effect of monochromatic lights on nitrogen fixation and hydrogen evolution in the isolated...
TRANSCRIPT

Int. J. Hydrogen Energy, Vol. 16, No. 6, pp. 397401, 1991. 036(~3199/91 $3.00 + 0.00 Printed in Great Britain. Pergamon Press plc.
International Association for Hydrogen Energy.
EFFECT OF MONOCHROMATIC LIGHTS ON NITROGEN FIXATION AND HYDROGEN EVOLUTION IN THE ISOLATED
HETEROCYSTS OF A N A B A E N A SP. STRAIN CA
D. KUMAR and H. D. KUMAR Department of Botany, Banaras Hindu University, Varanasi 221005, India
(Received for publication 13 December 1990)
Abstraet--Photostimulation of nitrogen fixation (acetylene reduction) and nitrogenase catalysed H 2 evolution was investigated using monochromatic lights in the heterocysts isolated from a mutant (hydrogen uptake-less N9C) strain of Anabaena CA. Nitrogenase activity was strictly light dependent, being maximum in fluorescent light, followed by 620 and 650 nm light respectively. Isolated heterocysts under fluorescent light evolved H 2 in a biphasic manner. However, when the heterocysts were exposed to wavelengths of 620 and 650 nm, the pattern changed to monophasic without any initial burst. Fluorescence-emission spectra were also studied in the context of energy transfer from phycobilisomes to chlorophyll a.
1. INTRODUCTION illuminated with four F36 TI2/C/HO fluorescent lamps at an average intensity of 250/~E m -2 s -~.
In recent years, the heterocysts of filamentous nitrogen- fixing cyanobacteria have become an attractive model 2.2. Experimental techniques
system for studying the process of dinitrogen fixation. The growth rate was determined turbidimetrically Several cyanobacteria produce heterocysts when grown with Bausch and Lomb Spectronic-20 colorimeter, under diazotrophic conditions, and it is these heterocysts equipped with a broad band pass filter set with a peak that, by virtue of their unique physiological and bio- transmission centered at 660 nm. Dry weights were chemical constitution, allow the reduction of dinitrogen determined by harvesting the whole filaments or hetero- to the level of NH~- [I]. Nitrogen fixation requires both ATP and a source of reducing power. During nitrogen cysts on 0.4/~m polycarbonate filters and drying to a fixation, there is obligatory hydrogen formation [2-5]. constant weight in a vacuum oven at 45°C over P205.
The effects of light intensity and quality on nitroge- 2.3. Mutant and heterocysts isolation nase [6-11] and on nitrogenase-mediated photoproduc- For isolating the hydrogen uptake-less strain, desig- tion of H 2 [12] have been reported. There appears to nated as N9C, mutagenesis was performed as per the be no previous report on the effect of monochromatic method of Smith et al. [4]. The procedure for heterocyst lights on nitrogenase mediated hydrogen production isolation was essentially that of Kumar et al. [14] with by isolated heterocysts. We describe the effect of mono- a few modifications. A suspension of 20-30 ml of cells at chromatic lights on the acetylene reducing activity, a density of 0.10-0.12 mg dry wt ml- ~ was washed twice nitrogenase-mediated H2 production, and energy trans- in ASP-2 medium with the total concentration of KCI fer in the isolated heterocysts in a mutant strain (N9C) of raised from 0.008 to 0.03 M and NaCI from 0.085 to Anabaena sp. strain CA that lacks uptake hydrogenase 0.37 M (assay medium). The cells were resuspended activity, in 5 ml of the assay medium containing 1 mg ml-~ of
lysozyme and sparged for 5 min with 100% Ar. Cells 2. EXPERIMENTAL were then transferred to a 27 ml serum stoppered tube
containing the same gas mixture, using a gas tight 2.1. Organism and culture conditions syringe, and the tube was placed in a 39°C light bath
