draft - · pdf filedraft 2 abstract: fluorapatite-solubilizing bacteria were isolated from...
TRANSCRIPT

Draft
Phosphate-solubilizing bacteria isolated from
ectomycorrhizal mycelium of Picea glauca are highly efficient at fluorapatite weathering
Journal: Botany
Manuscript ID cjb-2016-0089.R1
Manuscript Type: Article
Date Submitted by the Author: 08-Jun-2016
Complete List of Authors: Fontaine, Laurent; Université Laval, Département des sciences du bois et de la forêt Thiffault, Nelson; Ministère des Forêts, de la Faune et des Parcs, Direction de la recherche forestière Paré, David; Natural Resources Canada, Canadian Forest Service Fortin, J.-André; Université Laval, Département des sciences du bois et de la forêt Piché, Yves; Université Laval, Département des sciences du bois et de la forêt
Keyword: Ectomycorrhizal fungi, Mineral weathering, Phosphate solubilizing bacteria, Apatite, Burkholderia
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
1
Phosphate-solubilizing bacteria isolated from ectomycorrhizal mycelium of Picea
glauca are highly efficient at fluorapatite weathering
Laurent Fontainea,*, Nelson Thiffaultb, David Paréc, J.-André Fortina, and Yves Pichéa
a Département des sciences du bois et de la forêt, Faculté de foresterie, de géographie et
de géomatique, Université Laval, 2405 de la Terrasse, Québec, QC G1V 0A6, Canada
b Direction de la recherche forestière, Ministère des Forêts, de la Faune et des Parcs, 2700
Einstein, Québec, QC G1P 3W8, Canada
c Natural Ressources Canada, Canadian Forest Service, Laurentian Forestry Centre, 1055
du PEPS, P.O. Box 10380 Stn. Sainte-Foy, Québec, QC G1V 4C7, Canada
* Corresponding author: Laurent Fontaine (email: [email protected])
Page 1 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
2
Abstract: Fluorapatite-solubilizing bacteria were isolated from the hyphosphere of the
ectomycorrhizal (ECM) fungus Wilcoxina sp., a dominant species in the Picea glauca
rhizosphere. Hundreds of strains isolated from the ascomycete Wilcoxina sp. could
dissolve tricalcium phosphate, while only 27 of them could produce clarification halos on
fluorapatite-amended solid medium. Most of the fluorapatite-solubilizing strains
belonged to the Burkholderia genus. Scanning electron microscopy observations have
shown that these efficient phosphate-solubilizing bacteria (PSB) were able to completely
solubilize fluorapatite crystals within 22 h. The efficient PSB Burkholderia sp. strain 205
and Curtobacterium sp. strain 168 were tested for their ability to associate with a
genetically distant fungal host while fulfilling their phosphate solubilizing function.
Burkholderia sp. strain 205 successfully associated with the Basidiomycete Laccaria
bicolor when hydroxyapatite was the only phosphorus source available to the fungus,
while there was no bacterial development when L. bicolor could access soluble
phosphorus as well. Optical microscopic observation of L. bicolor associated with
Burkholderia sp. revealed extensive colonization of fungal hyphae by the bacterium.
These results suggest an important role of bacteria-ECM fungi associations in white
spruce phosphate nutrition.
Key words: Ectomycorrhizal fungi, apatite, mineral weathering, phosphate solubilizing
bacteria, hyphosphere, Burkholderia.
Page 2 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
3
Résumé: Des bactéries solubilisatrices de fluorapatite ont été isolées à partir de
l’hyphosphère du champignon ectomycorhizien (ECM) Wilcoxina sp., une espèce
dominant la rhizosphère de Picea glauca. Des centaines de souches isolées de
l’ascomycète Wilcoxina sp. ont dissout le phosphate tricalcique, alors que seulement 27
d’entre elles ont clarifié le milieu amendé en fluorapatite. La majorité des souches
dissolvant la fluorapatite appartiennent au genre Burkholderia. Des observations réalisées
en microscopie électronique à balayage ont montré que ces bactéries solubilisatrices de
phosphates (BSP) efficaces pouvaient dissoudre complètement des cristaux de
fluorapatite en 22 h. Les souches de BSP efficaces Burkholderia sp. 205 et
Curtobacterium sp. 168 ont été testées quant à leur capacité de s’associer avec un hôte
génétiquement distant tout en remplissant leur fonction de BSP. La souche 205 de
Burkholderia sp. s’est associée avec succès au basidiomycète Laccaria bicolor lorsque
l’hydroxyapatite était la seule source de phosphore accessible au champignon, alors
qu’une source de phosphore soluble inhibait la croissance bactérienne. Des observations
en microscopie à fond clair de L. bicolor en association avec Burkholderia sp. ont révélé
une importante colonisation bactérienne des hyphes. Ces résultats suggèrent une
importance des associations bactéries-champignons ECM dans la nutrition phosphatée de
l’épinette blanche.
Mots-clés: Champignons ectomycorhiziens, apatite, alteration minérale, bactérie
solubilisatrice de phosphate, hyphosphère, Burkholderia.
Page 3 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
4
Introduction
In boreal forests, soil properties shape the composition of plant communities,
which in turn enable the development of diverse communities of ectomycorrhizal fungi
(ECM) and their associated bacteria. The availability of nutrients in the soil as well as the
composition of soil minerals influence the development of ECM fungi (Rosling et al.
2004) and bacterial diversity (Uroz et al. 2012). For example, carbon allocation to ECM
mycelium formed by Pinus sylvestris L. and Paxillus involutus S.L. (Batsch) Fries was
significantly higher in soils amended with calcium phosphate compared with a control
treatment amended with quartz (Smits et al. 2012). Moreover, P. involutus releases large
amounts of organic acids when host growth is limited by a phosphorus deficiency (van
Schöll et al. 2006). Exudation of these organic acids by the ECM mycelium should
provide associated bacteria with a significant source of carbon (Olsson and Wallander
1998). This probably explains why mineral weathering bacteria are more abundant in the
ECM hyphosphere than in bulk soil (e.g. Scleroderma citrinum; Uroz et al. 2007).
Therefore, these bacteria closely associated with ECM hyphae are more effective in
mineral weathering than those isolated from the rest of the soil (Calvaruso et al. 2007).
This suggests a tripartite relationship, in which the ECM fungus receives photosynthates
that are then released as organic acids, which are substrates that can be assimilated by
certain soil bacteria. In exchange, these bacteria would mobilize ions from minerals
(Sokolova 2011), which are immediately absorbed by the ECM fungus and then
transmitted to the host (Calvaruso et al. 2010). According to this model, soil fertility in
boreal forest would be influenced by the content and weatherability of soil minerals as
well as by the mineral weathering capacities of fungal and bacterial communities
Page 4 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
5
associated with forest stands. The existence and role of this tripartite relationship in the
nutrition of boreal forest tree species has been observed for Pinus resinosa Sol. ex. Aiton
in microcosm experiments (Balogh-Brunstad et al. 2008).
