effects of carbonsources antibiotic resistance …aac.asm.org/content/15/1/59.full.pdf · effects...
TRANSCRIPT

Vol. 15, No. 1ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Jan. 1979, p. 59-660066-4804/79/01-0059/08$02.00/0
Effects of Carbon Sources on Antibiotic Resistance inPseudomonas aeruginosa
ROBERT S. CONRAD,* R. GARY WULF, AND DAMONDA L. CLAYDepartment ofMedical Biology, The Oklahoma College of Osteopathic Medicine and Surgery, Tulsa,
Oklahoma 74101
Received for publication 26 October 1978
The metabolism of branched-chain amino acids, branched-chain acyl deriva-tives, D-glucose, L-glutamate, and Mueller-Hinton medium was investigated todetermine their effects on the growth, lipid composition, and antibiotic suscepti-bility of Pseudomonas aeruginosa. The unsaturated fatty acid content of thereadily extractable lipids was altered by growth on selected branched-chain aminoacids and their acyl derivatives. Bacteria grown on branched-chain acyl deriva-tives became more susceptible to polymyxin B and colistin. The effect acylderivatives had on increasing susceptibility was also manifest in mixed mediawhich contained both an acyl derivative and a carbon source which did notincrease susceptibility. Growth on branched-chain amino acids gave mixed resultswhich were dependent on a number of factors, including unique manifestations ofindividual amino acids, growth conditions, and availability of other carbonsources. The cultural conditions which altered susceptibility to polymyxin anti-biotics did not correlate with similar effects on susceptibility to carbenicillin andgentamicin. An adaptive resistance to polymyxin B was observed when the solecarbon source was D-glucose or L-glutamate.
The recognition of Pseudomonas aeruginosaas a major opportunistic pathogen in compro-mised patients has given increased relevance tostudying its renowned lack of susceptibility toantibiotics. The pseudomonads differ from mostbacteria in that antibiotic-inactivating enzymesare not the primary mechanism of resistance.There is substantial evidence that pseudomo-nads are refractory to most chemotherapeuticregimens by excluding the antimicrobial agentsfrom the target site (4). This intrinsic lack ofsusceptibility presents a formidable therapeuticproblem because the antibiotic must first cir-cumvent a sophisticated exclusion mechanismbefore reaching its site of action. Although pseu-domonads may acquire antibiotic resistancefrom other species at a low frequency (15), theoccurrence of resistant strains which lack anti-biotic-inactivating enzymes indicates that exclu-sion is the primary mechanism of resistance (4).The polymyxins are one of the few groups ofantibiotics that can be used successfully againstpseudomonal infections. This success is attrib-uted to its detergent-like mechanism of actionwhich attacks the protective envelope directly(16). It is then logical that changes in envelopecomponents such as phospholipids which inter-act with polymyxins would elicit changes in sus-ceptibility to polymyxin antibiotics.A number of workers have demonstrated that
microbial lipids may be altered by the carbon
sources provided to the microbe (7, 16). Thesedifferences in cellular components no doubt con-tribute to differences in antibiotic susceptibilityamong different species and strains. Pseudomo-nads possess extraordinary nutritional capabili-ties which enable them to catabolize a diversearray of organic compounds, such as thebranched-chain amino acids and their resultantcatabolites (14). This metabolic prowess facili-tates studying the effects ofmany carbon sourcesnot metabolized by species lacking this physio-logical versatility. If these metabolic conditionscan cause alterations in the lipid composition ofthe protective envelope of P. aeruginosa, wethought it reasonable that changes in antibioticsusceptibility may result.The purpose of this study was to determine
what effects growth on selected carbon source(s)would have on the susceptibility of P. aerugi-nosa to antibiotics commonly used in the treat-ment of pseudomonal infections. This reportdescribes a series of experiments in whichbranched-chain amino acids and their acyl de-rivatives were used as carbon sources to deter-mine their effects on growth, lipid composition,and antibiotic susceptibility in P. aeruginosa.