The organism used for these studies was the wild type equipped with a linear rotating bar having 2.54 cm Anabaena sp. strain CA (ATCC 33047), a filamentous, stroke at 48 rpm. The bath was illuminated from the heterocystous, marine cyanobacterium. Cultures were bottom by six F48 T12/CW/HO fluorescent lamps at an routinely grown in Borosil culture tubes (22 x 175 mm) average intensity of 300/rE m -2 s -~. After 35 min, the containing 20 ml of ASP-2 medium [13] with no com- suspension was sonicated for 18 s on a model W-385 bined nitrogen and a NaCI content of 5 g 1-2. The sonicator (Heat System Ultrasonics, Inc., Plainview cultures were grown in 2% CO 2 in air at 39 + 1 °C and N.Y.) at output setting of 5. The sonicated suspension
397

398 D. KUMAR and H. D. KUMAR
was transferred to an 8-ml serum stoppered vacutainer spectrofluorimeter equipped with a differential corrected tube and centrifuged at 2000 rpm for 5 min. The final spectral unit, using a slit width of 8 nm. Samples for pellet was resuspended in either a 2-ml volume for fluorescence spectra were taken into the quartz cuvettes acetylene reduction assay or 3-ml volume for use on the (1 cm light path) with a perforation at the top cover to hydrogen electrode, sparge with the desired gas. Biochemicals were pur-
For acetylene reducing activity, a suspension of 2 ml chased from Sigma Chemical Co., St. Louis, Missouri. of isolated heterocysts was placed in an 8-ml serum stoppered tubes containing either 10% C2H2 + 90% H2, 3. RESULTS or 10% C2Hz ÷ 90% Ar, and placed in the same shaker bath as that used for heterocyst isolation. The ethylene The cellular nitrogenase activities found in Anabaena formed was measured by injection of 0.2 ml of the sp. strain CA and its hydrogen uptake-less (N9C) mu- gas-phase into an Antek model 464-IPC gas chromato- tant strain grown diazotrophically in 2% CO2-in-air graph equipped with a 182-cm Chromosorb 140 column were 1.4 _ 0.2 and 1.3 _+ 0.15 p moles C 2H 4 mg dry wt- (Johns-Manville, Celite, Division, Denver, Colorado). h 1 (80 + 10 and 75 _+ 10/~moles C2H4 mg Chl a- I h - l )
Amperometric measurements of hydrogen were per- respectively. In both the cases about 10% of the cells formed as described previously [4], using one 5331 differentiated as heterocysts. The isolated heterocysts electrode (Yellow Springs Instruments Co., Yellow from the Ngc strain showed 1.6_ 0.1/~moles C2H 4 mg Springs, Ohio) fitted into a 1.8ml water-jacketed drywt -~ h -~ under either 90% H2+ 10% C2H2or90% chamber held at 39°C. The suspensions were continu- Ar ÷ 10% C2H2 and nickel supplementation had no ously mixed with a magnetic stirring bar. The electrode effect on the acetylene reducing activity (Table 1). How- signal was monitored and amplified by a Keithley ever, the heterocysts isolated from the wild type strain model 617 microvoltammeter (Keithley Instruments, showed significant stimulation of acetylene reducing Inc., Cleveland, Ohio) and recorded. Actinic light activity under hydrogen incubation with or without was provided by a projector with DAY-DAK 500-W nickel (100 nM NiClz.6H20 ) supplementation in light. lamp (Sylvania, Winchester, Kentucky) screened by a Fluorescent light (300 #E m-2 s- l ) was conducive to No. 34-01-2 hot mirror (Baird Atomic Inc., Bedford, maximum acetylene reducing activity (1.6 + 0.1/~moles Massachusetts). The light intensity incident upon the C 2H 4 mg dry wt ~ h ~) followed by 620 nm and 650 nm electrode chamber was 1200/~E m 2 s ~. lights (0.3 _ 0.01 and 0.20 _ 0.01 #mole C2H 4 mg dry
