dyslexia: bridging the gap between hearing and reading
TRANSCRIPT

Current Biology Vol 19 No 6R260
natural route of ploidy reduction aftermating in a host. And if it is, why stop atdiploidy? Thus, sorting out the linkbetween the laboratory-generatedmating and the process occurring in themammalian host promises to continueto provide interesting insights into thebiology of this important humanpathogen.
References1. Tavanti, A., Gow, N.A., Maiden, M.C.,
Odds, F.C., and Shaw, D.J. (2004). Geneticevidence for recombination in Candida albicansbased on haplotype analysis. Fungal Genet.Biol. 41, 553–562.
2. Hull, C.M., and Johnson, A.D. (1999).Identification of a mating type-like locus in theasexual pathogenic yeast Candida albicans.Science 285, 1271–1275.
3. Hull, C.M., Raisner, R.M., and Johnson, A.D.(2000). Evidence for mating of the ‘‘asexual’’yeast Candida albicans in a mammalian host.Science 289, 307–310.
4. Magee, B.B., and Magee, P.T. (2000). Inductionof mating in Candida albicans by construction
of MTLa and MTLalpha strains. Science 289,310–313.
5. Miller, M.G., and Johnson, A.D. (2002). White-opaque switching in Candida albicans iscontrolled by mating-type locus homeodomainproteins and allows efficient mating. Cell 110,293–302.
6. Zordan, R.E., Miller, M.G., Galgoczy, D.J.,Tuch, B.B., and Johnson, A.D. (2007).Interlocking transcriptional feedback loopscontrol white-opaque switching in Candidaalbicans. PLoS Biol. 5, e256.
7. Huang, G., Srikantha, T., Sahni, N., Yi, S., andSoll, D.R. (2009). CO2 regulates white-to-opaque switching in Candida albicans. Curr.Biol. 19, 330–334.
8. Ramirez-Zavala, B., Reuss, O., Park, Y.N.,Ohlsen, K., and Morschhauser, J. (2008).Environmental induction of white-opaqueswitching in Candida albicans. PLoS Pathog.4, e1000089.
9. Dumitru, R., Navarathna, D.H., Semighini, C.P.,Elowsky, C.G., Dumitru, R.V., Dignard, D.,Whiteway, M., Atkin, A.L., and Nickerson, K.W.(2007). In vivo and in vitro anaerobic matingin Candida albicans. Eukaryot. Cell 6,465–472.
10. Klengel, T., Liang, W.J., Chaloupka, J.,Ruoff, C., Schroppel, K., Naglik, J.R.,Eckert, S.E., Mogensen, E.G., Haynes, K.,
Tuite, M.F., et al. (2005). Fungal adenylylcyclase integrates CO2 sensing with cAMPsignaling and virulence. Curr. Biol. 15,2021–2026.
11. Pendrak, M.L., Yan, S.S., and Roberts, D.D.(2004). Hemoglobin regulates expression ofan activator of mating-type locus alphagenes in Candida albicans. Eukaryot. Cell 3,764–775.
12. Odds, F.C. (1987). Candida infections: anoverview. Crit. Rev. Microbiol. 15, 1–5.
13. Forche, A., Alby, K., Schaefer, D.,Johnson, A.D., Berman, J., and Bennett, R.J.(2008). The parasexual cycle in Candidaalbicans provides an alternative pathway tomeiosis for the formation of recombinantstrains. PLoS Biol. 6, e110.
Genetics Group, Biotechnology ResearchInstitute, National Research Council ofCanada, 6100 Royalmount Ave., Montreal,Quebec, Canada H4P 2R2 and Department ofBiology, McGill University, Montreal, Quebec,Canada H3A 1B1.E-mail: [email protected]
DOI: 10.1016/j.cub.2009.01.043
l
changes (and model) are centered ondeficits in phonological encoding anddecoding and the networks thatsupport these processes [10–13]. Butdespite the presence of a stronglinkage between disruptedphonological abilities and poor readingskills in dyslexia, there has remaineda fundamental gap in ourunderstanding of how problems inencoding speech sounds ultimatelytranslate into reading difficulties. A keystep in this process must be the rapidand accurate matching of thecomponent speech sounds(phonemes) with their appropriatewritten representations (graphemes).Despite the intuitive nature of thismultisensory transformation process,there is little empirical evidence thatrelates across these domains andspecifically bridges speech processingand reading.
