ecological rhythms and the management of humid tropical
TRANSCRIPT

University of Pennsylvania University of Pennsylvania
ScholarlyCommons ScholarlyCommons
Departmental Papers (EES) Department of Earth and Environmental Science
December 2001
Ecological rhythms and the management of humid tropical Ecological rhythms and the management of humid tropical
forests. Examples from the Caribbean National Forest, Puerto forests. Examples from the Caribbean National Forest, Puerto
Rico Rico
Frederick N. Scatena University of Pennsylvania, [email protected]
Follow this and additional works at: https://repository.upenn.edu/ees_papers
Recommended Citation Recommended Citation Scatena, F. N. (2001). Ecological rhythms and the management of humid tropical forests. Examples from the Caribbean National Forest, Puerto Rico. Retrieved from https://repository.upenn.edu/ees_papers/19
Postprint version. Published in Forest Ecology and Management, Volume 154, Issue 3, 2001, pages 453-464. Publisher URL: http://www.sciencedirect.com/science/journal/03781127 NOTE: At the time of publication, author Frederick N. Scatena was affiliated with the USDA Forest Service. Currently (September 2005) he is a faculty member in the Department of Earth and Environmental Science at the University of Pennsylvania.
This paper is posted at ScholarlyCommons. https://repository.upenn.edu/ees_papers/19 For more information, please contact [email protected].

Ecological rhythms and the management of humid tropical forests. Examples Ecological rhythms and the management of humid tropical forests. Examples from the Caribbean National Forest, Puerto Rico from the Caribbean National Forest, Puerto Rico
Abstract Abstract A common premise in modern forest management is that land management should operate over large enough spatial and temporal scales that common natural disturbances are present and implicitly considered. Less emphasis has been focused on managing humid tropical forest ecosystems with the periodic ecological processes that occur between disturbances. The central premise of this paper is that timing management activities to periodic ecological processes that occur between disturbances is an additional prerequisite for the effective management of humid tropical forests. Ecological rhythms are defined here as biological or biogeochemical processes that have definable periodicities and include phenological, circadian, biogeochemical, and behavioral processes. The paper documents the use of ecological rhythms in the management of endangered species and water resources in the Caribbean National Forest of Northeastern Puerto Rico. While this type of dynamic management has proven benefits, managers and regulatory agencies have been hesitant to utilize complex, ecologically based dynamic management schedules because they can be difficult to monitor and regulate. Fortunately, recent technological advantages greatly increase the ability to conduct complex real-time, spatially explicit management. Identifying important ecological rhythms and developing administrative structures that can integrate them into management will be a major challenge in both tropical and temperate environments in the coming decades.
Keywords Keywords Tropical forest management, Municipal water supply, Endangered species
Comments Comments Postprint version. Published in Forest Ecology and Management, Volume 154, Issue 3, 2001, pages 453-464. Publisher URL: http://www.sciencedirect.com/science/journal/03781127
NOTE: At the time of publication, author Frederick N. Scatena was affiliated with the USDA Forest Service. Currently (September 2005) he is a faculty member in the Department of Earth and Environmental Science at the University of Pennsylvania.
This journal article is available at ScholarlyCommons: https://repository.upenn.edu/ees_papers/19

Ecological rhythms and the management of humid tropical forests. Examples from the Caribbean National Forest, Puerto Rico
F. N. ScatenaInternational Institute of Tropical Forestry, USDA Forest Service, P.O. Box 25,000, San Juan, PR 00928-5000, USA
Abstract
A common premise in modern forest management is that land management should operate over large enough spatial and temporal scales that common natural disturbances are present and implicitly considered. Less emphasis has been focused on managing humid tropical forest ecosystems with the periodic ecological processes that occur between disturbances. The central premise of this paper is that timing management activities to periodic ecological processes that occur between disturbances is an additional prerequisite for the effective management of humid tropical forests. Ecological rhythms are defined here as biological or biogeochemical processes that have definable periodicities and include phenological, circadian, biogeochemical, and behavioral processes. The paper documents the use of ecological rhythms in the management of endangered species and water resources in the Caribbean National Forest of Northeastern Puerto Rico. While this type of dynamic management has proven benefits, managers and regulatory agencies have been hesitant to utilize complex, ecologically based dynamic management schedules because they can be difficult to monitor and regulate. Fortunately, recent technological advantages greatly increase the ability to conduct complex real-time, spatially explicit management. Identifying important ecological rhythms and developing administrative structures that can integrate them into management will be a major challenge in both tropical and temperate environments in the coming decades.
1. Introduction
A common premise in modern forest management is that land use management should operate over large enough spatial and temporal scales that common natural disturbance patterns are present and implicitly considered in management plans (Kaufmann et al., 1994). The recent fires in Amazonia and Borneo, and the devastating flooding in Venezuela and Honduras are clear examples of the costs of not recognizing the importance of natural disturbances and their synergistic interactions with anthropogenic activities. While the need to explicitly incorporate natural disturbances in decision making is widely recognized, less emphasis has been focused on managing humid tropical forests with implicit consideration to the ecological processes that periodically occur between disturbances. In fact, many view humid tropical forests as relatively seasonless environments. The central premise of this paper is that timing management activities to periodic ecological and biogeochemical processes that occur between

