effects of beauveria bassiana on embryos of the inland silverside fish (menidia beryllina)
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Sept. 1992, p. 2840-2845 Vol. 58, No. 90099-2240/92/092840-06$02.00/0Copyright X) 1992, American Society for Microbiology
Effects of Beauveria bassiana on Embryos of the InlandSilverside Fish (Menidia beryllina)tFRED J. GENTHNER* AND DOUGLAS P. MIDDAUGH
U.S. Environmental Protection Agency, Environmental Research Laboratory,Gulf Breeze, Florida 32561
Received 6 April 1992/Accepted 8 June 1992
A chemical toxicity and teratogenicity test was adapted to assess potential adverse effects of a microbial pestcontrol agent on a nontarget fish. Developing embryos of the inland silverside, Menidia beryllina, were exposedto conidiospores of the insect-pathogenic fungus Beauveria bassiana. Embryo rupture and death were observed.Embryo rupture did not always result in death, nor was death always associated with embryo rupture.Adherence of spores to the chorion, followed by germination and penetration by the germ tube, probablycaused the embryos to rupture. Statistically significant (P < 0.05) responses were observed in tests in whichconidiospore concentrations were .8.3 x 104 or .1.5 x 106/ml. Conidiospores treated with a dispersant(biological detergent) showed significantly less binding (P c 0.01) to embryos than did untreated spores. Bothdetergent-treated and heat-killed spores failed to cause significant adverse effects.
Concern over potential adverse effects of chemical pesti-cides on our environment and health has prompted industryto resume or accelerate development of microbial pestcontrol agents. Applicants desiring to register microbial pestcontrol agents with the U.S. Environmental ProtectionAgency are required by the Federal Insecticide, Fungicideand Rodenticide Act to perform a battery of tests to evaluaterisks associated with the environmental application of thesebiological products. These tests are described in the Subdi-vision M Guidelines (36). Since introduced organisms haveoccasionally disrupted or killed nontarget organisms (12),our objective was to develop and validate protocols forassessing nontarget effects of microbial pest control agents.We adapted a chemical toxicity and teratogenicity test that
utilizes developing embryos of the inland silverside, Menidiaberyllina, to evaluate potential toxicity and pathogenicity ofa fungal microbial pest control agent to a nontarget organ-ism. We selected this test system because juvenile forms arefrequently more sensitive and because mycosis is a commonpathological condition of fish embryos (2, 28). In addition,embryonic and larval silversides have been used success-fully for assessment of toxic and teratogenic effects ofpesticides, oil, and groundwater contaminants (11, 21, 24,26, 27). The inland silverside is found in estuaries, coastalrivers, and lakes from Massachusetts to Vera Cruz, Mexico,and in the Mississippi River basin (4). Thus, it is widelyapplicable for nontarget testing. Moreover, embryos andlarvae of this fish are readily obtained from populations ofadults maintained in the laboratory (22).The test organism, Beauveria bassiana, is an ento-
mopathogenic deuteromycete fungus currently being testedand developed as a cost-effective insect biocontrol agent (6,15, 19). B. bassiana has been used predominately in terres-trial applications by spraying wettable powders or broad-casting dry preparations to control insect pests such asColorado potato beetles, fire ants, white fly larvae, Euro-pean corn borers (19), and grasshoppers (18). However, B.
* Corresponding author.t Contribution no. 774 from the Environmental Research Labo-
ratory, U.S. Environmental Protection Agency, Gulf Breeze, Fla.
bassiana and a related entomopathogenic fungus, Metarhi-zium anisopliae, have also been tested in aquatic systems forthe control of mosquito larvae (1, 5, 29). B. bassiana isgenerally applied at very high field doses in the range ofabout 1 x 10O3 to 3 x 1013 spores per ha (6, 19) or 5 x 105spores per ml of water (29).
Despite the massive field doses required for pest control,the safety of entomogenous fungi for vertebrates has notbeen completely resolved (6). The results of most studieshave indicated that B. bassiana was relatively nontoxic ornoninfectious to vertebrates (13, 20). However, these studieswere reported in homothermic vertebrates. Cases of beau-veriosis in a captive alligator (7) and a giant land tortoise (9)suggested that additional safety testing of this fungus beperformed with poikilothermic vertebrates. Thus, we ex-posed developing embryos of the inland silverside to conid-iospores of B. bassiana to determine whether any adverseeffects would occur.
