eagle owl (bubo bubo) pellets from roman sagalassos (sw turkey): distinguishing the prey remains...
TRANSCRIPT

International Journal of OsteoarchaeologyInt. J. Osteoarchaeol. 19: 1–22 (2009)Published online 30 April 2008 in Wiley InterScience
65
(www.interscience.wiley.com) DOI: 10.1002/oa.9* Correspondence to: RoyalVautierstraat 29, B-1000 Bruse-mail: wvanneer@naturalscie
Copyright # 2008 Joh
Eagle Owl (Bubo bubo) Pelletsfrom Roman Sagalassos(SW Turkey): Distinguishingthe Prey Remains from Nestand Roost Sites
B. DE CUPERE,a S. THYS,a,b W. VAN NEER,a,b* A. ERVYNCK,c
M. CORREMANSd AND M. WAELKENSd
a Royal Belgian Institute of Natural Sciences, Vautierstraat 29, B-1000 Brussels, Belgiumb Katholieke Universiteit Leuven, Laboratory of Comparative Anatomy and Biodiversity, Ch.
Deberiotstraat 32, B-3000 Leuven, Belgiumc Flemish Heritage Institute, Phoenix Building, Koning Albert II-laan 19 Box 5, B-1210
Brussels, Belgiumd Katholieke Universiteit Leuven, Sagalassos Archaeological Research Project, Blijde
Inkomststraat 21, B-3000 Leuven, Belgium
ABSTRACT Two concentrations of animal bones, almost exclusively from small mammals and wild birds,were found within the destruction debris of a Roman bath complex in Sagalassos (SW Turkey).The overall species spectrum, skeletal element representation, fragmentation and preser-vation condition of the bones indicate that they represent the prey remains of a large nocturnalavian predator, more precisely the eagle owl (Bubo bubo). Differences in skeletal elementrepresentation and in prey species’ spectrum show that the two bone clusters derive frompellets deposited near a nest site and a roost site, respectively. Radiocarbon dates obtainedfrom the bones indicate that eagle owls lived in the collapsing bath complex during the secondhalf of the 6th to the beginning of the 7th century AD, before the final abandonment of the town.The MNI of the prey animals found at the nest site, confronted with the daily dietary needs of afemale eagle owl and its young, indicates repetitive use of the same place during severalyears. Copyright � 2008 John Wiley & Sons, Ltd.
Key words: eagle owl; pellets; prey choice; skeletal element representation; fragmentation;
nest site; roost site
Provenance of the faunal material
Since 1991, the beginning of excavations of theClassical town of Sagalassos (SW Turkey), faunalremains have been analysed, mainly documentingthe subsistence economy of the occupants (DeCupere, 2001). The urban contexts sometimes
Belgian Institute of Natural Sciences,sels, Belgium.nces.be
n Wiley & Sons, Ltd.
yielded remains of small vertebrates that representpenecontemporaneous and, occasionally, lateintrusives (sensu Gautier, 1987), but these findsalmost never occurred in real concentrations. Thepresent contribution describes two remarkableclusters of small animal remains that were foundin the destruction debris of the Roman Baths(Figures 1–2). Attention will focus on theidentification of the accumulator of the concen-trations, and on the establishment of the timeperiod that was needed for the assemblages to
Received 28 June 2007Revised 2 October 2007
Accepted 22 October 2007
Figure 1. Location of Sagalassos and its main buildings.
2 B. De Cupere et al.
accumulate. The elaborate archaeological inter-pretation of these finds – in view of thereconstruction of catastrophic events that mayhave taken place in the 6th and 7th centuriesAD – will be published elsewhere.The bath complex, located to the east of the
Lower Agora in the southern part of the city, hasbeen excavated since 1995. During the 2005campaign, excavations concentrated on the north-eastern section of the bath building, including thefrigidarium 1 (Figure 2). Three parts could be definedhere: the large ‘central hall’ formed by four massivepiers, and two adjoining ‘six piered halls’, one to
Copyright # 2008 John Wiley & Sons, Ltd.
the south and one to the north of the ‘central hall’.A high density of bones (hereafter referred to as‘cluster 1’) was found in the sediments around oneof the central piers of the ‘northern six piered hall’.A second concentration (‘cluster 2’) was locatedabout 20 m to the south of cluster 1, next to thesouthwestern pier of the ‘central hall’.
Frigidarium 1 was filled with several layers ofdestruction material, which were the result ofthe gradual collapse of this part of the building.The bone material of cluster 2 was recoveredfrom layer 5 from one and the same sector(2415–2385) and was inventoried under find
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Figure 2. The Roman baths with indication of the find spots of clusters 1 and 2.
Eagle Owl Pellets: Nest and Roost Sites 3
number SA-2005-RB2-80. The bones of cluster 1were collected from the two superimposed layers2 and 5 in two adjacent sectors (2415–2405 and2420–2405). The delimitation of these twoneighbouring sectors is unrelated to the archi-tecture of the building, but completely defined bythe fixed excavation grid system used for thewhole site. According to their provenance (i.e.layer and sector), three different find numbershave been distinguished in the field (Table 1).The material north of the pier roughly corre-sponds with find numbers SA-2005-RB1-43 and
Table 1. Provenance data of clusters 1 and 2
Sample number Cluste
SA-2005-RB1-26 SA-2005-R
Sector 2420–2405 2415–2Layer 2 2
Copyright # 2008 John Wiley & Sons, Ltd.
SA-2005-RB1-59, while the bone material to theeast and northeast of the pier correspondsroughly with find number SA-2005-RB1-26.Although the excavators considered the materialfrom these three find numbers as representing asingle set of events (and therefore designated it as‘cluster 1’), the three contexts were kept separateduring the faunal analyses and the subsequentprocessing of the data. The relevance of doingthis will be demonstrated below.Retrieval of the small bones in the trench was
facilitated by the rather loose structure of the
r 1 Cluster 2
B1-43 SA-2005-RB1-59 SA-2005-RB2-80
405 2415–2405 2415–23855 5
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

4 B. De Cupere et al.
sediment and by the conspicuous yellow-brownishcolour of the bones. A sediment sample of 69 litreswas taken of SA-2005-RB1-43 and was water-sieved on 4, 2 and 1mmmeshes. In addition to thematerial from the two clusters, some dispersedbones were also hand collected from neighbouringsectors and layers. These showed characteristicscomparable to the bones from the clusters, butsince their numbers were very low, they have beenomitted from this report.Two radiocarbon dates were obtained from
bones of cluster 1, one from a sample ofSA-2005-RB1-43 (layer 2) and another one fromSA-2005-RB1-59 (layer 5) (Table 2). When bothdates are calibrated and combined, using Oxcalv.3.10 (Bronk Ramsey, 1995, 2001), it appearsthat the bones were deposited during the periodof the second half of the 6th to the first quarter ofthe 7th century AD (95.4% probability), whichcorresponds to a period when the town was stillinhabited by the Sagalassians (Waelkens et al.,2006). The combination of the two radiocarbondates was statistically justifiable (x2 test: df¼ 1,T¼ 0.7 (5% 3.8)). The peak of the distribution ofthe combined calibrated dates lies around AD590. The radiocarbon dates indicate that layers 2and 5, at least in the area of cluster 1, do not differin date significantly and that therefore theaccumulation of debris in this part of the bathcomplex must have been rather rapid. Cluster 2,located in layer 5, has not been radiocarbondated, but is believed to be contemporaneousbecause of its stratigraphical position.
Species compositionof the assemblages
During the sorting of the faunal remains on site, itbecame clear that the assemblages consisted
Table 2. Radiocarbon dates for two bone samples from clu
Lab number Uncalibrated date
SA-2005-RB1-43 KIA-29165 1500� 25 BP
SA-2005-RB1-59 KIA-29164 1470� 25 BPCombined 1485� 18 BP
Calibrations were carried out using Oxcal v.3.10 (Bronk Rams2004).
Copyright # 2008 John Wiley & Sons, Ltd.
exclusively of the skeletal elements of smallerbirds and mammals. These were identified usingthe reference collection housed at the RoyalBelgian Institute of Natural Sciences (Brussels).Most difficulties were encountered with thepostcranial elements of rodents, since only someof them were very diagnostic (e.g. specificelements of jerboa, Dipodidae sp., and blindmole rat, Spalacinae sp.). Of the bird remains,small elements such as phalanges were also onlyrarely brought to species, and were usually leftunidentified. The identified skeletal elementswere recorded, and whenever possible assigned tothe left or right side of the body. The state offragmentation of the bones was assessed bynoting the part preserved as complete bone,proximal fragment, distal fragment or shaftfragment. The material was quantified by count-ing the number of identified specimens (NISP),and evaluating both the minimum mumber ofelements (MNE) and the minimum number ofindividuals (MNI). The MNI estimation wasbased on the best-represented element, treatingbones of either the right or the left body sideseparately, but, in addition, age differences(juvenile/subadult/adult) were considered.
The taxonomic composition of the two clustersis listed in Table 3; both NISP andMNI are given.The amount of identified remains in the 1mmfraction of the sieved sample of RB1-43 was verylow, and has therefore been omitted. Only half ofthe 2mm sieved residue was sorted and studied.
The best-represented species are chukar(Alectoris chukar), followed by hare (Lepus europaeus),jerboa (Allactaga euphratica) and hedgehog (Erina-ceus concolor). Duck remains are from smallerspecies, of which white-headed duck (Oxyuraleucocephala) and teal (Anas crecca) were identifiedwith certainty, as well as from larger species oftwo sizes, i.e. that of tufted duck (Aythya fuligula)
ster 1
Calibrated (68.2% prob.) Calibrated (95.4% prob.)