Among the nutrients essential for tree development, the abundance and forms of
phosphate minerals found in the soil define the fertility of a site (Hahm et al. 2014). The
crystalline properties of phosphate minerals determine their reactivity (Elliott 1994) and
thus their availability as phosphorus sources for soil microorganisms. In turn, apatite
crystalline structures are determined by calcium, phosphate and fluoride substitutions
(McDonnell 1973). An amorphous mineral such as tricalcium phosphate (TCP) is easily
weathered by microorganisms, while crystalline phosphate minerals are much more
resistant to corrosion (Bashan et al. 2012). Igneous rocks contain apatites that can be
classified into the three subgroups of hydroxyapatite, chlorapatite and fluorapatite, the
last one being the most recalcitrant to chemical weathering (Dorozhkin 2011). Access to
the nutrients contained in soil minerals by plants and their associated microorganisms is
of particular interest in boreal and mountainous regions because of the relatively early
stage of soil development and, hence, the availability of unweathered minerals due to the
recent glacial retreat (Milner et al. 2007). We chose to study the microbial flora of white
spruce (Picea glauca (Moench) Voss), a species typical of the boreal zone, known for its
sensitivity to phosphorus availability (Quesnel and Côté 2009).
Based on our assumption of the tripartite relationship between the host, ECM
fungi and phosphate solubilizing bacteria (PSB), we predicted that the most abundant
ECM fungi in the rhizosphere of white spruce would be associated with potentially
cultivable PSB. Our hypothesis also suggests that PSB receive organic carbon from ECM
Page 5 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
6
mycelium when the only source of phosphorus is insoluble. Finally, it predicts that the in
vitro weathering of fluorapatite by PSB could be observed in less than a day when
crystals are incubated in a bacterial suspension.
Therefore, our overall objective was to verify the role of the bacterial microbiota
associated with ECM fungi in fluorapatite weathering, an important source of phosphorus
in mineral soils of igneous origin. To do so, our specific objectives were to i) isolate and
identify PSB from the mycelium of an ubiquitous ECM fungus associated with P. glauca,
ii) demonstrate in vitro fluorapatite solubilization by PSB and iii) validate in vitro the
close association between PSB and ECM fungi.
Materials and methods
PSB source
Potential fluorapatite-solubilizing bacteria were isolated from the rhizosphere of
four P. glauca seedlings. The host plants were aged between 5 and 15 years and located
in plantations. The collection site is located approximately 75 km north of Québec City
(Canada) (lat. 46°57'55.3'' N; long 71°30 55.9'' W) within the balsam fir (Abies balsamea
(L.) Mill.) – white birch (Betula papyrifera Marsh.) bioclimatic domain described by
Saucier et al. (2009). The seedlings were taken along with a layer of MOR organic
material (approx. 3 cm thick). The B-horizon from the surrounding stands (Humo-Ferric
podzol; Soil Classification Working Group 1998) was collected to serve as the mineral
soil substrate for 8 L transplantation pots.
Page 6 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
7
Maintenance of host plants
After transplantation, seedlings were maintained in a greenhouse for 18 months
(September 2012 – April 2014). Seedlings were submitted to a 16 h photoperiod at
temperatures ranging from 18–23 °C. The plants were watered once a week with tap
water. No weeding was made during this period; thus, herbaceous species (e.g.
Chamaerion angustifolium (L.) Holub, Poa pratensis L., Pilosella aurantiaca (L.)
F.W.Schultz & Schultz-Bip.) proliferated in the pots. After the first trimester in the
greenhouse, the seedlings became dormant and were lifted from that state by being kept
for 4 months at 4 °C in complete darkness.
Characterization of PSB hosting ECM morphotypes
ECM morphotypes were collected in order to use them as a PSB inoculum source
to be grown on solid and liquid media. Mycorrhizal morphotypes were collected in June
2013 for liquid cultures and in February 2014 for solid cultures. While growing in the
greenhouse, the seedlings produced large amounts of long roots running along the
perimeter of the pots, which made it possible to harvest short roots without disturbing the
substrate. The most abundant ECM were identified visually, then collected and screened
under a dissecting microscope to ensure only one species was present in the sample.
Molecular identification of the main ectomycorrhizal morphotype
Five ectomycorrhizas of the dominant morphotype were randomly selected from
three seedlings to proceed with the ECM fungal DNA extraction. They were ground with
Page 7 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
8
mortar and pestle in 100 µl of TTE buffer (0.002 M EDTA, 0.02 M Tris, 0.0008 M Triton
X-100) and heated at 95 °C for 10 min. The resulting product was centrifuged and the
supernatant retained as total DNA.
A polymerase chain reaction (PCR) (Saiki et al. 1988) was performed on the
internal transcribed spacers (ITS) of the ribosomal genes in a reaction volume of 25 µl
containing 0.2 mM dNTP mix, 0.5 µM primer ITS-1F and ITS-4, 2.5 µl PCR buffer
(Qiagen, Hilden, Germany), 1.5 mM MgCl2, BSA 0.2 mg·ml-1, 3 U Taq (Qiagen) and
2 µl of total DNA. Amplification was performed on a PTC-225 Thermal Cycler (MJ
Research, Waltham, Massachusetts) under the following conditions: 94 °C for 4 min,
followed by 34 cycles at 94 °C, 55 °C and 72 °C for 1 min, and finally at 72 °C for 10
min.
Amplification success was confirmed by migration of the PCR product on agarose
gel before proceeding to sequencing. The sequencing was performed on a 16 capillary
genetic analyzer (3130XL model, Applied Biosystems, Thermo Fisher Scientific Inc.,
Burlington, ON).
Isolation of PSB on solid selective medium
We isolated BSP closely associated with the identified ECM morphotype. In order
to obtain a diversity of strains, we used the spread plate method on agar media containing
only insoluble phosphorus sources. A subsample of ectomycorrhizas subjected to
molecular analysis was selected to form the bacterial inoculum. Ectomycorrhizas were
washed vigorously under running tap water to remove particles and bacteria not firmly
bound to the fungal sheath. The washing was performed on a 1 mm mesh sieve. The
Page 8 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
9
freshly cleaned ectomycorrhizal samples were then transferred to a sterile aqueous
solution of NaCl 8.5 g·L-1 (9x their fresh mass). In this saline solution, ectomycorrhizas
were finely ground with a sterile set of mortar and pestle. The resulting suspension was
considered as a 10-1 dilution. Serial dilutions were carried out in the same saline solution
to obtain a bacterial suspension factor of 10-4. That 10-4 dilution was used as inoculum for
spread plating on selective medium at 300 µl per plate. The medium was double layered
to facilitate the observation of clarification halos. The bottom layer, based on the NBRIP
medium (Nautiyal 1999), contained 10 g glucose, 5 g MgCl2·6H2O, 250 mg
MgSO4·7H2O, 200 mg KCl, 200 mg KH2PO4, 100 mg (NH4)2SO4, 200 mg yeast extract
(Difco Laboratories, Detroit, Michigan), 50 mg cycloheximide (Sigma Chemical Co., St.