MATERIALS AND METHODS
Cultivation of fungi and recovery of spores. B. bassiana(UF1 5789) was obtained from D. Boucias, Entomology andNematology Department, The University of Florida, Gaines-ville. Strain UF1 5789 was isolated in 1989 from a bananaweevil, Cosmopolitis sordidas, near Homestead, Fla. Thefungus was cultured at 25°C on glucose-yeast extract-basalsalts agar medium (GYBS) (3). At 10 to 14 days postinocu-lation, progeny conidiospores were harvested by scrapingconfluent mycelial mats with a sterile spatula. The conidiawere freeze-dried and stored frozen at -80°C.Numbers of conidiospores were determined by plate
counts. Spores were suspended at a concentration of 1mg/ml in a detergent solution (0.03% Triton X-100; UnionCarbide Chemicals and Plastics Co., Inc.) by gentle aspira-tion in a hand-held tissue homogenizer. Biological detergentsare routinely used to prevent clumping of the spores. Dilu-tions of this suspension were spread onto the surface ofGYBS-nalidixic acid (0.5 mg/ml) plates with a sterile bentglass rod. Plates were incubated at 25'C. Fungal colonieswere counted after 6 days.
B. bassiana was recovered from exposed embryos and
2840
on Novem
ber 18, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

EFFECTS OF B. BASSIANA ON FISH EMBRYOS 2841
larvae by extensively washing them (four times in 10 ml ortwice in 100 ml of sterile water) to eliminate the sporescarried over in water from the exposure tubes and thenhomogenizing the tissue and spreading dilutions onto thesurface of GYBS-nalidixic acid plates.Embryo tests. Inland silverside embryos were obtained
from adults that spawned naturally in the laboratory at 5 %osalinity and 25°C. The photoperiod was 16 h of light and 8 hof darkness (light intensity, approximately 17 microeinsteinsper m2 per s). Details of the laboratory spawning systemwere described by Middaugh et al. (23, 25).
Exposures were initiated by washing blastula-stage em-bryos five times with sterile moderately hard water (37). Thehardness was equivalent to 80 to 100 mg of CaCO3 per liter.Single embryos were placed in each of 120 randomly orderedLeighton culture tubes. Tubes were then reordered, and 6 mlof sterile moderately hard water was added to each of thefirst 30 embryo-containing tubes, which served as controls.For embryo exposures, a 1-mg/ml conidiospore suspensionwas prepared in sterile moderately hard water. This suspen-sion yielded a culturable count of approximately 108 CFU/ml. Spore clumps were dispersed in a hand-held tissuehomogenizer. This concentrated stock was used to prepare10-fold dilutions in moderately hard water. Serial dilutionscontaining 10, 1, and 0.1 ,ug of conidiospores per ml wereused for testing. Thirty tubes, each containing a singleembryo, were filled with 6 ml of each of the three dilutions.This design yielded four treatments, a control and three10-fold dilutions of conidiospores, with 30 replicate embryosfor each treatment. In additional tests, embryos were ex-posed, as described above, to conidiospores (10 ,ug/ml) thathad been sterilized in an autoclave (20 min, 15 lb/in2),washed in 0.03% Triton X-100 to aid in spore dispersal, orleft untreated.