Monochromatic light of wavelengths 620 and 650 nm wt 1 h ~). All measurements were performed under 90% were obtained by passing the light from one DAY-DAK H2+ 10% CzH 2. Equal energy levels (100/IE m 2 s ~) 500 W lamp screened by 34-01-2 hot mirror through were obtained by a Variac potentiometer attached with a 620 nm (44-78-62) and 650 nm (44-78-65) Bausch and light source. At the same (100/rE m-2 s-~) light intensity Lomb interference second order filters having half fluorescent light showed 0.5 + 0.10/tmoles C2H4mg dry band width of 2 nm each. Sharp cut yellow (CS-2-63) wt ~ h J. As might be expected, the capacity to photo- and red (CS-2-62) filters (Corning Glass Works, produce H2 varies widely depending on the prevailing Corning, New York) were placed separately before levels of nitrogenase and hydrogenase. To see the effect reference filters. Equal energy levels of all lights were of monochromatic lights only on the nitrogenase- obtained by changing the applied voltage to the lamp catalysed H 2 evolution, one must have a system where with a Variac-Duratrak potentiometer. Radiant energy there is no hydrogen uptake or hydrogenase enzyme. So was measured with a quantum meter (model 185 A, it was desirable to isolate a mutant lacking uptake Li-Cor Inc., Lincoln, Nebraska) calibrated with a hydrogenase. The direct kinetic response of different Hilger-Schwartz thermophile exposing a diaphragm monochromatic lights on nitrogenase-catalysed H 2 pro- area of 1 cm 2. Fluorescence spectra were determined as duction and H 2 uptake in the isolated heterocysts of described by Gray et al. [15] with a Perkin-Elmer MPF4 Hup- (a hydrogen uptake-less) mutant strain is shown
Table 1. Physiological properties of isolated heterocysts of Anabaena CA and mutant strain N9C
Acetylene reducing activity (ltmoles C2H 4 mg dry wt ~ h ~) H 2 evolution H 2 uptake
(/~1 H: produced/consumed mg dry wt -~ h 1) Without nickel With nickel
90% H2+ 90% Ar+ 90% H2+ 90% Ar+ Without With Without With 10% C2H2 10% C2H2 10% C2H 2 10% C2H2 nickel nickel nickel nickel
Anabaena CA 2.1 1.0 3.8 1.0 48.0, 28.0 0.0 2.5 18.0 Mutant Ngc N2-fixing 1.6 1.6 1.6 1.6 46.0, 24.0 46.0, 24 0 0 KNO 3 (10mM) grown 0.5 0.5 0.5 0.5 10.0, 4.0 10.0, 4.0 0 0
Effect of nickel-supplementation (100riM) on the acetylene reducing, hydrogen evolution and uptake activities of isolated heterocysts from parent and mutant strain of Anabaena CA. Cultures were grown for five generation under nickel supplementation for isolation of heterocysts. H 2 evolution and uptake activities were measured under 100% argon. First value for hydrogen evolution represents hydrogen burst followed by steady rate.
N2 FIXATION AND H 2 EVOLUTION IN ANABAENA 399
l _ _ F t type Anabaena CA showed significant stimulation of o.41pt Ha ~ nitrogenase (acetylene reducing) activity and complete
disappearance of hydrogen evolution under nickel sup- 1" ~o FL f ~250 plementation. However, when the heterocysts isolated
j from the hydrogen uptake less (N9C) mutant strain were ~ ' 7 2 / ~ / ~ " / * subjected to nickel supplementation, acetylene reducing
/ O ~ ~ J activity remained unaffected under argon or hydrogen incubation, and hydrogen evolution continued till the
/ o / . _ ~ ~ . j o experiment was over. The mutant strain described above represents a novel
/ , ~ . o / ~ . . ~ . ~ mutant not reported in any cyanobacteria. A similar g / ~ . ~ type of a mutant strain, N9AR (a hydrogen uptake less
mutant strain with lower generation time than the wild ~ , ~°'~, t i ~ J i ~ ~ i , j type, and other different physiological properties) had
0 2 4 6 a lo 12 been isolated and studied [4]. However, this mutant strain differs from N9AR in having a higher generation
TIME (minl time and producing heterocysts even in nitrate sup-
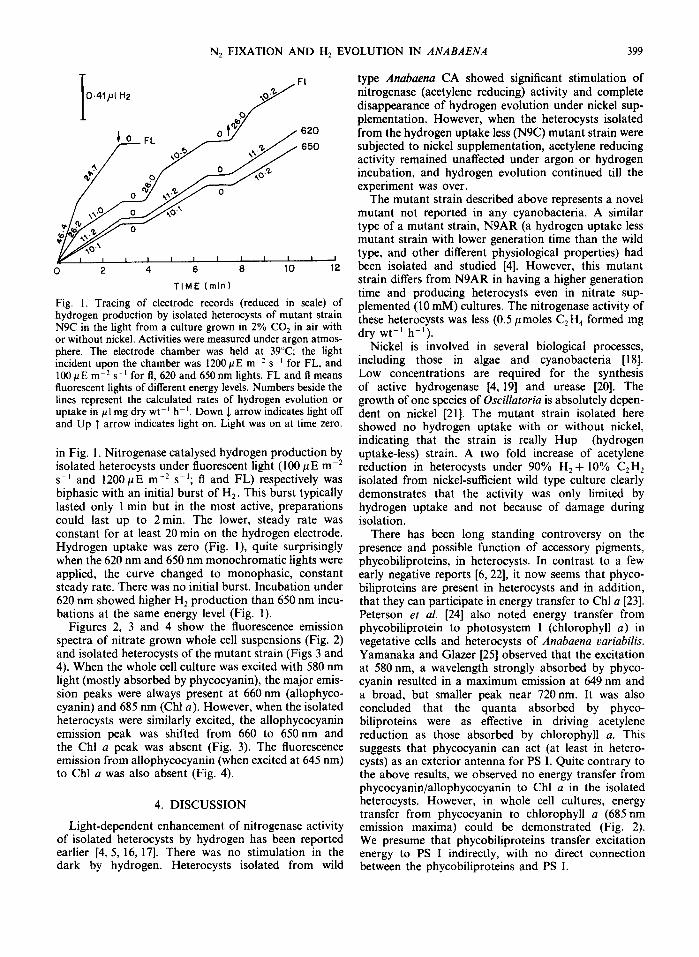
