
Available at: http://www.ictp.it/˜pub_off IC/2006/010
United Nations Educational Scientific and Cultural Organization and
International Atomic Energy Agency
THE ABDUS SALAM INTERNATIONAL CENTRE FOR THEORETICAL PHYSICS
WATER EXTRACTABLE PHYTOCHEMICALS FROM CAPSICUM PUBESCENS (TREE PEPPER) INHIBIT LIPID PEROXIDATION INDUCED BY
DIFFERENT PRO-OXIDANT AGENTS IN BRAIN
G. Oboh* Biochemistry Department, Federal University of Technology,
P.M.B. 704 Akure, Ondo State, Nigeria, Departamento de Quimica, Universidade Federal de Santa Maria (UFSM),
Campus Universitário – Camobi, 97105-900 Santa Maria RS, Brasil and
The Abdus Salam International Centre for Theoretical Physics, Trieste, Italy
and
J.B.T. Rocha
Departamento de Quimica, Universidade Federal de Santa Maria (UFSM), Campus Universitário – Camobi, 97105-900 Santa Maria RS, Brasil.
MIRAMARE – TRIESTE
March 2006
___________________________ * Junior Associate of ICTP. Corresponding author: [email protected]

1
ABSTRACT
Reactive oxygen species (ROS) is the cause of neurodegenerative disorders such as Lou
Gehrig's disease, Parkinson's disease and Huntington's disease; one practical way to
prevent and manage neurodegenerative diseases is through the eating of food rich in
antioxidants (dietary means). In this study, the antioxidant and neuroprotective properties
of aqueous extract of ripe and unripe Capsicum pubescens (popularly known as tree
pepper) on different pro-oxidant induced lipid peroxidation in Rat’s brain (in vitro) is
been investigated. Aqueous extract of freshly harvested pepper was prepared, and the
total phenol content, vitamin C, ferric reducing antioxidant property (FRAP) and Fe (II)
chelating ability was determined. In addition, the ability of the extracts to protect the
Rat’s brain against some pro-oxidant (FeSO4, Sodium nitroprusside and Quinolinic acid)
– induced oxidative stress was also determined. The results of the study revealed that
ripe Capsicum pubescens had a significantly higher (P<0.05) total phenol [ripe
(113.7mg/100g), unripe (70.5mg/100g)] content and ferric reducing antioxidant property
than the unripe pepper. However, there was no significant difference in the vitamin C
[ripe (231.5µg/g), unripe (224.4µg/g)] content and Fe (II) chelating ability. Furthermore,
the pepper extracts caused a significant decrease (P<0.05) in 25µM Fe(II), 7µM Sodium
Nitroprusside and 1mM Quinolinic acid induced lipid peroxidation in the Rat’s brain in a
dose-dependent manner. However, the ripe pepper inhibited MDA (Malondialdehyhide)
production in the Rat’s brain than the unripe pepper. Conversely, both extract did not
significantly inhibit Fe (II)/ H2O2 induced decomposition of deoxyribose. Therefore, ripe
and unripe Capsicum pubescens would inhibit lipid peroxidation in vitro. However, the
ripe potent was a more potent inhibitor of lipid peroxidation, which is probably due to its
higher vitamin C and phenol content, reducing power and Fe (II) chelating ability .

2
INTRODUCTION
Pepper is an important agricultural crop, not only because of its economic importance,
but also for the nutritional value of its fruits, mainly because they are an excellent source
of natural colours and antioxidant compounds (Howard et al. 2000; Lee et al. 1995). The
intake of these compounds in food is an important health-protecting factor. They have
been recognized, as being beneficial for prevention of widespread human diseases. A
wide spectrum of antioxidant vitamins and phenolic compounds are present in pepper
fruits. Phenol compounds retard or inhibit lipid autoxidation by acting as radical
scavengers (Namiki, 1990); the antiradical activity of phenolics is principally based on
the redox properties of their hydroxy groups and the structural relationships between
different parts of their chemical structure (Rice-Evans et al. 1996; Rice-Evans et al.
1997). Consequently, they are essential antioxidants that protect against propagation of
the oxidative chain. It is also known that pepper fruits contain vitamin C. Vitamin C can
chelates heavy metal ions (Namiki, 1990), reacts with singlet oxygen and other free
radicals, and suppresses peroxidation (Bielski et al. 1975), reducing the risk of
arteriosclerosis, cardiovascular diseases, and some forms of cancer (Harris, 1996).
Reactive oxygen species (ROS), which include free radicals such as superoxide
anion radicals, hydroxyl radicals and non free-radical species such as H2O2 and singled
oxygen, are various forms of activated oxygen (Gulcin et al. 2002; Halliwell and
Gutteridge, 1999; Yıldırım et al. 2001). These molecules are exacerbating factors in
cellular injury and aging process (Lai et al. 2001). In living organisms, various ROS can
be formed in different ways. Normal aerobic respiration stimulates polymorphonuclear
leukocytes and macrophages, and peroxisomes appear to be the main endogenous sources
of most of the oxidants produced by cells.
Exogenous sources of ROS include tobacco smoke, certain pollutants, organic
solvents, and pesticides (Davies, 1994; Halliwell and Gutteridge, 1989; Robinson et al.
1997). ROS can cause lipid peroxidation in foods, which leads to the deterioration of the
food (Miller et al. 1995; Sasaki et al. 1996). In addition, reactive oxygen species induce
some oxidative damage to biomolecules like lipids, nucleic acids, proteins and
carbohydrates. Their damage causes ageing, cancer, and many other diseases (Aruoma,
1994). As a result, ROS have been implicated in more than 100 diseases, including