545 (68.2%)–595 AD 460 (1.9%)–490 AD530 (93.5%)–640 AD
565 (68.2%)–620 AD 550 (95.4%)–645 AD560 (68.2%)–605 AD 540 (95.4%)–620 AD
ey, 1995, 2001, with the atmospheric data of Reimer et al.,
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Table 3. Taxonomic composition of clusters 1 and 2, quantified as NISP and MNI
Cluster 1 Cluster 2
RB1-26 RB1-43 RB1-59 RB2-80
Trench Trench 4 mm 2 mm (50%) Trench Trench
NISP MNI NISP MNI NISP MNI NISP MNI NISP MNI NISP MNI
AvesIxobrychus minutus 2 1Anatinae:Anas crecca 1 1 34 5Oxyura leucocephala 1 1Size Aythya fuligula 2 1 8 2Size Anas plathrynchos 4 1 20 3
Accipiter sp. 5 1 5 1 1 1Falco tinnunculus 2 1Alectoris chukar 82 9 129 8 297 15 295 22 382 18Coturnix coturnix 5 2Fulica atra 14 3 2 1 20 2 3 1 44 4Gallinulus chloropus 1 1 2 1 6 1 8 1Rallus aquaticus 1 1 1 1 4 1 6 1Crex crex 1 1 10 2 5 1 20 3Philomachus pugnax 2 1Columba livia/C. oenas 2 1 5 1Columba palumbus 1 1 2 1 25 3Streptopelia sp. 4 1 2 1Otus scops 3 2 2 1 7 2Asio otus/A. flammeus 2 1 3 1 1Coracias garrulus 2 1Picus viridis 1 1Passeriformes:
Size Passer domesticus 1 1Size Sturnus vulgaris 4 1Size Turdus merula 3 1 2 1 2 1 1 1Size Pica pica 1 1 4 1Size Garrulus glandarius 2 1Corvus monedula 2 1 5 2 2 1C. c.cornix/C. frugilegus 7 2 3 1 16 4 9 3 3 1
MammaliaErinaceus concolor 120 12 1 1 24 2 3 2 7 1 15 3Mustela nivalis 17 2 4 1Lepus europaeus 140 6 163 9 919 27 182� 2 410 17 133 3Sciurus anomalus 5 2 3 1 5 2 9 2 5 1 2 1Allactaga euphratica 2 1 19 4 254 17 60� 10 75 11 2 1Dryomys nitedula 1 1Spalax leucodon 5 2 3 1 7 2 2 1 9 1Apodemus sp. 13 6 1 2Rattus rattus 2 1 4 2 1 2 1 1 1 1Mus musculus 6 4 2 2Cricetulus migratorius 24 6 7 4Microtidae sp. 6 2 5 4 1 1Rodentia sp. 4 2 254 375 14 12Total of Aves 124 26 149 17 386 37 0 0 321 32 573 51Total of Mammalia 278 24 195 18 1512 68 403 30 531 34 180 13Unidentified 52 75 2550 2210 288 519
� n
Eagle Owl Pellets: Nest and Roost Sites 5
and mallard (Anas plathyrhynchos). Other well-represented aquatic birds belong to the familyRallidae, of which the largest, the coot (Fulicaatra), is the most frequent. Among the pigeons
Lepus: almost exclusively metapodials, skull fragments a
Copyright # 2008 John Wiley & Sons, Ltd.
and the passerines the largest species, woodpigeon (Columba palumbus) and crows (Corvus coronecornix/Corvus frugilegus), are again the mostcommon. Raptor birds, including common scops
d loose teeth; Allactaga: about 50% are loose teeth.
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

6 B. De Cupere et al.
owl (Otus scops), long-eared and/or short-earedowl (Asio otus/flammeus), hawk (Accipiter sp.) andcommon kestrel (Falco tinnunculus) are alsoidentified. Mammals other than those mentionedabove include mainly small rodents, constitutingonly a minor part of the assemblage.The hare remains all represent juvenile animals
(Figure 3). Using the ageing criteria establishedby Broekhuizen &Maaskamp (1979) for the radiusand the ulna, most of the animals appear to havebeen killed at an age of three months or younger.Subadult or adult hares are completely absent inthe bone assemblages. The hedgehog remains, incontrast, are all from adult animals, while thejerboa is also represented by full-grown speci-mens. Among the bird remains, mainly adultindividuals were represented. Cluster 2 containsthe remains of some subadult individuals ofchukar, but their MNI is low (4 subadult versus
Figure 3. Three ulnae from hare (Lepus europaeus),belonging to different age classes. The scale bar is 5 cm.This figure is available in colour online at www.interscience.wiley.com/journal/oa.
Copyright # 2008 John Wiley & Sons, Ltd.
14 adult). No remains of subadult chukar werefound in cluster 1.
In the paragraphs below, the species compo-sition of the various assemblages will becompared on the basis of the material collectedin the trench, since wet sieving was not carriedout systematically. The effect of wet sieving onthe proportion of the major taxa in sampleRB1-43 is shown in Table 4 where separate valuesare provided for the NISP of: (1) all the materialretrieved from the trench; (2) the remains fromthe sieved sample collected from the 4mm sieve;and (3) the remains from the sieved samplecollected from both the 4 and the 2mm sieves.For the 2mm fraction, the original NISPs havebeen doubled to correct for the fact that only halfof the sieved residue was analysed. It is obviousfrom these data that especially the proportions ofrodents and chukar are heavily influenced by therecovery methods. As could be expected (cf.Thomas, 1969), the rodents are heavily under-represented in the hand-collected samples.Comparison of the various fractions has alsobeen done for the MNI, albeit in a slightlydifferent way, since for a correct establishment ofthe MNI all the bones from all the subsamplesshould be taken into account simultaneously. Itwas also felt that the original figures of the 2mmfraction cannot simply be doubled. Therefore theMNIs are given of the material collected in thetrench, those found in the 4mm fraction, andthose in half of the 2mm sieved residue. It appearsagain that sieving results in an increase of thesmall rodents.
Although the results from sample RB1-43 arethe ones that reflect most accurately the originalcomposition, it should be kept in mind thatdifferential preservation may also have had aninfluence on the species spectrum. As will beshown below, the information obtained from thesamples collected in the trench is sufficient toestablish the taphonomic agent that was respon-sible for their deposition.
A comparison of the frequency of the four mainspecies in the samples collected in the trenchshows differences between the two clusters, andeven variation within cluster 1 (Figure 4). Morethan 70% of the MNIs in cluster 2 belong tochukar, while hare and hedgehog are aboutequally frequent, and jerboa is only represented
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa
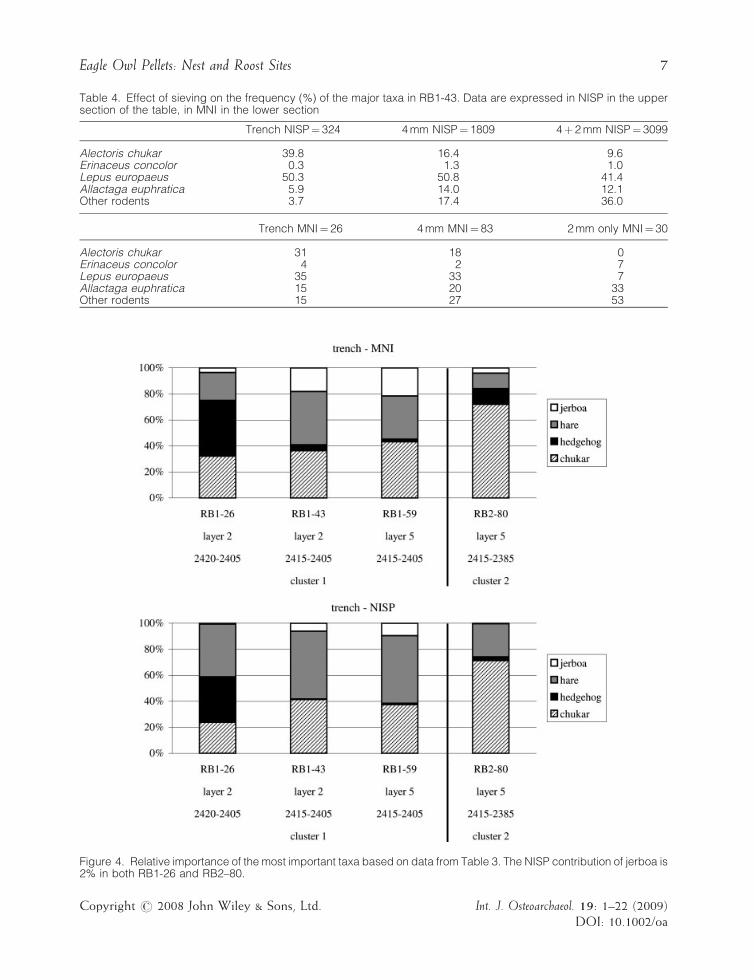
Table 4. Effect of sieving on the frequency (%) of the major taxa in RB1-43. Data are expressed in NISP in the uppersection of the table, in MNI in the lower section
Trench NISP¼ 324 4 mm NISP¼ 1809 4þ 2 mm NISP¼ 3099
Alectoris chukar 39.8 16.4 9.6Erinaceus concolor 0.3 1.3 1.0Lepus europaeus 50.3 50.8 41.4Allactaga euphratica 5.9 14.0 12.1Other rodents 3.7 17.4 36.0
Trench MNI¼ 26 4 mm MNI¼ 83 2 mm only MNI¼ 30
Alectoris chukar 31 18 0Erinaceus concolor 4 2 7Lepus europaeus 35 33 7Allactaga euphratica 15 20 33Other rodents 15 27 53
Figure 4. Relative importance of the most important taxa based on data from Table 3. The NISP contribution of jerboa is2% in both RB1-26 and RB2–80.
