ecological and evolutionary determinants of host species choice in mosquito vectors
TRANSCRIPT

Ecological and evolutionarydeterminants of host specieschoice in mosquito vectorsIssa N. Lyimo1,2 and Heather M. Ferguson1,2
1 Public Health Entomology Unit, Ifakara Health Institute, Off Mlabani Passage, PO Box 53, Ifakara, Tanzania2 Faculty of Biomedical and Life Sciences, 120 University Place, University of Glasgow, Glasgow G12 8TA, UK
Review
Glossary
Anthropophilic: mosquitoes exhibiting a preference for blood feeding on
humans.
Autogeny: the ability to produce one or more egg batches without feeding on
blood. Some mosquitoes are autogeneous, with energetic resources for egg
production being taken from larval nutritional reserves instead of from
bloodfeeding at the adult stage. By contrast, the egg production of
anautogenous mosquitoes depends upon consumption of blood at the adult
stage.
Human blood index (HBI): the proportion of bloodfed mosquitoes in a given
population sample that test positive for human blood.
Fitness: this term is notoriously difficult to define and is given numerous
different definitions in the literature. In this article, it is defined as the total
number of offspring produced by a certain type (of phenotype or genotype) of
individual relative to another type.
Insects exhibit diverse resource-exploitation strategies,including predation, herbivory and parasitism. The eco-logical and evolutionary factors that influence theresource selection of some insects (e.g. herbivores) havebeen extensively investigated because of their agricul-tural importance. By contrast, there has been little inves-tigation of the selective forces that mediate host choice inhaematophagous insects, despite their importance asvectors of disease. Here, we review potential determi-nants of host species choice in mosquitoes, the mostimportant insect vectors of human disease, and discusswhether thesecouldbemanipulatedtoyield new disease-control strategies based on vector behavioural change.
Epidemiological relevance of mosquito host specieschoiceInsects are the most abundant and diverse group ofanimals [1]. Due to the massive economic impact of herbi-vorous insects on agriculture and forestry, the determi-nants of their feeding preferences have been investigatedextensively [2]. By contrast, little is known about thedeterminants of host choice in insects that feed onvertebrate blood. These organisms transmit numeroushuman and animal diseases; mosquitoes have the greatestimpact on public health because of their role in malaria,arboviral and filariasis transmission [1]. Although not allmosquitoes require blood for reproduction (some arecapable of autogeny), the host species choice of those thatdo is a crucial determinant of the transmission intensity ofpathogens they transmit [3] (Box 1). The ecological andevolutionary factors that may determine host choice inhaematophagous mosquitoes are reviewed here. The cen-tral aim is to discuss the ultimate selective forces respon-sible for driving the evolution of host species choice andhow they may be manipulated to reduce vector-bornedisease. We do not focus on the proximate mechanismsused to detect and distinguish between host species orindividuals (for a review, see Ref. [4]) but rather thepotential ultimate evolutionary factors that have shapedthese choices.
Evolution of host choiceA substantial body of theory and empirical research hasaccrued to predict how organisms should select dietary
Corresponding author: Lyimo, I.N. ([email protected])
1471-4922/$ – see front matter � 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.pt.2009.01
resources to maximize their fitness (see Glossary) [5]. Thewidespread existence of dietary specialization presents aparadox: why choose a limited variety of resources whenmany are available? Dietary specialization is predicted toevolve when there are trade-offs between the fitnessobtained from feeding on different resources, such thatnet energy gain is higher when only a limited subset isconsumed [6]. By contrast, generalism should evolve whenthere are only moderate differences in energetic gainsbetween resources [6].
By extension tomosquitoes, generalism should evolve inenvironments where the chances of host encounter are lowand the advantage of waiting for an optimal host is tradedoff against the risk of death before feeding. Conversely,specialism should arise as the frequency of encounters withfavorable host species increases. The host breadth of hae-matophagous insects is probably the product of bothoptimal foraging on currently available hosts [7] andhistorical patterns of host availability, as reflected byphylogenetic congruence with their vertebrate hosts [8].
For mosquitoes to evolve generalist or specialist host-feeding strategies as predicted, there must be geneticvariation for host choice. Although rarely investigated,this phenomenon has been documented in the Africanmalaria vector Anopheles gambiae, in which divergentfeeding preferences for humans or cows were generatedwithin 5–6 generations of selection [9], and in cross-matingexperiments with zoophilic Aedes simpsoni and anthropo-philic Aedes aegypti, which generated offspring of inter-mediate preference [10]. Thus, selection has the potential
Zoophilic: mosquitoes exhibiting a preference for blood feeding on non-human
animals.
.005 Available online 9 March 2009 189
Box 1. Epidemiological significance of mosquito host
choice to malaria transmission
Malaria transmission is a function of the three-way interaction
between humans, Plasmodium parasites and the anopheline
mosquito vectors that transmit them between hosts. Transmission
depends on several aspects of the outcome of their interactions and
life history including human–vector contact rate, vector and parasite
survival, parasite-development rate inside vectors, and human and
vector population size. The manner in which these parameters
combine to determine malaria transmission was first proposed by
Ronald Ross [53] and later modified by MacDonald [54], who
developed the now standard Ross–MacDonald mathematical model
of malaria transmission. This model predicts malaria transmission
intensity in terms of the reproductive rate, R0, which is defined as
the number of new cases generated by one infected person in a
population of susceptibles, or:
R0 ¼ma2bce�dn
rd
where m is the number of vectors per person, a the number of
bites vectors take from humans each day, b and c the infectious-
ness of vectors to humans and vice versa, respectively, n the
incubation period of the parasite, and 1/r and 1/d the duration
of human infectiousness and mosquito longevity. The parameter
‘a’ best captures the impact of host choice on malaria transmission
rates. As the propensity of mosquitoes to feed on humans
increases, so does the value of ‘a’. Because this variable is expo-
nentially related to malaria transmission, even small changes in
mosquito preference for humans have a sizeable impact on
malaria transmission. No biological parameter other than mos-
quito survival has a greater impact on malaria transmission than
the feeding rate on humans. As summarized in Table 1, the
vertebrate species from which mosquitoes feed can also have a
direct impact on their survival; thus, host species choice could
make at least two independent and sizeable contributions to
malaria transmission intensity.