3
malaria, acquired immunodeficiency syndrome, heart disease, stroke, arteriosclerosis,
diabetes, and cancer (Alho and Leinonen, 1999; Duh, 1998; Hertog et al. 1993; Tanizawa
et al. 1992; Yıldırım et al. 2001).
Iron an essential metal for normal cellular physiology can result in cell injury
when in excess. This is because it plays a catalytic role in the initiation of free radical
reactions. The mechanism by which iron can cause this deleterious effect is that Fe (II)
can react with hydrogen peroxide (H2O2) to produce the hydroxyl radical (OH•) via the
Fenton reaction, whereas superoxide can react with iron(III) to regenerates iron(II) that
can participate in the Fenton reaction (Harris et al. 1992; Fraga and Oteiza, 2002).
Sodium nitroprusside is an anti-hypertensive drug, it acts by relaxation of vascular
smooth muscle; consequently it dilates peripheral arteries and veins. However, Sodium
nitroprusside (SNP) has been reported to cause cytotoxicity through the release of
cyanide and/or nitric oxide (NO) (Arnold et al 1984; Bates et al. 1990). NO is a free
radical with short half-life (< 30 s). Although NO acts independently, it also may cause
neuronal damage in cooperation with other reactive oxygen species (ROS) (Halliwell and
Gutteridge, 1999).
Quinolinic acid (QA) is a neuroactive metabolite of the tryptophan-kinurenine
pathway, which can be produced by macrophages and microglia (Cammer, 2001). It is
present in both the human and rat brain (Wolfensberger et al. 1983) and it has been
implicated in the pathogenesis of a variety of human neurological diseases (Belle et al.
2004). Quinolinic acid is recognized pharmacologically as an endogenous glutamate
agonist with a relative selectivity for the NMDA receptor in the brain (Vega-Naredo et al.
2005). Since it is not readily metabolized in the synaptic cleft, it stimulates the NMDA
receptor for prolonged periods. This sustained stimulation results in opening of calcium
channels causing Ca2+ influx followed by Ca2+-dependent enhancement of free radical
production leading to molecular damage and often to cell death (Cabrera et al. 2000).
The brain and nervous system are thought to be particularly vulnerable to
oxidative stress due to limited antioxidant capacity (Vega-Naredo et al. 2005). The brain
is responsible for about two percent of a person's mass but consumes 20 percent of their
metabolic oxygen. The vast majority of this energy is used by the neurons (Shulman et al.
2004). Of particular importance, neurons cannot synthesize glutathione, a fundamental

4
component of aerobic cell antioxidant machinery, but instead rely on surrounding
astrocyte cells to provide useable glutathione precursors (Shulman et al. 2004). Because
the brain has limited access to the bulk of antioxidants produced by the body, neurons are
the first cells to be affected by a shortage of antioxidants, and are most susceptible to
oxidative stress (Shulman et al. 2004).
Capsicum pubescens commonly known as tree pepper, its distinctive thick-fleshed
pungent fruits are used as a vegetable condiment or made into a sauce (Brown, 1995;
Facciola, 1990). As a hot pungent flavour, it is mainly used as flavouring in cooked foods
(Brown, 1995). In Peru the seeds are removed, the fruit stuffed with a savoury filling and
then baked (Facciola, 1990). The fruit can be dried and ground into a powder for use as a
pepper-like condiment (Brown, 1995). The hot and pungent fruit is antihaemorrhoidal
when taken in small amounts, antirheumatic, antiseptic, diaphoretic, digestive, irritant,
rubefacient, sialagogue and tonic (Chiej, 1984; Brown, 1995). It is taken internally in the
treatment of the cold stage of fevers, debility in convalescence or old age, varicose veins,
asthma and digestive problems (Brown, 1995). Externally it is used in the treatment of
sprains, unbroken chilblains, neuralgia, pleurisy etc. (Brown, 1995). However, there are
limited information with regard to the ability of the pepper to protect brain against
oxidative stress, this study therefore sought to determine and compare the antioxidant
activity and neuro-protective ability of ripe and unripe Capsicum pubescens against
different pro-oxidant induced oxidative stress in Rat’s brain- In vitro.
MATERIALS AND METHODS
Materials
Fresh samples of ripe and unripe Capsicum pubescens were collected from a vegetable
garden in Camobi, Santa Maria RS, Brazil. The authentication of the pepper was carried
out in Departamento de Biologia, Universidade Federal de Santa Maria, Santa Maria RS,
Brazil. All the chemicals used were analytical grade, while the water was glass distilled.
The handling and the use of the animal were in accordance with NIH Guide for the care
and use of laboratory animals. In the experiments Wister strain albino rats weighing 200
– 230g were collected from the breeding colony of Departamento de Biologia,

5
Universidade Federal de Santa Maria, Santa Maria RS, Brazil. They were maintained at
25oC, on a 12h light/12h dark cycle, with free access to food and water.
Aqueous extract preparation
The inedible portion of the pepper were removed from the edible portion, this edible
portion was subsequently washed in distilled water. The aqueous extract of the fresh
pepper was subsequently prepared by homogenizing 1g of the pepper in 20ml-distilled
water, and the homogenates were centrifuge at 2000rpm for 10 minutes. The supernatant
was used for the assay.
Total Phenol Determination
The total phenol content was determined by adding 0.5ml of the aqueous extract of the
pepper to 2.5ml, 10% Folin-Cioalteu’s reagent (v/v) and 2.0ml of 7.5% Sodium carbonate
was subsequently added. The reaction mixture was incubated at 45oC for 40min, and the
absorbance was measured at 765nm in the spectrophotometer, Gallic acid was used as
standard phenol (Singleton et al. 1999).
Vitamin C Determination
The vitamin C was determined using the method of Benderitter et al. (1998), briefly 75µl
of DNPH (2g of dinitrophenyl hydrazine, 230mg thiourea and 270mg in 100ml 5M
H2SO4) was added to 500µl reaction mixture ( 300µl of the pepper extracts with 100µl
13.3% trichloroacetic acid and water respectively). The reaction mixture was
subsequently incubated for 3 hr at 37oC, then 0.5 ml H2SO4 65% (v/v) was added to the
medium, and the absorbance was measured at 520nm, and the Vitamin C content of the
sample was subsequently calculated, using vitamin C standard curve.
Fe2+ Chelation assay
The ability of the aqueous extract to chelate Fe2+ was determined using a modified
method of Minotti and Aust (1987) with a slight modification by Puntel et al. (2005).
Briefly 150µl of freshly prepared 500µM FeSO4 was added to a reaction mixture
containing 168µl of 0.1M Tris-HCl (pH 7.4), 218µl saline and the aqueous extract of the