Copyright # 2008 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa
Eagle Owl Pellets: Nest and Roost Sites 7

8 B. De Cupere et al.
by a single individual (Figure 4, upper panel). Theproportion of the MNIs of chukar in the threeassemblages of cluster 1 is more or lesscomparable (32% to 43%), and significantlylower than in cluster 2. However, the proportionof hedgehog is much higher in sector 2420–2405than in sector 2415–2405 (43% versus 5%), whilethe highest number of jerboa is found in the lattersector (18% versus 4%). Differences betweenlayers 2 and 5 within the same sector are minimal.Similar results are obtained when the NISPs areconsidered (Figure 4, lower panel).
Figure 5. Traces of etching caused by gastric fluids on aproximal ulna of chukar (Alectoris chukar).
Preservation, skeletal elementrepresentation and fragmentation
The study of the preservation, skeletal elementrepresentation and fragmentation of bones hasbeen limited to the long bones of chukar, sinceit is believed to be the most informative forthe interpretation of the material in general.Chukar is a frequent taxon of relatively large sizein all the studied assemblages, and is mainlyrepresented by the remains of adult individuals.The chukar bones show a peculiar state of
preservation. While generally being in a goodcondition, clear traces of damage can be observed(Figure 5), mainly on the articular ends of thelong bones, irrespective of the skeletal elementconsidered. In contrast, the surfaces of the shaftsof the long bones do not seem to have beenattacked. Most of the fractures, however, showrounded edges or are thinned. Such clearlyrecognisable traces have been described in theliterature as being the result of digestion, of theimpact of the acid environment of an animal’sdigestive tract (Andrews, 1990; Bochenski &Tomek, 1997) through which the chukar remainsmust have passed. Dissolution of bone in acidsoils or sediments would have corroded all of thebone surface, and not only the articular ends.Complete bones going through the digestivetract of a predator show most damage at thearticular ends, less protected by surrounding softtissues (Andrews, 1990). In general, the effectof the gastric juices must not have been veryintense: the surface of the bone was partlydissolved but not altered in a severe way(Figure 5). Moreover, that even fragile or small
Copyright # 2008 John Wiley & Sons, Ltd.
bones have survived, and ended up in the bonedeposits, suggests a not too violent ‘chemicalattack’ on these remains.
The intraskeletal distribution of the long bonesof chukar can be interpreted from the NISP andMNE in Table 5. The subsamples from cluster 1were again not lumped together but treatedseparately, according to the sector and layer fromwhich they originate. It should be noted thatcomplete skulls were totally absent in bothclusters. Skull fragments were also seldom foundand included mainly fragments of the beak.Mandibles were somewhat better represented butwere not preserved completely either.
The frequency of wing bones (i.e. humerus,radius, ulna, carpometacarpus) within the assem-blage of wing and leg bones (femur, tibiotarsus,tarsometatarsus) has been calculated for bothclusters on the basis of the MNE counts(Figure 6). Within cluster 1, leg bones show aslight, almost negligible prevalence to wingbones (52% versus 48% for the total cluster), apattern that is present in all but one of thesubsamples. The inclusion of sieved material doesnot have an influence on the proportions. Cluster 2,on the other hand, shows a higher proportion ofwing bones (34% leg bones versus 66% wing
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa
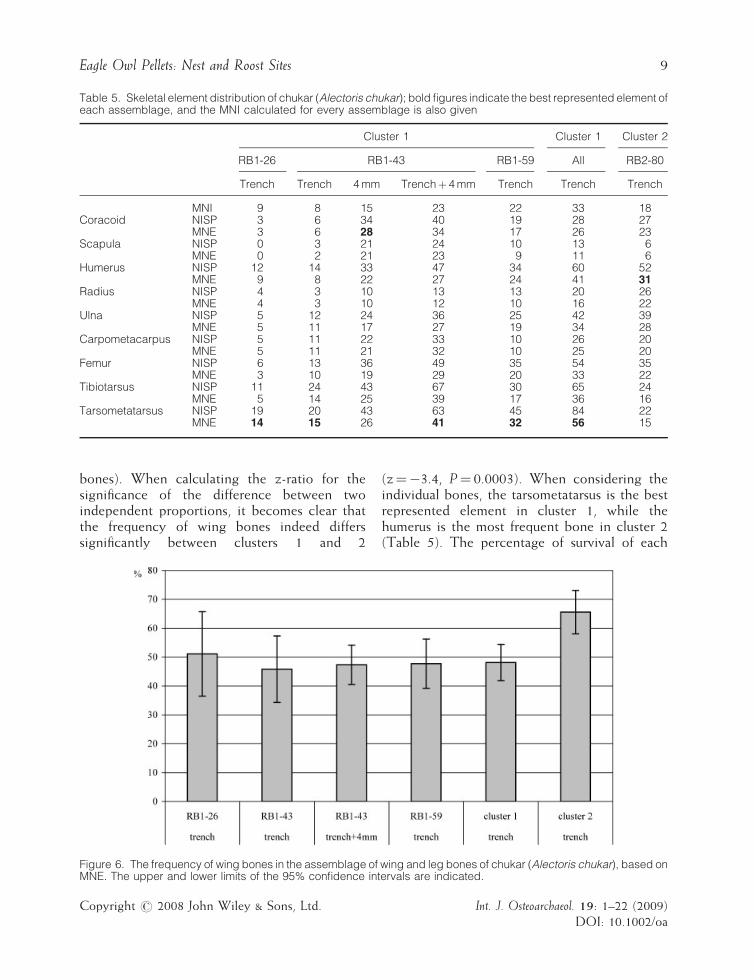
Table 5. Skeletal element distribution of chukar (Alectoris chukar); bold figures indicate the best represented element ofeach assemblage, and the MNI calculated for every assemblage is also given
Cluster 1 Cluster 1 Cluster 2
RB1-26 RB1-43 RB1-59 All RB2-80
Trench Trench 4 mm Trenchþ 4 mm Trench Trench Trench
MNI 9 8 15 23 22 33 18Coracoid NISP 3 6 34 40 19 28 27
MNE 3 6 28 34 17 26 23Scapula NISP 0 3 21 24 10 13 6
MNE 0 2 21 23 9 11 6Humerus NISP 12 14 33 47 34 60 52
MNE 9 8 22 27 24 41 31Radius NISP 4 3 10 13 13 20 26
MNE 4 3 10 12 10 16 22Ulna NISP 5 12 24 36 25 42 39
MNE 5 11 17 27 19 34 28Carpometacarpus NISP 5 11 22 33 10 26 20
MNE 5 11 21 32 10 25 20Femur NISP 6 13 36 49 35 54 35
MNE 3 10 19 29 20 33 22Tibiotarsus NISP 11 24 43 67 30 65 24
MNE 5 14 25 39 17 36 16Tarsometatarsus NISP 19 20 43 63 45 84 22
MNE 14 15 26 41 32 56 15
Eagle Owl Pellets: Nest and Roost Sites 9
bones). When calculating the z-ratio for thesignificance of the difference between twoindependent proportions, it becomes clear thatthe frequency of wing bones indeed differssignificantly between clusters 1 and 2
Figure 6. The frequency of wing bones in the assemblage ofMNE. The upper and lower limits of the 95% confidence int
Copyright # 2008 John Wiley & Sons, Ltd.
(z¼�3.4, P¼ 0.0003). When considering theindividual bones, the tarsometatarsus is the bestrepresented element in cluster 1, while thehumerus is the most frequent bone in cluster 2(Table 5). The percentage of survival of each
wing and leg bones of chukar (Alectoris chukar), based onervals are indicated.
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa
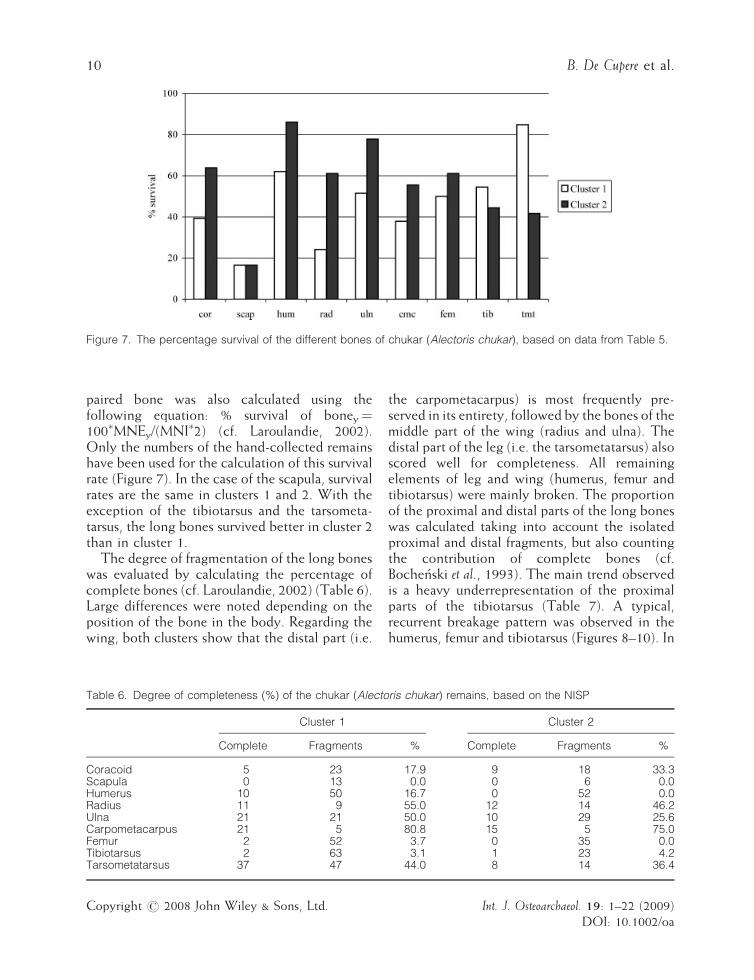
Figure 7. The percentage survival of the different bones of chukar (Alectoris chukar), based on data from Table 5.