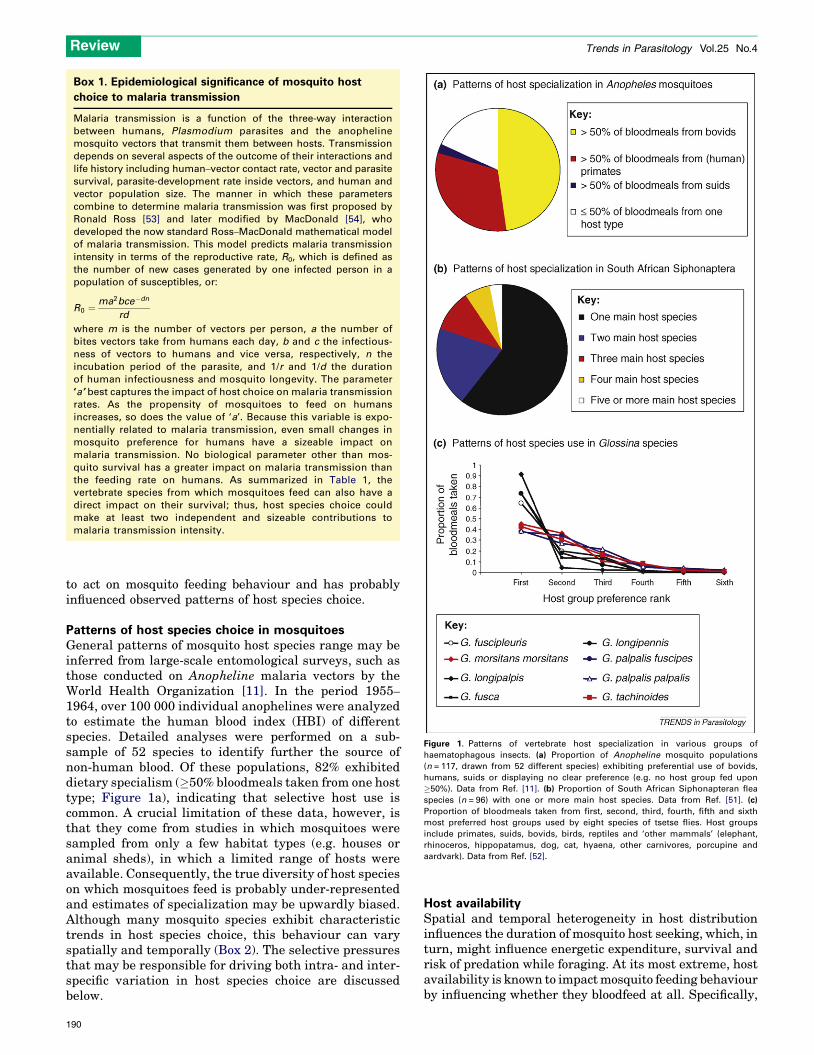
Figure 1. Patterns of vertebrate host specialization in various groups of
haematophagous insects. (a) Proportion of Anopheline mosquito populations
(n = 117, drawn from 52 different species) exhibiting preferential use of bovids,
humans, suids or displaying no clear preference (e.g. no host group fed upon
�50%). Data from Ref. [11]. (b) Proportion of South African Siphonapteran flea
species (n = 96) with one or more main host species. Data from Ref. [51]. (c)
Proportion of bloodmeals taken from first, second, third, fourth, fifth and sixth
most preferred host groups used by eight species of tsetse flies. Host groups
include primates, suids, bovids, birds, reptiles and ‘other mammals’ (elephant,
rhinoceros, hippopatamus, dog, cat, hyaena, other carnivores, porcupine and
aardvark). Data from Ref. [52].
Review Trends in Parasitology Vol.25 No.4
to act on mosquito feeding behaviour and has probablyinfluenced observed patterns of host species choice.
Patterns of host species choice in mosquitoesGeneral patterns of mosquito host species range may beinferred from large-scale entomological surveys, such asthose conducted on Anopheline malaria vectors by theWorld Health Organization [11]. In the period 1955–
1964, over 100 000 individual anophelines were analyzedto estimate the human blood index (HBI) of differentspecies. Detailed analyses were performed on a sub-sample of 52 species to identify further the source ofnon-human blood. Of these populations, 82% exhibiteddietary specialism (�50% bloodmeals taken from one hosttype; Figure 1a), indicating that selective host use iscommon. A crucial limitation of these data, however, isthat they come from studies in which mosquitoes weresampled from only a few habitat types (e.g. houses oranimal sheds), in which a limited range of hosts wereavailable. Consequently, the true diversity of host specieson which mosquitoes feed is probably under-representedand estimates of specialization may be upwardly biased.Although many mosquito species exhibit characteristictrends in host species choice, this behaviour can varyspatially and temporally (Box 2). The selective pressuresthat may be responsible for driving both intra- and inter-specific variation in host species choice are discussedbelow.
190
Host availabilitySpatial and temporal heterogeneity in host distributioninfluences the duration of mosquito host seeking, which, inturn, might influence energetic expenditure, survival andrisk of predation while foraging. At its most extreme, hostavailability is known to impactmosquito feeding behaviourby influencing whether they bloodfeed at all. Specifically,

Box 2. Source and examples of within-species variation in
mosquito host species choice
Geographical
In the Northern Hemisphere, the mosquito Culex pipiens in the
Mediterranean region feeds on mammals or on both mammals and
birds, whereas those further north are entirely ornithophilic [55].
The malaria vector An. gambiae s.s. is highly anthropophilic in
mainland Africa but prefers to feed on dogs on the island of Sao
Tome [56]. Although the mosquito Anopheles arabiensis is known
to prefer to feed on cows, it can be highly anthropophilic when
livestock are absent [57]. Similar spatial variation in host choice has
been observed within Aedes bromeliae (reported as Ae. Simpsoni)
in Uganda, where rodents are the preferred hosts at one site and
humans at another [55]. The underlying causes of such spatial
variation in mosquito host choice are unclear but in some cases,
may be linked to local host availability.