MATERIALS AND METHODSOrganisms. P. aeruginosa 39229, a strain resistant
to tobramycin, gentamicin, and carbenicillin and sus-ceptible to polymyxin B and colistin, was obtained
59
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

60 CONRAD, WULF, AND CLAY
from the Center for Disease Control, Atlanta, Ga. P.aeruginosa 015, a clinical isolate furnished by Univer-sity Hospital, Oklahoma City, Okla., was susceptibleto these same antibiotics. Stock cultures were main-tained on Mueller-Hinton slants and transferred every3 weeks.Growth conditions. Mueller-Hinton agar medium
(Difco Laboratories; lot 645524) and Mueller-Hintonbroth (Difco; lot 6483831) were used as controlsthroughout this study. All references to resistance andsusceptibility were made relative to values obtainedwith Mueller-Hinton medium. Jacobson medium wasused as the base medium when individual carbonsources were used to determine their effects on sus-ceptibility (10). This was a defined medium in whichthe final concentrations of trace elements and nitrogensource (NH4Cl) were not growth limiting. Final pHwas 6.8 to 7.0. Carbon source concentration in definedmedium was 25 mM with the exception of isoleucine-leucine-valine (ILV) and DL-2-methylbutyrate-isoval-erate-isobutyrate (MII) media, in which each respec-tive component was 10 mM. Solid medium was pre-pared from the defined medium by the addition of0.15% agar (Difco).
Doubling times for individual carbon sources werecalculated from 50-ml samples grown in 250-ml sidearm flasks. Doubling times for mixtures of carbonsources were determined from 40-ml samples grown in250-ml side arm flasks. Controls for the mixed mediumcontained 40 ml of either D-glucose or Mueller-Hintonmedium. Test flasks contained 20 ml of the controlmedium plus 20 ml of supplemental carbon medium.Inoculum was 1 ml of an overnight culture grown incontrol medium. Optical density for all experimentswas read at 660 nm with a Bausch and Lomb Spec-tronic 20 spectrophotometer. Aeration was providedby vigorous shaking in a Lab-Line Environ-Shaker3597-2 shaker. All growth was accomplished at 37°C.
Lipid analyses of cells grown on different car-bon sources. Approximately 3 g (wet weight) of P.aeruginosa 015 cells grown on either L-vahne, L-iso-leucine, DL-2-methylbutyrate, or Mueller-Hinton me-dium were used in each extraction. Growth was mon-itored spectrophotometrically and allowed to reachearly stationary phase. Cells were harvested by cen-trifugation and washed twice in 50 mM phosphatebuffer, pH 7.5.
Readily extractable lipids were extracted andwashed by the method of Folch et al. (9). The ex-tracted lipids were transmethylated with boron tri-fluoride methanol reagent (Applied Science Labora-tories) by the procedure of Morrison and Smith (13).The fatty acid methyl esters were extracted by hexaneand separated from the nonesterified fatty acids byusing thin-layer chromatography. The methylatedfatty acids were then dissolved and stored in cyclo-hexane. Identification of fatty acids was accomplishedwith a Hewlett-Packard 5840A gas chromatographequipped with a flame ionization detector. The glasscolumn (6 feet [ca. 183 cm] by 4 mm) was packed with10% SP-2330 on 100/120 Chromosorb. The tempera-tures of the injection port and detector were 200 and225°C, respectively. The column temperature was pro-grammed to run for 4 min at 100°C, followed by a10°C increase per min to a final temperature of 2000C.
The flow rate of the nitrogen carrier gas was 40ml/min. Bacterial fatty acids were identified by com-paring peak retention times with fatty acid standards.Disk diffusion susceptibilities. Disk diffusion
susceptibility tests were performed by a modificationof the method of Bauer et al. (2). Overnight culturesin Mueller-Hinton broth were transferred to freshmedium (1 ml/100 ml). Growth was followed spectro-photometrically to mid-exponential growth phase atwhich time the cultures were diluted in physiologicalsaline to an optical density of 0.10. Bacterial lawnswere prepared by spreading 0.1-ml samples with aglass rod. Analysis with a Coulter Channelyzer indi-cated that the inoculum contained 7 x 106 cells. Zonesizes were read after 24- and 48-h incubations at 37°C.This growth period was longer than what normallywould be expected because growth on many of thecarbon sources used in this study was not sufficientfor susceptibility determinations after 24 h of growth.Zone diameters on medium supporting faster growthdid not significantly change between 24 and 48 h. DifcoDispens-O/Discs of the following concentrations wereused: carbenicillin, 100 ,ug; colistin, 10 ug; gentamicin,10 jug; and polymyxin B, 200 U. Data regarding zonediameters in this study were the mean of 6 to 10separate experiments.
Determination of MIC. Minimal inhibitory con-centration (MIC) was measured by a modification ofthe method of Washington and Barry (17). Antibioticswere dissolved in double-distilled water at a concen-tration of 1 mg/ml. All solutions were filter-sterilizedthrough membrane filters (0.45 M; Millipore Corp.)and frozen until used. Each antibiotic was seriallydiluted in growth medium for a final volume of 0.5ml/tube. Overnight cultures of the test strain weretransferred to fresh Mueller-Hinton broth (1 ml/100ml of fresh medium). Growth was monitored to mid-exponential growth phase (optical density, 0.70 to0.90). Inoculum for the MIC determinations was pre-pared by diluting these fresh cultures to an opticaldensity of 0.05 in the same medium in which theantibiotics had been diluted. The diluted antibioticsolutions were brought to a volume of 1.0 ml by theaddition of 0.5 ml of this inoculum. The inoculumcontained 1.6 x 107 cells as indicated by a CoulterChannelyzer.
