ecological evaluation of transitional and coastal waters...
TRANSCRIPT

Medit. Mar. Sci., 2/2, 2001, 45-65 45
Ecological evaluation of transitional and coastal waters:A marine benthic macrophytes-based model
S. ORFANIDIS1, P. PANAYOTIDIS2 and N. STAMATIS1
1 National Agricultural Research Foundation, Fisheries Research Institute, 640 07 Nea Peramos,Kavala, Greecee-mail: [email protected]
2 National Center for Marine Research, 16604 Athens, Greece
Abstract
A model to estimate the ecological status and identify restoration targets of transitional and coastal waterswas developed. Marine benthic macrophytic species (seaweeds, seagrasses) were used to indicate shifts in theaquatic ecosystem from the pristine state with late-successional species (Ecological State Group I) to thedegraded state with opportunistic (ESG II) species. The first group comprises species with a thick or calcareousthallus, low growth rates and long life cycles (perennials), whereas the second group includes sheet-like andfilamentous species with high growth rates and short life cycles (annuals). Seagrasses were included in thefirst group, whereas Cyanophyceae and species with a coarsely branched thallus were included in the secondgroup.
The evaluation of ecological status into five categories from high to bad includes a cross comparison ina matrix of the ESGs and a numerical scoring system (Ecological Evaluation Index). The model could allowcomparisons, ranking and setting of priorities at regional and national levels fulfilling the requirements of theEU Water Frame Directive. A successful application of the model was realized in selected lagoons of theMacedonian and Thrace region (North Greece) and in the Saronic Gulf coastal ecosystems (Central Greece).
Key words: Ecological status, Water quality, Phytobenthos, Seaweeds, Seagrasses, Functionalgroups, Water Framework Directive
Introduction
Mediterranean Marine Science
Vol. 2/2, 2001, 45-65
The term "classes of ecological status" isused in the text of the EU frame Directive forWater Policy (WFD, 2000/60/EC) in order todescribe the degree of human impact on thebiological communities living in a water body.Five classes of quality (high, good, moderate,low and bad) are foreseen, the high class
reflecting pristine, undisturbed conditions ofthe ecosystem.
The concept of "ecological status" (orecological quality status) was elaborated duringthe early 90’s, to be used in a new frame ofEuropean water policy. For the purposes of theDirective "ecological water quality is an overall

expression of the structure and function of thebiological community taking into accountnatural physiographic, geographical andclimatic factors as well as physical and chemicalconditions, including those resulting fromhuman activities" (EEC, 1994).
Transitional and coastal waters are someof the most productive ecological systems onEarth and are recognized to be of extremelyhigh "value" to human society. However, theyare being severely threatened by anthropogenicpressure and climate change induced sea levelrise (CROOKS & TURNER, 1999). For themanagement of these ecosystems it is criticalto identify the key biological signals (impacts)that indicate the intensity of anthropogenicstress or ecological status.
Cumulative evidence indicates that impactsare best investigated at the population orcommunity level (LOBBAN & HARRISON,1994; CROWE et al., 2000) and this requiresan approach that integrates an ecologicalassessment into the more traditional chemicaland physical evaluation (GIBSON et al., 2000).However, the diagnosis of the ecological statusis often a difficult task because of spatial andtemporal variability in community features asa result of changes in physical and chemicalconditions (ORFANIDIS et al., 2001). Onepossibility for overcoming this complexity isto study communities from a functional pointof view (groups of functionally similar species).At a functional level, communities appear tobe much more temporally stable andpredictable than when examined at the specieslevel (STENECK & WALTING, 1982;STENECK & DETHIER, 1994). For example,anthropogenic stress shifts the communitystructure towards dominance of opportunisticspecies (BOROWITZKA M. A., 1972;REGIER & COWELL, 1972).
Marine benthic macrophytes (phyto-benthos) are mentioned in the WFD as a"quality element" for the classification ofmarine coastal areas. They include twofundamentally different groups of plants, theseaweeds (macroscopic algae) and the
seagrasses (vascular plants). These macrophytesform the structural base for some of the mostproductive ecosystems in the world (MANN,1973; McROY & LLOYD, 1981), includingrocky and soft bottom intertidal and subtidalzones, coral reefs, lagoons and salt marshes.
The three major taxonomic groups ofseaweeds, Chlorophyceae, Phaeophyceae andRodophyceae, although representing distinctevolutionary lines show similar ranges ofmorphologies. It would thus seem likely thatthis similarity of form is adaptive, conferringfitness on phyletically diverse organismsgrowing in a common habitat. The recognitionof the importance of morphology has led toecological classifications of seaweeds basedon thallus morphology, longevity and lifehistory (FELDMANN, 1951; CHAPMAN &CHAPMAN, 1976; RUSSEL, 1977). Morerecently, LITTLER & LITTLER (1980) haveproposed a functional-form model. This modelwas tested and verified experimentally: thefunctional characteristics of plants, such asphotosynthesis, nutrient uptake, and grazersusceptibility, are related to morphology andsurface area: volume ratios (LITTLER, 1980;LITTLER & ARNOLD, 1982; LITTLER &LITTLER 1984). Seaweeds and seagrassescomprise two evolutionary and physiologicallydifferent groups (LARKUM et al., 1989;HEMMINGA & DUARTE, 2000; LOBBAN& HARRISON, 1994) but have often beenexamined together because of morphological-functional similarities and the apparentoverlap in habitats.
Because marine benthic macrophytes aremainly sessile organisms, they respond directlyto the abiotic and biotic aquatic environment,and thus represent sensitive indicators of itschanges. A good example is water eutro-phication. It is well documented that elevatedconcentrations of nitrogen and phosphorus inthe water column do not necessarily indicatehighly eutrophic conditions, neither do lowconcentrations necessarily indicate absence ofeutrophication (CLOERN, 2001). The reasonis that nutrient concentrations in the water
46 Medit. Mar. Sci., 2/2, 2001, 45-65

column are related to nutrient load as well asto other biological and chemical processes. Areliable signal of increasing eutrophication isthe replacement of late successional, perennialseaweeds, like Cystoseira spp. and Fucus spp. byopportunistic species like Ulva spp. andEnteromorpha spp. (HARLIN, 1995;SCHRAMM & NIENHUIS, 1996;SCHRAMM, 1999). Several examples of impactsof anthropogenic stress on marine phytobenthiccommunities are shown in Table 1.
Marine benthic macrophytes, in particularseagrasses, also provide substrate, habitat andshelter for plants and animals, includingeconomically important species (HARMELIN-VINIEN et al., 1995; POLLARD, 1984;EDGAR, 1999 a, b). Since the canopy of leavesdiminishes wave energy and currents(FONSECA & CALAHAN, 1992), they alsosignificantly affect sediment stability(FONSECA, 1996) and the retention ofparticles (BULTHUIS et al., 1984; DAUBYet al., 1995).
The appliance of the WFD obligates all themembers of European community first toevaluate the ecological status and then toidentify restoration targets of their transitionaland coastal waters. The aim of this paper was(1) to develop a model for the estimation ofecological status and identification ofrestoration targets of transitional and coastalwaters, and (2) to apply this model in selectedlagoons of the Eastern Macedonia and Thraceregion (North Greece) and in the Saronic Gulf(Central Greece) coastal waters.
Materials and MethodsA. Development of the model
In the context of developing methodologysuitable for evaluation of ecological status intransitional and coastal waters, a series of stepswas followed:
(1) Identification of the basic concept thatbest describes the impact of anthropogenic
stress to community from a synthesis of existingliterature in Table 1.
(2) Identification a set of bio-indicatorsthat respond directly to anthropogenic stress.A synthesis of existing literature in Table 1.
(3) Identification of functional groups thatpredict function of communities in pristineand degraded ecosystems. The existence ofthe functional-form model for seaweeds(LITTLER, 1980) and its relevance fordescribing functional characteristics ofecosystem, e.g. productivity, was used as a basicmilestone. However, a new synthesis thatincorporates seagrasses with seaweeds in afunctional type scheme was needed.
(4) Development of a numerical scoringsystem that expresses the ecosystem status toa number. A good example of this procedurewas the methodology followed by BRICKERet al. (1999) to assess eutrophic conditions inUSA estuaries.
B. Case studies
The model was tested in selected lagoonsof the Macedonian and Thrace region (NorthGreece) and in the Saronic coastal ecosystems(Central Greece).
Eastern Macedonian and Thrace lagoons
The investigated lagoons are located in theEastern Macedonia and Thrace region, whereone of the most extensive Greek fresh water-estuarine systems exists. They belong to theNestos River (Vassova, Eratino, Agiasma,Keramoti) and Vistonida estuarine system(Fanari) catchments. Their value in ecological(Natura 2000 network) and economic (fishaquaculture) terms is high. Along the westerncoasts of the Nestos River Delta a series offour elongated bar-built lagoons exist (Figure1). They consist of a shallow (up to 1.5 m) areaand several artificially constructed channels(up to 3 m in depth). The fresh water sourcesof the lagoons are mainly agricultural run-offs
Medit. Mar. Sci., 2/2, 2002, 00-00 47

