effects of crayfish introduction on otter lutra lutra food in the doñana national park, sw spain
TRANSCRIPT

Biological Conservation 42 (1987) 153 159
Effects of Crayfish Introduction on Otter Lutra lutra Food in The Dofiana National Park, SW Spain
Miguel Delibes & Isabel Adrifin
Est. Biol. Dofiana, CSIC, Apartado 1056, Sevilla 41080, Spain
(Received 2 April 1987; accepted 26 April 1987)
A BSTRA CT
American red swamp crayfish were introduced into the Guadalquivir marismas, close to the Dohana National Park, in 1974, and are now thriving. Food habits of otters in a small stream by the marsh were compared before the arrival of cra)fish and three,five and eight years later. Fish were their staple prey at this locality in 1976, with amphibians and insects as secondary foods. In 1979 crayfish were the second most important food and the frequency of .fish in the spraints decreased significantly. Later the role offish did not change, but crayfish became the most common prey category in 1981 and 1984. The occurrence of amphibians decreased statistically from 1979 (probably because of their scarcity in the habitat, due to high mortality in fyke nets) and that of insects increased (probably because they are found more often by the predator when searching for crayfish). We conclude that at present crayfish are a sign(ficant food Jbr most of the marsh predators, but their spread and the related commercial fishing affect the faunistic diversity of the marismas.
INTRODUCTION
The introduction of exotic species is usually associated with risks of environmental disturbance; introduced species have frequently reached plague proportions, have disturbed trophic webs and/or exterminated some native species (Jungius, 1985). American red swamp crayfish (Procambarus spp., mainly P. clarki) were introduced into the Guadalquivir marismas,
153 Biol. Conserv. 0006-3207/87/$03'50 ~) Elsevier Applied Science Publishers Ltd, England, 1987. Printed in Great Britain

154 Miguel Delibes, Isabel Adrihn
40km north of the Dofiana National Park (SW Spain), in 1974, and their effects on the Park ecosystems are becoming evident. In this paper we report the changes in the diet of an aquatic and endangered predator, the otter Lu t ra lutra, subsequent to crayfish introduction.
Much information is available on otter dietary changes with seasonal or spatial changes in food abundance or vulnerability (e.g. Erlinge, 1967; Wise et al., 1981), but as far as we know, information is lacking on the trophic response of an otter population to the dramatic appearance in its habitat of a new potential prey species.
STUDY AREA AND NOTES ON CRAYFISH POPULATION
The Dofiana National Park is situated on the west bank of the mouth of the River Guadalquivir (approximately 37°00'N, 6°30'W) and occupies 700km 2 (including the peripheral zone of protection). The marshes or marismas, which are usually flooded from October-November to June-July, cover about 55% of this area, 30% being Mediterranean maquis and 15% sand dunes with pine forests. A detailed description of the area can be found in Valverde (1958) and Aguilar-Amat et al. (1979). The climate is Mediterranean with a slight Atlantic influence. The summers are warm and dry and the winters mild and wet. Average annual rainfall is 500-600 mm, 87% falling from October to April.
Otter faeces (spraints) were collected at two localities in the Park: the lower stretch of Arroyo Rocina (AR) and the Lucio Bolin (LB). The former is a small tributary of the marismas and the second an artificial pond situated 14 km south by the border of the marsh. They have been described elsewhere (Adrian & Delibes, 1987).
In 1974 red swamp crayfish were introduced on private property in the Guadalquivir marismas, 40 km north of the Park. From there they began a spontaneous and fishermen-induced spread. In 1976 they reached Arroyo Rocina. By the end of 1979 they occupied most of the marismas, except for the southern saltier zones. A summary of this process can be found in Algarin-V61ez (1981). In 1983-84 crayfish reached Lucio Bolin.
The abundance of crayfish has resulted in an important fishery. According to Molina-V~zquez (1984), five metric tonnes (mt) of crayfish were captured in 1976 (only individuals bigger than 8 cm were considered), 15 mt in 1977, 50 mt in 1978, 150 mt in 1979, 510 mt in 1980, 1700 mt in 1981 and 3000 mt in 1982. We lack data for the last four years, but it seems that an asymptotic trend (even a decrease) is likely. The available data fit a sigmoid curve which can be considered as representative of the crayfish population growth in the whole marismas.