Advances in non-invasiveneuroimaging methods, particularlyfunctional magnetic resonanceimaging (fMRI), have made thisproblem more tractable by allowinga view into the neural correlates ofreading and phonological processes[1,14–17]. With fMRI, changes in theblood oxygenation level dependent(BOLD) signal, an indirect measure ofneural activity, can be measured whilethe participant is presented with certainstimuli and/or engaged in a specifictask. Differences in the BOLD signalcan then be compared between two
Dyslexia: Bridging the Gap betweenHearing and Reading
Recent work with dyslexic subjects provides the first empirical evidence linkingchanges in the brain networks subserving phonological processing to deficitsin the matching of speech sounds to their appropriate visual representations.
Mark T. Wallace
Although most children rapidly developa strong facility to read the printed andwritten word, a surprisingly largenumber fail to acquire good readingskills, even after intensive instruction.When these reading difficulties areseen in the presence of normal orabove-normal intelligence, and whenthere are measurable deficits inphonological processing — the abilityto store, retrieve and manipulatespeech sounds — the child (or adult) istypically diagnosed with dyslexia,a term first coined in the late 19th
century. The most common form ofdyslexia is seen in a developmentalcontext as children fail to meet certainbenchmark measures of ‘normal’reading ability. Although there appearto be cultural and orthography-relateddifferences in its prevalence, someestimates suggest that the incidence ofdevelopmental dyslexia may be as highas 10% in the general population [1].Not surprisingly given this highprevalence, the monetary and societalimpacts of reading disabilities are
staggering. The work by Blau et al. [2]reported in this issue of CurrentBiology provides important newinsights into the neural bases ofdevelopmental dyslexia, by showingchanges in brain activation patterns indyslexic readers that are associatedwith the matching of speech soundswith their appropriate visualrepresentations (letters). Suchletter–speech matching must be bothrapid and accurate for the emergenceof fluent reading abilities.
Although its diagnosis is stillconsidered to be controversial in somedomains, there is a growing consensusthat dyslexia has a neurobiologicalbasis, with strong evidence that there isa genetic component to the disability[3]. Numerous theories abound as tothe physiological processes and neurasystems that are affected in dyslexia,with several of the more prominentmodels focusing on alterations inrapid auditory processing [4–6],disturbances in the magnocellularvisual pathway [7,8], and cerebellardysfunction [9]. As alluded to above,however, the best-established
DispatchR261
Auditory ‘t’visual ‘g’
(incongruent)
Dyslexic reader
Auditory ‘t’
Audiovisual ‘t’(congruent)
Typical reader
STG/STS
Auditory ‘t’visual ‘g’
(incongruent)
Auditory ‘t’
Audiovisual ‘t’(congruent)
Current Biology
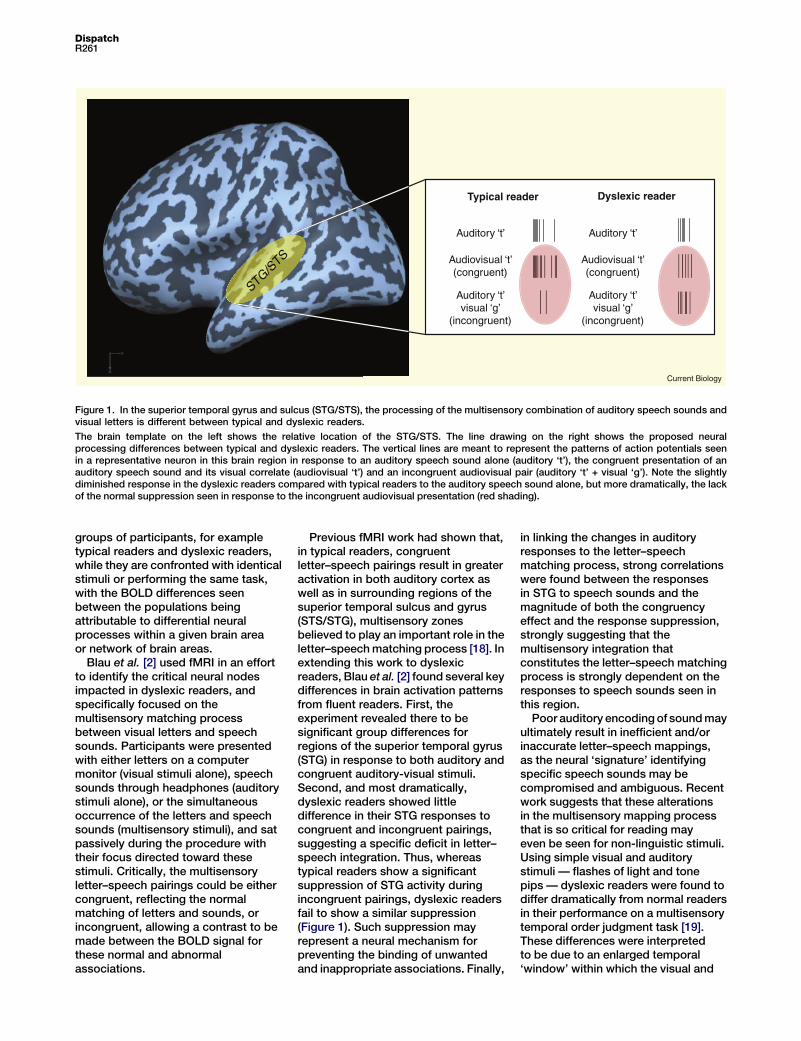
Figure 1. In the superior temporal gyrus and sulcus (STG/STS), the processing of the multisensory combination of auditory speech sounds andvisual letters is different between typical and dyslexic readers.