48 Medit. Mar. Sci., 2/2, 2001, 45-65
Ant
hrop
ogen
icB
enth
icIm
pact
Lit
erat
ure
stre
ssm
acro
phyt
esE
utro
phic
atio
nSe
awee
dsD
omin
ance
of o
ppor
tuni
stic
spec
ies,
HA
RIT
ON
IDIS
(197
8), D
IAPO
UL
IS &
HA
RIT
ON
IDIS
(198
7),
seaw
eed
bloo
ms,
dec
line
of d
iver
sity
CH
RY
SSO
VE
RG
IS &
PA
NA
YO
TID
IS (1
995)
, LA
ZA
RID
OU
et a
l.(1
997)
, SC
HR
AM
M &
NIE
NH
UIS
(199
6), S
CH
RA
MM
(199
9), L
OT
ZE
et a
l.(1
999)
, LO
TZ
E &
SC
HR
AM
M (2
000)
Seag
rass
esL
arge
scal
e an
d re
gion
al d
eclin
e of
mea
dow
s,L
AR
KU
M e
t al.
(198
9), H
EM
MIN
GA
& D
UA
RT
E (2
000)
dom
inan
ce o
f fle
shy
seaw
eeds
Org
anic
mat
ter,
Seaw
eeds
Lig
ht re
duct
ion
and
alte
ratio
n of
har
d su
bstr
ate
LO
BB
AN
& H
AR
RIS
ON
(199
4)Si
ltatio
naf
fect
s com
mun
ity st
ruct
ure
Seag
rass
esD
eclin
e of
mea
dow
s thr
ough
redu
ctio
n of
ligh
tH
EM
MIN
GA
& D
UA
RT
E (2
000)
and
accu
mul
atio
n of
org
anic
mat
ter i
n se
dim
ent
Hea
vy m
etal
sSe
awee
dsIn
hibi
tion
of re
prod
uctio
n an
d de
velo
pmen
t,L
OB
BA
N &
HA
RR
ISO
N (1
994)
, CO
EL
HO
et a
l. (2
000)
,ch
ange
s in
com
mun
ity st
ruct
ure
CR
OW
E e
t al.
(200
0)Se
agra
sses
No
dire
ct e
ffec
t has
bee
n ob
serv
ed
LA
RK
UM
et a
l.(1
989)
Oil
split
sSe
awee
dsSh
ort t
erm
gro
wth
redu
ctio
n in
inte
rtid
al sp
ecie
sL
OB
BA
N &
HA
RR
ISO
N (1
994)
Seag
rass
esN
o di
rect
eff
ect h
as b
een
docu
men
ted
Glo
bal w
arm
ing
Seaw
eeds
Cha
nges
in d
istr
ibut
ion
patt
erns
are
exp
ecte
dB
RE
EM
AN
(199
0), P
AK
KE
R &
BR
EE
MA
N (1
994)
Seag
rass
esC
hang
es in
dis
trib
utio
n pa
tter
ns a
re e
xpec
ted
HE
MM
ING
A &
DU
AR
TE
(200
0)In
crea
seSe
awee
dsF
urth
er e
xpan
sion
in e
stua
rine
eco
syst
ems
LO
BB
AN
& H
AR
RIS
ON
(199
4)of
salin
itySe
agra
sses
Spec
ies d
ispl
acem
ent,
e.g.
Cym
odoc
eain
stea
d of
Rup
pia
KA
ME
RM
AN
S et
al.
(199
9), p
rese
nt st
udy
Tra
wlin
gSe
awee
dsD
amag
e of
subl
ittor
al st
ands
BL
AD
ER
et a
l. (2
000)
Fis
hing
Seag
rass
esF
ragm
enta
tion
- dec
line
of m
eado
ws
SAN
CH
EZ
-JE
RE
Z &
RA
MO
S (1
996)
, BL
AD
ER
et a
l. (2
000)
Tab
le 1
Exa
mpl
es o
f im
pact
of a
nthr
opog
enic
str
ess
on m
arin
e be
nthi
c m
acro
phyt
ic c
omm
unit
ies.

coming in from surrounding drainage channelsand the old bed of the Nestos River. TheFanari lagoon consists of a uniform shallowarea (up to 2 m in depth) having a narrowconnection to the sea (Figure 1). The mainfresh water sources of the lagoon are theautumn-winter rainfalls.
The studied lagoons were visited betweensummer 1999 and summer 2001. The lagoonsof Agiasma and Keramoti were visited inAugust 2000; the lagoon of Eratino was visitedin July 1999 and in winter 2001; the lagoon ofVassova was visited in summer 2000 and inwinter 2001; the lagoon of Fanari was visitedin November 2000 and in July 2001. In eachlagoon representative stations (32 in total)were sampled taking into account hydrologicaland morphological characteristics (Figure 1).
At stations with obvious patchiness of thebenthic vegetation a second relevé (55 in total)was taken for verification. The sampling wasdestructive, by using a 30 cm x 50 cm x 100 cm(width x length x height) metallic stainlessframe and a net. Each sample was carefullysorted and identification at species level,except Cyanophyceae mats, was attempted.For the identification and classification ofspecies the following literature was used:COPPEJANS (1983), ATHANASIADIS(1987), BURROWS (1991), RIBERA et al.(1992), GALLARDO et al. (1993), MAGGS& HOMMERSAND (1993), LAZARIDOU(1994) and FLETCHER (1995). Theabundance of species was estimated as %coverage in the sampling area (15 cm2 = 1%of the sampling area) in horizontal projection
Medit. Mar. Sci., 2/2, 2001, 45-65 49
Fig. 1: A GIS-based map of the studied lagoons in the Eastern Macedonia and Thrace region, NorthGreece (St. = sampling station).

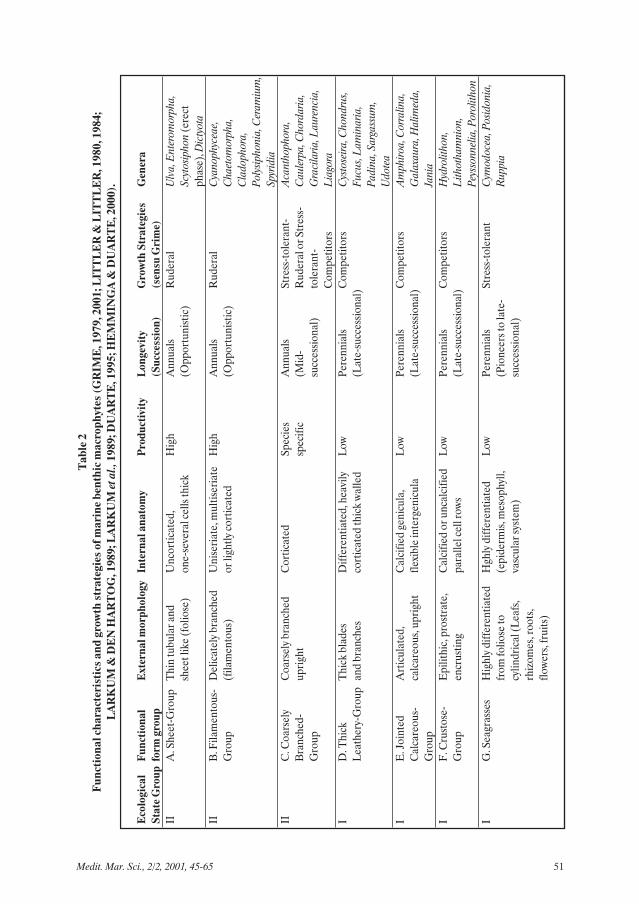
(BOUDOURESQUE, 1971; VERLAQUE,1987). Species were classified into functionalform groups (FFg) according to LITTLER &LITTLER (1980, 1984).
Saronic Gulf
The Saronic Gulf is an embayment of theAegean Sea (northeast Mediterranean) lyingbetween 37Æ30’N and 38Æ00’N (Figure 2). Thelength of the coastline is about 744 km, thesurface about 2.866 km2, the maximum depthabout 400 m and the mean depth about 100 m.The Saronic Gulf is bordered by the coasts ofAttica in the north and by the coasts of thePeloponnise in the west and south west. Thegulf communicates with the Aegean in the eastthrough an approximately 50 km wide openconnection. Rocky calcareous shores are thedominant element of the coastline. Thepresent study refers to the coastline of Attica,about 182 km long.
Four sampling stations were chosen in theupper infralittoral zone (0.5 to 1 meter indepth) across two axes with a gradualattenuation of pollution (Figure 2). Stations 1and 2 were located on an axis starting from the
outfall towards the western part of the gulf,whereas stations 3 and 4 were located on anaxis starting from the outfall towards theeastern part of the gulf. The choice of thesampling stations aimed to reflect the ecologicalstatus of the Inner and Outer Gulf as they aredescribed based on hydrological data(COAHMAN et al., 1976). Five samplings werecarried out from August 1998 to September2001 in order to monitor different seasonalaspects of the vegetation. The sampling wasdestructive, using a quadrat size of 20 cm x 20cm (400 cm2), which is considered to be therepresentative minimal sampling area forinfralittoral communities in the Mediterranean(DHOND & COPPEJANS, 1977;BOUDOURESQUE & BELSHER, 1979).Additionally, underwater photos of thevegetation at each sampling station were taken.
Each sample was carefully sorted andidentification at species level was attempted.Phaeophyceae, Chlorophyceae and Rhodo-phyceae were identified and classified accordingto RIBERA et al. (1992), GALLARDO et al.(1993) and ATHANASIADIS (1987),respectively. The abundance of species wasestimated as % cover in the sampling area (4
50 Medit. Mar. Sci., 2/2, 2001, 45-65
Fig. 2: Benthic macroalgal sampling stations in the Saronic Gulf.
Medit. Mar. Sci., 2/2, 2001, 45-65 51
Eco
logi
cal
Fun
ctio
nal
Ext
erna
l mor
phol
ogy
Inte
rnal
ana
tom
yP
rodu
ctiv
ity
Lon
gevi
tyG
row
th S
trat
egie
sG
ener
aSt
ate
Gro
upfo
rm g
roup
(Suc
cess
ion)
(sen
su G
rim
e)
IIA
. She
et-G
roup
Thi
n tu
bula
r and
Unc
ortic
ated
, H
igh
Ann
uals
Rud
eral
U
lva,
Ent
erom
orph
a,sh
eet l
ike
(fol
iose
)on
e-se
vera
l cel
ls th
ick
(Opp
ortu
nist
ic)
Scyt
osip
hon
(ere
ctph
ase)
, Dic
tyot
a II
B. F
ilam
ento
us-
Del
icat
ely
bran
ched
U
nise
riat
e, m
ultis
eria
teH
igh
Ann
uals
Rud
eral
C
yano
phyc
eae,
G
roup
(fila
men
tous
)or
ligh
tly c
ortic
ated
(Opp
ortu
nist
ic)
Cha
etom
orph
a,C
lado
phor
a,Po
lysi
phon
ia, C
eram
ium
,Sp
yrid
iaII
C. C
oars
ely
Coa
rsel
y br
anch
edC
ortic
ated
Spec
ies
Ann
uals
Stre
ss-t
oler
ant-
Aca
ntho
phor
a,B
ranc
hed-
upri
ght
spec
ific
(Mid
-R
uder
al o
r Str
ess-
Cau
lerp
a, C
hord
aria
,G
roup
succ
essi
onal
)to
lera
nt-
Gra
cila
ria, L
aure
ncia
,C
ompe
titor
sL
iago
ra
ID
. Thi
ckT
hick
bla
des
Diff
eren
tiate
d, h
eavi
ly
Low
Pere
nnia
lsC
ompe
titor
sC
ysto
seira
, Cho
ndru
s,L
eath
ery-
Gro
upan
d br
anch
esco
rtic
ated
thic
k w
alle
d(L
ate-
succ
essi
onal
)Fu
cus,
Lam
inar
ia,
Padi
na, S
arga
ssum
,U
dote
aI
E. J
oint
edA
rtic
ulat
ed,
Cal
cifie
d ge
nicu
la,
Low
Pere
nnia
lsC
ompe
titor
sA
mph
iroa,
Cor
ralin
a,C
alca
reou
s-ca
lcar
eous
, upr
ight
flexi
ble
inte
rgen
icul
a(L
ate-
succ
essi
onal
)G
alax
aura
, Hal
imed
a,G
roup
Jani
aI
F. C
rust
ose-
Epi
lithi
c, p
rost
rate
, C
alci
fied
or u
ncal
cifie
d L
owPe
renn
ials
Com
petit
ors
Hyd
rolit
hon,
G
roup
encr
ustin
gpa
ralle
l cel
l row
s(L
ate-
succ
essi
onal
)L
ithot
ham
nion
,Pe
ysso
nnel
ia, P
orol
ithon
IG
. Sea
gras
ses
Hig
hly
diff
eren
tiate
dH
ghly
diff
eren
tiate
d L
owPe
renn
ials
Stre
ss-t
oler
ant
Cym
odoc
ea, P
osid
onia
,fr
om fo
liose
to(e
pide
rmis
, mes
ophy
ll,(P
ione
ers t
o la
te-
Rup
pia
cylin
dric
al (L
eafs
,va
scul
ar sy
stem
)su
cces
sion
al)
rhiz
omes
, roo
ts,
flow
ers,
frui
ts)
Tab
le 2
Fun
ctio
nal c
hara
cter
isti
cs a
nd g
row
th s
trat
egie
s of
mar
ine
bent
hic
mac
roph
ytes
(G
RIM
E, 1
979,
200
1; L
ITT
LE
R &
LIT
TL
ER
, 198
0, 1
984;
LA
RK
UM
& D
EN
HA
RT
OG
, 198
9; L
AR
KU
M e
t al.,
198
9; D
UA
RT
E, 1
995;
HE
MM
ING
A &
DU
AR
TE
, 200
0).