Introduced crayfish and the otter 155
METHODS
Using standard methods (e.g. Webb, 1976) we analysed 334 spraints collected in LB in 1981 (before crayfish reached the pool) and 264 spraints collected in AR in 1979 (AR79, n = 54), 1981 (AR81, n = 167) and 1984 (AR84, n = 43). Although small, sample sizes were big enough to allow statistical comparisons. Due to the small sample sizes and small seasonal variation in the Dofiana otter food (Adri~.n & Delibes, 1987) we pooled spraints from each locality for all seasons.
For purposes of comparison, otter food in LB will be considered as representative of the situation in AR before the arrival of crayfish to the area (i.e. 1976), although this may not be exactly correct, mainly because of the lower fish species diversity in the artificial pond compared with AR (Hernando, 1978).
Results are presented as relative frequency of occurrence (i.e. number of occurrences of each food × 100 divided by the total number of spraints) and proportion of occurrence (i.e. number of occurrences of each food x 100 divided by the total number of occurrences of all the foods in all the spraints). These methods do not accurately reflect the weight of ingested material (e.g. Wise et al., 1981) but they are quick and simple and supply a good guide to the relative importance of the various items in the diet of otters (Erlinge, 1968; Rowe-Rowe, 1977).
Frequencies of the same food in different periods were compared by chi- square tests. Prey species were lumped into taxonomic groupings to facilitate statistical analysis. These groups were mammals, birds, reptiles, amphibians, fish, crayfish and insects. Only large insects (diving beetles such as Hydrous, Dytiscus, etc., and the Orthoptera Gryllotalpa) were considered as otter prey, the remaining ones (small beetles, ants, small larvae) probably being indirect prey (Mason & Macdonald, 1986). Trophic diversity was assessed by applying the Shannon-Weaver expression (Margalef, 1957) to the relative frequency of occurrences.
RESULTS
The trophic diversity and the relative frequency of occurrence of each food in the four samples (LB81, AR79, AR81, AR84), as well as statistical comparisons, are given in Table 1. Proportions of occurrence are shown in Fig. 1.
Fish occurred in 94% of the spraints in LB, being the staple prey for otters in this locality. This was probably also the situation in AR before the crayfish introduction. Frequency of occurrence offish decreased statistically
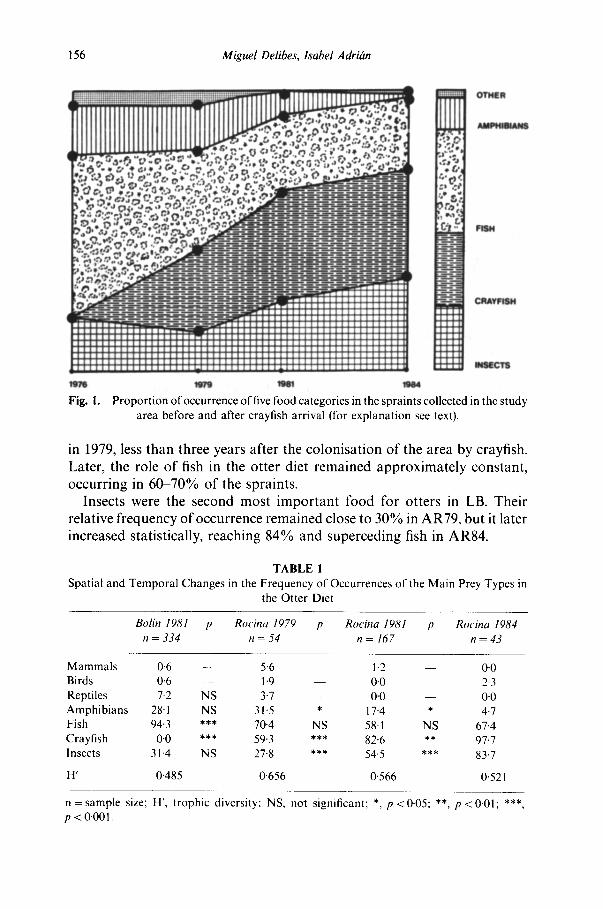
156 M i g u e l Delibes, Isabel Adr ihn
r.J . * I , ¢~ -J
DF_~ \-]
OTHER
AMPHIBIANS
FISH
CRAYFISH
1976
Fig. I.
~ INSECTS
1 9 7 9 1981 19.84
Propor t ion of occurrence of five food categories in the spraints collected in the study area before and after crayfish arrival (for explanat ion see text).
in 1979, less than three years after the colonisation of the area by crayfish. Later, the role of fish in the otter diet remained approximately constant, occurring in 60-70% of the spraints.