The brain template on the left shows the relative location of the STG/STS. The line drawing on the right shows the proposed neuralprocessing differences between typical and dyslexic readers. The vertical lines are meant to represent the patterns of action potentials seenin a representative neuron in this brain region in response to an auditory speech sound alone (auditory ‘t’), the congruent presentation of anauditory speech sound and its visual correlate (audiovisual ‘t’) and an incongruent audiovisual pair (auditory ‘t’ + visual ‘g’). Note the slightlydiminished response in the dyslexic readers compared with typical readers to the auditory speech sound alone, but more dramatically, the lackof the normal suppression seen in response to the incongruent audiovisual presentation (red shading).
in linking the changes in auditoryresponses to the letter–speechmatching process, strong correlationswere found between the responsesin STG to speech sounds and themagnitude of both the congruencyeffect and the response suppression,strongly suggesting that themultisensory integration thatconstitutes the letter–speech matchingprocess is strongly dependent on theresponses to speech sounds seen inthis region.
Poor auditory encoding of sound mayultimately result in inefficient and/orinaccurate letter–speech mappings,as the neural ‘signature’ identifyingspecific speech sounds may becompromised and ambiguous. Recentwork suggests that these alterationsin the multisensory mapping processthat is so critical for reading mayeven be seen for non-linguistic stimuli.Using simple visual and auditorystimuli — flashes of light and tonepips — dyslexic readers were found todiffer dramatically from normal readersin their performance on a multisensorytemporal order judgment task [19].These differences were interpretedto be due to an enlarged temporal‘window’ within which the visual and
Previous fMRI work had shown that,in typical readers, congruentletter–speech pairings result in greateractivation in both auditory cortex aswell as in surrounding regions of thesuperior temporal sulcus and gyrus(STS/STG), multisensory zonesbelieved to play an important role in theletter–speech matching process [18]. Inextending this work to dyslexicreaders, Blau et al. [2] found several keydifferences in brain activation patternsfrom fluent readers. First, theexperiment revealed there to besignificant group differences forregions of the superior temporal gyrus(STG) in response to both auditory andcongruent auditory-visual stimuli.Second, and most dramatically,dyslexic readers showed littledifference in their STG responses tocongruent and incongruent pairings,suggesting a specific deficit in letter–speech integration. Thus, whereastypical readers show a significantsuppression of STG activity duringincongruent pairings, dyslexic readersfail to show a similar suppression(Figure 1). Such suppression mayrepresent a neural mechanism forpreventing the binding of unwantedand inappropriate associations. Finally,
groups of participants, for exampletypical readers and dyslexic readers,while they are confronted with identicalstimuli or performing the same task,with the BOLD differences seenbetween the populations beingattributable to differential neuralprocesses within a given brain areaor network of brain areas.