Short-term attachment experiments were performed todetermine whether the detergent Triton X-100 decreasedconidiospore attachment and whether the number of conid-iospores attached per embryo was dependent upon theconcentration of conidiospores in the water. In the firstattachment experiment, two treatments (10 p,g of unwashedand detergent-treated conidiospores per ml) were used. Inthe second attachment experiment, three concentrations (10,1, and 0.1 ,ug/ml) of untreated conidiospores were used.After 18 h, embryos were removed from exposure tubes andwashed to remove unattached spores. Three groups of fiveembryos from each treatment were homogenized. The num-bers of conidiospores attached per embryo were determinedby plate counts.Exposure tubes were sealed with Teflon-lined screw caps,
placed in racks, and incubated in a horizontal position at25°C with a photoperiod of 14 h of light and 10 h of darkness.Observations were made daily with a Zeiss Axiovert model35 inverted microscope equipped for photomicrography todetermine the response of developing embryos to conidio-spore suspensions. The observed responses of individualembryos were scored daily as follows: 0, no effect detected;1, advanced spore germination and growth on chorion sur-face; 2, rupture of chorion with embryonic tissue extendingthrough the chorionic membrane; 3, death of embryo. At theend of each 7- to 9-day experiment the scores for eachembryo were summed. This procedure resulted in 30 indi-vidual ranks for each of the four treatments in each experi-ment. The nonparametric Friedman analysis of variance(ANOVA) by ranks was conducted to determine whether therank scores for each treatment within an experiment weresignificantly different (P c 0.05) (35). The Friedman
ANOVA by ranks test assumes that the variables (respectiveembryo responses at differing conidiospore concentrations)under consideration were measured on at least an ordinal(rank order) scale. The null hypothesis for the procedure wasthat the different treatments (variables) contained embryoswith responses (rank scores) that resulted in identical me-dian values. The Friedman ANOVA is an alternative to theparametric one-way within-subject (repeated-measures)ANOVA, and the interpretation of the null hypothesis issimilar to that in the parametric ANOVA (35). In this study,when the median values for treatment were not identical(i.e., when a significant difference [P c 0.05] existed be-tween embryo responses in at least two of the treatments),the null hypothesis was rejected.
In cases where statistically significant differences weredetected with the Friedman ANOVA, post hoc analyseswere conducted to determine whether the rank scores for thecontrol embryos and each of the conidiospore concentra-tions were significantly different from each other. Thisprocedure enables comparison of the response of controlembryos with the response of other treatments after asignificant difference(s) has been detected by rejection of thenull hypothesis in the Friedman ANOVA (32).The Kendall coefficient of concordance, a measure of the
agreement in responses among treatments within each ex-periment, was also determined. In the attachment experi-ment a two-way ANOVA (30) was used to test for significantdifferences (P c 0.01) in the number of conidiospores thatadhered to the chorion.Embryos were prepared for microscopy by fixing in 10%
neutral buffered formalin, embedding in paraffin, sectioningat 7 ,um, staining with Harris' hematoxylin and eosin, andmounting on microscope slides.
RESULTS
There was substantial variability in the observed effects ofB. bassiana on embryonic M. beryllina in the four separateexperiments (Table 1). This variability precluded among testcomparisons of embryonic responses. Individual responseswithin a given treatment were not mutually exclusive. Forexample, many embryos that exhibited advanced hyphalgrowth did not rupture, and ruptured embryos often com-pleted development and successfully hatched.The Friedman ANOVA revealed significant differences (P
c 0.05) among the treatments within each experiment (Table2). Post hoc analyses showed statistically significant re-sponses in treatments in which conidiospore concentrationswere >8.3 x 104 or c1.5 x 106/ml. Spore concentrations ofc4.4 x 104 or .9.6 x 106/ml failed to produce responses thatwere different from those of the controls (Table 2). TheKendall coefficient of concordance demonstrated that therewas significant agreement (P c 0.05) for the observedembryo responses across treatment levels within experi-ments 1 through 5.Responses attributable to spore viability and attachment
were investigated in experiment 5 by exposing embryos toconidiospores (1.5 x 10'/ml) that had been (i) untreated, (ii)killed by heat sterilization in an autoclave, or (iii) pretreatedwith the biological detergent Triton X-100. Heat-killed ordetergent-treated conidiospores at a concentration sufficientto elicit a significant pathogenic response (P c 0.05) withuntreated, viable spores failed to cause statistically signifi-cant responses in this experiment (Tables 3 and 4).