10 B. De Cupere et al.
paired bone was also calculated using thefollowing equation: % survival of boney¼100�MNEy/(MNI�2) (cf. Laroulandie, 2002).Only the numbers of the hand-collected remainshave been used for the calculation of this survivalrate (Figure 7). In the case of the scapula, survivalrates are the same in clusters 1 and 2. With theexception of the tibiotarsus and the tarsometa-tarsus, the long bones survived better in cluster 2than in cluster 1.The degree of fragmentation of the long bones
was evaluated by calculating the percentage ofcomplete bones (cf. Laroulandie, 2002) (Table 6).Large differences were noted depending on theposition of the bone in the body. Regarding thewing, both clusters show that the distal part (i.e.
Table 6. Degree of completeness (%) of the chukar (Alecto
Cluster 1
Complete Fragments
Coracoid 5 23Scapula 0 13Humerus 10 50Radius 11 9Ulna 21 21Carpometacarpus 21 5Femur 2 52Tibiotarsus 2 63Tarsometatarsus 37 47
Copyright # 2008 John Wiley & Sons, Ltd.
the carpometacarpus) is most frequently pre-served in its entirety, followed by the bones of themiddle part of the wing (radius and ulna). Thedistal part of the leg (i.e. the tarsometatarsus) alsoscored well for completeness. All remainingelements of leg and wing (humerus, femur andtibiotarsus) were mainly broken. The proportionof the proximal and distal parts of the long boneswas calculated taking into account the isolatedproximal and distal fragments, but also countingthe contribution of complete bones (cf.Bochenski et al., 1993). The main trend observedis a heavy underrepresentation of the proximalparts of the tibiotarsus (Table 7). A typical,recurrent breakage pattern was observed in thehumerus, femur and tibiotarsus (Figures 8–10). In
ris chukar) remains, based on the NISP
Cluster 2
% Complete Fragments %
17.9 9 18 33.30.0 0 6 0.0
16.7 0 52 0.055.0 12 14 46.250.0 10 29 25.680.8 15 5 75.03.7 0 35 0.03.1 1 23 4.2
44.0 8 14 36.4
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Table 7. NISP for proximal and distal ends of long bones of chukar (Alectoris chukar)
Cluster 1 Cluster 2 Total
RB1-26 RB1-43 RB1-59 RB2-80
Trench Trench 4 mm Trench Trench
Humerus Proximal 6 4 22 12 20 64Distal 9 8 9 24 31 81
Radius Proximal 4 2 10 10 22 48Distal 3 13 6 9 16 47
Ulna Proximal 4 11 17 13 20 65Distal 5 10 17 19 30 81
Carpometacarpus Proximal 4 11 21 10 20 66Distal 5 9 18 8 15 55
Femur Proximal 3 10 19 20 13 65Distal 3 4 18 16 22 63
Tibiotarsus Proximal 3 7 17 7 9 43Distal 5 14 25 17 16 77
Tarsometatarsus Proximal 14 15 23 27 14 93Distal 12 12 18 32 15 89
Eagle Owl Pellets: Nest and Roost Sites 11
many cases, the proximal part of the humerus wasseparated from the shaft just below the articula-tion, while the shaft itself was still attached to thedistal end. Just below the articular end a hole cansometimes be seen in the shaft. In the case of thefemur, the proximal and distal ends were bothseparated from the shaft. The breakage patternfor the tibiotarsus was more similar to thatobserved in the humerus.
Figure 8. Fragmentation pattern in the humerus of chukaravailable in colour online at www.interscience.wiley.com/jou
Copyright # 2008 John Wiley & Sons, Ltd.
Digestion traces have also been observed onthe bones of other bird species, but becauseNISPs per species were too low, a taphonomicanalysis as presented for chukar could not bemade. The mammal remains also presentedinterpretation problems. Damage due to diges-tion was observed on the bones of adult mammals(Figure 11), but in the case of the juvenile hare itwas difficult to decide whether the porous state of
(Alectoris chukar). The scale bar is 5 cm. This figure isrnal/oa.
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Figure 9. Fragmentation pattern in the femur of chukar (Alectoris chukar). The scale bar is 5 cm. This figure is availablein colour online at www.interscience.wiley.com/journal/oa.
12 B. De Cupere et al.
preservation was due to the young age of theanimals (between one and three months) or todigestion. The immature nature of the hare bonesmay have had a significant impact during pre- andpost-depositional processes (Behrensmeyer,1978) resulting in an underrepresentation of
Figure 10. Fragmentation pattern in the tibiotarsus of chukaavailable in colour online at www.interscience.wiley.com/jou
Copyright # 2008 John Wiley & Sons, Ltd.
certain (parts of) skeletal elements and greaterfragmentation. Also the fact that the agedifferences resulted in different sizes of the longbones must have influenced the preservation (cf.Cochard, 2004). The immature nature of the harebones make them thus less suitable for attributing
r (Alectoris chukar). The scale bar is 5 cm. This figure isrnal/oa.
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa
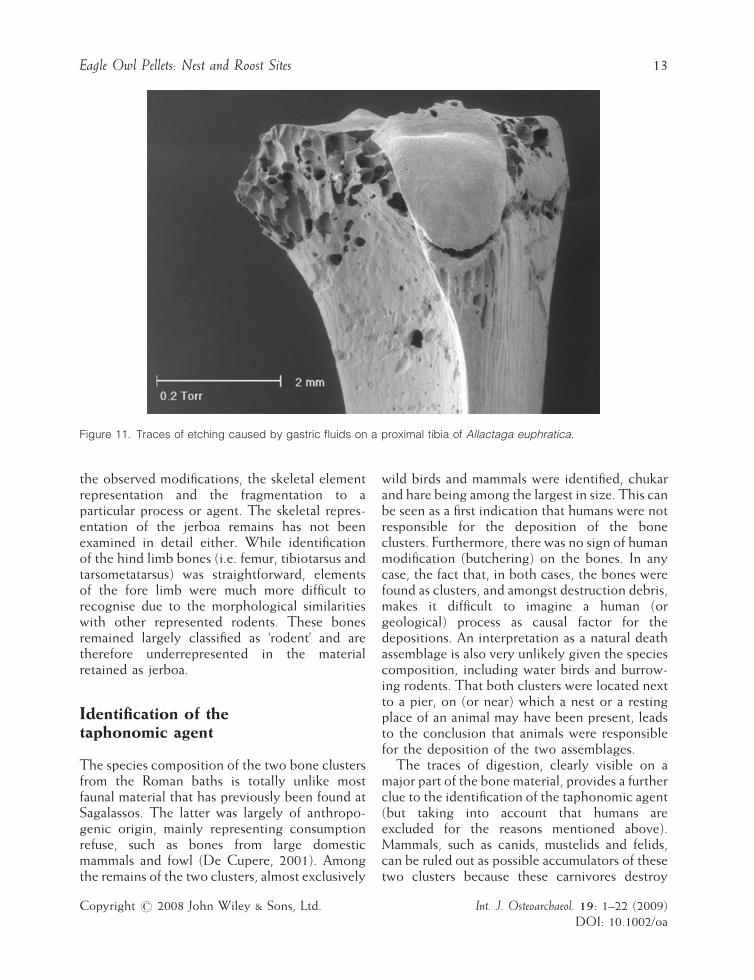
Figure 11. Traces of etching caused by gastric fluids on a proximal tibia of Allactaga euphratica.
Eagle Owl Pellets: Nest and Roost Sites 13
the observed modifications, the skeletal elementrepresentation and the fragmentation to aparticular process or agent. The skeletal repres-entation of the jerboa remains has not beenexamined in detail either. While identificationof the hind limb bones (i.e. femur, tibiotarsus andtarsometatarsus) was straightforward, elementsof the fore limb were much more difficult torecognise due to the morphological similaritieswith other represented rodents. These bonesremained largely classified as ‘rodent’ and aretherefore underrepresented in the materialretained as jerboa.