Seasonal
In North America, culicine mosquitoes including Culex tarsalis and
Culex nigripalpus switch their host species choice from birds in
spring and early summer to mammals in late summer and autumn
[58]. In addition, in Kenya, the feeding pattern of Culex univittatus
switches from mammals to avians during the long rains [59]. In
some cases, it has been hypothesised that these shifts are due to
seasonal changes in host availability [59] and climatic conditions
[55,59], and in other cases, the potential cause remains unclear [55].
Microhabitat
In western Kenya, the HBI of several culicine species was found to
vary with their site of capture; human blood was more prevalent in
mosquitoes caught inside houses, and bovid blood was more
common in those captured resting outdoors [60]. Similarly, the HBI
of An. arabiensis caught in houses was three times higher than in
those caught outdoors [61]. Thus, feeding preference of mosquitoes
may be influenced by availability of potential hosts in the particular
microhabitat in which they are found.
Foraging experience
Mosquito host choice may also be influenced by prior foraging
experience that causes them to learn which hosts are most
successfully fed upon. Several studies have attempted to test this
possibility by marking and releasing mosquitoes to assess their
tendency to return to their original location (for a review, see Ref.
[62]). Although several studies indicated that mosquitoes return to
their initial point of capture at higher than expected rates, it is
unclear whether this is evidence of learning [62] or genetic and
environmental factors that influence attraction to particular sites. To
avoid these confounding factors, an experimental study was
conducted in which mosquitoes known to have fed on either pigs
or cows during their first meal were allowed to choose between
these hosts on their second feed [63]. In this study, Aedes species
showed no host fidelity, whereas Culex tritaeniorhynchus, Culex
gelidus and Culex vishnui were substantially more likely to choose
the host species that was first fed upon for their second feed [63].
Because the offspring of these mosquitoes did not display a similar
host choice to their parents, it was concluded that the observed host
fidelity was more likely a product of learning than of genetic
predisposition. Further investigation with similarly robust experi-
mental designs are required to confirm the role of learning as a
determinant of mosquito host species choice.
Review Trends in Parasitology Vol.25 No.4
autogeneous egg production inmosquitoes has been associ-ated with environments in which vertebrate host avail-ability is severely limited [12].
In haematophagous species, two approaches have beenused to assess the dependence of host species choice on hostavailability. The first (andmost common) is to qualitativelycompare whether the proportion of bloodmeals taken fromparticular host species varies with their relative abun-dance in the environment, which almost always finds a
positive relationship (see, for example, Refs [13,14]).Although this approach is useful for evaluating the sensi-tivity of host choice to availability, a more quantitativeapproach is required to test how extensively patterns ofhost species choice can be attributed to availability. Onesuch measure is the forage ratio (FR), which is calculatedas the proportion of bloodmeals taken from a particularhost species divided by its relative abundance within thehost community [15]. FR values of >1 imply a preferencefor the host species, values of <1 are indicative of avoid-ance, whereas values of�1 indicate random foraging on allavailable hosts [15]. Probably because of the difficulty inconducting accurate host censuses, this technique is rarelyused in practice and has been criticized for its assumptionthat the presence of a host implies it is ‘available’ [16]. Kayet al. [16] proposed the ‘feeding index’ (FI), which requiresonly an approximation of the relative abundance of the twohost species on which feeding is compared, as an alterna-tive. As with the FR, values of the FI that are substantiallydifferent to 1 indicate non-random host choice. In the fewstudies that have calculated either an FI [15,17] or an FR[16,18], values are almost always considerably lower orhigher than 1. Taken together with results from qualita-tive studies, this indicates that although mosquito hostspecies choices are sensitive to host availability, true pre-ferences are evident that cannot simply be attributed torandom foraging on available host species.
Nutritional value of blood and energetic costs ofdigestionNon-random feeding of mosquitoes may be explained byvariation in nutritional rewards and corresponding fitnessaccruing from different host types. Experimental studiesconsistently indicate that the reproduction and survival ofmosquitoes after bloodfeeding varies with host species(Table 1). Several haematological properties vary betweenvertebrate species, which could influence the nutritivevalue of their blood [19,20]. During feeding, mosquitoespass ingested blood through the pyloric armature situatedat the posterior of the gut [21]. These sclerotized teeth-likestructures enable the passage of serum but trap red bloodcells [21], which are the primary source of protein for eggproduction. The number and complexity of these teeth varyamong mosquito species [22], as does red blood-cell sizeamong vertebrate species [20]. It has been hypothesizedthatmosquitoesmay specialize on host species whose bloodcan be most efficiently filtered by their particular pyloricarmature [22], although this remains to be demonstrated.The isoleucine content of vertebrate blood has been associ-ated with mosquito egg production, and its variation be-tween vertebrate species has been hypothesized toinfluence host selection [19]. However, artificial manipula-tion of isoleucine in blood did not influence the fecundity ofAe. aegypti, indicating this amino acid may be a correlaterather than cause of fecundity variation in nature [19].