disturbances is an additional prerequisite to minimize the effects of management on aquatic and terrestrial communities.
2. Methods
2.1. Ecological rhythms
Ecological rhythms are broadly defined here as biological or environmental processes that repeatedly occur at definable intervals. These rhythms have definable periodicities and can operate at the level of individuals, populations, or communities (Odum, 1971). They are commonly related to climate (i.e. phenological patterns), daily processes (i.e. diel or circadian cycles), life histories (i.e. reproduction, feeding behavior, etc.) or physical and biogeochemical processes (i.e. tidal cycles). While rhythms can occur over a wide range of spatial and temporal scales, the emphasis in this paper is towards adapting management strategies to ecological or biogeochemical rhythms that occur on daily or seasonal time scales.
2.2. Study area
The Luquillo Mountains are the dominant physiographic feature of Northeastern Puerto Rico and rise abruptly from a narrow coastal plain to an elevation of 1000 m over a distance of 10–20 km. The upper elevations of the mountains are managed by the USDA Forest Service Caribbean National Forest (CNF). The surrounding lowlands support a mix of urban to suburban land use. The CNF is a major source of recreation, municipal water, and a site of much research on tropical forest ecology. It is also a regional center of biodiversity and contains old growth forests that have been under some form of protection since Columbus landed on the island over 500 years ago.
The Luquillo Mountains are within the path of the easterly trade winds and has a subtropical maritime climate characterized by high frequency, locally derived orographic precipitation and large-scale synoptic disturbances. Rain and runoff occur in every month (Fig. 1). Although average monthly rainfall does not show strong seasonal patterns, a drier period typically occurs between late January and April, and a wetter season from September to December. Seasonal patterns are present in the occurrence of annual peak discharges and annual low flow. There are also recognized seasonal variations in ecological processes (Fig. 1). In general, the drier periods of the year coincide with increased leaf fall and reproduction in many terrestrial animals and plants ( Odum and Pigeon, 1970). The aquatic community tends to have increased reproduction and downstream migration during the wetter part of the year and increased upstream migration during the drier part of the year ( Covich and McDowell, 1996 and Johnson). There are also seasonal differences in the amount of recreation ( Fig. 2). Peak visitation of both forest and surrounding beaches occurs during summer months. Mostly, island residents visit the area during the hot summer months. The peak season for cruise ships, resort hotels, and other off-island residents occurs between January and March.

Fig. 1. Annual calendar of ecological processes for the Tabonuco type forest of the CNF, modified from Odum and Pigeon (1970). Including mean monthly rainfall (Scatena, unpublished) and the percent of annual peak daily discharges (Rivera, 1999) and percentage of annual minimum discharges (Scatena, unpublished) for streams draining the National Forest.
Hurricanes are the dominant natural disturbances affecting the Luquillo Mountains (Scatena and Boose). Forest structure and composition, productivity, spatial and temporal distribution of plants and animals, nutrient cycles, stream water chemistry, the abundance of aquatic organisms, and many other ecosystem processes have been related to hurricanes (see examples in Lugo and Scatena (1995) and Walker et al. (1996)). Several hurricanes pass through the Caribbean region each year ( Scatena and Larsen, 1991). On average, a hurricane will pass close enough to the island to cause severe but localized damage approximately once every 10 years. Once in every 50–60 years, one will pass directly over the CNF and cause widespread defoliation and subsequent ecosystem reorganization. The implication of these large-scale disturbances on the design and management of natural reserves ( Wunderle and Wiley, 1996) and plantations ( Wang and Scatena, 2000) has been discussed elsewhere. Additional information on the ecology and management of the site can be found elsewhere ( Odum and Pigeon, 1970; Lugo and Lowe, 1995 and Reagan and Waide, 1996).

Fig. 2. Recreational use of Puerto Rican beaches by month, average daily visits to a major visitor center in the CNF, and average monthly stream water flow in a principal river draining of the CNF.
3. Results
3.1. Examples of ecological rhythms and resource management in the Caribbean National Forest
Coordinating management activities to seasonal or reproductive cycles has long been a basic precept of agriculture, husbandry and wildlife management. Recent examples of managing with ecological rhythms at the ecosystem level include: (1) timing reservoir operations to minimize impacts on the migration of aquatic organisms (Bistal and Ruff, 1996); (2) timing fertilizer application to the growth phases of vegetation to minimize nutrient export ( Matson et al., 1998); and (3) scheduling insecticide application to the life cycles of pests ( Balogh and Walker, 1992). In the CNF, ecological rhythms have

been used to enhance the management of endangered species, municipal water supplies, recreational demand, natural forests and plantations ( Table 1).
Table 1. Examples of ecological rhythms used in natural resource management in the Luquillo Mountains of Puerto Rico
3.1.1. Endangered Puerto Rican parrot
Probably, the oldest example of adapting management activities to ecological rhymes concerns the endangered Puerto Rican parrot. When Columbus first arrived on the island, there may have been several hundred thousand to over a million Puerto Rican parrots (Snyder et al., 1987). By 1975, the islands wild population had dropped to 13 free flying birds that were all living in the CNF. Subsequent research-based management by the US Fish and Wildlife Service and the USDA Forest Service have increased the wild population to over 40 wild birds. During this recovery effort, several land management rules have been based on the parrot’s biological rhymes.
During non-breeding season, the typical day for a adult parrots begins shortly after sunrise with series of short territorial flights in the vicinity of their roosts (Snyder et al., 1987). After spending the day feeding and resting, they return to their roost at dusk, about 1900 h. During periods of nest selection and breeding, this behavior varies slightly. Nest selection usually occurs between mid-December and mid-March. Egg laying is highly synchronized among pairs with all clutches being produced during late February or early March. This egg laying coincides with the peak fruiting of Sierra Palm tree, the dominant food source of the bird. Chicks usually fledge between 8 and 11 weeks. During this