Pretreatment of conidiospores with Triton X-100 signifi-cantly decreased the number of conidiospores found at-
VOL. 58, 1992
on Novem
ber 18, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2842 GENTHNER AND MIDDAUGH
TABLE 1. Responses of embryonic M. beryllina exposed toB. bassiana in experiments 1 through 4
Treatment Observed response (%)aExpt (spores/ml) Embryo Embryo Embryo Larval
(n = 30) adv gr rupture death hatch
1 Control (0) 0 0 10 908.3 x103 0 13 13 878.3 x104 0 53 63 378.3 x 105 0 0 90 10
2 Control (0) 0 0 7 934.4 x103 0 1 7 934.4 x 104 10 10 10 874.4 x 105 10 27 27 73
3 Control (0) 0 0 7 939.6 x 104 33 67 3 939.6 x 105 33 77 13 879.6 x106 3 3 10 87
4 Control (0) 0 0 0 1001.2 x105 37 23 3 771.2 x106 17 17 7 831.2 x107 17 0 0 100
aThe sum of values in each row may exceed 100% because individualresponses were not always mutually exclusive. adv gr, advanced hyphalgrowth on chorion surface.
tached per embryo. Embryos exposed to untreated conid-iospores (7.2 x i05 1.2 x 105/ml) had an average of 2,084+ 440 spores attached per embryo. In contrast, embryosexposed to the detergent-treated conidiospores (6.5 x 105 +0.6 x 105/ml) had an average of 620 + 408 spores attachedper embryo.The number of conidiospores attached per embryo was
directly related to the concentration of conidiospores in thewater. Embryos incubated in water containing 1.2 x 106 +0.4 x 106, 1.2 x 105 + 0.4 x 105, and 1.2 x 104 + 0.4 x 104
TABLE 2. Summary of the Friedman ANOVA and post hocanalyses of data for experiments 1 through 4
Post hoc analysesExpt X2 df ANOVA P Treatment X
(spores/ml) rank
1 4.9 3 .0.01 Control (0) 49.58.3 x 103 58.0 NSa8.3 x 104 85.0 <0.018.3 x 10-5 107.5 .0.01
2 8.8 3 c0.05 Control (0) 67.54.4 x 103 66.0 NS4.4 x 104 74.0 NS4.4 x 105 92.5 .0.05
3 42.4 3 .0.01 Control (0) 50.09.6 x 104 96.0 <0.019.6 x 105 100.0 .0.019.6 x 106 54.0 NS
4 15.7 3 .0.01 Control (0) 57.01.2 x 105 94.0 .0.011.2 x 106 81.5 c0.051.2 x 107 67.5 NS
a NS, not significant.
TABLE 3. Responses of embryonic M. beryllina exposedto B. bassiana in experiment 5
Observed response" (%)Treatment'(n = 30) Embryo Embryo Embryo Larval
adv gr rupture death hatch
Control 0 0 3 97Viable spores 43 3 7 93Killed spores 0 0 3 97Detergent-treated spores 0 0 0 100
a In each treatment with spores, 1.5 x 106 spores were used. In the control,no spores were added.
b See footnote a of Table 1.
conidiospores per ml were found to have 3,120, 520, and 48conidiospores attached per embryo, respectively.
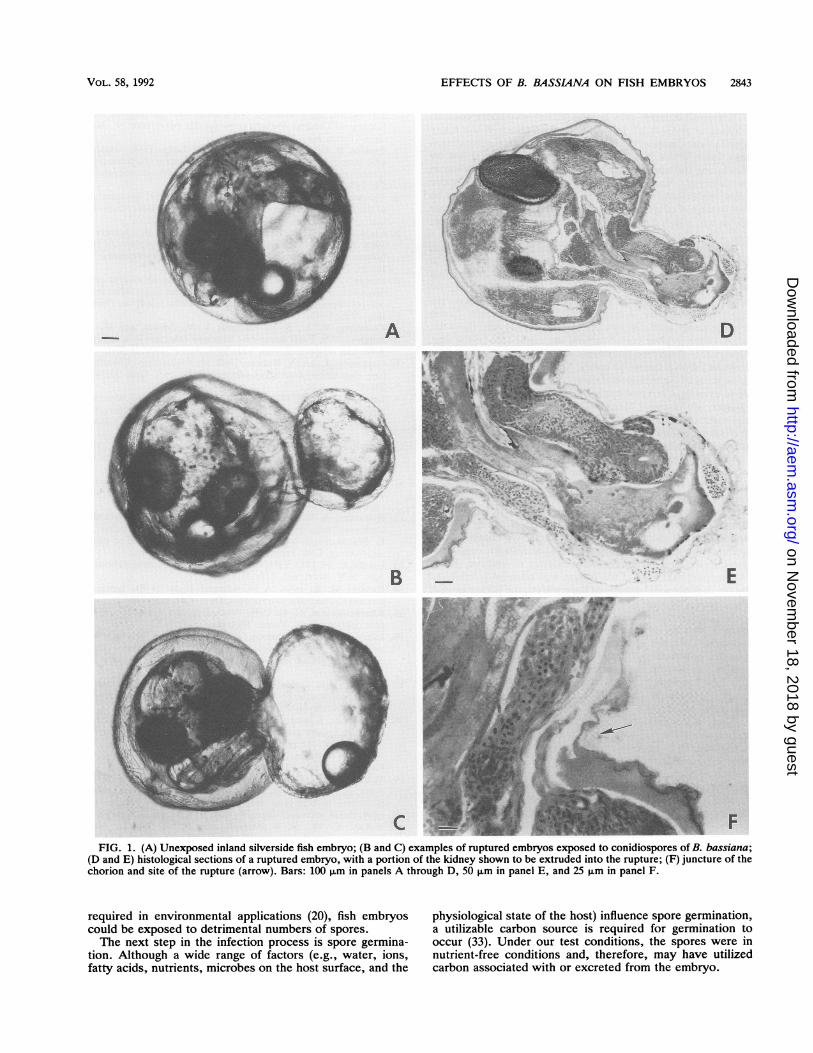
B. bassiana was consistently isolated from washed em-bryos that had been exposed to spores at all concentrations.B. bassiana was not isolated from unexposed embryos orlarvae. B. bassiana was isolated from all newly emergedlarvae exposed as embryos to 4.4 x 106 spores per ml andfrom 60% of the newly emerged larvae exposed as embryosto 8.3 x 103 spores per ml.An unexposed control embryo and examples of a rupture