Even if the base nutritional value of blood from differenthost sources is similar, the energetic costs of digesting itmight not be. Blood digestion requires a substantial expen-diture of energy; the metabolic rate of the mosquito Culextarsalis, for example, doubles when it is consuming blood incomparison to when it is consuming sugar water [23]. The
191
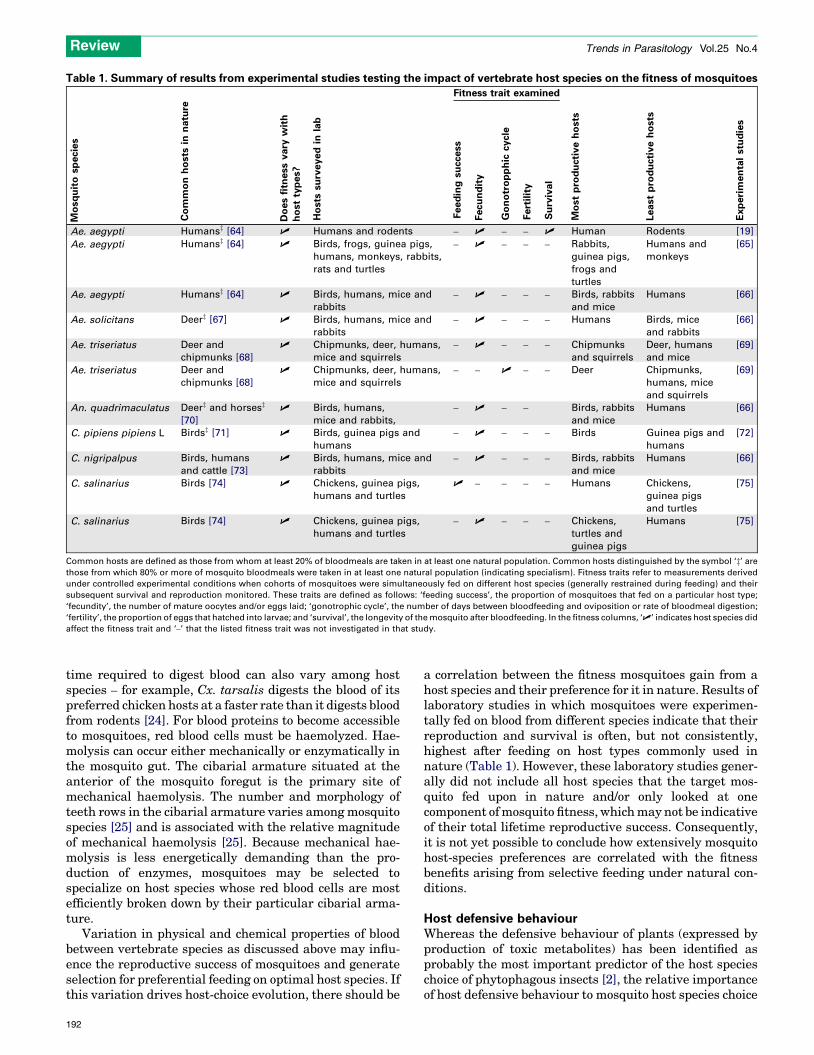
Table 1. Summary of results from experimental studies testing the impact of vertebrate host species on the fitness of mosquitoes
Fitness trait examined
Mo
squ
ito
speci
es
Co
mm
on
ho
sts
inn
atu
re
Do
es
fitn
ess
vary
wit
hh
ost
typ
es?
Ho
sts
surv
eyed
inla
b
Feed
ing
succ
ess
Fecu
nd
ity
Go
no
tro
pp
hic
cycl
e
Fert
ilit
y
Su
rviv
al
Mo
stp
rod
uct
ive
ho
sts
Least
pro
du
ctiv
eh
ost
s
Exp
eri
men
tal
stu
die
s
Ae. aegypti Humansz [64] U Humans and rodents – U – – U Human Rodents [19]
Ae. aegypti Humansz [64] U Birds, frogs, guinea pigs,
humans, monkeys, rabbits,
rats and turtles
– U – – – Rabbits,
guinea pigs,
frogs and
turtles
Humans and
monkeys
[65]
Ae. aegypti Humansz [64] U Birds, humans, mice and
rabbits
– U – – – Birds, rabbits
and mice
Humans [66]
Ae. solicitans Deerz [67] U Birds, humans, mice and
rabbits
– U – – – Humans Birds, mice
and rabbits
[66]
Ae. triseriatus Deer and
chipmunks [68]
U Chipmunks, deer, humans,
mice and squirrels
– U – – – Chipmunks
and squirrels
Deer, humans
and mice
[69]
Ae. triseriatus Deer and
chipmunks [68]
U Chipmunks, deer, humans,
mice and squirrels
– – U – – Deer Chipmunks,
humans, mice
and squirrels
[69]
An. quadrimaculatus Deerz and horsesz
[70]
U Birds, humans,
mice and rabbits,
– U – – Birds, rabbits
and mice
Humans [66]
C. pipiens pipiens L Birdsz [71] U Birds, guinea pigs and
humans
– U – – – Birds Guinea pigs and
humans
[72]
C. nigripalpus Birds, humans
and cattle [73]
U Birds, humans, mice and
rabbits
– U – – – Birds, rabbits
and mice
Humans [66]
C. salinarius Birds [74] U Chickens, guinea pigs,
humans and turtles
U – – – – Humans Chickens,
guinea pigs
and turtles
[75]
C. salinarius Birds [74] U Chickens, guinea pigs,
humans and turtles
– U – – – Chickens,
turtles and
guinea pigs
Humans [75]
Common hosts are defined as those from whom at least 20% of bloodmeals are taken in at least one natural population. Common hosts distinguished by the symbol ‘z’ are
those from which 80% or more of mosquito bloodmeals were taken in at least one natural population (indicating specialism). Fitness traits refer to measurements derived
under controlled experimental conditions when cohorts of mosquitoes were simultaneously fed on different host species (generally restrained during feeding) and their
subsequent survival and reproduction monitored. These traits are defined as follows: ‘feeding success’, the proportion of mosquitoes that fed on a particular host type;
‘fecundity’, the number of mature oocytes and/or eggs laid; ‘gonotrophic cycle’, the number of days between bloodfeeding and oviposition or rate of bloodmeal digestion;
‘fertility’, the proportion of eggs that hatched into larvae; and ‘survival’, the longevity of the mosquito after bloodfeeding. In the fitness columns, ‘U’ indicates host species did
affect the fitness trait and ‘–’ that the listed fitness trait was not investigated in that study.
Review Trends in Parasitology Vol.25 No.4
time required to digest blood can also vary among hostspecies – for example, Cx. tarsalis digests the blood of itspreferred chicken hosts at a faster rate than it digests bloodfrom rodents [24]. For blood proteins to become accessibleto mosquitoes, red blood cells must be haemolyzed. Hae-molysis can occur either mechanically or enzymatically inthe mosquito gut. The cibarial armature situated at theanterior of the mosquito foregut is the primary site ofmechanical haemolysis. The number and morphology ofteeth rows in the cibarial armature varies amongmosquitospecies [25] and is associated with the relative magnitudeof mechanical haemolysis [25]. Because mechanical hae-molysis is less energetically demanding than the pro-duction of enzymes, mosquitoes may be selected tospecialize on host species whose red blood cells are mostefficiently broken down by their particular cibarial arma-ture.