cm2 = 1% of the sampling area) in horizontalprojection (BOUDOURESQUE, 1971;VERLAQUE, 1987). In cases where thecoverage of morphologically similar speciescould not be measured precisely, the specieswere grouped together (as spp.) in order toavoid artificial dissimilarity between stations.Species were classified into FFg according toLITTLER & LITTLER (1980, 1984).
ResultsA. The ecological evaluation modelThe concept - Ecological State Groups
Anthropogenic stress shifts the ecosystemfrom pristine to degraded state, whereopportunistic species dominate. Table 2 showsa synthesis of the functional characteristics andgrowth strategies of marine benthic macro-phytes. Marine benthic macrophytic specieswere classified into two ecological state groups(ESG), the late successional (I) and theopportunistic (II). The first group includesspecies of FFg D, E and F, i.e. species with athick or calcareous thallus, low growth rates andlong life cycles (perennials), whereas the secondof FFg A and B, i.e. sheet-like and filamentousspecies with high growth rates and short lifecycles (annuals). Seagrasses were included inthe first group, whereas Cyanophyceae andspecies with a coarsely branched thallus wereincluded in the second group.
Ecological Evaluation Index
The ecological evaluation index (EEI) is anumber ranging from 2 to 10, indicating theoverall ecological status of transitional andcoastal waters (Table 3). To determine theEEI of an ecosystem the following procedureis used:
1. The area under examination is dividedinto relatively large representative non-overlapping permanent-polygons or -lines (PPor PL) and several relevées of benthicvegetation are obtained from each. All
protocols can preferably include seasonalsampling following either destructive or non-destructive designs.
2. In each relevé the absolute abundance(%) of each ESG is estimated by its coverage.In cases where abundance is estimated asbiomass or number of individuals the data haveto be transformed to a comparable form.
3. The average abundance (%) of ESG I andII are cross compared in a matrix to determinethe ecological status of the PPs or PLs in a rangeof five categories from high to bad (see Figure3). A numerical scoring system was developedto correspond the ecological status categoriesto a numerical value (Table 3).
4. The surface area of each PP or the lengthof each PL is multiplied by their ecologicalstatus value and then divided by the sum ofsurface areas of PPs or lengths of the PLs. Thearea- or length-weighted values are thensummed to obtain EEI and the ecologicalstatus category of the ecosystem (Table 3).
Example
A theoretical water system (WS) is dividedin two PP (WSa, WSb), which cover 30 and70% of its total area, respectively. The meanabsolute coverage (%) of ESG I and II ofsampled relevé in WSa was 45 and 20,respectively. This corresponds to the "good"ecological category (Figure 3) and to score 8(Table 3). The mean absolute coverage (%) ofESG I and II of sampled relevé in WSb was 35and 75, respectively. This corresponds to the
52 Medit. Mar. Sci., 2/2, 2001, 45-65
Numerical value of Ecological Evaluationecological categories Index (EEI)High = 10 [≤10 - >8] = HighGood = 8 [≤8 - >6] = GoodModerate = 6 [≤6 - >4] = ModerateLow = 4 [≤4 - >2] = LowBad = 2 [2] = Bad
Table 3A numerical scoring system for the evaluation
of ecological status of transitional andcoastal waters.

"low" ecological category (Figure 3) and toscore 4 (Table 3). The EEI of WS is following:
EEI = (8 x 0.3) + (4 x 0.7) = 2.4 + 2.8 =5.2, which corresponds to the "moderate"ecological category.
B. Case studies
Macedonian and Thrace lagoons
Eighteen (18) seaweed (7 Chlorophyceae,2 Phaeophyceae and 9 Rhodophyceae) andtwo seagrass species were identified in total(Table 4). The highest number of species wasrecorded in the Vassova lagoon (12 seaweedsand 2 seagrasses) and the lowest in the Fanarilagoon (4 seaweeds and one seagrass). By usingthe functional-form model the species wereclassified into seven FFg and two ESG. Threespecies and the Cyanophyceae mats wereclassified into ESG I, seventeen species intoESG II. The abundance of ESG II was higherthan that of ESG I in all Delta Nestos lagoons.This trend was mainly due to high dominanceof ESG II species in winter. In the Fanarilagoon there was a clear dominance of ESG Ispecies. The ecological status of the lagoonswas evaluated by using the EEI: three lagoonswere classified into the low (Eratino,
Keramoti, Vassova), one lagoon into the good(Agiasma) and one lagoon into the high(Fanari) ecological status category (Table 6).The overall ecological status of Delta Nestoscatchments was evaluated as moderate(EEI=4.9).
The Saronic Gulf
Forty-seven (47) taxa were identified (10Chlorophyceae, 14 Phaeophyceae and 23Rhodophyceae) in total (Table 5). The numberof taxa per sampling varied from 13 to 22 atStation (St.) 1, from 9 to 19 at St. 2, from 12-20 at St. 3 and from 16 to 28 at St. 4. A one-way ANOVA test showed that only at St. 2 thenumber of taxa was statistically different fromthe others. Coverage values varied from 132to 310 at St. 1, from 132 to 331 at St. 2, from117 to 289 at St. 3 and from 143 to 281 at St. 4.A one-way ANOVA test showed that thecoverage values where not statisticallydifferent. By using the functional-form modelthe species were classified into six FFg and twoESG. Seventeen species were classified intoESG I, thirty species into ESG II. Species ofESG II were dominant in St. 2 and 3 whereasspecies of ESG I were dominant at St. 1 and4. The ecological status of the Saronic Gulf
Medit. Mar. Sci., 2/2, 2001, 45-65 53
Fig. 3: A matrix based on the mean abundance (%) of ESGs to determine the ecological status oftransitional and coastal waters.