This protocol was somewhat modified for thoseexperiments which measured susceptibility to antibi-otics with multiple carbon sources. The antibiotic wasdiluted in the growth medium as previously described.However, in these experiments the inoculum was pre-pared by diluting the freshly grown cultures in mediumcontaining another carbon source (supplemental me-dium). Equal volumes of growth medium and supple-mental medium (0.5 ml each) resulted in a final con-centration of approximately 12.5 mM of each carbonsource. The MIC obtained on each carbon source wasdetermined at 24 and 48 h. All MIC values were themean of four to six separate determinations.
RESULTSThe utilization of either L-valine, L-isoleucine,
or DL-2-methylbutyrate as the sole carbonsource resulted in substantial changes in the
ANTIMICROB. AGENTS CHEMOTHER.
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

CARBON SOURCES AND P. AERUGINOSA RESISTANCE
extractable lipids of P. aeruginosa 015 (Table1). The comparative fatty acid profile of cellsgrown in Mueller-Hinton medium indicated thatthese differences were both quantitative andqualitative. Cells grown on L-isoleucine, L-valine,and DL-2-methylbutyrate had peaks correspond-ing to pentadecanate (C15) with a relative reten-tion time (RRT) of 0.95. All three of these car-bon sources are catabolized to propionyl-coen-zyme A, which might enhance the possibility ofincreased odd-chain fatty acid synthesis. Thesepossibilities in substrate ambiguity would beaccentuated by growth conditions in which alllipids are derived from odd-chain precursors,such as propionyl-coenzyme A, which are nor-mally found only in minute quantities. Palmitate(C16) appeared to be synthesized at a constantrate regardless of carbon source (RRT, 1.000).Mueller-Hinton medium-grown cells had higherconcentrations of palmitoleate (C06:1; RRT, 1.04),and oleate (Ci8s:; RRT, 1.17) than did cells grownon the other carbon sources. Cells grown on L-valine, L-isoleucine, and DL-2-methylbutyratehad higher levels of linoleate (018:2; RRT, 1.25)than did Mueller-Hinton medium-grown cells.Effects ofcarbon source on susceptibility
to antibiotics. The phenotypic expression ofsusceptibility and resistance to antibiotics in P.aeruginosa was influenced significantly by thecarbon source. The effects that growth on singleor multiple carbon source(s) had on susceptibil-ity to antibiotics in strain 39229 are illustratedin Table 2. The MICs of polymyxin B and col-istin indicated that growth in D-glucose, L-glu-tamate, and ILV medium enabled cells to resisthigher concentrations of these antibiotics thandid growth in a rich medium, such as Mueller-Hinton medium. Conversely, growth in L-isoleu-cine, L-valine, isovalerate, isobutyrate, and MIImedium rendered the cells more vulnerable tothe action of polymyxin B than did growth inMueller-Hinton medium. The MICs for genta-micin remained greater than 500 ,ug/ml in thisresistant strain regardless of growth medium.The apparent development of resistance in
strain 015 was related to exposure time betweenbacteria and antibiotic (Table 3). A comparisonof MICs at 24 h indicated that cells grown withD-glucose, L-glutamate, and ILV medium weremore susceptible to polymyxin B than were cellsgrown in Mueller-Hinton medium. After 48 h ofgrowth, the MICs of polymyxin B in these samecultures were 3 to 11 times higher than theirMueller-Hinton medium counterparts. This ap-parent increase in resistance noted in these me-dia was not due to slower growth per se, asshown by the sensitizing effects of acyl deriva-tives which support an even slower growth rate(Table 4). Subsequent investigations found this
TABLE 1. Effects of carbon source on readilyextractable lipids of P. aeruginosa 015
Area % with the following as carbon source'RTTa Mueller-Hin L-V ine L-Isoleu- DL-2-Methyl-
ton medium cine butyrate
0.76 0.09 0.05 0.210.86 0.27 1.46 0.30 0.590.89 1.22 0.95 0.54 1.000.95 0.62 2.28 3.97 2.650.97 0.35 2.91 0.64 0.261.00 28.94 25.85 27.56 26.261.04 19.14 14.69 9.73 8.021.06 2.65 1.591.09 2.00 4.36 4.86 9.251,12 1.20 1.19 1.16 1.991.17 40.58 32.64 36.72 21.491.25 4.64 8.17 11.80 23.70
a RTT was calculated by dividing peak retentiontime by retention time of a methyl palmitate standard.Standard methyl esters were chromatographed underthe same conditions which yielded RTTs as follows:methyl pentadecanate, 0.95; methyl palmitate, 1.00;methyl palmnitoleate, 1.04; methyl oleate, 1.17; andmethyl linoleate, 1.25.