Variation in physical and chemical properties of bloodbetween vertebrate species as discussed above may influ-ence the reproductive success of mosquitoes and generateselection for preferential feeding on optimal host species. Ifthis variation drives host-choice evolution, there should be
192
a correlation between the fitness mosquitoes gain from ahost species and their preference for it in nature. Results oflaboratory studies in which mosquitoes were experimen-tally fed on blood from different species indicate that theirreproduction and survival is often, but not consistently,highest after feeding on host types commonly used innature (Table 1). However, these laboratory studies gener-ally did not include all host species that the target mos-quito fed upon in nature and/or only looked at onecomponent ofmosquito fitness, whichmay not be indicativeof their total lifetime reproductive success. Consequently,it is not yet possible to conclude how extensively mosquitohost-species preferences are correlated with the fitnessbenefits arising from selective feeding under natural con-ditions.
Host defensive behaviourWhereas the defensive behaviour of plants (expressed byproduction of toxic metabolites) has been identified asprobably the most important predictor of the host specieschoice of phytophagous insects [2], the relative importanceof host defensive behaviour to mosquito host species choice

Review Trends in Parasitology Vol.25 No.4
remains unclear. Animals defend themselves from insectbites by a variety of means, including protective tissues(such as skin and feathers), physical movements andbehavioural avoidance (for a review, see Ref. [1]).Additional artificial defensive measures developed byhumans, such as house screening [26] and insecticide-treated nets (ITNs), are also highly effective means ofreducing the feeding success of mosquitoes [27].
Such defensive behaviour can influence the feedingsuccess of mosquitoes by physically blocking and/or killingthemwhile they are attempting to feed [28] and by causingthem to abandon feeding attempts [29]. Numerous studieshave demonstrated that defensive reactions to mosquitobiting vary among host species [1,30]. However, the defen-siveness of particular host species usually varies as afunction of mosquito density [31,32]; thus, there may beno consistent host-specific defense phenotype on whichnatural selection can act. Further investigation of therelative magnitude of variation in defensiveness withinand between host species is required to test whether thisbehaviour can explain patterns of mosquito host choice.
Other potential determinants of host species choiceOther physiological, behavioural and ecological factorsmay also play a part in mosquito host-choice evolution.Vertebrate hosts mount immune responses to arthropodsaliva, which can impede the feeding success, reproductionand survival of insects that subsequently bite them [33,34].Variation in immunocompetence among host species couldgenerate selection for specialization on less-responsivehost types. Biochemical and physiological properties ofmosquito saliva could also influence their ability to exploitdifferent hosts. Mosquito saliva consists of a complexmixture of vasodilators, anti-platelet compounds andimmune-modulating compounds [35]. The efficiency withwhich the saliva from a particular mosquito species cananaesthetize and extract blood may vary among hostspecies and, again, generate selection for preferential feed-ing on those whose blood can be most efficiently exploited.To our knowledge, these possibilities have not been inves-tigated.
Finally, mosquito host species choice may, in part, be aby-product of favourable environmental conditions thatdrawmosquitoes to habitats where only a particular subsetof hosts are available, and not the direct properties of thehosts themselves. Manipulative experiments in which therelative fitness obtained from feeding on hosts in commonand novel environments are required to test this possib-ility.
Contrasting mosquitoes with other haematophagousinsectsAs in mosquitoes, specialization on a limited number ofhost species is common in other haematophagous insectssuch as fleas, where many taxa feed principally on only onehost species (e.g. Figure 1b). Similarly, although the hostchoice of tsetse flies seems to be more evenly distributedacross different host species, some types are still fed uponsubstantially more often than others (Figure 1c).
Most information on the relationship between hostavailability and host species choice comes from studies
of mosquitoes (see above). However, there is evidenceindicating that the host species choice of triatomids andtsetse flies varies with host availability and proximity[36,37]. Like mosquitoes, however, these insects still exhi-bit preferences that cannot simply be explained by hostavailability.
Blood nutritive value and the ‘cost’ of digestion seems tobe clearly associated with the host species choice of somehaematophagous insects. The flea Parapulex chephrenisrequires less energy and time to digest the blood of itspreferred mouse host [38] than that of non-preferred gerbilhosts, and it also has higher fecundity and survival afterfeeding on preferred host species [39]. Similarly, the fleaCtenocephalides felis also has higher reproductive successon common rather than atypical host species [40]. Hostdefensive behaviour has been also linked to the feedingsuccess of other haematophagous insects, including tsetseflies and horse flies [32,41]. As is the case with mosquitoes,it is unclear whether intrinsic variation in defensive beha-viour between host species is linked to host choice in thesesystems.
Mosquito host choice evolution in a dynamic worldCurrent patterns of host species choice by mosquitoes arethe product of millions of generations of natural selection.Change in the abundance and diversity of host speciesrelative to insect generation length may have occurredslowly through much of this period but has escalatedrecently because of rapid human population growth andassociated changes in vertebrate species diversity. Recentchanges in vertebrate species communities that increasemosquito preference for humans could dramaticallyincrease the transmission of diseases such as malaria,dengue and filariasis and undermine currently effectivecontrol strategies. Unfortunately, preliminary evidencesuggests such changes in human preference may alreadybe occurring in response to some land-use activities. Forexample, the proportion of bloodmeals taken from humansby the malaria vector Anopheles fluviatilis in Uttar Pra-desh increased from 1.4% in 1938–1939 to 41.2% in 1949–
1952, which was attributed to forest clearing and theintensification of rural agriculture [11]. Similar changesare being observed around the Amazon basin, where thehuman biting rate of Anopheles darlingi is 278 timesgreater in deforested areas than in forested areas [42].