Louis, Missouri) and 10 g agarose (Amresco, Solon, Ohio) per litre. The upper layer
contained, per litre of medium, 2.5 g Ca3PO4 (TCP) (Fisher Science Education, Hannover
Park, Illinois) and 10 g of agarose. The media were poured into 80 mm Petri dishes at a
rate of 20 ml for the bottom layer and 5 ml for the upper layer. The initial isolation of
phosphate-solubilizing strains was carried out on a medium containing TCP as an
insoluble phosphorus source. The stability of the weathering phenotype was verified with
three subcultures on TCP. Strains exhibiting stable expression of the desired phenotype
were subcultured on an amended layer containing 2.5 g·L-1 hydroxyapatite (Sigma-
Aldrich, St. Louis, Missouri). The strains having successfully dissolved hydroxyapatite
were finally tested on a medium amended with 2.5 g·L-1 carbonated fluorapatite from the
Grenville geological province (Arianne Phosphate, Saguenay, Québec) containing 1%
fluorine.
Page 9 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
10
PSB isolation by enrichment in selective liquid medium
PSB were obtained by enrichment in selective liquid medium using the same
inoculum as for isolation on solid media. The enrichment was initiated with 200 µl of 10-2
dilution of ground ectomycorrhizas. Again, the NBRIP selective medium (Nautiyal 1999)
was used, but this time with fluorapatite ore rather than TCP as an amendment in order to
immediately select bacterial strains able to grow on igneous apatite. The bacteria were
grown in 125 ml flasks containing 50 ml of medium, sealed with foam plugs, and
incubated at 26 °C with agitation at 100 rpm. The medium was sterilized in an autoclave
with a liquid cycle for 20 min. A benomyl solution was added to each experimental unit
after autoclaving to obtain a medium concentration of 3 mg·L-1. After adding the
inoculum, the flasks were incubated for one week and subcultured in fresh media.
Transplanting was done with an inoculation loop so that subculture would favour the
most abundant strains in the selection of colony forming units inoculating the fresh
medium. In turn, this should promote the selection of the most competitive fluorapatite
weathering bacteria. After seven subcultures, the bacterial communities were transferred
to solid medium to allow the isolation of strains. The strains unable to weather TCP on
solid medium were rejected.
Ultrastructural observations of fluorapatite weathering by an efficient PSB
Fluorapatite weathering by an efficient bacterial strain was observed using
scanning electron microscopy (SEM). The elemental composition of the fluorapatite ore
was determined by X-ray photoelectron spectroscopy. The bacterial strain PSB 205,
Page 10 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
11
selected because of its effectiveness in fluorapatite dissolution, was propagated in flasks
containing the liquid medium Tryptic Soy Broth (Difco Laboratories, Detroit, Michigan)
maintained at 26 °C under agitation at 180 rpm. The microbial cultures were centrifuged,
the supernatant removed, followed by washing of the bacteria by resuspension in a
solution of NaCl 0.15 M. This suspension was centrifuged again to remove the
supernatant. The bacterial pellet was resuspended in phosphorus-free NBRIP medium to
produce a bacterial concentration of 1×106 CFU per ml. Two fluorapatite ores were used,
with fluorine concentrations of 1% (Arianne Phosphate, Saguenay, Québec) and 4%
(Mine Arnaud, Sept-Îles, Québec), respectively. The fluorapatite ore was washed with
distilled water, open air dried, and then autoclaved for 20 min. Ferromagnetic particles
were removed from the ore using a magnet. Carbon tape adhered to a stainless steel SEM
stub was used as a specimen holder for the ores (Goldstein 2003). Fluorapatite was
deposited on the carbon tape and loose particles were removed with an air jet.
Two methods were used to incubate the bacterial strain PSB 205 in the presence
of fluorapatite. The first was limited to covering the ore attached to the SEM stub with a
drop of bacterial suspension. The second method consisted in placing the fluorapatite-
bearing stub in a cavity of a sterile multiwell plate (24 Well Multiwell, Becton Dickinson,
Franklin Lakes, NJ) and then filling the well with the bacterial suspension to cover the
fluorapatite and incubating under agitation at 60 rpm. The first and second methods were
used after 6 and 22 h of incubation, respectively. Controls involving incubation in
bacteria-free growth medium were used to confirm that mineral weathering did not occur
spontaneously.
Page 11 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
12
Observations were made using a JEOL JSM-7600F SEM model equipped with an
energy-dispersive X-ray spectrometer (EDS) with a resolution of 1 nm at 15 kV and
1.4 nm at 1 kV. The conventional secondary electron detector (LEI) was used. No
conductive coatings (Au, C, Pt) were applied during sample preparation. Initial
observations were conducted on virgin fluorapatite crystals. The most conspicuous
crystals were identified in order to be easily located after incubation with PSB strain 205.
Micrographs of the whole crystals and their edges were taken. After incubation,
fluorapatite was washed under a stream of distilled water for 1 min. The sample was
dried under vacuum before being inserted back into the SEM column. The crystals
observed in their virgin state were found, photographed again and subjected to qualitative
observations. The mineralogical composition was also determined by X-ray
photoelectron spectroscopy.
Resynthesis of close associations between three PSB strains and an ECM fungus
The ECM fungus Laccaria bicolor (Maire) P.D.Orton strain B3-25#2 was
selected for its symbiotic properties, especially its inability to solubilize apatites. As for
bacteria, PSB strains (168, 205 and 260) were chosen for their ability to efficiently
weather fluorapatite. The reconstitution of associations between PSB strains and L.
bicolor was carried out in a bicompartmented Petri dish culture system. Initially, the
phosphate-free proximal compartment was inoculated with L. bicolor to promote
exploration of the distal compartment containing a phosphorus source when the proximal
compartment contained all essential nutrients, with the exception of phosphorus (Fig. 1).
Meeting of the ECM fungus with each PSB strain occurred in the distal compartment,
Page 12 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
13
comprising one of three sources of phosphorus: i) soluble phosphorus (soluble P), ii)
hydroxyapatite (HA), or iii) soluble phosphorus + hydroxyapatite (soluble P + HA). Each
treatment was inoculated or not with three bacterial strains able to solubilize fluorapatite.
The experiment also included two controls with bacteria growing without the fungus on a
medium of soluble phosphorus or hydroxyapatite. Each treatment was repeated three
times.