6
pepper (0 - 25µl). The reaction mixture was incubated for 5min, before the addition of
13µl of 0.25% 1, 10-Phenanthroline (w/v). The absorbance was subsequently measured at
510nm in the spectrophotometer
Degradation of Deoxyribose (Fenton´s Reaction)
The ability of the aqueous extract of the pepper to prevent Fe2+/ H2O2 induced
decomposition of deoxyribose was carried out using the method of Halliwell and
Gutteridge (1981). Briefly, freshly prepared aqueous extract (0 – 100µl) was added to a
reaction mixture containing 120µl 20mM deoxyribose, 400µl 0.1M phosphate buffer,
40µl 20mM hydrogen peroxide and 40µl 500µM FeSO4, and the volume for made to
800µl with distilled water. The reaction mixture was incubated at 37oC for 30min, and
the reaction was stop by the addition of 0.5ml of 2.8% TCA (Trichloroacetic acid), this
was followed by the addition of 0.4ml of 0.6% TBA solution. The tubes were
subsequently incubated in boiling water for 20min. The absorbance was measured at
measured at 532nm in spectrophotometer.
Reducing Property
The reducing property of the pepper extracts were determined by assessing the ability of
the extract to reduce FeCl3 solution as described by Pulido et al. (2000), briefly 2.5ml
aliquot was mixed with 2.5ml, 200mM Sodium phosphate buffer (pH 6.6) and 2.5ml of
1% Potassium ferricyanide. The mixture was incubated at 50oC for 20minutes, thereafter
2.5ml, 10% Trichloroacetic acid was added, and subsequently centrifuged at 650rpm for
10 minutes, 5ml of the supernatant was mixed with equal volume of water and 1ml of
0.1% ferric chloride, the absorbance was later measured at 700nm, a higher absorbance
indicates a higher reducing power.
Preparation of Brain Homogenates
The rats were decapitated under mild diethyl ether anaesthesia and the cerebral tissue
(whole brain) was rapidly dissected and placed on ice and weighed. This tissue were
subsequently homogenized in cold saline (1/10 w/v) with about 10-up-and –down strokes
at approximately 1200 rev/min in a Teflon –glass homogenizer. The homogenate was
centrifuge for 10min at 3000xg to yield a pellet that was discarded and a low-speed

7
supernatant (S1) containing mainly water, proteins and lipids (cholesterol, galactolipid,
individual phospholipids, gangliosides), DNA and RNA was kept for lipid peroxidation
assay (Belle et al. 2004).
Lipid peroxidation and thiobarbibutric acid reactions
The lipid peroxidation assay was carried out using the modified method of Ohkawa et al.
(1979), briefly 100µl S1 fraction was mixed with a reaction mixture containing 30µl of
0.1M pH 7.4 Tris-HCl buffer, pepper extract (0 - 100µl) and 30µl of the pro-oxidant
solution (250µM freshly prepared FeSO4, 70µM Sodium nitroprusside and 1mM
Quinolinic acid). The volume was made up to 300µl by water before incubation at 37oC
for 1h. The colour reaction was developed by adding 300µl 8.1% SDS (Sodium doudecyl
sulphate) to the reaction mixture containing S1, this was subsequently followed by the
addition of 600µl of acetic acid/HCl (pH 3.4) mixture and 600µl 0.8% TBA
(Thiobarbituric acid). This mixture was incubated at 100oC for 1h. TBARS
(Thiobarbituric acid reactive species) produced were measured at 532nm and the
absorbance was compared with that of standard curve using MDA (Malondialdehyde).
Analysis of data
The result of the replicates were pooled and expressed as mean±standard error (S.E.)
(Zar, 1984); student t-test was carried out. Significance was accepted at P≤ 0.05. The
EC50 (extract concentration that will cause 50% inhibition of lipid peroxidation in the
Rat’s brain) of the pepper extracts were determined using the method of Dixon & Massey
(1969).
RESULTS AND DISCUSSION
Oxidative stress has been extensively studied in neurologic disease including Alzheimer’s
disease, Parkinson’s disease, multiple sclerosis, AIDS dementia and so on. This is not
surprising since the brain (neural cells) is especially susceptible to oxidative stress and
subsequent damage to cells (including cell death). In a disease such as Alzheimer’s,
oxidative stress / oxidative damage are felt to play a key role in the loss of neurons and
the progression to dementia. The most likely and practical way to fight against