The tendency for increasedmosquito feeding on humansas a consequence of landscape and demographic changes,however, may be counteracted by the increased uptake ofvector-control tools that selectively protect people. Thewidespread usage of ITNs has been associated with sub-stantial shifts in mosquito bloodfeeding from humans toanimals in some [43], but not all, areas [44]. Similarreductions in the HBI of malaria vectors have beenobserved in areas undergoing indoor residual spraying(IRS), although observance of this phenomenon variedbetween geographical areas, between mosquito speciesand with the type of insecticides used [45]. It remainsunclear whether observed reductions in anthropophily inresponse to ITNs or IRS were the result of phenotypicplasticity or an underlying genetic change in the vectorpopulation.
193

Review Trends in Parasitology Vol.25 No.4
Can shifts in host-species use be manipulated?The idea of reducing pathogen transmission by altering thehost species choice of mosquito vectors is not new. Thisconcept, known as zooprophylaxis, has been advocated as amalaria control strategy by the WHO since 1982 [46], withthe guiding principle that transmission will be lowered bydiverting mosquitoes from humans onto livestock hoststhat cannot transmit Plasmodium. There have been fewrigorous large-scale trials of zooprophylaxis as a malariacontrol tool. Available empirical and theoretical evidence ismixed and indicates that the presence of livestock couldreduce [47], enhance [48] or have no effect on transmission[49]. Although there has been some discussion of howlivestock could actually increase malaria risk (indirectly,by attracting more mosquitoes and/or creating more larvalhabitats [48]), what has so far been missed from thezooprophylaxis debate is the potential evolutionary andecological consequences of enforcing a mass host-speciesshift on vector populations.
As discussed here, many mosquitoes (including import-ant vectors of human disease) are highly selective in theirhost species choice. Accepting the evolutionary principlethat such specificity arises only when there are fitnessadvantages from being selective, it follows that divertingmosquitoes onto less-preferred hosts will change their netreproductive output. Evidence reviewed here (Table 1)confirms that mosquito host species choice routinely influ-ences their subsequent survival and reproduction. It is,thus, unlikely that mosquito fitness will remain constant iftheir host species use is switched by zooprophylactic diver-sion. From a disease-control perspective, this has twoconsequences. First, it indicates zooprophylactic strategiescould bring ‘added value’ by lowering mosquito reproduc-tion and survival, which, in turn, could reduce the growthrate and stability of their populations. Second and lessbeneficially, it indicates that any inherent fitness advan-tages from feeding on humans will always provide anincentive for anthropophilic mosquitoes to evade controlmeasures that reduce human contact.
A more effective means of conducting zooprophylaxismay be not only to divert mosquitoes from humans but alsoto decrease the fitness benefits gained from them. Toachieve this, it is first necessary to prove anthropophilicspecies gain an advantage from feeding on humans andidentify the mechanism through which it arises: greateravailability, higher blood nutritional value and/or lowerdefensiveness. Once identified, the advantage of humanfeeding could potentially be reduced through selectiveapplication of control measures. For example, there isevidence that the host choice of some mosquito species iscorrelated with the reproductive success obtained fromthat host. Successful development of a vaccine that reducesthe survival and fecundity of mosquitoes after feeding onhumans [33] could be one means of reducing thisadvantage. Similarly, if it can be proven that humans havepoorer defensive behaviour than other available hosts,control measures that increase mortality associated withtrying to feed on people (such as ITNs) may be the quickestway to select against anthropophily. Efforts must also bemade to identify the genes involved in mosquito hostchoice. If a mosquito host-species shift cannot be generated
194
through selection, a transgenic approach based on drivinggenes for zoophily into vector populations could also beeffective.
A starting point for either of these enterprises is identi-fication of the selective forces that cause mosquitoes tospecialize on humans. This will require experimentalinvestigation of mosquito–host interactions of similar thor-oughness to those conducted on herbivore–plant systems[2]. Assessing the mosquito-fitness consequences of inter-actions with human hosts presents logistical and ethicalchallenges not applicable in studies of plants, especially ifthe mosquitoes are disease vectors. However, the devel-opment of large-scale semi-field systems [50] will make itpossible to observe human, animal and mosquito beha-viour under relatively natural experimental conditionswithout risking exposure to parasites. These systems willmake it possible to compare the defensive behaviour ofhumans and animals to mosquito biting and measuremosquito fitness under varying host availability andenvironmental conditions. Use of such tools, in combi-nation with rigorous field investigation of genetic andenvironmental variation inmosquito feeding behavior, willhelp reveal the basis of their host species choice and guidethe development of new disease-control strategies based onsustained modification of mosquito behaviour.
AcknowledgementsThis work was funded by a Faculty of Biomedical and Life SciencesStudentship (UG) to I.L. and a BBSRC Fellowship to H.M.F.
References1 Lehane, M.J. (2005) The biology of blood-sucking in insects. Cambridge
University Press2 Jaenike, J. (1990) Host specialization in phytophagous insects. Annu.
Rev. Ecol. Syst. 21, 243–2733 Kiszewski, A. et al. (2004) A global index representing the stability of
malaria transmission. Am. J. Trop. Med. Hyg. 70, 486–4984 Takken, W. and Knols, B.G.J. (1999) Odor mediated behaviour of
Afrotropical malaria mosquitoes. Annu. Rev. Entomol. 44, 131–1575 Begon, M. et al. (2006) Ecology: From Individuals to Ecosystems.