Agias
ma (S
)St.
1St.
2St.
3St.
4St.
5St.
6St.
7RL
.1RL
.2RL
.3RL
.4RL
.5RL
.6RL
.7
00
00
00
00
00
00
00
010
2560
6070
00
1025
6060
700
00
00
00
00
05
55
00
00
00
00
00
00
25
102
00
00
00
00
00
00
025
00
00
00
090
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
060
400
00
00
00
00
00
01
10
00
00
00
00
00
01
00
00
00
00
00
00
9062
467
1010
27
Speci
esFF
gEc
ologic
al Sta
te Gr
oup I
Cymo
docea
nodo
sa(U
cria)
Asch
erson
(Se)
GCy
stosei
ra ba
rbata
C. Ag
ardh (
Ph)
DRu
ppia
mariti
ma Li
nnaeu
s(Se)
GTo
tal co
verag
e (%)
Ecolo
gical
State
Grou
p II
Acan
thoph
ora na
jadifo
rmis (
Delile
)Pap
enfus
s (Rh
)C
Chond
ria ten
uissim
a(Good
enough
&Wood
ward)
C. Ag
ardh (
Rh)
CCe
ramium
bysso
ideum
Harv
ey (R
h)B
Chaet
omorp
ha ae
rea(D
illwyn
) K¸tz
ing (C
h)B
Chaet
omorp
ha m
edite
rrane
a(K¸
tzing
) K¸tz
ing (C
h)B
Clad
opho
ra da
lmati
caK¸
tzing
(Ch)
BCl
adop
hora
linifo
rmisK
¸tzing
(Ch)
BCy
anop
hyceae
AEc
tocarp
us sp.
(P)
BEn
termo
rpha i
ntestin
alis(L
innae
us) Li
nk (C
h)A
Enter
morph
a linz
a(Lin
naeu
s) J. A
gardh
(Ch)
AGr
acila
ria ar
mata
(C. A
gardh
) J. A
gardh
(Rh)
CGr
acila
ria bu
rsa-pa
storis
(Gme
lin) S
ilva (
Rh)
CHe
rposip
honia
secu
nda f
. tene
lla(C
. Aga
rdh) W
ynne
(Rh)
BLo
phosi
phon
ia sco
pulor
um(H
arvey)
Wom
ersley
(Rh)
BLo
phosi
phon
ia sub
adun
ca(K
¸tzing
) Falk
enbe
rg (R
h)B
Polys
iphon
ia elo
ngata
(Hud
son) S
preng
el (R
h)B
Ulva
rigida
C. Ag
ardh (
Ch)
ATo
tal co
verag
e (%)
Erati
no (S
)Er
atino
(W)
St.1
St.2
St.3
St.4
St.5
St.6
St.8
St. 1
St. 2
St. 3
St. 3
St. 4
St.5
St.5
St. 6
St. 6
St. 7
St. 7
RL.8
RL.9
RL.10
RL.11
RL.12
RL.13
RL.14
RL.15
RL.16
RL.17
RL.18
RL.19
RL.20
RL.21
RL.22
RL.23
RL.24
RL.25
00
00
00
00
00
00
00
00
00
4030
2570
4555
253
30
015
0,52
1015
00
00
00
00
00
00
00
00
00
00
4030
2570
4555
253
30
015
0.52
1015
00
00
00
00
00
00
00
00
00
00
57
00
40
04
30
0.510
2525
1015
210
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
02
10
0.51
8585
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00.5
470
700.5
22
11
3530
2535
350
400
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
05
2520
0.51
12
315
151
0.50
00
00
00
00
00
00
00
00
00
00
00
030
0.50.5
00.5
150.5
00.5
10
030
4235
044
035
3027
.570
.572
28.5
30.5
3027
3312
312
6
Tab
le 4
Com
posi
tion
and
abu
ndan
ce o
f mar
ine
bent
hic
mac
roph
ytes
in s
elec
ted
Mac
edon
ian
and
Thr
ace
lago
ons.
Ch=
Chl
orop
hyce
ae,
Ph=
Pha
eoph
ycea
e, R
h=R
hodo
phyc
eae,
Se=
Seag
rass
. S=
Sum
mer
, W=
Win
ter.
St=
Sam
plin
g st
atio
n, R
L=
rel
evé.
Mor
e de
tail
s fo
r th
efu
ncti
onal
-for
m g
roup
s (F
Fg)
and
the
ecol
ogic
al s
tate
gro
ups
(ESG
) in
Tab
le 2
.
54 Medit. Mar. Sci., 2/2, 2001, 45-65

Speci
esFF
gEc
ologic
al Sta
te Gr
oup I
Cymo
docea
nodo
sa(U
cria)
Asch
erson
(Se)
GCy
stosei
ra ba
rbata
C. Ag
ardh (
Ph)
DRu
ppia
mariti
ma Li
nnaeu
s(Se)
GTo
tal co
verag
e (%)
Ecolo
gical
State
Grou
p II
Acan
thoph
ora na
jadifo
rmis (
Delile
)Pap
enfus
s (Rh
)C
Chond
ria ten
uissim
a(Good
enough
&Wood
ward)
C. Ag
ardh (
Rh)
CCe
ramium
bysso
ideum
Harv
ey (R
h)B
Chaet
omorp
ha ae
rea(D
illwyn
) K¸tz
ing (C
h)B
Chaet
omorp
ha m
edite
rrane
a(K¸
tzing
) K¸tz
ing (C
h)B
Clad
opho
ra da
lmati
caK¸
tzing
(Ch)
BCl
adop
hora
linifo
rmisK
¸tzing
(Ch)
BCy
anop
hyceae
AEc
tocarp
us sp.
(P)
BEn
termo
rpha i
ntestin
alis(L
innae
us) Li
nk (C
h)A
Enter
morph
a linz
a(Lin
naeu
s) J. A
gardh
(Ch)
AGr
acila
ria ar
mata
(C. A
gardh
) J. A
gardh
(Rh)
CGr
acila
ria bu
rsa-pa
storis
(Gme
lin) S
ilva (
Rh)
CHe
rposip
honia
secu
nda f
. tene
lla(C
. Aga
rdh) W
ynne
(Rh)
BLo
phosi
phon
ia sco
pulor
um(H
arvey)
Wom
ersley
(Rh)
BLo
phosi
phon
ia sub
adun
ca(K
¸tzing
) Falk
enbe
rg (R
h)B
Polys
iphon
ia elo
ngata
(Hud
son) S
preng
el (R
h)B
Ulva
rigida
C. Ag
ardh (
Ch)
ATo
tal co
verag
e (%)
Fana
ri (S)
Fana
ri (W)
Kera
moti (
S)St.
1St.
2St.
3St.
4St.
1St.
2St.
3St.
4St.
1St.
2St.
3St.
4St.
5St.
6St.
7RL
.26RL
.27RL
.28RL
.29RL
.30RL
.31RL
.32RL
.33RL
.34RL
.35RL
.36RL
.37RL
.38RL
.39RL
.40
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
100
100
8580
9095
600
00
060
4040
2510
010
085
8090
9560
00
00
6040
4025
00
00
00
00
00
00
00
00
00
00
32
150
00
00
00
00
00
00
00
00
00
00
00
00
00
01
30
500
00
00
00
00
00
00
20
00
00
00
00
05
15
604
00
0.560
4050
00
00
00
00
00
00
00
00
00
00
00
00
020
200
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
050
00
00
00
00
00
00
00
00
00
00
00
00
00
00
30
100
50
00
00
00
00
00
00
00
00
00
00
00
00
01
00
10
00
00
00
00
00
00
00
00
00
00
05
25
04
00
00
00
00
00
00
00
00
00
00
00
00
010
613
7814
100
120
26.5
6040
50
Vasso
va (S
)Va
ssova
(W)
St. 1
St. 2
St.3
St. 4
St. 5
St. 6
St. 6
St. 1
St. 2
St. 3
St. 3
St. 4
St. 4
St. 5
St. 6
RL.41
RL.42
RL.43
RL.44
RL.45
RL.46
RL.47
RL.48
RL.49
RL.50
RL.51
RL.52
RL.53
RL.54
RL.55
00
00
00
01
055
10
00
055
055
6060
00
00
00
00
00
00
00
025
500
00
00
00
455
055
6060
2550
10
551
00
04
00
00
00
00
00
00
00.5
00
00
04
00
22
0.53
1515
102
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00.5
00
00
00
00
00
00
00
00
00
00
00
00
00
10
00
00
00
00
00
00
00
00
0.50
00
00
00
00
00
00
00
00
00
00
00
00
0.50
00
0.50
00
00
34
140
8080
153
0.535
410
1515
200
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
34
01
21
1525
400.5
3015
100
00
00
00
00
00
650
00
00
600
1015
10
01
40.5
102
3425
2915
2068
1011
.559
9896
.541
.531
Tab
le 4
(co
ntin
ued)
Medit. Mar. Sci., 2/2, 2001, 45-65 55

56 Medit. Mar. Sci., 2/2, 2001, 45-65
Stat
ion
1St
atio
n 2
Stat
ion
3 St
atio
n 4
Spec
ies
F
Fg
aug.
mar
. ju
n.ju
n.
sep.
au
g.
mar
. ju
n.
jun.
se
p.
aug.
m
ar.
jun.
ju
n.
sep.
au
g.
mar
.ju
n.
jun.
se
p.
9899
9901
0198
9999
0101
9899
9901
0198
9999
0101
Eco
logi
cal S
tate
Gro
up I
Am
phir
hoa
rigi
da (R
h)
E0
00
00
00
00
00
00
00
50
00
12M
onta
gne
Ana
dyom
ene
stel
lata
(Ch)
E0
00
00
00
00
00
00
00
10
00
0(W
ulfe
n) K
ützi
ngC
oral
lina
spp.
(Rh)
E1
83
0.01
01
4544
7340
114
20.
010
40.
010
0.01
0C
ysto
seir
a cr
inita
(Ph)
(Des
font
aine
s) B
orry
D
00
00
00
00
00
400
460
00
00
00
Cys
tose
ira
crin
itoph
ylla
(Ph)
D
6545
5450
900
00
000
00
00
090
6457
9090
Erc
egov
icC
ysto
seir
a co
mpr
essa
(Ph)
D
1520
1210
00
00
00
00
00
015
342
1019
(Esp
.) G
erlo
ff &
Niz
am.
Cys
tose
ira
med
iterr
anea
(Ph)
D0
00
00
00
00
00
00
00
100
00
0Sa
uvag
eau
D
erm
atol
ithon
spp.
(Rh)
E0.
010.
010.
010.
0120
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
0.01
15F
labe
llia
petio
lata
(Ch)
E0
00
00
00
00
00
00
00
21
00
4(T
ura)
Niz
anud
din
Fos
liella
spp.
(Rh)
E
0.01
0.01
0.01
0.01
100.
010.
010.
010.
010.
011
0.01
0.01
0.01
10.
010.
010.
010.
0125
Hal
imed
a tu
na (C
h)
E
00
00
00
00
00
04
00
204
00
00
(Elli
s & S
olan
der)
Lam
.
Jani
a sp
p. (R
h)
E16
2015
780
10
012
280
018
1680
2616
340
22L
ithot
ham
nion
spp.
(Rh)
F0
00
01
00
00
00
00
00
0.01
0.01
0.01
04
Pad
ina
pavo
nica
(Ph)
D
45
32
01
06
200
300
210
017
216
56
(Lin
naeu
s) L
amou
roux
P
eyss
onne
lia ro
sa-m
arin
a(R
h)F
00
00
00
00
00
00
00
02
00
01
(Bou
dour
esqu
e) D
eniz
.
Sarg
assu
m v
ulga
re(P
h)
D
7515
105
80
30
130
60
036
2551
012
1018
C. A
gard
hT
aoni
a at
omar
ia(P
h)
D0
03
00
00
00
00
00
00
00
04
0(W
oodw
ard)
J. A
gard
hE
colo
gica
l Sta
te G
roup
II
Aca
ntho
phor
a de
lilei
(Rh)
C
00
30
00
00
00
00
069
250
00
00
Lam
ouro
uxB
ryop
sis p
enna
ta (C
h)
B
11
00
00
00
00
00
00
00
00
00
Lam
ouro
ux
Cau
lerp
a ra
cem
osa
(Ch)
C0
00
00
00
00
00
00
00
282
1010
5(F
orsk
al) L
amou
roux
Cer
amiu
m d
iaph
anum
(Rh)
B0
40
00
00
00.
010.
010
00
0.01
0.01
0.01
00
0.01
1(R
oth)
Har
vey
Tab
le 5
Com
posi
tion
and
abu
ndan
ce o
f mar
ine
bent
hic
mac
roph
ytes
in th
e Sa
roni
c G
ulf.
Mor
e de
tail
s in
Tab
les
2 an
d 3.