Insects were the second most important food for otters in LB. Their relative frequency of occurrence remained close to 30% in AR79, but it later increased statistically, reaching 84% and superceding fish in AR84.
TABLE 1 Spatial and Tempora l Changes in the Frequency of Occurrences of the Main Prey Types in
the Otter Diet
Bolin 1981 p Rocina 1979 p Rocina 1981 p Rocina 1984
n = 334 n = 54 n = 167 n = 43
Mammal s 0.6 - - 5.6 1.2 0.0 Birds 0"6 - - 1"9 0"0 --- 2"3 Reptiles 7.2 NS 3"7 - - 0.0 - - 0.0 Amphib ians 28.1 NS 31"5 * 17.4 * 4-7 Fish 94.3 *** 70"4 NS 58"1 NS 67.4 Crayfish 0"0 *** 59"3 *** 82.6 ** 97'7 Insects 31-4 NS 27.8 *** 54.5 *** 83.7
H' 0"485 0-656 0'566 0"521
n = s a m p l e size; H', t rophic diversity; NS, not significant; *, p < 0 " 0 5 : **, p <0"01; *** p < 0"001.

Introduced crayfish and the otter 157
Amphibians occurred in approximately 30% of the samples from LB and AR79. Later, their frequency decreased statistically, and was only 5% in AR84.
Obviously crayfish did not appear in the spraints from LB. In AR79 they were the second most important prey, following fish, and in AR81 and AR84 the most common, appearing in 98% of the spraints collected in 1984. Frequency of crayfish occurrence increased statistically in all the pairs of consecutive samplings.
The remaining food items were unimportant, although there seems to have been a decrease in the frequency of reptiles. Trophic diversity increased following crayfish introduction, decreasing later as crayfish became the dominant item.
DISCUSSION
Changes in the food of otters in Dofiana following the spread of crayfish have been demonstrated. Two related points emerging from these results can be discussed: (1) the fast response of otters to crayfish availability and the changes in the role of other food items; and (2) the growing importance of the introduced crayfish in the trophic webs of the Dofiana National Park ecosystem.
Otters did not find crayfish in the study area and its surroundings before 1974. In spite of this, they learned in a short time to recognise crayfish as an attractive food. This would confirm the dietary adaptability of otters and their efficiency at shifting quickly to the most abundant and/or vulnerable prey in their aquatic habitat.
Fish were the only prey item whose frequency of occurrence decreased statistically in 1979, suggesting that the rising predation of otters upon crayfish was accompanied by a decrease in their interest for the former principal prey (i.e. fish). On the other hand, other prey (i.e. insects, amphibians) would be eaten when found, without any specific searching, and their frequency of occurrence did not change in the early years. Later, the importance offish stabilised, probably because they are preferred to crayfish when both prey are available (Erlinge, 1968).
The role of amphibians (and maybe of reptiles) in the otter diet decreased from 1979 to 1981 and 1984, probably as the consequence of a marked decrease in the numbers of these animals in the habitat. This reduction has been observed by naturalists and fishermen in all areas where crayfish exist and it is generally accepted in the region. Two reasons are given (1) predation of eggs and tadpoles by crayfish, and (2) high mortality of adult amphibians and aquatic reptiles in the fyke nets used by fishermen. Probably both factors are operating, but the second seems to be more important, as red

! 58 Miguel Delibes, Isabel Adriim
swamp crayfish are mainly plant-eaters in the area (Librero-Ruiz, 1981) and significant mortalities of non-target animals in fyke nets have been noted (Molina-Vfizquez, 1984).
The growing role of large insects in the otter diet could be explained if otters foraging for crayfish encountered insects more often than when hunting fish. If this hypothesis were correct, as the association of crayfish and insects in the otter diet found in other places by Adrifin (1985) seems to confirm, it would mean that insects are usually captured opportunistically.
The role of crayfish in the trophic webs of Dofiana and its implications for the conservation of otters and other predators and for the management of the National Park must also be considered.