Fig. 1. Tracing of electrode records (reduced in scale) of plemented (10 mM) cultures. The nitrogenase activity of hydrogen production by isolated heterocysts of mutant strain these heterocysts was less (0.5/tmoles C2H4 formed mg N9C in the light from a culture grown in 2% CO2 in air with dry wt-l h-i). or without nickel. Activities were measured under argon atmos- phere. The electrode chamber was held at 39°C; the l igh t Nickel is involved in several biological processes, incident upon the chamber was 1200/~E m 2 s ~ for FL, and including those in algae and cyanobacteria [18]. 100 ttEm -2 s -t for fl, 620 and 650 nm lights. FL and fl means Low concentrations are required for the synthesis fluorescent lights of different energy levels. Numbers beside the of active hydrogenase [4, 19] and urease [20]. The lines represent the calculated rates of hydrogen evolution or growth of one species of Oscillatoria is absolutely depen- uptake in #1 mg dry wt -~ h ~. Down J. arrow indicates light off dent on nickel [21]. The mutant strain isolated here and Up T arrow indicates light on. Light was on at time zero. showed no hydrogen uptake with or without nickel,
indicating that the strain is really Hup (hydrogen in Fig. 1. Nitrogenase catalysed hydrogen production by uptake-less) strain. A two fold increase of acetylene isolated heterocysts under fluorescent light (100/~E m -2 reduction in heterocysts under 90% H2 + 10% C2H 2 s -~ and 1200/~E m -2 s-I; fl and FL) respectively was isolated from nickel-sufficient wild type culture clearly biphasic with an initial burst of H2. This burst typically demonstrates that the activity was only limited by lasted only 1 min but in the most active, preparations hydrogen uptake and not because of damage during could last up to 2 min. The lower, steady rate was isolation. constant for at least 20 min on the hydrogen electrode. There has been long standing controversy on the Hydrogen uptake was zero (Fig. I), quite surprisingly presence and possible function of accessory pigments, when the 620 nm and 650 nm monochromatic lights were phycobiliproteins, in heterocysts. In contrast to a few applied, the curve changed to monophasic, constant early negative reports [6, 22], it now seems that phyco- steady rate. There was no initial burst. Incubation under biliproteins are present in heterocysts and in addition, 620 nm showed higher H2 production than 650 nm incu- that they can participate in energy transfer to Chl a [23]. bations at the same energy level (Fig. 1). Peterson et al. [24] also noted energy transfer from
Figures 2, 3 and 4 show the fluorescence emission phycobiliprotein to photosystem I (chlorophyll a) in spectra of nitrate grown whole cell suspensions (Fig. 2) vegetative cells and heterocysts of Anabaena variabilis. and isolated heterocysts of the mutant strain (Figs 3 and Yamanaka and Glazer [25] observed that the excitation 4). When the whole cell culture was excited with 580 nm at 580 nm, a wavelength strongly absorbed by phyco- light (mostly absorbed by phycocyanin), the major emis- cyanin resulted in a maximum emission at 649 nm and sion peaks were always present at 660 nm (allophyco- a broad, but smaller peak near 720 nm. It was also cyanin) and 685 nm (Chl a). However, when the isolated concluded that the quanta absorbed by phyco- heterocysts were similarly excited, the allophycocyanin biliproteins were as effective in driving acetylene emission peak was shifted from 660 to 650 nm and reduction as those absorbed by chlorophyll a. This the Chl a peak was absent (Fig. 3). The fluorescence suggests that phycocyanin can act (at least in hetero- emission from allophycocyanin (when excited at 645 nm) cysts) as an exterior antenna for PS I. Quite contrary to to Chl a was also absent (Fig. 4). the above results, we observed no energy transfer from
phycocyanin/allophycocyanin to Chl a in the isolated 4. DISCUSSION heterocysts. However, in whole cell cultures, energy
transfer from phycocyanin to chlorophyll a (685 nm Light-dependent enhancement of nitrogenase activity emission maxima) could be demonstrated (Fig. 2).
of isolated heterocysts by hydrogen has been reported We presume that phycobiliproteins transfer excitation earlier [4, 5, 16, 17]. There was no stimulation in the energy to P S I indirectly, with no direct connection dark by hydrogen. Heterocysts isolated from wild between the phycobiliproteins and PS I.