Medit. Mar. Sci., 2/2, 2001, 45-65 57
Stat
ion
1St
atio
n 2
Stat
ion
3 St
atio
n 4
Spec
ies
F
Fg
aug.
mar
. ju
n.ju
n.
sep.
au
g.
mar
. ju
n.
jun.
se
p.
aug.
m
ar.
jun.
ju
n.
sep.
au
g.
mar
.ju
n.
jun.
se
p.
9899
9901
0198
9999
0101
9899
9901
0198
9999
0101
Cer
amiu
m c
iliat
um(R
h)
B0
00
00
00
00
00
00
00
03
00
0(E
llis)
Duc
luze
auC
hond
ria
dasy
phyl
la(R
h)C
00
00
00
150
00
00
00
00
00
00
(Woo
dwar
d) C
. Aga
rdh
Cla
doph
ora
spp.
(Ch)
B1
20
0.01
00
00
00
220
50
10
10
00
Cod
ium
frag
ile (C
h)B
00
42
00
00
50
00
00
00
00
00
(Sur
inga
r) H
ario
tC
olpo
men
ia si
nuos
a (P
h)C
09
02
00
06
100
00
05
00
00
00
(Rot
h) d
e N
otar
isE
nter
omor
pha
spp.
(Ch)
A
020
00
00
00
00
00
00
00
00
00
Dic
tyop
teris
mem
bran
acea
(Ph)
A
670
25
099
2821
6080
024
030
50
06
100
(Sta
ukho
use)
Bat
ters
Dic
tyot
a sp
p. (P
h)
A1
00
20.
010
642
0.01
01
400
0.01
015
00
0.01
0E
ryth
rotr
ichi
a sp
p. (R
h)A
0.01
0.01
0.01
0.01
0.01
0.01
100.
010.
010.
010.
010.
010.
010.
010.
010.
010.
010.
010.
010.
01G
elid
iella
ram
ello
sa(R
h)C
00
00
00
00
00
00
00
00
30
00
(Küt
z.) F
eldm
. & H
amel
Gig
artin
a ac
icul
aris
(Rh)
C0
214
20
00
332
00
01
00
00
00
0(W
ulfe
n) L
amou
roux
Gig
artin
a te
dii(
Rh)
C
00
00
00
010
00
00
00
00
00
00
(Rot
h) L
amou
roux
Gra
cila
ria
cora
llico
la (R
h)C
00
00
40
00
00
00
00.
010.
010
00
00
Z
anar
dini
Gon
iotr
ichu
m sp
p. (R
h)B
0.01
0.01
0.01
0.01
0.01
0.01
100.
010.
010.
010.
010.
010.
010.
010.
010.
010.
010.
010.
010.
01G
riff
ithsi
a sc
hous
boei
(Rh)
C3
00
00
20
01
00
00
00
10
00
0M
onta
gne
Hal
opte
ris s
pp. (
Ph)
C10
2510
32
00
01
060
200
110
722
355
0H
ypne
a m
usci
form
is(R
h)C
152
00
01
00
00
552
00
01
00
00
(Wul
fen)
Lam
ouro
uxL
aure
ncia
obt
usa
(Rh)
C1
122
252
10
02
06
501
20
10
21
0(H
udso
n) L
amou
roux
Lom
enta
ria
clav
ello
sa(R
h)C
00
00
00
120
00
00
00
00
00
00
(Tur
ner)
Gai
llon
Lop
hosip
honi
a sc
opul
orum
(Rh)
B0
00
0.01
40
00
00
00
00.
010
0.01
0.01
0.01
0.01
0.01
(Har
v.) W
orm
esl.
Pet
alon
ia fa
scia
(Ph)
A
050
210
00
580
00
00
00
00
00
00
(Mül
ler)
O. K
untz
eSp
hace
lari
asp
p. (P
h)
B
00
00
00
00
00
00
01
00.
017
0.01
21
Scyt
osip
hon
lom
enta
ria
(Ph)
A0
60
00
00
00
00
00
00
00
00
0(L
yngb
ye) L
ink
Pol
ysip
honi
a sp
p. (R
h)
B1
00
10
16
02
00
100
30
0.01
20
0.01
0.01
Ulv
a ri
gida
(Ch)
A
346
86
074
8013
3310
764
4274
51
340
00
C. A
gard
hV
alon
ia u
tric
ular
is (C
h)
C0
00
00
00
00
00
00
02
00
00
1C
. Aga
rdh
Tab
le 5
(co
ntin
ued)

was evaluated by using the EEI: one station(St. 2) was classified into the low, two stationsinto the moderate (St. 1 and 3) and one stationinto the good (St. 4) ecological status category(Table 7). The overall ecological status of theInner and Outer Saronic Gulf was evaluated asmoderate (EEI=4.95) and good (EEI=6.38),respectively.
Discussion
The present model was developed toevaluate shifts from pristine to degraded statesin transitional and coastal ecosystems by usinga new ecological index (EEI). The EEIindicates the overall ecological status oftransitional and coastal waters (Table 3) andallows water managers to compare, to rankand to set management priorities at regionaland national levels.
The model used marine benthic macro-phytic communities as bio-indicators because,as sessile primary producers, they responddirectly to anthropogenic stress (Table 1).Since they are important structural and
functional components of the transitional andcoastal waters, inhabiting sediment (roots ofseagrasses) as well as water (seaweeds andleaves of seagrasses), they can potentiallyprovide an integrated measure of ecologicalstatus. Several other models to predictecological status of different aquaticecosystems by using one (HOLMES et al.,1998; RITZ & TRUDGILL, 1999; AMOROSet al., 2000) or several types of bio-indicators(BRICKER et al. 1999; GIBSON et al., 2000;LAFONT et al., 2001) have recently beendeveloped. The main advantages of thesemodels are: (1) Anthropogenic stress isevaluated from the community response(CROWE et al., 2000) and (2) The evaluationdoes not include antagonistic responseparameters like nutrient concentrations(driving force) and phytoplankton or seaweedbiomass (impact) in the same matrix.
The marine benthic macrophytes wereclassified into two ESGs (Table 2) representingcontrasting, alternative ecological states, e.g.pristine and degraded. Degraded marineenvironments are inhabited by annual specieswith high growth rates and reproductive
58 Medit. Mar. Sci., 2/2, 2001, 45-65
Catchments Lagoon Sampling time Area ESG I ESG II Ecological (km2) [Mean total [Mean total Status
(%) coverage] (%) coverage] Category Nestos River 1. Agiasma Summer 3.92 32.1 36 Moderate
(EEI=6)2. Eratino Summer 2.96 41.4 26.6 Good
Winter 4.41 54.3 LowTotal 18.8 43.5 Low
(EEI=4)3. Keramoti Summer 0.974 23.6 58.6 Low
(EEI=4)4. Vassova Summer 0.785 43.6 37.9 Moderate
Winter 7.63 51.9 LowTotal 24.4 45.4 Low
(EEI=4)Summary 8.644 Moderate
(EEI=4.9)Vistonida 5. Fanari Summer 1.93 91.3 0 Highestuarine Autumn 61.3 26.8 High
system Total 76.3 13.4 High(EEI=10)
Table 6Ecological evaluation of the studied Macedonian and Thrace lagoons.