"Average of three separate determinations. Thecarbon source was the sole source of carbon and en-ergy. Cells were harvested at an optical density of 0.90to 1.10.
TABLE 2. Relationship between carbon source andsusceptibility to antibiotics in P. aeruginosa 39229
MIC at 48 h" (,Lg/ml)Growth mediuma Gentami
Polymyxin B Colistin cmin
Mueler-Hinton 13 27 <500D-Glucose 167t 291T <500L-Glutamate 375T 250T <500L-Isoleucine 21 10 <500DL-2-Methylbutyr- 0.54 0.54 <500
ateL-Leucine 20 41 <500Isovalerate 11 21 <500L-Valine 31 13 <500Isobutyrate 0.54 0.51 <500ILV mixture 156T 217T <500MII mixture 0.54 0.54 <500
a The growth medium was the sole carbon source.b All MICs were compared with those obtained in
Mueller-Hinton broth, which served as the controls.Differences in susceptibility to each antibiotic whichwere greater than twofold for each respective antibi-otic when another carbon source was used are indi-cated by the following: T, increase in resistance; 4,increase in susceptibility.
resistance to be adaptive and transitory in na-ture. Bacteria subcultured in various other me-dia from these apparently resistant cultures ex-hibited the same susceptibility to antibiotics as
61VOL. 15, 1979
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
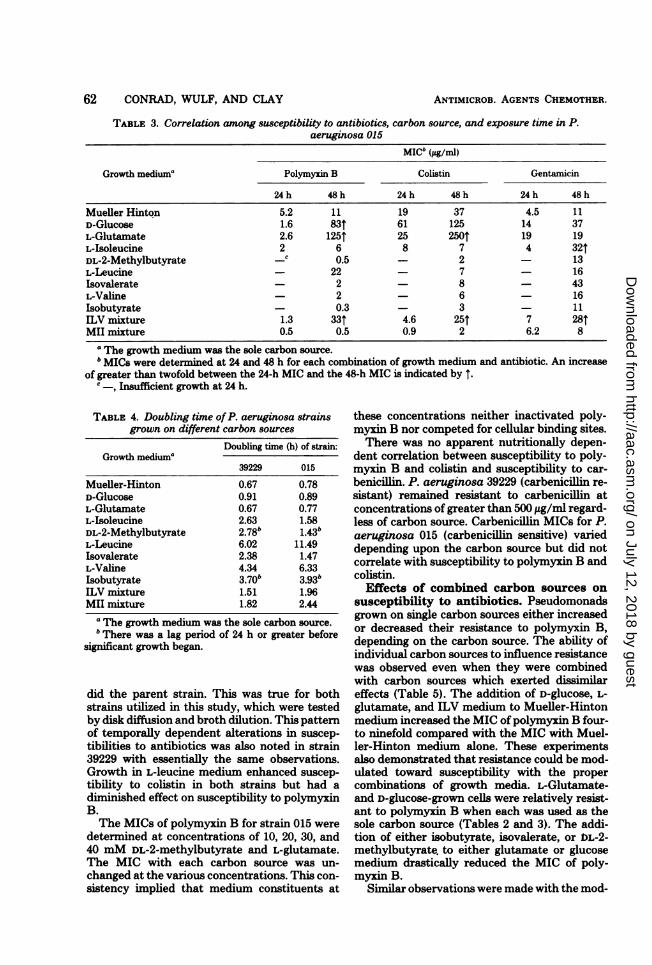
TABLE 3. Correlation among susceptibility to antibiotics, carbon source, and exposure time in P.aeruginosa 015
MICb (ug/m)
Growth mediuma Polymyxin B Colistin Gentamicin
24 h 48 h 24 h 48 h 24 h 48 h
Mueller Hinton 5.2 11 19 37 4.5 11D-Glucose 1.6 83t 61 125 14 37L-Glutamate 2.6 125T 25 2501 19 19L-Isoleucine 2 6 8 7 4 32tDL-2-Methylbutyrate -c 0.5 - 2 - 13L-Leucine - 22 - 7 - 16Isovalerate - 2 - 8 - 43L-Valine - 2 - 6 - 16Isobutyrate - 0.3 - 3 - 11ILV mixture 1.3 33T 4.6 25T 7 281MII mixture 0.5 0.5 0.9 2 6.2 8
aThe growth medium was the sole carbon source.b MICs were determined at 24 and 48 h for each combination of growth medium and antibiotic. An increase
of greater than twofold between the 24-h MIC and the 48-h MIC is indicated by 1.c -, Insufficient growth at 24 h.