400 D. K U M A R and H. D. K U M A R
o~ tO
z ,~ ~ I~ LU , 0
o " ~ I I I I 1 I i I I I 0 - 0 . .
0 0 0 0 0 0 0 0 0 0 ~0 ._
AJ.ISN31NI 33N30S3~IOn7& 3AIIV73~I
g ~
o ~
_ ~ o ~ "~~ e~
0 J ~ o o
> ~ ~
o ~ N I I I f I I I I I ~ O ~
0 0 0 0 0 0 0 0 0 0 ~ m " ~ 0 O~ O0 I~- tO ~ ~i" rO 0,1 ~ ~ . ~
A I I S N 3 1 N I 3 0 N 3 0 S 3 ~ I O n 7 4 3 A 1 1 ~ ' 7 3 ~ I ~ ~ u o
o 0 '~ -~ 0 ~ c~ "~_~ o ~.~
~ o~ o~ - r 0 0
I I I I , I I I I I ~ r ~ . 0 0 0 0 0 0 0 0 0 0 0 O~ ~0 ~ ~0 ~0 ~ ~ ~ ~ ~0
A I I S N ~ / N I 3 0 N ~ S ] ~ o n 7 ~ = I ^ I I V 7 3 E I

N 2 FIXATION AND H 2 EVOLUTION IN ANABAENA 401
When we purified aUophycocyanin and phycocyanin 7. P. Fay, Factors influencing dark nitrogen fixation in a from isolated heterocysts, they showed characteristics of blue-green alga. Appl. Environ. Microbiol. 31, 376-379 whole cell cultures, i.e. al lophycocyanin showed absorp- (1976). tion maxima at 278, 350, 645 nm with shoulders at 288, 8. M. Lex and W. D. P. Stewart, Algal nitrogenase, reductant
pools and photosystem I activity. Bioehirn. Biophys. Acta 600 and 620nm, and emission maxima at 660nm. 292,436-443 (1973). Phycocyanin showed absorption maxima at 278, 350 and 9. P. Rowell, R. H. Reed, M. J. Hawkesford, A. Ernst, J. Diez 610 nm with shoulder at 288 nm and emission maxima and W. D. P. Stewart, Some aspects of the regulation of at 645 nm. The significance of this observation is that nitrogenase activity in the cyanobacterium Anabaena vari- apparently one allophycocyanin species can exhibit abilis. In A. H. Gibson and W. E. Newton (eds), Current different emission maxima depending upon the state of Perspectives in Nitrogen Fixation, pp. 186-189. Australian aggregation. We presume that in vivo allophycocyanin Acad. Sci., Canberra (1981). tends to exist in a state with emission maxima shifted 10. P. M. Mullineaux, A. E. Chaplin and J. R. Gallon, Effects toward that of phycocyanin which would enhance of light to dark-transition on carbon reserves, nitrogen
fixation and ATP concentrations in cultures of Gloeocapsa the energy transfer efficiency among the pigments, sp. 1430. J. Gen. Microbiol. 120, 227-232 (1980). The presence of phycobiliproteins in heterocysts and 11. D. C, Yoch and J. W. Gotto, Effect of light intensity their involvement in the action spectrum of acetylene and inhibitors of nitrogen assimilation on NH~- inhibition reduction strongly suggests that the light reactions of nitrogenase activity in Rhodospirillum rubrum and in the heterocysts are not simple as is commonly Anabaena sp. J. Bacteriol. 151, 800-806 (1982). supposed. 12. D. Kumar and H. D. Kumar, Light dependent acetylene
We [5] have shown the biphasic nature of H 2 evolution reduction and hydrogen production by the cyanobacterium, from isolated heterocysts of Anabaena CA. The present Anabaena sp. strain CA. Int. J. Hydrogen Energy 13, observations on the effect of 620 nm and 650 nm mono- 573-575 (1988).