potential (opportunistic, ESG II), whereaspristine environments are dominated byperennial species with low growth rates andreproductive potential (late-successional, ESGI). Many authors (REGIER & COWELL,1972; MURRAY & LITTLER, 1978;SOUSA, 1980; DUARTE, 1995; HARLIN,1995; SCHRAMM & NIENHUIS, 1996;SCHRAMM, 1999) have documented thispattern. LITTLER & LITTLER (1980) haveextensively discussed the attributes that seemto improve the fitness of opportunistic andlate-successional seaweeds and theirhypothetical costs and benefits. Seaweeds witha sheet-like and filamentous thallus (FFg Aand B) are generally more productive and growin more temporally unstable habitats thanthicker and calcareous seaweeds (FFg D, Eand F), which are conspicuous in moreconstant environments (LITTLER &LITTLER, 1980; 1981; 1984, see also Table
2). Some species, however, throughmorphologically or ecologically dissimilaralternate phases (e.g. Scytosiphon, Colpomenia,Petalonia), have attributes of both extremes.
Prediction of function, like productivity,from morphology is not always simple, becausein several cases there is not a clear relationship.For example, Caulerpa van-bosseae had muchlower photosynthetic rates than othermembers of FFg-B and Sargassumherporhizum had much higher photosyntheticrates than other members of FFg-D(LITTLER & LITTLER, 1984). Similarly,species belonging to FFg-C had photosyntheticrates similar to members of FFg-B or FFg-Dor were intermediate. In the present modelthe species belonging FFg C were classifiedinto ESG II because several of its members,such as Gracilaria spp., Acanthophora spp.,Gigartina spp., are dominant species indegraded marine ecosystems (CASABIANCA
Medit. Mar. Sci., 2/2, 2001, 45-65 59
Sampled Area Sampling Coastline Sampling time ESG I ESG II Ecological Statusstation length (% Coverage) (% Coverage) Category
(Km)Inner August 1998 176 134 Moderate
Saronic Peristeria March 1999 113 139 ModerateGulf (St. 1) 10 June 1999 100 45 Good
June 2001 74 58 GoodSeptember 2001 209 12 High
mean 134.4 77.6 Moderate (EEI=6)August 1998 3 178 Bad
Kaki Vigla March 1999 48 283 Low(St. 2) 51 June 1999 50 85 Low
June 2001 118 116 ModerateSeptember 2001 42 90 Low
mean 52.2 150.4 Low (EEI=4)August 1998 158 131 Moderate
Agios Kosmas March 1999 18 230 Low(St.3) 36 June 1999 68 49 Good
June 2001 62 185 ModerateSeptember 2001 126 48 Good
mean 86.4 128.6 Moderate (EEI=6)Summary 97 Moderate
(EEI=4.95)Outer August 1998 227 54 Good
Saronic Agios March 1999 117 74 ModerateGulf Nikolaos 85 June 1999 90 53 Good
(St. 4) June 2001 159 28 HighSeptember 2001 216 8 High
mean 161.8 43.4 Good (EEI=8)
Table 7Ecological evaluation of the studied stations in the Saronic Gulf.

et al., 1997; LAZARIDOU et al., 1997;ORFANIDIS et al., 2001). However, we agreewith the conclusion that "functional groupranking realistically should be regarded asrecognizable units along a continuum, eachcontaining considerable variation of form andconcomitant functional responses" (LITTLER& LITTLER, 1984).
Comparing growth rates (DUARTE,1995), resistance to herbivory (CEBRIAN &DUARTE, 1994), longevity and canopy heightthere were many similarities betweenseagrasses and seaweeds belonging to FFg-D(Table 2). In addition, both groups are late-successional forming climax communities. Forexample, Posidonia oceanica is a late-successional species in undisturbed areas ofthe Mediterranean Sea and Cymodoceanodosa is a late successional species inrelatively more stressful habitats. Zosteramarina is considered a colonizer as well as aclimax species on the North European coasts(McROY & LLOYD, 1981).
Another often used model in plant ecologyis GRIME’S (1977, 2001) C-S-R triangle, inwhich species can be categorized as: (1)Ruderal (opportunists), (2) Stress-tolerant,(3) Competitors (late-successional). While thethree strategies are the extremes, many plantswill show tradeoffs of the threetrait syndromesto some extent. Obviously, the sheet-like andfilamentous species of LITTLER & LITTLER(1980) can be classified as ruderals in Grime’sscheme and the thick and calcareous speciesas competitors. However, morphology seemsnot to be a criterion in identifying the seaweedsthat follow a stress-tolerant strategy. Althoughin terrestrial environments, a stress-tolerantplant tends to have late-successionalcharacteristics, the more stress-resistantmarine algae, such as blue-greens, Ulva andEnteromorpha, are opportunistic species(LOBBAN & HARRISON, 1994; COELHOet al., 2000). Although Grime’s model has beenextended and modified by STENECK &DETHIER (1994) in order to apply tofunctional groups of marine algae, it seems
that it has limited usage as a predictive modelin applied marine ecology. This conclusion isfurther strengthened by the categorization ofseagrasses into the stress-tolerant group(LARKUM & DEN HARTOG, 1989)because of low diversity and the fact that theycolonize habitats (shallow bays and estuaries)with low competition. This is, however, incontradiction with the high sensitivity ofseagrasses to anthropogenic stress (Table 1).
On the axes of the matrix, the % ofabsolute abundance of the ecological groups(Figure 3) is represented. This was donebecause ecosystem function or processes, e.g.nutrient cycling, productivity, can be closelyrelated to species abundance. To give anexample, the particle and nutrient sink in twoZostera meadows in Sylt, North Sea, waspositively correlated with their density(ASMUS & ASMUS, 2000). A positiverelation between shoot density of seagrassesand fish and shrimp density was alsodocumented (FONSECA et al., 1996a, b).Because the stratification of marine benthicmacrophytic communities includes severalgrowth forms (canopies, understorey etc.) totalcoverage can be higher than 100% (see below).Consistent low values of total coverage,especially in seagrass meadows, are indicativeof habitat destruction and fragmentation.
The model can classify the transitional andcoastal waters into five ecological categories,from high to bad (EEC WFD, 2000/60), basedon a cross comparison in a matrix of the twoESGs (Figure 3). In contrast to the availabletypologies, e.g. EEC Directive 92/43, thistypology is based on ecological processes whichcan also predict restoration potentialities.According to the model, a restoration goal ofa degraded aquatic environment could includean improvement of hydrological and ecologicalconditions to allow growth of seagrass in softsubstratum (e.g. Posidonia) and seaweedcommunities of FFg-D (e.g. Laminaria, Fucus,Cystoseira) on hard substratum. Restorationof kelp communities and, in particular seagrassmeadows in degraded aquatic environments
60 Medit. Mar. Sci., 2/2, 2001, 45-65

is a well-known practice on the NorthAmerican coast (FONSECA et al., 1998;ROBLEDO et al., 2000). These communitiesnot only form the basis of natural, pristinemarine environments (PANAYOTIDIS et al.,2001) but also support many ecosystemservices, e.g. sustain biodiversity (DENHARTOG, 1970; PHILLIPS & MENEZ,1988; NIENHUIS, 1992), maintain fish habitat(HARMELIN-VIVIEN et al., 1995;POLLARD, 1984; BELL et al., 1992;FRANCOUR, 1997; EDGAR, 1999b;BLADER et al., 2000), offer detritus to thetrophic chain (EDGAR, 1999a), maintainwater quality (BULTHUIS et al., 1984;DAUBY et al., 1995; GACIA et al., 1999;ASMUS & ASMUS, 2000), stabilize sedimentand control erosion (FONSECA, 1996;ASMUS & ASMUS, 2000).
The test of the model in estuarine (Table6) and coastal waters (Table 7) was successfulbecause the results are in accordance withexisting ecological conditions as documentedbefore. During previous decades, the NestosDelta Lagoons (moderate ecological status)were severely influenced by eutrophicationbecause of point (phosphorus industry) andnon-point (agriculture) sources of effluents(THEOCHARIS et al., 2000; SYLAIOS &THEOCHARIS, 2002). Some ten years agothe problem was so severe that local watermanagers decreased the inflow of fresh-watersources into the lagoons. In contrast, theFanari lagoon (high ecological status) seemsto be un-affected by eutrophication becauseof limited fresh-water sources.
The inner parts of the Saronic Gulf(Central Greece) are considered among themost polluted Greek aquatic ecosystemsbecause of proximity to the densely populatedAthens basin. Earlier studies showed a gradualattenuation of the pollution caused by thecentral outfall of urban wastes towards thesouthern and eastern parts of the gulf(DIAPOULIS & HARITONIDIS, 1987;SIMBOURA et al., 1995). This pattern wasalso documented in this study.
The ecological status of the tested areasshowed temporal and spatial changes (Table6 and 7). This is in agreement with ecologicaltheory, which regards brackish waters andpolluted areas as unpredictable environments(SANDERS, 1968; COGNETTI &MALTAGLIATI, 2000) with spatial andtemporal changes in the intensity of disturbance(ORFANIDIS et al., 2001). In the model, thefinal ecological status designated to an area isbased on many seasonally sampled relevés,which reflects an average of environmentalconditions existing during the investigation.
Conclusion
The ecological evaluation modeldeveloped in this study can be a valuable toolfor transitional and coastal water managers inEurope and worldwide. It gives them thepossibility to compare, to rank and to setpriorities at regional and national levels quicklyand without a demand for specializedknowledge of seaweed or seagrass taxonomy.Usage of the model could provide acomprehensive and objective picture of currentecological status, whereas a monitoringprogram could allow analysis of environmentaldegradation or improvement.
Marine benthic macrophytes wereclassified from functional form characteristicsinto two ecological groups that representcontrasting ecological states. Because ofunclear limits between the proposed ESGs, acertain amount of research requirement isneeded. The proposed EEI classified waterecosystems into five ecological categories fromhigh to bad, where their exact limits still haveto be tested.
Acknowledgements
Authors S.O and N.S. are very grateful toE. Tsiagga (McS) for her significant supportin evaluating composition and abundance ofmarine benthic macrophytic communities of
Medit. Mar. Sci., 2/2, 2001, 45-65 61