TABLE 4. Doubling time of P. aeruginosa strainsgrown on different carbon sources
Doubling time (h) of strain:Growth mediuma
39229 015
Mueller-Hinton 0.67 0.78D-Glucose 0.91 0.89L-Glutamate 0.67 0.77L-Isoleucine 2.63 1.58DL-2-Methylbutyrate 2.78b 1.43bL-Leucine 6.02 11.49Isovalerate 2.38 1.47L-Vahne 4.34 6.33Isobutyrate 3.70b 3.93bILV mixture 1.51 1.96MII mixture 1.82 2.44
a The growth medium was the sole carbon source.b There was a lag period of 24 h or greater before
significant growth began.
did the parent strain. This was true for bothstrains utilized in this study, which were testedby disk diffusion and broth dilution. This patternof temporally dependent alterations in suscep-tibilities to antibiotics was also noted in strain39229 with essentially the same observations.Growth in L-leucine medium enhanced suscep-tibility to colistin in both strains but had adiminished effect on susceptibility to polymyxinB.The MICs of polymyxin B for strain 015 were
determined at concentrations of 10, 20, 30, and40 mM DL-2-methylbutyrate and L-glutamate.The MIC with each carbon source was un-changed at the various concentrations. This con-sistency implied that medium constituents at
these concentrations neither inactivated poly-myxin B nor competed for cellular binding sites.There was no apparent nutritionally depen-
dent correlation between susceptibility to poly-myxin B and colistin and susceptibility to car-benicillin. P. aeruginosa 39229 (carbenicillin re-sistant) remained resistant to carbenicillin atconcentrations of greater than 500 Ig/ml regard-less of carbon source. Carbenicillin MICs for P.aeruginosa 015 (carbenicillin sensitive) varieddepending upon the carbon source but did notcorrelate with susceptibility to polymyxin B andcolistin.Effects of combined carbon sources on
susceptibility to antibiotics. Pseudomonadsgrown on single carbon sources either increasedor decreased their resistance to polymyxin B,depending on the carbon source. The ability ofindividual carbon sources to influence resistancewas observed even when they were combinedwith carbon sources which exerted dissimilareffects (Table 5). The addition of D-glucose, L-glutamate, and ILV medium to Mueller-Hintonmedium increased the MIC ofpolymyxin B four-to ninefold compared with the MIC with Muel-ler-Hinton medium alone. These experimentsalso demonstrated that resistance could be mod-ulated toward susceptibility with the propercombinations of growth media. L-Glutamate-and D-glucose-grown cells were relatively resist-ant to polymyxin B when each was used as thesole carbon source (Tables 2 and 3). The addi-tion of either isobutyrate, isovalerate, or bL-2-methylbutyrate. to either glutamate or glucosemedium drastically reduced the MIC of poly-myxin B.
Similar observations were made with the mod-
62 CONRAD, WULF, AND CLAY ANTIMICROB. AGENTS CHEMOTHER.
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

CARBON SOURCES AND P. AERUGINOSA RESISTANCE
TABLE 5. Effects of carbon source combinations on P. aeruginosa 015 susceptibility to polymyxin B andcolistin
Supplemental medium'Mueller-HintonD-GlucoseL-GlutamateL-LeucineIsovalerateL-IsoleucineDL-2-MethylbutyrateL-ValineIsobutyrateILV mixtureMII mixtureL-GlutamateL-ValineIsobutyrateL-LeucineIsovalerateL-IsoleucineDL-2-MethylbutyrateD-GlucoseL-ValineIsobutyrateL-LeucineLsovalerateL-IsoleucineDL-2-MethylbutyrateMuller-HintonD-GlucoseL-GlutamateL-ValineIsobutyrateD-GlucoseL-ValineIsobutyrate
MIC at 48 hb (g/ml)6
3758205
24T3
63T3
39t3
12535419441
12541324271
4741
4721
47375T500T12524
250375161
'Each tube contained 0.5 ml each of base medium and supplemental medium prepared as described in thetext.
b All MICs were compared with susceptibility demonstrated when base and supplemental media were identical(Mueller-Hinton, L-glutamate, D-glucose). Differences in MICs of greater than twofold are compared with theircontrols as indicated by the following: t, increase in resistance; 4, increase in susceptibility.