8
degenerative diseases is to improve the body’s antioxidant status, which could be
achieved by a greater consumption of vegetables (Oboh, 2004; Oboh, 2005a). Ability of
Capsicum pubescens to prevent different pro-oxidant induced oxidative stress in Rat’s
brain (In vitro) are highlighted below.
The results of the total phenol and vitamin C content of the pepper are presented
in table 1. The results revealed that there was no significant difference (P>0.05) in the
vitamin C content of the ripe and unripe Capsicum pubescens [ripe (231.5 µg/g), unripe
(224.4µg/g)]. However, the vitamin C content was lower than that of Capsicum annum,
Tepin [ripe (332.5 µg/g) & unripe (250.2 µg/g)], but higher than that of Capsicum
chinese, Habanero [ripe (152.5 µg/g) & unripe (103.6µg/g)] (Oboh et al. 2005).
Moreover, these values were generally lower than the vitamin C content of Capsicum
annuum L cv. Vergasa (Marin et al. 2004) and what Chu et al. (2002) reported for red
pepper. Nevertheless, the vitamin C content of the pepper was above that of lettuce,
cucumber, celery, cabbage, onion, carrot and some tropical green vegetables (Chu et al.
2002; Oboh, 2005b). Furthermore, the vitamin C content of this pepper is above that of
pineapple, banana, apple, but lower than that of orange, lemon and strawberry (Sun et al.
2002).
However, the ripe pepper had higher vitamin C content than the unripe pepper,
the trend agrees with what Marin et al. (2004) reported for Capsicum annuum, L.cv.
Vergasa, and what Oboh et al. (2005) reported for Capsicum annuum, Tepin and
Capsicum chinese, Habanero. This goes a long way to confirm that ripe pepper will
likely be a better source of dietary vitamin C than unripe pepper. Vitamin C is found in
fruits, particularly fruits and juices and green leafy vegetables, they protect the body
against cancer of the oesophagus, oral cavity and stomach, it also helps to maintain the
blood vessel flexibility and improves blood circulation in the arteries of smokers as well
as facilitating the absorption of iron in the body. As an antioxidant, vitamin C’s primary
role is to neutralize free radicals; it can work for both inside and outside the cells to
combat free radical damage (Block et al. 1992; Umar et al. 1998).
However, Pietta (2000) reported that the phenolic compounds are responsible for
most of the antioxidant activity in plants; the presence of derivatives of cinnamic acid and
flavonoids has been found in pepper fruit. However, numerous studies have conclusively

9
shown that the majority of the antioxidant activity may be from compounds such as
flavonoids, isoflavone, flavones, anthocyanin, catechin and isocatechin rather than from
vitamins C, E and β-carotene (Sun et al. 2002; Chu et al. 2002; Marin et al. 2004;
Materska & Perucka, 2005). In addition, to their role as antioxidant, some of them are
known to have anti-inflammatory and antiproliferative activities (Sun et al. 2002; Chu et
al. 2002). Polyphenols are capable of removing free radicals, chelate metal catalysts,
activate antioxidant enzymes, reduce α-tocopherol radicals, and inhibit oxidases (Amic et
al. 2003; Alia et al. 2003; Oboh, 2006a). The results of the total phenol content of the
pepper as shown in table 1 revealed that the ripe pepper had significantly higher (P<0.05)
total phenol content than the unripe pepper [ripe (117.7mg/100g) & unripe
(70.5mg/100g)]
However, the total phenol content of this pepper (ripe and unripe) is below that of
Capsicum annuum, Tepin, but within the same range with that of Capsicum chinese,
Habanero (Oboh et al. 2005). Furthermore, the trend in the phenol content with ripening
agrees with the trend in Capsicum chinese, Habanero (Oboh et al. 2005) and what
Materska and Perucka (2005) reported for some varieties of hot pepper [Capsicum annum
L (Bronowicka Ostra, Cyklon, Tomado and Tajfun)], in that the ripe pepper had higher
total phenol content than the unripe pepper. Furthermore, the total phenol content was
within the same range with what Chu et al. (2002) reported for broccoli, spinach, onion
and red pepper, but above that of carrot, cabbage, potato, lettuce, celery and cucumber.
As presented in figure 1 and table 2, aqueous extract of the ripe and unripe
Capsicum pubescens (1.7 – 8.3mg/ml) caused a significant decrease (P<0.05) in the lipid
peroxidation in the Rat’s brain homogenates in a dose-dependent manner. However, the
ripe pepper (EC50 = 3.5mg/ml) caused a significantly higher (P<0.05) inhibition in the
MDA production in the brain tissues than the unripe pepper (EC50 = 7.2mg/ml). The
inhibition of lipid peroxidation in the Rat’s brain by the aqueous extract could be ascribed
to the presence of both antioxidant vitamins and polyphenol compounds present in the
pepper. Furthermore, it is worth noting that ripening increases the ability of the pepper to
prevents MDA production in the brain tissue homogenates, unlike Capsicum annuum,
Tepin, that ripening decreases the protective ability of the pepper on brain tissue (Oboh et
al. 2005).

10
This increasing in the protective ability of the aqueous extracts on the brain tissue
could be attributed to the fact that pepper undergoes some physiological changes during
ripening that leads to changes in the phytochemicals (Marin et al. 2004; Materska and
Perucka, 2005). This change in phytochemicals during ripening may have lead to an
increase in the total phenol and vitamin C content of the pepper. This may have
accounted for the enhanced protective ability of the ripe pepper, as many studies had
conclusively reported that there are correlation between plant phenol content and
antioxidant properties (Chu et al. 2002; Sun et al. 2002; Alia et al. 2003; Oboh, 2005b;
Oboh, 2006b). In addition, the EC50 of the pepper was higher than Capsicum annuum,
Tepin and Capsicum chinese, Habanero that are considered to be the hottest pepper
(Pepper Gallery, 2005)
Figure 2 shows the interaction (inhibition) of the pepper extract with Fe (II)-
induced lipid peroxidation in Rat’s brain. The result revealed that incubation of the brain
tissue in the presence of 25µM Fe (II) caused 260% increase in the MDA content of the
brain homogenates when compared with the basal brain homogenates. The increased lipid
peroxidation in the presence of Fe2+ could be attributed to the fact that Fe2+ can catalyze
one-electron transfer reactions that generate reactive oxygen species, such as the reactive
OH·, which is formed from H2O2 through the Fenton reaction. Iron also decomposes lipid
peroxides, thus generating peroxyl and alkoxyl radicals, which favors the propagation of
lipid oxidation (Zago et al. 2000). High levels Fe play a crucial role in brain cancer and in
degenerative diseases of the brain (Parkinson’s and Alzheimer's diseases, multiple
sclerosis, etc.) (Johnson, 2001).
However, aqueous extracts of Capsicum pubescens caused a significant inhibition
(P<0.05) in Fe (II) induced lipid peroxidation in the Rat’s brain in a dose-pedendent.
Nevertheless, the ripe pepper caused a significantly higher (P<0.05) inhibition in the
MDA production in the Rat’s brain tissue than the unripe pepper. The decrease in the
Fe2+ induced lipid peroxidation in the Rat’s brain homogenates in the presence of the
extracts could be as result of the ability of the extracts to chelate Fe2+. In addition,
scavenge free radicals produced by the Fe2+ catalyzed production of reactive oxygen
species (ROS) in the Rat’s brain homogenates by the antioxidant phytochemicals in the
extract of the peppers. Peppers are rich in antioxidants vitamin as well as phenols;