The proximal compartment contained a modified MNM medium (Marx 1969)
without phosphorus. This medium consisted of 0.15 g MgSO4 · 7H2O, 0.025 g NaCl, 5 g
glucose, 0.26 g KCl, 0.05 g CaCl2, 1 ml trace elements solution, 0.5 ml FeEDTA , 100 ug
thiamine HCl, 0.340 g l-alanine, and 10 g agar per litre of medium. Distal compartments
contained 10 g agar, 0.5 g KH2PO4 (soluble P), 2.5 g hydroxyapatite (HA) or 0.5 g
KH2PO4 (soluble P), and 2.5 g hydroxyapatite (HA) per litre. The media were sterilized
in an autoclave with a liquid cycle for 20 min.
The proximal compartments were each inoculated with two 5 mm diameter fungal
plugs taken at the edge of a 1-month-old colony of L. bicolor grown on GYME medium.
The plugs were formed using a cork borer and placed at the centre of the proximal
compartment. The morphologically distinct PSB strains 168, 205 and 260 were deposited
in the distal compartment under a laminar flow hood. The bacteria were deposited once L.
bicolor colonies had grown 1.5 cm past the septum. The bacteria were then deposited in
the proximal compartment using a micropipette as three 2 µl drops of an aqueous
suspension containing 1×106 CFU per ml and set 1.5 cm apart, a few mm from the edge
of the fungal colonies.
Page 13 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
14
The diameters of the bacterial colonies were used as a proxy for growth and
analyzed according to a complete block experimental design using an analysis of variance
(ANOVA). In the presence of a significant F value, the means were compared using a
priori orthogonal contrasts to discern meaningful differences in growth for the bacterial
strains 168, 205 and 260 in terms of i) the soluble or insoluble nature of the phosphorus
source available to L. bicolor in the distal compartment, and ii) the presence or absence
of a soluble phosphorus source when an insoluble phosphorus source was always
available to L. bicolor in the distal compartment.
The data were examined to verify normality and homogeneity of variance with
standard graphical methods. All analyses were performed using the GLM procedure of
SAS v.9.32 software (SAS Institute Inc., Cary, North Carolina). Effects were considered
significant at the α = 0.05 threshold.
Molecular identification of PSB strains
PSB strains selected by culture on solid medium and in liquid medium were
identified based on 16S rRNA genes. Molecular identification was performed on 27
strains of PSB having produced clarification halos on fluorapatite ore on solid media and
on the nine strains isolated by enrichment in liquid media displaying a stable weathering
phenotype. The strains were grown on Tryptic Soy Broth (Difco Laboratories, Detroit,
Michigan) for a period of 7 days at 26 °C, with agitation at 180 rpm. We collected and
centrifuged 100 µl of culture medium containing the bacteria. The supernatant was
removed and the pellet resuspended in 100 µl of TTE buffer. This suspension was then
heated at 95 °C for 10 min. The resulting product was centrifuged and the supernatant
Page 14 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
15
retained as total DNA. A polymerase chain reaction (PCR) was performed in the same
way as for the ectomycorrhizal morphotype, except for the primers (358E and 907R,
Weidner et al. 1996) and the use of three U Taq instead of one. After validation of the
amplification success, the sequences were edited using the Bioedit software v.7.1.3 (Ibis
Biosciences, Carlsbad, California). Identity of the taxa was determined by comparing the
sequences obtained with those deposited in GenBank
(http://www.ncbi.nlm.nih.gov/genbank/). A phylogenic tree was created with the Mega5
software (Tamura et al. 2011) using the UPGMA method with 100 bootstrap replicates.
Results
Isolation of PSB closely associated with the dominant ECM morphotype in the P. glauca
rhizosphere
The most abundant ECM morphotype found in association with P. glauca, used as
our source of PSB, matched the fungus Wilcoxina sp. with 95-98% similarity to ITS of
the ribosomal genes with sequences returned by GenBank.
The spread plating of crushed Wilcoxina sp. mycorrhizae produced 455 colonies
displaying clarification halos on TCP-amended solid medium. Only 140 strains were
retained based on their capacity to produce clarification halos after being subcultured
three times (Table 1). Of these 140 phenotypically stable strains, 98 clarified the
hydroxyapatite amended medium. Finally, 27 strains produced clarification halos on the
fluorapatite ore amendment. Clarification halos typically became visible on TCP within
24-48 h, while those on fluorapatite required a minimum of 1 week to be discernible.
Page 15 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
16
Enrichment in liquid medium produced 39 strains that were later grown on solid
media. Of these 39 strains, nine showed a stable weathering phenotype on three
subcultures on TCP-amended medium. The weathering phenotype remained after the
strains were cultured on rich media.
Identification of PSB taxa
The strains isolated on solid media and whose DNA was successfully amplified
mostly belong to the genus Burkholderia (Fig. 2). Burkholderia spp. are divided into two
taxa with a support of 1 (Fig. 2). The other bacterial strains obtained on solid media
belong to the genus Curtobacterium, while those obtained by enrichment in liquid media
were identified as Leifsonia sp. The phylogenic tree is rooted by Pseudozyma aphidis, an
environmental yeast isolated by enrichment in liquid media.
Ultrastructural observations of fluorapatite weathering by Burkholderia sp. 205
Fluorapatite weathering by the PSB Burkholderia sp. 205 was successfully
observed with both incubation methods. With the drop method, the edges of fluorapatite
crystals containing 1% fluorine showed signs of corrosion after only 6 h of incubation
(Fig. 3b), compared to intact fluorapatite (Fig. 3a). Complete dissolution of fluorapatite
crystals containing 1 % fluorine was observed after 22 hours of incubation with the
immersion method (Figs. 3c and 3d). There were no signs of weathering for the controls
incubated in bacteria-free growth medium. The surface study of a fluorapatite crystal
shows an elemental composition consisting mainly of phosphorus, calcium and oxygen
Page 16 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
17
while also showing traces of fluorine, silica and iron1. These fluorapatite crystals are
commingled with dust and layers of complex oxides resistant to weathering. Furthermore,
the crystals of fluorapatite containing 4% fluorine (Figs. 3e and 3f) are more resistant to
weathering and have different patterns of etching than those containing 1% fluorine (Fig.
3e).
Close association between PSB and ECM fungi
The association between L. bicolor and PSB strains in the distal compartment of
bicompartmented plates demonstrated that the growth of Burkholderia sp. 205 depended
on the nature of the phosphorus source made available to the fungus. Indeed, when L.
bicolor could acquire phosphorus on its own, (Fig. 4a), the growth of Burkholderia sp.
205 could not be observed on the hyphae of the ECM fungus located in the distal portion
of the plate (Fig. 4a inset). In contrast, when L. bicolor was deprived of a soluble source
of phosphorus (Fig. 4b), Burkholderia sp. 205 displayed growth on the hyphae of the
symbiotic fungus (Fig. 4b inset). Thus, the presence of hydroxyapatite in the absence of
soluble phosphorus produced a significantly increased growth for Burkholderia sp. 205,
for the unknown strain 260 and, to a lesser, non-significant level, for Curtobacterium sp.