period, the female guards the eggs and does not leave the roosting site during daily forging.
The wild parrot’s sensitivity to human presence and human-generated noise during nest selection and breeding is considered extremely high (Caribbean National Forest, 1997). Nevertheless, water intakes, electrical power lines, and roads occur within hearing distance of nesting areas and often require maintenance related construction activities. To mitigate the impacts of these activities, construction within hearing distance of the breeding areas has been regulated to coincide with non-breeding periods. Moreover, the land management plan requires that human activities in sensitive areas be timed to occur outside the nest selection and the breeding season (15 December–15 July). During the rest of the year, all activities in sensitive areas are planned to occur while the parrots are forging for food away from their roosts. That is, between 0730 and 1900 h.
3.1.2. Water supply management
The CNF supplies municipal water to all of Northeastern Puerto Rico and approximately 20% of the islands population. Nearly, all this water is withdrawan from low head dams that exist within or near the forest boundary. These streams also support fishing, swimming, scenic recreation, eels, fish, snails, 11 species of crustaceans, and more than 60 aquatic insects (Covich and McDowell, 1996). In an effort to maintain the multiple uses of the forest, these water extractions are permitted and regulated. Historically, water extraction was only limited by the size of the permitted intakes and instream flow requirements were not explicitly required ( Scatena and Johnson, 2001). More recently, the increased pressures for the mountains water resources has also increased the need and scrutiny of water management activities. In response, the CNF has mandated the need to maintain some level of instream flow ( Caribbean National Forest, 1997). Knowledge of the ecological rhythms, specifically the diurnal feeding and migratory behavior of the dominant aquatic organisms, has been invaluable in developing these management protocols ( March; Benstead and Johnson).
Observation of the density of individuals in specific habitats and flow conditions are often used to develop predictive models of the impacts of water diversions on habitat abundance (Johnson and Covich, 1999). Due to their abundance and ecological importance, freshwater shrimp have been the target organisms of instream flow models used in the Luquillo Mountains ( Sutton and Scatena and Johnson, 2001). Nevertheless, understanding the diurnal behavior of these shrimp is a prerequisite to successfully use their behavior to model instream flows. In general, these shrimp are active at night and hide within the substrates during daylight, apparently to avoid predation ( Fig. 3). Moreover, some species are never observed during the day. Those that are observed are typically found in intermediate water depths and relatively high velocities. In contrast, many more individuals are observed at night during which time they tend to use lower velocity microhabitats and a greater range of channel conditions.
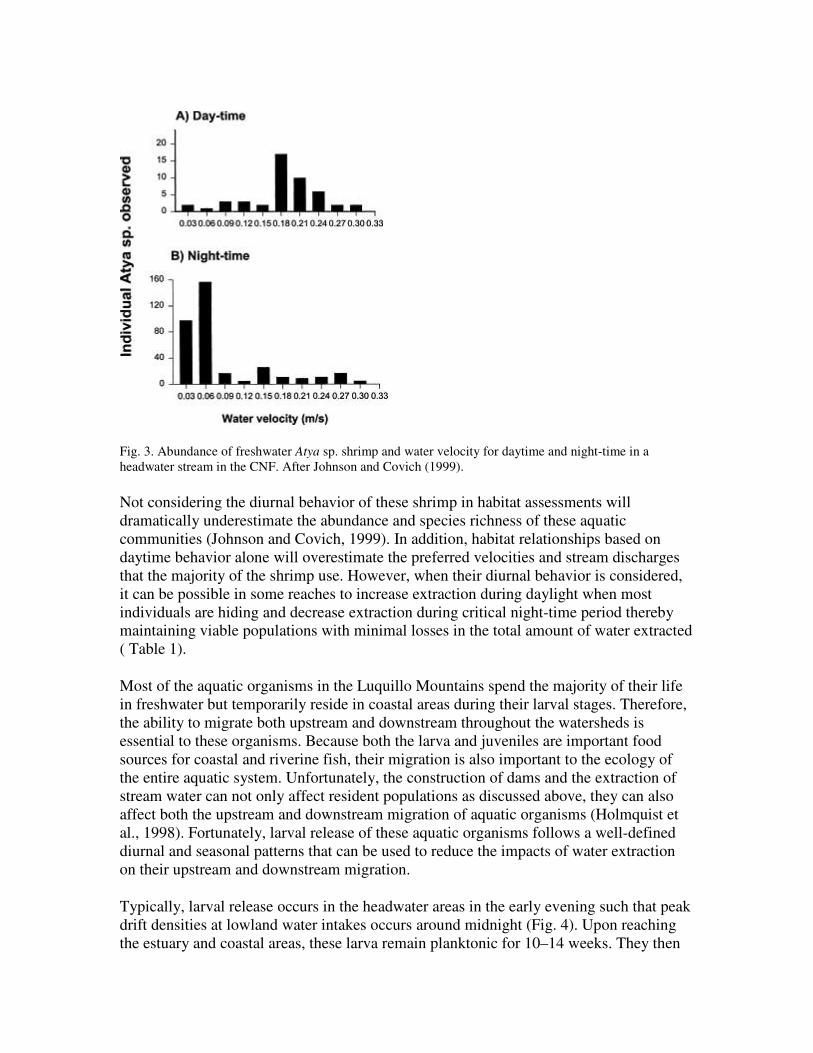
Fig. 3. Abundance of freshwater Atya sp. shrimp and water velocity for daytime and night-time in a headwater stream in the CNF. After Johnson and Covich (1999).
Not considering the diurnal behavior of these shrimp in habitat assessments will dramatically underestimate the abundance and species richness of these aquatic communities (Johnson and Covich, 1999). In addition, habitat relationships based on daytime behavior alone will overestimate the preferred velocities and stream discharges that the majority of the shrimp use. However, when their diurnal behavior is considered, it can be possible in some reaches to increase extraction during daylight when most individuals are hiding and decrease extraction during critical night-time period thereby maintaining viable populations with minimal losses in the total amount of water extracted ( Table 1).
Most of the aquatic organisms in the Luquillo Mountains spend the majority of their life in freshwater but temporarily reside in coastal areas during their larval stages. Therefore, the ability to migrate both upstream and downstream throughout the watersheds is essential to these organisms. Because both the larva and juveniles are important food sources for coastal and riverine fish, their migration is also important to the ecology of the entire aquatic system. Unfortunately, the construction of dams and the extraction of stream water can not only affect resident populations as discussed above, they can also affect both the upstream and downstream migration of aquatic organisms (Holmquist et al., 1998). Fortunately, larval release of these aquatic organisms follows a well-defined diurnal and seasonal patterns that can be used to reduce the impacts of water extraction on their upstream and downstream migration.
Typically, larval release occurs in the headwater areas in the early evening such that peak drift densities at lowland water intakes occurs around midnight (Fig. 4). Upon reaching the estuary and coastal areas, these larva remain planktonic for 10–14 weeks. They then
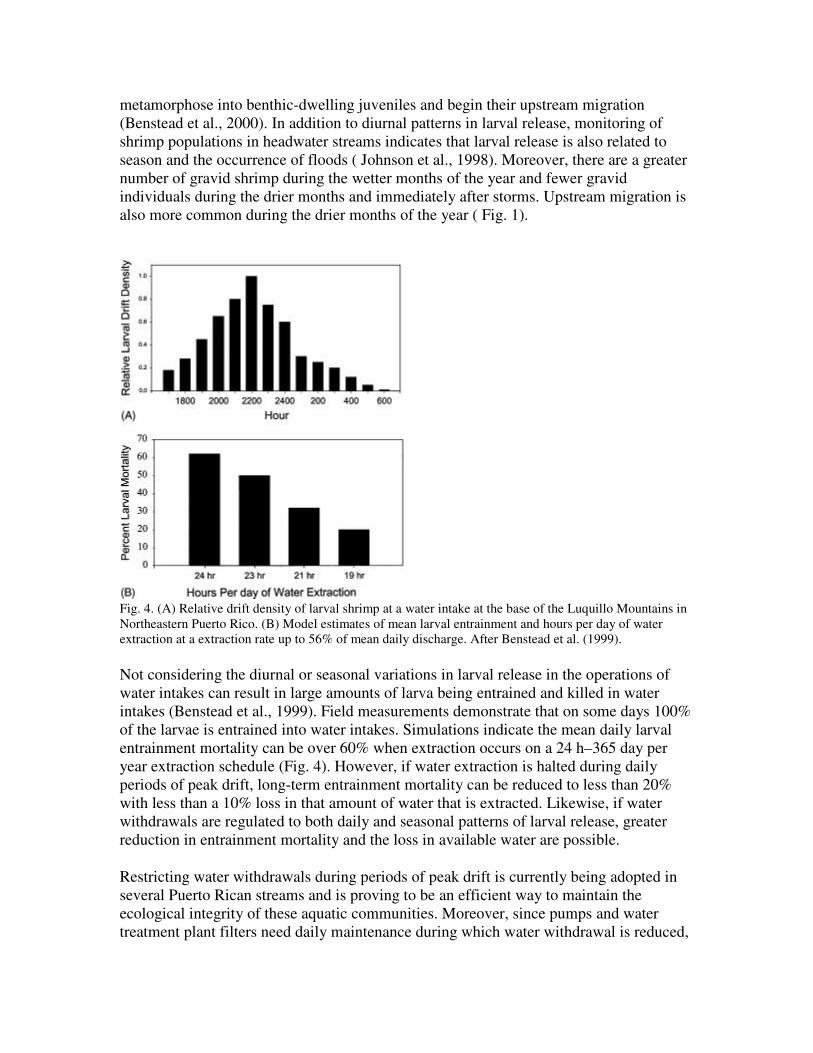
metamorphose into benthic-dwelling juveniles and begin their upstream migration (Benstead et al., 2000). In addition to diurnal patterns in larval release, monitoring of shrimp populations in headwater streams indicates that larval release is also related to season and the occurrence of floods ( Johnson et al., 1998). Moreover, there are a greater number of gravid shrimp during the wetter months of the year and fewer gravid individuals during the drier months and immediately after storms. Upstream migration is also more common during the drier months of the year ( Fig. 1).
Fig. 4. (A) Relative drift density of larval shrimp at a water intake at the base of the Luquillo Mountains in Northeastern Puerto Rico. (B) Model estimates of mean larval entrainment and hours per day of water extraction at a extraction rate up to 56% of mean daily discharge. After Benstead et al. (1999).
Not considering the diurnal or seasonal variations in larval release in the operations of water intakes can result in large amounts of larva being entrained and killed in water intakes (Benstead et al., 1999). Field measurements demonstrate that on some days 100% of the larvae is entrained into water intakes. Simulations indicate the mean daily larval entrainment mortality can be over 60% when extraction occurs on a 24 h–365 day per year extraction schedule (Fig. 4). However, if water extraction is halted during daily periods of peak drift, long-term entrainment mortality can be reduced to less than 20% with less than a 10% loss in that amount of water that is extracted. Likewise, if water withdrawals are regulated to both daily and seasonal patterns of larval release, greater reduction in entrainment mortality and the loss in available water are possible.
Restricting water withdrawals during periods of peak drift is currently being adopted in several Puerto Rican streams and is proving to be an efficient way to maintain the ecological integrity of these aquatic communities. Moreover, since pumps and watertreatment plant filters need daily maintenance during which water withdrawal is reduced,