Identification of thetaphonomic agent
The species composition of the two bone clustersfrom the Roman baths is totally unlike mostfaunal material that has previously been found atSagalassos. The latter was largely of anthropo-genic origin, mainly representing consumptionrefuse, such as bones from large domesticmammals and fowl (De Cupere, 2001). Amongthe remains of the two clusters, almost exclusively
Copyright # 2008 John Wiley & Sons, Ltd.
wild birds and mammals were identified, chukarand hare being among the largest in size. This canbe seen as a first indication that humans were notresponsible for the deposition of the boneclusters. Furthermore, there was no sign of humanmodification (butchering) on the bones. In anycase, the fact that, in both cases, the bones werefound as clusters, and amongst destruction debris,makes it difficult to imagine a human (orgeological) process as causal factor for thedepositions. An interpretation as a natural deathassemblage is also very unlikely given the speciescomposition, including water birds and burrow-ing rodents. That both clusters were located nextto a pier, on (or near) which a nest or a restingplace of an animal may have been present, leadsto the conclusion that animals were responsiblefor the deposition of the two assemblages.The traces of digestion, clearly visible on a
major part of the bone material, provides a furtherclue to the identification of the taphonomic agent(but taking into account that humans areexcluded for the reasons mentioned above).Mammals, such as canids, mustelids and felids,can be ruled out as possible accumulators of thesetwo clusters because these carnivores destroy
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

14 B. De Cupere et al.
bones to a high degree while handling anddigesting their prey (see Andrews, 1990). Thediagnostic tooth marks that they leave on theirfood remains are lacking. The bone assemblageshave more likely been accumulated by apredatory bird, since these animals cause lessdamage to the bones of their prey. Some taxashow the habit of eating their prey always at thesame locations, where they swallow (parts of) theanimals caught, digest the soft parts andregurgitate the inedible parts (bone, hair, claws,teeth) as a pellet. Accumulations of pellets atfeeding places may eventually lead to archae-ological clusters of numerous small bones. For theSagalassos finds, this points towards diurnalraptor birds such as falcons and buzzards, ornocturnal birds of prey (owls) (cf. Andrews,1990). Even raven is known to produce pelletswhich may contain bones (Laudet & Selva, 2004,2005), but this species can immediately beexcluded as a possible accumulator. Ravensgenerally gather in large numbers at roostingplaces, with about 100 individuals or even more.Their pellets are scattered around and notconcentrated in isolated clusters. Other speciesthat can be excluded a priori are vultures such asthe bearded vulture (Gypaetus barbatus), whose nestaccumulations are characterised by the dom-inance of medium-sized ungulates (especiallytheir phalanges). These elements are completelylacking in both clusters of Sagalassos. In addition,the bones from vulture nest sites show very severedigestion marks which we did not observe (seebelow) (Robert & Vigne, 2002).The diet of owls and diurnal raptors includes a
wide range of species (Cramp & Simmons, 1980;Cramp, 1985). The list of the identified taxa(Table 3) indicates that the predator to a largeextent preferred medium-sized birds (chukar,ducks, crows) and similar-sized mammals (younghares and hedgehogs). That small mammals suchas rodents are represented to a much lesser degreemay also be significant. Although the low overallfrequency of rodents in the assemblages is partly aresult of the sampling technique, a correction forfine sieving indicates that the non-rodentcomponent is still the major fraction (Table 4).The jerboa and unidentified rodents would makeup about 45% of the remains (in NISP). Becausestill more than half of the prey consisted of
Copyright # 2008 John Wiley & Sons, Ltd.
medium-sized birds and mammals, smaller pre-datory birds, both diurnal and nocturnal, can beexcluded as possible predators (for an overview ofdietary preferences of birds of prey, see Cramp &Simmons, 1980; Cramp, 1985; Bochenski, 2005).The material has most likely been deposited bylarge birds of prey, such as the golden eagle(Aquila chrysaetos) or the eagle owl (Bubo bubo),which can indeed take medium-sized birds andmammals but more rarely feed on smaller prey.
Criteria exist to distinguish the bone assem-blages accumulated by diurnal birds of prey fromthose deposited by owls. Around the nests ofdiurnal birds of prey, not only pellet contents arefound but also food remains that were notingested by the raptor (Bochenski et al., 1999;Laroulandie, 2002; Bochenski & Tornberg, 2003).Diurnal birds of prey have the tendency toconsume their prey bite by bite, starting at thehead and systematically working their waycaudally. However, they often leave part of thecarcass uneaten when hunger is satisfied (Dodson& Wexlar, 1979, and references therein), and thiswill result in an accumulation of bones withouttraces of digestion at the feeding place(Bochenski et al., 1998; Bochenski and Nekrasov,2001). In addition to those unmodified remains,there are bones that show a heavier alterationcompared with those deposited by owls(Andrews, 1990; Bochenski et al., 1997, 1998).Articular ends may dissolve completely, oftenleaving only shafts in the pellets of the diurnalraptors (Mayhew, 1977).
The type of food handling described above isvery unlike the behaviour seen in owls, who oftenswallow their prey whole. Even when this is notthe case (as with rather large prey items), speciessuch as the eagle owl regularly hide large prey, orparts of it, until the prey is completely eaten(Blondel & Badan, 1976). Therefore, most of thebones of the prey will ultimately show signs ofacid attack. The bone clusters of the Roman bathsvery frequently show clear digestion traces.Among the bird bones, all the different skeletalelements have been attacked by gastric juices andthere seems to be no specimens undamaged at all.Similar observations have been made for the adultmammals, indicating that the bone clusters fromSagalassos must be attributed to the activities ofowls. Finally, it is also striking that the digestion
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Eagle Owl Pellets: Nest and Roost Sites 15
traces occur only on the articular ends and not onthe shafts of long bones. As stated, in diurnalbirds these shafts are also affected (Bochenskiet al., 1998, 1999).Several species of owl occur in the region of
Sagalassos: the barn owl (Tyto alba), the commonscops owl (Otus scops), the eagle owl (Bubo bubo),the little owl (Athene noctua), the tawny owl (Strixaluco), the long-eared owl (Asio otus) and theshort-eared owl (Asio flammeus) (Kasparek, 1992;Kirwan et al., 1998). With the exception of theeagle owl, these species mainly hunt on rathersmall animals (insects, amphibians, reptiles, andsmall birds and rodents), although larger preymay exceptionally be killed, such as birds the sizeof mallard (Cramp, 1985). The species compo-sition of the prey identified from the two clustersat Sagalassos, with the dominance of medium-sized birds and mammals, indicates that the eagleowl must have been responsible for the deposit.The eagle owl indeed hunts on a wide variety ofanimals, ranging from insects to medium-sizedmammals and birds, depending on their avail-ability (e.g. Bezzel et al., 1976; Blondel & Badan,1976; Donazar & Ceballos, 1989; Papageorgiouet al., 1993; Rifai et al., 2000; Alivizatos et al., 2005;for more references see Cramp, 1985). Lago-morphs, hedgehogs and partridges are among theprey frequently taken (e.g. Orsini, 1985),although smaller and larger animals can also becaptured, such as fox-cubs (Blondel & Badan,1976). In the case of hedgehogs, the skin withspines is sometimes ripped off first, but often thespines are eaten as well (Cramp, 1985). Spineswere not preserved in the material of Sagalassos.Since the soil conditions at the site areunfavourable for the preservation of such organicsubstances, no inferences can be made about thelocal behaviour of the raptor.Besides the composition of the diet, the
fragmentation and modification of bird bonesfound in pellets can also be used to determinetheir producer, as the various owls show differentpatterns of damaging their prey (e.g. Bochenskiet al., 1993; Bochenski & Tomek, 1994). Usefulinformation on the skeletal element representa-tion, fragmentation and modification of birdbones typically deposited by eagle owl can befound in Bochenski et al. (1993), Bochenski &Nekrasov (2001) and Laroulandie (2002). Skulls
Copyright # 2008 John Wiley & Sons, Ltd.
and mandibles are rare in eagle owl pellets, bonesare often broken, and with the exception of thetibiotarsus, the proximal ends of the skeletalelements have been observed to outnumber thedistal ends (Bochenski et al., 1993; Bochenski &Nekrasov, 2001). A predominance of distal partsof tibiotarsus is indeed observed in the twoclusters from Sagalassos (Table 7), but in theother long bones the proximal parts do not alwaysoutnumber the distal parts, as expected byBochenski and colleagues (Figure 12). However,the scarcity of cranial elements and the frag-mented condition of the bones is in agreementwith the observations in the literature.The modifications observed by macroscopic
observation of the chukar bones from Sagalassosindeed also indicate that the animals werehandled by eagle owl. All humeri from cluster2 and 83% from cluster 1 are fragmented, and thebones exhibit a very typical breakage pattern(Figure 8). The proximal part of the humerus isusually severed from the rest of the bone, and insome specimens where this is not the case, a holeis seen in the shaft just below the articular end.Rather than tearing the prey asunder, eagle owltypically cut off the wings, whereby the proximalpart of the humerus stays attached to the scapulaand the scapular part of the coracoid (Bochenski,1960; Bochenski et al., 1993; Laroulandie, 2000,2002). Laroulandie (2002) mentioned, besidesthe humerus, also the femur and tibiotarsus aselements exhibiting heavy fragmentation in herstudy on the damage to modern pigeon bones(Columba sp.) found in pellets of eagle owl.Although prey size influences the patterns of birdbone fragmentation (Bezzel et al., 1976;Bochenski et al., 1993), the size differencebetween pigeon and chukar is considered hereto be negligible, and indeed the archaeologicalsamples and the recent study show the samepatterns of handling. According to Laroulandie(2002) the frequent fragmentation of the femurand tibiotarsus (and that of the humerus) isrelated to the fact that these bones bear a lot ofmeat and that they therefore have to be reducedin size before digesting them. This reduction insize of the meat-bearing bones can be illustratedby the special breakage pattern that was observedin the femur and tibiotarsus of the Sagalassosmaterial (Figures 9 and 10).