Eastern Macedonian and Thrace lagoons.Thanks are also due to members of theAgricultural Fisheries Cooperation of Kavalaand especially Dr V. Ragias for their help insample collection. Many thanks also to P.Manitsa for her technical assistance in theimplementation of GIS technology. Finally,we all thank Dr A. M. Breeman and Dr I.Bartsch, as well as the two anonymous referees,for constructive comments on the manuscript.
References
AMOROS, C., BORNETTE, G. & HENRY, C. P.,2000. A vegetation-based method for the ecologicaldiagnosis of riverine wetlands. EnvironmentalManagement 25: 211-227.
ASMUS, H. & ASMUS, R., 2000. Material exchangeand food web of seagrass beds in the Sylt-Ro/ mo/Bight: how significant are community changes atthe ecosystem level? Helgoland Marine Research,54: 137-150.
ATHANASIADIS A., 1987. A survey of the seaweedof the Aegean Sea with taxonomic studies on thespecies of the tribe Antithamnieae (Rhodophyta).Ph.D. Thesis, University of Göthenburg, 174 p.
BELL, J. D., FERRELL, D. J., McNEILL, S. E. &WORTHINGTON, D. G., 1992. Variation inassemblages of fish associated with deep andshallow margins of the seagrass Posidonia australis.Marine Biology, 114: 667-676.
BOROWITZKA M. A., 1972. Intertidal algal speciesdiversity and the effects of pollution. AustralianJournal of Marine and Freshwater Research, 25: 73-84.
BLADER, S. J. M., CYRUS, D. P., ALBARET, J.–J., CHONG VING CHING, DAY, J. W.,ELLIOTT, M., FONSECA, M. S., HOSS D. E.,ORENSANZ, J., POTTER, I. C. and SILVERT,W., 2000. Effects of fishing on structure andfunctioning of estuarine and nearshore ecosystems.ICES Journal of Marine Science, 57: 590-602.
BOUDOURESQUE C.F. & BELSHER T., 1979.Le peuplement algal du port de Port-Vendres:Recherches sur l' aire minimale qualitative. Cahiersde Biologie Marine, Fr.20: 259-269.
BOUDOURESQUE, C.F., 1971. Méthodes d' étudequalitative et quantitative du benthos (en particulierdu phytobenthos). Téthys, (1): 79-104.
BREEMAN, A. M., 1990. Expected effects ofchanging seawater temperatures on the geographicdistribution of seaweed species, p. 69-76. In:Expected effects of climatic change on marine coastalecosystems, edited by J. J. Beukema, W. J. Wolff,J. J. M. W. Brouns, Kluwer Academic Publishers,The Netherlands.
BRICKER, S. B., CLEMENT, C. G., PIRHALLA,D. E., ORLANDO, S. P. & FARROW, D. R. G.,1999. National estuarine eutrophicationassessment: Effects of nutrient enrichment in thenation’s estuaries. NOAA, National Ocean Service,Special Projects Office and the National Centersfor Coastal Ocean Science. Silver Spring, MD: 71pp.
BULTHUIS, D. A., BRAND, G. W. & MOBLEY,M. C., 1984. Suspended sediments and nutrientsin water ebbing form seagrass-covered anddenuded tidal mudflats in a southern Australianembayment. Aquatic Botany, 20: 257-256.
BURROWS, E. M., 1991. Seaweeds of the BritishIsles, Volume 2 Chlorophyta. Natural HistoryMuseum, London.
CASABIANCA, M. L. De, MARINHO-SORIANO,E. & LAUGIER, T., 1997. Growth of Gracilariabursa-pastoris in a Mediterranean lagoon: Thau,France. Botanica Marina, 40: 29-37.
CEBRIAN, J. & DUARTE, C. M, 1994. Thedependence of herb ivory on growth rate in naturalplant communities. Functional Ecology, 8: 518-525
CHAPMAN, V.J. & CHAPMAN, D. J., 1976. Lifeforms in the algae. Botanica Marina, 19: 65-74.
CHRYSSOVERGIS, F. & PANAYOTIDIS, P.,1995. Communities of macrophytobenthos alongan eutrophication gradient (Maliakos Gulf, AegeanSea, Greece). Oceanologica Acta, 18 (6): 649-658.
CLOERN, J. E, 2001. Our evolving conceptual modelof the coastal eutrophication problem. MarineEcology Progress Series, 210: 223-253.
COACHMAN, L., HOPKINS, T. & DUGDALER., 1976. Water masses of the Saronikos gulf inwinter. Acta Adriatica, 18: 133-162.
COELHO, S. M., RIJSTENBIL, J. W. & BROWN,M. T., 2000. Impacts of anthropogenic stress onthe early development stages of seaweeds. Journalof Aquatic Ecosystem Stress and Recovery, 7: 317-333.
COGNETTI, G. & MALTAGLIATI, F., 2000.Biodiversity and adaptive mechanisms in brackishwater fauna. Marine Pollution Bulletin, 40: 7-14.
COPPEJANS, E., 1983. Iconographie d’ AlguesMéditerranéennes. J. Cramer.
62 Medit. Mar. Sci., 2/2, 2001, 45-65

CROOKS, S. & TURNER, R. K., 1999. Intergradedcoastal management: sustaining estuarine naturalresources, p. 241-289. In: Estuaries, edited byNedwell D. B., Raffaelli D. G., Advances inecological research vol. 29, Academic Press.
CROWE, T. P., THOMPSON, R. C., BRAY, S. &HAWKING, S. J., 2000. Impacts of anthropogenicstress on rocky intertidal communities. Journal ofAquatic Ecosystem Stress and Recovery. 7: 273-297
DAUBY, P., BALE, A. J., BLOOMER, N.,CANON, C., LING, R. D., NORRO, A.,ROBERTSON, J. E., SIMON, A., THEATE, J.M., WATSON, A. J. & FRANKIGNOULLE, M.,1995. Particle fluxes over the Mediterraneanseagrass beds: a one-year case study. Marine EcologyProgress Series, 126: 233-246.
DIAPOULIS A. & HARITONIDIS S., 1987. Aqualitative and quantitative study of the marinealgae in the Saronikos Gulf (Greece). P.S.Z.N.I.,Marine Ecology. 8 (2): 175-189.
DHOND, F. & COPPEJANS, E., 1977. Résultatsd'une étude d' aire minima des peuplements algauxphotophiles sur substrat rocheux à Port-Cros et àBanyuls (France). Rapport Comm. int. Mer Médit.,24 (4): 141-142.
DUARTE, C. M, 1995. Submerged aquaticvegetation in relation to different nutrient regimes.Ophelia, 41: 87-112.
EDGAR, G. J. 1999a. Experimental analysis of thestructural versus trophic importance of seagrassbeds. I. Effects on macrofaunal and meiofaunalinvertebrates. Vie et Mileu, 49 (4): 239-248.
EDGAR, G. J. 1999b. Experimental analysis of thestructural versus trophic importance of seagrassbeds. II. Effects on fishes, decapods andcephalopods. Vie et Mileu, 49 (4): 249-260.
EEC, 1992. Council Directive for the Habitats,92/43/EEC, Official Journal of the E.C. 22/7/1992.
EEC, 1994. Proposal for a Council Directive on theecological quality of water. 94/C 222/06, OfficialJournal of the E.C. 10/8/94.
EEC, 2000. Council Directive for a legislative frameand actions for the water policy, 2000/60/EC,Official Journal of the E.C. 22/12/2000.
FELDMAN, J., 1951. Ecology of marine algae, p.313-334. In: Manual of Phycology-an Introductionto the algae and their biology, edited by Smith G.M.,Waltham, Massachusetts.
FLETCHER, R. L., 1995. Seaweeds of the BritishIsles, volume 3, Part 1 Fucophyceae(Phaeophyceae). Natural History Museum,London.
FONSECA, M. S., 1996. The role of seagrasses innearshore sedimentary processes: A review. pp.261-286. In: Estuarine Shores: Hydrological,Geomorphological and Ecological Interactions,edited by C. Roman, K. Nordstrom. Blackwell,Boston, pp. 261-286.
FONSECA, M. S. & CALAHAN, J. A., 1992.Preliminary evaluation of wave attenuation by fourspecies of seagrass. Estuarine Coastal Shelf Science,35: 565-576.
FONSECA, M. S., KENWORTHY, W. J. &COURTNEY F. X., 1996a. Development ofplanted seagrass beds in Tampa Bay, Florida,U.S.A.: I. Plant components. Marine EcologyProgress Series, 132: 127-139.
FONSECA, M. S., MEYER, D. L. & HALL, M. O.,1996b. Development of planted seagrass beds inTampa Bay, Florida, U.S.A.: II. Faunalcomponents. Marine Ecology Progress Series, 132:141-156.
FONSECA, M. S., KENWORTHY, W. J. &THAYER, G. W., 1998. Guidelines for theconservation and restoration of seagrasses in theUnited States and Adjacent waters. NOAA CoastalOcean Program Decision Analysis Series No. 12.NOAA Coastal Ocean Office, Silver Spring, MD.222 p.
FRANCOUR, P., 1997. Fish assemblages ofPosidonia oceanicabeds at Port-Cros (France, NWMediterranean): assessment of composition andlong-term fluctuations by visual census. P.S.Z.N.I.,Marine Ecology, 18: 157-173.
GACIA, E., GRANATA, T. C. & DUARTE, C. M.,1999. An approach to measurement of particle fluxand sediment retention within seagrass (Posidoniaoceanica) meadows. Aquatic Botany, 65: 255-268.
GALLARDO, T., GOMEZ – GARRETA, A.,RIBERA, M.A., CORMACI, M., FURNARI, G.,GIACCONE, G. & BOUDOURESQUE, C.F.,1993. Checklist of Mediterranean Seaweeds. II.Chlorophyceae (Wille s.l.). Botanica Marina, 36:399-421.
GIBSON, G. R., BOWMAN, M. L., GERRITSEN,J. and SNYDER, B. D., 2000. Estuarine and coastalmarine waters: Bioassessment and biocriteriatechnical guidance. EPA 822-B-00-024. U.S.Environmental Protection Agency, Office ofWater, Washington, DC.
GRIME, J. P., 1977. Plant strategies and vegetationprocesses. John Wiley & Sons, New York.
Medit. Mar. Sci., 2/2, 2001, 45-65 63