timing these maintenance activities to the diurnal behavior of these organisms can be a very efficient way to reduce impacts on the aquatic community without greatly affecting municipal water supplies. Nevertheless, this technique is not without problems. First, because of differences in travel time between headwater streams and water intakes, the average timing of peak larval drift will vary within and between watersheds and must be evaluated for each intake. Secondly, although water withdrawals are usually limited for several hours per day to perform routine plant maintenance, senior plant mangers usually preform this maintenance during daylight working hours. Changing their work schedule to time these maintenance activities to the night-time migration can increase the cost of labor and supervision.
3.1.3. Sewage treatment
Nearly 60% of all water delivered to municipal users in Puerto Rico eventually passes through sewage treatment plants and is discharged as sewage effluent (Dopazo and Molina-Rivera, 1995). The management of this nutrient rich effluent can be a costly and complex endeavor. In Puerto Rico, the standard practice has been secondary treatment of raw sewage and the subsequent release of the effluents into lower elevation reaches of streams that are at, or near, a highly stratified estuary. In undisturbed conditions the water leaving the highlands has relatively low nutrient concentrations ( McDowell and Asbury, 1994). In contrast, the practice of releasing N- and P-rich sewage effluent waters into lower elevation streams can result in eutrophic water bodies ( Kent, 1996 and Kent). This can be extremely severe in water bodies that also receive non-point urban runoff and are unnaturally low because of upstream water diversions. Fortunately, diurnal and seasonal variations in the biogeochemistry of these streams provide opportunities to reduce the impacts of these nutrient releases.
In many of the lower elevation streams in Puerto Rico, dissolved oxygen concentrations follow a well-defined diurnal pattern (Fig. 5; Ortiz-Zaya, 1998). The highest oxygen levels are found during the day when the submerged aquatic plants are producing oxygen during photosynthesis. The lowest dissolved oxygen values are found at night when oxygen is being consumed during the decomposition of organic matter. In addition to this diurnal dissolved oxygen cycle, there exist a seasonal pattern in the dissolved oxygen profiles and the abundance of submerged aquatic vegetation (personal observation). In general, the lowest dissolved oxygen values are found during the low flow periods of the late summer months when the lower elevation water bodies are covered with submerged aquatic vegetation ( Fig. 1). Higher dissolved oxygen values are found during wetter months of the year after large storms have remove the submerge aquatics from the channels and water temperatures are at their seasonal lows.
Knowledge of these annual and diurnal dissolved oxygen cycles provides opportunities to minimize the impacts of effluent releases. Moreover, reducing effluent discharges during low oxygen night-time periods and increasing releases during the early part of the day when the assimilation capacity of the receiving water bodies are at their highest can potentially reduce some eutrophication. In contrast, only sampling dissolved oxygen concentrations during daylight hours will result in unrealistically high estimates of the