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa
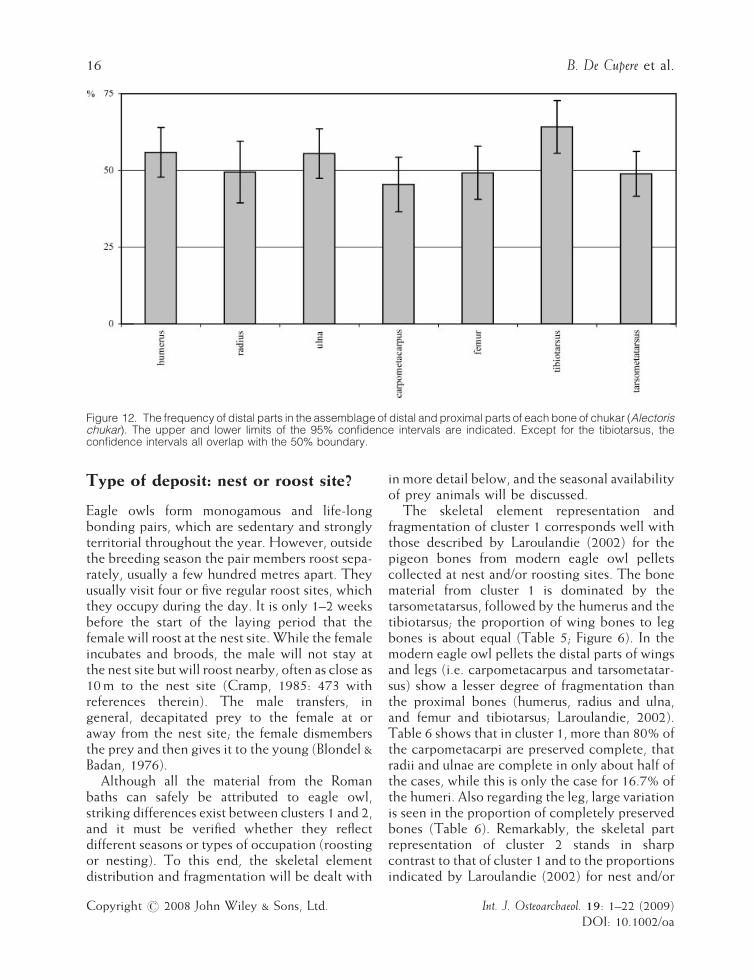
Figure 12. The frequency of distal parts in the assemblage of distal and proximal parts of each bone of chukar (Alectorischukar). The upper and lower limits of the 95% confidence intervals are indicated. Except for the tibiotarsus, theconfidence intervals all overlap with the 50% boundary.
16 B. De Cupere et al.
Type of deposit: nest or roost site?
Eagle owls form monogamous and life-longbonding pairs, which are sedentary and stronglyterritorial throughout the year. However, outsidethe breeding season the pair members roost sepa-rately, usually a few hundred metres apart. Theyusually visit four or five regular roost sites, whichthey occupy during the day. It is only 1–2 weeksbefore the start of the laying period that thefemale will roost at the nest site. While the femaleincubates and broods, the male will not stay atthe nest site but will roost nearby, often as close as10m to the nest site (Cramp, 1985: 473 withreferences therein). The male transfers, ingeneral, decapitated prey to the female at oraway from the nest site; the female dismembersthe prey and then gives it to the young (Blondel &Badan, 1976).Although all the material from the Roman
baths can safely be attributed to eagle owl,striking differences exist between clusters 1 and 2,and it must be verified whether they reflectdifferent seasons or types of occupation (roostingor nesting). To this end, the skeletal elementdistribution and fragmentation will be dealt with
Copyright # 2008 John Wiley & Sons, Ltd.
in more detail below, and the seasonal availabilityof prey animals will be discussed.
The skeletal element representation andfragmentation of cluster 1 corresponds well withthose described by Laroulandie (2002) for thepigeon bones from modern eagle owl pelletscollected at nest and/or roosting sites. The bonematerial from cluster 1 is dominated by thetarsometatarsus, followed by the humerus and thetibiotarsus; the proportion of wing bones to legbones is about equal (Table 5; Figure 6). In themodern eagle owl pellets the distal parts of wingsand legs (i.e. carpometacarpus and tarsometatar-sus) show a lesser degree of fragmentation thanthe proximal bones (humerus, radius and ulna,and femur and tibiotarsus; Laroulandie, 2002).Table 6 shows that in cluster 1, more than 80% ofthe carpometacarpi are preserved complete, thatradii and ulnae are complete in only about half ofthe cases, while this is only the case for 16.7% ofthe humeri. Also regarding the leg, large variationis seen in the proportion of completely preservedbones (Table 6). Remarkably, the skeletal partrepresentation of cluster 2 stands in sharpcontrast to that of cluster 1 and to the proportionsindicated by Laroulandie (2002) for nest and/or
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Eagle Owl Pellets: Nest and Roost Sites 17
roost sites: in cluster 2 the humerus is the bestrepresented element, and a prevalence of wingbones is noted (Table 5; Figure 6).Bochenski et al. (1993) showed that material
collected at recent nest sites differs significantlyfrom that of roost sites.While the tarsometatarsusis the best preserved element at nest sites, thehighest numbers in the case of roost sites arereached by the humerus. Although calculatedsomewhat differently (as an MNI on the basis ofeach skeletal element), Bochenski et al. (1993) alsopointed out that, with the exception of thetibiotarsus and tarsometatarsus, all long bones arebetter or similarly represented at the roostingsites than at the nest sites. This dichotomy is alsoobserved when the survival rates in clusters 1 and2 from Sagalassos are compared (Figure 6;Table 5). Therefore, it is assumed that cluster 1represents the pellet material of a nest site, whilecluster 2 corresponds to the remains of pelletsnear a roost site.Some of the bird species listed in Table 3 occur
only seasonally in the southwestern part ofTurkey and may therefore give information onthe period of the year for which the nest and roostsites in the Roman baths were occupied. Thepresence of common teal (Anas crecca) in Turkeyis, with the exception of Central and EastAnatolia, restricted to winter. During this seasonlarge numbers of teals are seen on the lakes ofsouthern Turkey (Kasparek, 1992: 34). The ruff(Philomachus pugnax) and the corn crake (Crex crex)pass through Anatolia in spring and autumn,during migration (Heinzel et al., 1972: 132, 114;Kasparek, 1992: 63). Some other birds, such asthe little bittern (Ixobrychus minutus), commonquail (Coturnix coturnix) and European roller(Coracias garrulus), breed in Anatolia but areabsent in winter (Heinzel et al., 1972). All otherbirds identified occur in Anatolia throughoutthe year (cf. Heinzel et al., 1972). Amongthe mammals, few seasonality indicators arepresent. The hedgehog, a hibernating species,was most probably captured outside the winterperiod. The juvenile hares are not goodseasonality indicators because in this speciesseveral litters a year can occur, from January toSeptember (Lange et al., 1986). However, verysmall specimens are unlikely to be found duringwinter.
Copyright # 2008 John Wiley & Sons, Ltd.
Both clusters 1 and 2 thus include species thatlive in the area all year round, as well as taxa thatare typical of either summer, spring or autumn, orwinter. The only striking difference between theclusters is the abundance of the common teal, atypical winter visitor, in cluster 2. In thatassemblage the species represents more than halfof the Anatinae (NISP¼ 34 out of 62; MNI¼ 5out of 8). In cluster 1, Anatinae are less numerousin general, and, in addition, only one bone out ofeight is from common teal (MNI¼ 1 out of 4).This means that cluster 2 more clearly illustratesthat there was also occupation during winter,outside the reproduction season. This observa-tion supports the idea that the bones of cluster 2are the remains of pellets near a roost site ratherthan those from a nest site.As mentioned above, male eagle owl roost
within the vicinity of the nest site during thebreeding season. As cluster 2 is located at adistance of about 25m from cluster 1, this mayrepresent such a place, although it is possible thatthe roost site was also used outside the breedingperiod by one of the pair members. Whether thenest and roost site were occupied by exactly thesame couple, is hard to prove, but cannot be ruledout (see below).