11
phenolics compound are more potent antioxidant in foods than vitamin C and E (Marin
et al. 2004; Materska and Perucka, 2005; Oboh and Akindahunsi, 2004, Oboh, 2005a;
Oboh, 2006b). The higher ability of the ripe pepper to protect the rat’s brain could be
attributed to its higher phenol and vitamin C content.
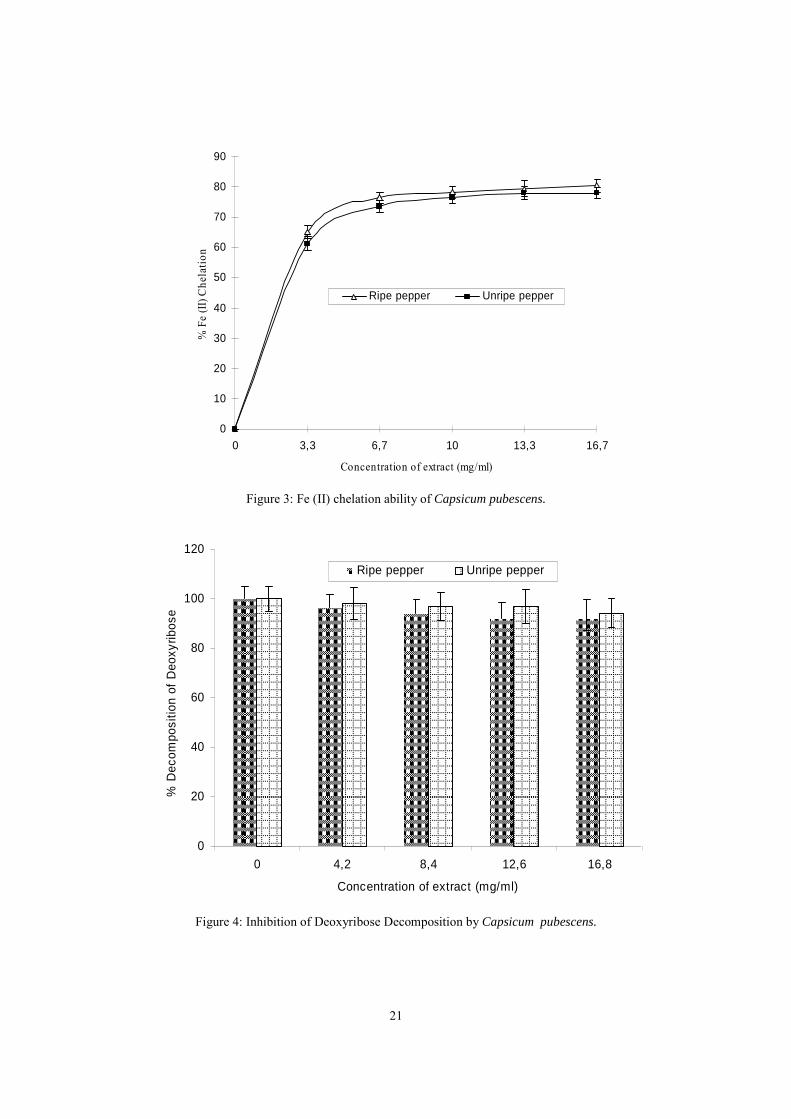
The use of Fe chelator is a popular therapy for the management of Fe (II)
associated oxidative stress in brain. The Fe (II) chelating ability of the pepper extracts
are presented in figure 3. The results revealed that both extract significantly (P<0.05)
chelate Fe (II) in a dose-dependent manner. However, there was no significant (P>0.05)
difference in the Fe (II) chelating ability of the ripe and unripe pepper, nevertheless the
ripe pepper extract had higher Fe (II) chelating ability. It is worth noting that, correlation
exists between the total phenol content and Fe (II) chelating ability. Conversely, the
extracts did not significantly inhibits (P>0.05) OH radical (Fe (II) / H2O2) -induced
decomposition of deoxyribose as shown in figure 4. The reason for the inability of the
extract to prevent deoxyribose decomposition could not be categorically stated.
However, it is likely that the complex formed between the pepper phytochemicals and
Fe, could not initiate lipid peroxidation, but active in Fenton reaction.
Furthermore, the ripe pepper had a significantly higher (P<0.05) ability to reduce
Fe (III) to Fe (II) than the unripe pepper (figure 5). This result agrees with the trend in
vitamin C content presented in table 1. It could reason that phenol and vitamin C are
actively involved in the protection of brain tissue from Fe (II) induced lipid peroxidation,
the mechanism through they possibility do this is by their Fe(II) chelating ability and
reducing, and not through hydroxyl radical scavenging ability.
The interaction of the aqueous extract of Capsicum pubescens with sodium
nitroprusside and quinolinic acid induced lipid peroxidation in rat’s brain are presented
in figures 6 & 7 respectively. The results revealed that the extract caused a significant
decrease (P<0.05) in the brain MDA content in sodium nitroprusside and quinolinic acid
induced lipid peroxidation in a dose dependent manner. However, the ripe pepper had a
significantly higher (P<0.05) inhibitory effect in the lipid peroxidation induced by both
pro-oxidants than the unripe pepper. The higher inhibitory effect of the ripe pepper on
lipid peroxidation in the Rat’s brain is in line with the vitamin C and total phenol
content, reducing power and Fe chelating ability of the pepper.