assimilation capacity of these water bodies. Likewise, releasing effluent 24 h per day on the basis of daytime measurements of assimilation capacity will also result in unexpected eutrophication.
Fig. 5. Diurnal temperature and dissolved oxygen concentrations by time of day for the lower Rio Mameyes, Puerto Rico (Scatena, unpublished).
Like all management activities, timing effluent releases to diurnal oxygen cycles is not without problems. As effluent is continually being generated, variable releases with a constant supply create the need to temporarily store sewage or have alternate disposal sites. In many areas in Northeastern Puerto Rico, the effluent can be used for the irrigation of golf courses (Rio Mar Golf Course, pers. commun.). In other areas, wetland release, or the irrigation of degraded lands are possible.
4. Discussion
4.1. Potential management applications of other ecological rhythms
As the above examples illustrate timing management activities with ecological rhythms can be an efficient way to reduce adverse impacts of some land management activities. In addition to those described above, many other examples and possibilities exist. These include timing management activities to temporal patterns in recreational use (Fig. 2), plant phenology, growth cycles of native and non-native species, lunar cycles, El Niño events, and decadal scale climatic variability (Table 2). Links between climate and populations of insect related tropical diseases have also been established elsewhere and can be used to predict epidemics several months in advance (Epstein, 1999 and