The eagle owl and the Roman baths
Radiocarbon dating of the bone remains of cluster1 has indicated that eagle owl lived in the Romanbaths during the second half of the 6th century tothe first quarter of the 7th century AD (Table 2).Although, by that time, the human population ofthe city was already considerably reduced due tothe continuous warfare in the Byzantine empire, aseries of natural disasters and a general economicdecline (Waelkens et al., 2006), large areas of thetown were still inhabited. For example, bothporticos of the Lower Agora, at the western sideof the Roman baths, remained in use until theirfinal destruction in the middle of the 7th centuryAD (Putzeys, 2007: 215). Also the macellum, to thenorth of the Roman baths, and the urban mansion(Domestic Area 1) to the northeast, wereoccupied well into the 7th century AD (Waelkenset al., 2006: 232). The presence of eagle owl nearhuman occupation seems surprising, especially
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

18 B. De Cupere et al.
since they nowadays only nest or roost in regionsof sparse human settlement or in topographicallyinaccessible terrain (Cramp, 1985). Suitablenesting places are avoided when roads are at adistance of less than 400m (Bergerhausen et al.,1989). However, there are some rare observationsof eagle owl breeding very close to humanhabitation, such as in an empty house in thecentre of a village (Cugnasse, 1983; Doucet,1989). During the nesting period couples of eagleowl can be so discrete that they will not be heardor seen at all (Doucet, 1989), which may explainhow they could co-exist with humans atSagalassos in the period under consideration.On the other hand, it is not excluded that peopletolerated the animal’s presence because of itssymbolic status or its predation on vermin. It isalso possible that the eagle owl’s behaviour wasdifferent compared with today, and that theanimal had adopted a less shy, even commensalattitude towards humans. Such behaviouraldifferences have been noted on the basis ofarchaeological material for other species (O’Con-nor, 1993; Mulkeen & O’Connor, 1997).Eagle owls were living in the Roman baths
when the western part of the building was out ofuse and at least part of its roof had collapsed, thusoffering the birds the possibility to use piers as aroosting or nesting place. Since the eagle owlshows fidelity to its breeding area and uses thesame nest site over many years (Cramp, 1985:473), it is possible that such birds inhabited theRoman baths for a long period of time. A firstargument that supports this scenario is the factthat the bone material from cluster 1 was found intwo superimposed layers of destruction materialwith a different texture. This suggests subsequentphases of deposition of debris that may indicatethat the collapse of the building was a gradualprocess (or at least not a single event). Thecollapse may have taken several years, withincreased deposit of debris after winter when, asin recent times, ruined buildings suffered fromthawing and percolation of water. The archae-ozoological material also yields some indicationsfor an extended period of deposition. As shown inthe paragraphs above, differences show up in thespecies composition when the spatially dividedassemblages of cluster 1 are considered separately(Figure 4). It is remarkable that the material
Copyright # 2008 John Wiley & Sons, Ltd.
collected in each of the two sectors (2420–2405and 2415–2405) differs significantly since thedelimitation between these sectors is arbitrary.The place where the pellets were deposited, andlater disintegrated, must have depended on theposition and the slope of the pile of destructionmaterial at the foot of the columns. Pellets may,after they fell to the ground, have tumbled in aparticular direction and have accumulated fartheraway. If the collapse in this part of the Romanbaths was a gradual process, pellets may at acertain moment have concentrated in one area,while in a following period they may havetumbled in another direction. The gradualaccumulation of bone from pellets in varyingdirections, combined with seasonal or annualvariation in prey availability and prey choice, mayhave provoked the observed differences inspecies composition between the two sectors.Indeed, the prey of the eagle owl is known toinclude a large variety of mammals and birds, butwhen these large owls are forced by environ-mental conditions, they can easily adapt to asmaller number of prey species. Following theprinciples of optimal foraging, eagle owls will alsochange their diet composition as a reactionto changes in the abundance of prey species. As aconsequence, within a single population, thepercentages of a prey species can vary consider-ably from year to year and from territory toterritory (Cramp, 1985).
The length of occupation represented by thematerial from cluster 1 can be roughly estimatedby taking into account the number of consumedprey animals and the daily dietary needs of afemale eagle owl and its young. In Table 8 theaverage value of body mass is listed that isretained per individual of each of the major preyspecies. Table 9 calculates the weight that isprovided in cluster 1 by the various prey species,taking into account their MNI’s and their averagebody mass. An average of 400 g of food is taken asthe daily need for an adult eagle owl, althoughdata in the literature on the food consumption ofcaptive and wild eagle owl include both lower andhigher values (Cramp, 1985). It is supposed herethat during the period the female is taking care ofits two or three young, double the amount wouldbe sufficient. The female occupies the nest site1–2 weeks prior to the actual egg laying and then
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Table 8. Average total weight in grams retained for the major prey species, with indication of the variation mentioned inthe literature
Average weight (g) Variation Reference
Fulica atra 800 <: 580–1000 (745) (spring) Il’icev & Flint (1989: 340)780–1040 (930) (summer)
780–1100 (965) (autumn & winter),: 540–870 (660) (spring)630–930 (760) (summer)
510–980 (780) (autumn & winter)Alectoris chukar 500 <: 450–700 Il’icev & Flint (1989: 57)
,: 360–550Corvus frugilegus 450 <: 433 (trapped)–545 (shot)� Patterson et al. (1988)
,: 379 (trapped) –475 (shot)�
Erinaceus concolor 800 <: 400 (Apr)–1100 (Sept) Ozen (2006),: 795 (June)
Lepus europaeus 1700 800–2400 (1 to 3 months) Broekhuizen & Maaskamp (1979)Sciurus anomalus 340 <: 350 Yigit et al. (2006)
<: 250–401 (336) Albayrak & Arslan (2006),: 274–410 (349)
Allactaga euphratica 90 92 (A. e. kivanci holotype) Colak & Yigit (1998)Spalax leucodon 300 306 Yigit et al. (2006)
� Variation of mean weight values of trapped and shot rooks during different seasons in subsequent years.
Eagle Owl Pellets: Nest and Roost Sites 19
she broods, beginning with the laying of the firstegg, during 34–36 days (Cramp, 1985). Theyoung stay on the nest for 55 to 65 days (Blondel& Badan, 1976). Therefore, the amount of foodneeded from the very beginning (i.e. the femalefrequenting the nest site before egg-laying) to themoment that the family leaves the nest wouldbe about 72 kg (50 � 0.4 kg for the female plus65 � 0.8 kg for female and young).The total weight provided by the prey animals
considered from cluster 1 is about 164 kg, whichwould be sufficient to feed the female and itsyoung during at least two breeding seasons. Thecalculation above has to be considered aminimum estimate since, for obvious reasons
Table 9. Estimated weight in grams contributed by themajor prey species in cluster 1
MNI Weightper individual
Total weight
Fulica atra 7 800 5600Alectoris chukar 54 500 27,000Corvus frugilegus 10 450 4500Erinaceus concolor 18 800 14,400Lepus europaeus 61 1700 103,700Sciurus anomalus 8 340 2720Allactaga euphratica 43 90 3870Spalax leucodon 6 300 1800
Copyright # 2008 John Wiley & Sons, Ltd.
(taphonomic loss, sampling techniques), theactual MNI must have been higher. Moreover,not all of the small rodent taxa were taken intoaccount in the calculations above. The data fromthe pellets hence indicate that the Roman bathswere occupied by eagle owl for an extendedperiod of time, showing again that the collapse ofthe western part of the building was a gradualprocess.
Conclusion
Concentrations of animal remains found in thedestruction debris filling the frigidarium 1 of theRoman bath complex consist almost exclusivelyof wild birds and mammals, among which chukar,hare, jerboa and hedgehog are the bestrepresented species. The species compositionand the common presence of digestion traces andtheir nature indicate that both clusters are theremains of pellets from a large owl. Consideringthe owl species that occur in the region, the eagleowl is the only plausible taphonomic agent. Thisis supported by the strong under-representationof cranial elements, typical breakage patterns andthe skeletal element representation. The twoclusters are revealed to be the remains of pellets
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

20 B. De Cupere et al.
near a nest site (cluster 1) and a nearby roost site(cluster 2), a distinction that could be made onthe basis of the best represented element and thesurvival rates of the bones. The presence ofremains of common teal, a winter visitor in theregion, in cluster 2 and the near absence of suchwinter elements in cluster 1 supports theinterpretation as a roost and a nest site. Thespecies composition of cluster 1 shows differ-ences in its horizontal distribution which can belinked to the occupation of the nest duringsubsequent seasons or years during the secondhalf of the 6th to the beginning of the 7th centuryAD. The occupation by eagle owl pre-dates thefinal human abandonment of the town, andillustrates how this large predator was surprisinglyable to live and breed in the close vicinity of man.
Acknowledgements
This paper presents research results of the Inter-university Attraction Poles Programme, BelgianScience Policy. SEM photographs were made byJ. Cillis (Royal Belgian Institute of NaturalScience).
References
Albayrak I, Arslan A. 2006. Contribution to the tax-onomical and biological characteristics of Sciurusanomalus in Turkey (Mammalia: Rodentia). TurkishJournal of Zoology 30: 111–116.
Alivizatos H, Goutner V, Zogaris S. 2005. Contri-bution to the study of the diet of four owl species(Aves, Strigiformes) from mainland and island areasof Greece. Belgian Journal of Zoology 135: 109–118.
Andrews P. 1990. Owls, Caves and Fossils. NaturalHistory Museum Publications: London.
Behrensmeyer AK. 1978. Taphonomic and ecologicinformation from bone weathering. Paleobiology 4:150–162.
Bergerhausen W, Radler K, Willems H. 1989. Besie-dlungspraferenzen des Uhus (Bubo bubo L.) in derEifel. Charadrius 25: 157–178.
Bezzel E, Obst J, Wickl KH. 1976. Zur Ernahrung undNahrungswahl des Uhus (Bubo bubo). Journal ofOrnithology 117: 210–238.
Blondel J, Badan O. 1976. La biologie du Hibougrand-duc en Provence. Nos Oiseaux 33: 189–219.
Copyright # 2008 John Wiley & Sons, Ltd.
Bochenski Z. 1960. The diet of the eagle owl Bubo bubo(L.) in the Pieniny Mts. Acta Zoologica Cracoviensia 5:311–333.
Bochenski ZM. 2005. Owl, diurnal raptors andhumans: signatures on avian bones. In Biosphere toLithosphere. New Studies in Vertebrate Taphonomy,O’Connor T (ed.). Proceedings of the 9th Con-ference of the International Council of Archaeo-zoology, Durham, August 2002. Oxbow Books:Oxford; 31–45.
Bochenski ZM, Tomek T, Boev Z, Mitev I. 1993.Patterns of bird bone fragmentation in pellets ofthe TawnyOwl (Strix aluco) and the Eagle Owl (Bubobubo) and their taphonomic implications. Acta Zool-ogica Cracoviensia 36: 313–328.
