytes. Proc. Natl. Acad. Sci. USA
78, 2559-2561.
Rock, M. J., Despot, J., and Lemanske, R. F. (1990). Mast cell granulesmodulate alveolar macrophage respiratory burst activity and eicosanoid
metabolism. / Allergy Clin. Immunol. 86, 452-461.
Rodgers, K. (1995). Measurement of the respiratory burst of leukocytes forimmunotoxicologic analysis. Mod. Methods Immunotoxicol, in press.
Rodgers, K. E., and Ellefson, D. D. (1988). Effects of acute administrationof O,O,5-trimethyl phosphorothioate on the respiratory burst and phago-cytic activity of splenic and peritoneal leukocytes. Agents Actions 24,152-160.
Rodgers, K. E., and Ellefson, D. D. (1990). Modulation of respiratory burstactivity and mitogenic response of human peripheral blood mononuclearcells and murine splenocytes by malathion. Fundam. Appl. Toxicol. 14,309-317.
Rodgers, K., and Ellefson, D (1992). Mechanism of the modulation ofmurine peritoneal cell function and mast cell degranulation by low dosesof malathion. Agents Actions 35, 57-63.
Rodgers, K. E., Imamura, T., and Devens, B. H. (1985). Investigations intothe mechanism of immunosuppression caused by acute treatment withO,O,S-trimethyl phosphorothioate. I. Characterization of the immune cellpopulation affected. Immunopharmacology 10, 171-180.
Rodgers, K E., Leung, N., Ware, C. F., Devens, B. H., and Imamura,T. (1986). Lack of immunosuppressive effects of acute and subacuteadministration of malathion. Pestic. Biochem Physiol. 25, 358.
Schwartz, L. B., and Austen, K. F. (1984). Structure and function of thechemical mediators of mast cells. Prog. Allergy 34, 271-321.
Schultz, R. M., Papmatheakis, J. D., and Chirigos, M. A. (1977). Interferonand induces of macrophage activation by polyanions. Science 197, 674.
Umetsu, N., Grose, F. H., Allahyari, R., Abu-El-Haj, S., and Fukuto, T. R.(1977). Effect of impurities on mammalian toxicity of technical malathionand acephate. J. Agric. Food Chem. 25, 946-953.
Umetsu, N., Mallipudi, N. M., Toia, R. F., March, R. B., and Fukuto,T. R. (1981). Toxicologic properties of phosphorothioate and relatedesters present in organophosphate insecticides. J. Toxicol. Environ.Health 7,481-497.
Warbrick, E. V., Taylor, A. M., Botchkarev, V. A., and Coleman, J. W.(1995). Rat connective tissue-type mast cell express MHC II: Up-regula-tion by IFN-G. Cell. Immunol. 163, 222-228.
Weeks, M. M., Lawson, M. A., Angerhofer, R. A., Davenpod, C. D., andPennington, N. E. (1977). Preliminary assessment of the acute toxicityof malathion in animals. Arch. Environ. Contam. Toxicol. 5, 23—31.
Xiong, S., and Rodgers, K. E. (1996). Effects of malathion metabolites ondegranulation of and mediator release by human and rat basophilic cells.Submitted for publication.
Yoffee, J. R., Taylor, D. J., and Wooley, D. E. (1985). Mast-cell productsand heparin stimulate the production of mononuclear cell factor by cul-tured human monocytes/macrophages. Biochem. J. 230, 83.
Dow
nloaded from https://academ
ic.oup.com/toxsci/article/31/2/252/1720614 by guest on 18 February 2022