ulation of susceptibility to colistin. The additionof D-glucose, L-glutamate, and L-valine to Muel-ler-Hinton medium increased resistance to col-istin (Table 5). The addition of isobutyrate toMueller-Hinton medium resulted in an increaseof susceptibility to colistin. Colistin resistance inD-glucose medium was increased by L-valine anddecreased by isobutyrate.The role of branched-chain amino acids in
influencing susceptibility to polymyxin B wassomewhat erratic, compared with the effects oftheir acyl derivatives. The addition of either L-isoleucine, L-leucine, or L-valine to either Muel-ler-Hinton or D-glucose medium increased re-sistance (Table 5). The addition of L-leucine andL-isoleucine had little effect in L-glutamate me-dium, although L-valine decreased the MIC ofpolymyxin B. This was in sharp contrast to theutilization of L-isoleucine and L-vahne as the
sole carbon source, which increased the suscep-tibility of the cell to polymyxin B.
It was noted that the individual branched-chain amino acids had dissimilar effects on sus-ceptibility to antibiotics depending on whetherthey were (i) the sole source of carbon, (ii) asupplemental carbon source, or (iii) a combina-tion carbon source (e.g., ILV medium). This wasnot the case for the branched-chain acyl acidswhich increased cellular susceptibility to poly-myxin B and colistin regardless of the experi-mental protocol.
Effects of carbon source as measured bydisk diffusion. Determination of susceptibilityto antibiotics by the disk diffusion techniqueindicated that susceptibility and resistance toantibiotics on solid medium was influenced bythe carbon source in a manner analogous to thatobserved in liquid cultures. Cells grown with
Antibiotic
Polymyxin B
Colistin
Base medium'Mueller-Hinton
L-Glutamate
D-Glucose
Mueller-Hinton
D-Glucose
VOL. 16, 1979 63
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

64 CONRAD, WULF, AND CLAY
branched-chain amino acids, branched-chainacyl acids, or combinations thereof were moresusceptible to polymyxin B and colistin thanwere cells grown on Mueller-Hinton agar. Thezones of inhibition for polymyxin B ranged from13 mm for Mueller-Hinton agar to 36 mm forisobutyrate medium. Other representative zonesincluded: D-glucose, 16 mm; L-glutamate, 15 mm;L-isoleucine, 20 mm; DL-2-methylbutyrate, 23mm; L-leucine, 28 mm; isovalerate, 24 mm; L-valine, 28 mm; ILV mixture, 18 mm; and MIImixture, 23 mm. The response to colistin resem-bled that of polymyxin B on all media. Therewere no significant differences between the re-sponses of P. aeruginosa 015 and 39229 to eitherpolymyxin B or colistin. The adaptive resistanceto the polymyxins noted in liquid media between24 and 48 h was not observed on solid media.Gentamicin resistance in strain 39229 (carben-
icillin and gentamicin resistant) was not alteredby the carbon source in solid medium, althoughthe branched-chain acyl derivatives appeared toenhance resistance to some degree. Susceptibil-ity testing for carbenicillin in strain 39229 re-sulted in confluent growth on all media. Suscep-tibility to gentamicin in strain 015 was not influ-enced by the carbon source and did not correlatewith susceptibility or resistance to polymyxin Band colistin. Carbenicillin susceptibility testingindicated that P. aeruginosa 015 was slightlymore susceptible when the carbon source wasD-glucose, L-isoleucine, DL-2-methylbutyrate, L-
leucine, isovalerate, L-valine, and isobutyrate,compared with susceptibility on Mueller-Hintonmedium. Susceptibilities to carbenicillin were
unchanged when the growth medium was L-glu-tamate, ILV, and MII.Influence of carbon source on growth
rates. The doubling times of both strains were
less than 1 h when the carbon source was Muel-ler-Hinton medium, D-glucose, or L-glutamate(Table 4). Doubling times were much slowerwhen either L-leucine or L-valine was the solecarbon source. The cells grew on all three of thebranched-chain acyl derivatives, but a lag periodof 24 h or longer before exponential growthcommenced was observed when the carbonsource was DL-2-methylbutyrate or isobutyrate.L-Valine was growth inhibitory to strain 015grown in D-glucose medium but had no effectwhen added to Mueller-Hinton medium (Fig. 1).L-Isoleucine and L-leucine did not affect growthof strain 015 in either Mueller-Hinton or D-glu-cose medium. DL-2-Methylbutyrate inhibitedgrowth in the same media. Isovalerate and iso-butyrate inhibited growth in D-glucose mediumbut had little effect when added to Mueller-Hinton medium. All inhibitions were more pro-nounced in D-glucose medium than in Mueller-Hinton medium. Combinations of either
E
(09w
cD
zw
0
0
a-
0
HOURS
FIG. 1. Effects of branched-chain amino acid andacyl derivative media on growth ofP. aeruginosa 015in D-glucose and Mueller-Hinton media. Control andtest media werepresent in equal volumes at zero time.Symbols: A- - -A, control; *..A 0e , L-valine;0-.---0, L-isoleucine; 0- L-leucine;
-0, DL-methylbutyrate; A..--.--A, isovalerate; and*-U, isobutyrate.