13. C. Van Baalen, Studies on marine blue-green algae. Bot. chromatic lights on the burst pattern of hydrogen evol- Mar. 4, 129 139 (1962). ution constitute a novel observation. The burst pattern 14. A. Kumar, F. R. Tabita and C. Van Baalen, Isolation and was not seen with monochromat ic lights even at the characterization of heterocysts from Anabaena sp. strain same energy level. A burst of hydrogen can be seen only CA. Arch. Microbiol. 133, 103-109 (1982). when the two subunits of the nitrogenase complex are 15. B. H. Cray, J. Cosner and E. Gantt, Phycocyanins brought together under the conditions of low electron with absorption maxima at 637 nm and 623nm from flux, or when Fe-protein is limiting [3]. In our case, Agmenellum quadruplicatum. Photochem. Photobiol. 24, the burst pattern was only observed under fluorescent 299-302 (1976). light, and it changed to monophasic during 620 and 16. J. P. Houchins and G. Hind, Pyridine nucleotides and H 2
as electron donors to the respiratory and photosynthetic 650 nm light incubations. The change in hydrogen electron transfer chains and to nitrogenase in Anabaena evolution from biphasic to monophasic pattern is very heterocysts. Biochim. Biophys. Acta 682, 86-96 (1982). interesting at the same energy level and requires further 17. G. Neuer and H. Bothe, Electron donation to nitrogenase study, in heterocysts of cyanobacteria. Arch. Microbiol. 143,
185-191 (1985). Acknowledgements--D.K. thanks the University Grants Com- 18. R. M. Welch, The biological significance of nickel. J. Plant mission for financial help. Nutrition 3, 345-356 (1981).
19. A. Daday and G. D. Smith, The effect of nickel on the hydrogen metabolism of the cyanobacterium Anabaena
R E F E R E N C E S cylindrica. FEMS Microbiol. Lett. 20, 327-330 (1983). 20. P. J. Weathers, H. L. Chee and M. M. Allen, Arginine
1. W. D. P. Stewart, Some aspects of structure and function catabolism in Aphanocapsa 6308. Arch. Microbiol. 118, 1~5 in N2-fixing cyanobacteria. Ann. Rev. Microbiol. 34, (1978). 497-536 (1980). 21. C. Van Baalen and R. O'Donnell, Isolation of a nickel-
2. J. Chatt, Chemistry relevant to biological nitrogen dependent blue-green alga. J. Gen. Microbiol. 105, 351-353 fixation. In W. D. P. Stewart and J. R. Gallon (eds), (1978). Nitrogen Fixation, pp. 1-18. Academic Press, San Francisco 22. J. Thomas, Absence of pigments of photosystem II of (1980). photosynthesis in heterocysts of a blue-green algae. Nature,
3. F. B. Simpson and R. H. Burris, A nitrogen pressure of Lond. 228, 181-183 (1970). 50 atmospheres does not prevent evolution of H 2 by 23. V. V. S. Tyagi, T. B. Ray, B. C. Mayne and G. A. Peters, nitrogenase. Nature, Lond. 224, 1095-1097 (1984). The Azolla-Anabaena azollae relationship. XI. Phyco-
4. R. L. Smith, D. Kurnar, Z. Xiankong, F. R. Tabita and biliproteins in the action spectrum for nitrogenase catalyzed C. Van Baalen, H 2, N 2 and 02 metabolism by isolated acetylene reduction. Plant Physiol. 68, 1479 1484 (1981). heterocysts from Anabaena sp. strain CA. J. Bacteriol. 162, 24. R. B. Peterson, E. Drlan, H. E. Calvert and B. Ke, 565-570 (1985). Energy transfer from phycobiliproteins to photosystem I
5. D. Kumar, Effect of light quality on the acetylene in vegetative cells and heterocysts of Anabaena variabilis. reducing activity of the isolated heterocysts of Anabaena Biochim. Biophys. Acta 634, 237-248 (1981). sp. strain CA. J. Gen. Appl. Microbiol. 35, 369-375 25. G. Yamanaka and A. N. Glazer, Phycobiliproteins in (1989). Anabaena 7119 heterocysts. In G. P. Papageorgiou and
6. P. Fay, Photostimulation of nitrogen fixation in Anabaena L. Packer (eds), Photosynthetic Prokaryotes: Cell Differen- cylindrica. Biochim. Biophys. Acta 216, 353-356 (1970). tiation and Function. Elsevier, Amsterdam.