Blau et al. [2] used fMRI in an effortto identify the critical neural nodesimpacted in dyslexic readers, andspecifically focused on themultisensory matching processbetween visual letters and speechsounds. Participants were presentedwith either letters on a computermonitor (visual stimuli alone), speechsounds through headphones (auditorystimuli alone), or the simultaneousoccurrence of the letters and speechsounds (multisensory stimuli), and satpassively during the procedure withtheir focus directed toward thesestimuli. Critically, the multisensoryletter–speech pairings could be eithercongruent, reflecting the normalmatching of letters and sounds, orincongruent, allowing a contrast to bemade between the BOLD signal forthese normal and abnormalassociations.

Current Biology Vol 19 No 6R262
auditory stimuli were bound intoa unitary construct.
Taken together, these recent studiesare providing important insights intothe neurobiological bases of specificreading disabilities, and are convergingon a multisensory model that betterlinks auditory processing deficits withthe visual functions that mediatereading. This knowledge providesa better conceptual framework forunderstanding reading disabilities,and holds great promise for thedevelopment of more effectiveremediation strategies for thetreatment of those suffering from theseoften debilitating disabilities.
References1. Habib, M. (2000). The neurological basis of
developmental dyslexia: an overview andworking hypothesis. Brain 123, 2373–2399.
2. Blau, V., van Atteveldt, N., Ekkebus, M.,Goebel, R., and Blomert, L. (2009). Reducedneural integration of letters and speech soundslinks phonological and reading deficits in adultdyslexia. Curr. Biol. 19, 503–508.
3. Paracchini, S., Scerri, T., and Monaco, A.P.(2007). The genetic lexicon of dyslexia. Annu.Rev. Genomics Hum. Genet. 8, 57–79.
4. Tallal, P. (1980). Auditory temporal perception,phonics, and reading disabilities in children.Brain Lang. 9, 182–198.
5. Tallal, P., Miller, S., and Fitch, R.H. (1993).Neurobiological basis of speech: a case for thepreeminence of temporal processing. Ann. NYAcad. Sci. 682, 27–47.
6. Temple, E., Poldrack, R.A., Protopapas, A.,Nagarajan, S., Salz, T., Tallal, P.,Merzenich, M.M., and Gabrieli, J.D. (2000).Disruption of the neural response to rapidacoustic stimuli in dyslexia: evidence fromfunctional MRI. Proc. Natl. Acad. Sci. USA 97,13907–13912.
7. Livingstone, M.S., Rosen, G.D., Drislane, F.W.,and Galaburda, A.M. (1991). Physiological andanatomical evidence for a magnocellular defectin developmental dyslexia. Proc. Natl. Acad.Sci. USA 88, 7943–7947.
8. Stein, J., and Walsh, V. (1997). To see but notto read; the magnocellular theory of dyslexia.Trends Neurosci. 20, 147–152.
9. Nicolson, R.I., Fawcett, A.J., and Dean, P.(2001). Developmental dyslexia: the cerebellardeficit hypothesis. Trends Neurosci. 24,508–511.
10. Shaywitz, S.E., and Shaywitz, B.A. (2005).Dyslexia (specific reading disability). Biol.Psychiatry 57, 1301–1309.
11. Shaywitz, S.E., and Shaywitz, B.A. (2003).Dyslexia (specific reading disability). Pediatr.Rev. 24, 147–153.
12. Ramus, F. (2003). Developmental dyslexia:specific phonological deficit or generalsensorimotor dysfunction? Curr. Opin.Neurobiol. 13, 212–218.
13. Ramus, F., Rosen, S., Dakin, S.C., Day, B.L.,Castellote, J.M., White, S., and Frith, U. (2003).Theories of developmental dyslexia: insightsfrom a multiple case study of dyslexic adults.Brain 126, 841–865.
14. Pekkola, J., Laasonen, M., Ojanen, V., Autti, T.,Jaaskelainen, I.P., Kujala, T., and Sams, M.
(2006). Perception of matching and conflictingaudiovisual speech in dyslexic and fluentreaders: an fMRI study at 3T. Neuroimage 29,797–807.