_
r ~~D-GLUCOSE~~~~/ss¢ f v-
2 3 4 1
Isl l
S MUELLER-HINTON -6__5_
4 2p
ANTIMICROB. AGENTS CHEMOTHER.
1%
sE7f9
4
2
I
EI4I1.
4
09
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

CARBON SOURCES AND P. AERUGINOSA RESISTANCE
branched-chain amino (ILV medium) or acyl(MII medium) acids supported growth betterthan did any of these compounds used individ-ually.
DISCUSSIONThe metabolism of branched-chain amino
acids and their acyl derivatives had profoundeffects upon P. aeruginosa susceptibility to pol-ymyxin B and colistin. The metabolism of se-lected carbon sources also had marked effectson the quantitative and qualitative fatty acidcomposition of the readily extractable lipidsfound in P. aeruginosa (Table 1). Cells grownon branched-chain amino acids as the sole car-bon source did not have the same susceptibilityto antibiotics as did cells grown on the acylanalogs. Growth on L-isoleucine and L-valine asthe sole carbon source resulted in increased sus-ceptibility to antibiotics. The addition of eitherL-isoleucine, L-leucine, or L-vahne medium toMueller-Hinton and D-glucose media increasedresistance to polymyxin B (Table 5). Cells grownon combinations of branched-chain amino acids(ILV medium) were also more resistant to pol-ymyxin B.The metabolism of branched-chain acyl deriv-
atives resulted in increased susceptibility to pol-ymyxin B and colistin regardless of experimentalprotocol. The sensitizing effects of acyl deriva-tives was expressed even when they were incor-porated into either D-glucose or L-glutamate me-dium. Either of these media increased resistancewhen used as the sole carbon source. The dissim-ilarities in the effects of branched-chain aminoacids and their acyl derivatives indicate thatthe metabolism of the amino acid and/or itsketo acid analog plays a pivotal role in determin-ing susceptibility to polymyxin B and colistin.The catabolism of branched-chain amino acidsproceeds via a common catabolic pathway inwhich the amino acids are deaminated to ketoacids, followed by decarboxylation to the acylderivatives (12). The growth of bacteria onbranched-chain amino acids as the sole carbonsource would undoubtedly result in abnormallyhigh intracellular concentrations of the respec-tive amino and keto acids. These accumulationswould not be found in cells grown on acyl deriv-atives because the oxidative decarboxylation tothe acyl derivative is an irreversible metabolicreaction.The mechanism of action of polymyxin B in-
volves interaction between antibiotic and cellu-lar phospholipids (15). Therefore, alterations incellular phospholipids should result in changesin susceptibility if the target site has been suffi-ciently modified. The growth conditions used inthis study resulted in significant changes in
chain lengths and degree of saturation. The ad-dition or elimination of double bonds to long-chain fatty acids results in significant stereo-chemical modifications to biological membranes.It is most likely not coincidental that alterationsin cellular lipids are accompanied by changes insusceptibility to antibiotics. It should be notedthat the carbon source exerting the greatestsensitizing effect (DL-2-methylbutyrate) also re-sulted in the greatest changes in fatty acid com-position. Additional proof of the relationshipbetween changes in lipids and susceptibility topolymyxin B was indicated by the fact that thecarbon source did not significantly alter suscep-tibility to either gentamicin or carbenicillin. Thisincongruity would be consistent with the diversemechanisms of action for the three classes ofantibiotics represented here. This distinctive-ness in susceptibility suggests that alteration insusceptibility to polymyxin is a separate phe-nomenon not necessarily related to metabolicvulnerability in peptidoglycan biosynthesis (car-benicillin) or protein synthesis (gentamicin).
It is not a novel idea that susceptibility toantibiotics is related to cellular lipid content.Anderess et al. noted differences in extractablelipids of pseudomonal strains both susceptibleand resistant to quatarnary ammonium com-pounds and chloramphenicol (1). Similar obser-vations have been made in the enterics with avariety of antibiotics (5). It has been establishedthat the lipid composition of gram-negative bac-teria varies as a function of other parameters,such as temperature (8), carbon source (6), pres-ence of plasmids (11), and cation concentrations(3).Our results indicate that growth on the carbon
sources used in this study resulted in quantita-tive and qualitative changes in readily extract-able lipids of P. aeruginosa. We are presently inthe process of further analyzing the effects ofthe nutritional environment on P. aeruginosa,with particular emphasis on the phospholipidsof the readily extractable lipid fraction. If theselipid alterations can be correlated to changes insusceptibility to antibiotics, it may be possibleto demonstrate a causal relationship betweenresistance or susceptibility to polymyxin B andthe metabolism of specific lipids or classes oflipids.