caused by B. bassiana are shown in Fig. 1A, B, and C.Eleven spore-treated embryos were examined as histologicalsections. Tissues that extruded into the ruptures varied. Aportion of the kidney that extruded into the rupture is shownin Fig. 1D and E, and the juncture of the chorion and the siteof rupture is shown in Fig. 1F.
DISCUSSION
The initial developmental event in an entomopathogenicfungal mycosis is attachment of spores to the host cuticle.Al-Aidroos and Roberts (1) reported that the failure of aentomopathogen to attach was considered a feature of avir-ulent isolates. Boucias et al. (3) found that the hydrophobic-ity of B. bassiana conidia was responsible for their adhesionto insect cuticle and that detergents known to neutralizehydrophobicity reduced conidial binding. Our results sup-port those of Boucias et al. (3) and extend adhesion of thehydrophobic spores to include the chorion of the inlandsilverside embryo. Thus, the fact that detergent treatment ofconidiospores did not significantly affect our inland silver-side embryo assay may be due to a reduction in conidialbinding. Our finding that the number of conidiospores foundattached per embryo was directly proportional to the con-centration of conidiospores in the water supports the dose-response data. Specifically, conidiospore concentrations of.8.3 x 104 were required to show a significant effect.Considering the massive doses of B. bassiana conidiospores
TABLE 4. Significant differences in responses by post hocanalysis in experiment 5a
Treatment X rank P
Control 68.5Viable spores 97.5 <0.05Killed spores 68.0 NSbDetergent-treated spores 66.0 NS
a In the Friedman ANOVA, x2 = 13.6, the df = 3, and P c 0.05.b NS, not significant.
APPL. ENVIRON. MICROBIOL.
on Novem
ber 18, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
EFFECTS OF B. BASSL4NA ON FISH EMBRYOS 2843
A
B
CFIG. 1. (A) Unexposed inland silverside fish embryo; (B and C) examples of ruptured embryos exposed to conidiospores of B. bassiana;
(D and E) histological sections of a ruptured embryo, with a portion of the kidney shown to be extruded into the rupture; (F) juncture of thechorion and site of the rupture (arrow). Bars: 100 ,um in panels A through D, 50 ,um in panel E, and 25 pm in panel F.
required in environmental applications (20), fish embryoscould be exposed to detrimental numbers of spores.The next step in the infection process is spore germina-
tion. Although a wide range of factors (e.g., water, ions,fatty acids, nutrients, microbes on the host surface, and the
physiological state of the host) influence spore germination,a utilizable carbon source is required for germination tooccur (33). Under our test conditions, the spores were innutrient-free conditions and, therefore, may have utilizedcarbon associated with or excreted from the embryo.