Bochenski ZM, Tomek T. 1994. Patterns of bird bonefragmentation in pellets of the long-eared owl Asiootus and its taphonomic implications. Acta ZoologicaCracoviensia 37: 177–190.
Bochenski ZM, Tomek T. 1997. Preservation of birdbones: erosion versus digestion by owls. InternationalJournal of Osteoarchaeology 7: 372–387.
Bochenski ZM, Korovi VA, Nekrasov AE, Tomek T.1997. Fragmentation of bird bones in food remainsof Imperial Eagles (Aquila heliaca). International Journalof Osteoarchaeology 7: 165–171.
Bochenski ZM, Huhtala K, Jussila P, Pulliainen E,Tornberg R, Tunkkari PS. 1998. Damage to birdbones in pellets of Gyrfalcon Falco rusticolus. Journal ofArchaeological Science 25: 425–433.
Bochenski ZM, Huhtala K, Sulkava S, Tornberg R.1999. Fragmentation and preservation of bird bonesin food remains of the Golden Eagle Aquila rusticolus.Archaeofauna 8: 31–39.
Bochenski ZM, Nekrasov AE. 2001. The taphonomy ofSub-Atlantic bird remains from Bazhukovo III, Uralmountains, Russia.Acta Zoologica Cracoviensia44: 93–106.
Bochenski ZM, Tornberg T. 2003. Fragmentation andpreservation of bird bones in uneaten food remainsof the Gyrfalcon Falco rusticolus. Journal of Archaeolo-gical Science 30: 1665–1671.
Broekhuizen S, Maaskamp F. 1979. Age determinationin the European hare (Lepus europaeus Pallas) in theNetherlands. Zeitschrift fur Saugetierkunde 44: 162–175.
Bronk Ramsey C. 1995. Radiocarbon calibration andanalysis of stratigraphy: the OxCal program. Radio-carbon 37: 425–430.
Bronk Ramsey C. 2001. Development of the radio-carbon program OxCal. Radiocarbon 43: 355–363.
Cochard D. 2004. Influence de l’age des proies sur lescaracteristiques des accumulations de leporides pro-duites par le hibou grand-duc. In Petits animaux etsocietes humaines. Du complement alimentaire aux ressources
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

Eagle Owl Pellets: Nest and Roost Sites 21
utilitaires, Brugal JP, Desse J (eds). XXIVe rencontresinternationales d’archeologie et d’histoire d’Antibes.Editions APDCA: Antibes; 313–316.
Cramp S. 1985. The Birds of the Western Paleartic, Vol. IV.Oxford University Press: Oxford.
Cramp S, Simmons KEL. 1980. The Birds of theWestern Paleartic, Vol. II. Oxford University Press:Oxford.
Cugnasse J-M. 1983. Contribution a l’etude du Hibougrand-duc, Bubo bubo, dans le sud du Massif Central.Nos Oiseaux 37: 117–128.
Colak E, Yigit N. 1998. A new subspecies of jerboafrom Turkey, Allactaga euphratica kivanci subsp. n.Turkish Journal of Zoology 22: 93–98.
De Cupere B. 2001. Animals at Ancient Sagalassos. Evidenceof the Faunal Remains, Studies in Eastern Mediterra-nean Archaeology 4. Brepols: Turnhout.
Dodson P, Wexlar D. 1979. Taphonomic investi-gations of owl pellets. Paleobiology 5: 275–284.
Donazar JA, Ceballos O. 1989. Selective predation byeagle owls Bubo bubo on rabbits Oryctolagus cuniculus:age and sex preferences. Ornis Scandinavica 20:117–122.
Doucet J. 1989. Reapparition de la nidification duHibou grand-duc (Bubo bubo) en Wallonie. Sa rein-troduction en Europe occidentale. Aves 26:137–258.
Gautier A. 1987. Taphonomic groups: how and why?ArcheoZoologia 1: 47–52.
Heinzel H, Fitter R, Parslow J. 1972. Oiseaux d’Europe,d’Afrique et du Moyen-Orient. Delachaux et Niestle:Neuchatel.
Il’icev VD, Flint VE. 1989. Handbuch der Vogel derSowjetunion. Aula Verlag: Wiesbaden.
Kasparek M. 1992. Die Vogel der Turkei. Max KasparekVerlag: Heidelberg.
Kirwan GM, Martins RP, Eken G, Davidson P. 1998.A checklist of the birds of Turkey. SandgrouseSuppl. 1: 1–32.
Lange R, van Winden A, Twisk P, de Laender J, SpeerC. 1986. Zoogdieren van de Benelux. Herkenning en onder-zoek. ERLA: Amsterdam.
Laroulandie V. 2000. Taphonomie et archeozoologie desoiseaux en grotte: applications aux sites paleolithiques duBois-Rgot (Vienne), de Combe Sauniere (Dordogne) et dela Vache (Ariege). PhD thesis, Universite Sciences etTechnologies, Bordeaux.
Laroulandie V. 2002. Damage to pigeon long bones inpellets of the Eagle Owl Bubo bubo and food remainsof Peregrine Falcon Falco peregrinus: zooarchaeologi-cal implications. In Proceedings of the 4th meeting of theICAZ bird working group, Bochenski ZM, BochenskiZ, Stewart JR (eds). Acta Zoologica Cracoviensia 45(special issue): 331–339.
Copyright # 2008 John Wiley & Sons, Ltd.
Laudet F, Selva N. 2004. Small mammal taphonomy ofraven (Corvus corax) pellets: archaeological perspect-ives. In Petits animaux et societes humaines. Du complementalimentaire aux ressources utilitaires, Brugal JP, Desse J(eds). XXIVe rencontres internationales d’archeo-logie et d’histoire d’Antibes. Editions APDCA: Anti-bes; 317–320.
Laudet F, Selva N. 2005. Raven as small mammal boneaccumulators: first taphonomic study on mammalremains in raven pellets. Palaeogeography, Palaeoclima-tology, Palaeoecology 226: 272–286.
Mayhew DF. 1977. Avian predators as accumulators offossil mammal material. Boreas 6: 25–31.
Mulkeen S, O’Connor TP. 1997. Raptors in towns:towards an ecological model. International Journal ofOsteoarchaeology 7: 440–449.
O’Connor TP. 1993. Birds and the scavenger niche.Archaeofauna 2: 155–162.
Orsini P. 1985. The diet of the eagle owl Bubo bubo inProvence, southern France. Alauda 53: 11–28.
Ozen AS. 2006. Some biological, ecological andbehavioural features of Erinaceus concolor Martin,1838 (Mammalia: Insectivora) in Turkey. G.U. Jour-nal of Science 19: 91–97.
Papageorgiou NK, Vlachos CG, Bakaloudis DE. 1993.Diet and nest site characteristics of Eagle Owl (Bubobubo) breeding in two different habitats in north-eastern Greece. Avocetta 17: 49–54.
Patterson IJ, Dunnet GM, Goodbody SR. 1988. Bodyweight and juvenile mortality in rooks Corvus frugi-legus. Journal of Animal Ecology 57: 1041–1052.
Putzeys T. 2007. Contextual analysis at Sagalassos. Devel-oping a methodology for classic archaeology. PhD thesis,Katholieke Universiteit Leuven.
Reimer PJ, Baillie MGL, Bard E, Bayliss A, Beck JW,Bertrand C, Blackwell PG, Buck CE, Burr G, CutlerKB, Damon PE, Edwards RL, Fairbanks RG, Frie-drich M, Guilderson TP, Hughen KA, Kromer B,McCormac FG, Manning S, Bronk Ramsey C,Reimer RW, Remmele S, Southon FR, Stuiver M,Talamo S, Taylor FW, van der Plicht J, Weyhen-meyer CE. 2004. INTCAL04 terrestrial radiocarbonage calibration, 0-26 cal kyr BP. Radiocarbon 46:1029–1058.
Rifai LB, Al-Melhim WN, Gharaibeh BM, Amr ZS.2000. The diet of the Desert Eagle Owl, Bubo buboascalaphus, in the Eastern Desert of Jordan. Journal ofArid Environments 44: 369–372.
Robert I, Vigne J-D. 2002. The Bearded Vulture(Gypaetus barbatus) as an accumulator of archaeolo-gical bones. Late Glacial assemblages and present-day reference data in Corsica (Western Mediterra-nean). Journal of Archaeological Science 29: 763–777.
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa

22 B. De Cupere et al.
Thomas DH. 1969. Great basin hunting patterns: aquantitative method for treating faunal remains.American Antiquity 34: 392–401.
Waelkens M, Vanhaverbeke H, Martens F, Talloen P,Poblome J, Kellens N, Putzeys T, Degryse P, VanThuyne T, Van Neer W. 2006. The Late Antique toEarly Byzantine city in Southwest Anatolia. Saga-lassos and its territory: a case study. In Die Stadt in der
Copyright # 2008 John Wiley & Sons, Ltd.
Spatantike – Niedergang oder Wandel? Akten des Interna-tionalen Kolloquiums in Munchen am 30. und 31. Mai 2003,Krause JU, Witschel C (eds). Historia Einzelschriften190: 199–255.
Yigit N, Demirsoy A, Karatas A, Ozkurt S, Colak E.2006. Notes on the mammals found in KazdagıNational Park and its environs. Turkish Journal ofZoology 30: 73–82.
Int. J. Osteoarchaeol. 19: 1–22 (2009)DOI: 10.1002/oa