12
The protective properties of the pepper against sodium nitroprusside induced
lipid peroxidation on the brain could be because of the ability of the antioxidant
phytochemicals present in the aqueous extract to quench-scavenge the nitrous radical
and Fe produced from the decomposition of sodium nitroprusside. While the inhibition
of the quinolinic acid induced lipid peroxidation in the brain tissues could at least in part,
be by preventing over stimulation of NMDA receptor and free radical scavenging ability
of the phenol content.
CONCLUSION
Capsicum pubescens could prevent Fe (II), sodium nitroprusside and quinolinic acid
induced lipid peroxidation in brain, this they do through the antioxidant activity of their
phytochemicals (Phenols and vitamin C). However, ripening which brings about a
change in the pigment of pepper caused an increase in the phenol and vitamin C content;
and consequently cause an increase in the protective ability of the Capsicum pubescens
ability to protect rats brain from the pro-oxidant induced lipid peroxidation by enhancing
Fe (II) chelating and reducing power of the pepper.
ACKNOWLEDGMENTS
The authors wish to acknowledge the Conselho Nacional de Desenvolvimento Científico
e Tecnológico (CNPq) Brazil and Academy of Science for the Developing World
(TWAS), Trieste Italy; for granting Dr G. Oboh, Post-Doctoral fellowship tenable at
Biochemical Toxicology Unit of the Department of Chemistry, Federal University of
Santa Maria, Brazil. This study was also supported by CAPES, FIPE/UFSM, VITAE
Foundation and FAPERGS. In addition, the authors apppreciate the support of The Abdus
Salam International Centre for Theoretical Physics, Trieste, Italy. Financial support from
the Swedish International Development Cooperation Agency is acknowledged.

13
REFERENCES
Alho H, Leinonen J. 1999. Total antioxidant activity measured by chemiluminescence
method. Methods of Enzymology 299: 3–15.
Alia M, Horcajo C, Bravo L, Goya L. 2003. Effect of Grape Antioxidant Dietary Fiber
on the Total Antioxidant Capacity and the Activity of Liver Antioxidant Enzymes
in Rats. Nutrition Research 23: 1251 – 1267.
Amic D, Davidovic-Amic D, Beslo D, Trinajstic N. 2003. Structure-Radical
Scavenging Activity Relationship of Flavonoids. Croatia Chemical Acta 76 (1): 55 –
61.
Arnold WP, Longneeker DE, Epstein RM. 1984. Photodegradation of sodium
ntroprusside: biologic activity and cyanide release, Anesthesiology 61: 254– 260.
Aruoma OI.1994. Nutrition and health aspects of free radicals and antioxidants. Food
and Chemical Toxicology 62: 671–683.
Bates JN, Baker MT, Guerra R. Harrison DG. 1990. Nitric oxide generation from
nitroprusside by vascular tissue, Biochemical Pharmacolology 42: S157–S165.
Belle NAV, Dalmolin GD, Fonini G, Rubim MA, Rocha JBT. 2004. Polyamines reduces
lipid peroxidation induced by different pro-oxidant agents. Brain Research 1008:
245 – 251.
Benderitter M, Maupoil V, Vergely C, Dalloz F, Briot F, Rochette L.1998. Studies by
electron paramagnetic resonance of the importance of iron in the hydroxyl
scavenging properties of ascorbic acid in plasma: Effects of iron chelators.
Fundamentals of Clinical Pharmacology 12: 510–516.
Bielski BH, Richter HW, Chan PC. 1975. Some properties of the ascorbate free radical.
Annals of the New York Academy of Sciences, 258, 231–237.
Block G, Patterson B, Subar A. 1992. Fruits, Vegetables and Cancer Prevention. A
review of epidemiological evidence. Nutrition and Cancer 18: 1- 29.
Brown D. 1995. Encyclopaedia of Herbs and their Uses. Dorling Kindersley, London.
Cabrera J, Reiter RJ, Tan D, Qi W, Sainz RM, Mayo JC, Garcia JJ, Kim SJ,
El-Sokkary G. 2000. Melatonin reduces oxidative neurotoxicity due to
quinolinic acid: in vitro an in vivo findings, Neuropharmacology 39: 507– 514.
Cammer W. 2001. Oligodendrocyte killing by quinolinic acid in vitro, Brain Research:

14
896, 157– 160.
Chiej R. 1984. Encyclopaedia of Medicinal Plants. MacDonald
Chu Y, Sun J, Wu X, Liu RH. 2002. Antioxidant and antiproliferative activity of
common vegetables. Journal of Agricultural and Food Chemistry 50: 6910 –
6916.
Davies KJA. 1994. Oxidative stress: the paradox of aerobic life. Biochemistry Society
Symphosium 61: 1–34.
Dixon WJ, Massey, FJ 1969. Introduction to statistical analysis. McGraw-Hill,
third edition
Duh PD. 1998. Antioxidant activity of burdock (Arctium lappa Linne): its scavenging
effect on free radical and active oxygen. Journal of the American Oil Chemist’s
Society 75: 455–465.
Facciola. S. 1990. Cornucopia - A Source Book of Edible Plants. Kampong
Publications
Fraga CG, Oteiza PI. 2002. Iron toxicity and antioxidant nutrients, Toxicology
180: 23 – 32.
Gulcin I, Oktay M, Kufrevioglu OI, Aslan A. 2002. Determination of Antioxidant
activity of Lichen Cetraria islandica (L) Ach. Journal of Ethnopharmacology,
79(3): 325–329.
Halliwell B, Gutteridge JMC. 1981. Formation of a thiobarbituric-acid-reactive
substance from deoxyribose in the presence of iron salts: The role of superoxide
and hydroxyl radicals. FEBS Letter 128: 347–352.
Halliwell B, Gutteridge JM. 1989. Free radicals in biology and medicine. Clarendon
Press.
Halliwell B, Gutteridge JM. 1999. Free radicals in biology and medicine. Oxford:
Oxford University Press.
Harris JR. 1996. Subcellular Biochemistry, Ascorbic acid: Biochemistry and
Biomedical Cell Biology (Vol. 25). New York:Plenum.
Harris ML, Shiller HJ, Reilly PM, Donowitz M, Grisham MB, Bulkley GB. 1992.
free radicals and other reactive oxygen metabolites in inflammatory bowel
disease: cause, consequence or epiphenomenona? Pharmacology & Therapeutics