VOL. 58, 1992
'W- -F 4 'R tv, 7t,-Aw*#F%wJ,.
-.,*, 1, -4
on Novem
ber 18, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

2844 GENTHNER AND MIDDAUGH
Nutrient limitation may be one reason why spore concen-trations of .9.6 x 106 failed to show an effect. If too manyspores were competing for limited carbon, spore germinationmay have been inhibited. Effects might be enhanced byincreasing the concentration of dissolved organic carbon inour system. Another explanation as to why high sporeconcentrations of .9.6 x 106 failed to show an effect may bethe presence of germination inhibitors that reach effectivelevels when high concentrations of conidiospores are present(16, 17).The developmental events in fish embryos infected with
fungi have not been described as fully as have those ininsects. Most reports have described events caused by fungibelonging to the class Oomycetes. Gajduesek et al. (8)reported that fungi of the genus Saprolegnia weakened carpegg membranes by damaging their microstructure. Somehyphae penetrated through the egg membrane into the yolk,while other hyphae coiled around the outer side of the egg.Others (31) described the events as a pigment change (whiteto black) with a failure to hatch. Smith et al. (34) found thatzoospores of Saprolegnia diclina and S. ferax only infecteddead trout eggs but that hyphae of both species colonized thechorion of live eggs. Neish and Hughes (28) concluded thatthe most common observation with saprolegnian fungi wasthe colonization of dead eggs, with the eventual smotheringof adjacent living eggs.The proteinaceous nature of the chorion (14) probably
rendered the inland silverside embryo susceptible to diges-tion by the extracellular enzymes of this fungus. The rup-tured chorion is a phenomenon never before observed withthis test system. The effects were observed on living eggs;fungal growth was not merely saprophytic. Since heat-killedspores did not cause any adverse effects, we conclude thatgermination and penetration were responsible for rupture ofthe chorion and the increased incidence of embryo death.We suspect that even if the fungus was effectively inhibitedby the defense mechanisms of the embryo, the penetration ofthe chorion by hyphae was sufficient to cause a rupture.Although several toxic compounds have been identified
and/or isolated from culture filtrates or mycelium of B.bassiana (19), we are uncertain as to whether these com-pounds contributed to the observed adverse effects. Ananalysis of a nonpolar extract from the conidiospore inocu-lum failed to detect beauvaricin (10), the most studied of thetoxic metabolites produced by this organism (data notshown).There was substantial variability in the observed effects in
experiments 1 through 4 (Table 1); the most severe responsewas obtained in experiment 1. We have no definitive expla-nation of this occurrence, which may be attributable to thefungus, embryos, or test conditions. It is conceivable thatembryos used in experiment 1 were inadvertently stressed,rendering them more susceptible to the fungus. Furthertesting may explain the variability.
ACKNOWLEDGMENTSWe thank Donna Crosby, Cherie Snyder-Heard, and Jeanne
Micari for technical assistance. We acknowledge John Fournie forphotomicrographs of the histological sections and preparation ofplates. We also thank George Ryan for statistical assistance.
REFERENCES1. Al-Aidroos, K., and D. W. Roberts. 1978. Mutants of Metarhi-
zium anisopliae with increased virulence toward mosquito lar-vae. Can. J. Genet. Cytol. 20:211-219.
2. Alderman, D. J. 1982. Fungal diseases of aquatic animals, p.
189-242. In R. J. Roberts (ed.), Microbial diseases of fish.Society of General Microbiology special publication 9. Aca-demic Press, Inc., New York.
3. Boucias, D. G., J. C. Pendland, and J. P. Latge. 1988. Nonspe-cific factors involved in attachment of entomopathogenic deu-teromycetes in host insect cuticle. Appl. Environ. Microbiol.54:1795-1805.