At present, crayfish are the main food in Dofiana not only for otters but also for many other aquatic predators (e.g. the gull-billed tern Gelochelidon nilotica (Costa, 1984), the white stork Ciconia ciconia (Rubio-Garcia et al., 1983) and they are frequently preyed on by almost all predators in the area. We have recorded crayfish in the diet of the fox Vulpes vulpes, badger Meles meles, polecat Mustela putorius, genet Genetta genetta and mongoose Herpestes ichneumon, most of the herons, ducks and Rallidae, as well as some raptors, such as the black kite Milvus migrans and tawny owl Strix aluco. Amphibians, aquatic reptiles and probably fish (J. A. Hernando, pers. comm.) seem to be decreasing, so that marsh predators depend more and more on invertebrates as a food source. Under these conditions, the hypothetical elimination of the introduced crayfish, as has been debated (Junta de Andalucia, 1981), could be catastrophic in the short term for otters and other predators in the area. On the other hand, the decreasing faunistic diversity in the marismas following the crayfish introduction, and the resultant intensive commercial fishing, strongly demand a control of the crayfish populations and of fishing volume and methods.
ACKNOWLEDGEMENTS
We are grateful to Drs S. M. Macdonald, C. F. Mason, S. Moreno and E. Collado, N. Bustamante and R. Laffitte for their support and criticisms of the manuscript. Financial support was obtained from the CSIC-CAICYT (Project 944).
REFERENCES
Adrifin, M. I. (1985). Distribuci6n y alimentacion de la nutria, Lutra lutra (L.) en Andalucia Occidental. Tesina de licenciatura, University of Oviedo.

Introduced crayfish and the otter 159
Adri~,n, M. I. & Delibes, M. (in press). Food habits of the otter Lutra lutra in two habitats of the Dofiana National Park, SW Spain. J. Zool., Lond., 212, 399406.
Aguilar-Amat, J., Montes del Olmo, C., Ramirez-Diaz, L. & Torres-Martinez, A. (1979). Parque Nacional de Dohana. Mapa ecol6gico. Madrid, ICONA, Ministerio de Agricultura.
Algarin-V61ez, S. (1981). Problem~tica y perspectivas de la introducci6n del cangrejo. In El cangrejo rojo de la marisma. Jornadas de estudio, ed. by Junta de Andalucia, 25-31. Madrid, Publicaciones Agrarias.
Costa, L. (1984). Alimentaci6n de la pagaza piconegra Gelochelidon nilotica en las M arismas del Guadalquivir. Dohana, A cta Vert., 11(2), 185-95.
Erlinge, S. (1967). Food habits of the fish otter Lutra lutra L. in south Swedish habitats. Viltrevy, 4, 371~443.
Erlinge, S. (1968). Food studies on captive otters Lutra lutra. Oikos, 19, 259--70. Hernando, J. A. (1978). Estructura de una comunidad de peces de la Marisma del
Guadalquivir. Doctoral thesis, University of Seville. Jungius, H. (1985). Prospects for re-introduction. In Advances in animal
conservation, ed. by J. P. Hearn & J. K. Hodges, 47 55 (Zool. Soc. Lond. Symp. no. 54). Oxford, Clarendon Press.
Junta de Andalucia (1981). El cangrejo rojo de la marisma. Jornadas de estudio. Madrid, Publicaciones Agrarias.
Librero-Ruiz, M. (1981). Biologia y pesca del cangrejo. In El cangrejo rojo de la marisma. Jornadas de estudio, ed. by Junta de Andalucia, 17-23, Madrid, Publicaciones Agrarias.
Margalef, R. (1957). La teoria de la informaci6n en ecologia. Mem. R. Acad. Cien. Art. Barcelona, 32, 373~449.
Mason, C. F. & Macdonald, S. M. (1986). Otters, ecology and conservation. Cambridge, University Press.
Molina-Vhzquez, F. (1984). La pesca del cangrejo rojo americano y su influencia en el entorno del Parque de Dofiana. Rev. Est. Andaluces, 3, 151 60.
Rubio-Garcia, J. C., Rodriguez de los Santos, M. & Santo-Rosa, R. (1983). Reproduction de la cigogne blanche Ciconia ciconia dans les Marismas du Guadaiquivir. Alauda, 51,251-8.
Rowe-Rowe, D. T. (1977). Food ecology of otters in Natal, South Africa. Oikos, 2g, 210-19.
Valverde, J. A. (1958). An ecological sketch of the Coto Dofiana. British Birds, 51, 1-23.
Webb, J. B. (1976). Otter spraint analysis. London, Mammal Society. Wise, M. H., Linn, I. J. & Kennedy, C. R. (1981). A comparison of the feeding
biology of mink Mustela vison and otter Lutra lutra. J. Zool., Lond., 195, 181 213.