GRIME, J. P., 2001. Plant strategies, vegetationprocesses and ecosystem properties. John Wiley& Sons, New York.
HARITONIDIS, S., 1978. Contribution to theresearch of marine plant macrophyceae (Chloro,Phaeo-, and Rodophyceae) of the ThermaikosGulf. Phd Thesis, Aristotle University ofThessaloniki (in Greek).
HARLIN, M. M., 1995. Changes in major plantgroups following nutrient enrichment. p. 173-187.In: Eutrophic shallow estuaries and lagoons, editedby A. J. Mc Comb. Institute for EnvironmentalScience, Murdoch University, Murdoch, Australia,CRC Press.
HARMELIN-VIVIEN, M. L, HARMELIN, J. G,& LEBOULLEUX, V., 1995. Microhabitatrequirements for settlement of juvenile sparid fisheson Mediterranean rocky shores. Hydrobiologia,300/301: 309-320.
HARTOG, C. den, 1970. The sea-grasses of theworld. Verhandlingen der KoninklijkeNederlandse Akademie van Wetenschappen Afd.Natuurkunde Tweede Reeks Deel 59 No. 1. North-Holand, Amsterdam.
HEMMINGA, M. A. & DUARTE, C. M., 2000.Seagrass ecology. Cambridge University Press.
HOLMES, N. T. H., BOON, P. J. & ROWELL, T.A., 1998. A revised classification system for BritishRivers based on their aquatic plant communities.Aquatic Conservation, 8 (4): 555-578.
KAMERMANS, P., HEMMINGA, M. A. & DEJONG, D. J. (1999). Significance of salinity andsilicon levels for growth of a formerly estuarineeelgrass (Zostera marina) population (LakeGrevelingen). Marine Biology, 133: 527-539.
LAFONT, M., CAMUS, J-C., FOURNIER, A. &SOURP, E., 2001. A practical concept for theecological assessment of aquatic ecosystems:application on the River Dore in France. AquaticEcology, 35 (2): 195-205.
LARKUM, A. W. D. & HARTOG, C. den, 1989.Evolution and biogeography of seagrasses, p. 112-156. In: Biology of seagrasses. A treatise on the biologyof seagrasses with special references to the Australianregion, edited by A. W. D., Larkum, A. J. McComb,and S. A. Shepherd, Elsevier, Amsterdam.
LARKUM, A. W. D., McCOMB, A. J. &SHEPHERD, S. A., (Eds), 1989. Biology ofseagrasses. A treatise on the biology of seagrasseswith special references to the Australian region.Elsevier, Amsterdam.
LAZARIDOU, E. 1994. Systematics, bionomy andecological study of the marine phytobenthos ofMilos Island (Cyclades). PhD Thesis, AristotleUniversity of Thessaloniki (in Greek).
LAZARIDOU, E., ORFANIDIS, S.,HARITONIDIS, S. & SEFERLIS, M., 1997.Impact of eutrophication on species compositionand diversity of macrophytes in the Gulf ofThessaloniki, Macedonia, Greece: First evaluationof the results of one year study. FreseniusEnvironmental Bulletin, 6: 54-59.
LITTLER, M. M., 1980. Morphological form andphotosynthesis performance of marine macroalgae:tests of a functional/form hypothesis. BotanicaMarina, 22: 161-165.
LITTLER, M. M. & ARNOLD, K. E., 1982. Primaryproductivity of marine macroalgal functional-formsgroups from southwestern North America. Journalof Phycology, 18: 307-311.
LITTLER, M. M. & LITTLER, D. S., 1980. Theevolution of thallus form and survival strategies inbenthic marine macroalgae: field and laboratorytests of a functional form model. AmericanNaturalist, 116: 25-44.
LITTLER, M. M. & LITTLER, D. S., 1981. Intertidalmacrophyte communities from Baja Californiaand upper gulf of California relative constant vs.environmentally fluctuating systems. MarineEcology Progress Series, 4: 145-158.
LITTLER, M. M. & LITTLER, D. S. 1984.Relationships between macroalgal functional formgroups and substrata stability in a subtropical rocky-intertidal system. Journal of Experimental MarineBiology and Ecology, 74: 13-34.
LOBBAN, C. & HARRISON, P. J., 1994. Seaweedecology and physiology. Cambridge UniversityPress. 366 p.
LOTZE, H. K. & SCHRAMM, W., 2000.Ecophysiological traits explain species dominancepatterns in macroalgal blooms. Journal of Phycology36: 287-295.
LOTZE, H. K., SCHRAMM, W. & SCHORIES, D,1999. Control of macroalgal blooms at earlydevelopmental stages: Pilayella littoralis versusEnteromorpha spp. Oecologia, 119: 46-54.
MAGGS, C. A. & HOMMERSAND, M. H., 1993.Seaweeds of the British Isles, Volume 1Rhodophyta Part 3A Ceramiales. Natural HistoryMuseum, London.
MANN, K. H., 1973. Seaweeds: Their productivityand strategy for growth. Science, 182: 975-981.
64 Medit. Mar. Sci., 2/2, 2001, 45-65

McROY, C. P & LLOYD, D. S, 1981. Comparativefunction and stability of macrophyte-basedecosystems, p. 473-489. In: Analysis of Marineecosystems, edited by A. R Longhurst, AcademicPress.
MURRAY, S. N & LITTLER, M. M, 1978. Patternsof algal succession in a perturbated marineintertidal community. Journal of Phycology, 14: 506-512.
NIENHUIS, P. H., 1992. Ecology of coastal lagoonsin the Netherlands (Veerse Meer andGrevelingen). Vie et Milieu, 42: 59-72.
ORFANIDIS, S., STAMATIS, N., TSIAGGA, E. &SCHRAMM, W., 2001. Variability of thecharacteristics of seaweed communities in aeutrophic lagoon (Vassova, N. Greece). Proc. 7thInt. Phycological. Cong. Phycologia 40 (4),supplement, 117 p.
PAKKER, H. & BREEMAN, A. M., 1994.Temperature ecotypes in seaweeds-adaptivesignificance and biogeographic implications.Botanica Marina, 37: 171-180.
PANAYOTIDIS, P., SIAKAVARA, A.,ORFANIDIS, S. & HARITONIDIS, S., 2001.Identification and description of habitat types atsites of interest for conservation. Study 5: Marinehabitats. Final Technical Report, Athens October2001.
PHILLIPS, R. C. & ME_EZ, E. G., 1988. Seagrasses.Smithsonian Contribution to the Marine Sciences.Smithsonian Institution Press, Washington, D.C.no 34, pp. 1-104.
POLLARD, D. A., 1984. A review of ecologicalstudies on seagrass-fish communities, withparticular reference to recent studies in Australia.Aquatic Botany, 18: 3-42.
REGIER, H. A & CORWELL, E. B., 1972.Applications of ecosystem theory, succession,diversity, stability, stress and conservation.Biological Conservation, 4 (2): 83-88.
RIBERA, M.A., GOMEZ-GARRETA, A.,GALLARDO,T., CORMACI, M., FURNARI,G. & GIACCONE, G. 1992. Check-list ofMediterranean Seaweeds. I. Fucophyceae(Warming, 1884). Botanica Marina, 35: 109-130.
RITZ, K. & TRUDGILL, D. L. 1999. Utility ofnematode community analysis as an integratedmeasure of the functional state of soils: perspectivesand challenges. Plant and soil, 212 (1): 1-11.
ROBLEDO, D., GARCIA, O., FOSTER, M. &HERNANDEZ-CARMONA, G., 2000.Restoration techniques for Macrocystis pyrifera
(Phaeophyceae) populations at the southern limitof their distribution in Mexico. Botanica Marina,43 (3): 273-284.
RUSSEL, G., 1977. Vegetation on rocky shores atsome north Irish sea sites. Journal of Ecology, 65:485-495.
SANCHEZ-JEREZ, P. & RAMOS, A., 1996.Detection of environmental impacts by bottomtrawling on Posidonia oceanica (L.) Delilemeadows: sensitivity of fish and macroinvertebratescommunities. Journal of Aquatic Ecosystem Health,5: 239-253.
SANDERS, H. L., 1968. Marine benthic diversity: acomparative study. American Naturalist, 102: 243-282.
SCHRAMM, W., 1999. Factors influencing seaweedresponses to eutrophication: some results fromEU-project EUMAC. Journal of Applied Phycology,11(1): 69-78.
SCHRAMM, W. & NIENHUIS, P. H., (Eds), 1996.Marine benthic vegetation: recent changes and theeffects of eutrophication. Ecological studies 123.Springer-Verlag, 470 p.
SIMBOURA, N., ZENETOS, A., PANAYOTIDIS,P. & MAKRA, A., 1995. Changes in benthiccommunity structure along an environmentalpollution gradient. Marine Pollution Bulletin, 30(7): 470-474.
SOUSA, W. P, 1980. The responses of a communityto disturbance: the importance of successional ageand species' life histories. Oecologia, 45: 72-81.
STENECK, R. & DETHIER, M.N., 1994. Afunctional group approach to the structure of algal-dominated communities. Oikos, 69 (3), 476-498.
STENECK, R. S & WATLING, L, 1982. Feedingcapabilities and limitation of herbivorous molluscs:A functional group approach. Marine Biology, 68:299-319.
SYLAIOS, G. & THEOCHARIS, V. A., 2002.Hydrology and nutrient enrichment at two coastallagoon systems in Northern Greece. WaterResources Management (in press).
THEOCHARIS, V., SYLAIOS, G. & STAMATIS,N., 2000. Water quality variability at two coastallagoons in Northern Greece. FreseniusEnvironmental Bulletin, 9: 30-35.
VERLAQUE, M., 1987. Contribution à l’étude duphytobenthos d’un écosystème photophilethermophile marin en Méditerranée Occidentale.Thèse Doctorat d’Etat-Sciences, Univ. Aix-Marseille II, Vol. 1 (texte): 389 p. & Vol. 2(illustrations & annexe): 260 p.
Medit. Mar. Sci., 2/2, 2001, 45-65 65