15
53: 375–408.
Hertog MGL, Feskens EJM, Hollman PCH, Katan MB, Kromhout D.1993. Dietary
antioxidant flavonoids and risk of coronary heart disease: the zupthen elderly
study. Lancet 342: 1007–1014.
Howard LR, Talcott ST, Brenes CH, Villalon B. 2000. Changes in phytochemical and
antioxidant activity of selected pepper cultivars (Capsicum species) as influenced
by maturity. Journal of Agricultural of Food Chemistry 48: 1713–1720.
Johnson S. 2001.The possible crucial role of iron accumulation combined with low
tryptophan, zinc and manganese in carcinogenesis, Medical Hypotheses 57(5):
539-543
Lai LS, Chou ST, Chao WW. 2001. Studies on the antioxidative activites of Hsian-tsao
(Mesona procumbens Hemsl) leaf gum. Journal of Agricultural and Food
Chemistry 49: 963–968.
Lee Y, Howard LR, Villalon B. 1995. Flavonoids and antioxidant activity of fresh
pepper (Capsicum annum) cultivars. Journal of Food Science 60: 43–476.
Marin A, Ferreres F, Tomas-Barberan FA, Gil MI. 2004. Characterization and
quantitation of antioxidant constituents of Sweet pepper (Capsicum annuum L.).
Journal of Agricultural and Food Chemistry 52: 3861-3869.
Materska M, Perucka I. 2005. Antioxidant activity of the main phenolic compounds
Isolated from Hot pepper fruit (Capsicum annuum L.). Journal of Agricultural
and Food Chemistry 53: 1750 – 1756
Miller NJ, Diplock AT, Rice-Evans CA. 1995. Evaluation of the total antioxidant
activity as a marker of the deterioration of apple juice in storage.
Journal of Agricultural and Food Chemistry, 43, 1794–1801.
Minotti G, Aust SD. 1987. An investigation into the mechanism of citrate-Fe2+ -
dependent lipid peroxidation. Free Radical Biology and Medicine 3: 379-387.
Namiki M. 1990. Antioxidants/antimutagens in food. CRC Critical Reviews in Food
Science and Nutrition 29: 273–300.
Oboh G. 2004. Prevention of Garlic Induced Haemolytic Aneamia by Some
Commonly Consumed Green Leafy Vegetables in Nigeria. Journal of Medicinal
Food 7 (4): 500 - 503

16
Oboh G. 2005a. Hepatoprotective property of ethanolic and aqueous extracts of
Telfairia occidentalis (Fluted Pumpkin) leaves against garlic-induced oxidative
stress. Journal of Medicinal Food 8 (4): 560 - 563
Oboh G. 2005b. Effect of blanching on the antioxidant properties of some tropical green
leafy vegetables. Lebensm.-Wiss.u.-Technol 38 (5): 513 – 517.
Oboh G. 2006a. Antioxidant and antimicrobial properties of ethanolic extract of
Ocimum gratissimum leaves. Journal of Pharmacology and Toxicology 1 (1): 47
– 53
Oboh G. 2006b. Antioxidant properties of some commonly consumed & underutilized
tropical legumes. European Food Research and Technology – In press
Oboh G, Akindahunsi AA. 2004. Change in the ascorbic acid, total phenol and
antioxidant activity of sun-dried commonly consumed green leafy vegetables in
Nigeria. Nutrition & Health 18: 29-36.
Oboh G, Puntel, RL, Rocha JBT. 2005. Hot pepper (Capsicum annuum, Tepin
& Capsicum chinese, Habanero) Prevents Fe2+-induced Lipid Peroxidation in
Brain- In vitro. A Report Submitted to Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) Brazil, 31p
Ohkawa H, Ohishi N, Yagi K. 1979. Assay for lipid peroxides in animal tissues by
thiobarbituric acid reaction. Analytical Biochemistry 95: 351 – 358.
Pepper Gallery 2005. http://www.floridata.com/tracks/peppergallery/pepper_menu.htm
Pietta PG. 2000. Flavonoids as Antioxidants. Journal of Natural Products 63: 1035-
1042
Pulido R, Bravo L, Saura-Calixto F. 2000. Antioxidant activity of dietary polyphenols as
determined by a modified ferric reducing/antioxidant power assay.
Journal of Agricultural and Food Chemistry 48: 3396 – 3402
Puntel RL, Nogueira CW, Rocha JBT. 2005. Krebs cycle intermediates modulate
thiobarbituric acid reactive species (TBARS) production in rat brain in
vitro. Neurochemical Research 30 (2): 225-235
Rice-Evans CA, Miller NJ, Paganga G. 1996. Structure-antioxidant activity
relationships of flavonoids and phenolic acids. Free Radical Biology and
Medicine 20: 933-956.