Blackwell Publishing6 Egas, M. et al. (2004) Evolution restricts the coexistence of specialists
and generalists: The role of trade-off structure. Am. Nat. 163, 518–
5317 Kelly, D.W. and Thompson, C.E. (2000) Epidemiology and optimal
foraging: modelling the ideal free distribution of insect vectors.Parasitology 120, 319–327
8 Page, R.D.M. (2003) Tangled Trees: Phylogeny, Cospeciation, andCoevolution. University of Chicago Press
9 Gillies, M.T. (1964) Selection for host preference in Anopheles gambiae.Nature 203, 852–854
10 Mukwaya, L.G. (1977) Genetic control of feeding preferences inmosquitoes Aedes simpsoni (Stegomyia) and Aedes aegypti(Stegomyia). Physiol. Entomol. 2, 133–145
11 Bruce-Chwatt, L.J. et al. (1966) Ten years’ study (1955-64) of hostselection by anopheline mosquitos. Bull. World Health Organ. 35, 405–
43912 Corbet, P.S. (1967) Facultative autogeny in arctic mosquitoes. Nature
215, 662–66313 Zimmerman, R.H. et al. (2006) Bloodmeal hosts of Anopheles species
(Diptera: Culicidae) in a malaria-endemic area of the BrazilianAmazon. J. Med. Entomol. 43, 947–956
14 White, G.B. (1974) Anopheles gambiae complex and diseasetransmission in Africa. Trans. R. Soc. Trop. Med. Hyg. 68, 278–301
15 Hess, A.D. et al. (1968) The use of the forage ratio technique inmosquito host preference studies. Mosq. News 28, 386–389
16 Kay, B.H. et al. (1979) Application of the ‘feeding index’ concept tostudies of mosquito host-feeding patterns. Mosq. News 39, 68–72

Review Trends in Parasitology Vol.25 No.4
17 Lardeux, F. et al. (2007) Host choice and human blood index ofAnopheles pseudopunctipennis in a village of the Andean valleys ofBolivia. Malar. J. 6, 1–14
18 Loyola, E.G. et al. (1993) Anopheles albimanus (Diptera, Culicidae)host selection patterns in 3 ecological areas of the coastal plains ofChiapas, southern Mexico. J. Med. Entomol. 30, 518–523
19 Harrington, L.C. et al. (2001) Why do female Aedes aegypti (Diptera:Culicidae) feed preferentially and frequently on human blood? J. Med.Entomol. 38, 411–422
20 Wintrobe,M.M. (1933) Variations in the size and hemoglobin content oferythrocytes in the blood of various vertebrates. Folia Haematol.(Frankf.) 51, 32–49
21 Trembley, H.L. (1951) Pyloric spines in mosquitoes. J. Natl. Malar.Soc. 10, 213–215
22 Vaughan, J.A. et al. (1991) Concentration of human erythrocytes byanopheline mosquitoes (Diptera: Culicidae) during feeding. J. Med.Entomol. 28, 780–786
23 Gray, E.M. and Bradley, T.J. (2003) Metabolic rate in female Culextarsalis (Diptera: Culicidae): Age, size, activity, and feeding effects. J.Med. Entomol. 40, 903–911
24 Downe, A.E.R. and Archer, J.A. (1975) Effects of different blood mealsources on digestion and egg production inCulex tarsalisCOQ (DipteraCulicidae). J. Med. Entomol. 12, 431–437
25 Colluzi, M. et al. (1982) Effect of cibarial armature of mosquitoes(Diptera, Culicidae) on blood-meal haemolysis. J. Insect Physiol. 28,885–888
26 Lindsay, S.W. et al. (2002) Reducing malaria by mosquito-proofinghouses. Trends Parasitol. 18, 510–514
27 Takken, W. (2002) Do insecticide-treated bednets have an effect onmalaria vectors? Trop. Med. Int. Health 7, 1022–1030
28 Waage, J.K. and Nondo, J. (1982) Host behaviour andmosquito feedingsuccess: an experimental study. Trans. R. Soc. Trop. Med. Hyg. 76,119–122
29 Walker, E.D. and Edman, J.D. (1985) The influence of host defensivebehaviour on mosquito (Diptera: Culicidae) biting persistence. J. Med.Entomol. 22, 370–372
30 Edman, J.D. and Scott, T.W. (1987) Host defensive behavior and thefeeding success of mosquitos. Insect Sci. Appl. 8, 617–622
31 Kelly, D.W. et al. (1996) Density-dependent feeding success in a fieldpopulation of the sandfly, Lutzomyia longipalpis. J. Anim. Ecol. 65,517–527
32 Waage, J.K. and Davies, C.R. (1986) Host-mediated competition in abloodsucking insect community. J. Anim. Ecol. 55, 171–180
33 Billingsley, P.F. et al. (2008) Mosquitocidal vaccines: a neglectedaddition to malaria and dengue control strategies. Trends Parasitol.24, 396–400
34 Tschirren, B. et al. (2007) Host condition and host immunity affectparasite fitness in a bird-ectoparasite system. Funct. Ecol. 21, 372–378
35 Ribeiro, J.M.C. and Francischetti, I.M.B. (2003) Role of arthropodsaliva in blood feeding: sialome and post-sialome perspectives.Annu. Rev. Entomol. 48, 73–88
36 Gurtler, R.E. et al. (1997) Shifting host choices of the vector of Chagasdisease, Triatoma infestans, in relation to the availability of hosts inhouses in north-west Argentina. J. Appl. Ecol. 34, 699–715
37 Clausen, P.H. et al. (1998) Host preferences of tsetse (Diptera:Glossinidae) based on bloodmeal identifications. Med. Vet. Entomol.12, 169–180
38 Sarfati, M. et al. (2005) Energy costs of blood digestion in a host-specifichaematophagous parasite. J. Exp. Biol. 208, 2489–2496
39 Krasnov, B.R. et al. (2003) Host specificity and foraging efficiency inblood-sucking parasite: feeding patterns of the flea Parapulexchephrenis on two species of desert rodents. Parasitol. Res. 90, 393–399
40 Williams, B. (1993) Reproductive success of cat fleas, Ctenocephalidesfelis, on calves as unusual hosts. Med. Vet. Entomol. 7, 94–98
41 Torr, S.J. et al. (2001) Application of DNA markers to identify theindividual-specific hosts of tsetse feeding on cattle. Med. Vet. Entomol.15, 78–86
42 Vittor, A.Y. et al. (2006) The effect of deforestation on the human-bitingrate of Anopheles darlingi, the primary vector of falciparummalaria inthe Peruvian Amazon. Am. J. Trop. Med. Hyg. 74, 3–11
43 Bogh, C. et al. (1998) Permethrin-impregnated bednet effects on restingand feeding behaviour of lymphatic filariasis vector mosquitoes inKenya. Med. Vet. Entomol. 12, 52–59
44 Quinones, M.L. et al. (1997) Anopheles gambiae gonotrophiccycle duration, biting and exiting behaviour unaffected bypermethrin-impregnated bednets in The Gambia. Med. Vet.Entomol. 11, 71–78
45 Garrett-Jones, C. et al. (1980) Feeding habits of anophelines (Diptera:Culicidae) in 1971–1978, with reference to the human blood index: areview. Bull. Entomol. Res. 70, 165–185
46 WHO (1982) Manual on environmental management for mosquitocontrol: with special emphasis on malaria vectors. WHO Offset Publ.66, 2–284
47 Service, M.W. (1991) Agricultural development and arthropod-bornediseases: a review. Rev. Saude Publica 25, 165–178
48 Saul, A. (2003) Zooprophylaxis or zoopotentiation: the outcome ofintroducing animals on vector transmission is highly dependent onthe mosquito mortality while searching. Malar. J. 2, 32
49 Bogh, C. et al. (2001) Effect of passive zooprophylaxis on malariatransmission in the Gambia. J. Med. Ent. 38, 822–828
50 Ferguson, H.M. et al. (2008) Establishment of a large semi-field systemfor experimental study of African malaria vector ecology and control inTanzania. Malar. J. 7, 158
51 Segerman, J. (1995) Siphonaptera of southern Africa: handbook for theidentification of fleas. Publ. S. Instit. Med. Res. 57, 1–264
52 Weitz, B. (1963) The feeding habits of Glossina. Bull. World HealthOrgan. 28, 711–729
53 Ross, R. (1911) The prevention of malaria. John Murray, (London)54 Macdonald, G. (1957) The Epidemiology and Control of Malaria.