ACKNOWLEDGMENTSThis investigation was supported by a faculty research
grant from The Oklahoma College of Osteopathic Medicineand Surgery, Tulsa, and by Public Health Service grant1ROlAM19927-01 from the National Institute of Arthritis,Metabolism and Digestive Diseases to R.S.C.
LITERATURE CITED1. Anderess, E. A., W. E. Sandine, and R. R. Elliker.
1971. Lipids of antibiotic-sensitive and -resistant strains
65VOL. 15, 1979
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

66 CONRAD, WULF, AND CLAY
of Pseudomonas aeruginosa. Can. J. Microbiol. 17:1357-1365.
2. Bauer, A. W., W. M. M. Kirby, J. C. Sherris, and M.Turek. 1966. Antibiotic susceptibility testing by a singledisc method. Am. J. Clin. Pathol. 45:493-496.
3. Brown, M. R. W. 1972. Relation between cation and lipidcontent of cell walls of Pseudomonas aeruginosa, Pro-teus vulgaris and Klebsiella aerogenes and their sen-sitivity to polymyxin B and other antibacterial agents.J. Pharm. Pharmacol. 24:215-218.
4. Brown, M. R. W. 1975. The role of the cell envelope inresistance, p. 71-107. In M. R. W. Brown (ed.), Resist-ance of Pseudomonas aeruginosa. John Wiley & Sons,Inc., New York.
5. Dunnick, J. K., and W. M. O'Leary. 1970. Correlationof bacterial lipid composition with antibiotic resistance.J. Bacteriol. 101:892-900.
6. Edmonds, P., and J. J. Cooney. 1969. Lipids of Pseu-domonas aeruginosa cells grown on hydrocarbons andon Trypicase soy broth. J. Bacteriol. 98:16-22.
7. Ellwood, D. C., and D. W. Tempest. 1972. Environmen-tal effects on bacterial walls. Adv. Microb. Physiol. 7:83-117.
8. Farrel, J., and A. H. Rose. 1967. Temperature effectson microorganisms. Annu. Rev. Microbiol. 21:101-120.
9. Folch, J., M. Lees, and G. H. S. Stanley. 1957. A simplemethod for the isolation and purification of total lipidsfrom animal tissue. J. Biol. Chem. 226:497-509.
10. Jacobson, L. A., R. C. Bartholomaus, and I. C. Gun-
ANTIMICROB. AGENTS CHEMOTHER.
salus. 1966. Represssion of malic enzyme by acetate inPseudomonas. Biochem. Biophys. Res. Commun. 24:955-960.
11. Kenward, M. A., M. R. W. Brown, S. R. Hesslewood,and C. Dillon. 1978. Influence of R-plsmid RP1 ofPseudomonas aeruginosa on cell wall composition,drug resistance, and sensitivity to cold shock. Antimi-crob. Agents Chemother. 13:446-453.
12. Massey, L. K., J. R Sokatch, and R. S. Conrad. 1976.Branched-chain amino acid catabolism in bacteria. Bac-teriol. Rev. 40:42-54.
13. Morrison, W. R., andL M. Smith. 1964. Preparation offatty acid methyl esters and dimethyl acetals from lipidswith boron trifluoride methanol. J. Lipid Res. 5:600-608.
14. Ornston, L. N. 1971. Regulation of catabolic pathways inPseudomonas. Bacteriol. Rev. 35:87-116.
15. Roe, E. A., R. J. Jones, and E. J. L Lowbury. 1971.Transfer of antibiotic resistance between Pseudomonasaeruginosa, Escherichia coli, and other gram-negativebacilli in bums. Lancet i:149-152.
16. Tueber, M., and S. R Miller. 1977. Selective binding ofpolymyxin B to negatively charged lipid monolayers.Biochim. Biophys. Acta 467:280-289.
17. Washington, J. A., I, and A. L Barry. 1974. Dilutiontest procedures, p. 410-417. In E. H. Lennette, E. H.Spaulding, and J. P. Truant (ed.), Manual of clinicalmicrobiology, 2nd ed. American Society for Microbiol-ogy, Washington, D. C.
on July 12, 2018 by guesthttp://aac.asm
.org/D
ownloaded from