168 (Fig.5; Table 2).
Discussion
The PSB we isolated from ECM fungi were able to rapidly dissolve fluorapatite.
The enrichment in liquid medium yielded the most competitive strains for growth in
defined medium whose sole source of phosphorus was fluorapatite ore. In contrast, 1 See supplementary figure 1.
Page 17 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
18
enrichment on solid medium better expressed the diversity of species able to grow in
these conditions. However, the low diversity of bacterial strains obtained either by
isolation on solid media or by enrichment in liquid medium does not match the
complexity of bacterial communities typically found in soils (Sharma et al. 2005). The
PSB community potentially sensitive to culture-dependent isolation is also limited
because it comes from the immediate environment of the hyphae of one ECM fungus,
Wilcoxina sp. (Scheublin et al. 2010). The use of a selective medium and the removal of
bacterial strains that did not produce clarification halos also limited the isolation of
strains able to solubilize phosphates. We further limited our estimation of the
taxonomical diversity of isolated PSB strains by restricting 16S rRNA-based
identification to the strains that were successful in dissolving fluorapatite under in vitro
conditions only.
Our results confirm the presence of PSB in the hyphosphere of Wilcoxina sp., a
dominant and abundant ectomycorrhizal fungus in P. glauca stands (Sokolski et al.,
unpublished results). Obviously, the role of phosphate solubilization in boreal forests is
not restricted to the three genera isolated in this study (Uroz and Frey-Klett 2011).
Moreover, the strains found to be the most effective for in vitro phosphate solubilization
might display a lessened weathering activity when grown in hyphospheric communities
maintained in microcosms (Calvaruso et al. 2006). A semi-quantitative estimation of
biomass under natural conditions of the Burkholderia sp. isolated in this study could be
established by quantifying the fraction of bacterial DNA they represent within
hyphospheric communities.
Page 18 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
19
We observed that the bacterial strains capable of dissolving fluorapatite were
fewer than those that were capable of dissolving TCP. This variation in the PSB
community appears to be modulated by the lower reactivity of apatites characterized by
the presence of a crystal lattice that is more resistant to bacterial chemical attack
compared with precipitated amorphous phosphates such as TCP (Zhao et al. 2011). Out
of 203 TCP-solubilizing strains, 98 dissolved hydroxyapatite and 27 succeeded in
dissolving fluorapatite ore. Fluorapatite is the most resistant type of apatite to weathering
(Bashan et al. 2012).
Ultrastructural observations of fluroapatite weathering by Burkholderia sp. 205
support that biotic factors contributing to mineral weathering could achieve impressive
rates; Burkholderia sp. 205 growing in a static drop of NBRIP medium succeeded in
corroding the edges of fluorapatite crystals within six hours. When this same fluorapatite
was incubated under agitation in a Burkholderia sp. 205 suspension of NBRIP medium,
less than 1 day was sufficient to observe the dissolution of whole crystals. However,
complete dissolution in 22 h was observed on fluorapatite with a fluorine content of 1%
per mass. When the experiment was repeated for 23 h with fluorapatite containing 4%
fluorine, we only observed partial dissolution. The fluorine content therefore appears to
be a determining factor in the availability of phosphorus from fluorapatite crystals when
the time scale remains substantially the same. The first signs of weathering were
observed on crystal edges. This is due to the energy of lattice edges being greater than
that of flat surfaces; edges are thus the most reactive regions of particles (G. L'Espérance
(personal communication, 2015)).
Page 19 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
20
Whereas some ECM fungi were ineffective at mineral weathering when grown
axenically in vitro, they are still abundant in nature (Calvaruso et al. 2010). This apparent
contradiction could be explained by an ecological role that is possibly largely limited to
the absorption and transport of nutrients. Those ECM fungi receive carbon from host
photosynthesis, which they exude in the immediate vicinity of minerals containing useful
elements for plant growth (Paris et al. 1996). This release supports the activity of bacteria
possessing mineral weathering capabilities. In return, the fungus absorbs minerals and
transfers them to its host. The model ECM fungus L. bicolor alone is ineffective in
apatite weathering but, in nature, this ECM fungus is associated with effective apatite
weathering bacteria (Frey-Klett 2005; Uroz et al. 2007) and has a tendency to increase
plants’ Mg and K budgets in microcosm experiments when co-inoculated with efficient
biotite weathering bacteria (Koele et al. 2009). Among the Glomeromycota, several
bacterial species preferentially attach to active fungal hyphae (Toljander et al. 2005;
Taktek et al. 2015). This situation also occurs in Basidiomycetes. For example, the
apatite weathering bacterium Burkholderia terrae migrates on the hyphae of Lyophyllum
sp. and carries other bacterial species (Warmink et al. 2010). This ability to assist the
comigration of strains unable to achieve it on their own is widespread among the genus
Burkholderia (Nazir et al. 2012). In light of our isolation of strains from the hyphosphere
of Wilcoxina sp., the ubiquity of the Burkholderia genus in forest soils (Lepleux et al.
2012) has been confirmed as encompassing the boreal forest.
Our study innovates by demonstrating the non-specificity of the fungal host for
Burkholderia sp. 205, which hails from the hyphosphere of an Ascomycete (Wilcoxina
sp.) and then associates with a Basidiomycete (L. bicolor). The fungal host exuded
Page 20 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
21
substances necessary for the growth of bacteria and their hydroxyapatite solubilizing
activity, providing a source of phosphorus available to L. bicolor. Bacteria developed and
completed their alleged ecological function as they migrated on the hyphae of their
fungal host. This comigration undoubtedly facilitates the colonization of large amounts of
substrates by Burkholderia sp. 205 due to the exploration capacity of the fungus host
which greatly exceeds that of the bacterium.
In conclusion, efficient PSB were isolated from the ECM ascomycete Wilcoxina
sp., hosted by P. glauca, and identified as belonging to the Burkholderia, Curtobacterium
and Leifsonia genera. Our study was essentially observational and qualitative in nature
though. For example, our observations would have been strengthened by assessing the
number of bacteria in the distal portion of the plate by extracting the contents,
homogenizing and preparing dilution series for plate counts, measuring P contents within
the fungal hyphae, the supernatant of the well or the microbial biomass. Nevertheless, our
ultrastructural observations demonstrated rapid in vitro fluorapatite solubilization by
Burkholderia sp. 205, and the close association between PSB and ECM fungi was
validated in monoxenic conditions. Although our results have neither been reproduced
with L. bicolor in symbiosis nor under field conditions, and are thus not to be
generalized, they suggest that the efficiency of ECM symbiosis in acquiring P from soil
minerals relies on hyphospheric PSB.