4. Chernoff, B., J. V. Conner, and C. F. Bryan. 1981. Systematicsof the Menidia beryllina complex (Pisces: Atherinidae) from theGulf of Mexico and its tributaries. Copeia 2:319-335.
5. Clark, T. B., W. R. Kellen, T. Fukuda, and J. E. Lindegren.1968. Field and laboratory studies on the pathogenicity of thefungus Beauvena bassiana to three genera of mosquitoes. J.Invert. Pathol. 11:1-7.
6. Ferron, P. 1981. Pest control by the fungi Beauveria andMetarhizium, p. 465-482. In H. D. Burges (ed.) Microbialcontrol of pests and plant diseases 1970-1980. Academic Press,Inc., New York.
7. Fromtling, R. A., S. D. Kosnake, J. M. Jensen, and G. S.Bulmer. 1979. Fatal Beauveria bassiana infection in a captiveAmerican alligator. J. Am. Vet. Med. Assoc. 175:934-936.
8. Gajduesek, J., and V. Rubcov. 1985. Microstructure of moultedcarp eggs infested with Saprolegnia. Folia Zool. Brno 34:349-355.
9. Georg, I. K., W. M. Williamson, and F. B. Tilden. 1962. Mycoticpulmonary disease of captive giant tortoises due to Beauveriabassiana and Paecilomyces fumoro-roseus. Sabouraudia 2:80-86.
10. Hamill, R. L., C. E. Higgens, H. E. Boaz, and M. Gorman. 1969.The structure of beauvericin, a new depsipeptide antibiotictoxic to Artemia salina. Tetrahedron Lett. 49:4255-4258.
11. Hemmer, M. J., D. P. Middaugh, and J. C. Moore. 1990. Effectsof temperature and salinity on Menidia beryllina embryosexposed to terbufos. Dis. Aquat. Org. 8:127-136.
12. Hollander, A. K. 1991. Environmental impacts of geneticallyengineered microbial and viral biocontrol agents, p. 251-266. InK. Maramorosch (ed.), Biotechnology for biocontrol of pestsand vectors. CRC Press, Inc., Ann Arbor, Mich.
13. Ignoffo, C. M. 1973. Effects of entomopathogens on vertebrates.Ann. N.Y. Acad. Sci. 217:141-172.
14. Kaighn, M. E. 1964. A biochemical study of the hatchingprocess in Fundulus heteroclitus. Dev. Biol. 9:56-80.
15. Khachatourians, G. G. 1986. Production and use of biologicalpest control agents. Trends Biotechnol. 4:120-124.
16. Lax, A. R., G. E. Templeton, and W. L. Meyer. 1985. Isolationpurification and biological activity of a self-inhibitor fromconidia of Colletotrichum gloeosporioides. Phytopathology 75:386-390.
17. Macko, V., R. C. Staples, P. J. Allen, and J. A. A. Renwick.1971. Identification of the germination self-inhibitor from wheatstem rust uredospores. Science 173:835-836.
18. Marcandier, S., and G. G. Khachatoruians. 1987. Susceptibilityof the migratory grasshopper, Melanoplus sanguinipes (Fab.)(Orthoptera: Acrididae), to Beauveria bassiana (Bals.)Vuillemin (Hyphomycete): influence of relative humidity. Can.Entomol. 119:901-907.
19. McCoy, C. W. 1990. Entomogenous fungi as microbial pesti-cides, p. 139-159. In R. R. Baker and P. E. Dunn (ed.), Newdirections in biological control: alternatives for suppressingagricultural pests and diseases. Alan R. Liss, Inc., New York.
20. McCoy, C. W., R. A. Samson, and D. G. Boucias. 1988.Entomogenous fungi, p. 192. In C. M. Ignoffo (ed.), CRChandbook of natural pesticides, vol. 5. CRC Press, Inc., BocaRaton, Fla.
21. Middaugh, D. P., J. W. Fournie, and M. J. Hemmer. 1990.Vertebral abnormalities in larval Menidia beryllina exposed toterbufos during embryogenesis. Dis. Aquat. Org. 9:109-116.