Linthicum). Likewise, relationships between temperature, rainfall, river flow, tidal cycles, and the abundance of fish and shrimp populations have also been identified and can be used to manage fishing pressure ( Goulding, 1984 and Teicher-Coddington, 1999).
Table 2. Examples of ecological rhythms and life history traits that can potentially be used for in tropical forest management
4.2. Constraints to managing with ecological rhythms
While timing management activities to the ecological rhythms that occur between disturbances has definable benefits, this type of dynamic management is not without costs or tradeoffs. For example, in an effort to reduce potentially disturbing noise during the parrot breeding season, construction activities in the CNF must be done under an accelerated pace during the rainier periods of the year. As a consequence, soil erosion and or construction costs may increase. Likewise, the cost of lost water withdrawals or sewage releases caused by night-time reduction in operations can be offset by scheduling daily pump and filter maintenance during the ecologically sensitive night-time periods. However, this often requires that senior plant managers work in evening rather than daily periods and therefore can increase administrative costs. Additional water storage facilities and complex distribution systems that can be managed on a dynamic basis may also be needed.
One of the largest problems associated with mandating dynamic management systems based on ecological rhythms concerns compliance monitoring. Similar problems have been associated with effluent taxes and tradable effluent permits (Brookshire and Neill,

1992). Moreover, regulatory agencies have been hesitant to approve complex operation schedules that follow ecological rhythms or market-based effluent trading schemes because they are difficult to monitor and regulate. Fortunately, recent advantages in automatic environmental data acquisition systems can greatly increase the ability to conduct complex real-time, spatially explicit monitoring ( Scatena, 2000). Moreover, inexpensive automatic water level recorders, telemetering systems, and water quality sensors now allow continual, real-time monitoring of both management operations and important ecological processes. Although the technology exist, identifying relationships between ecological processes and environmental conditions and then developing monitoring systems and operation schemes that integrate these processes into management is a major challenge in both tropical and temperate environments.
Regardless of the monitoring technology used, the success of ecologically based management schemes requires specific knowledge of the response of ecosystems and organisms to environmental conditions. A generic problem encountered when initiating this type of management is the lack of ecosystem-specific information on the spatial and temporal variability of ecological and pollution generating processes (Graham et al., 1991). This need for site-specific knowledge can be expected to increase as tropical forests are increasingly used for multiple purposes, like water supplies ( Postel, 1998), carbon and nutrient sequestration ( DeLucia et al., 1999) and wastewater recycling ( Sopper and Kardos, 1973).
The success of processes-based dynamic management also requires institutional memory and an administrative commitment to long-term environmental monitoring, data analysis and synthesis. A major challenge for watershed management in the 21st century will be the development of spatially explicit analytical methods and institutional structures that rapidly synthesize information about environmental conditions so managers can make informed, defensible decisions in a timely fashion (Scatena, 2000). This will be especially true in a changing global environment where community structure and ecological rhythms are also changing ( Wuethrich, 2000).
4.3. Research needs and opportunities
Public and private tropical forests will continue to be managed for multiple uses including water supplies, timber products, recreation, mining, fisheries, grazing, and the conservation of biodiversity. How these forests will be managed in this increasingly competitive, multi-use environment will be affected by local environmental change, technological change, social and administrative considerations? Nevertheless, conducting mechanistic studies to identify and understand stand scale ecological rhythms in humid tropical forests will be essential to develop ecosystem-based management strategies and operating procedures.

Acknowledgements
This research was supported by the USDA Forest Service with additional support from the National Science Foundation through the Luquillo Long-term Ecological Research Program and the University of Puerto Rico. The reviews by A.E. Lugo and two anonymous reviewers are greatly appreciated and significantly improved the manuscript.
References
Balogh, J.C., Walker, W.J., 1992. Golf Course Management and Construction. Environmental Issues. Lewis Publishers, Chelsea, MN, 951 pp.
Benstead, J.P., March, J.G., Pringle, C.M. and Scatena, F.N., 1999. Effects of a low-head dam and water abstraction on migratory tropical stream biota. Ecol. Appl. 9 2, pp. 656–668.
Benstead, J.P., March, J.G. and Pringle, C.M., 2000. Estuarine larval development and upstream post-larval migration of freshwater shrimp in two tropical rivers of Puerto Rico. Biotropica 32 3, pp. 545–548.
Bistal, G.A. and Ruff, J.D., 1996. Quality of 1995 spring total dissolved gas data: Columbia and Snake rivers. Water Resour. Bull. 32 6, pp. 1177–1186.
Black, D.E., Peterson, L.C., Overpeck, J.T., Kaplan, A., Evans, M.N. and Kashgarian, L.P., 1999. Eight centuries of North Atlantic Ocean atmosphere variability. Science 286, pp. 1709–1713.
Boose, E.R., Foster, D.R. and Fluet, M., 1994. Hurricane impacts to tropical and temperate forest landscapes. Ecol. Monogr. 64, pp. 369–400
Brookshire, D.S. and Neill, H.R., 1992. Benefit transfers: conceptual and empirical issues. Water Resour. Res. 28 3, pp. 651–655.
Caribbean National Forest, 1997. Final environmental impact statement for the revised land and resource management plan, Appendix F. Puerto Rican Parrot Management Situation. USDA Forest Service, Southern Region, 1997.
Covich, A.P., McDowell, W.H., 1996. The stream community. In: Reagan, D.P., Waide, R.B. (Eds.), The Food Web of a Tropical Rain Forest. University of Chicago Press, Chicago, 616 pp. (Chapter 13).
Curran, L.M., Caniago, I., Paoli, G.D., Astiant, D., Kusneti, M., Leighton, M., Nirarita, C.E. and Haeruman, H., 1999. Impact of the El Niño and logging on canopy tree recruitment in Borneo. Science 286, pp. 2184–2188.