Acknowledgements
We thank Philippe Plamondon (Centre for Characterization and Microscopy of Materials,
(CM)²), Serge Sokolski and Franck Stefani for the quality of their technical work as well
Page 21 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
22
as Isabelle Lamarre for text editing. We are thankful for the advice provided by Salma
Taktek and Jean-Guy Catford, and grateful for the help of interns Gwenola Plougoulen
and Mathieu Paradis. Special thanks also extended to Armand Séguin, two anonymous
reviewers and the Associate Editor, whose relevant comments served to improve a
preliminary version of this manuscript. This study was funded by the Fonds de recherche
du Québec – Nature et technologies.
Page 22 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
23
References
Bashan, Y., Kamnev, A.A., and de-Bashan, L.E. 2012. Tricalcium phosphate is
inappropriate as a universal selection factor for isolating and testing phosphate-
solubilizing bacteria that enhance plant growth: a proposal for an alternative procedure.
Biol. Fertil. Soils 49 (4): 465-479. doi: 10.1007/s00374-012-0737-7.
Balogh-Brunstad, Z., Keller, C.K., Gill, R.A., Bormann, B.T., and Li, C.Y. 2008. The
effect of bacteria and fungi on chemical weathering and chemical denudation fluxes in
pine growth experiments. Biogeochemistry 88 (2): 153-167. doi: 10.1007/s10533-008-
9202-y.
Calvaruso, C., Turpault, M.P., and Frey-Klett, P. 2006. Root-associated bacteria
contribute to mineral weathering and to mineral nutrition in trees: a budgeting analysis.
Appl. Environ. Microbiol. 72: 1258-1266. doi: 10.1128/AEM.72.2.1258–1266.2006.
Calvaruso, C., Turpault, M.P., Leclerc, E., Frey-Klett P. 2007. Impact of
ectomycorrhizosphere on the functional diversity of soil bacterial and fungal
communities from a forest stand in relation to nutrient mobilization processes. Microb.
Ecol. 54: 567-577. doi: 10.1007/s00248-007-9260-z.
Calvaruso, C., Turpault, M.P., Leclerc, M., Ranger, J., Garbaye, J., Uroz, S., and Frey-
Klett, P. 2010. Influence of forest trees on the distribution of mineral weathering-
Page 23 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
24
associated bacterial communities of the Scleroderma citrinum mycorrhizosphere. Appl.
Environ. Microbiol.76: 4780-4787. doi: 10.1128/AEM.03040-09.
Dorozhkin, S.V. 2011. Calcium orthophosphates. Biomatter 1(2): 121-164. doi:
10.4161/biom.18790.
Elliott, J.C. 1994. Structure and chemistry of the apatites and other calcium
orthophosphates. Stud. Inorg. Chem.: 18: 1-389.
Frey-Klett, P., Chavatte, M., Clausse, M.L., Courrier, S., Le Roux, C., Raaijmakers, J.,
Martinotti, M.G., Pierrat, J.C., and Garbaye, J. 2005. Ectomycorrhizal symbiosis affects
functional diversity of rhizosphere fluorescent pseudomonads. New Phytol. 165: 317–
328. doi: 10.1111/j.1469-8137.2004.01212.x.
Goldstein, J. 2003. Scanning electron microscopy and x-ray microanalysis. Kluwer
Academic/Plenum Publishers, New York.
Hahm, W.J., Riebe, C.S., Lukens, C.E., and Araki,S. 2014. Bedrock composition
regulates mountain ecosystems and landscape evolution. Proc. Natl. Acad. Sci. U.S.A.
111: 3338-3343. doi: 10.1073/pnas.1315667111.
Koele, N., Turpault, M.P., Hildebrand, E.E., Uroz, S., and Frey-Klett, P. 2009.
Interactions between mycorrhizal fungi and mycorrhizosphere bacteria during mineral
Page 24 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
25
weathering: Budget analysis and bacterial quantification. Soil Biol. Biochem. 41: 1935–
1942. doi: 10.1016/j.soilbio.2009.06.017.
Lepleux, C., Turpault, M.P., Oger, P., Frey-Klett, P., and Uroz, S. 2012. Correlation of
the abundance of betaproteobacteria on mineral surfaces with mineral weathering in
forest soils. Appl. Environ. Microbiol. 78: 7114-7119. doi: 10.1128/AEM.00996-12.
Marx, D.H. 1969. Influence of ectotrophic mycorrhizal fungi on resistant of pine roots to
pathogenic infections. I. Antagonism of mycorrhizal fungi to root pathogenic fungi and
soil bacteria. Phytopathology 59 (2): 153.
McDonnell, D. 1973. Apatite – Its crystal chemistry, mineralogy, utilization, and
geologic and biological occurences. Springer-Verlag, Vienna. doi: 10.1007/978-3-7091-
8314-4.
Milner, A., Fastie, C.L., Chapin, F.S., Engström, D.R., and Sharman, L.C. 2007.
Interactions and linkages among ecosystems during landscape volution. Biosci. 57: 237-
247. doi: 10.1641/B570307.
Nautiyal, C.S. 1999. An efficient microbiological growth medium for screening
phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 170: 265-270. doi:
10.1111/j.1574-6968.1999.tb13383.x.
Page 25 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
26
Nazir, R., Zhang, M., de Boer, W., and van Elsas, J.D. 2012. The capacity to comigrate
with Lyophyllum sp. strain Karsten through different soils is spread among several
phylogenetic groups within the genus Burkholderia. Soil Biol. Biochem. 50: 221-233.
doi: 10.1016/j.soilbio.2012.03.015.
Olsson, P.A., and Wallander, H. 1998. Interactions between ectomycorrhizal fungi and
the bacterial community in soils amended with various primary minerals. FEMS
Microbiol. Ecol. 27: 195-205. doi: 10.1111/j.1574-6941.1998.tb00537.x.
Paris, F., Botton, B., and Lapeyrie, F. 1996. In vitro weathering of phlogopite by
ectomycorrhizal fungi. Plant Soil 179: 141-150. doi: 10.1007/BF00011651.
Quesnel, P.O. and Côté, B. 2009. Prevalence of phosphorus, kotassium, and calcium
limitations in white spruce across Canada. J. Plant Nutr. 32: 1290–1305. doi:
10.1080/01904160903006002.
Rosling, A., Lindhal, B.D., Taylor, A., and Finlay, R.D. 2004. Mycelial growth and
substrate acidification of ectomycorrhizal fungi in response to different minerals. FEMS
Microbiol. Ecol. 47: 31-37. doi: 10.1016/S0168-6496(03)00222-8.
Saiki, R.K., Gelfand, D.H., Stoffel, S., Scharf, S.J., Higuchi, R., Horn, G.T., Mullis, K.B.,
and Erlich, H.A. 1998 Primer-directed enzymatic amplification of DNA with a
Page 26 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
27
thermostable DNA polymerase. Science 239 (4839): 487-491. doi:
10.1126/science.2448875.