22. Middaugh, D. P., L. R. Goodman, and M. J. Hemmer. Methodsfor spawning, culturing, and conducting toxicity-tests with earlylife-stages of esturarine and marine fishes. In P. Calow (ed.),Handbook of ecotoxicology, in press. Blackwell Publishers,Inc., London.
23. Middaugh, D. P., M. J. Hemmer, and L. R. Goodman. 1987.
APPL. ENVIRON. MICROBIOL.
on Novem
ber 18, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

EFFECTS OF B. BASSL4NA ON FISH EMBRYOS 2845
Methods for spawning, culturing and conducting toxicity testswith early life stages of four Atherinid fishes: the inland silver-side, Menidia beryllina, Atlantic silverside, Menidia menidia,tidewater silverside, Menidia peninsulae and the Californiagrunion, Leuresthes tenuis. Report EPA-600/8-87/004. U.S.Environmental Protection Agency, Gulf Breeze, Fla.
24. Middaugh, D. P., M. J. Hemmer, and E. M. Lores. 1988.Teratological effects of 2,4-dinitrophenol, "produced water"and naphthalene on embryos of the inland silverside Menidiaberyllina. Dis. Aquat. Org. 4:53-65.
25. Middaugh, D. P., M. J. Hemmer, and Y. L. Rose. 1986.Laboratory spawning cues in Menidia beryllina and Menidiapeninsulae (Pisces: Atherinidae) with notes on survival andgrowth of larvae at different salinities. Environ. Biol. Fishes18:107-117.
26. Middaugh, D. P., J. G. Mueller, R. L. Thomas, S. E. Lantz,M. J. Hemmer, G. T. Brooks, and P. J. Chapman. 1991.Detoxification of pentachlorophenol and creosote contaminatedgroundwater by physical extraction: chemical and biologicalassessment. Arch. Environ. Contam. Toxicol. 21:233-244.
27. Mueller, J. G., D. P. Middaugh, S. E. Lantz, and P. J. Chapman.1991. Biodegradation of creosote and pentachlorophenol incontaminated groundwater: chemical and biological assess-ment. Appl. Environ. Microbiol. 57:1277-1285.
28. Neish, G. A., and G. C. Hughes. 1980. Fungal diseases of fish, p.159. In S. F. Snieszko and H. R. Axelrod (ed.), Diseases offishes, book 6. T.F.H. Publications, Inc., Neptune, N.J.
29. Pinnock, D. E., R. Garcia, and C. M. Cubbin. 1973. Beauveriatenella as a control agent for mosquito larvae. J. Invert. Pathol.22:143-147.
30. SAS Institute Inc. 1985. SAS user's guide: statistics, version 5edition. SAS Institute Inc., Cary, N.C.
31. Shah, K. L., B. C. Jha, and A. G. Jhingran. 1977. Observationson some aquatic phycomycetes pathogenic to eggs and fry offreshwater fish and prawn. Aquaculture 12:141-147.
32. Siegel, S., and N. J. Castellan, Jr. 1988. Nonparametric statisticsfor the behavioral sciences. McGraw-Hill Book Co., Inc., NewYork.
33. Smith, R. J., and E. A. Grula. 1981. Nutritional requirementsfor conidial germination and hyphal growth of Beauveria bassi-ana. J. Invert. Pathol. 37:222-230.
34. Smith, S. N., R. A. Armstrong, J. Springate, and G. Barker.1985. Infection and colonization of trout eggs by Saprolegni-aceae. Trans. Br. Mycol. Soc. 85:719-723.
35. StatSoft, Inc. 1991. Complete statistical system: statistica, ver-sion 3.0, p. 237-270. StatSoft, Inc., Tulsa, Okla.
36. U.S. Environmental Protection Agency. 1988. Pesticide assess-ment guidelines-Subdivision M. PB 83-153965. National Tech-nical Information Service, Springfield, Va.
37. U.S. Environmental Protection Agency. 1991. Methods for mea-suring the acute toxicity of effluents and receiving waters tofreshwater and marine organisms, p. 293. Report EPA/600/4-90/027. U.S. Environmental Protection Agency, Washington, D.C.
VOL. 58, 1992
on Novem
ber 18, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from