Oxford University Press55 Clements, A.N. (1999) Biology of mosquitoes. CABI Publishing56 Sousa, C.A. et al. (2001) Dogs as a favored host choice of Anopheles
gambiae sensu stricto (Diptera: Culicidae) of Sao Tome West Africa. J.Med. Entomol. 38, 122–125
57 Mwangangi, J.M. et al. (2003) Blood-meal analysis for anophelinemosquitoes sampled along the Kenyan coast. J. Am. Mosq. ControlAssoc. 19, 371–375
58 Kilpatrick, A.M. et al. (2006) West Nile virus epidemics in NorthAmerica are driven by shifts in mosquito feeding behaviour. PLoSBiol. 4, 606–610
59 Chandler, J.A. et al. (1977) Seasonal variation in the proportion ofmosquitoes feeding on mammals and birds at a heronry in westernKenya. J. Med. Entomol. 14, 233–240
60 Beier, J.C. et al. (1990) Relative abundance and blood feeding behaviorof nocturnally active culicine mosquitoes in western Kenya. J. Am.Mosq. Control Assoc. 6, 207–212
61 Petrarca, V. et al. (1991) Species composition of the Anopheles gambiaecomplex (diptera: Culicidae) at two sites in western Kenya. J. Med.Entomol. 28, 307–313
62 Alonso, W.J. and Schuck-Paim, C. (2006) The ‘ghosts’ that pesterstudies on learning in mosquitoes: guidelines to chase them off.Med. Vet. Entomol. 20, 157–165
63 Mwandawiro, C. et al. (2000) Heterogeneity in the host preference ofJapanese encephalitis vectors in Chiang Mai, northern Thailand.Trans. R. Soc. Trop. Med. Hyg. 94, 238–242
64 Ponlawat, A. and Harrington, L.C. (2005) Blood feeding patterns ofAedes aegypti and Aedes albopictus in Thailand. J. Med. Entomol. 42,844–849
65 Woke, P.A. (1937) Comparative effects of the blood of different speciesof vertebrates on egg production of Aedes aegypti. Am. J. Trop. Med.Hyg. 17, 729–745
66 Nayar, J.K. and Sauerman, D.M., Jr (1977) The effects of nutritionon survival and fecundity in Florida mosquitoes. Part 4. Effects ofblood source on oocyte development. J. Med. Entomol. 14, 167–
17467 Crans, W.J. et al. (1990) The blood feeding habits of Aedes sollicitans
Walker in relation to eastern equine encephalitis virus in CoastalAreas of New Jersey U.S.A.I. Host selection in nature determinedby precipitin tests onwild caught specimens.Bull. Soc. Vector. Ecol. 15,144–148
68 Nasci, R.S. (1982) Differences in host choice between the sibling speciesof treehole mosquitoes Aedes triseriatus and Aedes hendersoni. Am. J.Trop. Med. Hyg. 31, 411
69 Mather, T.N. and DeFoliart, G.R. (1983) Effect of host blood source onthe gonotrophic cycle of Aedes triseriatus. Am. J. Trop. Med. Hyg. 32,189
195

Review Trends in Parasitology Vol.25 No.4
70 Robertson, L.C. et al. (1993) Bionomics of Anopheles quadrimaculatusand Culex erraticus (Diptera: Culicidae) in the falls lake basin, NorthCarolina: Seasonal changes in abundance and gonotrophic status andhost feeding patterns. J. Med. Entomol. 30, 689–698
71 Hamer, G.L. et al. (2008) Culex pipiens (Diptera: Culicidae): a bridgevector of West Nile virus to humans. J. Med. Entomol. 45, 125–128
72 Shroyer, D.A. and Siverly, R.A. (1972) A comparison of egg productionof Culex pipiens pipiens L fed on avian and mammalian hosts. Mosq.News 32, 636–637
196
73 Gomes, A.C. et al. (2003) Host-feeding patterns of potential humandisease vectors in the ParaibaValley Region, State of Sao Paulo, Brazil.J. Vector Ecol. 28, 74–78
74 Cupp, E.W. and Stokes, G.M. (1976) Feeding patterns of Culexsalinarius Coquillett in Jefferson Parish, Louisiana. Mosq. News 36,332–335
75 Shelton, R.M. (1972) The effect of blood source and quantity onproduction of eggs by Culex salinarius coquillett (Diptera:Culicidae).Mosq. News 32, 31–38