Saucier, J.P., Robitaille, A., and Grondin, P. 2009. Cadre bioclimatique du Québec. In
Manuel de foresterie, 2e édition. Edited by R. Doucet and M. Côté. Ordre des ingénieurs
forestiers du Québec, Éditions Multimondes, Québec, QC. pp. 186-205.
Scheublin, T.R., Sanders, I.R., Keel, C., and van der Meer, J.R. 2010. Characterisation of
microbial communities colonising the hyphal surfaces of arbuscular mycorrhizal fungi.
The ISME Journal 4: 752-763. doi: 10.1038/ismej.2010.5.
Sharma, R., Ranjan, R., Kapardar, R.K., and Grover, A. 2005. ‘Unculturable’ bacterial
diversity: An untapped resource. Curr. Sci. 89 (1): 72-77.
Smits, M.M., Bonneville, S., Benning, L.G., Banwart, S.A., and Leake, J.R. 2012. Plant-
driven weathering of apatite – the role of an ectomycorrhizal fungus. Geobiology 10:
445-456. doi: 10.1111/j.1472-4669.2012.00331.x.
Sokolova, T.A. 2011. The role of soil biota in the weathering of minerals: A review of
literature. Eurasian Soil Sci. 44 (1): 56-72. doi: 10.1134/S1064229311010121.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. 2011.
MEGA5: Molecular evolutionary genetics analysis using maximum likelihood,
Page 27 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
28
evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28: 2731-
2739. doi: 10.1093/molbev/msr121.
Soil Classification Working Group. 1998. The Canadian System of Soil Classification.
3rd ed. Agriculture and Agri-Food Canada, Ottawa. Publication. No. 1646.
Taktek, S., Trépanier, M., Magallon-Servin, P., St-Arnaud, M., Piché, Y., Fortin, J.A.,
and Antoun H. 2015. Trapping of phosphate solubilizing bacteria on hyphae of the
arbuscular mycorrhizal fungus Rhizophagus irregularis DAOM 197198. Soil Biol.
Biochem. 90: 1-9. doi: 10.1016/j.soilbio.2015.07.016.
Toljander, J.F., Artursson, V., Paul, L.R., Jansson, J.K., and Finlay, R.D. 2005.
Attachment of different soil bacteria to arbuscular mycorrhizal fungal extraradical hyphae
is determined by hyphal vitality and fungal species. FEMS Microbiol. Lett. 254: 34-40.
doi: 10.1111/j.1574-6968.2005.00003.x.
Uroz, S., Calvaruso, C., Turpault, M.P., Pierrat, J.C., Mustin, C., and Frey-Klett, P. 2007.
Effect of the mycorrhizosphere on the genotypic and metabolic diversity of the bacterial
communities involved in mineral weathering in a forest soil. Appl. Environ. Microbiol.
79: 3019–3027. doi: 10.1128/AEM.00121-07.
Uroz, S. and Frey-Klett, P. 2011. Linking diversity to function: highlight on the mineral
weathering bacteria. Cent. Eur. J. Biol. 6: 817-820. doi: 10.2478/s11535-011-0053-5.
Page 28 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
29
Uroz , S., Turpault, M.P., Delaruelle, C., Mareschal, L., Pierrat, J. C., Frey-Klett, P. 2012.
Minerals affect the specific diversity of forest soil bacterial communities. Geomicrobiol.
J. 29: 88-98. doi: 10.1080/01490451.2010.523764.
van Schöll, L., Hoffland, E., and van Breemen, N. 2006. Organic anion exudation by
ectomycorrhizal fungi and Pinus sylvestris in response to nutrient deficiency. New
Phytol. 170: 153–163. doi: 10.1111/j.1469-8137.2006.01649.x.
Warmink, J.A., Nazir, R., Corten, B., and van Elsas, J.D. 2010. Hitchhikers on the fungal
highway: the helper effect for bacterial migration via fungal hyphae. Soil Biol. Biochem.
43: 760-765. doi: 10.1016/j.soilbio.2010.12.009.
Weidner, S., Arnold, W., and Pühler, A. 1996. Diversity of uncultured microorganisms
associated with the seagrass Halophila stipulacea estimated by restriction fragment
length polymorphism analysis of PCR-amplified 16S rRNA genes. Appl. Environ.
Microbiol. 62: 766-771.
Zhao, J., Liu, Y., Sun, W., and Zhang, H. 2011. Amorphous calcium phosphate and its
application in dentistry. Chem. Cent. J. 5: 40. doi: 10.1186/1752-153X-5-40.
Page 29 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
30
Tables
Table 1. Number of bacterial strains grown on amended solid media successful at
dissolving tricalcium phosphate (TCP), hydroxyapatite and fluorapatite.
Amendment Subculture
Nb.
Colonies
TCP - 455
TCP 1 428
TCP 2 203
TCP 3 140
Hydroxyapatite - 98
Fluorapatite (< 45µm) - 27
Page 30 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
31
Table 2. A priori orthogonal contrasts comparing colony diameters of Burkholderia sp.
strain 205, unknown species strain 260, and Curtobacterium sp. strain 168 growing in the
presence of L. bicolor in the distal compartment containing various sources of
phosphorus (soluble, insoluble, or both).
Phosphorus source Strain P
Soluble P vs. Hydroxyapatite
205 < 0.0001
260 < 0.0001
168 0.0691
Soluble P + hydroxyapatite vs.
Hydroxyapatite
205 < 0.0001
260 < 0.0001
168 0.0143
Page 31 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
32
Figure captions
Fig. 1. Experimental set-up of dual compartment culture media driving L. bicolor to
explore both compartments in order to obtain all the essential nutrients for its growth.
Proximal compartment: agar medium without phosphates (P), inoculum L. bicolor
represented by grey circles. Distal compartment: agar with either i) soluble phosphorus
(soluble P), ii) hydroxyapatite (HA), or iii) soluble phosphorus + hydroxyapatite (soluble
P + HA). Bacterial inocula represented by three black dots: strains 168, 205 and 260.
Fig. 2. Phylogenic tree illustrating the relationship between the PSB strains associated
with Wilcoxina sp. that were isolated on selective solid and liquid media and that are
capable of solubilizing fluorapatite.
Fig. 3. SEM images of fluorapatite crystals. (a) Virgin fluorapatite crystal with sharp
edges. (b) Edge of a fluorapatite crystal after 6 h of incubation in the presence of
Burkholderia sp. strain 205. Edges are corroded and the etching pattern does not follow
crystalline axes. (c) and (d) Fluorapatite crystal before (c) and after (d) incubation by
immersion for 22 h in the presence of Burkholderia sp. strain 205. In image d, the
remnants of the particle still present on the carbon tape are complex oxides. (e) and (f)
Fluorapatite crystals after immersion for 23 h in the presence of Burkholderia sp. strain
205. Those crystals were part of an ore containing 4% fluorine. The crystals are partially
weathered, and weathering patterns follow crystalline axes.