17
Rice-Evans CA, Miller J, Paganga G. 1997. Antioxidant properties of phenolic
compounds. Trends in Plant Science 2: 152-159.
Robinson EE, Maxwell SRJ, Thorpe GHG. 1997. An investigation of antioxidant activity
of black tea using enhanced chemiluminescence. Free Radical Research, 26, 291–
302.
Sasaki S, Ohta T, Decker EA. 1996. Antioxidant activity of water-soluble fraction of
salmon spremary tissue. Journal of Agricultural and Food Chemistry 44: 1682–
1686.
Shulman RG, Rothman DL, Behar KL, Hyder F.2004. Energetic basis of brain
activity: implications for neuroimaging. Trends in Neuroscience 27(8): 489-95.
Singleton VL Orthofer R, Lamuela-Raventos RM 1999. Analysis of total phenols and
other oxidation substrates and antioxidants by means of Folin-Ciocalteu Reagent.
Methods of Enzymology 299: 152 – 178.
Sun J, Chu Y, Wu X, Liu R. 2002.Antioxidant and antiproliferative activities of common
fruits. Journal of Agricultural and Food Chemistry 50: 7449 – 7454 .
Tanizawa H, Ohkawa Y, Takino Y, Ueno A, Kageyama T, Hara S. 1992. Studies on
natural antioxidants in citrus species. I. Determination of antioxidant activities of
citrus fruits. Chemical and Pharmaceutical Bulletin 40: 1940–1942.
Umar IA, Arjinoma ZG, Gidado A, Hamza HH. 1998. Prevention of garlic (Allium
sativum Linn)-induced anaemia in rats by supplementation with ascorbic
acid and vitamin E. Nigerain Journal of Biochemistry and Molecular
Biology 13:31–40.
Vega-Naredo I, Poeggeler B, Sierra-Sanchez V, Caballero B, Tomas-Zapico C,
Alvarez-Garcıa O, Tolivia D, Rodrıguez-Colunga MJ, Coto-Montes A
2005. Melatonin neutralizes neurotoxicity induced by quinolinic acid in brain
tissue culture. Journal of Pineal Research 39:266–275
Wolfensberger M, Amsler U, Cuenod M, Foster AC, Whetsell Jr. WO, Schwarcz
R. 1983. Identifications of quinolinic acid in rat and human brain tissue,
Neuroscience Letter 41:247– 252.
Yildirim A, Mavi A, Kara AA. 2001. Determination of antioxidant and antimicrobial
activities of Rumaxs crispus L. extracts. Journal of Agricultural and Food

18
Chemistry 49: 4083–4089.
Zago MP, Verstraeten SV, Oteiza PI. 2000. Zinc in the prevention of Fe2+initiated lipid
and protein oxidation. Biological Research.33 (2): 143-150.
Zar J. H. 1984. Biostatistical Analysis, Prentice-Hall, Inc. USA, p620.

19
Table 1: Total phenol content and Vitamin C content of Capsicum pubescens Sample Vitamin C Total Phenol (µg/g) (mg/100g gallic acid equivalent) Ripe 231.5±11.5a 117.7±10.5a Unripe 224.4±9.20a 70.5±7.8b Values represent means of triplicate. Values with the same alphabet along the same column are not significantly different (P>0.05)
Table 2: EC50 of Aqueous extract of Capsicum pubescensi Sample EC50 (mg/ml) Ripe 3.5 ±1.2 b Unripe 7.2±0.8a Values represent means of triplicate. Values with the same alphabet along the same column are not significantly different (P>0.05)

20
0
20
40
60
80
100
120
0 1,7 3,3 5 6,7 8,3
Concentration of extract (mg/ml)
MD
A (%
Con
trol)
Ripe pepper Unripe pepper
Figure 1: Capsicum pubescens inhibits lipid peroxidation in Rat´s brain.
200
210
220
230
240
250
260
270
280
0 4,2 8,4 12,6 16,8
Concentration of extract (mg/ml)
MD
A (%
Con
trol)
Ripe pepper Unripe pepper
Figure 2: Capsicum pubescens inhibits Fe (II) induced lipid peroxidation in Rat´s brain.
21
0
10
20
30
40
50
60
70
80
90
0 3,3 6,7 10 13,3 16,7
Concentration of extract (mg/ml)
% F
e (II
) Che
latio
n
Ripe pepper Unripe pepper
Figure 3: Fe (II) chelation ability of Capsicum pubescens.
0
20
40
60
80
100
120
0 4,2 8,4 12,6 16,8
Concentration of extract (mg/ml)
% D
ecom
posi
tion
of D
eoxy
ribos
e
Ripe pepper Unripe pepper
Figure 4: Inhibition of Deoxyribose Decomposition by Capsicum pubescens.

22
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0 3,3 6,7 10
Concentration of extract (mg/ml)
OD
700
Ripe pepper Unripe pepper
Figure 5: Reducing power of Capsicum pubescens.
0
20
40
60
80
100
120
140
160
180
200
0 4,2 8,4 12,6 16,8
Cocentration of extract (mg/ml)
MD
A (%
Con
trol)
Ripe pepper Unripe pepper
Figure 6: Capsicum pubescens inhibits Sodium nitroprusside induced lipid peroxidation
in Rat´s brain.

23
0
50
100
150
200
250
0 4,2 8,4 12,6 16,8
Concentration of extract (mg/ml)
MD
A (%
Con
trol)
Ripe pepper Unripe pepper
Figure 7: Capsicum pubescens inhibits Quinolinic acid induced lipid peroxidation in
Rat´s brain.






![THE SYNTHESIS CONDITIONS, CHARACTERIZATIONS ...users.ictp.it/~pub_off/preprints-sources/2008/IC2008030P.pdfphase separation method using dimethyl sulphoxide [15], synthesis of 2,3-di-O-carboxymethyl](https://cdn.vdocuments.us/doc/165x107/6065cdf14abbd1491559fe10/the-synthesis-conditions-characterizations-usersictpitpuboffpreprints-sources2008.jpg)