15. Eden, G.F., Jones, K.M., Cappell, K., Gareau, L.,Wood, F.B., Zeffiro, T.A., Dietz, N.A.,Agnew, J.A., and Flowers, D.L. (2004).Neural changes following remediation inadult developmental dyslexia. Neuron 44,411–422.
16. Shaywitz, B.A., Lyon, G.R., and Shaywitz, S.E.(2006). The role of functional magneticresonance imaging in understandingreading and dyslexia. Dev. Neuropsychol. 30,613–632.
17. Temple, E., Deutsch, G.K., Poldrack, R.A.,Miller, S.L., Tallal, P., Merzenich, M.M., andGabrieli, J.D. (2003). Neural deficits in childrenwith dyslexia ameliorated by behavioralremediation: evidence from functional MRI.Proc. Natl. Acad. Sci. USA 100, 2860–2865.
18. van Atteveldt, N., Formisano, E., Goebel, R.,and Blomert, L. (2004). Integration of letters andspeech sounds in the human brain. Neuron 43,271–282.
19. Hairston, W.D., Burdette, J.H., Flowers, D.L.,Wood, F.B., and Wallace, M.T. (2005). Alteredtemporal profile of visual-auditory multisensoryinteractions in dyslexia. Exp. Brain Res. 166,474–480.
Vanderbilt Brain Institute, VanderbiltUniversity, 465 21st Avenue South,Nashville, TN 37232, USA.E-mail: [email protected]
DOI: 10.1016/j.cub.2009.01.025
Innate Immunity: Cytoplasmic DNASensing by the AIM2 Inflammasome
Cytoplasmic double-stranded DNA triggers cell death and secretion of the pro-inflammatory cytokine IL-1b in macrophages. Recent reports now describe themechanism underlying this observation. Upon sensing of DNA, the HIN-200family member AIM2 triggers the assembly of the inflammasome, culminatingin caspase-1 activation, IL-1b maturation and pyroptotic cell death.
analysis of TLR9-deficient micerevealed the existence of alternativeDNA-sensing pathways, includingthose regulating IL-1b maturationand IRF-3-dependent type I IFNexpression [2,5]. Furthermore, whileTLR9 recognizes foreign DNA inendo-lysosomal compartments [6],DNA delivered to the cytoplasmtriggers a TLR-independent innateimmune response that includes thesecretion of IFNb and IL-1b frommacrophages [2,5,7]. By comparison,IFNb is not induced in response to theTLR9 ligand, CpG DNA, inmacrophages [8].
The hunt for the cytoplasmic DNAsensor(s) then began in earnest. Thefirst DNA sensor identified was DAI(also known as DLM-1/ZBP1),which was shown to trigger a robustTBK-1/IRF-3-dependent type I IFNresponse. Subsequent reportssuggested, however, thatDAI-independent mechanisms alsooperate [9,10]. A recent report fromour group implicated the‘inflammasome’ pathway in thesensing of cytoplasmic DNA, leadingto caspase-1 activation and IL-1b
maturation [5].
Many years of intense researchhave only begun to explain theimmunostimulatory effects of DNA. Thefirst clue came with the discovery of themembrane-bound Toll-like receptor(TLR) family of PRRs. Upon recognitionof PAMPs, TLR pathways triggerprofound changes in gene regulation,including the induction of manypro-inflammatorycytokinesdownstreamof NFkB, mitogen-activated protein(MAP) kinases, and IFN regulatoryfactors (IRFs). Our understanding ofthe immunostimulatory properties offoreign DNA was advancedsignificantly by the identification ofTLR9, which specifically recognizesunmethylated CpG sequences that arepresent in prokaryotic DNA butsuppressed in mammalian DNA [4]. The
Kate Schroder1, Daniel A. Muruve2,and Jurg Tschopp1,*
The innate immune system recognizespathogens through an extensive arrayof pattern recognition receptors (PRRs)that detect invariant microbial motifscalled pathogen-associated molecularpatterns (PAMPs). DNA is one suchPAMP that is highly immunostimulatorywhen internalized or delivered into thecytoplasm of cells [1,2]. Prokaryotic,viral and non-microbial DNA triggersa number of innate immune pathwaysthat result in the secretion of pro-inflammatory cytokines, in particularinterleukin-1b (IL-1b), the induction ofanti-viral type I interferons (IFNa/b) [2]and cell death in susceptible cells, suchas macrophages [3].