Page 32 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
33
Fig. 4. Reconstitution of the association between L. bicolor and Burkholderia sp. strain
205. Arrows indicate where the bacterial inoculum was deposited. (a) Laccaria bicolor in
the presence of Burkholderia sp. strain 205 in the distal compartment with the soluble
P+hydroxyapatite treatment. Inset: the hyphae of L. bicolor devoid of PSB. (b) Laccaria
bicolor in the presence of Burkholderia sp. strain 205 on the distal compartment of the
hydroxyapatite treatment. Inset: the hyphae of L. bicolor covered with PSB.
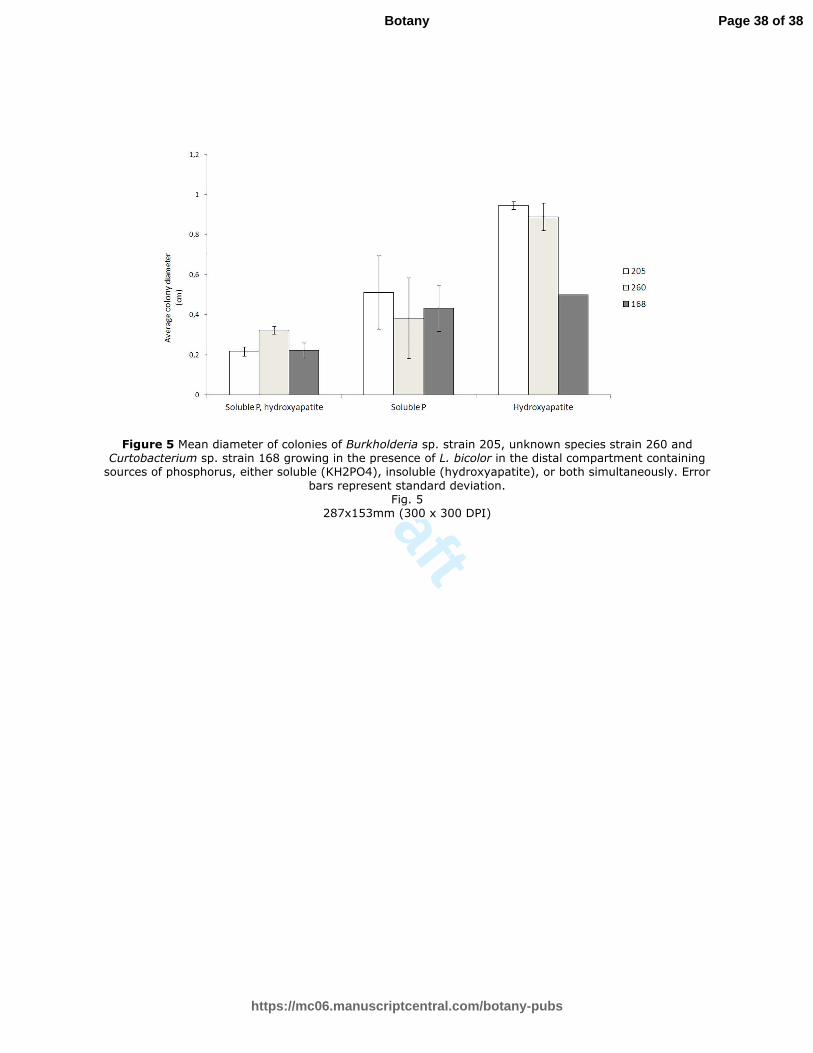
Fig. 5. Mean diameter of colonies of Burkholderia sp. strain 205, unknown species strain
260 and Curtobacterium sp. strain 168 growing in the presence of L. bicolor in the distal
compartment containing sources of phosphorus, either soluble (KH2PO4), insoluble
(hydroxyapatite), or both simultaneously. Error bars represent standard deviation.
Page 33 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Figure 1 Experimental set-up of dual compartment culture media driving L. bicolor to explore both compartments in order to obtain all the essential nutrients for its growth. Proximal compartment: agar
medium without phosphates (P), inoculum L. bicolor represented by grey circles. Distal compartment: agar with either i) soluble phosphorus (soluble P), ii) hydroxyapatite (HA), or iii) soluble phosphorus +
hydroxyapatite (soluble P + HA). Bacterial inocula represented by three black dots: strains 168, 205 and 260. (Fig. 1)
123x129mm (300 x 300 DPI)
Page 34 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Figure 2 Phylogenic tree illustrating the relationship between the PSB strains associated with Wilcoxina sp. that were isolated on selective solid and liquid media and that are capable of solubilizing fluorapatite.
(Fig. 2) 247x158mm (300 x 300 DPI)
Page 35 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Figure 3 SEM images of fluorapatite crystals. (a) Virgin fluorapatite crystal with sharp edges. (b) Edge of a fluorapatite crystal after 6 h of incubation in the presence of Burkholderia sp. strain 205. Edges are corroded and the etching pattern does not follow crystalline axes. (c) and (d) Fluorapatite crystal before (c) and after
(d) incubation by immersion for 22 h in the presence of Burkholderia sp. strain 205. In image d, the remnants of the particle still present on the carbon tape are complex oxides. (e) and (f) Fluorapatite crystals after immersion for 23 h in the presence of Burkholderia sp. strain 205. Those crystals were part of an ore
containing 4% fluorine. The crystals are partially weathered, and weathering patterns follow crystalline axes. Fig. 3
338x406mm (300 x 300 DPI)
Page 36 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany

Draft
Figure 4 Reconstitution of the association between L. bicolor and Burkholderia sp. strain 205. Arrows indicate where the bacterial inoculum was deposited. (a) Laccaria bicolor in the presence of Burkholderia sp. strain 205 in the distal compartment with the soluble P+hydroxyapatite treatment. Inset: the hyphae of L.
bicolor devoid of PSB. (b) Laccaria bicolor in the presence of Burkholderia sp. strain 205 on the distal compartment of the hydroxyapatite treatment. Inset: the hyphae of L. bicolor covered with PSB.
Fig. 4 398x203mm (300 x 300 DPI)
Page 37 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany
Draft
Figure 5 Mean diameter of colonies of Burkholderia sp. strain 205, unknown species strain 260 and Curtobacterium sp. strain 168 growing in the presence of L. bicolor in the distal compartment containing sources of phosphorus, either soluble (KH2PO4), insoluble (hydroxyapatite), or both simultaneously. Error
bars represent standard deviation. Fig. 5
287x153mm (300 x 300 DPI)
Page 38 of 38
https://mc06.manuscriptcentral.com/botany-pubs
Botany