DeLucia, E.H., Hamilton, J.G., Naidu, S.L., Thomas, R.B., Andrews, J.A., Finzi, A., Lavive, M., Matamala, R., Mohan, J.E., Hendrey, G.R. and Schlesinger, W.H., 1999. Net primary production of a forest ecosystem with experimental CO2 enrichment. Science 284, pp. 1177–1179.
Dopazo, T., Molina-Rivera, W.L., 1995. Estimated water use in Puerto Rico, 1988–1989. US Geological Survey Open-file Report 95-380, 31 pp.
Epstein, P.R., 1999. Climate and health Science 285, p. 348.
Goulding, M., 1984. Forest fish of the Amazon. In: Prance, G.T., Lovejoy, T.E. (Eds.), Amazonia, Key Environments. Pergamon Press, New York, pp. 267–276 (Chapter 14).
Graham, R.L., Hunsaker, C.T., O’Neill, R.V. and Jackson, B.L., 1991. Ecological risk assessment at the regional scale. Ecol. Appl. 1 2, pp. 196–206.
Gray, W.M., Sheaffer, J.D., Landsea, C.W., 1997. Climate trends associated with multidecadal variability of Atlantic hurricane activity. In: Diaz, H.F., Pulwarty, R.S. (Eds.), Hurricanes Climate and Socioeconomic Impacts. Springer, Berlin, pp. 15–53 (Chapter 2).
Holmquist, J.G., Schmidt-Gengenbach, J.M. and Yoshioka, B.B., 1998. High dams and marine–freshwater linkages: effects on native and introduced fauna in the Caribbean. Conserv. Biol. 12, pp. 621–630.
James, T.R. and Havens, K.E., 1996. Algal bloom probability in a large subtropical lake. Water Resour. Bull. 32 5, pp. 995–1006.
Johnson, S.L. and Covich, A.P., 1999. The importance of night-time observations for determining habitat preferences of stream biota. Regul. River. 16, pp. 91–99.
Johnson, S.L., Covich, A.P., Crowl, T.A., Estrada, A., Bithorn, J. and Wurtsbaugh, W.A., 1998. Do seasonality and disturbance influence reproduction in freshwater atyid shrimp in headwater streams in Puerto Rico?. Proc. Int. Assoc. Theoret. Appl. Limnol. 26, pp. 2076–2081.
Kaufmann, M.R., Graham, R.T., Boyce, D.A., Moir, W.H., Perry, L., Reynolds, R.T., Bassett, R.L., Mehlhop, P., Edminster, C.B., Block, W.M., Corn, P.S., 1994. An Ecological basis for ecosystem management. General Technical Report RM-246, USDA Forest Service, 22 pp.
Kent, R., 1996. Seedling survival and colonizing vegetation in wetland plots receiving pig wastes in Puerto Rico. M.S. Thesis. Department of Environmental Engineering Sciences, University of Florida, Gainesville, 328 pp.

Kent, R., Odum, H.T. and Scatena, F.N., 2000. Eutrophic overgrowth in the self-organization of tropical wetlands illustrated with a study of swine wastes in rainforest plots. Ecol. Eng. 16, pp. 255–269.
Linthicum, K.J., Anyamba, A., Tucker, C.J., Kelley, P.W., Myers, M.F. and Peters, C.J., 1999. Climate and satellite indicators to forecast Rift valley fever epidemics in Kenya. Science 285, pp. 397–400.
Lugo, A.E., Lowe, C. (Eds.), 1995. Tropical Forests: Management and Ecology. Springer, Berlin, New York.
Lugo, A.E., Scatena, F.N., 1995. Ecosystem-level properties of the Luquillo experimental forest with emphasis on the tabonuco forest. In: Lugo, A.E., Lowe, C. (Eds.), Tropical Forests: Management and Ecology. Springer, Berlin, New York.
March, J.G., Benstead, J.P., Pringle, C.M. and Scatena, F.N., 1998. Migratory drift of larval freshwater shrimps in two tropical streams, Puerto Rico. Freshw. Biol. 40, pp. 261–273.
Matson, P.A., Naylor, R. and Ortiz-Monasterio, I., 1998. Integration of environmental, agronomic, and economic aspects of fertilizer management. Science 280, pp. 112–115.
McDowell, W.H. and Asbury, C.E., 1994. Export of carbon, nitrogen, and major ions from three tropical montane watersheds. Limnol. Oceanogr. 39, pp. 111–125.
Neeling, J.D., Latif, M., 1998. El Niño dynamics. Physics Today (December), 32–36.
Odum, E.P., 1971. Fundamentals of Ecology, 3rd Edition. Saunders, Philadelphia, PA, 545 pp.
Odum, H.T., 1996. Tropical forest systems and the human economy. In: Lugo, A.E., Lowe, C. (Eds.), Tropical Forests: Management and Ecology, Ecological Studies, Vol. 112. Springer, Berlin, pp. 343–396 (Chapter 14).
Odum, H.T., Pigeon, R.F., 1970. A tropical rain forest: a study of irradiation and ecology at El Verde, Puerto Rico. Division of Technical Information, US Atomic Energy Commission, Washington, DC.
Ortiz-Zaya, J.R., 1998. The metabolism of the Rio Mameyes, Puerto Rico: carbon fluxes in a tropical rain forest river. Ph.D. Thesis. University of Colorado, p. 344.
Postel, S.L., 1998. Water for food production: will there be enough in 2025?. Bioscience48 8, pp. 629–637.
Pringle, C.M. and Hamazaki, T., 1997. Effects of fishes on algal response to storms in a tropical stream. Ecology 78, pp. 2432–2442.

Reagan, D.P., Waide, R.B., 1996. The Food Web of a Tropical Rain Forest. University of Chicago Press, Chicago, 616 pp.
Scatena, F.N., 2000. Future trends and research needs in managing forests and grasslands as drinking water sources. Chapter 12, pages 197–201. In: Dissmeyes G.E. (Ed.), Drinking Water from Forests and Grasslands. USFS General Technical Report SRS-39, 246 pages.
Scatena, F.N. and Larsen, M.C., 1991. Physical aspects of hurricane Hugo in Puerto Rico. Biotropica 23, pp. 317–323.
Scatena, F.N. and Lugo, A.E., 1995. Geomorphology, disturbance, and the soil and vegetation of two subtropical wet steepland watersheds of Puerto Rico. Geomorphology13, pp. 199–213.
Scatena, F.N., Johnson, S.L., 2001. Instream-flow Analysis for the Luquillo Experimental Forest Puerto Rico: Methods and Analysis. General Technical Report, USDA Forest Service, in press.
Schmidt, K.F., 1997. No-take zones spark fisheries debate. Science 277, pp. 489–491.
Snyder, N.F.R., Wiley, J.W., Kepler, C.B., 1987. The parrots of Luquillo: natural history and conservation of the Puerto Rican Parrot. Western Foundation of Vertebrate Zoology, Los Angles, CA, 384 pp.
Sopper, W.E., Kardos, L.T. (Eds.), 1973. Recycling Treated Municipal Wastewater and Sludge through Forest and Cropland. Pennsylvania State University Press, University Park, 479 pp.
Sutton, R.J., Miller, W.J. and Patti, S.J., 1997. Application of the instream flow incremental methodology to a tropical river in Puerto Rico. Rivers 6 1, pp. 1–9.
Teicher-Coddington, D., 1999. Shrimp farming in southern Honduras: a case for sustainable production. In: Hatch, L.U., Swisher, M.E. (Eds.), Managed Ecosystems. The Mesoamerican Experience. Oxford University Press, Oxford, pp. 222–230 (Chapter 25).
Vogt, K.A., Beard, K.H., Hammann, S., O’Hara, J.L., Vogt, D.J., Scatena, F.N., Parry, B.A., 2000. Carbon fluxes in plants correlate with lunar cycles: Sierra Palm case study, in review.
Wadsworth, F.H., 1997. Forest Production for Tropical America. Agriculture Handbook, Vol. 710, USDA Forest Service, 563 pp.
Walker, L.R., Silver, W.L., Willig, M.R., Zimmerman, J.K., 1996. Long-term Response of Caribbean Ecosystems to Disturbance (Special Issue). Biotropica 28, (4a) 613 pp.

Wang, H.H., Scatena, F.N., 2000. Regeneration dynamics of big-leaf and hybrid mahogany plantations in Puerto Rico following hurricane disturbance. In: Lugo, A.E. (Ed.), Management of Mahogany, in press.
Woods, P., 1989. Effects of logging, drought, and fire on structure and composition of tropical forests in Sabah, Malaysia. Biotropica 21 4, pp. 290–298.
Wootton, J.T.M., Parker, M.S. and Power, M.E., 1996. Effects of disturbance on river food webs. Science 273, pp. 1558–1561.
Wuethrich, B., 2000. How climate change alters rhythms of the wild?. Science 287, pp. 793–795.
Wunderle, J.M., Wiley, J.W., 1996. Effects of hurricanes on wildlife: implications and strategies for management. In: DeGraff, R.M., Miller, R.I. (Eds.), Conservation of Faunal Diversity in Forested Landscapes. Chapman & Hall, London, pp. 253–264 (Chapter 9).